

Abstract
The American oyster Crassostrea virginica, an ecologically and economically important estuarine organism, can suffer high mortalities in areas in the Northeast United States due to Roseovarius Oyster Disease (ROD), caused by the gram-negative bacterial pathogen Roseovarius crassostreae. The goals of this research were to provide insights into: 1) the responses of American oysters to R. crassostreae, and 2) potential mechanisms of resistance or susceptibility to ROD. The responses of oysters to bacterial challenge were characterized by exposing oysters from ROD-resistant and susceptible families to R. crassostreae, followed by high-throughput sequencing of cDNA samples from various timepoints after disease challenge. Sequence data was assembled into a reference transcriptome and analyzed through differential gene expression and functional enrichment to uncover genes and processes potentially involved in responses to ROD in the American oyster. While susceptible oysters experienced constant levels of mortality when challenged with R. crassostreae, resistant oysters showed levels of mortality similar to non-challenged oysters. Oysters exposed to R. crassostreae showed differential expression of transcripts involved in immune recognition, signaling, protease inhibition, detoxification, and apoptosis. Transcripts involved in metabolism were enriched in susceptible oysters, suggesting that bacterial infection places a large metabolic demand on these oysters. Transcripts differentially expressed in resistant oysters in response to infection included the immune modulators IL-17 and arginase, as well as several genes involved in extracellular matrix remodeling. The identification of potential genes and processes responsible for defense against R. crassostreae in the American oyster provides insights into potential mechanisms of disease resistance.
Introduction
The American or eastern oyster Crassostrea virginica is an estuarine molluscan bivalve species fished and cultured from Texas, USA to New Brunswick, Canada. Oyster production is an important sector of United States agriculture and the American oyster was estimated in 2012 to have a farm gate value of $104 million in the United States [1]. Ecologically, the American oyster provides biogenic habitat and filters large quantities of plankton, having a great impact on the coastal ecosystems it inhabits [2], [3]. Several oyster diseases, both protozoan and bacterial, have expanded in range and increased in severity during the latter half of the twentieth century, often causing staggering losses [4]. Juvenile or Roseovarius Oyster Disease (ROD), an emerging disease caused by the gram-negative bacterium Roseovarius crassostreae, was first reported in 1988 and presently affects oysters from the Long Island Sound north to Maine [5], [6]. As high as 90–100% of oyster juveniles in a farm may succumb to this disease during mortality events that often coincide with peak summer water temperatures. Gross clinical signs include uneven shell margins, soft tissue emaciation, and conchiolin depositions (a mix of shell material and organic molecules) on the inner shell surfaces [5]–[7].
The host-pathogen interactions between C. virginica and R. crassostreae are poorly understood. This extracellular bacterial pathogen colonizes the oyster’s inner shell surface before lesions develop in the epithelial mantle. Colonization of the inner side of the oyster shell by R. crassostreae likely stimulates oysters to deposit conchiolin [8]. It has been hypothesized that smaller juvenile oysters (<25 mm in shell length) are most susceptible to ROD because they lack adequate metabolic resources to fuel immune responses, including conchiolin deposition, leading to emaciation [5], [6], [8]. Roseovarius crassostreae may produce a toxin with ciliostatic activity [9] and extracellular products from R. crassostreae have a cytotoxic effect on oyster hemocytes that cannot be solely attributed to lipopolysaccharide (LPS), a component of the membrane of gram-negative bacteria [10].
Traditional selective breeding practices have led to the production of ROD-resistant oysters [11], [12], but the genetic basis of resistance is presently unknown. The identification of potential genes and pathways responsible for an effective host defense response in the American oyster to R. crassostreae is important not only to provide a basis for enhanced breeding techniques [13]–[15], but also advances the understanding of immunity in a member of Lophotrochozoa, a superphylum that has been poorly represented among genomic and transcriptomic datasets until recently e.g. [16]–[19]. Invertebrate hosts lack the classical adaptive immune system, yet they successfully combat widely varied types of microbes and parasites. To mount effective and flexible defense responses to diverse pathogens, invertebrate hosts have developed diversified repertoires of receptors, regulators, and/or effectors including Toll-like receptors (TLRs), fibrinogen-related proteins (FREPs), scavenger receptor cysteine-rich (SRCRs), and antimicrobial proteins, as well as many other molecules involved in the key processes of agglutination, phagocytosis, and encapsulation [20]–[22].
In order to identify genes and processes potentially involved in: 1) the responses of American oysters to challenge with the bacterial pathogen R. crassostreae, and 2) potential mechanisms of resistance or susceptibility to ROD, cDNA sequences from ROD-resistant and susceptible families of oysters exposed to the bacterial pathogen Roseovarius crassostreae were assembled into a reference transcriptome. A targeted differential gene expression analysis, followed by evaluation of functional categories enriched among differentially expressed genes, were used to identify genes and processes involved in the response of oysters exposured to R. crassostreae. This targeted analysis was also used to identify a list of genes and molecular processes potentially involved in resistance/susceptibility to ROD.
Materials and Methods
Bacterial challenge of American oysters
Juvenile American oysters from 2 families with known differential susceptibility to ROD (F3L and GX09) were kindly provided by X. Guo (Haskin Shellfish Research Laboratory, Rutgers University). Susceptible F3L oysters were F3 generation progeny from a single pair mating of a female oyster from the Rutgers NEH (Northeastern High-survival) line [23] and a male oyster from Louisiana (LA). Resistant GX09 (GX) oysters were an F3 generation containing germline material from the NEH, DBH (Delaware Bay High-survival line), LA, and the ROD-resistant lines UMFS (University of Maine Flowers Select) and FMF (Frank M. Flowers) lines [11], [12]. Oysters (10–15 mm in shell length) were labeled on the outside of the shell using non-toxic paint (to distinguish each family) and placed into two replicate 250 l tanks with filtered sterile seawater (FSSW) for bacterial challenge (experimental groups GX and F3L, about 120 oysters per family per tank). Additional groups of 50 (F3L) and 2×50 (GX) oysters were kept in 50 l tanks as unchallenged controls (CGX and CF3L). Oysters were acclimated during a period of 2 weeks to experimental conditions (salinity 28–30‰, temperature 19°C). Oysters in the challenge tanks were exposed to R. crassostreae, strain CV919-312T [7] by addition of bacteria to the tank at a final concentration of 7.5×106 colony forming units (CFU) ml−1 (day 0 of challenge). Oysters were fed Instant Algae (Reed Mariculture) every other day and water was partially changed (50%) weekly. Oysters were monitored weekly for 93 days for mortalities and for the presence of clinical signs of ROD (uneven valves and conchiolin deposits in shells of dead oysters). Infection by R. crassostreae was confirmed by PCR [24].
Sample collection, cDNA preparation, and sequencing
Oyster whole body tissue was collected from 15 randomly sampled oysters each from CGX, CF3L (unchallenged controls), GX, and F3L (challenged resistant and susceptible oysters) at days 1, 5, 15, and 30 following challenge and stored in RNAlater until time of RNA isolation. All RNA molecules >200 nucleotides were purified using Qiagen RNAeasy Mini Kit. Samples were checked for RNA purity using a Nanodrop 8000 spectrophotometer and a subset of the extracts were checked using an Agilent 2100 Bioanalyzer. Due to limitations in funding, control non-challenged susceptible (CF3L) oysters were not included in the sequencing analysis. Equal amounts of total RNA from 5 oysters from each treatment and time point (excluding C3FL) were pooled for a total of 12 experimental samples (3 treatments×4 time points). Samples of RNA were selectively enriched for poly-A containing mRNA and cDNA libraries for sequencing were prepared using the Illumina mRNA-Seq-8 Sample Prep Kit. The cDNA libraries were sequenced on the Illumina GAIIx platform (1 lane per sample for a total of 12 lanes, Genome Quebec, Canada).
Read processing and de novo assembly
Raw sequencing reads of 108 bp from all lanes (SRP042090) were pooled, processed, and filtered for contamination of mitochondrial and ribosomal sequences by mapping to all Crassostrea spp. rRNA and mtDNA in NCBI Genbank database. Reads were filtered for vector sequences by mapping to Univec (ftp://ftp.ncbi.nih.gov/pub/univec) using bowtie2 [25]. Low-complexity artifacts were removed, and Illumina adapters and the 5′-ends of reads were trimmed using the fastx-toolkit (http://hannonlab.cshl.edu/fastx_toolkit/). Adaptor trimming was performed on reads using the btrim software package [26]. Reads less than 20 bp in length were discarded. Processed transcriptome reads from the 12 lanes were assembled into a reference transcriptome using Trinity (release 20111126) with default options [27]. Only those assembled contigs ≥200 bp were retained. Transcriptome contigs were compared to the RefSeq protein database [28]. A custom python script, created by L. Dong (Brown University), was used to parse BLAST output and identify possible contaminants. Contigs that had all top blast hits (a maximum of 10) with associated e-value≤1e-06 to proteins from Archaebacteria, Bacteria, or Protozoa were discarded. Additional mitochondrial and ribosomal contaminants were identified and discarded through text searching of BLAST results. Phage integrase sequences were identified and discarded by comparing the transcripts to Pfam_A using Pfam scan (version 1.3) and HMMER (version 3.0) with hits retained where e-value≤1e-05 [29], [30]. DNA transposons were identified and discarded using RepeatMasker [31].
Differential Gene Expression
Reads from individual treatment-days samples (e.g. GX-1d) were aligned to the reference transcriptome using bowtie [32] with parameters “-v 3–a –best –strata,” such that 3 mismatches were allowed per read to account for the high rate of polymorphism in oysters [33]. Transcript abundances in reads per kilobase per million reads mapped (RPKM) were estimated using RSEM (RNA-Seq by Expectation Maximization) through the Trinity plug-in, run_RSEM.pl. [27], [34], [35]. To reduce bias from differential sequencing depth across lanes, the trimmed mean of M values (TMM) method was used to calculate normalization factors for each lane [36]. Only those contigs with at least 1 count-per-million in at least 2 samples were tested for differential expression.
