

Abstract
Background
Although telomere shortening occurs as a natural part of aging, there is now a robust body of research that suggests that there is a relationship between psychosocial, environmental, and behavioral factors and changes in telomere length. These factors need to be considered when integrating telomere measurement in biobehavioral research studies.
Objectives
This article provides a brief summary of the known facts about telomere biology and an integrative review of current human research studies that assessed relationships between psychosocial, environmental, or behavioral factors and telomere length.
Methods
An integrative review was conducted to examine human research studies that focused on psychosocial, environmental, and behavioral factors affecting telomere length and telomerase activity using the electronic databases PubMed/Medline and CINAHL from 2003 to the present. In addition to the known individual factors that are associated with telomere length, the results of the integrative review suggest that perceived stress, childhood adversities, major depressive disorder, educational attainment, physical activity, and sleep duration should also be measured.
Discussion
Multiple factors have been shown to affect telomere length. To advance understanding of the role of telomere length in health and disease risk, it will be important to further elucidate the mechanisms that contribute to telomere shortening.
Keywords: aging, biobehavior, nursing research, oxidative stress, psychological stress, telomere
The identification of telomeres as an important structure for maintaining genetic stability and description of their role in regulating cellular senescence provided insight on the mechanisms of aging at a molecular level. Pivotal findings from research dating back to the early 20th century (Table 1) fostered an intense area of research focused on understanding the relationships among telomere biology, aging, and disease. More recently, telomere length (TL) has emerged as a promising bio-marker to assess the cumulative influence of psychosocial, environmental, and behavioral factors on complex disease risk (especially cancer). However, a majority of human studies that comprise this growing body of research entail associations between TL and complex multidimensional variables that make it difficult to elucidate the specific physiological mechanisms undermining telomere maintenance. To design clinically relevant bio-behavioral research studies with findings that can be translated into novel diagnostic and treatment approaches, it is important to understand and be able to quantify the psychosocial–behavioral contexts in which alterations in telomere biology occur.
TABLE 1.
Historical Milestones in the Science of Telomere Structure and Function
| Year | Milestone |
|---|---|
| 1930s | Hermann Muller and Barbara McClintock observed that fragmented chromosomes tended to fuse together whereas normal chromosomes were stable and proposed that the “sealed ends” of chromosomes help to maintain their integrity. |
| 1938 | Muller names the “sealed ends” of chromosomes telomeres. |
| 1941 | McClintock hypothesized that telomeres stabilize chromosome ends by preventing them from being recognized as DNA double strand breaks. |
| 1961 | Leonard Hayflick and Paul Moorhead discovered that cells cultured in vitro undergo a type of “counting process” that establishes the limit for the number of divisions that a cell can complete, which is now known as replicative senescence or the “Hayflick limit.” |
| 1973 | Alexey Olovnikov and James Watson recognize a fundamental problem in the mechanics of DNA replication with loss of genetic information during each cycle of cell division; Olovnikov proposed the theory of marginotomy to explain the function of telomeres by reasoning that the repeated noncoding telomeric nucleotide sequences act as a buffer to protect gene coding sequences. |
| 1979 | Elizabeth Blackburn and Joseph Gall discover the structure of telomeres in the ciliated protozoan Tetrahymena. |
| 1982 | Blackburn and Jack Szostak determine how telomeres in yeast serve as a buffer for the coding DNA during replication and postulated the existence of an enzyme that could rebuild telomeres. |
| 1985 | Blackburn and Carol Greider provide the first report of an enzymatic activity in Tetrahymena extracts that is capable of extending a telomeric oligonucleotide in vitro. |
| 1987 | The enzyme is named telomerase and shown to contain both protein and RNA as necessary components (based on sensitivity of the activity to protease and RNase). |
| 1989 | The RNA is purified and shown to contain a sequence with complementarity to the Tetrahymena telomeric repeat; the mechanism of telomerase function as a reverse transcriptase is proposed. |
| 1990 | Yu et al. show that introducing a mutated telomerase RNA into Tetrahymena results in synthesis of abnormal telomeric repeats, proving that the RNA acts as a template and that telomerase synthesizes telomeric repeats in vivo; although a role for telomerase in vivo had been speculated since the original 1985 article, this article provided the first clear evidence for an in vivo role in telomere synthesis. |
The purpose of this article was to summarize and critically evaluate the current state of knowledge regarding TL in humans. Known facts about telomere biology are summarized, followed by an integrative review of current human research studies that assessed relationships between psychosocial, environmental, or behavioral factors and TL. Definitions of key terms are listed in Table 2.
TABLE 2.
Definitions of Key Terms
| Term | Definition |
|---|---|
| Apoptosis | Process of programmed cell death that is characterized by changes in cell morphology followed by death |
| Mitosis | A form of cell division by which a cell that has replicated its chromosomes separates the chromosomes into two identical sets |
| Oxidative stress | An abnormality in the normal mechanisms of cellular signaling that is caused by an imbalance between the reactive oxygen species present and the body’s ability to detoxify the toxic effects of these intermediate species |
| p53 activation | Transcriptional activation of p53, which is a tumor suppressor, encoded by the TP53 gene |
| Senescence | Process of biological aging that could be either organismal senescence (at the level of the whole organism) or cellular senescence (at the cellular level) |
| Telomere | The region of repetitive nucleotide sequences at the chromatid ends of eukaryotic chromosomes |
| Telomerase | A ribonucleoprotein, an enzyme that adds DNA sequence repeats at the 3′ end of the DNA strands in the telomere regions |
| Telomere attrition | The process in which the terminal caps of the chromosomes become shorter due to reasons such as aging, tumorigenesis, oxidative stress amongst others. |
Telomere Structure and Function
Telomeres are protein-bound DNA repeat structures at the end of chromosomes that regulate cellular replicative capacity, protect chromosomes from fusing together during mitosis, and prevent the loss of genetic data (Blackburn, Greider, & Szostak, 2006). During cell division, the telomere shortens by 30 to 200 base pairs because of the inability of DNA polymerase to fully replicate the 3′ end of the DNA strand (Aubert & Lansdorp, 2008). This is known as the end replication problem and results in a gradual decline in TL over time. Consequently, leukocyte TL (LTL) shortens in a predictable way with age by roughly 20–40 base pairs per year (Cesare & Reddel, 2010). Cell division may occur 50–70 times before the telomere reaches a critically short length (Hayflick limit), in which the telomere becomes too close to the coding DNA region and the loop structures cannot hold the telomere structure together (Figure 1). Ultimately, the exposed telomere is recognized by tumor suppressors (such as p53 protein), which activate DNA damage checkpoints and induce cellular senescence or apoptosis (Zakian, 2012). In this manner, telomere shortening plays a protective role by limiting the proliferative capacity of cells that have sustained DNA damage and are vulnerable to oncogenic transformation.
FIGURE 1.
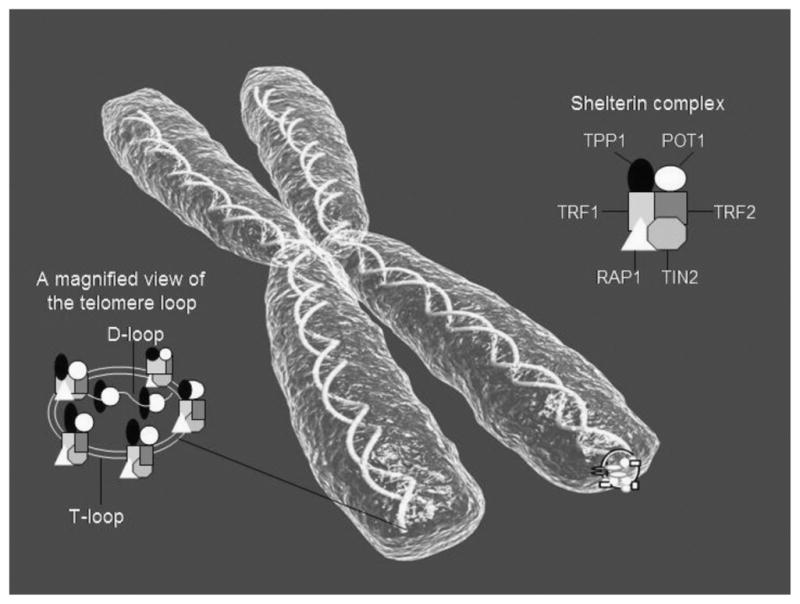
Telomere biology. The telomere protects adjacent chromosomes by creating a cap at the end in conjunction with specialized proteins to compose a telomere-shelterin complex. In humans, the telomere is composed of arrays of guanine-rich base pair repeats that terminate with a 3′ single-stranded DNA overhang, known as the G-tail. When the cell is not undergoing mitosis, the double-stranded DNA forms a loop structure called a telomere loop (T-loop). At the very end of the T-loop, the single-stranded telomere DNA is bound to a region of double-stranded DNA to form the displacement loop (D-loop). The T-loop and D-loop are stabilized by telomere-binding proteins, which are collectively referred to as the shelterin complex. Proteins of the shelterin complex include telomeric repeat binding factor 1 (TRF1) and 2 (TRF2), protein protection of telomeres 1 (POT1), repressor activator protein 1 (Rap1), tripeptidyl peptidase 1 protein (TPP1), and TRF1-interacting nuclear factor 2 (TIN2). With telomere shortening, these proteins cannot be recruited to form the T-loops, thus leaving the chromosome ends uncapped and vulnerable to erosion and fusion. ©iStockphoto.com/Virginia Commonwealth University.
