

Abstract
α-Herpesvirinae subfamily members, including herpes simplex virus type 1 (HSV-1) and bovine herpes virus 1 (BHV-1), initiate infection in mucosal surfaces. BHV-1 and HSV-1 enter sensory neurons by cell-cell spread where a burst of viral gene expression occurs. When compared to non-neuronal cells, viral gene expression is quickly extinguished in sensory neurons resulting in neuronal survival and latency. The HSV-1 latency associated transcript (LAT), which is abundantly expressed in latently infected neurons, inhibits apoptosis, viral transcription, and productive infection, and directly or indirectly enhances reactivation from latency in small animal models. Three anti-apoptosis genes can be substituted for LAT, which will restore wild type levels of reactivation from latency to a LAT null mutant virus. Two small non-coding RNAs encoded by LAT possess anti-apoptosis functions in transfected cells. The BHV-1 latency related RNA (LR-RNA), like LAT, is abundantly expressed during latency. The LR-RNA encodes a protein (ORF2) and two microRNAs that are expressed in certain latently infected neurons. Wild-type expression of LR gene products is required for stress-induced reactivation from latency in cattle. ORF2 has anti-apoptosis functions and interacts with certain cellular transcription factors that stimulate viral transcription and productive infection. ORF2 is predicted to promote survival of infected neurons by inhibiting apoptosis and sequestering cellular transcription factors which stimulate productive infection. In addition, the LR encoded microRNAs inhibit viral transcription and apoptosis. In summary, the ability of BHV-1 and HSV-1 to interfere with apoptosis and productive infection in sensory neurons is crucial for the life-long latency-reactivation cycle in their respective hosts.
Keywords: alpha-herpesviruses, latency in sensory neurons, apoptosis, non-coding RNAs
α-Herpesvirinae Subfamily Members are Important Pathogens in their Respective Hosts
Herpes simplex virus type 1 (HSV-1) and bovine herpes virus 1 (BHV-1) are both important pathogens in their respective natural hosts and both are α-herpesvirinae subfamily members. For example, HSV-1 is the cause of one of the most frequent and serious viral eye infections in the United States, with over 400,000 affected individuals.1 Following primary infection of the eye, latency is established in sensory neurons within trigeminal ganglia (TG).2,3 HSV-1 reactivates sporadically from TG and the infectious virus can be detected on surfaces of the eye, where it can cause recurrent ocular disease. Reactivation from latency is necessary for recurrent ocular HSV-1 infections.4,5 Long-term oral acyclovir treatment only reduces ocular HSV-1 recurrences by 41%.6 Herpes simplex virus type 2 (HSV-2) is the cause of the most common sexually transmitted disease, and sporadic recurrent genital lesions occur periodically. Two genital herpes vaccine trials failed7,8 indicating there is a need for new and effective therapies that will reduce the incidence of recurrent HSV-1 and HSV-2 disease.
Bovine herpes virus 1 (BHV-1) is an important pathogen of cattle as it induces clinical signs in the upper respiratory tract of cattle and is immunesuppressive. BHV-1 establishes latency in sensory neurons, but periodically reactivates from latency, and thus is widespread in cattle.2,9–11 BHV-1 infection inhibits cell-mediated immunity,12–15 CD8+ T cell recognition of infected cells,16–19 and induces apoptosis in CD4+ T cells.20,21 Two viral regulatory proteins, bICP0 and bICP27, inhibit interferon dependent transcription.10,22–25 Infection also erodes mucosal surfaces of the upper respiratory tract, which can allow bacterial pathogens to colonize the lower respiratory tract.26–28
Acute Infection Results in High Levels of Infectious Virus and Apoptosis
Binding and entry of HSV-1 and BHV-1 to mammalian cells are mediated by viral glycoproteins and cellular factors.29–31 A cellular receptor (HveA or HVEM) is primarily expressed in activated T cells and belongs to the tumor necrosis factor receptor family.32 Entry of HSV-1 into epithelial and fibroblasts is mediated by another membrane glycoprotein, HveB or HveC.33 HveC is an entry mediator for HSV-1 and BHV-1 and is abundantly expressed in neurons. Additionally, soluble HveC blocks viral entry in neuronal-like cell lines.33 After uncoating, the viral genome enters the nucleus and productive infection is initiated.
HSV-1 and BHV-1 gene expression is tightly regulated in three distinct phases during productive infection of cultured cells: immediate early (IE), early (E), or late (L).34 IE RNA expression does not require protein synthesis and is stimulated by VP16, a viral structural protein.35 E gene expression requires at least one IE protein, and E genes encode nonstructural proteins that stimulate viral DNA replication. L gene expression is maximal after viral DNA replication, requires IE protein production, and L proteins comprise the virion particle. Although a vigorous immune response leads to viral clearance following primary infection, BHV-1 and HSV-1 establish a life-long latent infection in ganglionic sensory neurons, primarily TG, or sacral dorsal root ganglia.2,3,9,36 Approximately 40% of sensory neurons appear to harbor viral genomes during latency.37–41
Five HSV-1 IE genes encode ICP0, ICP4, ICP22, ICP27, or ICP47. ICP442–45 and ICP2746–48 are required for virus growth in tissue culture. ICP4 represses IE gene expression44,49–53 but activates E or L gene expression by interacting with RNA polymerase II transcription factors and specifically binding viral DNA.49,54 ICP27 redistributes small nuclear ribonucleoprotein complexes, interferes with splicing of IE transcripts, and promotes E and L poly A site selection.55–58 ICP47 prevents transport of antigenic peptides into the endoplasmic reticulum59 and inhibits CD8+ T cell responses.60 ICP22 enhances viral gene expression, in part by modifying RNA polymerase II.61 ICP0 increases steady-state levels of viral mRNA and stimulates all viral promoters.62 ICP0 also binds several cellular proteins: (1) elongation factor 1-α63; (2) cyclin D364; (3) an ubiquitin-specific protease65,66; and (4) promyelocytic leukemia (PML) protein.67–69 Interactions between ICP0 and chromatin-remodeling enzymes activate viral transcription by multiple mechanisms, including sequestering histone deacetylase (HDAC) inhibitors.70,71 Secondly, HSV-1 ICP0 interacts with HDAC272 and blocks histone deacetylation to stimulate viral gene expression.73,74 ICP0 also alters a complex that inhibits gene expression (REST/CoREST/HDAC repressor complex).73 Since ICP0 can remove histones from viral chromatin during productive infection,75 ICP0 may have similar functions during reactivation from latency. These activities of ICP0 promote virus replication in differentiated cells.76 BHV-1 encoded ICP0 (bICP0) has similar functions as ICP0.10
Viral infection routinely leads to apoptosis in cultured cells.77–80 Killing of infected cells by apoptosis in vivo can reduce inflammation, alter immune recognition, reduce burst size, and thus prevent virus spread. Premature apoptosis of infected cells limits production of infectious virus and limits viral spread. Members of the α-herpesvirinae subfamily induce apoptosis after infection of cultured cells.81–86 HSV-183,84,87–89 and BHV-190 can also inhibit apoptosis in a cell type dependent manner after infection of cultured cells. HSV can induce DNA damage, and consequently apoptosis, even in the absence of productive infection.91–95 Two viral proteins, US1.5 and UL13, activate caspase 3 in the absence of other viral proteins, indicating these viral proteins play an important role during virus mediated apoptosis.96 Finally, ICP0 is also a trigger for apoptosis in the context of productive infection, in part because it activates viral gene expression.
HSV-1 encodes several proteins (ICP27, Us3, Us5, gJ, gD, and LAT) that have anti-apoptosis activity.83–85,87,88,97–105 Us3 is a serine/threonine protein kinase that inhibits cleavage and activation of the pro-apoptotic Bcl-2 family member, Bad. Us3 protein expression in cultured cells, in the absence of other viral proteins, inhibits caspase 3 activation, a crucial executioner caspase that commits cells to apoptosis. As expected, US3 inhibits the pro-apoptotic activity of US1.5 and UL13 by blocking caspase 3 activation.96 These anti-apoptotic genes play an important role in the pathogenic properties of HSV-1.
Viral Genes Expressed During Latency Regulate the Latency-Reactivation Cycle
The HSV-1 latency associated transcript is abundantly expressed during latency and regulates the latency-reactivation cycle
The HSV-1 latency associated transcript (LAT) is abundantly expressed in sensory gangionic neurons of mice, rabbits, or humans that are latently infected.106–114 LAT is predominantly expressed in the nucleus of latently infected neurons suggesting it is a non-protein coding regulatory RNA. LAT is antisense to ICP0 and overlaps ICP0 mRNA sequences (Fig. 1B), suggesting LAT inhibits ICP0 expression by an anti-sense mechanism. Although the ability of LAT to repress ICP0 expression may be important, LAT sequences that promote spontaneous reactivation in a rabbit ocular model of infection do not overlap ICP0 mRNA sequences.115
Figure 1.

Location of genes within the HSV-1 repeats. (Panel A) UL and U denote the unique sequences of the long (L) and short (S) components of the genome. The boxes depict repeat sequences. (Panel B) Transcription map of the repeat region. Location and orientation of LAT,111,112 ICP0, α134.5,253,254 ORFP,150 L/STs255 are indicated by solid lines.