In order to identify general patterns of variation driving differences between treatment groups, two analyses were performed in the R programming environment [37] using the Z-score centered log2-transformed RPKM for each transcript in each of the treatment groups: a) a principal components analysis (PCA); and b) a heatmap analysis. For the heatmap analysis, transcripts were hierarchically clustered (Euclidean distance, complete linkage) using the fastcluster [38] package. Results were visualized using gplots [39]. Based on the results from the mortality curves and the principal components and heatmap analyses, differential gene expression analysis was performed by comparing read abundances for contigs in each of the samples to read abundances in a control pool (CGX 15 and 30 d, see results for rationale) using edgeR [40]. Significance values yielded by hypergeometric test were adjusted using the False Discovery Rate (FDR) correction and a contig was considered differentially expressed (DE) if it had an FDR-adjusted p-value≤0.05 [41].
Annotation and Functional Enrichment
Transcriptome contigs were compared to the NCBI protein non-redundant (NR) database using BLASTx [42]. Hits with e-value≤1e-6 were retained. Gene Ontology (GO) terms were mapped to the best BLASTx hits for each contig using the Blast2GO pipeline (version 2.3.5) [43]. In order to identify which functional categories were enriched within differentially expressed transcripts for the selected treatment groups, functional enrichment was performed using the R package topGO by comparing the numbers of GO terms associated with annotations of differentially expressed transcripts within each selected treatment to the numbers of terms associated with all transcripts not differentially expressed [44]. Fisher’s exact test was used to determine significance of enrichment of each GO term, with Bonferroni-adjusted p-values≤0.05 taken as significant. Functionally enriched GO terms were visualized in semantic space using SimRel functional similarity measure [45] and the REViGO online visualization tool [46] modified with the R package ggplot2 [47] (scripts available upon request).
Results
Oyster survival in response to bacterial challenge
Oysters from the F3L family (susceptible to ROD) experienced a constant rate of mortality of about 7% per week after challenge with the bacterial pathogen R. crassostreae, reaching over 90% cumulative mortality by the end of the 93-day challenge period (Figure 1). The survival curve of the challenged susceptible F3L oysters was significantly different from all other groups (log-rank survival, p<0.01). At day 28, F3L had a significantly higher cumulative mortality than C3FL and GX (resistant to ROD), but not CGX control (p<0.01, Pearson’s chi-squared test with Bonferroni corrections). At day 93, F3L had a significantly higher cumulative mortality than GX, CGX, and C3FL (p<0.01). No significant differences in mortality were observed between unchallenged control oysters (CF3L and CGX) and oysters from the resistant challenged family GX at day 28 and day 93 after challenge. Challenged resistant oysters did not show any of the clinical signs of ROD, suggesting that the pathogen is eliminated rapidly and does not cause an active infection in these oysters. Oysters from the control resistant family CGX suffered a mortality event of unresolved origin between days 1 and 7 (20% cumulative percent mortality by day 7; Figure 1). Due to the potential confounding effect of this mortality event on gene expression at early time points after challenge, samples from CGX at days 1 and 5 were not included in the gene expression analysis.
Figure 1. Mortality in resistant and susceptible oysters after challenge with Roseovarius crassostreae.

Cumulative percent mortality in resistant GX and susceptible F3L oysters following bacterial challenge compared to mortality in non-challenged (CGX, CF3L) oysters. Arrows on the x-axis indicate the timepoints at which RNA was isolated for RNA-seq.
Oyster transcriptome assembly
From a total of 4.1×108 Illumina GAIIx-sequenced cDNA reads of 108 bp, the final set consisted of 3.8×108 reads of 94±5 bp after filtering and adaptor trimming. After the Trinity assembly of 374,029 contigs was filtered for contaminants, 356,237 contigs remained with a mean length of 440 bp and an N50 of 487 bp (link to assembly available at Text S1). A BLASTx search to the NCBI NR protein database led to annotation of 19.8% of the transcriptome. Of the total transcriptome, 22,934 contigs (16.3%) were at least 1 Kb in length. When the final set of processed reads (3.8×108) were mapped to the transcriptome, 58% of the reads mapped to at least one alignment (Table 1).
Table 1. Assembly metrics, annotation information, and reads mapped for transcriptome assembly.
| Number of contigs | 356,237 |
| Total span (bp) | 156,920,694 |
| Number of contigs >1Kb | 22,934 |
| Max Contig Length (bp) | 16,256 |
| Mean Contig Length (bp) | 440 |
| N50 (bp) | 487 |
| Number of contigs with BLAST hits* | 70,621 |
| % of contigs with BLAST hits* | 19.8 |
| % of reads mapped to transcriptome (bowtie [32]) | 58.13 |
*Contigs compared to NCBI’s non-redundant protein database using BLASTx, hits with e-value≤1e-06 retained.
General patterns of gene expression in oysters in response to bacterial challenge
Principal components and heat map analyses were performed to evaluate general patterns of variation in gene expression between treatment groups. Principal components (PC) analysis showed that 93% of the variation in gene expression between groups is explained by 8 principal components, with 24%, 14%, and 12% of the variance explained by PC1, PC2, and PC3. The first PC separated treatments by family (F3L from GX), suggesting that the largest component of the variation (24%) in gene expression patterns can be attributed to genetic differences between the two families. The second component PC2 (explaining 14% of the variance) separated treatments by time after challenge (Figure 2). Gene expression patterns for GX-15d and CGX-15d, as well as GX-30 and CGX-30 clustered together in the projection of PC1 and PC2, showing relatively higher similarity between control and challenged resistant oysters at these time points. Based on similarity, comparisons of gene expression between resistant control and challenged oysters at days 15 and 30 were not included in further analyses of differential gene expression.
Figure 2. Principal components (PC) analysis of gene expression in resistant and susceptible oysters experimentally challenged with Roseovarius crassostreae: Spatial projection of PC1 and PC2.

The Z-score centered log2-transformed RPKM for each transcript in challenged susceptible oysters at days 1, 5, 15, and 30 after challenge (F3L_1 to F3L_30), challenged resistant oysters at days 1 to 30 (GX_1 to GX_30), and unchallenged resistant oysters at days 15 and 30 (CGX_15, CGX_30) was used in the PCA. Data from unchallenged resistant oysters at days 1 and 5 were not included in the analysis due to the potential confounding effect of an unrelated mortality event observed before day 7. Gene expression in unchallenged susceptible oysters (CF3L) was not studied.
Consistent with the results from the PCA, heat map cluster analysis showed two major clusters separating F3L and GX/CGX treatments, suggesting that a major portion of the variation in gene expression is due to genetic differences between the resistant and susceptible families (Figure 3). Within these major clusters, the following subclusters were detected that separated treatments within family based on time: GX 1 and 5d (designated GX_early), F3L 1 and 5d (F3L_early), F3L 15 and 30d (F3L_late), and CGX 15 and 30d (control) (Figure 3).
Figure 3. Heatmap of differentially expressed transcripts in resistant and susceptible oysters experimentally challenged with Roseovarius crassostreae.
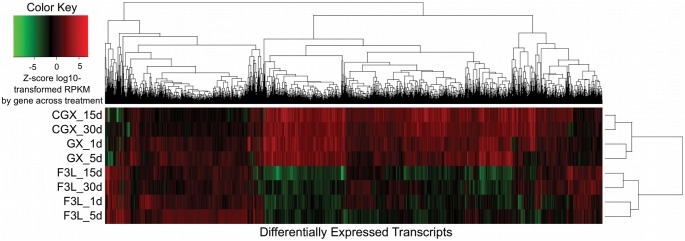
The Z-score centered log2-transformed RPKM for each transcript in each of eight sample groups is shown using a color scale. Genes are hierarchically clustered using Euclidean distance and complete linkage of the Z-score-transformed gene expression. Sample groups are clustered using the complete linkage Euclidean distance of the Spearman correlation of the Z-score-transformed gene expression.
Based on: a) the unexplained mortality observed in the unchallenged resistant oysters by day 7, which precluded the use of the data from the control unchallenged resistant oysters (CGX) on days 1 and 5 oysters as controls for differential gene expression; and b) results from the PC (Figure 2) and heat map (Figure 3) analyses, which clustered samples first by family, and then by early and late time points, we decided to strengthen the statistical analysis of differential gene expression by considering data within each family at days 1 and 5 as replicates (GX_early, F3L_early). The same was done for the data from days 15 and 30 for the susceptible oysters (F3L_late). As a first step in identifying genes and processes differentially expressed in response to bacterial challenge, as well as genes that may be involved in disease resistance, differential gene expression of challenged resistant GX and susceptible F3L oysters at the early and late time points relative to the control unchallenged resistant oysters collected at 15 and 30 days (CGX_late) was determined.
Of the 356,237 total transcripts tested for differential expression relative to CGX_late, 6,097 (1.7%) transcripts were differentially expressed in F3L_early and/or F3L_late, compared to only 552 (0.15%) transcripts differentially expressed in GX_early (Figure 4). This is consistent with the expectation that a relatively large component of the variation in gene expression is due to genetic differences between the oyster families. While differential gene expression in resistant (GX_early) oysters can be attributed to responses of oysters to challenge, as well as some temporal differences in gene expression, the larger amount of transcripts differentially expressed in susceptible F3L oysters is probably due to both differences in gene expression in response to challenge and to genetic differences between families.
Figure 4. Differentially expressed transcripts in response to bacterial challenge shared and unique between resistant and susceptible oysters.