In cell types that continue to replicate over the lifespan, TL is maintained by the enzyme telomerase, a ribonucleoprotein that contains the RNA template TERC and a reverse transcriptase TERT (Blackburn et al., 2006). Telomerase replenishes the lost base pairs that are not replicated by DNA polymerase during transcription. In humans, telomerase activity is present in germ cells, stem cells, and a subset of somatic cells (e.g., active fibroblasts). However, most somatic cells lack or have very minimal telomerase activity (Zanni & Wick, 2011). Peripheral blood mononuclear cells (PBMCs) express telomerase at low levels, which can be measured over a short duration (hours) to demonstrate immediate, short-term changes, unlike TL, which takes months for changes to be detected (Epel et al., 2010).
In most cancers, telomerase activity is increased due to genetic mutations (or polymorphisms in TERC and/or TERT) that reactivate telomerase despite DNA damage and chromosomal instability within the cell (Zanni & Wick, 2011). In addition, mechanisms known as alternate lengthening of telomeres can be activated that lengthen telomeres in the absence of telomerase and allow abnormal proliferation of cells (Chen & McLeskey, 2010). Mutations in telomerase and telomere genes resulting in abnormalities in telomere maintenance are responsible for a spectrum of syndromes ranging from dyskeratosis congenita to a progressive phenotype of idiopathic pulmonary fibrosis (Ballew & Savage, 2013).
In addition to the end replication problem, telomere shortening can occur as a consequence of oxidative stress and inflammation that cause DNA damage and breaks with potential loss of large telomeric parts during cell division (Sahin & DePinho, 2010). Oxidative stress occurs when there is an increased production of reactive oxygen species (ROS) beyond the body’s ability to detoxify the reactive intermediates or repair the damage they cause to proteins, lipids, and DNA. Due to the high guanine content of the telomere, they are particularly susceptible to single- and double-strand DNA breaks caused by ROS, and telomere breaks are less efficiently repaired than any other location in the genome (Shen et al., 2009).
Telomere length is measured in chromosomes, cells, or genomic DNA. There are several different methods used to measure TL, and each approach has distinct advantages and disadvantages. The first method described—Terminal Restriction Fragment (TRF) length analysis—uses Southern blotting or in-gel hybridization with a labeled probe specific for telomere DNA. The TRF method provides a mean TL of a cell population. The TRF method is technically challenging, requires substantial amounts of DNA, and is relatively insensitive to detect very short telomeres; however, it is considered to be the gold-standard technique (Aubert, Hills, & Lansdorp, 2012). Advancements in polymerase chain reaction (PCR), a technique for amplifying the number of copies of a specific region of DNA, have allowed for measurement of TL using smaller amounts of DNA. Methods based on PCR include single TL analysis, which measures the specific length of a single chromosome end, as well as quantitative PCR and monochrome multiplex quantitative PCR, which amplify telomeres to generate a ratio between the telomere and a standard single copy gene. The PCR-based methods require very few cells; however, variability within and between samples remains high. Other methods use fluorescent in situ hybridization (FISH) using digital microscopy (quantitative FISH) or flow cytometry (flow FISH) to determine cell average TL. The FISH methods require fresh cell samples, and the required calibrations can be both time-consuming and technically demanding; however, they are highly accurate and can detect subtle changes in TL (Canela, Vera, Klatt, & Blasco, 2007).
Telomere length is genetically determined, with heritability estimates ranging from 44% to 80%, and is highly variable between individuals. There is evidence that an individual’s mean LTL and TL in other tissues are highly correlated (Zakian, 2012). A strong maternal inheritance and paternal age effect on TL was recently reported in a meta-analysis that included 19,713 subjects (Broer et al., 2013). Although there is no gender difference at birth, in some cohorts, female adults have been shown to have longer telomeres than male adults did. In addition, differences in TL have been noted among different racial and ethnic groups (Schaefer et al., 2012; Zanni & Wick, 2011). Thus, age, sex, race, ethnicity, paternal age at birth, and genetic mutations of telomerase and telomere maintenance genes are nonmodifiable factors that contribute to TL.
Shorter age- and sex-adjusted mean LTL has been advanced as a potentially important biomarker in studies focused on accelerated aging, disease risk, and longevity. Support for the premise that TL is involved in aging and disease comes from studies in which TLs have been positively associated with longevity (longer telomeres in centenarians; Atzmon et al., 2010; Garagnani et al., 2013) and negatively associated increased risk of cancer (patients with these conditions have shortened telomeres; Codd et al., 2013; Ma et al., 2011). A recent multiethnic epidemiological study of 100,000 people found that individuals whose telomeres were in the shortest 10% were approximately 23% more likely to die in the 3 years after measurement of their telomeres (Schaefer et al., 2012). They also found that although telomeres did get shorter with age, men older than 75 years and women older than 80 years tended to have longer telomeres than did their slightly younger counterparts, suggesting that those who survive to older age carry longer telomeres.
Although TL assessments related to aging and cancer have been among the most prominent areas of telomere biology investigations, recent evidence, albeit not conclusive, supports the notion that psychosocial conditions, as well as environmental and behavioral exposures, can also impact TL. To summarize and critically evaluate the current state of knowledge regarding the assessment of TL in humans, an integrative review was performed on current human research studies that assessed the relationships between psychosocial, environmental, or behavioral factors and TL.
Methods
An integrative review was conducted to examine human research studies that focused on factors affecting TL and telomerase activity. The search of the electronic databases PubMed/ Medline and CINAHL, from 2003 to the present, used the key words telomere length, telomerase, psychological stress, mental health, social environment, and behavior. Manual searches were also completed using references from previously published studies and literature reviews. The 284 articles that were retrieved via these multiple search methods were reviewed for duplication and whether they met the following inclusion criteria:
Research studies that involved human participants with a primary aim of assessing the relationship between psychosocial, environmental, and/or behavioral (physical activity and sleep) factors and TL and/or telomerase activity.
The study reported clear methodologies for measuring TL and/or telomerase activity.
The study used a clinical diagnosis of psychiatric disorders (as opposed to self-report).
It was written in English (see Figure 2 for the process of the literature selection). A total of 50 articles met the inclusion criteria and are included in this review (Tables 3–11).
FIGURE 2.

Literature review process.
TABLE 3.
Psychosocial Factors: Chronic Psychological Stress
| Author | Date | Samples | Method | Findings |
|---|---|---|---|---|
| Epel et al. | 2004 | 39 mothers with chronically ill children 19 mothers with healthy children |
qPCR | Chronicity and perceived stress associated with telomere attrition (Equivalent to up to 17 years accelerated aging) |
| Damjanovich et al. | 2007 | 41 caregivers of patients with AD 41 controls, age and gender matched |
Southern blot | Telomere attrition in caregivers (loss of 6.2–6.4 kb compared with controls) |
| O’Donovan et al. | 2009 | 36 female healthy postmenopausal caregivers and noncaregivers | qPCR | Pessimism independently associated with shorter LTL |
| Parks et al. | 2009 | 647 women Sisters of women with breast cancer |
qPCR | Perceived stress modestly associated with telomere attrition and stress hormones |
| Humphreys et al. | 2011 | 61 women exposed to interpartner violence 41 healthy female controls |
qPCR | Telomere attrition in those exposed to interpartner violence Length of time in the abusive relationship and having children associated with shortened telomeres |
Note. qPCR = quantitative polymerase chain reaction; AD = Alzheimer’s disease; kb = kilobases; LTL = leukocyte telomere length.
TABLE 11.
Behavioral Factors: Sleep Quality and Duration
| Authors | Date | Sample | Method | Findings |
|---|---|---|---|---|
| Liang et al. | 2011 | 4,117 women Nurses’ Health Study |
qPCR | Women G50: significant association between sleep duration and LTL ≤6 hours sleep per night associated with shorter LTL |
| Prather et al. | 2011 | 245 community-dwelling women 49–66 years |
qPCR | Poor self-reported sleep quality associated with shorter LTL |
| Jackowska et al. | 2012 | 434 men and women Whitehall II Cohort Study 63.3 years (average) |
qPCR | In men only, shorter sleep duration associated with shorter telomeres |
Note. qPCR = quantitative polymerase chain reaction; LTL = leukocyte telomere length.
Results
Psychosocial Factors
Chronic Psychological Stress
Information about studies assessing associations between chronic stress and TL are summarized in Table 3. Seminal findings that linked TL with chronic psychological stress were first reported by Epel et al. (2004). In this study, LTL was evaluated in PBMCs along with telomerase activity and an index of oxidative stress in 58 mothers who were caring for either a healthy or a chronically ill child. There was no difference in LTL between the two groups; however, on comparison of extreme high and low levels of psychological stress, the high-stress group had significantly shorter telomeres, lower telomerase activity, and higher oxidative stress. Although the findings were based on cross-sectional analysis and the sample size was small, the length of caregiving was significantly associated with shorter LTL, suggesting that chronic stress was important in this relationship.
Similar findings were reported by Damjanovic et al. (2007), who compared depressive symptoms, PBMC composition, cytokine production, LTL, and telomerase activity between caregivers of individuals with Alzheimer’s disease and a group of age- and gender-matched controls. Depressive symptoms were significantly higher in caregivers along with higher production of immune-regulatory cytokines, and care-givers had significantly shorter LTL in PBMCs than controls did; however, basal telomerase activity in PBMCs and T cells was significantly higher in caregivers. A weakness of this study was that perceived stress, as well as the caregiver’s health status and level of caregiving, was not assessed.