Note: Partially mapped transcripts (αX and αX) are denoted by dashed arrows.256,257
Splicing of the 8.5 kb LAT transcript yields a stable 2 kb LAT and an unstable 6.5 kb LAT (Fig. 1B).107,111,116 Correct splicing of the 2 kb LAT is necessary for establishment and maintenance of latency.117,118 In general, the 2 kb LAT is not capped, is poly A-, appears to be circular, and is a stable intron.119,120 A subset of LAT is detected in the cytoplasm97,121,122 and is associated with polyribosomes or splicing factors.97,123 Small non-coding RNAs can regulate gene expression,124,125 promote neuronal differentiation,126 or inhibit apoptosis127 suggesting LAT is a non-coding regulatory RNA.
A study by Umbach et al128 concluded that LAT is a microRNA (miRNA) precursor which encodes four miRNAs, two within LAT promoter sequences (Fig. 2A and B). LAT miR-H6, reduces ICP4 protein steady state levels but not ICP4 RNA levels. Protein levels, not RNA levels, of ICP0 are inhibited by the LAT miRNA, miR-H2-3p. Within the first 1.5 kb of LAT coding sequences, two small RNAs (sRNAs), LAT sRNA1 and sRNA2, were identified (Fig. 2B). The sRNAs are larger than mature miRNAs (typically 23 nucleotides long) and the sequence of both possess extensive secondary structure. The sRNAs can be detected in TG of mice latently infected with wild-type HSV-1, but not in TG of mice latently infected with a LAT null mutant.79,129 LAT sRNA2, but not LAT sRNA1, reduced ICP4 protein levels in transient transfection assays. Both LAT sRNAs inhibit productive infection in mouse neuroblastoma cells, however LAT sRNA1 inhibited productive infection more efficiently than LAT sRNA2.130 Collectively, these studies provide evidence that the LAT encoded miRNAs and sRNAs promote latency by interfering with expression of important viral transcriptional regulatory proteins.
Figure 2.

Schematic of putative factors encoded within the LAT locus. (Panel A) Schematic of genes within the long repeats that contain the LAT locus. The large arrow indicates the primary LAT transcript. The solid rectangle represents the sTable 2 kb LAT intron. Initiation of LAT transcription is denoted by the arrow at +1 (genomic nucleotide 118801). Several restriction enzyme sites and the relative locations of the ICP0 and ICP34.5 transcripts are shown for reference. The location of the 6 microRNAs (miR-H1-6) that is located within the 8.5 kb LAT128 are shown. (Panel B) Partial restriction map of LAT and position of LAT open reading frames (L1-8) within the first 1.5 Kb of strain McKrae LAT coding sequences, which were based on previous studies.148 The numbering system of the ORFs was consistent with a previous study.148 Only the ORFs with at least 30 amino acids are shown (the number of amino acids in each ORF is denoted by the numbers in brackets). Open circles denote the position of two LAT small RNAs that are encoded within the first 1.5 kb LAT coding sequences.258 Positions of UOL transcript, AL transcript, and ORFs located on the opposite strand of LAT (AL2 and AL3) are shown. The number of amino acids of AL2 and AL3 are in brackets. Nucleotide positions relative to the start of LAT transcription are not shown in parenthesis.
The LAT locus encodes additional transcripts (Fig. 2B). More than one transcript, including UOL (Upstream of LAT)131 are located within LAT promoter sequences. Expression of the UOL transcript or protein does not reduce reactivation from latency in rabbits.132 An antisense to LAT (AL) transcript is expressed from the first 1.5 kb of LAT coding sequences and appears to encode a protein.133 Two additional small open reading frames (ORFs) that are antisense to LAT (AL2 and AL3) are present in LAT coding sequences. An AL3 transcript is expressed during productive infection and in TG of mice latently infected with wild-type HSV-1, but not a LAT null mutant virus.134 An AL3-specific polyclonal antibody detected a protein in a subset of TG neurons in latently infected mice. A transcript encompassing AL2 has not been detected during productive infection or latency (unpublished data).
LAT null HSV-1 mutants have been examined in various small animal models.2,3 Although two studies concluded that LAT does not play a role in latency,135,136 most have provided evidence that LAT is important. This discrepancy may be due to the strain of virus or mouse that was used for specific studies. LAT enhances the establishment of latency in mice137,138 and in rabbit ocular infection models,139 in part by reducing lytic cycle viral gene expression in TG of mice.140,141 By enhancing the establishment of latency, LAT would increase the pool of latently infected neurons; thus indirectly increasing the incidence of reactivation from latency.
As a result of stress or other external stimuli, reactivation from latency can occur, resulting in virus shedding (Fig. 4). The McKrae strain of wild-type HSV-1, however not a LAT null mutant, is consistently detected in tears of infected rabbits, due to spontaneous reactivation.139,142–145 These same LAT null mutants grow with wild-type efficiency in cultured cells and in acutely infected rabbits. When just the first 1.5 kb of LAT coding sequences (Fig. 2B) is expressed from the HSV-1 genome, wild-type levels of spontaneous reactivation from latency occur in rabbits.139 Similar results were observed using another virulent strain of HSV-1 (17 syn+) in a rabbit eye model.146,147 The first 1.5 kb of LAT coding sequences does not overlap ICP0, suggesting that antisense repression of ICP0 expression by LAT is not important for spontaneous reactivation in the rabbit ocular model of infection. The factors encoded by the first 1.5 kb of LAT coding sequences that promote spontaneous reactivation have not been fully characterized.
Figure 4.

Putative steps that occur during the latency-reactivation cycle.
Note: For details, see text.
Although certain studies suggested LAT does not encode a protein,148 other studies have concluded that a protein encoded within LAT sequences is expressed.131,149–154 These proteins were suggested to either substitute for ICP0 functions,153,154 interfere with binding of ICP4 to DNA,152 or their functions were not described. The proposed LAT proteins are mapped downstream of the critical first 1.5 kb of the primary LAT transcript, a region that appears both sufficient and necessary for the wild type spontaneous reactivation phenotype in rabbit models.139,155 Within the first 1.5 kb of LAT coding sequences, 8 potential ORFs have been identified in the McKrae strain (Fig. 2B).148 The L2 ORF (Fig. 2B) appears to be expressed in TG of latently infected mice.156 Although LAT is not absolutely required for the latency-reactivation cycle in small animal models, its importance may be underestimated using small animal models and measuring latency in terms of weeks or months, not decades.
The BHV-1 latency related RNA is abundantly expressed in sensory neurons and is necessary for reactivation from latency
Latency related (LR) RNA is abundantly expressed in TG neurons of calves that are latently infected.111,157 Two different start sites of LR-RNA transcription (Fig. 3A) have been identifed suggesting this has functional significane. It is clear that the LR gene encodes more than one product.11,36 For example, the LR gene contains two-well defined ORFs (ORF2 and ORF1; Fig. 3B) and two reading frames that lack an initiating methionine (RF-B and RF-C). As a result of alternative splicing of polyA+ LR-RNA in TG of infected calves (Fig. 3A),158,159 ORF2 can be fused with ORF1 protein coding sequences or RF-B. The ORF2/ORF1 fusion protein stably interacts with the cellular transcription factor C/EBP-alpha.160 C/EBP-alpha RNA and protein levels increase in TG neurons during dexamethasone induced reactivation from latency. Over-expression of C/EBP-alpha enhanced productive infection,161 suggesting that ORF2 sequesters C/EBP-alpha and reduces the efficieny of productive infection during the latency-reactivation cycle.
Figure 3.
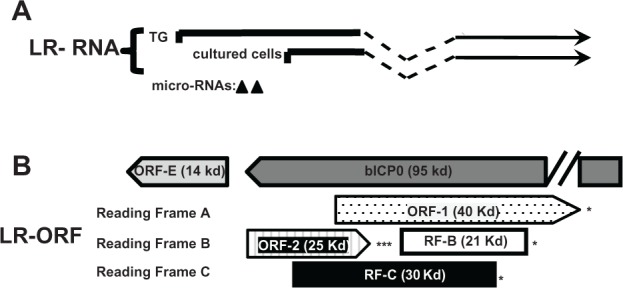
Schematic of the BHV-1 LR gene and surrounding genes. (Panel A) The start sites for LR transcription during latency and productive infection were previously described.159,259 (Panel B) Organization of LR ORFs and 3′ terminus of bICP0. ORF-1 and ORF-2 are located in the LR gene and have the potential to encode a 40 or 25 kd protein respectively.
Notes: Reading Frames B (RF-B) and C (RF-C) are open reading frames that lack an initiating Met. The (*) denotes the position of stop codons that are in frame with the respective ORF. The positions of ORF-E and bICP0, which are antisense to LR-RNA, are also shown.