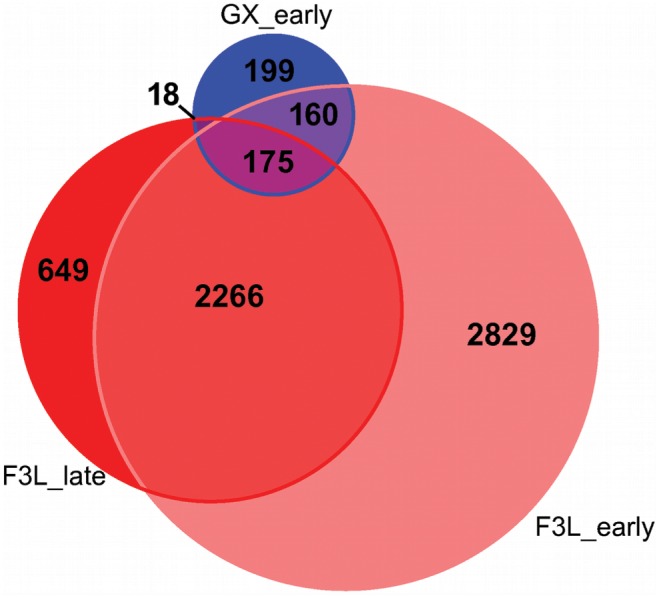
Venn diagram of shared and unique differentiall expressed transcripts in GX_early (resistant family–days 1 and 5), F3L_early (susceptible – days 1 and 5), and F3L_late (susceptible – days 15 and 30) oysters after challenge with the bacterial pathogen Roseovarius crassostreae.
Enriched Gene Ontology terms and differentially expressed genes common to resistant and susceptible oysters in response to challenge with R. crassostreae
A gene ontology (GO) enrichment analysis was also performed to determine which biological processes were most highly represented (significantly enriched) amongst differentially expressed genes in each of the treatment groups. This approach to differential gene expression analysis should account for the variability in gene expression derived from the effect of time (day of sampling) while providing: a) a broad overview of the responses of oysters to challenge with R. crassostreae (differentially expressed genes and enriched GO terms observed in F3L and GX oysters in response to challenge); b) a list of genes potentially involved in disease susceptibility to ROD (genes differentially expressed in F3L, but not GX, in response to challenge); and c) a list of genes potentially involved in disease resistance to ROD (genes differentially expressed in GX but not detected in F3L, in response to challenge). Limitations of this approach will be addressed in the discussion section.
Differentially expressed annotated genes and enriched Gene Ontology (GO) categories shared between resistant and susceptible oysters should provide a general view of the most conserved transcripts and molecular processes associated with host defenses to R. crassostreae in these 2 oyster families. Many of the genes diferentially expressed in among challenged oysters from both resistant GX and susceptible F3L families were associated with the gene ontology terms “defense response”, “defense response to bacterium”, “response to molecule of bacterial origin”, and “protein folding” (Figure 5A–C), as well as the related molecular functions “enzyme inhibitor activity”, “endopeptidase inhibitor activity”, “endopeptidase regulator activity”, and “peptidases” (Figure 5D–F). Examples of the most highly differentially expressed transcripts in response to bacterial challenge shared between resistant GX and susceptible F3L included several transcripts involved in immune recognition and signaling, such as C1q domain-containing (C1qDC) proteins, scavenger receptors cysteine-rich, c-type lectins, and dopamine-beta hydroxylase-like proteins. They also include several transcripts corresponding to the immune effectors serine protease inhibitors and a few annotated transcripts involved in detoxification, such as cytochrome p450 and glutathione S-transferase (Table 2).
Figure 5. Functionally enriched Gene Ontology terms in the transcriptome of resistant and susceptible oysters in response to bacterial challenge.

Functionally enriched Gene Ontology (GO) terms among differentially expressed transcripts in resistant oysters at 1 and 5 days after bacterial challenge (GX_early, A, D, G), susceptible oysters at 1 and 5 days after bacterial challenge (F3L_early, B, E, H), and susceptible oysters at 15 and 30 days after bacterial challenge (F3L_late, C, F, I) are displayed for biological processes (A–C), molecular function (D–F), and cellular component (G–I). Each GO term category is represented by a shape (circle or square) in the same x,y location in each of the graphs. The color of the shapes from cool (green) to warm (red) signifies increasing significance of enrichment as indicated in the color key. The size of shapes reflects whether the GO term is enriched among up-regulated DE transcripts (large) or down-regulated DE transcripts (small), while a GO term enriched among both up-regulated and down-regulated transcripts is represented by a square shape. Overlapping shapes corresponding to functionally similar categories have been labeled using a more general term, noted by the suffix “-related”.
Table 2. Annotated transcripts differentially expressed in both resistant (GX_early) and susceptible (F3L early and/or late) oysters in response to challenge with R. crassostreae.
| Contig | Type ofDE (GX) | Type ofDE (F3L) | Annotation | Accession # |
| comp3607_c0_seq3 | Up | Up | arylsulfatase a-like | XP_002607295 |
| comp1799_c0_seq1 | - | Down | c-type lectin | XP_002603342 |
| comp10350_c0_seq1 | Down | - | c-type lectin 2 | XP_002603342 |
| comp3136_c3_seq2 | - | Down | c1q domain containing protein 1q11 | CBX41660 |
| comp887_c0_seq1 | Up | Up | c1q domain containing protein 1q13 | CBX41662 |
| comp12483_c0_seq1 | Down | Down | c1q domain containing protein 1q13 | CBX41662 |
| comp4668_c0_seq2 | - | Down | c1q domain containing protein 1q40 | CBX41689 |
| comp1886_c0_seq5 | Up | c1q domain containing protein 1q83 | CBX41732 | |
| comp6091_c0_seq2 | Down | Down | camp responsive element binding 2 | AAU93879 |
| comp1102_c0_seq3 | Down | Down | collagen alpha-5 chain | XP_002595170 |
| comp4943_c0_seq1 | Up | Up | cytochrome family subfamily polypeptide 2-like | XP_002594971 |
| comp13170_c1_seq3 | Down | Down | deleted in malignant brain tumors 1 | XP_002833280 |
| comp7186_c1_seq4 | Up | Up | dna damage-regulated autophagy modulator protein 2 | NP_001230625 |
| comp631_c0_seq1,2 | Up | Up | dopamine beta hydroxylase-like | XP_002117561 |
| comp1893_c0_seq1 | Up | Up | dopamine beta hydroxylase-like | AAS92605 |
| comp8625_c0_seq1 | Up | Up | fatty acid synthase-like | ACZ55138 |
| comp2451_c0_seq15 | Up | Up | galactosamine (n-acetyl)-6-sulfate sulfatase-like | XP_002605064 |
| several (>10) contigs | Down | Down | gtpase imap family members | XP_001920359, AAH96680 |
| comp3498_c0_seq2 | Up | Up | heat shock protein 60 | ABN11936 |
| comp5396_c0_seq2 | Down | Down | melatonin receptor 1a | ADM73175 |
| comp3971_c0_seq4 | Down | Down | monocarboxylate transporter | XP_002573719 |
| comp3971_c0_seq1 | Down | Down | monocarboxylate transporter | XP_001606814 |
| comp3971_c0_seq2 | Down | Down | monocarboxylate transporter | EGI68511 |
| comp11520_c0_seq1 | Up | Up | nose resistant to fluoxetine family member (nrf-6)-like | XP_002600112 |
| comp50794_c0_seq1 | Down | Down | novel protein human megf11 | EGW04058 |
| comp30091_c0_seq2 | Down | Down | nudt9 | EGD73755 |
| comp9303_c0_seq5 | Up | Up | omega class glutathione s-transferase | CAD89618 |
| comp11276_c0_seq3 | Up | Up | polyketide synthase pks2 | XP_002734101 |
| comp25817_c0_seq1 | Down | Down | protein tyrosine phosphatase | ACH42087 |
| several contigs | Up | Up | scavenger receptor cysteine-rich protein | XP_001186391 |
| comp2875_c0_seq2 | Up | Up | serine protease | XP_002593726 |
| comp3584_c0_seq1 | Down | Up | serine protease inhibitor cvsi-1 | Q30HU9 |
| comp619_c0_seq1 | Up | Up | serine protease inhibitor cvsi-2 | B9A8D7 |
| comp928_c0_seq1 | Up | Up | serine protease inhibitor cvsi-2 | B9A8D7 |
| comp28180_c0_seq1 | Down | Down | sushi-repeat-containing x-linked 2 | XP_002932840 |
| comp869_c0_seq2 | Down | Down | x-box binding | XP_002732738 |
Contig number, direction of differential expression (up- or down-regulation), and name and accession number for the best BLASTx hits are shown.
Consistent with the extracellular nature of the infection by the bacterial pathogen R. crasssotreae, the most commonly differentially expressed transcripts in both resistant and susceptible oysters annotate to genes corresponding to the cellular component GO terms corresponding to membrane or extracellular regions (Figure 5G–I).
Enriched Gene Ontology terms and differentially expressed genes unique to susceptible oysters in response to challenge with R. crassostreae
Annotated transcripts that are differentially expressed in challenged susceptible F3L (but not in challenged resistant GX oysters) relative to unchallenged CGX oysters, should include, among others, transcripts characteristic of the responses of oysters to an acute infection that may not be effective in removing the pathogen. These transcripts could be candidates for markers of disease susceptibility to ROD. Gene Ontology terms enriched in this group provide a general overview of the processes associated with the response of susceptible oysters to the bacterial pathogen. Many of the processes highly differentially regulated in susceptible F3L oysters (but not in resistant oysters) in response to challenge were related to metabolic functions, including hexose, carboxylic acid, and carbohydrate metabolic processes (Figure 5B,C), suggesting that infection with R. crassostreae may place a large metabolic demand on susceptible oysters. Examples of transcripts in these categories included several genes involved in detoxification, such as several transcripts for genes of the cytochrome p450 family (Table 3). Consistent with this, the terms “monooxygenase activity” (Fig. 5E) and “oxidoreductase activity” (Fig. 5F) were significantly enriched in susceptible oysters. The most significantly enriched F3L_early biological process term was “cholesterol transport” (Figure 5B), corresponding to epididymal secretory protein E1 (Table 3). Annotated transcripts showing the highest degree of differential expression in susceptible oysters, but not resistant oysters, included those coding for several heat shock proteins, several fibrinogen c domain-containing proteins, cadherin, legumain, vgd3, dermatopontin-2, and apextrin (Table 3).
