

Abstract
To determine whether ITS sequences in the rrn operon are suitable for identifying individual Acinetobacter Acb complex members, we analysed length and sequence differences between multiple ITS copies within the genomes of individual strains. Length differences in ITS reported previously between A. nosocomialis BCRC15417T (615 bp) and other strains (607 bp) can be explained by presence of an insertion (indel 13i/1) in the longer ITS variant. The same Indel 13i/1 was also found in ITS sequences of ten strains of A. calcoaceticus, all 639 bp long, and the 628 bp ITS of Acinetobacter strain BENAB127. Four additional indels (13i/2–13i/5) were detected in Acinetobacter strain c/t13TU 10090 ITS length variants (608, 609, 620, 621 and 630 bp). These ITS variants appear to have resulted from horizontal gene transfer involving other Acinetobacter species or in some cases unrelated bacteria. Although some ITS copies in strain c/t13TU 10090 are of the same length (620 bp) as those in Acinetobacter strains b/n1&3, A. pittii (10 strains), A. calcoaceticus and A. oleivorans (not currently acknowledged as an Acb member), their individual ITS sequences differ. Thus ITS length by itself can not by itself be used to identify Acb complex strains. A shared indel in ITS copies in two separate Acinetobacter species compromises the specificity of ITS targeted probes, as shown with the Aun-3 probe designed to target the ITS in A. pitti. The presence of indel 13i/5 in the ITS of Acinetobacter strain c/t13TU means it too responded positively to this probe. Thus, neither ITS sequencing nor the currently available ITS targeted probes can distinguish reliably between Acb member species.
Introduction
Acinetobacter spp. are important nosocomial pathogens and epidemiological survey data from around the world suggest that multi-antibiotic resistant Acinetobacter baumannii (genomic species 2), A. pittii (genomic species 3) and A. nosocomialis (genomic species TU13) are the most important clinically [1], [2], [3]. These three species were grouped with A. calcoaceticus (genomic species BG1), a soil organism, into the A. calcoaceticus-A. baumannii (Acb) complex because of their close genetic similarities [4], [2]. Based on DNA-DNA hybridization and ribotyping data, Gerner-Smidt and Tjernberg [5] proposed that two additional genomic species be added to the Acb complex, namely ‘close to 13TU’ and ‘between 1 & 3’. The taxonomic relationships of these strains to the other members of the Acb complex still have to be resolved [6].
Interest in using chemotaxonomic and molecular methods to distinguish between Acb complex members has increased, as phenotypic characters have generally proved to be unreliable for this task. Many such methods have been described [7], and include exploiting the sequence variations in the 16S–23S rRNA gene internal transcribed spacer region (ITS sequences) in the rrn operons [8]. In their analyses of Acinetobacter ITS regions, Chang et al. [9] suggested those of individual Acb member species were highly conserved in both their lengths and sequences. As intra-genomic ITS sequence variance had never been confirmed then in member species of this group, they proposed that the direct sequencing of Acb ITS sequences provided a promising method for their identification [9]. Although all Acinetobacter spp. are known to possess multiple ITS copies [10], [11], ITS targeted probes have been designed to identify Acb members [12]–[17] on the premise that the extent of Acb intra-species ITS copy sequence variance is low.
Methods based on ITS features include using multiplex PCR [13], oligonucleotide arrays [14], [15], oligonucleotide hybridisation probes [16], and microsphere-based arrays [17]. While nine of 12 currently available Acb probes have been claimed to give unequivocal identifications, the remaining three published ITS-targeted probes are more problematic. The A. nosocomialis-targeted P-13 probe [17] cross hybridized with the ITS sequences of some A. pittii strains, while the A. pittii-targeted Aci3 [16] and P-3 [17] probes cross hybridized with the ITS sequences of some A. nosocomialis strains. An examination of these three probe sequences against the ITSs of the type strain of A. nosocomialis (BCRC 15417T) and A. pittii (BCRC 15420T) revealed such cross hybridisation events should not have occurred.
The known ITS lengths of most Acb members appear to be highly conserved with A. calcoaceticus (6 strains) all 638 bp; A. baumannii (27 strains) all 607 bp; A. pittii (6 strains) all 619 bp [9]. However, while the majority of the publicly available ITS sequences of the A. nosocomialis strains are 607 bp, those of the type strain A. nosocomialis BCRC15417T are 615 bp (Table 1). Furthermore, the differences in sequence between the 607 bp and 615 bp A. nosocomialis ITSs must be substantial as such a small (8 bp) length difference between otherwise identical ITS >600 bp would give a similarity score of >98%, and not the reported 95% [18]. Whether this variance is the result of point mutations or recombination events has never been examined.
Table 1. ITS of A. calcoaceticus, A. baumannii, A. pittii and A. nosocomialis strains downloaded from GenBank and their ITS length (bp).
| Acinetobacterstrain | AccessionNumber | ITSLength | Author | ||
| A. nosocomialis | (1) | 00574 | AY510070 | 607 bp | Chang et al, 2005 [9] |
| v104-2 | AY510071 | 607 bp | Chang et al, 2005 [9] | ||
| DR25612/96 | EU030649 | 607 bp | Koh et al, unpublished | ||
| DR32226/96 | EU030656 | 599 bp(2) | Koh et al, unpublished | ||
| DB30014/96 | EU030653 | 599 bp(2) | Koh et al, unpublished | ||
| DM18619/96 | EU030654 | 572 bp(2) | Koh et al, unpublished | ||
| 74510 | FJ360743 | 607 bp | Zarrilli et al, 2009 [18] | ||
| Ab22222T(1) | AKAR00000000 | 608 bp | Murphy et al, unpublished | ||
| (2) | BCRC15417T | AY601830 | 615 bp | Chang et al, 2005 [9] | |
| BCRC15417T | U60281 | 609 bp(2) | Lagatolla et al, 1998 [16] | ||
| NCTC 8102(1) | AIEJ00000000 | 615 bp | Chan et al, 2012 [30] | ||
| Ab22222(1) | AKAR00000000 | 616 bp | Murphy et al, unpublished | ||
| TG21145(1) | AMJH00000000 | 615 bp | Sahl et al, unpublished | ||
| A. calcoaceticus | (3) | ATCC 23055 | U60278 | 628 bp(2) | Lagatolla et al, 1998 [16] |
| LMG 1046 | AY601820 | 638 bp | Chang et al, 2005 [9] | ||
| LMG 992 | AY601821 | 638 bp | Chang et al, 2005 [9] | ||
| BCRC 11562 | AY601822 | 637 bp | Chang et al, 2005 [9] | ||
| IH9 | FJ786266 | 629 bp(2) | Peix et al, 2009 [24] | ||
| OCI1 | FJ786267 | 632 bp(2) | Peix et al, 2009 [24] | ||
| PHEA-2(1) | NC_016603 | 620 bp | Zhan et al, 2011 [44] | ||
| A. baumannii | (4) | BCRC10591 | AY601823 | 607 bp | Chang et al, 2005 [9] |
| BCRC15884 | AY601824 | 607 bp | Chang et al, 2005 [9] | ||
| BCRC15886 | AY601825 | 607 bp | Chang et al, 2005 [9] | ||
| LMG984 | AY601826 | 607 bp | Chang et al, 2005 [9] | ||
| ATCC 17978(1) | CP000521 | 608 bp | Smith et al, 2007 [45] | ||
| ACICU(1) | CP000863 | 608 bp | Iacono et al, 2008 [46] | ||
| AYE(1) | CU459141 | 608 bp | Vallenet et al, 2008 [47] | ||
| SDF(1) | CU468230 | 608 bp | Vallenet et al, 2008 [47] | ||
| BCRC10591 | U60279 | 610 bp | Lagatolla et al, 1998 [16] | ||
| DM19034/01 | EU030648 | 607 bp(2) | Koh et al, unpublished | ||
| DU32993/01 | EU030651 | 607 bp(2) | Koh et al, unpublished | ||
| DM169/96 | EU030652 | 601 bp(2) | Koh et al, unpublished | ||
| DU54004/05 | EU030657 | 607 bp | Koh et al, unpublished | ||
| DM19305/01 | EU030660 | 602 bp(2) | Koh et al, unpublished | ||
| DU5377/05 | EU030658 | 601 bp(2) | Koh et al, unpublished | ||
| DB6474/05 | EU030659 | 607 bp | Koh et al, unpublished | ||
| DU16891/96 | EU030661 | 607 bp | Koh et al, unpublished | ||
| DB15354/07 | EU030662 | 601 bp(2) | Koh et al, unpublished | ||
| DB34441/07 | EU030663 | 600 bp(2) | Koh et al, unpublished | ||
| 25001 CMCC(B) | DQ108593 | 608 bp | Chen et al, 2007 [13] | ||
| 29108 CMCC(B) | DQ108594 | 608 bp | Chen et al, 2007 [13] | ||
| A. pittii | (5) | LMG1035T | AY601827 | 619 bp | Chang et al, 2005 [9] |
| CCUG26384 | AY601828 | 619 bp | Chang et al, 2005 [9] | ||
| BCRC15420 | AY601829 | 619 bp | Chang et al, 2005 [9] | ||
| BCRC15420 | U60280 | 622 bp | Lagatolla et al, 1998 [16] | ||
| DB60079/01 | EU030647 | 615 bp(2) | Koh et al, unpublished | ||
| DM22501/01 | EU030650 | 595 bp(2) | Koh et al, unpublished | ||
| DM21785/01 | EU030655 | 619 bp | Koh et al, unpublished | ||
| SH024 | NZ_GG753607 | 571 bp(2) | Peleg et al, 2012 [48] | ||
| DSM 9306(1) | AIEF00000000 | 620 bp | Chan et al, 2012 [30] | ||
| D499(1) | AGFH00000000 | 620 bp | Chen et al, 2012 [49] | ||
| DSM 21653(1) | AIEK00000000 | 620 bp | Chan et al, 2012 [30] | ||
| TG6411(1) | AMJI00000000 | 620 bp | Sahl et al, unpublished | ||
| A. oleivorans | DR1(1) | NC_014259 | 620 bp | Jung et al, 2010 [50] |
Whole genome sequences (from which all ITS were extracted in silico).