One day after calves are infected and during latency, splicing of LR-RNA in TG is such that ORF2 is intact,159 suggesting ORF2 expression is important for the latency-reactivation cycle. ORF2 interacts with Notch1 and Notch3, components of the Notch signaling pathway.162 Mammalian Notch receptor family members (Notch1-4) are membrane tethered transcription factors that regulate many developmental and physiological processes.163,164 For example, Notch promotes neuronal maintenance, development, and differentiation.165–167 Notch3168 and Notch1169,170 promote cell survival by activating a protein kinase, (AKT) which inhibits apoptosis. Notch family members can also induce apoptosis,163,164 suggesting Notch influences cell survival by cell-type dependent mechanisms. When the Notch receptor is engaged by one of its five transmembrane ligands (Jagged1, Jagged2, Delta-like1, Delta-like3, or Delta-like4), the Notch intracellular domain (ICD) is cleaved by specific proteases, and subsequently translocates to the nucleus. In the nucelus, Notch ICD interacts with members of the CSL family of transcriptional factors, CBF1, Su(H), or Lag1 (also referred to as RBP-J binding proteins) subsequently activating downstream genes. Notch1, but not Notch3, enhances BHV-1 productive infection163 and Notch1 activates the BHV-1 immediate-early transcription unit 1 (IEtu1) and bICP0 early promoters. Notch1 and Notch3 trans-activated the late glycoprotein C (gC) promoter. ORF2 interferes with the ability of Notch1 to trans-activate the bICP0 early promoter and Notch1 or Notch3 mediated activation of the gC promoter162 suggesting this function is important for establishing and/or maintaining latency. Notch3 RNA levels are higher during dexamethasone (DEX) induced reactivation from latency, suggesting Notch family members stimulate productive infection during reactivation from latency. Activation of Notch signaling in post-mitotic-neurons or neuroblastoma cells inhibits neurite sprouting165,171–174 and axon repair,175 which can lead to neuronal degeneration and apoptosis.176–178 Conversely, neurite sprouting correlates with regeneration of damaged axons and dendrites.175 ORF2 promotes neuruite sprouting and neuronal differentiation of mouse neuroblastoma cells when Notch1 or Notch3 is over-expressed.179 Collectively, these studies suggest that ORF2 interactions with Notch family members promote the establishment and maintenace of latency by (1) interfering with viral gene expression necessary for productive infection, (2) supporting a mature neuronal phenotype, and (3) overcoming the deleterious effects of Notch expression during stress-induced reactivation from latency.
Although the results from the LR mutant virus suggested that proteins encoded by the LR gene are necessary for the latency-reactivation cycle, non-protein coding functions within LR-RNA have also been identified. For example, the intact LR gene inhibits the ability of bICP0 to stimulate productive infection in a dose-dependent manner.180,181 Insertion of three in-frame stop codons at the amino-terminus of the first ORF within the LR gene (ORF2) inhibited bICP0 repression with similar efficiency as the wild-type LR gene, suggesting expression of a LR protein is not required.181 LR gene products also inhibit mammalian cell growth,182,183 and the cell growth inhibitory function of the LR gene maps to a 463-bp fragment that lacks a significant open reading frame.182 Two miRNAs located upstream of ORF2 are expressed during latency.184 These miRNAs, or larger sRNAs containing these miRNAs, reduced bICP0 protein levels in transient transfection assays.
A small ORF located within the LR promoter is designated ORF-E (Fig. 3B). ORF-E is antisense to the LR transcript and is downstream of bICP0 coding sequences, but does not overlap bICP0. A transcript that encompasses ORF-E is expressed during productive infection and in TG of latently infected calves.185 The LR promoter contains multiple cis-acting motifs, has a neuronal specific binding domain,186–188 and contains a long AT-rich motif (40/53 nucleotides are A or T) that may promote ORF-E transcription. When ORF-E protein coding sequences are fused in frame with green fluorescent protein (GFP) sequences, GFP protein expression is detected in the nucleus of mouse or human neuroblastoma cells. In contrast, the ORF-E-GFP fusion protein is detected throughout rabbit skin cells. In transient transfection assays, ORF-E promotes neurite formation in mouse neuroblastoma cells,189 which may support a mature neuronal phenotype following infection.
LAT and LR Gene Products Inhibit Apoptosis
LAT inhibits apoptosis
LAT expressing plasmids interfere with apoptosis in transiently transfected cells, and LAT expressing viruses inhibit apoptosis in TG of infected mice or rabbits.117,190–192 The anti-apoptotic functions of LAT correlate with promoting spontaneous reactivation from latency.191,193 In the context of promoting spontaneous reactivation from latency in the rabbit model (model), inhibiting apoptosis is the most important function of LAT as three different anti-apoptosis genes129,194–196 restore wild-type levels of spontaneous reactivation from latency to a LAT null mutant. LAT may encode other functions because the LAT null mutants that express cellular anti-apoptosis genes have reduced virulence, in spite of reactivating from latency with wild-type frequency. LAT expressing plasmids, in the absence of other viral genes, inhibit caspase 8- and caspase 9-induced apoptosis,193,197 the two major apoptotic pathways in mammals.198–200 LAT also inhibits caspase 3 activation.201
LAT sRNA1 and sRNA2 cooperate to inhibit cold-shock induced apoptosis in mouse neuroblastoma cells.130 Introduction of ATG→TTG mutations in ORFs within the first 1.5 kb of LAT coding sequences impairs the anti-apoptotic functions of LAT,202 suggesting that LAT either encodes a functional protein or alters RNA structure. Two of these ATG→TTG mutations are within LAT sRNA1 and sRNA2, and introducing these mutations into the small RNAs inhibits their ability to inhibit apoptosis.130 At this time, it is not clear how these sRNAs interfere with apoptosis. It will also be important to construct a recombinant virus with these same mutations and test whether the spontaneous reactivation incidence is affected.
LAT also inhibits GrzB induced apoptosis in transient transfection studies.203 GrzB is released from CD8+ T cells as well as other specific lymphocytes; GrzB has features similar to apical caspases, and can induce apoptosis in most cell types.204–207 Inhibiting GrzB induced apoptosis may be important for the latency-reactivation cycle because CD8+ T lymphocytes control HSV infection in sensory ganglia.208,209
The LR gene encodes more than one product that inhibits apoptosis
A mutant BHV-1 strain with 3 stop codons after the initiating methionine codon of ORF-2 (LR mutant virus) does not express detectable levels of ORF-2101 but expresses reduced levels of ORF1 in cultured cells during productive infection.210 The LR mutant virus grows less efficiently in the ocular cavity and TG, but grows almost as efficiently as wild-type BHV-1 in the nasal cavity, and does not reactivate from latency following DEX treatment.211,212 The LR mutant virus induces higher levels of apoptosis in TG neurons of infected calves,213 and a LR gene expressing plasmid with the same stop codon mutations does not effectively inhibit apoptosis.214,215 ORF2 expression in the absence of other viral genes inhibits apoptosis in transiently transfected cells,216,217 suggesting that ORF2 is a dominant function encoded by the LR gene. ORF2, like LAT, can inhibit caspase 8 and caspase 9 mediated apoptosis; however the mechanism by which it inhibits apoptosis is not known.
Two microRNAs encoded within the LR gene (Fig. 3A) interfere with bICP0 protein expression184 and cold shock induced apoptosis in transfected mouse neuroblastoma cells (Neuro-2A cells).218 Since cold shock induced apoptosis in Neuro-2A cells is inhibited by casapse 3 and caspase 9 inhibitors,219 the microRNAs must influence these apoptotic signaling pathways. The ability of the microRNAs to stimulate the anti-apoptotic transcription factor NF-αB220–223 seems to be important for inhibiting cold-shock induced apoptosis. In summary, these results provide additional evidence that interfering with apoptosis is crucial for a successful life-long latent infection.
Why is Inhibiting Neuronal Apoptosis Important During the Latency-Reactivation Cycle?
The latency-reactivation cycle has been operationally divided into three distinct steps: establishment, maintenance, and reactivation (Fig. 4). Following acute infection where high levels of infectious virus are produced, virus particles enter sensory neurons. Initial entry of the viral genome into a sensory neuron results in a burst of lytic cycle viral gene expression and infectious viruses are produced. Viral gene expression is then extinguished, with the exception of HSV-1 LAT and BHV-1 LR gene products. Neuronal cell factors,2,9 LAT encoded microRNAs plus sRNAs,128,130 and LR encoded functions162,184 interfere with various aspects of productive infection. During acute infection and establishment of latency, neuronal and satellite cells undergo apoptosis when small animal models are infected with HSV-2224 or HSV-1.225–227 BHV-1 replication and gene expression also occur in TG of acutely infected calves, resulting in apoptosis of neurons and non-neuronal cells.192,213,228
HSV-1 LAT192 and the LR gene213 enhance neuronal survival during the establishment of latency. The ability of LAT229 and the LR gene179 to promote a mature neuronal phenotype and sprout neurites may also promote establishment of latency by stimulating repair of damaged neurons following infection. Successful establishment correlates with an increase in the number of infected neurons that survive and enhances the probability that reactivation from latency occurs.
Maintenance of latency is a phase that lasts for the duration of the host’s life and is operationally defined as a period when infectious virus is not readily detected. In general, abundant expression of viral genes required for productive infection does not occur. LAT or LR gene products are abundantly expressed during the maintenance of latency. Expression of LAT correlates with an increase of latently infected neurons during the maintenance of latency,230 suggesting latently infected sensory neurons are exposed to apoptotic stimuli during the maintenance of latency. It is reasonable to predict that LAT and LR gene products actively participate in maintaining a latent infection in sensory neurons.
Reactivation from latency is initiated by external stimuli (stress, immunosuppression, or UV light for example), which ultimately must stimulate viral gene expression.36,231,232 Abundant viral gene expression can be detected in sensory neurons and infectious virus can be isolated from TG, ocular swabs, and/or nasal swabs. Stress leads to elevated corticosteriod levels, which has rapid effects on neural activity.233,234 DEX, a synthetic corticosteriod, induces viral gene expression,235 stimulates an HSV-1 origin of replication (Ori-L) in neuronal cells,55 and alters splicing patterns in the absence of protein synthesis.236 DEX and other apoptosis stimulators can also stimulate HSV-1 reactivation from latency.237,238 BHV-1 reactivation from latency is induced by DEX, in part because it stimulates expression of cellular transcription factors and viral gene expression while repressing expression of LR gene products.9,11,36 Prolonged exposure to corticosteroids can also induce immunosuppression, in part by inducing apoptosis in lymphocytes.239 A subset of neurons that successfully reactivate from latency to produce infectious virus may not survive;240–242 however it is not clear if this is the fate for all neurons that produce infectious virus during reactivation from latency. Most latently infected neurons that are exposed to reactivation stimuli re-establish latency and do not produce infectious virus.243,244 Given that sensory neurons are terminally differentiated cells, inhibiting apoptosis during the latency-reactivation cycle is crucial for life-long latent infections of α-herpesvirinae subfamily members.