To examine whether personality traits influence the relationship between psychological stress and LTL, O’Donovan et al. (2009) measured dispositional traits in 36 healthy post-menopausal women who were caregivers and noncaregivers. Pessimism was independently associated with shorter LTL and higher levels of interleukin-6 (IL-6); however, optimism was not associated with either measure. The association between pessimism and LTL remained significant when controlling for age, caregiver status, optimism, perceived stress, neuroticism, body mass index (BMI), exercise, and sleep but not for IL-6. As with the previous studies, the cross-sectional design and small sample size were major limitations.
Using a larger sample, Parks et al. (2009) evaluated LTL, current perceived stress, and urinary stress hormones in women enrolled in the National Institute of Environmental Health Sciences Sister Study, a cohort of women aged 35–74 years who had a sister with breast cancer. Women with higher perceived stress had somewhat shorter LTL, but it did not decrease monotonically with higher stress levels. Significantly shorter LTL was found in women 55 years or older, with recent major losses and above average urinary catecholamines. Finally, Humphreys et al. (2012) reported significantly shorter LTL in women with chronically elevated psychological stress related to interpartner violence and found that length of time in the abusive relationship and having children were associated with shortened telomeres.
Collectively, these studies suggest that there is a link between chronically high levels of perceived stress and shortened LTL in a subset of participants. Nonetheless, several aspects of the design and methodologies of these studies warrant caution in generalizing the findings, particularly because most of the studies included only women, and the psychological stress endured in most of the studies was related to the caregiving role. Most importantly, the cross-sectional nature of these studies provides only a glimpse of TL; therefore, it cannot be inferred that accelerated LTL shortening continues over time. Even with the large sample size in the study of Parks et al. (2009), the relationship between psychological stress and telomere shortening was modest, yet the small elevation in perceived stress in the subgroups with shorter LTL was comparable with associations with obesity, smoking, or diagnosis of cardiovascular disease (CVD; Table 3).
Acute Psychological Stress
Three studies addressing the association between acute stress and TL are summarized in Table 4. Following their study from 2004, Epel et al. (2006) assessed the relationship between acute psychological stress, TL, and telomerase activity in leukocytes of 62 healthy women. Low telomerase activity was associated with exaggerated autonomic reactivity to acute mental stress and elevated nocturnal epinephrine, as well as major risk factors for CVD, including smoking, poor lipid profile, high systolic blood pressure, high fasting glucose, and greater abdominal adiposity. Shorter LTL was related to elevated catecholamines and cortisol. These findings, along with those of a study by Bekaert et al. (2007), suggest an association between markers of hypothalamic–pituitary–adrenocortical (HPA) axis dysregulation, inflammation, and oxidative stress with decreased telomerase activity and shortened telomeres. Epel et al. (2010) also found that elderly highly stressed caregivers had lower levels of basal telomerase activity compared with low-stress controls, but during a stress task, telomerase activity increased by approximately 18% in both groups along with an increase in cortisol. This study suggests that increased telomere shortening associated with chronically high perceived stress (in caregivers) is related to decreased basal telomerase activity and not an altered response to acute stress.
TABLE 4.
Psychosocial Factors: Acute Psychological Stress
| Authors | Date | Samples | Method | Findings |
|---|---|---|---|---|
| Epel et al. | 2006 | 64 healthy women | qPCR | Shorter LTL associated with increased catecholamines and cortisol |
| Bekaert et al. | 2007 | 2,509 community-dwelling Caucasian women and men No overt CVD 35–55 years old |
qPCR | Greater age-dependent telomere attrition in men compared with women Associated with inflammation, oxidative stress markers Associated with higher BMI and lack of exercise |
| Epel et al. | 2010 | 22 high-stress dementia caregivers and 22 low-stress age-matched controls | qPCR | Baseline: lower levels of telomerase activity of PBMCs in caregivers than in controls Stress task: increase in telomerase activity by approximately 18% in both groups |
Note. qPCR = quantitative polymerase chain reaction; LTL = leukocyte telomere length; CVD = cardiovascular disease; BMI = body mass index; PBMC = peripheral blood mononuclear cell.
Maternal Stress
Two studies to date have assessed the relationship between stress during pregnancy and the offspring’s LTL (Table 5). In the first study, Entringer et al. (2011) assessed the association between maternal stress during pregnancy and LTL of adult offspring and found that prenatal stress exposure was a significant predictor of subsequent adult offspring LTL. Although maternal stress was evaluated retrospectively, the authors controlled for age, sex, BMI, birth weight percentile, postnatal early-life adversity, chronic stress, and depressive symptoms. The second study assessed maternal pregnancy-specific psychological stress and newborn LTL in a prospective study of 27 mother–newborn dyads (Entringer et al., 2013). The authors reported a significant, independent, linear effect of pregnancy-specific stress on newborn LTL that accounted for 25% of the variance in adjusted LTL after accounting for gestational age at birth, weight, sex, and exposure to antepartum obstetric complications. While the results are intriguing, other potential determinants of newborn LTL, particularly the mother’s CVD risk factors and paternal age at birth, were not included in the analyses (Broer et al., 2013).
TABLE 5.
Psychosocial Factors: Maternal Stress
| Authors | Date | Sample | Method | Findings |
|---|---|---|---|---|
| Entringer et al. | 2011 | 94 healthy adults (45 offspring of mothers with a severe stressor in the index pregnancy and 49 with mothers who had a healthy, uneventful index pregnancy) 25 years old (average) | qPCR | Prenatal stress exposure significantly predicted subsequent offspring LTL during adulthood. There was a 178-base pair difference between groups. |
| Entringer et al. | 2013 | 27 mother–newborn dyads (singleton pregnancies, no known pre-pregnancy major medical conditions or fetal anomalies) | qPCR | There was a significant, independent, linear effect of pregnancy-specific stress on newborn LTL that accounted for 25% of the variance in adjusted LTL. |
Note. qPCR = quantitative polymerase chain reaction; LTL = leukocyte telomere length.
Childhood Maltreatment
The notion that other forms of psychological or physical stress during critical time points of growth and development could lead to stress-induced changes in LTL in later life was also assessed by several investigators (Table 6). The association between childhood maltreatment and shortened telomeres was studied by Tyrka et al. (2010) in 31 adults who reported either no childhood maltreatment or moderate to severe levels of childhood maltreatment on any of the five subscales of the Childhood Trauma Questionnaire. Participants reporting a history of physical or emotional neglect had significantly shorter telomeres than did those who did not. However, the association did not reach statistical significance for physical or emotional abuse. Glass, Parts, Knowles, Aviv, and Spector (2010) reported no association between being a victim of physical, emotional, or sexual abuse and LTL in participants of the twins UK cohort. However, the assessment of abuse entailed only whether it had occurred (yes/no) and not duration or extent.
TABLE 6.
Psychosocial Factors: Childhood Maltreatment
| Authors | Date | Samples | Method | Findings |
|---|---|---|---|---|
| Tyrka et al. | 2010 | 31 adults with no current or past major Axis I psychiatric disorders (10 reported moderate to severe maltreatment and 21 reported no maltreatment) | qPCR | Telomere attrition in those exposed to childhood neglect (not abuse) |
| Glass et al. | 2010 | Reported childhood maltreatment (123 cases vs. 1751 controls) Reported childhood maltreatment and sexual abuse (34 cases vs. 516 controls) Reported childhood maltreatment and physical abuse (20 cases vs. 520 controls) |
Southern blot | Childhood maltreatment not linked to telomere attrition |
| Kananen et al. | 2010 | 321 anxiety disorder 653 matched controls 30–87 years |
qPCR | Shorter telomere length associated with a greater number of childhood adverse life events in both groups Significant age- and sex-adjusted effect of number of childhood adversities on LTL in adulthood |
| Surtees et al. | 2011 | 4,441 women UK European Prospective Investigation into Cancer–Norfolk Study |
qPCR | Adverse childhood experiences associated with shorter LTL Each additional childhood adversity equivalent to an approximate 3-year increase in aging |
| Kiecolt-Glaser et al. | 2011 | 132 healthy older adults Community sample (58 were dementia family caregivers and 74 were noncaregivers) |
qPCR | Multiple childhood adversities related to increased IL-6 Multiple childhood adversities related to shorter telomeres (telomere difference equivalent to 7–15 years shortened lifespan) |
| Shalev et al. | 2013 | 236 children Environmental-Risk Longitudinal Twin Study |
qPCR | Significantly more telomere erosion between the ages of 5 and 10 years in children who experienced two or more kinds of violence compared with participants exposed to one type or no violence |
Note. qPCR = quantitative polymerase chain reaction; LTL = leukocyte telomere length; IL-6 = interleukin-6.
Shorter TL was found to be associated with a greater number of childhood adverse life events among adults with and without anxiety disorder, and the number of childhood adversities was found to have a significant age- and sex-adjusted effect on LTL in adulthood (Kananen et al., 2010). The most significant childhood adversity was chronic or serious illness during childhood. No significant difference in LTL was evident overall between those with and without anxiety disorder; among older individuals, however, LTL was shorter among those with anxiety disorder. In a study by Surtees et al. (2011), social adversity, including traumatic circumstances experienced before age 17 years and lifetime occurrence of a set of stressful adverse life events, was associated with shorter LTL in 4,441 women. Similarly, multiple childhood adversities measured with the Childhood Trauma Questionnaire and a six-item tool were related to both increased IL-6 and shorter telomeres compared with the absence of adversity in a community sample of 132 healthy older adults (Kiecolt-Glaser et al., 2011). In the first prospective longitudinal study with repeated telomere measurements in children, Shalev et al. (2013) reported that exposure to two or more kinds of violence (maternal domestic violence, frequent bullying victimization, and physical maltreatment by an adult) was associated with significantly more telomere erosion between the ages of 5 and 10 years.