Partial sequence.
ITS sequences downloaded from GenBank were analysed using Geneious Pro (percentage similarity calculations not shown; accession numbers listed).
No instances of ITS allele sequence variance have been demonstrated for any member of this Acb complex. However, the unusual nature of the A. nosocomialis BCRC15417T ITS, together with the low specificities of some published Acb ITS targeted probes suggested that a more thorough examination of the ITS of Acb members’ was required. The outcomes are reported here.
Materials and Methods
Strains sequenced and source of ITS sequences
The ITS sequences were analysed from the following strain collections: Collection (A): consisted of strains whose ITS sequences were downloaded as individual sequences, or extracted from whole genome sequence data, with the GenBank accession numbers shown in Table 1. The ITS of type strains sequenced by Chang et al. [9] were designated by them as A. calcoaceticus LMG1046T, A. baumannii BCRC10591T and A. pittii LMG10350T, while the same strains sequenced here are designated as A. calcoaceticus ATCC23055T, A. baumannii ATCC19606T and A. pittii ATCC19004T (Table 1).
Collection (B): included were Acinetobacter strains in culture collections from which cloned ITS PCR amplicons were generated and sequenced here. They contained strains from three Acinetobacter species and two genomic species included in or closely related to the Acb complex. These were A. calcoaceticus strains ATCC23055T, 97366, 97420, 97424; A. baumannii strains ATCC19606T, 97429, 97434, 16842; A. pittii ATCC19004T; ‘close to 13TU’ strains 5804 and 10090 (hereafter referred to as c/t13TU 5804 and c/t13TU 10090); ‘between1&3’ strains 10095 and 10169 (hereafter referred to as b/n1&3 10095 and b/n1&3 10169); Acinetobacter BENAB127. Thirteen of these Acb strains were speciated by DNA-DNA hybridization [19], [20], [5], and 11 were isolated from clinical samples, as detailed by Maslunka et al. [10]. The exceptions were A. calcoaceticus strains 97366 and 97420, both isolated from soil, and Acinetobacter strain BENAB127, which was isolated from activated sludge. While Beacham et al. [21] identified BEN127 phenotypically as A. pittii, analysis of its 16S rRNA (data not shown: Accession number HE651911) and ITS sequences (see later) suggest it cannot be placed confidently in this species. Consequently, it is referred to here as Acinetobacter BENAB127.
Collection (C): These forty-one clinical isolates were ‘identified’ tentatively by pulsed field gel electrophoresis (PFGE) and riboprinting as either A. baumannii (7–8, 147–150, 153–8, 160–1, 163–8, 170–1, 176–8, 180–1, 248–9, 251, 253–7, 262) or A. calcoaceticus (250, 175, 151, 159, 169) at the Austin Hospital Microbiology Department, Melbourne, Australia. 16S rRNA sequencing performed in this study (data not shown) revealed that only five strains (7, 8, 147, 148, & 150) were considered to belong to A. baumannii. Strains 151, 159, 169, and 175 and 250 did not appear to be members of either of these species. Instead, 16S rRNA sequences of strains 151, 159 and 175 were most similar to those of A. pittii and A. oleivorans, while those of strains 169 and 250 clustered most closely to Acinetobacter b/n1&3 strain 10095 and A. calcoaceticus respectively (data not shown). The remaining 31 were considered most likely to belong to A. nosocomialis, a conclusion supported by ITS sequencing data (see later). The ITS sequences of all these were generated by direct sequencing as described next (accession numbers given below).
DNA extraction
Cells of each collection (B) strains were grown on R2A agar at 30°C. Genomic DNA was extracted using an UltraClean Soil DNA extraction kit (Mo Bio Laboratories Inc.) and stored in 10 mM Tris buffer at −20°C.
Direct sequencing of the ITS
ITS sequences were PCR amplified by adding genomic DNA to a reaction mixture containing the 1512f (GTC GTA ACA AGG TAG CCG TA) and 6r (GGG TTY CCC CRT TCR GAA AT) bacterial universal primers [9]. This protocol generated ITS amplicons containing 16S and 23S rDNA flanks of approximately 18 bp and 115 bp respectively. A GeneAmp PCR 2400 thermocycler (Applied Biosystems) was used, with initial denaturation at 94°C for 6 min, denaturation at 94°C for 30 sec, annealing at 53°C for 1 min, extension at 72°C for 30 cycles of 90 sec each and a final extension at 72°C for 10 min. Amplicons were cleaned using an Ultra-Clean PCR Cleanup Kit (Mo Bio Laboratories Inc.).
Product purity was checked by electrophoresis with 2% agarose gels stained with ethidium bromide, which were viewed with a UV transilluminator. DNA concentrations were calculated using Adobe Photoshop 7 to compare the band intensity of each ITS amplicon against that of similarly sized 2 Log ladder band (New England Biolabs) of known DNA concentration. Primer 1512f was used for forward reads while primer 6r was used for reverse reads. Samples were sequenced by the Australian Genome Research Facility (AGRF) (Brisbane, Australia).
ITS sequencing by colony PCR
Ligation (pGem-T Easy Vector kit) and transformation were carried out as described by Carr et al. [22]. Cells from individual colonies were transferred from the growth medium (LB plates with ampicillin/IPTG/X-gal) and resuspended in 10 mM Tris buffer. Cells were collected from clones for each strain, and shaken gently for 60 min before being stored at 4°C. The M13f (CGC CAG GGT TTT CCC AGT CAC GAC) and M13r (TCA CAC AGG AAA CAG CTA TGA C) primers were used to amplify the ITS sequence as above, but with denaturation occurring at 95°C for 30 sec. PCR products were cleaned, checked for purity on 2% agarose gels and prepared for sequencing as described earlier.
ITS sequence accession numbers
All sequences generated in this study were deposited in the DDBJ/EMBL/GenBank database (at EMBL-EBI, European Molecular Biology Laboratories, http://www.ebi.ac.uk/embl/) as follows: Strain collection (B). A. calcoaceticus strains, HE651573–HE651628 and HE651903–HE651906; A. baumannii strains, HE651629–HE651686 and HE651907–HE651910; A. pittii ATCC19004T, HE651687–HE651696 and HE651911; c/t13TU 5804 and c/t13TU 10090, FN669512–FN669527 and FN677952–FN677963; b/n1&3 10095 and b/n1&3 10169, HE651546–HE651572, HE651912 and HE651913; Acinetobacter BENAB127, HE651697–HE651710 and HE651914. Strain collection (C) strains 41 directly sequenced clinical isolates; 7–8, KC257006–7; 147–151, KC257008–12; 153–161, KC257013–21; 163–171, KC257022–30; 175–178, KC257031–34; 180–181, KC257035–36; 248–251, KC257037–40; 253–257, KC257041–45; 262, KC257046.
In silico analysis of ITS sequences
Geneious Pro (version 4.0.4: www.geneious.com) bioinformatics software (Biomatters Ltd) was used to construct ITS sequence alignments, contigs and consensus sequences. Similarity matrices were constructed with MegAlign (DNASTAR software, version 7.1) using a ClustalW alignment. Only a single ITS sequence was publicly available for most Acinetobacter species, including those with variable length ITS sequences [9].