Table 3. Top annotated transcripts differentially expressed in susceptible oysters at early and late timepoints after challenge with R. crassostreae.
| Contig | logFC_1d | logFC_5d | logFC_15d | logFC_30d | Annotation | Accession # |
| comp1465_c0_seq5 | −11.10 | −11.00 | - | - | adipose differentiation-related protein | XP_002595036 |
| comp1612_c0_seq1 | −10.62 | −4.16 | - | - | ankyrin unc44 | XP_001190300 |
| comp20450_c0_seq1 | - | - | −10.12 | −10.18 | ankyrin unc44 | XP_782809 |
| comp781_c0_seq4 | - | - | 4.05 | 4.99 | apextrin-like protein | AEK10749 |
| comp6246_c1_seq1 | 2.83 | 4.62 | - | - | cadherin- isoform h | BAD91058 |
| comp4943_c0_seq1 | - | - | 3.27 | 4.55 | cytochrome family subfamily polypeptide 2-like | XP_002594971 |
| comp4943_c0_seq4 | - | - | 2.27 | 4.06 | cytochrome p450 2b11 | BAD02925 |
| comp4943_c0_seq5 | - | - | 2.56 | 4.57 | cytochrome p450 2g1-like | NP_001106451 |
| comp4943_c0_seq6 | - | - | 3.54 | 4.74 | cytochrome p450 2k | XP_002594509 |
| comp1153_c0_seq7 | −5.74 | −7.45 | - | - | dermatopontin 2 | AAZ80787 |
| comp1153_c0_seq5 | −4.88 | −6.67 | - | - | dermatopontin 2 | XP_001628981 |
| comp1382_c0_seq1 | - | - | 7.21 | 8.66 | dermatopontin 2 | XP_001628981 |
| comp211_c0_seq2 | - | - | −9.54 | −4.24 | developmentally-regulated vdg3 | ABB76764 |
| comp211_c0_seq3 | - | - | - | 3.90 | developmentally-regulated vdg3 | ABB76764 |
| comp211_c0_seq5 | −12.84 | −12.73 | −8.48 | −7.23 | developmentally-regulated vdg3 | ABB76764 |
| comp7066_c0_seq1,2 | - | - | - | 4.14 | developmentally-regulated vdg3 | ABB76764 |
| comp84_c0_seq2 | 3.74 | 5.64 | - | - | developmentally-regulated vdg3 | ABB76764 |
| comp84_c0_seq3 | 2.82 | 4.73 | - | - | developmentally-regulated vdg3 | ABB76764 |
| comp14053_c0_seq4 | - | 2.26 | 2.24 | - | epididymal secretory protein e1 precursor | ACO09278 |
| comp1891_c0_seq1 | - | 3.02 | - | - | epididymal secretory protein e1 precursor | AAX61146 |
| comp1891_c0_seq2 | - | 1.98 | - | - | epididymal secretory protein e1 precursor | XP_003408814 |
| comp1234_c1_seq4 | −8.35 | −6.77 | - | - | fibrinogen c domain-containing protein 1-a-like | XP_003390678 |
| comp3625_c0_seq2 | −10.54 | −5.97 | - | - | fibrinogen c domain-containing protein 1-like | XP_003391179 |
| comp3814_c0_seq1 | - | - | −11.06 | −6.66 | fibrinogen-related protein | XP_002609404 |
| comp9922_c0_seq7 | 5.53 | 2.52 | heat shock 70 kda protein cognate 3 | BAD15288 | ||
| comp2543_c1_seq1 | - | - | −11.71 | −5.66 | hemicentin 2-like | XP_002731765 |
| comp7788_c0_seq2 | −10.40 | −5.31 | - | - | hippocalcin-like protein 1 | XP_001639635 |
| comp6419_c0_seq3 | −11.25 | - | - | - | ing (mammalian inhibitor of growth) homolog family member (ing-3)-like | XP_002165512 |
| comp8790_c1_seq1 | - | - | −11.57 | −5.16 | legumain [Haliotis discus discus] | ABO26629 |
| comp1637_c0_seq1 | −5.38 | −4.31 | −2.86 | −5.54 | low affinity immunoglobulin epsilon fc receptor | NP_001138689 |
| comp1890_c0_seq1 | −10.44 | −2.63 | - | - | oncoprotein-induced transcript 3 | XP_002595494 |
| comp40_c0_seq9 | 6.11 | 8.14 | - | - | period clock protein | ABM66066 |
| comp1593_c0_seq2 | −7.74 | −6.47 | - | - | proliferating cell nuclear antigen-like | XP_001631319 |
| comp4352_c1_seq3 | - | - | −10.54 | −10.86 | ring finger protein 145 | AAH55485 |
| comp11695_c0_seq2 | 5.11 | 4.01 | 7.25 | 6.17 | tyrosine-protein kinase fer- partial | XP_002945302 |
Magnitude of differential expression in each treatment group is expressed as log10 fold change (logFC) over expression in the control group (CGX15/30d). The name and accession number for the best BLASTx hits are shown.
Enriched Gene Ontology terms and differentially expressed genes unique to resistant oysters in response to challenge with R. crassostreae
Genes (annotations) differentially expressed in GX_early relative to unchallenged CGX_late that were not differentially expressed in susceptible oysters after bacterial challenge should include, among others, transcripts and processes contributing to host-defenses and disease resistance in the GX family. The biological process GO terms most significantly enriched among GX_early up-regulated transcripts and not present in susceptible oysters were the related terms “programmed cell death” and “apoptotic process,” corresponding to transcripts that annotated as inhibitor of apoptosis (IAP) proteins (Table 4). Other examples of differentially expressed annotated transcripts unique to GX_early included several transcripts that annotated to genes associated with the remodeling of the extracellular matrix (ECM), such as ADAMTS8 and furin, as well as several trancripts involved in immune recognition (scavenger receptor cysteine-rich), signaling (interleukin 17, rapunzel), and regulation of effector functions (arginase) (Table 4).
Table 4. Top annotated transcripts differentially expressed in resistant oysters at early time points after challenge with R. crassostreae (GX 1 and 5 days).
| Contig | logFC_1d | logFC_5d | Annotation | Accession # |
| comp1506_c0_seq4 | 2.71 | 3.08 | ADAMTS8 | XP_002940685 |
| comp4626_c0_seq4 | 3.34 | - | alpha-ketoglutarate-dependent hypophosphite dioxygenase-like | XP_002944900 |
| comp1285_c1_seq8 | - | −3.90 | arginase type-i-like, arginase-i | AEB70965 |
| comp1285_c1_seq3 | - | −3.54 | arginase type-i-like, arginase-i | XP_002130834 |
| comp5608_c0_seq1 | - | −6.92 | c-type lectin | ABB71672 |
| comp24124_c0_seq1 | - | 4.49 | ched related family member (ptr-19) | XP_002734100 |
| comp5722_c1_seq1,2 | - | −3.35 | collagen alpha | XP_001512734 |
| comp7972_c0_seq1 | - | −8.09 | cubilin | XP_002734392 |
| comp7972_c0_seq4 | - | −3.11 | cubilin | XP_002612977 |
| comp1788_c0_seq4 | - | 3.26 | fibrinolytic enzyme | CAA64472 |
| comp18756_c0_seq3 | 3.48 | − | fibropellin ia | XP_002601363 |
| comp6161_c0_seq11 | - | 2.48 | furin | AAA49718 |
| comp810_c1_seq1 | - | −3.40 | heat shock protein 22 | ACU83231 |
| comp18757_c0_seq1 | - | −7.14 | hla-b associated transcript 1 | XP_003217350 |
| comp2015_c0_seq13 | −7.19 | - | inhibitor of apoptosis (IAP) | AEB54799 |
| comp2015_c0_seq24 | 3.95 | - | inhibitor of apoptosis (IAP) | AEB54800 |
| comp15440_c0_seq1 | 3.18 | - | inhibitor of apoptosis (IAP) | XP_002426441 |
| comp6837_c0_seq1 | 3.02 | - | interleukin 17 | A9XE49 |
| comp3858_c0_seq5 | 3.63 | - | isoleucyl-trna synthetase | NP_001090690 |
| comp5396_c0_seq1 | - | −3.01 | melatonin receptor 1a | ADM73175 |
| comp7137_c0_seq2 | - | 3.26 | organic solute transporter subunit alpha | XP_002732822 |
| comp39520_c0_seq1 | - | −4.31 | polyprotein | XP_002740782 |
| comp24428_c0_seq1 | −8.68 | −8.64 | rapunzel 5 | NP_001103594 |
| comp18902_c0_seq1 | - | −3.43 | rho gtpase | XP_002739105 |
| comp1023_c0_seq2 | - | 2.62 | scavenger receptor cysteine-rich | ACT53266 |
| comp1023_c0_seq3 | 3.07 | 2.93 | scavenger receptor cysteine-rich | XP_001622238 |
| comp18756_c0_seq2 | 3.83 | - | sushi repeat-containing | XP_002664481 |
| comp25746_c0_seq4 | - | −7.60 | tenascin xb | XP_002741293 |
| comp6161_c0_seq5 | 3.49 | - | type 2 proinsulin processing endopeptidase | 2206277A |
Magnitude of differential expression in each treatment group is expressed as log10 fold change (logFC) over expression in the control group (CGX15/30d). The name and accession number for the best BLASTx hits are shown.