Numerous studies have demonstrated that infiltrating lymphocytes in TG regulate the latency-reactivation cycle. For example, a persistent cell-mediated immune response occurs in TG during latency and CD8+ T lymphocytes inhibit reactivation from latency.208,209,245–250 Release of granzyme B from CD8+ T cells into latently infected neurons helps to inhibit reactivation from latency by cleaving the viral transcriptional trans-activator, ICP4.251 Since it is well established that granzyme B activates caspase 3 and the intrinsic pathway of apoptosis,207 the ability of LAT and perhaps LR gene products to inhibit apoptosis is important to overcome the effects of granzyme B. The ability of HSV-1 to inhibit major histocompatibility complex (MHC) class I presentation in sensory neurons correlates with successful reactivation252 providing further evidence that CD8+ T cells monitor latently infected neurons. In conclusion, the ability of HSV-1, HSV-2, and BHV-1 to reactivate from latency is regulated by complex virus-host interactions.
Perspectives
Although genetic and functional studies have demonstrated that LAT and the LR gene regulate the latency-reactivation cycle, there are many unanswered questions. For example, identifying the functions of various transcripts, the small non-coding RNAs, and the ORFs within LAT are crucial to understand the role these various factors play in the latency-reactivation cycle. It would not be surprising to find that one or more of these factors encoded within the LAT locus regulate certain neuronal specific functions that maintain normal functions. It will be difficult to make additional LAT mutant viruses as many of these factors overlap and deletion of these sequences would likely interfere with expression of more than one LAT encoded factor. Consequently, many of these studies will have to be performed in transient transfection assays in primary neurons or neuroblastoma cells. Finally, examining LAT in small animal models in terms of weeks after acute infection may not accurately reflect the latency-reactivation cycle in the context of life-long latency in humans.
With respect to the LR gene, there are no studies that have determined whether ORF1, ORF-E, or the microRNAs play a role in the latency-reactivation cycle. Furthermore, functional analysis of ORF-1 and ORF-E has not been performed. Identifying the cellular proteins that interact with ORF-1 and ORF-E may provide insight into their functions. In summary, the finding that sequences encompassing LAT and the LR gene encode for more than one transcript and/or small non-coding RNAs implies many functions are necessary to successfully regulate the lifelong latency-reactivation in the natural host.
Footnotes
Author Contributions
Conceived and designed the experiments: CJ. Analyzed the data: CJ. Wrote the first draft of the manuscript: CJ. Contributed to the writing of the manuscript: CJ. Agree with manuscript results and conclusions: CJ. Jointly developed the structure and arguments for the paper: CJ. Made critical revisions and approved final version: CJ. All authors reviewed and approved of the final manuscript.
Funding
Research in the author’s laboratory is supported by a grant from the USDA, NIFA Competitive Grants Program (09-01653), a grant to the Nebraska Center for Virology (1P20RR15635), and the NE Research Initiative.
Competing Interests
Author disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication the author has provided signed confirmation of compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.
References
- 1.Smith RE, McDonald HR, Nesburn AB, Minckler DS. Penetrating keratoplasty: changing indications. Arch Ophthalmol. 1947 to 1978;1980;98(7):1226–9. doi: 10.1001/archopht.1980.01020040078009. [DOI] [PubMed] [Google Scholar]
- 2.Jones C. Herpes simplex virus type 1 and bovine herpesvirus 1 latency. Clin Micro Rev. 2003;16(1):79–95. doi: 10.1128/CMR.16.1.79-95.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wagner EK, Bloom DC. Experimental investigation of herpes simplex virus latency. Clin Microbiol Rev. 1997;10(3):419–43. doi: 10.1128/cmr.10.3.419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nesburn AB. Recurrent herpes simplex infection:pathogenesis and treatment. In: Barraquer J, Binder P, Buxton J, editors. Symposium on Medical and Surgical Disease of the Cornea. CV Mosby Company; St. Louis: 1980. pp. 48–68. [Google Scholar]
- 5.Nesburn AB, editor. Report of the Corneal Disease Panel: Vision Research: A National Plan 1983–1987. II. CV Mosby Co; St. Louis: 1983. [Google Scholar]
- 6.Nesburn AB, Ghiasi H, Wechsler SL. Ocular safety and efficacy of an HSV-1 gD vaccine during primary and latent infection. Invest Ophthalmol Vis Sci. 1990;31(8):1497–502. [PubMed] [Google Scholar]
- 7.Cohen J. Painful failure of promosing genital herpes vaccine. Science. 2010;330(6002):304. doi: 10.1126/science.330.6002.304. [DOI] [PubMed] [Google Scholar]
- 8.Stanberry LR, Spruance SL, Cunningham AL, et al. Glycoprotein-D-adjuvant vaccine to prevent genital herpes. N Engl J Med. 2002;347(21):1652–61. doi: 10.1056/NEJMoa011915. [DOI] [PubMed] [Google Scholar]
- 9.Jones C. Alphaherpesvirus latency: its role in disease and survival of the virus in nature. Adv. Virus Res. 1998;51:81–133. doi: 10.1016/s0065-3527(08)60784-8. [DOI] [PubMed] [Google Scholar]
- 10.Jones C. Regulation of innate immune responses by bovine herpesvirus 1 and infected cell protein 0. Viruses. 2009;1(2):255–75. doi: 10.3390/v1020255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jones C, Geiser V, Henderson G, et al. Functional analysis of bovine herpesvirus 1 (BHV-1) genes expressed during latency. Vet Micro. 2006;113:3–4. 199–210. doi: 10.1016/j.vetmic.2005.11.009. [DOI] [PubMed] [Google Scholar]
- 12.Carter JJ, Weinberg AD, Pollard A, Reeves R, Magnuson JA, Magnuson NS. Inhibition of T-lymphocyte mitogenic responses and effects on cell functions by bovine herpesvirus 1. J Virol. 1989;63(4):1525–30. doi: 10.1128/jvi.63.4.1525-1530.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Griebel P, Ohmann HB, Lawman MJ, Babiuk LA. The interaction between bovine herpesvirus type 1 and activated bovine T lymphocytes. J Gen Virol. 1990;71(Pt 2):369–77. doi: 10.1099/0022-1317-71-2-369. [DOI] [PubMed] [Google Scholar]
- 14.Griebel P, Qualtiere L, Davis WC, et al. T lymphocyte population dynamics and function following a primary bovine herpesvirus type-1 infection. Viral Immunol. 1987;1(4):287–304. doi: 10.1089/vim.1987.1.287. [DOI] [PubMed] [Google Scholar]
- 15.Griebel P, Qualtiere L, Davis WC, Lawman MJ, Babiuk LA. Bovine peripheral blood leukocyte subpopulation dynamics following a primary bovine herpesvirus-1 infection. Viral Immunol. 1987–1988;1(4):267–86. doi: 10.1089/vim.1987.1.267. [DOI] [PubMed] [Google Scholar]
- 16.Hariharan MJ, Nataraj C, Srikumaran S. Down regulation of murine MHC class I expression by bovine herpesvirus 1. Viral Immunol. 1993;6(4):273–84. doi: 10.1089/vim.1993.6.273. [DOI] [PubMed] [Google Scholar]
- 17.Hinkley S, Hill AB, Srikumaran S. Bovine herpesvirus-1 infection affects the peptide transport activity in bovine cells. Virus Res. 1998;53(1):91–6. doi: 10.1016/s0168-1702(97)00128-7. [DOI] [PubMed] [Google Scholar]
- 18.Koppers-Lalic EA, Reits EAJ, Ressing ME, et al. Varicelloviruses avoid T cell recognition by UL49.5-mediated inactivation of the transporter associated with antigen processing. Proc Natl Acad Sci U S A. 2005;102(14):5144–9. doi: 10.1073/pnas.0501463102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nataraj C, Eidmann S, Hariharan MJ, Sur JH, Perry GA, Srikumaran S. Bovine herpesvirus 1 downregulates the expression of bovine MHC class I molecules. Viral Immunol. 1997;10(1):21–34. doi: 10.1089/vim.1997.10.21. [DOI] [PubMed] [Google Scholar]
- 20.Eskra L, Splitter GA. Bovine herpesvirus-1 infects activated CD4+ lymphocytes. J Gen Virol. 1997;78(Pt 9):2159–66. doi: 10.1099/0022-1317-78-9-2159. [DOI] [PubMed] [Google Scholar]
- 21.Winkler MT, Doster A, Jones C. Bovine herpesvirus 1 can infect CD4(+) T lymphocytes and induce programmed cell death during acute infection of cattle. J Virol. 1999;73:8657–68. doi: 10.1128/jvi.73.10.8657-8668.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.da Silva LF, Sinani D, Jones C. The ICP27 protein encoded by bovine herpesvirus type 1 (bICP27) interferes with promoter activity of the bovine genes encoding beta interferon 1 (IFN-β1) and IFN-β3. Virus Res. 2012;169(1):162–8. doi: 10.1016/j.virusres.2012.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Henderson G, Zhang Y, Jones C. The bovine herpesvirus 1 gene encoding infected cell protein 0 (bICP0) can inhibit interferon-dependent transcription in the absence of other viral genes. J Gen Virol. 2005;86(Pt 10):2697–702. doi: 10.1099/vir.0.81109-0. [DOI] [PubMed] [Google Scholar]