Assessing A. pittii ITS targeted Aun3 probe specificities
ITS amplicon generation was performed with the same PCR protocol as described above, except that a 70 sec extension time was applied, and probe Aun3 [14] (GAT GAA GAA TCG CAC GGA CAA CA) as reverse primer. PCR products were separated on a 2% agarose gel stained with ethidium bromide.
Results
ITS sequence isolation and analysis
Organizational properties of 139 Acb ITS sequences with one or more distinguishing features are shown in Figure 1. When genomic DNA was used as PCR template for the Acinetobacter strains in strain collections (B) and (C), electrophoretic mobilities for ITS amplicons (approximately 800 bp long, data not shown) from A. calcoaceticus ATCC23055T, A. baumannii ATCC19606T and A. pittii ATCC19004T matched exactly those reported for these same strains by Chang et al. [9]. Furthermore, with total genomic DNA as PCR template, direct ITS sequencing of strains in collection (B) was successful for every strain except c/t13TU 10090 (discussed below), and their ITS sequences derived by this approach were identical to those derived from sequencing multiple individual clones of the same strains (see below).
Figure 1. Sequence properties of ITS Acinetobacter Acb member strain sequences with one or more distinctive properties.

ITS-Type, ITS-Subtype, Indel Subtype and Indel Sequence Subtype defined in Results. ITS sequences were plotted with species on y-axis with established species in bold and collection C isolates in regular type. Figure 1A shows their individual ITS sequence lengths, Figure 1B the number of ITS sequences of a distinctive length (black) or sequence (grey) and Fig. 1C the number of indels (see Results) within each ITS of a distinctive length. The Acb species and strains associated with each ITS sequence length, number, sequence and indel type are given below. On x-axis: (A) ITS-lengths (named ‘Type’ and symbols refer to: * A. c/tTU13; § A. calcoaceticus; † A. nosocomialis); (B) Number of ITS-Types and -SubTypes (ITS copies of same sequence lengths but different sequences). Black/grey bars (parallel to green bar in A) show 40 ITS Types and 5 SubTypes of A. nosocomialis 607 bp ITS; black and grey bars (parallel to purple bar in A) show 5 ITS Types and 5 SubTypes of A. nosocomialis 615 bp ITS; white lines show number of each SubType; (C) Number of specific Indel Subtypes and Sequence Subtypes as totals for each strain are shown by coloured bars. See Fig. 3 for sequences; each indel subtype name refers to strains with 13i/1 and 13i/4 indels. 13i/5 53/54 refers to homology in 53/54 nucleotides, while 13i/5 54/54 indicates sequence homology for all 54 nucleotides). ‡ Refers to A. baumannii strains in Table 1 and M&M, isolates 147–8, 7; A. nosocomialis 607/8 bp, 00574, v104-2, DR25612/96, DR3226/96, DM18619/96, 74510, AB22222, isolates 149, 153–6, 160, 163–8, 170–1, 176–8, 180–1, 248–9, 251–7; A. nosocomialis 615/6 bp, TG21145, BCRC15417T, AB22222, NCTC8102, isolate 157; A. pittii, DSM9306, D499, DSM21653, TG6411, SH024. * Property defined as: ‘Species’, ‘Strain’, ‘Type’, ‘SubType’, ‘Indel’, ‘Indel sequence subtype’, ‘rrn allele name’. ¶ With ITS sequence Types of different lengths (608 to 638 bp), depending on their length and/or sequence, between 90–100% of the sequence is homologous, allowing for precise alignment. The alignment program introduces indels into regions not homologous with high numbers of other sequences in these regions, depending on ITS sequence Type length. Details in Gürtler & Grando [42] and Gürtler et al [43].
After 16S and 23S rRNA gene flanks were removed, the resultant ITS consensus sequence lengths were as follows: 638 bp/639 bp, for all sequenced A. calcoaceticus strains; 608 bp, for all sequenced A. baumannii strains; 620 bp, for A. pittii ATCC19004T; 608 bp, for strain c/t13TU 5804; 620 bp, for both ‘b/n1&3’ strains; and 628 bp, for Acinetobacter BENAB127. All these ITS consensus sequences contained a tRNAile gene (position 61/62–137/138) and tRNA-ala gene (position 193/194–268/269) as shown in Figure 2.
Figure 2. Sequence alignments of distinctive ITS subtypes from ITS sequences of Acb members (see Figure 1).

Indels labeled as in Figure 1. Nucleotide substitutions [grey bars, homologous regions; vertical lines, red (T), green (A), black (G), blue (C), grey (N)]. For each indel, the closest Blast match is listed below its name. Sequences given only numbers are those of the collection C clinical isolates. The number of ITS sequences used in each consensus sequence is given in parentheses [ ]. The phylogenetic tree was constructed by Neighbor-Joining with 100 boot straps and numbers at nodes show percentage confidence of respective branches. Branch lengths are proportional to number of nucleotide substitutions but make no allowance for deletions (shown in white on the alignment).
Direct sequencing of the ITS sequence of strain c/t13TU 10090 generated sequence data that suggested amplicon contamination, where read quality deteriorated markedly downstream of the first 268 bp. As with the strain c/t13TU 5804 ITS sequence, tRNAile and tRNAala genes were located at positions 61 to 137 and 193 to 268 respectively. Subsequent cloning and sequencing revealed these initial results were not the result of contamination, but rather by the presence of multiple variable-length ITS sequences within the c/t13TU 10090 genome (discussed below).
Clone sequencing was performed on PCR products amplified from genomic DNA isolated from strain collection (B) strains, using between 10 and 15 clones for each strain. Apart from strain c/t13TU 10090, the degree of similarity between ITS sequences of individual clones for each strain was between 99.5 and 100%. Occasional single nucleotide polymorphisms (SNPs) were present in some ITS Types (see Figure 1 legend for definition). These are not considered to be sequencing artefacts since they were located repeatedly in the same positions in multiple replicate sequences of cloned ITS fragments generated here, and/or in retrieved deposited whole genome or ITS sequence data downloaded from GenBank. Furthermore, for the Acb strains whose ITS sequences were sequenced both directly and using colony PCR (see Materials and Methods), the similarities between the clone-based ITS consensus sequences and those derived from direct sequencing were always 100% [data not shown: accession numbers listed above].
The greatest number of Subtypes (defined in Figure 1 legend) were seen in A. baumannii (12 Subtypes), A. nosocomialis (5 Subtypes) and A. pittii (8 Subtypes), (Figures 1 & 2).
Features of A. nosocomialis ITS sequences
When ITS sequences from 12 A. nosocomialis collection A strains were downloaded from GenBank (see Table 1) and aligned, the sequences separated into one of two distinct Types (Figures 1 & 2): Type 607 (Group 1) and Type 615 (Group 2). Group 1 ITS sequences were all 607 bp and 99.7% similar to each other, while Group 2 sequences from the A. nosocomialis BCRC15417T 615 bp ITS sequences generated here were 99.8% similar to each other (Table 1). The whole genome sequence of A. nosocomialis (strain AB22222) possessed three copies of Group 1 and one copy of Group 2 ITS sequences, while A. nosocomialis strains NCTC8102 (the same strain as BCRC15417T) and TG21145 contain only the Group 2 615 bp ITS variant. Consensus sequences were generated for ITS sequences of each of these Groups, and are referred to subsequently as A. nosocomialis (Group 1) and A. nosocomialis (Group 2) ITS respectively. The degree of similarity between these Group 1 and 2 ITS consensus sequences was only 96.4%.
During this study, no A. nosocomialis ITS sequences were sequenced by clone analysis, but ITS sequences homologous to Group 1 (Type 607, Subtype A. nosocomialis) were found in the sequenced collection (C) isolates, considered likely to be strains of A. nosocomialis from the 16S rRNA sequence data discussed above (strains 149, 153–6, 160, 163–8, 170–1, 176–8, 180–1, 248–9, 251–7). On the other hand, the Group 2 (Type 615, Subtype A. nosocomialis) ITS was seen only in strain 157 from collection (C) isolates (see Figure 1A and Figure 1 legend).
Sequence analyses revealed a 37 bp region (TAA CAA AGA GAG ATG AAG TAA TTC TGA TCT TGG AGT T) distinguished the ITS sequences of ITS Group 1 (607 bp) from those of all Group 2 ITS (615 bp) A. nosocomialis. This 37 bp indel sequence is designated as indel 13i/1, and is present in the same location in the ITS of all Group 2 strains (Figure 3, position 436–472). Blast analysis showed its sequence is 91.9% similar to a region in the ITS sequence of A. ursingii LMG19575T, and 89.8% similar to a comparable region in the A. calcoaceticus ITS consensus sequence (Figures 2 & 3).