Overall, these studies provide some evidence of an association between childhood maltreatment and shortened telomeres, with both the type(s) of maltreatment or stress exposures and intensity of the exposures showing an association with TL. Similar to the studies on maternal stress, other influences on TL, such as paternal age at birth, as well as diet, exercise, and other health behaviors may have played a role in LTL shortening (Broer et al., 2013). Two challenges for this area of research entail identifying the most appropriate measures to assess childhood maltreatment and understanding the biological mechanisms that lead to shortened telomeres. Clearly, more longitudinal studies are needed to assess the long-term consequences of childhood maltreatment on LTL, as well as other outcomes of physical, emotional, and mental health. In addition, there has been a lack of research on aspects of resiliency that may play a protective role in mitigating the association between childhood maltreatment and LTL, which would help to inform the development of appropriate interventions.
Mental Health
Mental health is an integral component of successful aging. Several studies have assessed the relationship between a diagnosed mental health problem, mainly major depressive disorder (MDD), and LTL (Table 7). The first study to examine this association focused on mood disorders and LTL (Simon et al., 2006). Although there were no differences when comparing the three types of mood disorders individually (MDD and bipolar disorder with and without comorbid anxiety), when combined as a group, LTL was significantly shorter in those with mood disorders compared with the control group. In two other cross-sectional studies, LTL was compared between outpatients diagnosed with MDD (Lung, Chen, & Shu, 2007) and hospitalized patients receiving various treatment for MDD (Hartmann, Boehner, Groenen, & Kalb, 2010) and controls. Although these three studies reported an association between mood disorders/MDD and shorter telomeres, they were limited by a lack of ethnic/racial diversity, or data on paternal age at birth, and evaluation of stress levels.
TABLE 7.
Psychosocial Factors: Mental Health
| Authors | Date | Samples | Method | Findings |
|---|---|---|---|---|
| Simon et al. | 2006 | 15 major depressive disorder 15 bipolar disorder with comorbid anxiety 14 bipolar disorder without comorbid anxiety 44 age-matched controls |
Southern blot | LTL significantly shorter in those with mood disorders compared with controls Overall mean difference of 660 base pairs |
| Lung et al. | 2007 | 253 outpatients with MDD 411 healthy controls |
qPCR | Shorter telomeres in the group with MDD than in the control group (BMI, smoking, and exercise not reported) |
| Hartmann et al. | 2010 | 54 hospitalized with MDDa 20 age-matched controls |
Southern blot | LTL significantly shorter in the MDD group compared with the control group Mean difference of 350 base pairs |
| Hoen et al. | 2011 | 206 individuals with MDD and CAD 746 individuals without MDD |
qPCR | Shorter LTL in participants with MDD Insignificant when controlling for confounders |
| Malan et al. | 2011 | 64 women Survivors of rape |
qPCR | No association between relative LTL and development of MDD Weak association between LTL and PTSD |
| Wolkowitz et al. | 2011 | 18 unmedicated individuals with MDD 17 healthy age-, sex- and ethnicity-matched controls 17 healthy age-, sex- and ethnicity-matched controls |
qPCR | No significant difference between groups in LTL, oxidative stress, or inflammation (vitamin C, F2-isoprostanes, and IL-6) Significantly shorter LTL in those who experienced depression the longest than in the control group, by approximately 280 base pairs |
| Wolkowitz et al. | 2012 | 16 with MDD enrolled in open-label study with sertraline 18 healthy matched controls |
qPCR | Telomerase activity levels elevated in the MDD group compared with controls Higher baseline telomerase activity associated with higher levels of depression and stress but not LTL or measures of inflammation (IL-6, C-reactive protein) |
| Wikgren et al. | 2012 | 91 with diagnosed MDD 451 healthy controls |
qPCR | Significantly shorter LTL in participants with MDD who had low postdexamethasone suppression test cortisol levels than healthy controls |
| Puterman et al. | 2013 | 954 with stable CVD Heart and Soul Study |
qPCR | High multisystem resiliency predicted longer LTL whereas each individual factor did not; multisystem resiliency significantly moderated the MDD–LTL association |
Note. LTL = leukocyte telomere length; MDD = major depressive disorder; qPCR = quantitative polymerase chain reaction; BMI = body mass index; CAD = coronary artery disease; PTSD = posttraumatic stress disorder; IL-6 = interleukin-6; CVD = cardiovascular disease.
20 patients were receiving low doses of antidepressants, 16 were receiving high doses of antidepressants, and 18 were receiving antidepressants plus electroconvulsive therapy.
In patients with coronary artery disease, the relationship between shorter LTL and MDD became insignificant when controlling for age, gender, BMI, smoking status, physical inactivity, antidepressant use, anxiety, left ventricular ejection fraction, and statin use (Hoen et al., 2011). At 5 years after their baseline measures, MDD did not predict shorter TL; however, the study did not report whether MDD or the extent of CVD was reevaluated over that period of time. Malan, Hemmings, Kidd, Martin, and Seedat (2011) tested their hypothesis that shorter LTL would predict MDD or post-traumatic stress disorder (PTSD) in 64 women who were survivors of rape. They found a weak association between LTL and the development of PTSD and none with MDD at 3 months postbaseline measures (Malan et al., 2011). Still, this may have been too short of a time span to detect changes in TL (Epel et al., 2010).
Wolkowitz et al. (2011) reported a significant difference in LTL compared with a healthy control group only among individuals with MDD who experienced depression the longest. In a subsequent study (Wolkowitz et al., 2012), telomerase activity was higher in individuals with MDD compared with controls and was associated with higher levels of depression and stress. Although the sample sizes were small in both studies, the results suggest that the duration of MDD is an important factor and support a connection between telomerase activity and severity of MDD and perceived stress. A study by Wikgren et al. (2012) found that participants with MDD, who had low postdexamethasone suppression test cortisol levels, had shorter LTL, suggesting that the difference in LTL between MDD and control participants was due to hypocortisolemia. This finding is consistent with other studies that have described alterations in HPA axis dysfunction as one of the etiologies underlying MDD.
Thus, from the research that assessed the relationship between MDD and LTL, it appears that the duration and, possibly, severity of MDD are associated with some degree of increased LTL shortening compared with not having MDD. Similar to the other psychosocial factors reviewed, more longitudinal research on the mechanisms of MDD that may be responsible for increased telomere shortening is needed. In addition, there has been a lack of research on aspects of resiliency that may play a protective role in mitigating the association between depression and LTL. Recently, a study by Puterman et al. (2013) examined these aspects using a model of multisystem resiliency defined by health emotional regulation, strong social connections, and health behaviors in sleep and exercise. In this study of 954 participants with stable CVD, high multisystem resiliency predicted longer LTL, whereas each individual factor did not. Multisystem resiliency significantly moderated the MDD–LTL association, suggesting that biological outcomes should be examined within a psychosocial–behavioral context and supporting the notion that even small differences in resiliency and positive health behaviors can have a significant influence on TL.
Socioeconomic Status and Educational Attainment
Findings about associations between TL and socioeconomic status (SES)/educational attainment are summarized in Table 8. In 2006, Cherkas et al. reported an association between lower SES and shorter LTL in a cross-sectional study with 1,552 female twins. However, Adams et al. (2007) found no evidence of an association between any measure of SES and LTL in a homogeneous birth cohort at age 50 years. In addition, Batty et al. (2009) reported that unemployed men had significantly shorter telomeres than did those who were employed, but there was no association between LTL and educational attainment or residence within “area-based deprivation.” In contrast, Woo, Suen, Leung, Tang, and Ebrahim (2009) reported that a higher self-rated community standing was associated with shorter TL in men only.
TABLE 8.
Psychosocial Factors: Socioeconomic Status and Educational Attainment
| Authors | Date | Sample | Method | Findings |
|---|---|---|---|---|
| Cherkas et al. | 2006 | 1,552 female twins Subset discordant for low SES |
Southern blot | Telomere length significantly shorter in lower SES groups 14% difference accounted for by high BMI, smoking, and lack of exercise |
| Adams et al. | 2007 | 318 individuals Homogeneous birth cohort At age 50 years |
PCR | No evidence of association between any measure of SES and LTL when controlling for lifestyle variables, gender, and paternal age at birth |
| Batty et al. | 2009 | 1,542 men West of Scotland Coronary Prevention Study |
qPCR | Shorter LTL in unemployed than in employed men |
| Woo, Suen, et al. | 2009 | 958 men and 978 women Hong Kong community 65 years or older |
qPCR | Higher self-rated community standing associated with shorter telomere length in men only |
| Steptoe et al. | 2011 | 448 healthy men and healthy women Whitehall II cohort 53–76 years |
qPCR | Lower educational attainment associated with shorter LTL but not household income or employment |
| Surtees et al. | 2012 | 4,441 women Ethnically homogenous UK European Prospective Investigation Into Cancer–Norfolk study 41–80 years |
qPCR | Educational attainment but not social class associated with LTL Relationship not attenuated by social adversity, including adversity experienced during childhood |
| Needham et al. | 2012 | 70 children Community based Admixture Mapping for Ethnic and Racial Insulin Complex Outcomes study 7–13 years |
qPCR | Mean reduction of 1,178 base pairs in telomere length in children with parents who never attended college compared with children with at least one college-educated parent (difference equivalent to 6 years of additional aging) |
| Needham et al. | 2013 | 536 participants National Health and Nutrition Examination Survey, 1999–2002 |
qPCR | Significantly shorter LTL in those with less than a high school education Partially mediated by smoking and BMI but not drinking or sedentary behavior |
| Carroll, Diez-Roux, Adler, et al. | 2013 | 963 participants MESA: Stress Ancillary Study |
qPCR | Significant difference in LTL between home owners and renters in Hispanic and White participants but not African Americans |
| Carroll, Diez-Roux, Fitzpatrick, et al. | 2013 | 948 participants MESA 45–84 years at study entry |
qPCR | Low social support significantly associated with shorter LTL in adults 65 years or older but not younger participants after adjusting for sex, race, BMI, diabetes, pulse pressure, smoking, physical activity, and diet |
| Adler et al. | 2013 | 2,599 Black and White adults Health, Aging and Body Composition Study High-functioning 70–79 years old |
qPCR | Significantly shorter LTL in older adults with only a high school education compared with those with post-high school education |
Note. SES = socioeconomic status; BMI = body mass index; PCR = polymerase chain reaction; LTL = leukocyte telomere length; qPCR = quantitative polymerase chain reaction; MESA =Multi-Ethnic Study of Atherosclerosis.