Discussion
The two oyster families used in this study showed a dramatic difference in mortality to challenge with the bacterial pathogen R. crassostreae, causative agent of Roseovarius Oyster Disease (ROD) in juvenile oysters. While the susceptible oysters experienced constant levels of mortality due to ROD throughout the length of the challenge, oysters from the resistant family showed levels of mortality equal to the non-challenged oysters, even if they were continuously exposed to the pathogen through cohabitation with the susceptible oysters. We exploited these differences in mortality patterns, and ultimately differences in gene expression, to mine for genes and processes potentially involved in: 1) host-pathogen interactions in juvenile American oysters, and 2) disease resistance or susceptibility to ROD. Our analysis of the gene ontology terms most commonly represented (enriched) amongst the genes differentially expressed in challenged resistant and susceptible oysters relative to non-challenged oysters provides a broad view of the most conserved genes and processes involved in host responses of juvenile American oysters to R. crassostreae. We found that transcripts related to pathogen recognition, immune signaling and effector molecules, apoptosis, and detoxification were involved in the responses of the American oyster to bacterial challenge. In addition, we have identified several genes showing differential patterns of gene expression in either susceptible or resistant oysters in response to challenge, providing a useful foundation for the future identification of genes involved in disease resistance or susceptibility to ROD.
There are several limitations to this study that should be considered in the interpretation of the results. Although the levels of annotation achieved in this study (20% of the transcriptome) are comparable to the results of previous Illumina-generated transcriptome analyses in oysters (e.g. C. gigas, 16–23% annotated [48], [49]), our work should be viewed as an initial exploration of the most evolutionarily conserved aspects of the American oyster’s responses to R. crassostreae challenge. Furthermore, due to the limitations of the experimental design (differential gene expression in challenged F3L and GX was determined relative to that in unchallenged CGX oysters collected on days 15 and 30 after the start of the challenge), patterns of gene expression observed in this study could be due to either: a) true differences in gene expression between groups (for the most conserved genes); b) genetic differences between the families (transcripts corresponding to the same gene being identified as different genes in the assembly); and/or c) variability in gene expression between early and late time points (for the F3L_early and GX_early comparisons). Therefore, further work should be done in the future to validate the role of these genes in disease resistance/susceptibility to ROD and identify potential mechanisms of disease resistance. Constitutive and inducible differences in gene expression, as well as genetic differences between families are some of the mechanisms involved in disease resistance [13].
Juvenile oyster responses to challenge with R. crassostreae
Differentially expressed annotated transcripts shared between resistant and susceptible oysters may provide insights into the most common immune responses of oysters to challenge with R. crassostreae. For both susceptible and resistant oysters, major immune responses to R. crassostreae included pathogen recognition, signaling, serine protease inhibition, detoxification, and apoptosis. Transcripts differentially expressed in both resistant and susceptible oysters in response to challenge annotating to genes involved in pathogen recognition included scavenger receptors cysteine-rich (SRs), C1qDC proteins, and c-type lectins [20]–[22], [50], [51]. Scavenger receptors are differentially expressed in oyster species in response to summer mortality [52], [53] and hypoxia [54]. Recently, an SR protein representing a novel class of scavenger receptor has been characterized in the scallop Chlamys farreri that is up-regulated by exposure to Pathogen Associated Molecular Patterns (PAMPs) like LPS, peptidoglycan and β-glucan and can bind LPS and peptidoglycan [55]. The role of C1qDC proteins as pathogen recognition receptors (PRRs) in molluscs has been solidified by a demonstration of the ability of a recombinant C1qDC protein from the scallop Argopecten irradians to bind PAMPs from diverse pathogens including gram-negative and gram-positive bacteria and fungi [56], [57]. Recently, a c-type lectin from C. farreri was shown to act as a PRR, binding LPS and β-glucan, and as an opsonin, enhancing the phagocytic capabilities of C. farreri hemocytes [58]. Interestingly, transcripts for fibrinogen-related proteins (FREPs) were differentially expressed in susceptible oysters but not in resistant oysters. This was an unexpected finding, since FREPs, which function in invertebrates in pathogen recognition, agglutination, and parasite resistance [59], have been shown in B. glabrata to contribute to resistance against the parasite Schistosoma mansoni [60]. A FREP in the bay scallop Argopecten irradians has agglutinating activity against chicken and human erythrocytes and bacteria and is up-regulated following challenge by gram-negative bacteria [61]. It is possible that FREPs expression in response to R. crassostreae may involve very early and acute up-regulation (before 24 h) or constitutive expression in resistant oysters.
Multiple dopamine beta-hydroxylase (DBH) transcripts were highly differentially expressed early in both resistant and susceptible oysters. DBH produce/modify catecholamines, which have been shown to modulate both the immune and stress response in vertebrates and invertebrates [62] including the scallop C. farreri [63], [64] and the Pacific oyster C. gigas [65], [66].
The responses of resistant and susceptible oysters to bacterial challenge also included several effectors of immunity responsible for minimizing or preventing damage caused by virulence factors from the pathogen. The second most highly up-regulated transcript at early time points for resistant oysters (also up-regulated in susceptible oysters) annotated as serine protease inhibitor-2 (Cvsi-2). Serine proteases are key virulence factors of many pathogens of bivalves, inhibiting phagocytosis in clams [66] and causing cytotoxicity of bivalve hemocytes [67], [68]. Serine protease inhibitor-1 (Cvsi-1), which was also highly up-regulated in susceptible oysters at early timepoints in this study, has been shown to have a role in the immune response of American oysters against the protozoan parasite Perkinsus marinus, likely by inhibiting the parasite’s major extracellular protease [69]. Moreover, polymorphism in the promoter of Cvsi-1 is associated with disease resistance to P. marinus [70]. We hypothesize that serine protease inhibitors may also play a role in neutralizing serine proteases released by R. crassostreae.
Transcripts differentially expressed in both resistant and susceptible oysters also included transcripts annotated as glutathione s-transferase, cytochrome p450, and multiple heat shock proteins, which are involved in detoxification and preventing oxidative damage. Glutathione s-transferase is an anti-oxidant and is up-regulated in hemocytes of Pacific oysters challenged with a pathogenic Vibrio sp. [17]. Although cytochrome p450s have been best studied in detoxification of xenobiotics in bivalves [71], they have also been implicated in the host defense responses of the flat oyster Ostrea edulis to the parasite Bonamia ostreae [72] and the Manila clam Ruditapes phillipinarum to Vibrio tapetis (the causative agent of Brown Ring Disease, a disease with clinical signs similar to ROD) [73]. Heat shock protein 60 is involved in xenobiotic detoxification and the stress response in oysters [74] and has an important role in immunity in mammals [75].
Another process involved in the responses of both the resistant and susceptible oysters to bacterial challenge was apoptosis, a form of programmed cell death that plays an important role in many processes, including immunity and development [76], [77]. In our study, inhibitor of apoptosis (IAP) and GTPase of the immunity-associated protein (GIMAP) transcripts were differentially expressed in both resistant and susceptible oysters. GIMAP proteins are important regulators of apoptosis [78]. Exposure of human monocytes to LPS induces the down-regulation of GIMAP, which may serve to promote the survival of monocytes by negatively regulating apoptosis [79]. We hypothesize that GIMAP proteins may serve an analogous function in oyster hemocytes. Apoptosis in general and IAP proteins in particular are associated with molluscan immunity [80], participating in the defense response of clams to V. tapetis [73], [81] and oysters to the protozoan parasite P. marinus [82], [83]. Four apoptosis-related genes, including IAP, were induced in Pacific oysters in response to challenge with the bacterial pathogen V. anguillarum [84]. Further work is needed to evaluate the potential role of IAP and GIMAP genes in hemocyte activation and survival.
Potential genes involved in susceptibility to Roseovarius oyster disease
While resistant oysters appeared to be able to rapidly eliminate the pathogen, susceptible oysters suffered constant levels of morbidity and mortality throughout the challenge. Genes and processes activated in susceptible oysters in response to bacterial challenge and absent or present to a much lesser degree in resistant oysters may be used as indicators of an unsuccessful defense response and provide further insights on the molecular basis of disease susceptibility. Enrichment of transcripts corresponding to metabolic processes in susceptible oysters supports the hypothesis that a failed immune response against ROD places a large metabolic demand on these oysters, leading to mortality events [5], [6], [8]. Moreover, the unique enrichment in transcripts involved in detoxification (monooxygenase and oxidoreductase activities) in susceptible oysters could also be a reflection of the impact of ongoing acute infections in susceptible oysters, leading to upregulation of genes involved in minimizing the damage produced by the bacterial pathogen.
Other top genes differentially expressed in susceptible oysters but not in resistant oysters in our study that may warrant further investigation include epididymal secretory protein E1, cadherin, legumain, vdg3, dermatopontin 2, and apextrin. Epididymal secretory protein E1, also known as Niemann-Pick type C-2, facilitates cholesterol transport from lysosomes [85]. The up-regulation of epididymal secretory protein e1 uniquely in susceptible oysters at days 5 and 15 may represent lysomal turnover as an aspect of a continuous response to bacteria [86]. Cadherin is involved in cell adhesion [87], and legumain is a cysteine protease associated with response to bacteria [88], antigen processing [89], and ECM remodeling [90] in mammals. Transcripts annotating as dermatopontin-2, a shell matrix protein involved in cell adhesion and shell formation [91], [92] may be involved in bacterial encapsulation and conchiolin production (a hallmark clinical sign of ROD). Dermatopontin is strongly induced in amphioxus following bacterial challenge by gram-positive and gram-negative bacteria [93]. In the same bacterial challenge of amphioxus, another highly up-regulated gene was apextrin. Known primarily for its role in embryonal development [94], apextrin is a member of the membrane attack complex/perforin domain protein family [95] and is involved in innate antibacterial responses, possibly by sequestering or inactivating bacteria [96].