Figure 3. ITS consensus sequence alignments of Acb members including c/t13TU and Acinetobacter b/t 1 & 3 strains.

For each indel, coloured bars are shown below and behind each sequence in the alignment, and closest Blast match listed below its name. Number of sequences used for each in [ ]. Sequences conserved for all species are shown as grey bars, and single nucleotide differences within as vertical lines coloured red (T), blue (C), green (A), black (G) and grey (N). Numbers at branch nodes are % consensus support using HKY genetic distance, neighbor joining tree and bootstrap resampling 100 replicates. A. calcoaceticus (pink), A. baumannii (yellow), A. pittii (green) and A. nosocomialis (blue) sequences are labelled BG1, BG2, BG3 and 13TU respectively.
Features of Acinetobacter c/t13TU strains 5804 and 10090 ITS sequences
No publicly available ITS sequences were available for strains c/t13TU 5804 and 10090. Data generated here showed that the ITS of strain c/t13TU 5804 were all 608 bp in length, and an ITS consensus sequence (from 11 clones, 99.8% similarity) was 98.8% similar to that of the A. nosocomialis Group1 (607 bp) ITS consensus sequence (Figure 2, Figure S1).
Alignment of ITS sequences of 16 strain c/t13TU 10090 clones revealed their lengths were more diverse than the single length 608 bp ITS detected in strain c/t13TU 5804. Thus, they fell into five distinct Types (Figure 1A, 609, 608, 620, 621, 630, denoted by an asterisk) of 608 bp (4 identical clones, and 99.1% similar to the 608 bp ITS sequence in strain 5804), 609 bp (4 clones, 99.9% similarity, and 99.5% similar to the 608 bp ITS in strain 5804), 620/1 bp (5 clones, 99.5% similarity) and 630 bp (3 identical clones), with an overall sequence similarity of only 95.8% (Figures 1 & 2, Figure S1).
A consensus sequence for each c/t13TU 10090 clone collection was then generated, but clearly this information can not be used directly for ITS-based identification, as the results obtained would depend upon which ITS Group consensus sequence was used. Thus, the c/t13TU 10090 608 bp, 609 bp, 620/1 bp and 630 bp ITS consensus sequences were 98.7%, 98.7%, 93.4% and 93.0% similar respectively to the A. nosocomialis Group 1 ITS consensus sequence (Figure S1), while the c/t13TU 10090 620 bp ITS consensus sequence was 99.4% similar to that of the A. pittii 620 bp ITS consensus sequence, as discussed below (Figure S1). Sequence data revealed that variability between the ITS sequences of the c/t13TU 10090 clone groups did not result from accumulated single nucleotide polymorphisms, but rather from the presence of four indels, designated here as indel 13i/2 to 13i/5 respectively (Figures 1, 2 & 3).
Blast analyses of these indels showed the closest matching sequences of indels 13i/3, 13i/4 and 13i/5) are found in the ITS sequences of other Acinetobacter species (Figures 1, 2 & 3). They also revealed that indel 13i/2 (19 bp: GTA GAT AAA AAG ATA CAT G ) is absent from the ITS sequences of all other Acinetobacter isolates so far examined (Maslunka, unpublished data). Instead, its sequence is identical to a region within the AarF gene of Thermoanaerobacter tengcongensis strain MB4 (position 1,187,443 to 1,187,462) (Figure S1), and it shares 18 of 19 nucleotides with a region in an undetermined gene in the genomes of Bacillus cereus strains Q1, AH187 and E33L (Q1 locus tag: BCQ_2928).
While indel 13i/3 (2 bp, underlined: ATT AAT CTA GAT GAA TTG) was too short to Blast, alignment data showed this same adjacent pair of nucleotides (A and G) and conserved flanking regions occur at the same locale in the A. baumannii ITS consensus sequence, and ITS sequences of all A. pittii strains so far sequenced. It is also present in the 620 bp ITS sequence of Acinetobacter b/n1&3 strains 10095 and 10169 (Figures 1, 2 & 3). The only available ITS sequence for the non-Acb member A. radioresistens BCRC15425T also contains this indel sequence at the same locale (Figures 1, 2 & 3).
Indel 13i/4 is 75 bp long, and appears to have arisen from two recombination events. A Blast alignment of the first 43 nucleotides shows that all known ITS sequences of A. calcoaceticus strains have a homologous region at the same locale (83.7% to 86% similarity) (Figures 1, 2 & 3). The final 32 nucleotides are 93.9% similar to a region within Acinetobacter genomic species BJ14 CCUG34435T ITS at a similar locale (Figures 2 & 3). Importantly, the first 43 nucleotide region of the A. calcoaceticus ITS sequence and the 32 nucleotide Acinetobacter BJ14 CCUG34435T ITS region share a five nucleotide overlap (CAT TG) (Figure 3).
Indel 13i/5 is 54 bp long. It has a sequence identical to a comparable region in five of the seven A. pittii ITS sequences available in GenBank, and is 98.2% similar to those in the ITS sequence of A. pittii strains BCRC15420T and DM21785–01. Its presence makes the Acinetobacter c/t13TU 10090 620 bp ITS sequence copies which carry indel 13i/5 essentially identical to the ITS sequences of all six A. pittii strains examined here (99.4% average similarity) (Figures 2 & 3), with only three nucleotides distinguishing the strain c/t13TU 10090 620 bp ITS sequence carrying indel 13i/5 from the A. pittii ITS consensus sequence (Figure 3, positions 177, 316, 318). The position 177 ‘A’ nucleotide is unique to the ITS sequence of strain c/t13TU 10090, and could conceivably be used to differentiate it from A. pittii by methods like High Resolution Melt-PCR (HRM-PCR) [23]. However, the latter nucleotide pair (AGA GAT TTC GG ) are of limited use for identification purposes as they are also found in the ITS sequences of all known strains of A. nosocomialis, as well as those of Acinetobacter c/t13TU 5804 (Figures 2 & 3).
Thus, there appear to be at least four different ITS sequence Types within the rrn operons of the genome of Acinetobacter c/t13TU strain 10090. The exact number remains to be determined by whole genome sequencing, although it can be concluded that the variations seen here in its ITS sequences share homology with those of A. calcoaceticus (Figure 2 red background: 608 and 609 bp) and A. pittii (see below) (Figure 2 green background: indel 13i/5, 621 bp).
Features of A. baumannii ITS sequences
A. baumannii ITS sequences from 27 strains from GenBank (Table 1) and eleven whole genome sequences (Table 1) were aligned against those from 55 ITS clones from A. baumannii strains ATCC19606T, 97429, 97434, 16842 generated here. A single ITS consensus sequence was compiled. All 132 ITS sequences generated were 608 bp long and shared 99.7% homology. They represent 12 ITS Subtypes (See Figures 1 & 2, yellow background), with each differing by only a single nucleotide. The ITS sequences from A. baumannii are the shortest among the Acb members (608 bp), and do not contain the indels 13i/1, 13i/2, 13i/4 or 13i/5 (Figures 1 & 2).
Collection C clinical isolates 7–8, 150, 147–8 have the same ITS sequence Type and Subtype (608) as A. baumannii. Although those from isolates 8 and 150 differed from the other twelve A. baumannii subtypes by a single nucleotide (Figures 1 & 2), they would still fall into the A. baumannii Type (608) Group. Hence these data might suggest that collection C strains 7–8, 150 and 147–8 were probably identified correctly by 16S rRNA sequencing and pulse field electrophoresis as A. baumannii.
Features of A. pittii ITS sequences
ITS sequences downloaded from GenBank from 10 A. pittii strains (single ITS sequence (Table 1), were aligned against those from 10 ITS clones generated here from A. pittii ATCC19004T. A single consensus sequence was generated as each ITS was 620 bp (Type 620) and they shared 99.7% sequence similarity. Four different sequence ITS Subtypes were detected within the genome of A. pittii strain D499, and a further eight were present in five other A. pittii strains (see Figure 1). Both A. calcoaceticus NC016603 and the Acb complex non-member species A. oleivorans DR1 share the same ITS sequence Type as A. pittii (620), but they differ in their sequence Subtypes by single nucleotide differences (Figures 1 & 2). Importantly indel 13i/5 sequence (Subtype 54/54) was present in all A. pittii ITS sequences examined here, including the complete rrn operon set from the whole genome of A. pitti D449 (Figure 1 & 2).