A study involving participants of the Whitehall II epidemiological cohort (Steptoe et al., 2011) and another involving the UK European Prospective Investigation Into Cancer–Norfolk Study participants (Surtees et al., 2012) found that lower educational attainment was associated with shorter LTL. Both studies controlled for age, gender, blood pressure, glycated hemoglobin, high-density lipoprotein, cholesterol, smoking, BMI, and physical activity. Needham, Fernandez, Lin, Epel, and Blackburn (2012) found that parental education was positively associated with child LTL when controlling for sex, age, race/ethnicity, and family income. Using data from the National Health and Nutrition Examination Survey, 1999–2002, they also reported significantly shorter LTL in respondents who completed less than a high school degree compared with those who graduated from college (Needham et al., 2013).
Carroll, Diez-Roux, Adler, and Seeman (2013) reported a significant difference in LTL between homeowners and renters in Hispanic and White participants but not African Americans. There were no significant associations between educational attainment or family income and LTL, but there was an inverse association between the father’s educational level and LTL. In another study, Carroll, Diez-Roux, Fitzpatrick, and Seeman (2013) reported that low social support was significantly associated with shorter LTL in adults aged 65 years or older but not younger participants. In a different cohort of older adults, Adler et al. (2013) reported that those with only a high school education had significantly shorter LTL than did those with post-high school education, and a significant interaction of race and education revealed more beneficial effects of post-high school education for Blacks than for Whites.
Collectively, these studies suggest that there is a small but significant association between some aspects of SES, particularly educational attainment and social support with LTL. There is also some evidence of a relationship between parental educational attainment and LTL of their offspring. However, the connection between educational attainment and the behavioral and/or biological mechanisms that promote telomere maintenance remains to be elucidated.
Environment
Only two studies were identified that assessed the relationship between LTL and the environment, specifically, the amount of green space and neighborhood risk (Table 9). The association between LTL and living in an area of Hong Kong with more green spaces (near the river with more parks and markets) compared with residing in an older part of the city was assessed by Woo, Tang, Suen, Leung, and Wong (2009). They reported that shorter LTL was associated with living in the older regions of the city after adjusting for age, smoking, SES, and physical activity level. Theall, Brett, Shirtcliff, Dunn, and Drury (2013) evaluated the association between salivary TL (sTL) in children and neighborhood-level social environmental risk from data collected at the level of the child, family/household, and neighborhood. They found that children living in neighborhoods characterized by high disorder had lower sTL values than did children not living in a high disordered environment. The findings from these two studies, although limited, suggest that some characteristics of the environment may be related to shortened telomeres. Whether this association is due to perception, behavior (more or less opportunities to exercise), the effects of an unmeasured third variable on both environment and TL, or more direct mechanisms (pollutants) remains an area of investigation that requires greater attention.
TABLE 9.
Environmental Factors
| Authors | Date | Sample | Method | Findings |
|---|---|---|---|---|
| Woo, Tang, et al. | 2009 | 976 men Various regions of Hong Kong 65 years or older |
qPCR | Telomeres in participants living in older regions of Kowloon shorter than in those living in the newly renovated area with more green spaces |
| Theall et al. | 2013 | 99 children ages 4–14 from 87 neighborhoods who were recruited from 5 school districts in New Orleans | qPCR | Mean salivary TL value 7.4 telomere signal/single-copy signal ratio units and 4.7% of the variance attributed to differences across neighborhoods |
Note. qPCR = quantitative polymerase chain reaction; TL = telomere length.
Behavioral Factors
Oxidative stress and inflammation are two mechanisms that are known to cause accelerated telomere shortening. It follows that lifestyle behaviors that increase levels of oxidative stress and/or inflammation will also result in shortened telomeres. Smoking, which directly increases oxidative stress, is associated with a loss of 5 base pairs of telomeric sequence each year (Valdes et al., 2005). Clinically, these findings support that smoking one pack of cigarettes per day for 40 years could lead to telomeric attrition rates that would be equivalent to a loss of 7.4 years of life. Obesity is also a condition associated with increased oxidative stress and inflammation and is also associated with other CVD risk factors (arteriosclerosis, diabetes, hypertension, hyperlipidemia) that have been linked with accelerated telomere loss (Brouilette et al., 2008; Demissie et al., 2006; Ma, Zhu, Hu, Yu, & Yang, 2013). In contrast, lifestyle behaviors that decrease oxidative stress and inflammation will help to preserve TL (Cassidy et al., 2010). The health benefits of adequate physical activity and sleep are well established. Studies that assessed the relationship between these factors and LTL are reviewed (Tables 10 and 11 for physical activity and sleep, respectively).
TABLE 10.
Behavioral Factors: Physical Activity
| Authors | Date | Sample | Method | Findings |
|---|---|---|---|---|
| Ludlow et al. | 2008 | 34 men and 35 women 50–70 years |
qPCR | Moderate levels of physical activity (but not strenuous levels) significantly associated with longer LTL |
| Cherkas et al. | 2008 | 2,152 women and 249 men Healthy twin volunteers |
qPCR | LTLs of the most active participants 200 nucleotides longer than those of the least active subjects |
| Puterman et al. | 2010 | 63 healthy, postmenopausal women | qPCR | Among nonexercisers, a 1-unit increase in perceived stress related to a 15-fold increase in the odds of having short telomeres Among exercisers, perceived stress unrelated to TL |
| LaRocca et al. | 2010 | 15 sedentary young (18–32 years) 15 sedentary older (55–72 years) 10 endurance exercise-trained young 17 endurance exercise-trained older |
qPCR | LTL shorter in the older vs. young sedentary adults LTL of the older endurance-trained adults 900 base pairs greater than that of their sedentary peers and not significantly different from that of young exercise-trained adults |
| Savela et al. | 2012 | 204 men Helsinki Businessmen Study |
Southern blot | Longer LTL in men in the moderate physical activity group than in the low or high physical activity group |
| Denham et al. | 2013 | 67 ultra-marathon runners 56 healthy male controls |
qPCR | Significantly longer LTL in ultra-marathon runners compared with controls |
| Du et al. | 2012 | 7,813 women Nurses’ Health Study 43–70 years old |
qPCR | Greater moderate to vigorous intensity activity associated with increased LTL (especially calisthenics/aerobics) |
| Mathur et al. | 2013 | 17 marathon runners 15 sedentary controls Age- and sex-matched controls |
qPCR | Similar lymphocyte and granulocyte TL in athletes and sedentary controls |
Note. qPCR = quantitative polymerase chain reaction; LTL = leukocyte telomere length; TL = telomere length.
Physical Activity
Ludlow et al. (2008) evaluated the association between TL, telomerase activity, and level of physical activity using the Yale Physical Activity Survey in men and women aged 50–70 years. Their findings suggest that moderate levels of physical activity may protect PBMC TL compared with low or high exercise energy expenditure levels. Similarly, Cherkas et al. (2008) studied the association between physical activity during leisure time and LTL in 2,401 healthy twin volunteers. The LTLs of the most active participants were significantly longer compared with those of age- and sex-matched nonexercisers. Moderate exercise in midlife moderates the association between increased stress and shorter telomeres (Puterman et al., 2010) and is associated with longer telomeres in old age (Savela et al., 2013). Moderate to vigorous levels of physical activity or endurance exercise is associated with longer TL in most studies (Denham et al., 2013; Du et al., 2012; LaRocca, Seals, & Pierce, 2010), with one small study showing no difference when compared with nonexercisers (Mathur et al., 2013; Table 10).
Sleep Quality and Duration
Liang et al. (2011) reported a significant association between sleep duration and LTL in women under the age of 50 years, but not for those over 50 years, who were enrolled in the Nurses’ Health Study. Those with sleep duration of 6 hours or less had an equivalent to 9-year telomere attrition after adjustment for age, BMI, and cigarette smoking compared with women who had 9 hours or more of sleep per night. Although sleep duration and onset latency were unrelated to LTL in a community-dwelling sample of healthy women, poor sleep quality was associated with shorter LTL and predicted shorter LTL in participants who had chronic poor sleep quality (Prather et al., 2011). In contrast, a study among older adults found that shorter sleep duration was associated with significantly shorter telomeres in men only, independent of demographic, physical, and psychological covariates (Jackowska et al., 2012). On average, telomeres were 6% shorter in men sleeping 5 hours or less compared with those sleeping more than 7 hours per night (Table 11).