Potential genes involved in resistance to Roseovarius oyster disease
Oysters from the resistant family did not show clinical signs of infection and suffered mortalities comparable to non-challenged oysters, suggesting that these oysters were able to eliminate the pathogen rapidly. ROD-resistant oysters responded to the bacterial pathogen R. crassostreae mainly by the differential expression of transcripts annotating to proteins that modify the extracellular matrix (ECM) (e.g. ADAMTS8), proteins that bind self or non-self ligands including pathogens (e.g. scavenger receptor cysteine-rich), stress proteins (e.g. HSP20, 60, and 70), and proteins involved in signaling (e.g. IL-17, cubilin, rapunzel). The up-regulation in resistant oysters of furin suggests the possible involvement of neuroendocrine signaling and/or host defense-relevant protein processing. Furin is involved in the processing of von Willebrand Factor, antimicrobial peptides in invertebrates, and certain matrix metalloproteinases; which in turn affect cell migration, differentiation, inflammation control, and the restructuring of the ECM [97]. The importance of ECM restructuring in the response of resistant oysters to bacterial challenge is corroborated by the up-regulation of a transcript annotating as ADAMTS8, a matrix metalloproteinase that is activated through cleavage by furin and likely participates in ECM proteolysis [98], as well as the differential expression of transcripts coding for tenascin-xb, an anti-adhesive glycoprotein involved in wound healing and matrix maturation [99].
The early response in resistant oysters also involved the pro-inflammatory mediator, interleukin 17 (IL-17), and the nitric oxide modulator, arginase. Members of the IL-17 family of cytokines induce the expression of antimicrobial proteins [100], and previous research suggests that IL-17 is an important mediator of the pro-inflammatory response in oysters [101]. Our results are consistent with the role of IL-17 in the immune response of oysters against bacterial infection and suggest a potential role in disease resistance to ROD. Arginases have been shown in macrophages to modulate the production of nitric oxide [102], which is an immune effector in the American oyster [103]. Using microarray technology, a transcript annotating as arginase was shown to increase rapidly after 6 h of heat stress in C. gigas [104]. The down-regulation of arginase in resistant oysters on day 5 may signal a down-regulation of the inflammation and stress response following a successful defense response.
In conclusion, this study shows that transcripts involved in processes such as pathogen recognition, extracellular matrix remodeling, detoxification, apoptosis, and regulation of the inflammatory (i.e. hemocytic infiltration) response may have an important role in the immune defenses of American oysters against Roseovarius crassostreae, the causative agent of Roseovarius Oyster Disease. This work represents the first deep characterization of the transcriptome of American oysters in response to a bacterial pathogen, providing many candidates genes and processes that should be targeted in the future characterization of mechanisms of resistance and susceptibility to this important bacterial disease of juvenile American oysters. The present study has also generated a pool of genes to be considered for further evaluation as candidate markers for advanced genotypic selection regimes for disease resistance in oysters.
Supporting Information
Assembled transcriptome of American oysters in response to challenge with Roseovarius crassostreae.
(DOCX)
Acknowledgments
We are grateful to all Lane and Gomez-Chiarri lab members who have helped contribute to this project including I. Misner, J. Piesz, M. Karim, and S. Sohn (University of Rhode Island). We thank S. Siebert (Brown University) for helpful scripts and computational advice, and the staff at Brown University’s Center for Computing and Visualization, especially L. Dong and M. Howison, for their timely support. We thank D. Proestou, S. Roberts, and an anonymous reviewer for the useful comments on the manuscript. We also thank the support of R. Rheault and the East Coast Shellfish Growers Association.
Funding Statement
This research has been funded by USDA NIFA AFRI award 2009-38925-19971 to MGC, CEL, and DP, Rhode Island Agricultural Experiment Station Hatch13 901-INT to MGC, CEL, and RR, and RI-NSF EPSCoR grant no. 0554548. Work by DA and SI was funded by NSF awards 1048831 and 1321000 to SI. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.National Marine Fisheries Service (NMFS) (2012) Annual Commercial Landing Statistics, Fisheries Statistics. Available: http://www.st.nmfs.noaa.gov/commercial-fisheries/commercial-landings/annual-landings/index. Accessed 2014 Jan 17.
- 2. Newell RI (2004) Ecosystem influences of natural and cultivated populations of suspension-feeding bivalve molluscs: a review. J Shellfish Res 23: 51–62. [Google Scholar]
- 3. Gutiérrez JL, Jones CG, Strayer DL, Iribarne OO (2003) Mollusks as ecosystem engineers: the role of shell production in aquatic habitats. Oikos 101: 79–90. [Google Scholar]
- 4.Ford SE, Tripp MR (1996) Diseases and defense mechanisms. In: The American Oyster, Crassotrea virginica. 581–600.
- 5. Ford SE, Borrero FJ (2001) Epizootiology and Pathology of Juvenile Oyster Disease in the Eastern Oyster, Crassostrea virginica. J Invertebr Pathol 78: 141–154. [DOI] [PubMed] [Google Scholar]
- 6. Bricelj VM, Ford SE, Borerro FJ, Perkins FO, Rivara G, et al. (1992) Unexplained mortalities of hatchery-reared, juvenile oysters, Crassostrea virginica (Gmelin). J Shellfish Res 11: 331–347. [Google Scholar]
- 7. Boettcher KJ, Geaghan KK, Maloy AP, Barber BJ (2005) Roseovarius crassostreae sp. nov., a member of the Roseobacter clade and the apparent cause of juvenile oyster disease (JOD) in cultured Eastern oysters. Int J Syst Evol Microbiol 55: 1531–1537. [DOI] [PubMed] [Google Scholar]
- 8. Boardman CL, Maloy AP, Boettcher KJ (2008) Localization of the bacterial agent of juvenile oyster disease (Roseovarius crassostreae) within affected eastern oysters (Crassostrea virginica). J Invertebr Pathol 97: 150–158 10.1016/j.jip.2007.08.007 [DOI] [PubMed] [Google Scholar]
- 9. Boettcher KJ, Barber BJ, Singer JT (2000) Additional evidence that juvenile oyster disease is caused by a member of the Roseobacter group and colonization of nonaffected animals by Stappia stellulata-like strains. Appl Environ Microbiol 66: 3924–3930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Gómez-León J, Villamil L, Salger S, Sallum R, Remacha-Triviño A, et al. (2008) Survival of eastern oysters Crassostrea virginica from three lines following experimental challenge with bacterial pathogens. Dis Aquat Organ 79: 95–105. [DOI] [PubMed] [Google Scholar]
- 11. Davis CV, Barber BJ (1999) Growth and survival of selected lines of eastern oysters, Crassostrea virginica (Gmelin 1791) affected by juvenile oyster disease. Aquaculture 178: 253–271. [Google Scholar]
- 12. Barber BJ, Davis CV, Crosby MA (1998) Cultured oysters, Crassostrea virginica, genetically selected for fast growth in the Damariscotta River, Maine, are resistant to mortality caused by Juvenile Oyster Disease (JOD). J Shellfish Res 17: 1171–1176. [Google Scholar]
- 13.Cancela ML, Bargelloni L, Boudry P, Boulo V, Dias J, et al.. (2010) Genomic approaches in aquaculture and fisheries. Introduction to Marine Genomics. Springer. 213–286.
- 14.Guo X, Wang Y, Wang L, Lee J-H (2008) Oysters. Genome mapping and genomics in fishes and aquatic animals. Springer. 163–175.
- 15. Lande R, Thompson R (1990) Efficiency of marker-assisted selection in the improvement of quantitative traits. Genetics 124: 743–756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhang L, Li L, Zhu Y, Zhang G, Guo X (2014) Transcriptome Analysis Reveals a Rich Gene Set Related to Innate Immunity in the Eastern Oyster (Crassostrea virginica). Mar Biotechnol: 1–17. doi:10.1007/s10126-013-9526-z. [DOI] [PubMed]
- 17. De Lorgeril J, Zenagui R, Rosa RD, Piquemal D, Bachère E (2011) Whole Transcriptome Profiling of Successful Immune Response to Vibrio Infections in the Oyster Crassostrea gigas by Digital Gene Expression Analysis. PLoS ONE 6: e23142 10.1371/journal.pone.0023142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Zhang G, Zhang L, Li L (2011) Gene discovery, comparative analysis and expression profile reveal the complexity of the Crassostrea gigas apoptosis system. Dev Comp Immunol 35: 603–610 10.1016/j.dci.2011.01.005 [DOI] [PubMed] [Google Scholar]
- 19. Adema CM, Hanington PC, Lun C-M, Rosenberg GH, Aragon AD, et al. (2010) Differential transcriptomic responses of Biomphalaria glabrata (Gastropoda, Mollusca) to bacteria and metazoan parasites, Schistosoma mansoni and Echinostoma paraensei (Digenea, Platyhelminthes). Mol Immunol 47: 849–860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Loker ES (2012) Macroevolutionary Immunology: A Role for Immunity in the Diversification of Animal life. Front Immunol 3: 25 10.3389/fimmu.2012.00025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Ghosh J, Lun CM, Majeske AJ, Sacchi S, Schrankel CS, et al. (2011) Invertebrate immune diversity. Dev Comp Immunol 35: 959–974. [DOI] [PubMed] [Google Scholar]
- 22. Messier-Solek C, Buckley KM, Rast JP (2010) Highly diversified innate receptor systems and new forms of animal immunity. Semin Immunol 22: 39–47. [DOI] [PubMed] [Google Scholar]
- 23.Ford SE, Haskin HH (1987) Infection and mortality patterns in strains of oysters Crassostrea virginica selected for resistance to the parasite Haplosporidium nelsoni (MSX). J Parasitol: 368–376. [PubMed]
- 24. Maloy AP, Barber BJ, Boettcher KJ (2005) A PCR-based diagnostic assay for the detection of Roseovarius crassostreae in Crassostrea virginica affected by juvenile oyster disease (JOD). Dis Aquat Organ 67: 155–162. [DOI] [PubMed] [Google Scholar]
- 25. Langmead B, Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2. Nat Methods 9: 357–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Kong Y (2011) Btrim: a fast, lightweight adapter and quality trimming program for next-generation sequencing technologies. Genomics 98: 152–153. [DOI] [PubMed] [Google Scholar]
- 27. Grabherr MG, Haas BJ, Yassour M, Levin JZ, Thompson DA, et al. (2011) Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat Biotechnol 29: 644–652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Sayers EW, Barrett T, Benson DA, Bolton E, Bryant SH, et al. (2012) Database resources of the national center for biotechnology information. Nucleic Acids Res 40: D13–D25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Finn RD, Mistry J, Tate J, Coggill P, Heger A, et al. (2009) The Pfam protein families database. Nucleic Acids Res 38: D211–D222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Finn RD, Clements J, Eddy SR (2011) HMMER web server: interactive sequence similarity searching. Nucleic Acids Res 39: W29–W37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Smit AFA, Hubley R, Green P (1996) RepeatMaster Open-3.0.