Features of A. calcoaceticus ITS sequences
The A. calcoaceticus ITS sequences fell into two sequence lengths of 620 bp (Type 620) and 639 bp (Type 639) (Figure 1). Sequences from six A. calcoaceticus strains from GenBank (Table 1) were aligned against ITS sequences generated from clone sequencing by colony PCR of A. calcoaceticus strains ATCC23055T, 97366, 97420, and 97424. All were 639 bp and 99.5% similar to each other. ITS sequence Type (620) only included the two alleles extracted from the rrn loci of the whole genome sequence of A. calcoaceticus BG1 NC016603.
Again these ITS length differences arose from differences in their indel content. Indels 13i/1 Subtype BG1 and 13i/4 Subtype BG1 were present in all A. calcoaceticus Type (639) ITS sequences (Figures 1, 2 & 3). On the other hand, indel 13i/5 Subtype 54/54 occurred in all A. calcoaceticus Type (620) ITS sequences and those from collection (C) clinical isolates 151 and 159, but not in any of Type (639) ITS sequences (Figures 1 & 2). It may be that Type (620) and (639) ITS and their associated indels are mutually exclusive in A. calcoaceticus, a view substantiated by the observation that whole genome sequence data from strain A. calcoaceticus PHEA-2 with ITS Type (620) and associated indels, lacked ITS Type (639)
Features of Acinetobacter b/n1&3 Strains 10095 and 10169 ITS sequences
No publicly available ITS sequences were available for Acinetobacter b/n1&3 strains 10095 and 10169. Therefore, all the ITS sequences reported here were obtained by clone sequencing from strains in collection (B).
The 620 bp ITS consensus sequences of Acinetobacter b/n1&3 10095 (from 12 clones, 99.9% similarity) and b/n1&3 10169 (from 13 clones, 99.9% similarity) were 99% similar to each other. Furthermore, ITS sequences from both strains possess the same indel 13i/5 Subtype (Red, 53/54) as the six A. oleivorans DR1 ITS alleles (each identical to each other). (Figures 1 & 2; Table 1). The 13i/5 sequences and hence ITS from the two b/n1&3 strains have a different indel 13i/5 Subtype (Red, 53/54) to that seen in the ITS of A. pittii ATCC19004T and A. calcoaceticus NC016603. As described above, while A. pittii ATCC 19004T and A. calcoaceticus NC016603 possess indel 13i/5 (Subtype 54/54) (Purple, 54/54: Figures 1 & 2), their ITSs differ from those of b/n1&3 strains (Subtype 53/54) by one and three nucleotides respectively (Figures 1 & 2). On this basis, both b/n1&3 strains emerge more closely related to A. oleivorans DR1 than to either A. pittii ATCC19004T or A. calcoaceticus NC016603, even though A. oleivorans is not recognised currently as a member of the Acb complex.
Features of other Acinetobacter ITS sequences from closely related strains
The Acinetobacter strain BENAB127 628 bp ITS consensus sequence derived from 13 identical clones was most closely related to that of A. pittii strain 19004T (96.1% similar). It too contained indel 13i/5 SubType 53/54 (Figures 1 & 2) as do the ITS of Acinetobacter b/n1&3 strains 10095 and 10169 discussed above (96.3% and, 96.7% similarity respectively). Furthermore, the Acinetobacter BENAB127 ITS consensus sequence contains a variant of indel 13i/1 (Subtype BENAB) which has four single nucleotide substitutions compared to that of indel 13i/1 present in the A. calcoaceticus ITS consensus sequence (Figure 2, red background).
Analysis of the clone derived Acinetobacter BENAB127 ITS sequences would suggest the presence of only one ITS sequence Type (Type 628) although its rrn operon number is not known. Thus, it may be another closely related species to members of the A. pittii clade, but its ITS contains both indel 13i/1 (Subtype BENAB) (Figure 1C, light green) and indel 13i/5 Subtype 53/54 (Figure 1C, red), a combination not found in any other Acb strain ITS so far sequenced (Figures 2, 3 & 4).
Figure 4. Alignment of ITS sequences of Acb members showing oligonucleotide probe target sites.

These data show that existing oligonucleotide probes differentiate between the 620/1 bp ITS sequences of c/t13TU 10090, and those of genomic species A. pittii (BG3) and Acinetobacter b/n1&3 strains. Currently twelve probes have been designed to differentiate between Acb complex members: probes Acal, Abau and Aun3 [14]; probes Aci2 and Aci3 [16]; probes P-1, P-2, P-3, P-13 [17]; the two A. baumannii-targeted probes designed by Chen et al. [13] have not been designated names.
ITS sequences from collection (C) strains 151, 159 and 169, which could not be placed into any prexisting Acb complex species by 16S rRNA sequence analyses, were most closely related to those from A. pittii (98.9%), c/t13TU (99.5–99.7%) and A. oleivorans (99.5%) and b/n 1&3 strains 10169 and 10095 (99.5%) respectively, all of which had ITS lengths of 620/622 bp. As mentioned above, the ITS of strains 151 and 159 both contain the indel 13i/5 subtype 54/54, while those of strains 159 and 169 contain indel 13i/3 and13i/5 subtype 53/54.
With collection (C) strains 175 and 250, their ITS sequences were most closely related to those of the A. calcoaceticus consensus sequence, but only with 96.1% similarity. The ITS sequences of both these strains lack the 13i/1 indel, but possess indel 13i/4 (subtype BG1) also present in A. calcoaceticus. Although, the sequence of ITS region 150–178 in strain 250 was not present in the A. calcoaceticus ITS consensus sequence, it was seen in the ITS of two putative A. calcoaceticus strains 1H9 and OCI1 [24]. On the other hand the ITS of strain 175 very closely resembled those of every other Acb member at this location. At positions 549–512, the ITS sequences of strains 175 and 250 were identical to that of the A. calcoaceticus strains, but different to those of all other Acb members in this region. Thus, these data reveal further ITS sequence variations, although it is not clear whether either of these strains represent a new Acinetobacter species, especially since the 16S rRNA sequence of strain 250 is identical to that of A. calcoaceticus T (data not presented here).
Identification of Acb strains containing indel 13i/5 with the A. pittii targeted probe Aun3
When the A. pittii ITS-targeted Aun3 probe [14] was used against Acinetobacter c/t13TU strain 10090, a PCR amplicon identical in size to that generated from A. pittii ATCC19004T was obtained. Electrophoretic separation of the ITS amplicons on 2% agarose revealed that like A. pittii ATCC19004T, strain c/t13TU 10090 also produced a fragment of approximately 650 bp (Figure 5). The fragment size deduced from its ITS sequence alignment was 620 bp (Figure 3). No such fragment was generated from strain c/t13TU strain 5804, as its ITS sequence data would predict (Figure 4). Although not determined here, the same positive outcome would be expected for this probe when used against Acinetobacter b/n1&3 strains 10095 and 10169, and collection (C) strains 151, 159 and 169 too, since their ITS sequences also contain the Aun3 probe target site (Figure S1).
Figure 5. Electrophoretic separation of ITS amplicons from A. pittii ATCC19004T (A1), c/t13TU 5804 (A2), and c/t13TU 10090 (A3).
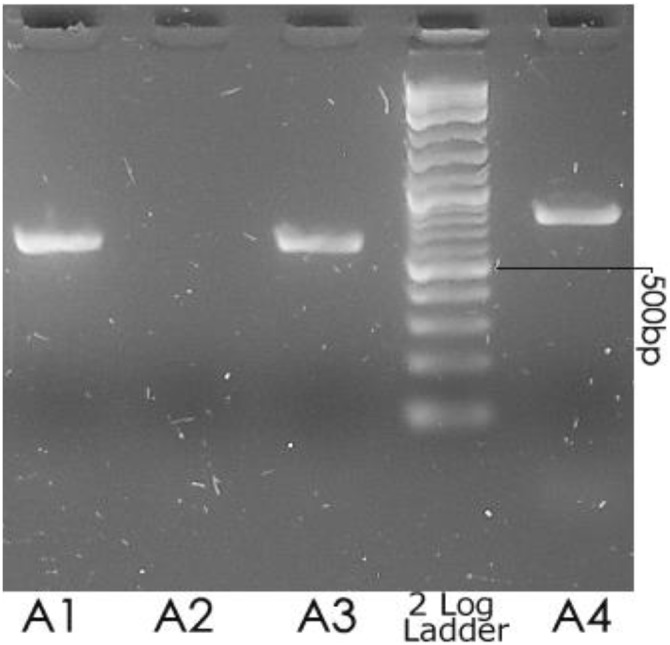
The amplification primers were 1512f and Aun3, designed to target only A. pittii strains [14]. A positive control was generated using primers 1512f and 6r to amplify the ITS sequences of strain c/t13TU 10090 (A4).