- 24.Saira K, Zhou Y, Jones C. The infected cell protein 0 encoded by bovine herpesvirus 1 (bICP0) induces degradation of interferon response factor 3 (IRF3), and consequently inhibits beta interferon promoter activity. J Virol. 2007;81(7):3077–86. doi: 10.1128/JVI.02064-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Saira K, Jones C. The infected cell protein 0 encoded by bovine herpesvirus 1 (bICP0) associates with interferon regulatory factor 7 (IRF7), and consequently inhibits beta interferon promoter activity. J Virol. 2009;83(8):3977–81. doi: 10.1128/JVI.02400-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Highlander SK. Molecular genetic analysis of virulence in Mannheimia (Pasteurella) haemolytica. Front Biosci. 2001:D1128–50. doi: 10.2741/highland. [DOI] [PubMed] [Google Scholar]
- 27.Highlander SK, Fedorova ND, Dusek DM, Panciera R, Alvarez LE, Renehart C. Inactivation of Pasteurella (Mannheimia) haemolytica leukotoxin causes partial attenuation of virulence in a calf challenge model. Infect Immun. 2000;68(7):3916–22. doi: 10.1128/iai.68.7.3916-3922.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zecchinon L, Fett T, Desmecht D. How Mannheimia haemolytica defeats host defense through a kiss of death mechanism. Vet Res. 2005;36(2):133–56. doi: 10.1051/vetres:2004065. [DOI] [PubMed] [Google Scholar]
- 29.Spear PG. Herpes simplex virus: receptors and ligands for cell entry. Cell Microbiol. 2004;6(5):401–10. doi: 10.1111/j.1462-5822.2004.00389.x. [DOI] [PubMed] [Google Scholar]
- 30.Spear PG, Manoj S, Yoon M, Jogger CR, Zago A, Myscofski D. Different receptors binding to distinct interfaces on herpes simplex virus gD can trigger events leading to cell fusion and viral entry. Virology. 2006;344(1):17–24. doi: 10.1016/j.virol.2005.09.016. [DOI] [PubMed] [Google Scholar]
- 31.Spear PG, Longnecker R. Herpesvirus entry: an update. J Virol. 2003;77(19):10179–85. doi: 10.1128/JVI.77.19.10179-10185.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Montgomery RI, Warner MS, Lum BJ, Spear PG. Herpes simplex virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor family. Cell. 1996;87(3):427–36. doi: 10.1016/s0092-8674(00)81363-x. [DOI] [PubMed] [Google Scholar]
- 33.Geraghty RJ, Krummenacher C, Cohen GH, Eisenberg RJ, Spear PG. Entry of alphaherpesviruses mediated by poliovirus receptor-related protein 1 and poliovirus receptor. Science. 1998;280(5369):1618–20. doi: 10.1126/science.280.5369.1618. [DOI] [PubMed] [Google Scholar]
- 34.Honess RW, Roizman B. Regulation of herpes virus macromalecular synthesis: Cascade regulation of three groups of viral proteins. J Virol. 1974;14(1):8–19. doi: 10.1128/jvi.14.1.8-19.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.O’Hare P. The virion transactivator of herpes simplex virus. Seminars Virol. 1993;4:145–55. [Google Scholar]
- 36.Jones C, da Silva LF, Sinani D. Regulation of the latency-reactivation cycle by products encoded by the bovine herpesvirus 1 (BHV-1) latency-related gene. J Neurovirol. 2011;17(6):535–45. doi: 10.1007/s13365-011-0060-3. [DOI] [PubMed] [Google Scholar]
- 37.Maggioncalda J, Mehta A, Su YH, Fraser NW, Block TM. Correlation between herpes simplex virus type 1 rate of reactivation from latent infection and the number of infected neurons in trigeminal ganglia. Virology. 1996;225(1):72–81. doi: 10.1006/viro.1996.0576. [DOI] [PubMed] [Google Scholar]
- 38.Mehta A, Maggioncalda J, Bagasra O, et al. In situ DNA PCR and RNA hybridization detection of herpes simplex virus sequences in trigeminal ganglia of latently infected mice. Virology. 1995;206(1):633–40. doi: 10.1016/s0042-6822(95)80080-8. [DOI] [PubMed] [Google Scholar]
- 39.Ramakrishnan R, Fink DJ, Jiang G, Desai P, Glorioso JC, Levine M. Competitive quantitative PCR analysis of herpes simplex virus type 1 DNAand latency-associated transcript RNA in latently infected cells of the ratbrain. J Virol. 1994 Mar;68(3):1864–73. doi: 10.1128/jvi.68.3.1864-1873.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ramakrishnan R, Levine M, Fink DJ. PCR-based analysis of herpes simplex virus type 1 latency in the rat trigeminal ganglion established with a ribonucleotide reductase-deficient mutant. J Virol. 1994;68:7083–91. doi: 10.1128/jvi.68.11.7083-7091.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sawtell NM. Comprehensive quantification of herpes simplex virus latency at the single-cell level. J Virol. 1997;71(7):5423–31. doi: 10.1128/jvi.71.7.5423-5431.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Carrozza MJ, DeLuca NA. Interaction of the viral activator protein ICP4 with TFIID through TAF250. Mol Cell Biol. 1996;16(6):3085–93. doi: 10.1128/mcb.16.6.3085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.DeLuca NA McCarthy AM, PA Schaffer. Isolation and characterization of deletion mutants of herpes simplex virus type 1 in the gene encoding immediate-early regulatory protein ICP4. J Virol. 1985;56(2):558–70. doi: 10.1128/jvi.56.2.558-570.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.DeLuca NA Schaffer PA. Activation of immediate-early, early, and late promoters by temperature-sensitive and wild-type forms of herpes simplex virus type 1 protein ICP4. Mol Cell Biol. 1985;5(8):1997–208. doi: 10.1128/mcb.5.8.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Dixon RA, Schaffer PA. Fine-structure mapping and functional analysis of temperature-sensitive mutants in the gene encoding the herpes simplex virus type 1 immediate early protein VP175. J Virol. 1980;36(1):189–203. doi: 10.1128/jvi.36.1.189-203.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.McCarthy AM McMahan L, PA Schaffer. Herpes simplex virus type 1 ICP27 deletion mutants exhibit altered patterns of transcription and are DNA deficient. J Virol. 1989;63(1):18–27. doi: 10.1128/jvi.63.1.18-27.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.McMahan L Schaffer PA. The repressing and enhancing functions of the herpes simplex virus regulatory protein ICP27 map to C-terminal regions and are required to modulate viral gene expression very early in infection. J Virol. 1990;64(7):3471–85. doi: 10.1128/jvi.64.7.3471-3485.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sacks WR, Greene CC, Aschman DP, Schaffer PA. Herpes simplex virus type 1 ICP27 is an essential regulatory protein. J Virol. 1985;55(3):796–805. doi: 10.1128/jvi.55.3.796-805.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gu B, DeLuca N. Requirements for activation of the herpes simplex virus glycoprotein C promoter in vitro by the viral regulatory protein ICP4. J Virol. 1994;68(12):7953–65. doi: 10.1128/jvi.68.12.7953-7965.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gu B, Rivera-Gonzalez R, Smith CA, DeLuca NA. Herpes simplex virus infected cell polypeptide 4 preferentially represses Sp1-activated over basal transcription from its own promoter. Proc Natl Acad Sci U S A. 1993;90(20):9528–32. doi: 10.1073/pnas.90.20.9528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Michael N, Roizman B. Repression of the herpes simplex virus 1 alpha 4 gene by its gene product occurs within the context of the viral genome and is associated with all three identified cognate sites. Proc Natl Acad Sci U S A. 1993;90(6):2286–90. doi: 10.1073/pnas.90.6.2286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.O’Hare P Hayward GS. Three trans-acting regulatory proteins of herpes simplex virus modulate immediate-early gene expression in a pathway involving positive and negative feedback regulation. J Virol. 1985;56(3):723–33. doi: 10.1128/jvi.56.3.723-733.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Roberts MS, Boundy A, O’Hare P, Pizzorno MC, Ciufo DM, Hayward GS. Direct correlation between a negative autoregulatory response element at the cap site of the herpes simplex virus type 1 IE175 (alpha 4) promoter and a specific binding site for the IE175 (ICP4) protein. J Virol. 1988;62(11):4307–20. doi: 10.1128/jvi.62.11.4307-4320.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Smith CA, Bates P, Rivera-Gonzalez R, Gu B, DeLuca NA. ICP4, the major transcriptional regulatory protein of herpes simplex virus type 1, forms a tripartite complex with TATA-binding protein and TFIIB. J Virol. 1993;67(8):4676–87. doi: 10.1128/jvi.67.8.4676-4687.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hardwicke MA, Schaffer PA. Differential effects of nerve growth factor and dexamethasone on herpes simplex virus type 1 oriL- and oriS-dependent DNA replication in PC12 cells. J Virol. 1997;71(5):3580–7. doi: 10.1128/jvi.71.5.3580-3587.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hardy WR, Sandri-Goldin RM. Herpes simplex virus inhibits host cell splicing, and regulatory protein ICP27 is required for this effect. J Virol. 1994;68(12):7790–9. doi: 10.1128/jvi.68.12.7790-7799.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sandri-Goldin RM, Hibbard MK, Hardwicke MA. The C-terminal repressor region of herpes simplex virus type 1 ICP27 is required for the redistribution of small nuclear ribonucleoprotein particles and splicing factor SC35; however, these alterations are not sufficient to inhibit host cell splicing. J Virol. 1995;69(10):6063–76. doi: 10.1128/jvi.69.10.6063-6076.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sandri-Goldin RM, Mendoza GE. A herpesvirus regulatory protein appears to act post-transcriptionally by affecting mRNA processing. Genes Dev. 1992;6(5):848–63. doi: 10.1101/gad.6.5.848. [DOI] [PubMed] [Google Scholar]