- 32. Langmead B, Trapnell C, Pop M, Salzberg SL (2009) Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol 10: R25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Zhang G, Fang X, Guo X, Li L, Luo R, et al. (2012) The oyster genome reveals stress adaptation and complexity of shell formation. Nature 490: 49–54. [DOI] [PubMed] [Google Scholar]
- 34. Li B, Dewey C (2011) RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 12: 323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Mortazavi A, Williams BA, McCue K, Schaeffer L, Wold B (2008) Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat Methods 5: 621–628. [DOI] [PubMed] [Google Scholar]
- 36. Robinson MD, Oshlack A (2010) A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol 11: R25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.R Core Team (2013) R: A Language and Environment for Statistical Computing. Vienna, Austria. Available: http://www.R-project.org.
- 38.Müllner D (2013) fastcluster: Fast hierarchical clustering routines for R and Python. Available: http://math.stanford.edu/muellner.
- 39.Warnes GR, Bolker B, Lumley T (2013) gplots: Various R programming tools for plotting data. R package version 2.6.0.
- 40. Robinson MD, McCarthy DJ, Smyth GK (2010) edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26: 139–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Ser B Methodol: 289–300.
- 42. Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389–3402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Conesa A, Götz S, García-Gómez JM, Terol J, Talón M, et al. (2005) Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21: 3674–3676. [DOI] [PubMed] [Google Scholar]
- 44. Alexa A, Rahnenführer J, Lengauer T (2006) Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 22: 1600–1607. [DOI] [PubMed] [Google Scholar]
- 45. Schlicker A, Domingues FS, Rahnenführer J, Lengauer T (2006) A new measure for functional similarity of gene products based on Gene Ontology. BMC Bioinformatics 7: 302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Supek F, Bošnjak M, Škunca N, Šmuc T (2011) REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 6: e21800 10.1371/journal.pone.0021800 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wickham H (2009) ggplot2: elegant graphics for data analysis. Springer New York. Available: http://had.co.nz/ggplot2/book.
- 48. Zhao X, Yu H, Kong L, Li Q (2012) Transcriptomic Responses to Salinity Stress in the Pacific Oyster Crassostrea gigas. PLoS ONE 7: e46244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Zhang G, Fang X, Guo X, Li L, Luo R, et al. (2012) The oyster genome reveals stress adaptation and complexity of shell formation. Nature 490: 49–54. [DOI] [PubMed] [Google Scholar]
- 50. Canton J, Neculai D, Grinstein S (2013) Scavenger receptors in homeostasis and immunity. Nat Rev Immunol 13: 621–634 10.1038/nri3515 [DOI] [PubMed] [Google Scholar]
- 51. Carland TM, Gerwick L (2010) The C1q domain containing proteins: Where do they come from and what do they do? Dev Comp Immunol 34: 785–790 10.1016/j.dci.2010.02.014 [DOI] [PubMed] [Google Scholar]
- 52. Fleury E, Moal J, Boulo V, Daniel J-Y, Mazurais D, et al. (2010) Microarray-Based Identification of Gonad Transcripts Differentially Expressed Between Lines of Pacific Oyster Selected to Be Resistant or Susceptible to Summer Mortality. Mar Biotechnol 12: 326–339 10.1007/s10126-009-9227-9 [DOI] [PubMed] [Google Scholar]
- 53. Huvet A, Herpin A, Dégremont L, Labreuche Y, Samain J-F, et al. (2004) The identification of genes from the oyster Crassostrea gigas that are differentially expressed in progeny exhibiting opposed susceptibility to summer mortality. Gene 343: 211–220 10.1016/j.gene.2004.09.008 [DOI] [PubMed] [Google Scholar]
- 54. David E, Tanguy A, Pichavant K, Moraga D (2005) Response of the Pacific oyster Crassostrea gigas to hypoxia exposure under experimental conditions. FEBS J 272: 5635–5652 10.1111/j.1742-4658.2005.04960.x [DOI] [PubMed] [Google Scholar]
- 55. Liu L, Yang J, Qiu L, Wang L, Zhang H, et al. (2011) A novel scavenger receptor-cysteine-rich (SRCR) domain containing scavenger receptor identified from mollusk mediated PAMP recognition and binding. Dev Comp Immunol 35: 227–239 10.1016/j.dci.2010.09.010 [DOI] [PubMed] [Google Scholar]
- 56. Kong P, Zhang H, Wang L, Zhou Z, Yang J, et al. (2010) AiC1qDC-1, a novel gC1q-domain-containing protein from bay scallop Argopecten irradians with fungi agglutinating activity. Dev Comp Immunol 34: 837–846 10.1016/j.dci.2010.03.006 [DOI] [PubMed] [Google Scholar]
- 57. Wang L, Wang L, Kong P, Yang J, Zhang H, et al. (2012) A novel C1qDC protein acting as pattern recognition receptor in scallop Argopecten irradians. Fish Shellfish Immunol 33: 427–435 10.1016/j.fsi.2012.05.032 [DOI] [PubMed] [Google Scholar]
- 58. Yang J, Wang L, Zhang H, Qiu L, Wang H, et al. (2011) C-Type Lectin in Chlamys farreri (CfLec-1) Mediating Immune Recognition and Opsonization. PLoS ONE 6: e17089 10.1371/journal.pone.0017089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Hanington PC, Zhang S-M (2011) The Primary Role of Fibrinogen-Related Proteins in Invertebrates Is Defense, Not Coagulation. J Innate Immun 3: 17–27 10.1159/000321882 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Hertel LA, Adema CM, Loker ES (2005) Differential expression of FREP genes in two strains of Biomphalaria glabrata following exposure to the digenetic trematodes Schistosoma mansoni and Echinostoma paraensei. Dev Comp Immunol 29: 295–303 10.1016/j.dci.2004.08.003 [DOI] [PubMed] [Google Scholar]
- 61. Zhang Y, Zhang H, Wang L, Wang B (2009) A fibrinogen-related protein from bay scallop Argopecten irradians involved in innate immunity as pattern recognition receptor. Fish Shellfish Immunol 26: 56–64 10.1016/j.fsi.2008.07.019 [DOI] [PubMed] [Google Scholar]
- 62. Ottaviani E, Franceschi C (1996) The neuroimmunology of stress from invertebrates to man. Prog Neurobiol 48: 421–440 10.1016/0301-0082(95)00049-6 [DOI] [PubMed] [Google Scholar]
- 63. Zhou Z, Wang L, Shi X, Yue F, Wang M, et al. (2012) The expression of dopa decarboxylase and dopamine beta hydroxylase and their responding to bacterial challenge during the ontogenesis of scallop Chlamys farreri. Fish Shellfish Immunol 33: 67–74 10.1016/j.fsi.2012.04.002 [DOI] [PubMed] [Google Scholar]
- 64. Zhou Z, Wang L, Shi X, Zhang H, Gao Y, et al. (2011) The modulation of catecholamines to the immune response against bacteria Vibrio anguillarum challenge in scallop Chlamys farreri. Fish Shellfish Immunol 31: 1065–1071 10.1016/j.fsi.2011.09.009 [DOI] [PubMed] [Google Scholar]
- 65. Lacoste A, Malham SK, Cueff A, Poulet SA (2001) Noradrenaline modulates oyster hemocyte phagocytosis via a β-adrenergic receptor–cAMP signaling pathway. Gen Comp Endocrinol 122: 252–259. [DOI] [PubMed] [Google Scholar]
- 66. Ordás MC, Ordás A, Beloso C, Figueras A (2000) Immune parameters in carpet shell clams naturally infected with Perkinsus atlanticus. Fish Shellfish Immunol 10: 597–609. [DOI] [PubMed] [Google Scholar]
- 67. Allam B, Ford SE (2006) Effects of the pathogenic Vibrio tapetis on defence factors of susceptible and non-susceptible bivalve species: I. Haemocyte changes following in vitro challenge. Fish Shellfish Immunol 20: 374–383. [DOI] [PubMed] [Google Scholar]
- 68. Borrego JJ, Luque A, Castro D, Santamaría JA, Martínez-Manzanares E (1996) Virulence factors of Vibrio P1, the causative agent of brown ring disease in the Manila clam, Ruditapes philippinarum. Aquat Living Resour 9: 125–136. [Google Scholar]
- 69. La Peyre JF, Xue Q-G, Itoh N, Li Y, Cooper RK (2010) Serine protease inhibitor cvSI-1 potential role in the eastern oyster host defense against the protozoan parasite Perkinsus marinus. Dev Comp Immunol 34: 84–92. [DOI] [PubMed] [Google Scholar]
- 70. Yu H, He Y, Wang X, Zhang Q, Bao Z, et al. (2011) Polymorphism in a serine protease inhibitor gene and its association with disease resistance in the eastern oyster (Crassostrea virginica Gmelin). Fish Shellfish Immunol 30: 757–762 10.1016/j.fsi.2010.12.015 [DOI] [PubMed] [Google Scholar]