These results demonstrate elegantly what the sequence data in Figure 2 predict, namely that more than one species will be amplified by the indel 13i/5 targeted Aun3 oligonucleotide PCR primer, including A. oleivorans, A. calcoaceticus NC016603, A. pittii, Acinetobacter BENAB127, Acinetobacter c/t13TU 10090 and Acinetobacter b/n1&3 (10095 and 10169).
Discussion
The feasibility of applying ITS sequences to identify reliably members of the Acinetobacter Acb complex has been addressed here. Earlier work had claimed that the length of the ITS region was an inherent characteristic for each Acb species, and hence could be used potentially as a primary tool for their individual identification [25]. This proposal seems to hold true for A. baumannii, where ITS copies of all strains appear to share the same length (608 bp), and where the only sequence differences are single nucleotide polymorphisms. Furthermore, both whole-ITS sequencing [9], [18] and ITS targeted oligonucleotide probes were considered as promising in being able to differentiate between members of the Acb and other Acinetobacter genomic species [13]–[17]. However, the potential complications caused by variable length ITS sequences in strains, as clearly demonstrated here, had not been addressed in these discussions.
Results presented here firmly establish for the first time that variable length ITS sequences in members of the Acinetobacter Acb complex result from the horizontal transfer of DNA fragments (i.e. indels), usually between the ITS sequences of species within this genus (Figure 3). In some instances, the presence of indels results in one or more ITS copy variants in strains of some Acb species being indistinguishable from those in others. Consequently, neither applying oligonucleotide probes nor whole-ITS sequencing, both still commonly used for identifying Acb members [26], [12], can be assumed always to distinguish between them.
Alignment analyses of ITS sequences from A. calcoaceticus (59 ITS sequences, 99.5% similarity) and A. baumannii (132 ITS sequences, 99.7% similarity) suggest the two A. calcoaceticus-targeted and six A. baumannii-targeted oligonucleotide probes available will differentiate them from other members of the Acb complex (Figure 4). In each case, multiple nucleotide variants distinguish the probe target sites from comparable regions in the ITS sequences of the other Acb species (Figure 4). While the high degree of similarity between each of the many A. baumannii ITS sequences analysed here may suggest indels are absent, this does not preclude them from acting as donors in horizontal gene transfer events.
The detection of indels like 13i/1 can be revealed only by comparing ITS sequences of the same strain/species. Thus, such analyses of ITS sequences of several A. nosocomialis strains revealed the 615 bp ITS sequences of strain BCRC15417T were 8 bp longer than the 607 bp ITS sequences of the seven other A. nosocomialis (Group1) strains, because of the presence of the 37 bp indel 13i/1 (Figure 3). This finding addresses the concerns expressed by Zarilli et al. [18] regarding the distinctively different ITS length of most of their A. nosocomialis strains and the type strain BCRC15417T. It now appears that each of the less frequently seen 615 bp ITS copy sequences, regardless of the site of isolation of the host strains, contains this same indel 13i/1 located always in the same ITS position, which may result from a process of gene homogenization following its initial incorporation into a single ITS copy [27], [28], [29].
In contrast to the A. nosocomialis BCRC15417T ITS sequences, where all copies contain the same indel 13i/1, the ITS sequences of Acinetobacter strain c/t13TU 10090 exhibit considerable intra-genomic variations, which arise from the variable insertion of four different indels (Figures 2 & 3). Relative to the shortest (608 bp) ITS in this strain, their presence increases the ITS length by 1 bp (13i/2), 0 bp (13i/3), 22 bp (13i/4) and 12 bp (13i/5) respectively (Figure 3). All these five indels appear to have arisen from horizontal gene transfer events between Acinetobacter species e.g. A. calcoaceticus, A. baumannii, A. ursingii, A. pittii, A. nosocomialis, Acinetobacter b/n1&3, Acinetobacter c/t13TU, A. ursingii, and Acinetobacter BJ14, or members of other genera (Figure 3). Establishing which species acts as donor/recipients is not possible, given the limited number of Acinetobacter ITS sequences available.
As a consequence of containing indels, some individual c/t13TU 10090 ITS sequences are less similar to each other than they are to the sequences of ITS copies in other Acinetobacter species. For example, while the Acinetobacter c/t13TU 10090 608 bp and 609 bp ITS sequences are 98.7% similar to those of A. nosocomialis Group 1, the presence of indel 13i/5 in the 620 bp ITS results in its sequence being 99.4% similar to the A. pittii 620 bp ITS consensus sequence (Figure 3). Thus, its presence makes it effectively impossible to distinguish between the 620 bp ITS sequences of Acinetobacter c/t13TU 10090 and A. pittii (Figures 4 & 5).
Based on phenotypic properties, ribotyping, plasmid profiling and DNA-DNA hybridization values, Gerner-Smidt & Tjernberg [5] suggested Acinetobacter c/t13TU strains 5804 and 10090 represented a new genomic species within the Acb complex. While Acinetobacter c/t13TU strain 5804 has only one ITS Type, 98.8% similar to that of the A. nosocomialis 13TU Group 1 (607) ITS consensus sequence, the variable presence of indels in ITS sequences of strain c/t13TU 10090 means its phylogenetic placement depends on whether 608 and 609 ITS Type copies are used (Figures. 2 & 3). In earlier attempts to clarify this, Nemec et al. [6] showed using rpoB gene sequences that both these strains clustered closely with A. baumannii and A. nosocomialis. Yet on the other hand, their fusA, gltA and rplB gene sequences suggested they were not closely related to either. Unfortunately, it may not be possible to use ITS sequencing to resolve this taxonomic question until whole genome sequence data from more c/t13TU strains strains of these become available [30]. The value of using ITS sequence data to distinguish between Acb species is further undermined when the sequences of Acinetobacter b/n1&3 strains 10095 and 10169 are examined. Both were also placed into a new genomic species in the Acb complex using criteria similar to those applied to the c/t13TU strains [5]. The suggestion is that the higher mutability (and hence variability) of ITS sequences may make them more suitable than 16S rRNA genes for distinguishing between closely related bacterial species or strains [31], [22], [32], [33]. However, no such variability was apparent in the 620 bp ITS sequences of b/n1&3 strains 10095 and 10169. All were essentially identical (99% and >99.2% similar respectively) to the A. pittii ATCC19004T ITS sequence (Figure 3). Three possible conclusions arise from this: both are members of A. pittii, despite earlier data suggesting they represented distinct genomic species [5]; the ITS sequences of b/n1&3 strains 10095 and 10169 and those of the six A. pittii strains have not changed substantially since all eight strains last shared a common ancestor; their ITS sequences are so similar because of horizontal transfer of DNA fragments between them, possibly involving indel 13i/5 (Figure 3). It is not possible to determine which of these is the case until more Acinetobacter b/n1&3 strain ITS are sequenced.
Equally, the ITS sequences of Acinetobacter BENAB127 do not assist in resolving its possible relationship to existing Acb complex members. An alignment of its 628 bp ITS sequence against those of other Acb complex strains produced outcomes interpretable in one of two ways. One is that this strain is a member of A. pittii, and that each of its ITS sequences contains a variant of indel 13i/1 (Figures 1, 2 & 3). Alternatively, it might be argued equally that this strain is a member of A. calcoaceticus, and that each of its ITS sequences contains a variant of indel 13i/5 (Figures 1, 2 & 3). Similar problems arise in determining the taxonomic destiny of collection (C) strains 150, 159, 169, 175 and 250 (Figures 1, 2 & 3).
The presence of indels further complicates selection and application of the currently available ITS targeted probes to Acb members. If only a single ITS copy in a recipient strain possesses an indel which includes a region of DNA targeted by any probe, then its specificity is compromised. For example, in silico analysis of Acinetobacter c/t13TU 10090 620 bp ITS sequences reveals that the A. pittii-targeted probes Aci3 [16], P-3 [17] and Aun3 [14] would each return a positive result for Acinetobacter c/t13TU 10090, even though the 13i/5 indel is only carried in some, or possibly only one of its ITS copies. This prediction was confirmed experimentally, with strain c/t13TU 10090 returning a positive result for the Aun3 probe, while as expected, strain c/t13TU 5804 did not (Figures 4 & 5).