- 59.Hill A, Jugovic P, York I, et al. Herpes simplex virus turns off the TAP to evade host immunity. Nature. 1995;375(6530):411–5. doi: 10.1038/375411a0. [DOI] [PubMed] [Google Scholar]
- 60.Goldsmith K, Chen W, Johnson DC, Hendricks RL. Infected cell protein (ICP)47 enhances herpes simplex virus neurovirulence by blocking the CD8+ T cell response. J Exp Med. 1998;187(3):341–8. doi: 10.1084/jem.187.3.341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Rice SA, Long MC, Lam V, Schaffer PA, Spencer CA. Herpes simplex virus immediate-early protein ICP22 is required for viral modification of host RNA polymerase II and establishment of the normal viral transcription program. J Virol. 1995;69(9):5550–9. doi: 10.1128/jvi.69.9.5550-5559.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Jordan R, Schaffer PA. Activation of gene expression by herpes simplex virus type 1 ICP0 occurs at the level of mRNA synthesis. J Virol. 1997;71(9):6850–62. doi: 10.1128/jvi.71.9.6850-6862.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kawaguchi Y, Bruni R, Roizman B. Interaction of herpes simplex virus 1 alpha regulatory protein ICP0 with elongation factor 1 delta: ICP0 affects translational machinery. J Virol. 1997;71(2):1019–24. doi: 10.1128/jvi.71.2.1019-1024.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kawaguchi Y, Van Sant C, Roizman B. Herpes simplex virus 1 alpha regulatory protein ICP0 interacts with and stabilizes the cell cycle regulator cyclin D3. J Virol. 1997;71(10):7328–36. doi: 10.1128/jvi.71.10.7328-7336.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Meredith M, Orr A, Elliott M, Everett R. Separation of sequence requirements for HSV-1 Vmw110 multimerisation and interaction with a 135-kDa cellular protein. Virology. 1995;209(1):174–87. doi: 10.1006/viro.1995.1241. [DOI] [PubMed] [Google Scholar]
- 66.Meredith M, Orr A, Everett R. Herpes simplex virus type 1 immediate-early protein Vmw110 binds strongly and specifically to a 135-kDa cellular protein. Virology. 1994;200(2):457–69. doi: 10.1006/viro.1994.1209. [DOI] [PubMed] [Google Scholar]
- 67.Everett R, O’Hare P, O’Rourke D, Barlow P, Orr A. Point mutations in the herpes simplex virus type 1 Vmw110 RING finger helix affect activation of gene expression, viral growth, and interaction with PML-containing nuclear structures. J Virol. 1995;69(11):7339–44. doi: 10.1128/jvi.69.11.7339-7344.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Everett RD, Lomonte P, Sternsdorf T, van Driel R, Orr A. Cell cycle regulation of PML modification and ND10 composition. J Cell Sci. 1999;112(Pt 24):4581–8. doi: 10.1242/jcs.112.24.4581. [DOI] [PubMed] [Google Scholar]
- 69.Everett RD, Meredith M, Orr A, Cross A, Kathoria M, Parkinson J. A novel ubiquitin-specific protease is dynamically associated with the PML nuclear domain and binds to a herpesvirus regulatory protein. EMBO J. 1997;16(7):1519–30. doi: 10.1093/emboj/16.7.1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hobbs WE, DeLuca NA. Perturbation of cell cycle progression and cellular gene expression as a function of herpes simplex virus ICP0. J Virol. 1999;73(10):8245–55. doi: 10.1128/jvi.73.10.8245-8255.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Poon AP, Liang Y, Roizman B. Herpes simplex virus 1 gene expression is accelerated by inhibitors of histone deacetylases in rabbit skin cellls infected with a mutant carrying a cDNA copy of the infected cell protein no. J Virol. 2003;77(23):12671–8. doi: 10.1128/JVI.77.23.12671-12678.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Lomonte P, Seigneurin-Berny D, Everett RD, Khochbin S, Epstein AL. Presented at the 26th International Herpesvirus Workshop; 2001. [Google Scholar]
- 73.Gu H, Liang Y, Mandel G, Roizman B. Components of the REST/CoREST/histone deacytlase repressor complex are disrupted, modified, and translocated in HSV-1-infected cells. Proc Nat Acad Sci U S A. 2005;102(21):7571–6. doi: 10.1073/pnas.0502658102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Poon APW, Gu H, Roizman B. ICP0 and the Us3 protein kinase of herpes simplex virus 1 independently block histone deactylation to enable gene expression. Proc Nat Acad Sci U S A. 2006;103(26):9993–8. doi: 10.1073/pnas.0604142103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Cliffe AR, Knipe DM. Herpes simplex virus ICP0 promotes both histone removal and acetylation on viral dna during lytic infection. J Virol. 2008;82(24):12030–8. doi: 10.1128/JVI.01575-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Cai W, Schaffer PA. A cellular function can enhance gene expression and plating efficiency of a mutant defective in the gene for ICP0, a transactivating protein of herpes simplex virus type 1. J Virol. 1991;65(8):4078–90. doi: 10.1128/jvi.65.8.4078-4090.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Hardwick JM. Viral interference with apoptosis. Semin Cell Dev Biol. 1998;9(3):339–49. doi: 10.1006/scdb.1998.0243. [DOI] [PubMed] [Google Scholar]
- 78.Razvi ES, Welsh RM. Apoptosis in viral infections. Adv Virus Res. 1995;45:1–60. doi: 10.1016/s0065-3527(08)60057-3. [DOI] [PubMed] [Google Scholar]
- 79.Shen Y, Shenk TE. Viruses and apoptosis. Curr Opin Genet Dev. 1995;5:105–11. doi: 10.1016/s0959-437x(95)90061-6. [DOI] [PubMed] [Google Scholar]
- 80.Teodoro JG, Branton PE. Regulation of apoptosis by viral gene products. J Virol. 1997;71(3):1739–46. doi: 10.1128/jvi.71.3.1739-1746.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Aiamkitsumrit B, Zhang X, Block TM, Norton P, Fraser NW, Su YH. Herpes simplex virus type 1 ICP4 deletion mutant virus d120 infection failed to induce apoptosis in nerve growth factor-differentiated PC12 cells. J Neurovirol. 2007;13(4):305–14. doi: 10.1080/13550280701361490. [DOI] [PubMed] [Google Scholar]
- 82.Devireddy LR, Jones C. Activation of caspases and p53 by bovine herpesvirus 1 infection results in programmed cell death and efficient virus release. J Virol. 1999;73(5):3778–88. doi: 10.1128/jvi.73.5.3778-3788.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Galvan V, Brandimarti R, Roizman B. Herpes simplex virus 1 blocks caspase-3-independent and caspase-dependent pathways to cell death. J Virol. 1999;73(4):3219–26. doi: 10.1128/jvi.73.4.3219-3226.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Galvan V, Roizman B. Herpes simplex virus 1 induces and blocks apoptosis at multiple steps during infection and protects cells from exogenous inducers in a cell-type-dependent manner. Proc Natl Acad Sci U S A. 1998;95(7):3931–6. doi: 10.1073/pnas.95.7.3931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Pradhan P, Nguyen ML. Early passage neonatal and adult keratinocytes are sensitive to apoptosis induced by infection with an ICP27-null mutant of herpes simplex virus 1. Apoptosis. 2013;18(2):160–70. doi: 10.1007/s10495-012-0773-7. [DOI] [PubMed] [Google Scholar]
- 86.Sadzot-Delvaux C, Thonard P, Schoonbroodt S, Piette J, Rentier B. Varicella-zoster virus induces apoptosis in cell culture. J Gen Virol. 1995;76(Pt 11):2875–9. doi: 10.1099/0022-1317-76-11-2875. [DOI] [PubMed] [Google Scholar]
- 87.Asano S, Honda T, Goshima F, et al. US3 protein kinase of herpes simplex virus type 2 plays a role in protecting corneal epithelial cells from apoptosis in infected mice. J Gen Virol. 1999;80(Pt 1):51–6. doi: 10.1099/0022-1317-80-1-51. [DOI] [PubMed] [Google Scholar]
- 88.Aubert M, Blaho JA. The herpes simplex virus type 1 regulatory protein ICP27 is required for the prevention of apoptosis in infected human cells. J Virol. 1999;73(4):2803–13. doi: 10.1128/jvi.73.4.2803-2813.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Leopardi R, Roizman B. The herpes simplex virus major regulatory protein ICP4 blocks apoptosis induced by the virus or by hyperthermia. Proc Natl Acad Sci U S A. 1996;93(18):9583–7. doi: 10.1073/pnas.93.18.9583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Geiser V, Rose S, Jones C. The bovine herpes virus 1 bICP0 protein regulates toxicity in a cell type dependent fashion. Molec Path. 2008;44:459–66. doi: 10.1016/j.micpath.2007.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Chenet-Monte C, Mohammad F, Celluzzi CM, Schaffer PA, Farber FE. Herpes simplex virus gene products involved in the induction of chromosomal aberrations. Virus Res. 1986;6(3):245–60. doi: 10.1016/0168-1702(86)90073-0. [DOI] [PubMed] [Google Scholar]