Discussion
In addition to individual factors that are known to affect LTL, including age, sex, race/ethnicity, paternal age at birth, and genetic mutations of telomerase and telomere maintenance genes, this integrative review identified several additional psychosocial, environmental, and behavioral factors that should be considered when integrating LTL into bio-behavioral research studies. The integrative review found evidence to support an association between shorter LTL and chronically high levels of perceived stress, the number of childhood adversities, duration of MDD, low educational attainment, low levels of physical activity, and short sleep duration. It should be noted that there were conflicting results identified for each of these factors. Explanations for the reason that conflicting conclusions have been observed by scientists regarding a purported association between psychosocial, environmental, and behavioral factors and LTL include, but are not limited to, heritable variation in LTL, study participant sampling, and methodological differences between investigations. Despite these limitations, the studies reviewed provide adequate justification for measuring perceived stress, childhood adversities, duration of MDD, educational attainment, physical activity, and sleep duration.
Consistent with the holistic lens of nursing, the integrative review found evidence to support the need for a multidimensional assessment of factors related to TL, which, in turn, has implications for biobehavioral research. Conceptual frameworks that incorporate the various factors related to LTL and plausible mechanisms underlying the changes are needed to guide future studies (Figure 3). A deeper understanding of the relationships between aspects of resiliency, coping, adaptation, behavioral change, and LTL would provide insight on the development of interventions to protect LTL. Carefully designed longitudinal studies are also needed to evaluate multidimensional nursing interventions and LTL, as well as potential underlying mechanisms.
FIGURE 3.
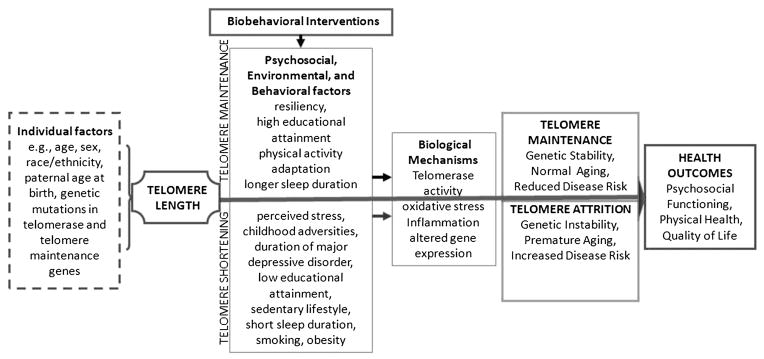
Model for the study of telomere length in biobehavioral research. This model is adapted from the general biobehavioral model developed out of the ongoing work in the Center for Biobehavioral Clinical Research at Virginia Commonwealth University School of Nursing (P20 NR008988, McCain; P30 NR011403, Grap). The model includes individual factors that influence telomere length, psychosocial, environmental and behavioral factors associated with telomere maintenance and attrition, and health outcomes.
As the science of telomere biology continues to expand, nurses need to be aware of the complexities in using LTL as a biomarker of health and disease risk. In addition to CVD and cancer, shorter TL has been associated with over 30 different metabolic and inflammatory diseases, and, at present, it remains unclear whether these relationships are bidirectional (Kong, Lee, & Wang, 2013). It was recently reported that a shorter mean LTL equivalent to one standard deviation in age-dependent length was associated with a 21% higher risk of CVD (Codd et al., 2013). Further research is needed to determine whether shorter telomeres play an etiological role in CVD or simply reflect an increased state of oxidative stress.
Alternatively, telomere shortening may directly contribute to alterations in the activity of genes that are adjacent to the proximal end of the telomere, allowing for changes in their expression that could lead to disease. These purported alterations in gene expression could be due to a more open chromatin conformation that occurs with shortening of the telomeric heterochromatic region with aging, also referred to as a telomere position effect (Mondoux & Zakian, 2005; Zakian, 2012). Indeed, this scenario has been observed in humans by experimental manipulation of TLs (Baur, Zou, Shay, & Wright, 2001), as well as in spontaneously occurring conditions, such as facioscapulohumeral muscular dystrophy, in which a deletion of telomeric sequences on chromosome 4 results in the expression of genes that are normally silenced (van Deutekom et al., 1996). A contrasting explanation for the observed relationship between TL and aging/disease risk is that this association may be simply correlative in nature, with TL serving as a “chronicle” (i.e., the “canary in the coal mine”) to document the occurrence of biologically relevant changes that could lead to an increased health risk (Effros, 2009).
Conclusions
There is now a robust body of research that has identified an association between LTL and various psychosocial, environmental, and biological factors. Although the evidence of a direct effect on TL is still lacking, the results of the integrative review suggest that perceived stress, childhood adversities, MDD, educational attainment, physical activity, and sleep duration should also be measured. A formidable challenge for this area of research will be to elucidate the exact mechanism(s) that underlie the associations with TL.
Acknowledgments
This research was supported by a grant to Dr. Lyon (NIH/NINR P30 NR011403; R01 NR012667). Dr. Montpetit (NIH/NINR K99/ R00NR012016) and Dr. Jackson-Cook (NIH/NINR R01 NR012667; NIH/NIA R01AG037986) are currently receiving grants.
Footnotes
The content of this publication is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Nursing Research, National Institute on Aging, or the National Institutes of Health.
The authors have no conflicts of interest to disclose.
Contributor Information
Angela R. Starkweather, Associate Professor, School of Nursing, Virginia Commonwealth University, Richmond.
Areej A. Alhaeeri, Graduate Student, School of Medicine, Virginia Commonwealth University, Richmond.
Alison Montpetit, Assistant Professor, School of Nursing, Virginia Commonwealth University, Richmond.
Jenni Brumelle, Postdoctoral Fellow, School of Medicine, Virginia Commonwealth University, Richmond.
Kristin Filler, Doctoral Student Fellow, School of Nursing, Virginia Commonwealth University, Richmond.
Marty Montpetit, Assistant Professor, School of Nursing, Virginia Commonwealth University, Richmond.
Lathika Mohanraj, Postdoctoral Fellow, School of Nursing, Virginia Commonwealth University, Richmond.
Debra E. Lyon, Associate Dean for Research, School of Nursing, Virginia Commonwealth University, Richmond.
Colleen K. Jackson-Cook, Professor, School of Medicine, Virginia Commonwealth University, Richmond.
References
- Adams J, Martin-Ruiz C, Pearce MS, White M, Parker L, Von Zglinicki T. No association between socioeconomic status and white blood cell telomere length. Aging Cell. 2007;6:125–128. doi: 10.1111/j.1474-9726.2006.00258.x. [DOI] [PubMed] [Google Scholar]
- Adler N, Pantell MS, O’Donovan A, Blackburn E, Cawthon R, Koster A, Epel E. Educational attainment and late life telomere length in the Health, Aging, and Body Composition Study. Brain, Behavior, and Immunity. 2013;27:15–21. doi: 10.1016/j.bbi.2012.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aubert G, Hills M, Lansdorp PM. Telomere length measurement—Caveats and a critical assessment of the available technologies and tools. Mutation Research. 2012;730:59–67. doi: 10.1016/j.mrfmmm.2011.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aubert G, Lansdorp PM. Telomeres and aging. Physiological Reviews. 2008;88:557–579. doi: 10.1152/physrev.00026.2007. [DOI] [PubMed] [Google Scholar]
- Atzmon G, Hao L, Pe’er I, Velez C, Pearlman A, Palamara PF, Ostrer H. Abraham’s children in the genome era: Major Jewish diaspora populations comprise distinct genetic clusters with shared Middle Eastern ancestry. American Journal of Human Genetics. 2010;86:850–859. doi: 10.1016/j.ajhg.2010.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ballew BJ, Savage SA. Updates on the biology and management of dyskeratosis congenita and related telomere biology disorders. Expert Review in Hematology. 2013;6:327–337. doi: 10.1586/ehm.13.23. [DOI] [PubMed] [Google Scholar]
- Batty GD, Wang Y, Brouilette SW, Shiels P, Packard C, Moore J, Ford I. Socioeconomic status and telomere length: The West of Scotland Coronary Prevention Study. Journal of Epidemiology & Community Health. 2009;63:839–841. doi: 10.1136/jech.2009.088427. [DOI] [PubMed] [Google Scholar]
- Baur JA, Zou Y, Shay JW, Wright WE. Telomere position effect in human cells. Science. 2001;292:2075–2077. doi: 10.1126/science.1062329. [DOI] [PubMed] [Google Scholar]
- Bekaert S, De Meyer T, Rietzschel ER, De Buyzere ML, De Bacqquer D, Langlois M Asklepios Investigators. Telomere length and cardiovascular disease risk factors in a middle-aged population free of overt cardiovascular disease. Aging Cell. 2007;6:639–647. doi: 10.1111/j.1474-9726.2007.00321.x. [DOI] [PubMed] [Google Scholar]
- Blackburn EH, Greider CW, Szostak JW. Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nature Medicine. 2006;12:1133–1138. doi: 10.1038/nm1006-1133. [DOI] [PubMed] [Google Scholar]