Unlike the A. pittii-targeted probes discussed above, neither of the A. nosocomialis-targeting Aun13TU [14] nor P-13 [17] probes appear to target ITS regions susceptible to recombination events (Figure 4). However, probe P-13 has been reported to cross hybridize with some A. pittii strains [17], which is not surprising given that ITS sequences of A. nosocomialis Group 1 strains differ from those of every other Acb strain by a single nucleotide only in the region targeted by this probe (Figure 4). Furthermore, since A. nosocomialis BCRC15417T lacks the P-13 target site this probe will not target ITS sequences in all A. nosocomialis strains (Figure 4). In contrast, the Aun13TU probe [14] targets an ITS region occurring in all known A. nosocomialis strains (Figure 4). Its specificity depends entirely upon a single nucleotide difference in the middle of its ITS target region that distinguishes A. nosocomialis strains from both A. baumannii (42 strains) and c/t13TU strains 5804 and 10090 (Figure 4). The value of the Aun13TU probe is further undermined when a Blast search reveals the ITS sequences of both A. radioresistens BCRC15425T (AY601839) and Acinetobacter genomic species BG6 BCRC15421 (AY601833) also contain its target site (Maslunka, unpublished).
One intriguing outcome of this work is that while individual ITS copies of many other Acinetobacter Acb species vary in their sequences from the presence of indels, those of the many different A. baumannii strains are all highly conserved. This is despite A. baumannii being naturally transformable [34], [35]. Diancourt et al. [36] also reported low levels of polymorphism in A. baumannii, which they suggested may indicate that A. baumannii either experienced a severe recent evolutionary bottleneck, or that the true diversity of this species is not represented by clinical isolates alone. While intra-genomic heterogeneity between bacterial ITS sequences resulting from homologous recombination is documented extensively [37], [38], [39], [27], [11], inter-genomic recombination events like those described here have never been reported previously for Acinetobacter ITS sequences. That the possibility of such events occurring has been overlooked in earlier studies is surprising, given the probable transformable nature of most Acinetobacter species [40], and that Acinetobacter ITS sequences contain, and are flanked by, highly conserved DNA regions (e.g. tRNAala, tRNAile, and the start/end of the 16S and 23S rRNA genes respectively). These data emphasize that a more cautious approach to using ITS sequences to identify individual Acinetobacter strains and species is needed, especially in those instances where variable length ITS sequences have been confirmed [41], [9]. An assessment of its usefulness as a target for species-specific probes/primers for other Acinetobacter species will only be possible when the whole genomes are sequenced in other strains [30] which contain variable length ITS copies.
Supporting Information
Alignments of c/t 13TU 10090 clone ITS sequences reveal presence of five indels. The number of ITS sequences used in each consensus sequence is given in parentheses [ ].
(PDF)
Acknowledgments
We wish to acknowledge the input of Dr. Simon McIlroy and the many helpful discussions on indels held with Dr. Daniel Tillett.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All sequences generated in this study were deposited in the DDBJ/EMBL/GenBank database (at EMBL-EBI, European Molecular Biology Laboratories, http://www.ebi.ac.uk/embl/) as follows: Strain collection (B). A. calcoaceticus strains, HE651573–HE651628 and HE651903–HE651906; A. baumannii strains, HE651629–HE651686 and HE651907–HE651910; A. pittii ATCC19004T, HE651687–HE651696 and HE651911; c/t13TU 5804 and c/t13TU 10090, FN669512–FN669527 and FN677952–FN677963; b/n1&3 10095 and b/n1&3 10169, HE651546–HE651572, HE651912 and HE651913; Acinetobacter BENAB127, HE651697–HE651710 and HE651914. Strain collection (C) strains 41 directly sequenced clinical isolates; 7–8, KC257006–7; 147–151, KC257008–12; 153–161, KC257013–21; 163–171, KC257022–30; 175–178, KC257031–34; 180–181, KC257035–36; 248–251, KC257037–40; 253–257, KC257041–45; 262, KC257046.
Funding Statement
CM was supported by a La Trobe University Postgraduate Scholarship. BG was supported by an Australian Post Graduate Award from the Australian Research Council. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. McConnell MJ, Actis L, Pachon J (2013) Acinetobacter baumannii: human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol Rev 37: 130–155. [DOI] [PubMed] [Google Scholar]
- 2. Dijkshoorn L, Nemec A, Seifert H (2007) An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat Rev 5: 939–951. [DOI] [PubMed] [Google Scholar]
- 3. Poirel L, Nordmann P (2006) Carbapenem resistance in Acinetobacter baumannii: mechanisms and epidemiology. Clin Microbiol Infect 12: 826–836. [DOI] [PubMed] [Google Scholar]
- 4. Gerner-Smidt P, Tjernberg I, Ursing J (1991) Reliability of phenotypic tests for identification of Acinetobacter species. J Clin Microbiol 29: 277–282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Gerner-Smidt P, Tjernberg I (1993) Acinetobacter in Denmark: II. Molecular studies of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex. APMIS 101: 826–832. [PubMed] [Google Scholar]
- 6. Nemec A, Krizova L, Maixnerova M, van der Reijden TJ, Deschaght P, et al. (2011) Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res Microbiol 162: 393–404. [DOI] [PubMed] [Google Scholar]
- 7.Dijkshoorn L, Nemec A (2008) The diversity of the genus Acinetobacter. In: Acinetobacter molecular biology, Edited by U. Gerisher. Norfolk, UK: Caister Academic Press. 1–34.
- 8. Gurtler V, Stanisich VA (1996) New approaches to typing and identification of bacteria using the 16S–23S rDNA spacer region. Microbiology 142: 3–16. [DOI] [PubMed] [Google Scholar]
- 9. Chang HC, Wei YF, Dijkshoorn L, Vaneechoutte M, Tang CT, et al. (2005) Species-level identification of isolates of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex by sequence analysis of the 16S–23S rRNA Gene Spacer Region. J Clin Microbiol 43: 1632–1639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Maslunka C, Carr E, Gurtler V, Kampfer P, Seviour R (2006) Estimation of ribosomal RNA operon (rrn) copy number in Acinetobacter isolates and potential of patterns of rrn operon-containing fragments for typing strains of members of this genus. Syst Appl Microbiol 29: 216–228. [DOI] [PubMed] [Google Scholar]
- 11. Maslunka C, Gürtler V, Carr E, Seviour R (2008) Unique organization of the 16S–23S intergenic spacer regions of strains of Acinetobacter baylyi provides a means for its identification from other Acinetobacter species. J Microbiol Meth 73: 227–236. [DOI] [PubMed] [Google Scholar]
- 12. Lai CC, Hsu HL, Tan CK, Tsai HY, Cheng A, et al. (2012) Recurrent bacteremia caused by the Acinetobacter calcoaceticus-baumannii complex. J Clin Microbiol 50: 2982–2986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Chen TL, Siu LK, Wu RC, Shaio MF, Huang LY, et al. (2007) Comparison of one-tube multiplex PCR, automated ribotyping and intergenic spacer (ITS) sequencing for rapid identification of Acinetobacter baumannii. Clin Microbiol Infect 13: 801–806. [DOI] [PubMed] [Google Scholar]
- 14. Ko WC, Lee NY, Su SC, Dijkshoorn L, Vaneechoutte M, et al. (2008) Oligonucleotide array-based identification of species in the Acinetobacter calcoaceticus-Acietobacter baumannii complex in isolates from blood cultures and antimicrobial susceptibility testing of the isolates. J Clin Microbiol 46: 2052–2059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Su SC, Vaneechoutte M, Dijkshoorn L, Wei YF, Chen Y, et al. (2009) Identification of non-fermenting Gram-negative bacteria of clinical importance by an oligonucleotide array. J Med Microbiol 58: 596–605. [DOI] [PubMed] [Google Scholar]
- 16. Lagatolla C, Lavenia A, Tonin E, Monti-Bragadin C, Dolzani L (1998) Characterisation of oligonucleotide probes for the identification of Acinetobacter spp. A. baumannii and Acinetobacter genomic species 3. Res Microbiol 149: 557–566. [DOI] [PubMed] [Google Scholar]
- 17. Lin YC, Sheng WH, Chang SC (2008) Application of a microsphere-based array for rapid identification of Acinetobacter spp. with distinct antimicrobial susceptibilities. J Clin Microbiol 46: 612–617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Zarrilli R, Giannouli M, Di Popolo A, Tomasone F, Chu YW (2009) Identification of Acinetobacter Genomic species 13TU by sequence Analysis of the 16S–23S rRNA gene spacer region. J Clin Microbiol 47: 1281–1282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Bouvet PJM, Grimont PAD (1986) Taxonomy of the genus Acinetobacter with the recognition of Acinetobacter baumannii sp. nov., Acinetobacter haemolyticus sp. nov., Acinetobacter johnsonii sp. nov., Acinetobacter junii sp. nov. and emended descriptions of Acinetobacter calcoaceticus and Acinetobacter lwoffii. Int J Syst Bacteriol 36: 228–240. [Google Scholar]