- 92.Ellison SA, Hampar B. Chromosomal aberrations induced by an animal virus. Nature. 1961;192:145–7. doi: 10.1038/192145a0. [DOI] [PubMed] [Google Scholar]
- 93.Heilbronn R, zur Hausen H. A subset of herpes simplex virus replication genes induces DNA amplification within the host cell genome. J Virol. 1989;63(9):3683–92. doi: 10.1128/jvi.63.9.3683-3692.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Pilon L, Langelier Y, Royal A. Herpes simplex virus type 2 mutagenesis: characterization of mutants induced at the hprt locus of nonpermissive XC cells. Mol Cell Biol. 1986;6(8):2977–83. doi: 10.1128/mcb.6.8.2977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Soengas MS, Alarcon RM, Yoshida H, et al. Apaf-1 and caspase-9 in p53-dependent apoptosis and tumor inhibition. Science. 1999;284(5411):156–9. doi: 10.1126/science.284.5411.156. [DOI] [PubMed] [Google Scholar]
- 96.Hagglund R, Munger J, Poon AP, Roizman B. U(S)3 protein kinase of herpes simplex virus 1 blocks caspase 3 activation induced by the products of U(S)1.5 and U(L)13 genes and modulates expression of transduced U(S)1.5 open reading frame in a cell type-specific manner. J Virol. 2002;76(2):743–54. doi: 10.1128/JVI.76.2.743-754.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Ahmed M, Fraser NW. Herpes simplex virus type 1 2-kilobase latency-associated transcript intron associates with ribosomal proteins and splicing factors. J Virol. 2001;75(24):12070–80. doi: 10.1128/JVI.75.24.12070-12080.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Blaho JA, Aubert M. Modulation of apoptosis during herpes simplex virus infection in human cells. Microbes Infect. 2001;3(10):859–66. doi: 10.1016/s1286-4579(01)01444-7. [DOI] [PubMed] [Google Scholar]
- 99.Jerome KR, Chen Z, Lang R, et al. HSV and Glycoprotein J inhibit caspase activation and apoptosis induced by Granzyme B or Fas. J Immunol. 2001;167(7):3928–35. doi: 10.4049/jimmunol.167.7.3928. [DOI] [PubMed] [Google Scholar]
- 100.Jerome KR, Fox R, Chen Z, Sears AE, Lee H, Corey L. Herpes simplex virus inhibits apoptosis through the action of two genes, Us5 and Us3. J Virol. 1999;73(11):8950–7. doi: 10.1128/jvi.73.11.8950-8957.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Jiang Y, Inman M, Zhang Y, Posadas NA, Jones C. A mutation in the latency related gene of bovine herpesvirus 1 (BHV-1) inhibits protein expression of a protein from open reading frame 2 (ORF-2) and an adjacent reading frame during productive infection. J Virol. 2004;78(6):3184–9. doi: 10.1128/JVI.78.6.3184-3189.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kather A, Raftery MJ, Devi-Rao G, et al. Herpes simplex virus type 1 (HSV-1)-induced apoptosis in human dendritic cells as a result of downregulation of cellular FLICE-inhibitory protein and reduced expression of HSV-1 antiapoptotic latency-associated transcript sequences. J Virol. 2010;84(2):1034–46. doi: 10.1128/JVI.01409-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Munger J, Chee AV, Roizman B. The U(S)3 protein kinase blocks apoptosis induced by the d120 mutant of herpes simplex virus 1 at a premitochondrial stage. J Virol. 2001;75(12):5491–7. doi: 10.1128/JVI.75.12.5491-5497.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Munger J, Roizman B. The US3 protein kinase of herpes simplex virus 1 mediates the posttranslational modification of BAD and prevents BAD-induced programmed cell death in the absence of other viral proteins. Proc Natl Acad Sci U S A. 2001;98(18):10410–5. doi: 10.1073/pnas.181344498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Nguyen ML, Blaho JA. Cellular players in the herpes simplex virus dependent apoptosis balancing act. Viruses. 2009;1(3):965–78. doi: 10.3390/v1030965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Croen KD, Ostrove JM, Dragovic LJ, Smialek JE, Straus SE. Latent herpes simplex virus in human trigeminal ganglia. Detection of an immediate early gene “anti-sense” transcript by in situ hybridization. N Engl J Med. 1987;317(23):1427–32. doi: 10.1056/NEJM198712033172302. [DOI] [PubMed] [Google Scholar]
- 107.Deatly AM, Spivack JG, Lavi E, O’Boyle DR, 2nd, Fraser NW. Latent herpes simplex virus type 1 transcripts in peripheral and central nervous system tissues of mice map to similar regions of the viral genome. J Virol. 1988;62(3):749–56. doi: 10.1128/jvi.62.3.749-756.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Deatly AM, Spivack JG, Lavi E, Fraser NW. RNA from an immediate early region of the type 1 herpes simplex virus genome is present in the trigeminal ganglia of latently infected mice. Proc Natl Acad Sci U S A. 1987;84(10):3204–8. doi: 10.1073/pnas.84.10.3204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Krause PR, Croen KD, Straus SE, Ostrove JM. Detection and preliminary characterization of herpes simplex virus type 1 transcripts in latently infected human trigeminal ganglia. J Virol. 1988;62(12):4819–23. doi: 10.1128/jvi.62.12.4819-4823.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Mitchell WJ, Lirette RP, Fraser NW. Mapping of low abundance latency-associated RNA in the trigeminal ganglia of mice latently infected with herpes simplex virus type 1. J Gen Virol. 1990;71(Pt 1):125–32. doi: 10.1099/0022-1317-71-1-125. [DOI] [PubMed] [Google Scholar]
- 111.Rock DL, Nesburn AB, Ghiasi H, et al. Detection of latency-related viral RNAs in trigeminal ganglia of rabbits latently infected with herpes simplex virus type 1. J Virol. 1987;61(12):3820–6. doi: 10.1128/jvi.61.12.3820-3826.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Stevens JG, Wagner EK, Devi-Rao GB, Cook ML, Feldman LT. RNA complementary to a herpesvirus alpha gene mRNA is prominent in latently infected neurons. Science. 1987;235(4792):1056–9. doi: 10.1126/science.2434993. [DOI] [PubMed] [Google Scholar]
- 113.Wagner EK, Devi-Rao G, Feldman LT, et al. Physical characterization of the herpes simplex virus latency-associated transcript in neurons. J Virol. 1988;62(4):1194–202. doi: 10.1128/jvi.62.4.1194-1202.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Wagner EK, Flanagan WM, Devi-Rao G, et al. The herpes simplex virus latency-associated transcript is spliced during the latent phase of infection. J Virol. 1988;62(12):4577–85. doi: 10.1128/jvi.62.12.4577-4585.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Perng GC, Ghiasi H, Slanina SM, Nesburn AB, Wechsler SL. The spontaneous reactivation function of the herpes simplex virus type 1 LAT gene resides completely within the first 1.5 kilobases of the 8.3-kilobase primary transcript. J Virol. 1996;70(2):976–84. doi: 10.1128/jvi.70.2.976-984.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Zwaagstra JC, Ghiasi H, Slanina SM, et al. Activity of herpes simplex virus type 1 latency-associated transcript (LAT) promoter in neuron-derived cells: evidence for neuron specificity and for a large LAT transcript. J Virol. 1990;64(10):5019–28. doi: 10.1128/jvi.64.10.5019-5028.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kang W, Mukerjee R, Fraser NF. Establishement and maintenance of HSV latent infection is mediated through correct splicing of the LAT primary transcript. Virology. 2003;312(1):233–44. doi: 10.1016/s0042-6822(03)00201-0. [DOI] [PubMed] [Google Scholar]
- 118.Mador N, Panet A, Latchman D, Steiner I. Expression and splicing of the latency-associated transcripts of herpes simplex virus type 1 in neuronal and non-neuronal cell lines. J Biochem (Tokyo) 1995;117(6):1288–97. doi: 10.1093/oxfordjournals.jbchem.a124857. [DOI] [PubMed] [Google Scholar]
- 119.Farrell MJ, Dobson AT, Feldman LT. Herpes simplex virus latency-associated transcript is a stable intron. Proc Natl Acad Sci U S A. 1991;88:790–4. doi: 10.1073/pnas.88.3.790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Krummenacher C, Zabolotny JM, Fraser NW. Selection of a nonconsensus branch point is influenced by an RNA stem-loop structure and is important to confer stability to the herpes simplex virus 2-kilobase latency-associated transcript. J Virol. 1997;71(8):5849–60. doi: 10.1128/jvi.71.8.5849-5860.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Nicosia M, Zabolotny JM, Lirette RP, Fraser NW. The HSV-1 2-kb latency-associated transcript is found in the cytoplasm comigrating with ribosomal subunits during productive infection. Virology. 1994;204(2):717–28. doi: 10.1006/viro.1994.1587. [DOI] [PubMed] [Google Scholar]