- Broer L, Codd V, Nyholt DR, Deelen J, Mangino M, Willemsen G, Boomsma DI. Meta-analysis of telomere length in 19,713 subjects reveals high heritability, stronger maternal inheritance and a paternal age effects. European Journal of Human Genetics. 2013;21:1163–1168. doi: 10.1038/ejhg.2012.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brouilette SW, Whitaker A, Stevens SE, van der Harst P, Goodall AH, Samani NJ. Telomere length is shorter in healthy offspring of subjects with coronary artery disease: Support for the telomere hypothesis. Heart. 2008;94:422–425. doi: 10.1136/hrt.2007.139675. [DOI] [PubMed] [Google Scholar]
- Canela A, Vera E, Klatt P, Blasco MA. High-throughput telomere length quantification by FISH and its application to human population studies. Proceedings of the National Academy of Sciences. 2007;104:5300–5305. doi: 10.1073/pnas.0609367104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carroll JE, Diez-Roux AV, Adler NE, Seeman TE. Socioeconomic factors and leukocyte telomere length in a multi-ethnic sample: Findings from the Multi-ethnic Study of Atherosclerosis (MESA) Brain, Behavior and Immunity. 2013;28:108–114. doi: 10.1016/j.bbi.2012.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carroll JE, Diez-Roux AV, Fitzpatrick AL, Seeman T. Low social support is associated with shorter leukocyte telomere length in late life: Multi-ethnic study of atherosclerosis. Psychosomatic Medicine. 2013;75:171–177. doi: 10.1097/PSY.0b013e31828233bf. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cassidy A, De Vivo I, Liu Y, Han J, Prescott J, Hunter DJ, Rimm EB. Associations between diet, lifestyle factors, and telomere length in women. American Journal of Clinical Nutrition. 2010;91:1273–1280. doi: 10.3945/ajcn.2009.28947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cesare AJ, Reddel RR. Alternative lengthening of telomeres: Models, mechanisms and implications. Nature Reviews Genetics. 2010;11:319–330. doi: 10.1038/nrg2763. [DOI] [PubMed] [Google Scholar]
- Chen M, McLeskey SW. Telomere-based cancer treatment: Emerging targeted therapies. Clinical Journal of Oncology Nursing. 2010;14:720–726. doi: 10.1188/10.CJON.720-726. [DOI] [PubMed] [Google Scholar]
- Cherkas LF, Aviv A, Valdes AM, Hunkin JL, Gardner JP, Surdulescu GL, Spector TD. The effects of social status on biological aging as measured by white-blood-cell telomere length. Aging Cell. 2006;5:361–365. doi: 10.1111/j.1474-9726.2006.00222.x. [DOI] [PubMed] [Google Scholar]
- Cherkas LF, Hunkin JL, Kato BS, Richards JB, Gardner JP, Surdulescu GL, Aviv A. The association between physical activity in leisure time and leukocyte telomere length. Archives of Internal Medicine. 2008;168:154–158. doi: 10.1001/archinternmed.2007.39. [DOI] [PubMed] [Google Scholar]
- Codd V, Nelson CP, Albrecht E, Mangino M, Deelen J, Buxton JL, Samani NJ. Identification of seven loci affecting mean telomere length and their association with disease. Nature Genetics. 2013;45:422–427. doi: 10.1038/ng.2528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Damjanovic AK, Yang Y, Glaser R, Kiecolt-Glaser JK, Nguyen H, Laskowski B, Weng NP. Accelerated telomere erosion is associated with a declining immune function of caregivers of Alzheimer’s disease patients. Journal of Immunology. 2007;179:4249–4254. doi: 10.4049/jimmunol.179.6.4249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demissie S, Levy D, Benjamin EJ, Cupples LA, Gardner JP, Herbert A, Aviv A. Insulin resistance, oxidative stress, hypertension, and leukocyte telomere length in men from the Framingham Heart Study. Aging Cell. 2006;5:325–330. doi: 10.1111/j.1474-9726.2006.00224.x. [DOI] [PubMed] [Google Scholar]
- Denham J, Nelson CP, O’Brien BJ, Nankervis SA, Denniff M, Harvey JT, Charchar FJ. Longer leukocyte telomeres are associated with ultra-endurance exercise independent of cardiovascular risk factors. PLoS One. 2013;8:e69377. doi: 10.1371/journal.pone.0069377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du M, Prescott J, Kraft P, Han J, Giovannucci E, Hankinson SE, De Vivo I. Physical activity, sedentary behavior, and leukocyte telomere length in women. American Journal of Epidemiology. 2012;175:414–422. doi: 10.1093/aje/kwr330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Effros RB. Kleemeier Award Lecture 2008—The canary in the coal mine: Telomeres and human healthspan. Journals of Gerontology Series A: Biological Sciences and Medical Sciences. 2009;64:511–515. doi: 10.1093/gerona/glp001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Entringer S, Epel ES, Kumsta R, Lin J, Hellhammer DH, Blackburn EH, Wust S, Wadhwa PD. Stress exposure in intrauterine life is associated with shorter telomere length in young adulthood. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:E513–E518. doi: 10.1073/pnas.1107759108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Entringer S, Epel ES, Lin J, Buss C, Shahbaba B, Blackburn EH, Simhan HN, Wadhwa PD. Maternal psychological stress during pregnancy is associated with newborn leukocyte telomere length. American Journal of Obstetrics and Gynecology. 2013;208:134.e1–134.e7. doi: 10.1016/j.ajog.2012.11.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epel ES, Blackburn EH, Lin J, Dhabhar FS, Adler NE, Morrow JD, Cawthon RM. Accelerated telomere shortening in response to life stress. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:17312–17315. doi: 10.1073/pnas.0407162101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epel ES, Lin J, Dhabhar FS, Wolkowitz OM, Puterman E, Karan L, Blackburn EH. Dynamics of telomerase activity in response to acute psychological stress. Brain, Behavior, and Immunity. 2010;24:531–539. doi: 10.1016/j.bbi.2009.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epel ES, Lin J, Wilhelm FH, Wolkowitz OM, Cawthon R, Adler NE, Blackburn EH. Cell aging in relation to stress arousal and cardiovascular disease risk factors. Psychoneuroendocrinology. 2006;31:277–287. doi: 10.1016/j.psyneuen.2005.08.011. [DOI] [PubMed] [Google Scholar]
- Garagnani P, Giuliani C, Pirazzini C, Olivieri F, Bacalini MG, Ostan R, Franceschi C. Centenarians, as super-controls to assess the biological relevance of genetic risk factors for common age-related diseases: A proof of principle on type 2 diabetes. Aging. 2013;5:373–385. doi: 10.18632/aging.100562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glass D, Parts L, Knowles D, Aviv A, Spector TD. No correlation between childhood maltreatment and telomere length. Biological Psychiatry. 2010;68:e21–e22. doi: 10.1016/j.biopsych.2010.02.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartmann N, Boehner M, Groenen F, Kalb R. Telomere length of patients with major depression is shortened but independent from therapy and severity of disease. Depression and Anxiety. 2010;27:1111–1116. doi: 10.1002/da.20749. [DOI] [PubMed] [Google Scholar]
- Hoen PW, de Jonge P, Na BY, Farzaneh-Far R, Epel E, Lin J, Blackburn E, Whooley MA. Depression and leukocyte telomere length in patients with coronary heart disease: Data from the Heart and Soul Study. Psychosomatic Medicine. 2011;73:541–547. doi: 10.1097/PSY.0b013e31821b1f6e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphreys J, Epel ES, Cooper BA, Lin J, Blackburn EH, Lee KA. Telomere shortening in formerly abused and never abused women. Biological Research for Nursing. 2012;14:115–123. doi: 10.1177/1099800411398479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackowska M, Hamer M, Carvalho LA, Erusalimsky JD, Butcher L, Steptoe A. Short sleep duration is associated with shorter telomere length in healthy men: Findings from the Whitehall II Cohort Study. PLoS One. 2012;7:e47292. doi: 10.1371/journal.pone.0047292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kananen L, Surakka I, Pirkola S, Suvisaari J, Lönnqvist J, Peltonen L, Hovatta I. Childhood adversities are associated with shorter telomere length at adult age both in individuals with an anxiety disorder and controls. PLoS One. 2010;5:e10826. doi: 10.1371/journal.pone.0010826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiecolt-Glaser JK, Gouin J-P, Weng N-P, Malarkey WB, Beversdorf DQ, Glaser R. Childhood adversity heightens the impact of later-life caregiving stress on telomere length and inflammation. Psychosomatic Medicine. 2011;73:16–22. doi: 10.1097/PSY.0b013e31820573b6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong CM, Lee XW, Wang X. Telomere shortening in human diseases. The FEBS Journal. 2013;280:3180–3193. doi: 10.1111/febs.12326. [DOI] [PubMed] [Google Scholar]
- LaRocca TJ, Seals DR, Pierce GL. Leukocyte telomere length is preserved with aging in endurance exercise-trained adults and related to maximal aerobic capacity. Mechanisms of Ageing and Development. 2010;131:165–167. doi: 10.1016/j.mad.2009.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang G, Schernhammer E, Qi L, Gao X, De Vivo I, Han J. Associations between rotating night shifts, sleep duration, and telomere length in women. PLoS One. 2011;6:e23462. doi: 10.1371/journal.pone.0023462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludlow AT, Zimmerman JB, Witkowski S, Hearn JW, Hatfield BD, Roth SM. Relationship between physical activity level, telomere length, and telomerase activity. Medicine & Science in Sports & Exercise. 2008;40:1764–1771. doi: 10.1249/MSS.0b013e31817c92aa. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lung F-W, Chen NC, Shu B-C. Genetic pathway of major depressive disorder in shortening telomeric length. Psychiatric Genetics. 2007;17:195–199. doi: 10.1097/YPG.0b013e32808374f6. [DOI] [PubMed] [Google Scholar]