- 20. Tjernberg I, Ursing J (1989) Clinical strains of Acinetobacter classified by DNA-DNA hybridisation. APMIS 97: 595–605. [DOI] [PubMed] [Google Scholar]
- 21. Beacham AM, Seviour RJ, Lindrea KC, Livingston I (1990) Genospecies diversity of Acinetobacter isolates obtained from a biological nutrient removal pilot plant of modified UCT configuration. Water Res 24: 23–29. [Google Scholar]
- 22. Carr EL, Gurtler V, Seviour RJ (2004) Variation of 16S–23S rRNA intergenic spacer regions (ISRs) in Acinetobacter baylyi (Strain B2) isolated from activated sludge. Syst Appl Microbiol 27: 478–491. [DOI] [PubMed] [Google Scholar]
- 23. Gurtler V, Grando D, Mayall BC, Wang J, Ghaly-Derias S (2012) A novel method for simultaneous Enterococcus species identification/typing and van genotyping by high resolution melt analysis. J Microbiol Meth 90: 167–181. [DOI] [PubMed] [Google Scholar]
- 24. Peix A, Lang E, Verbarg S, Spröer C, Rivas R, et al. (2009) Acinetobacter strains IH9 and OCI1, two rhizospheric phosphate solubilizing isolates able to promote plant growth, constitute a new genomovar of Acinetobacter calcoaceticus. Syst Appl Microbiol 32: 334–341. [DOI] [PubMed] [Google Scholar]
- 25. Vaneechoutte M, Dijkshoorn L, Chang TC (2009) Identification of Acinetobacter genomic species 13TU by sequence analysis of the 16S–23S rRNA gene spacer region: Authors’ reply. J Clin Microbiol 47: 1281–1282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Golanbar GD, Lam CK, Chu YM, Cueva C, Tan SW, et al. (2011) Phenotypic and molecular characterization of Acinetobacter clinical isolates obtained from inmates of California correctional facilities. J Clin Microbiol 49: 2121–2131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Liao D (2000) Gene conversion drives within genic sequences: concerted evolution of ribosomal RNA genes in bacteria and archaea. J Mol Evol 51: 305–317. [DOI] [PubMed] [Google Scholar]
- 28. Luz SP, Rodríguez-Valera F, Lan R, Reeves PR (1998) Variation of the ribosomal operon 16S–23S gene spacer region in representatives of Salmonella enterica subspecies. J Bacteriol 180: 2144–2151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Stewart FJ, Cavanaugh CM (2007) Intragenomic variation and evolution of the transcribed spacer of the rRNA operon in bacteria. J Mol Evol 65: 44–67. [DOI] [PubMed] [Google Scholar]
- 30. Chan JZM, Halachev MR, Loman NJ, Constantinidou C, Pallen MJ (2012) Defining bacterial species in the genomic era: insights from the genus Acinetobacter. BMC Microbiol 12: 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Barry T, Colleran G, Glennon M, Dunican LK, Gannon F (1991) The 16S/23S ribosomal spacer region as a target for DNA probes to identify eubacteria. PCR Methods Appl 1: 51–56. [DOI] [PubMed] [Google Scholar]
- 32. Goncalves ER, Rosato YB (2002) Phylogenetic analysis of Xanthomonas species based upon 16S–23S rDNA intergenic spacer sequences. Int J Syst Evol Microbiol 52: 355–361. [DOI] [PubMed] [Google Scholar]
- 33. Jensen MA, Webster JA, Straus N (1993) Rapid identification of bacteria on the basis of polymerase chain reaction-amplified ribosomal DNA spacer polymorphisms. Appl Environ Microbiol 59: 945–987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gerischer U (2008) Acinetobacter Molecular Biology. Norfolk, UK: Caister Academic Press.
- 35. Ramirez MS, Don M, Merkier AK, Soler Bistue AJ, Zorreguieta A, et al. (2010) Naturally competent Acinetobacter baumannii clinical isolate as a convenient model for genetic studies. J Clin Microbiol 48: 1488–1490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Diancourt L, Passet V, Nemec A, Dijkshoorn L, Brisse S (2010) The population structure of Acinetobacter baumannii: expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS One 5: e10034 doi:10.1371/journal.pone.0010034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Gogarten JP, Townsend JP (2005) Horizontal gene transfer, genome innovation and evolution. Nat Rev Microbiol 3: 679–687. [DOI] [PubMed] [Google Scholar]
- 38. Gurtler V (1999) The role of recombination and mutation in 16S–23S rDNA spacer rearrangements. Gene 238: 241–252. [DOI] [PubMed] [Google Scholar]
- 39. Lebuhn M, Bathe S, Achouak W, Hartmann A, Heulin T, et al. (2006) Comparative sequence analysis of the internal transcribed spacer 1 of Ochrobactrum species. Syst App Microbiol 29: 265–275. [DOI] [PubMed] [Google Scholar]
- 40. Jung J, Madsen EL, Jeon CO, Park W (2011) Comparative genomic analysis of Acinetobacter oleivorans DR1 to determine strain-specific genomic regions and gentisate biodegradation. Appl Environ Microbiol 77: 7418–7424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Barbe V, Vallenet D, Fonknechten N, Kreimeyer A, Oztas S, et al. (2004) Unique features revealed by the genome sequence of Acinetobacter sp. ADP1, a versatile and naturally transformation competent bacterium. Nucleic Acids Res 32: 5766–5779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Gürtler V, Grando D (2013) New opportunities for improved ribotyping of C. difficile clinical isolates by exploring their genomes. J Microbiol Meth. 93: 257–272. [DOI] [PubMed] [Google Scholar]
- 43.Gürtler V, Subrahmanyam G, Shekar M, Maiti B, Karunasagar I (2014) Bacterial typing and identification by genomic analysis of 16S–23S rRNA intergenic spacer (ITS) sequences. “In: New Approaches to Prokaryotic Systematics, Methods in Microbiology volume 41, Chapter 3”. in press. Elsevier Academic Press, San Diego, CA.
- 44. Zhan Y, Yan Y, Zhang W, Yu H, Chen M, et al. (2011) Genome sequence of Acinetobacter calcoaceticus PHEA-2, isolated from industry wastewater. J Bacteriol 10: 2672–2673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Smith MG, Gianoulis TA, Pukatzki S, Mekalanos JJ, Ornston LN, et al. (2007) New insights into Acinetobacter baumannii pathogenesis revealed by high-density pyrosequencing and transposon mutagenesis. Genes Dev 21: 601–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Iacono M, Villa L, Fortini D, Bordoni R, Imperi F, et al. (2008) Whole-genome pyrosequencing of an epidemic multidrug-resistant Acinetobacter baumannii strain belonging to the European clone II group. Antimicrob Agents Chemother 52: 2616–2625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Vallenet D, Nordmann P, Barbe V, Poirel L, Mangenot S, et al. (2008) Comparative analysis of Acinetobacters: three genomes for three lifestyles. PLoS One 3: e1805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Peleg AY, de Breij A, Adams MD, Cergueira GM, Mocali S, et al. (2012) The success of Acinetobacter species; genetic, metabolic and virulence attributes. PLos One 7: e46984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Chen Y, Cui Y, Pu F, Jiang G, Zhao X, et al. (2012) Draft genome sequence of an Acinetobacter genomic species 3 strain harbouring a bla (NDM-1) gene. J Bacteriol. 194: 204–205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Jung J, Baek J, Park W (2010) Complete genome sequence of the diesel-degrading Acinetobacter sp. strain DR1. J. Bacteriol 192: 4794–4795. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Alignments of c/t 13TU 10090 clone ITS sequences reveal presence of five indels. The number of ITS sequences used in each consensus sequence is given in parentheses [ ].
(PDF)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All sequences generated in this study were deposited in the DDBJ/EMBL/GenBank database (at EMBL-EBI, European Molecular Biology Laboratories, http://www.ebi.ac.uk/embl/) as follows: Strain collection (B). A. calcoaceticus strains, HE651573–HE651628 and HE651903–HE651906; A. baumannii strains, HE651629–HE651686 and HE651907–HE651910; A. pittii ATCC19004T, HE651687–HE651696 and HE651911; c/t13TU 5804 and c/t13TU 10090, FN669512–FN669527 and FN677952–FN677963; b/n1&3 10095 and b/n1&3 10169, HE651546–HE651572, HE651912 and HE651913; Acinetobacter BENAB127, HE651697–HE651710 and HE651914. Strain collection (C) strains 41 directly sequenced clinical isolates; 7–8, KC257006–7; 147–151, KC257008–12; 153–161, KC257013–21; 163–171, KC257022–30; 175–178, KC257031–34; 180–181, KC257035–36; 248–251, KC257037–40; 253–257, KC257041–45; 262, KC257046.