- 71. Snyder MJ (2000) Cytochrome P450 enzymes in aquatic invertebrates: recent advances and future directions. Aquat Toxicol 48: 529–547 10.1016/S0166-445X(00)00085-0 [DOI] [PubMed] [Google Scholar]
- 72. Morga B, Arzul I, Faury N, Segarra A (2011) Molecular responses of Ostrea edulis haemocytes to an in vitro infection with Bonamia ostreae. Dev Comp Immunol 35: 323 10.1016/j.dci.2010.10.005 [DOI] [PubMed] [Google Scholar]
- 73. Brulle F, Jeffroy F, Madec S, Nicolas J-L, Paillard C (2012) Transcriptomic analysis of Ruditapes philippinarum hemocytes reveals cytoskeleton disruption after in vitro Vibrio tapetis challenge. Dev Comp Immunol 38: 368–376 10.1016/j.dci.2012.03.003 [DOI] [PubMed] [Google Scholar]
- 74. Ivanina AV, Cherkasov AS, Sokolova IM (2008) Effects of cadmium on cellular protein and glutathione synthesis and expression of stress proteins in eastern oysters, Crassostrea virginica Gmelin. J Exp Biol 211: 577–586 10.1242/jeb.011262 [DOI] [PubMed] [Google Scholar]
- 75. Quintana FJ, Cohen IR (2011) The HSP60 immune system network. Trends Immunol 32: 89–95. [DOI] [PubMed] [Google Scholar]
- 76. Lettre G, Hengartner MO (2006) Developmental apoptosis in C. elegans: a complex CEDnario. Nat Rev Mol Cell Biol 7: 97–108 10.1038/nrm1836 [DOI] [PubMed] [Google Scholar]
- 77. Lopez J, Meier P (2010) To fight or die – inhibitor of apoptosis proteins at the crossroad of innate immunity and death. Curr Opin Cell Biol 22: 872–881 10.1016/j.ceb.2010.08.025 [DOI] [PubMed] [Google Scholar]
- 78. Nitta T, Takahama Y (2007) The lymphocyte guard-IANs: regulation of lymphocyte survival by IAN/GIMAP family proteins. Trends Immunol 28: 58–65 10.1016/j.it.2006.12.002 [DOI] [PubMed] [Google Scholar]
- 79. Dower K, Ellis DK, Saraf K, Jelinsky SA, Lin L-L (2008) Innate immune responses to TREM-1 activation: overlap, divergence, and positive and negative cross-talk with bacterial lipopolysaccharide. J Immunol 180: 3520–3534. [DOI] [PubMed] [Google Scholar]
- 80. Sokolova IM (2009) Apoptosis in molluscan immune defense. Invertebr Surviv J 6: 49–58. [Google Scholar]
- 81. Donaghy L, Lambert C, Choi K-S, Soudant P (2009) Hemocytes of the carpet shell clam (Ruditapes decussatus) and the Manila clam (Ruditapes philippinarum): Current knowledge and future prospects. Aquaculture 297: 10–24 10.1016/j.aquaculture.2009.09.003 [DOI] [Google Scholar]
- 82. Hughes FM, Foster B, Grewal S, Sokolova IM (2010) Apoptosis as a host defense mechanism in Crassostrea virginica and its modulation by Perkinsus marinus. Fish Shellfish Immunol 29: 247–257 10.1016/j.fsi.2010.03.003 [DOI] [PubMed] [Google Scholar]
- 83. Sunila I, LaBanca J (2003) Apoptosis in the pathogenesis of infectious diseases of the eastern oyster Crassostrea virginica. Dis Aquat Organ 56: 163–170 10.3354/dao056163 [DOI] [PubMed] [Google Scholar]
- 84. Zhang G, Zhang L, Li L (2011) Gene discovery, comparative analysis and expression profile reveal the complexity of the Crassostrea gigas apoptosis system. Dev Comp Immunol 35: 603–610 10.1016/j.dci.2011.01.005 [DOI] [PubMed] [Google Scholar]
- 85. Infante RE, Wang ML, Radhakrishnan A, Kwon HJ, Brown MS, et al. (2008) NPC2 facilitates bidirectional transfer of cholesterol between NPC1 and lipid bilayers, a step in cholesterol egress from lysosomes. Proc Natl Acad Sci U S A 105: 15287–15292 10.1073/pnas.0807328105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Mohandas A, Cheng TC, Cheng JB (1985) Mechanism of lysosomal enzyme release from Mercenaria mercenaria granulocytes: A scanning electron microscope study. J Invertebr Pathol 46: 189–197 10.1016/0022-2011(85)90148-X [DOI] [PubMed] [Google Scholar]
- 87. Shimoyama Y, Hirohashi S, Hirano S, Noguchi M, Shimosato Y, et al. (1989) Cadherin Cell-Adhesion Molecules in Human Epithelial Tissues and Carcinomas. Cancer Res 49: 2128–2133. [PubMed] [Google Scholar]
- 88. Matussek A, Strindhall J, Stark L, Rohde M, Geffers R, et al. (2005) Infection of Human Endothelial Cells with Staphylococcus aureus Induces Transcription of Genes Encoding an Innate Immunity Response. Scand J Immunol 61: 536–544 10.1111/j.1365-3083.2005.01597.x [DOI] [PubMed] [Google Scholar]
- 89. Manoury B, Hewitt EW, Morrice N, Dando PM, Barrett AJ, et al. (1998) An asparaginyl endopeptidase processes a microbial antigen for class II MHC presentation. Nature 396: 695–699 10.1038/25379 [DOI] [PubMed] [Google Scholar]
- 90. Morita Y, Araki H, Sugimoto T, Takeuchi K, Yamane T, et al. (2007) Legumain/asparaginyl endopeptidase controls extracellular matrix remodeling through the degradation of fibronectin in mouse renal proximal tubular cells. FEBS Lett 581: 1417–1424. [DOI] [PubMed] [Google Scholar]
- 91. Marxen JC, Nimtz M, Becker W, Mann K (2003) The major soluble 19.6 kDa protein of the organic shell matrix of the freshwater snail Biomphalaria glabrata is an N-glycosylated dermatopontin. Biochim Biophys Acta BBA-Proteins Proteomics 1650: 92–98. [DOI] [PubMed] [Google Scholar]
- 92. Sarashina I, Yamaguchi H, Haga T, Iijima M, Chiba S, et al. (2006) Molecular evolution and functionally important structures of molluscan dermatopontin: Implications for the origins of molluscan shell matrix proteins. J Mol Evol 62: 307–318. [DOI] [PubMed] [Google Scholar]
- 93. Huang G, Liu J, Liu H, Han Y (2007) Profile of acute immune response in Chinese amphioxus upon Staphylococcus aureus and Vibrio parahaemolyticus infection. Dev Comp Immunol 31: 1013–1023 10.1016/j.dci.2007.01.003 [DOI] [PubMed] [Google Scholar]
- 94. Haag ES, Sly BJ, Andrews ME, Raff RA (1999) Apextrin, a Novel Extracellular Protein Associated with Larval Ectoderm Evolution in Heliocidaris erythrogramma. Dev Biol 211: 77–87 10.1006/dbio.1999.9283 [DOI] [PubMed] [Google Scholar]
- 95. Rosado CJ, Kondos S, Bull TE, Kuiper MJ, Law RHP, et al. (2008) The MACPF/CDC family of pore-forming toxins. Cell Microbiol 10: 1765–1774 10.1111/j.1462-5822.2008.01191.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Dheilly NM, Haynes PA, Bove U, Nair SV, Raftos DA (2011) Comparative proteomic analysis of a sea urchin (Heliocidaris erythrogramma) antibacterial response revealed the involvement of apextrin and calreticulin. J Invertebr Pathol 106: 223–229 10.1016/j.jip.2010.09.008 [DOI] [PubMed] [Google Scholar]
- 97. Khokha R, Murthy A, Weiss A (2013) Metalloproteinases and their natural inhibitors in inflammation and immunity. Nat Rev Immunol 13: 649–665 10.1038/nri3499 [DOI] [PubMed] [Google Scholar]
- 98. Dunn JR, Reed JE, Plessis DG du, Shaw EJ (2006) Expression of ADAMTS-8, a secreted protease with antiangiogenic properties, is downregulated in brain tumours. Br J Cancer 94: 1186 10.1038/sj.bjc.6603006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Egging D, Berkmortel F van den, Taylor G, Bristow J (2007) Interactions of human tenascin-X domains with dermal extracellular matrix molecules. Arch Dermatol Res 298: 389. [DOI] [PubMed] [Google Scholar]
- 100. Gaffen SL (2011) Recent advances in the IL-17 cytokine family. Curr Opin Immunol 23: 613–619 10.1016/j.coi.2011.07.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Roberts S, Gueguen Y, de Lorgeril J, Goetz F (2008) Rapid accumulation of an interleukin 17 homolog transcript in Crassostrea gigas hemocytes following bacterial exposure. Dev Comp Immunol 32: 1099–1104 10.1016/j.dci.2008.02.006 [DOI] [PubMed] [Google Scholar]
- 102. Chang C-I, Liao JC, Kuo L (1998) Arginase modulates nitric oxide production in activated macrophages. AJP - Heart Circ Physiol 274: H342. [DOI] [PubMed] [Google Scholar]
- 103. Villamil L, Gómez-León J, Gómez-Chiarri M (2007) Role of nitric oxide in the defenses of Crassostrea virginica to experimental infection with the protozoan parasite Perkinsus marinus. Dev Comp Immunol 31: 968–977 10.1016/j.dci.2007.01.006 [DOI] [PubMed] [Google Scholar]
- 104. Lang RP, Bayne CJ, Camara MD, Cunningham C, Jenny MJ, et al. (2009) Transcriptome Profiling of Selectively Bred Pacific Oyster Crassostrea gigas Families that Differ in Tolerance of Heat Shock. Mar Biotechnol 11: 650–668 10.1007/s10126-0009-9181-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Assembled transcriptome of American oysters in response to challenge with Roseovarius crassostreae.
(DOCX)