- Ma D, Zhu W, Hu S, Yu X, Yang Y. Association between oxidative stress and telomere length in type 1 and type 2 diabetic patients. Journal of Endocrinological Investigation. 2013 doi: 10.3275/9036. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- Ma H, Zhou Z, Wei S, Liu Z, Pooley KA, Dunning AM, Wei Q. Shortened telomere length is associated with increased risk of cancer: A meta-analysis. PLoS ONE. 2011;6:e20466. doi: 10.1371/journal.pone.0020466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malan S, Hemmings S, Kidd M, Martin L, Seedat S. Investigation of telomere length and psychological stress in rape victims. Depression and Anxiety. 2011;28:1081–1085. doi: 10.1002/da.20903. [DOI] [PubMed] [Google Scholar]
- Mathur S, Ardestani A, Parker B, Cappizzi J, Polk D, Thompson PD. Telomere length and cardiorespiratory fitness in marathon runners. Journal of Investigative Medicine. 2013;61:613–615. doi: 10.2310/JIM.0b013e3182814cc2. [DOI] [PubMed] [Google Scholar]
- Mondoux MA, Zakian VA. Telomere position effect: Silencing near the end. In: de Lange T, Lundblad V, Blackburn E, editors. Telomeres. 2. Cold Spring Harbour, NY: Cold Spring Harbor Laboratory Press; 2005. pp. 261–316.pp. 261–316. [Google Scholar]
- Needham BL, Adler N, Gregorich S, Rehkopf D, Lin J, Blackburn EH, Epel ES. Socioeconomic status, health behavior, and leukocyte telomere length in the National Health and Nutrition Examination Survey, 1999–2002. Social Science & Medicine. 2013;85:1–8. doi: 10.1016/j.socscimed.2013.02.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Needham BL, Fernandez JR, Lin J, Epel ES, Blackburn EH. Socioeconomic status and cell aging in children. Social Science & Medicine. 2012;74:1948–1951. doi: 10.1016/j.socscimed.2012.02.019. [DOI] [PubMed] [Google Scholar]
- O’Donovan A, Lin J, Dhabhar FS, Wolkowitz O, Tillie JM, Blackburn E, Epel E. Pessimism correlates with leukocyte telomere shortness and elevated interleukin-6 in post-menopausal women. Brain, Behavior, and Immunity. 2009;23:446–449. doi: 10.1016/j.bbi.2008.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parks CG, Miller DB, McCanlies EC, Cawthon RM, Andrew ME, DeRoo LA, Sandler DP. Telomere length, current perceived stress, and urinary stress hormones in women. Cancer Epidemiology, Biomarkers & Prevention. 2009;18:551–560. doi: 10.1158/1055-9965.EPI-08-0614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prather AA, Puterman E, Lin J, O’Donovan A, Krauss J, Tomiyama AJ, Blackburn EH. Shorter leukocyte telomere length in midlife women with poor sleep quality. Journal of Aging Research. 2011;2011:1–6. doi: 10.4061/2011/721390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puterman E, Epel ES, Lin J, Blackburn EH, Gross JJ, Whooley MA, Cohen BE. Multisystem resiliency moderates the major depression-telomere length association: Findings from the Heart and Soul Study. Brain, Behavior, and Immunity. 2013 doi: 10.1016/j.bbi.2013.05.008. Epub ahead of print. Retrieved from http://www.chc.ucsf.edu/AME_lab/pdfs/PUterman_etal_2013.pdf. [DOI] [PMC free article] [PubMed]
- Puterman E, Lin J, Blackburn E, O’Donovan A, Adler N, Epel E. The power of exercise: Buffering the effect of chronic stress on telomere length. PLoS One. 2010;5:e10837. doi: 10.1371/journal.pone.0010837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sahin E, DePinho RA. Linking functional decline of telomeres, mitochondria and stem cells during ageing. Nature. 2010;464:520–528. doi: 10.1038/nature08982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savela S, Saijonmaa O, Strandberg TE, Koistinen P, Strandberg AY, Tilvis RS, Fyhrguist F. Physical activity in midlife and telomere length measured in old age. Experimental Gerontology. 2013;48:81–84. doi: 10.1016/j.exger.2012.02.003. [DOI] [PubMed] [Google Scholar]
- Schaefer C, Sciortino S, Kvale M, Lapham K, Lin J, Ranatunga D, Blackburn E. The Kaiser Permanente/UCSF Genetic Epidemiology Research Study on Adult Health and Aging: Demographic and behavioral influences on telomeres and relationship with all-cause mortality. Paper presented at The American Society of Human Genetics Annual Meeting; San Francisco, CA. 2012. Nov, [Google Scholar]
- Shalev I, Moffitt TE, Sugden K, Williams B, Houts RM, Danese A, Caspi A. Exposure to violence during childhood is associated with telomere erosion from 5 to 10 years of age: A longitudinal study. Molecular Psychiatry. 2013;18:576–581. doi: 10.1038/mp.2012.32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen J, Gammon MD, Terry MB, Wang Q, Bradshaw P, Teitelbaum SL, Santella RM. Telomere length, oxidative damage, antioxidants and breast cancer risk. International Journal of Cancer. 2009;124:1637–1643. doi: 10.1002/ijc.24105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon NM, Smoller JW, McNamara KL, Maser RS, Zalta AK, Pollack MH, Wong K-K. Telomere shortening and mood disorders: Preliminary support for a chronic stress model of accelerated aging. Biological Psychiatry. 2006;60:432–435. doi: 10.1016/j.biopsych.2006.02.004. [DOI] [PubMed] [Google Scholar]
- Steptoe A, Hamer M, Butcher L, Lin J, Brydon L, Kivimäki M, Erusalimsky JD. Educational attainment but not measures of current socioeconomic circumstances are associated with leukocyte telomere length in healthy older men and women. Brain, Behavior, and Immunity. 2011;25:1292–1298. doi: 10.1016/j.bbi.2011.04.010. [DOI] [PubMed] [Google Scholar]
- Surtbees PG, Wainwright NWJ, Pooley KA, Luben RN, Khaw K-T, Easton DF, Dunning AM. Life stress, emotional health, and mean telomere length in the European Prospective Investigation Into Cancer (EPIC)–Norfolk population study. Journals of Gerontology Series A: Biological Sciences and Medical Sciences. 2011;66:1152–1162. doi: 10.1093/gerona/glr112. [DOI] [PubMed] [Google Scholar]
- Surtees PG, Wainwright NWJ, Pooley KA, Luben RN, Khaw KT, Easton DF, Dunning AM. Educational attainment and mean leukocyte telomere length in women in the European Prospective Investigation into Cancer (EPIC)-Norfolk population study. Brain, Behavior, and Immunity. 2012;26:414–418. doi: 10.1016/j.bbi.2011.11.009. [DOI] [PubMed] [Google Scholar]
- Theall KP, Brett ZH, Shirtcliff EA, Dunn EC, Drury SS. Neighborhood disorder and telomeres: Connecting children’s exposure to community level stress and cellular response. Social Science & Medicine. 2013;85:50–58. doi: 10.1016/j.socscimed.2013.02.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tyrka AR, Price LH, Kao H-T, Porton B, Marsella SA, Carpenter LL. Childhood maltreatment and telomere shortening: Preliminary support for an effect of early stress on cellular aging. Biological Psychiatry. 2010;67:531–534. doi: 10.1016/j.biopsych.2009.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valdes AM, Andrew T, Gardner JP, Kimura M, Oelsner E, Cherkas LF, Spector TD. Obesity, cigarette smoking, and telomere length in women. The Lancet. 2005;366:662–664. doi: 10.1016/S0140-6736(05)66630-5. [DOI] [PubMed] [Google Scholar]
- van Deutekom JCT, Bakker E, Lemmers RJLF, van der Wielen MJR, Bik E, Hofker MH, Frants RR. Evidence for subtelomeric exchange of 3.3 kb tandemly repeated units between chromosomes 4q35 and 10q26: Implications for genetic counselling and etiology of FSHD1. Human Molecular Genetics. 1996;5:1997–2003. doi: 10.1093/hmg/5.12.1997. [DOI] [PubMed] [Google Scholar]
- Wikgren M, Maripuu M, Karlsson T, Nordfjäll K, Bergdahl J, Hultdin J, Norrback K-F. Short telomeres in depression and the general population are associated with a hypocortisolemic state. Biological Psychiatry. 2012;71:294–300. doi: 10.1016/j.biopsych.2011.09.015. [DOI] [PubMed] [Google Scholar]
- Wolkowitz OM, Mellon SH, Epel ES, Lin J, Dhabar FS, Su Y, Blackburn EH. Leukocyte telomere length in major depression: Correlations with chronicity, inflammation and oxidative stress—preliminary findings. PLoS One. 2011;6:e17837. doi: 10.1371/journal.pone.0017837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolkowitz OM, Mellon SH, Epel ES, Lin J, Reus VI, Rosser R, Blackburn EH. Resting leukocyte telomerase activity is elevated in major depression and predicts treatment response. Molecular Psychiatry. 2012;17:164–172. doi: 10.1038/mp.2010.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo J, Suen EWC, Leung JCS, Tang NLS, Ebrahim S. Older men with higher self-rated socioeconomic status have shorter telomeres. Age and Ageing. 2009;38:553–558. doi: 10.1093/ageing/afp098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo J, Tang NLS, Suen E, Leung JCS, Wong M. Green space, psychological restoration, and telomere length. The Lancet. 2009;373:299–300. doi: 10.1016/S0140-6736(09)60094-5. [DOI] [PubMed] [Google Scholar]
- Zakian VA. Telomeres: The beginnings and ends of eukaryotic chromosomes. Experimental Cell Research. 2012;318:1456–1460. doi: 10.1016/j.yexcr.2012.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zanni GR, Wick JY. Telomeres: Unlocking the mystery of cell division and aging. Consultant Pharmacist. 2011;26:78–90. doi: 10.4140/TCP.n.2011.78. [DOI] [PubMed] [Google Scholar]