- 122.Thomas DL, Lock M, Zabolotny JM, Mohan BR, Fraser NW. The 2-kilobase intron of the herpes simplex virus type 1 latency-associated transcript has a half-life of approximately 24 hours in SY5Y and COS-1 cells. J Virol. 2002;76(2):532–40. doi: 10.1128/JVI.76.2.532-540.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Goldenberg D, Mador N, Ball MJ, Panet A, Steiner I. The abundant latency-associated transcripts of herpes simplex virus type 1 are bound to polyribosomes in cultured neuronal cells and during latent infection in mouse trigeminal ganglia. J Virol. 1997;71(4):2897–904. doi: 10.1128/jvi.71.4.2897-2904.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Dykxhoorn DM, Novina CD, Sharp PA. Killing the messenger: short RNAs that silence gene expression. Nat Rev Mol Cell Biol. 2003;4(6):457–67. doi: 10.1038/nrm1129. [DOI] [PubMed] [Google Scholar]
- 125.Hannon GJ. RNA interference. Nature. 2002;418(6894):244–51. doi: 10.1038/418244a. [DOI] [PubMed] [Google Scholar]
- 126.Kuwabera T, Hsieh J, Nakashima K, Taira K, Gage FH. A small modulatory dsRNA specified the fate of adult neural stem cells. Cell. 2004;116(6):779–93. doi: 10.1016/s0092-8674(04)00248-x. [DOI] [PubMed] [Google Scholar]
- 127.Xu P, Guo M, Hay BA. MicroRNAs and the regulation of cell death. Trends Genet. 2004;20(12):617–24. doi: 10.1016/j.tig.2004.09.010. [DOI] [PubMed] [Google Scholar]
- 128.Umbach JL, Kramer MF, Jurak I, Karnowski HW, Coen DM, Cullen BR. MicroRNAs expressed by herpes simplex virus 1 during latent infection regulate viral mRNAs. Nature. 2008;454(7205):780–5. doi: 10.1038/nature07103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Perng GC, Maguen B, Jing L, et al. A gene capable of blocking apoptosis can substitute for the herpes simplex virus type 1 latency-associated transcript gene and restore wild-type reactivation levels. J Virol. 2002;76(3):1224–35. doi: 10.1128/JVI.76.3.1224-1235.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Shen W, Sa e Silva MSE, Jaber T, et al. Two small RNAs encoded within the first 1.5 kb of the herpes simplex virus type 1 (HSV-1) latency-associated transcript (LAT) can inhibit productive infection, and cooperate to inhibit apoptosis. J Virol. 2009;83(18):9131–9. doi: 10.1128/JVI.00871-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Naito J, Mukerjee R, Mott KR, et al. Identification of a protein encoded in the herpes simplex virus type 1 latency associated transcripti promoter region. Virus Research. 2005;108:1–2. 101–10. doi: 10.1016/j.virusres.2004.08.011. [DOI] [PubMed] [Google Scholar]
- 132.Chan D, Cohen J, Naito J, et al. A mutant deleted for most of the herpes simplex virus type 1 (HSV-1) UOL gene does not affect the spontaneous reactivation phenotype in rabbits. J Neurovirology. 2006;12(1):5–16. doi: 10.1080/13550280500516401. [DOI] [PubMed] [Google Scholar]
- 133.Perng GC, Maguen B, Jing L, et al. A novel herpes simplex virus type 1 (HSV-1) transcript (AL-RNA) antisense to the 5′ end of LAT (latency associated transcript) produces a protein in infected rabbits. J Virol. 2002;76(16):8003–10. doi: 10.1128/JVI.76.16.8003-8010.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Jaber T, Henderson G, Li S, et al. Identification of a novel herpes simplex virus type 1 (HSV-1) transcript and protein (AL3) expressed during latency. J Gen Virol. 2009;90(Pt 10):2342–52. doi: 10.1099/vir.0.013318-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Block TM, Spivack JG, Steiner I, et al. A herpes simplex virus type 1 latency-associated transcript mutant reactivates with normal kinetics from latent infection. J Virol. 1990;64(7):3417–26. doi: 10.1128/jvi.64.7.3417-3426.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Ho DY, Mocarski ES. Beta-galactosidase as a marker in the peripheral and neural tissues of the herpes simplex virus-infected mouse. Virology. 1988;167(1):279–83. doi: 10.1016/0042-6822(88)90079-7. [DOI] [PubMed] [Google Scholar]
- 137.Sawtell NM, Thompson RL. Herpes simplex virus type 1 latency-associated transcription unit promotes anatomical site-dependent establishment and reactivation from latency. J Virol. 1992;66(4):2157–69. doi: 10.1128/jvi.66.4.2157-2169.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Thompson RL, Sawtell NM. The herpes simplex virus type 1 latency-associated transcript gene regulates the establishment of latency. J Virol. 1997;71(7):5432–40. doi: 10.1128/jvi.71.7.5432-5440.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Perng GC, Ghiasi H, Slanina SM, Nesburn AB, Wechsler SL. The spontaneous reactivation function of the herpes simplex virus type 1 LAT gene resides completely within the first 1.5 kilobases of the 8.3 kilobase primary transcript. J Virol. 1996;70(2):976–84. doi: 10.1128/jvi.70.2.976-984.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Chen SH, Kramer MF, Schaffer PA, Coen DM. A viral function represses accumulation of transcripts from productive-cycle genes in mouse ganglia latently infected with herpes simplex virus. J Virol. 1997;71(8):5878–84. doi: 10.1128/jvi.71.8.5878-5884.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Garber DA, Schaffer PA, Knipe DM. A LAT-associated function reduces productive-cycle gene expression during acute infection of murine sensory neurons with herpes simplex virus type 1. J Virol. 1997;71(8):5885–93. doi: 10.1128/jvi.71.8.5885-5893.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Perng GC, Dunkel EC, Geary PA, et al. The latency-associated transcript gene of herpes simplex virus type 1 (HSV-1) is required for efficient in vivo spontaneous reactivation of HSV-1 from latency. J Virol. 1994;68(12):8045–55. doi: 10.1128/jvi.68.12.8045-8055.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Perng GC, Slanina S, Ghiasi H, Nesburn AB, Wechsler SL. The effect of latency-associated transcript on the herpes simplex virus type 1 latency-reactivation phenotype is mouse strain-dependent. J Gen Virol. 2002;82(Pt 5):1117–22. doi: 10.1099/0022-1317-82-5-1117. [DOI] [PubMed] [Google Scholar]
- 144.Perng GC, Slanina AM, Yukht A, et al. A herpes simplex virus type 1 latency-associated transcript mutant with increased virulence and reduced spontaneous reactivation. J Virol. 1999;73(2):920–9. doi: 10.1128/jvi.73.2.920-929.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Perng GC, Slanina SM, Ghiasi H, Nesburn AB, Wechsler SL. A 371-nucleotide region between the herpes simplex virus type 1 (HSV-1) LAT promoter and the 2-kilobase LAT is not essential for efficient spontaneous reactivation of latent HSV-1. J Virol. 1996;70(3):2014–8. doi: 10.1128/jvi.70.3.2014-2018.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Hill JM, Sedarati F, Javier RT, Wagner EK, Stevens JG. Herpes simplex virus latent phase transcription facilitates in vivo reactivation. Virology. 1990;174(1):117–25. doi: 10.1016/0042-6822(90)90060-5. [DOI] [PubMed] [Google Scholar]
- 147.Trousdale MD, Steiner I, Spivack JG, et al. In vivo and in vitro reactivation impairment of a herpes simplex virus type 1 latency-associated transcript variant in a rabbit eye model. J Virol. 1991;65(12):6989–93. doi: 10.1128/jvi.65.12.6989-6993.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Drolet BS, Perng GC, Cohen J, et al. The region of the herpes simplex virus type 1 LAT gene involved in spontaneous reactivation does not encode a functional protein. Virology. 1998;242(1):221–32. doi: 10.1006/viro.1997.9020. [DOI] [PubMed] [Google Scholar]
- 149.Doerig C, Pizer LI, Wilcox CL. An antigen encoded by the latency-associated transcript in neuronal cell cultures latently infected with herpes simplex virus type 1. J Virol. 1991;65(5):2724–7. doi: 10.1128/jvi.65.5.2724-2727.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Lagunoff M, Roizman B. Expression of a herpes simplex virus 1 open reading frame antisense to the gamma(1)34.5 gene and transcribed by an RNA 3′ coterminal with the unspliced latency-associated transcript. J Virol. 1994;68(9):6021–8. doi: 10.1128/jvi.68.9.6021-6028.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Lock M, Miller C, Fraser NW. Analysis of protein expression from within the region encoding the 2.0-kilobase latency-associated transcript of herpes simplex virus type 1. J Virol. 2001;75(7):3413–26. doi: 10.1128/JVI.75.7.3413-3426.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Randall G, Lagunoff M, Roizman B. The product of ORF O located within the domain of herpes simplex virus 1 genome transcribed during latent infection binds to and inhibits in vitro binding of infected cell protein 4 to its cognate DNA site. Proc Natl Acad Sci U S A. 1997;94:10379–84. doi: 10.1073/pnas.94.19.10379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Thomas SK, Gough G, Latchman DS, Coffin RS. Herpes simplex virus latency-associated transcript encodes a protein which greatly enhances virus growth, can compensate for deficiencies in immediate-early gene expression, and is likely to function during reactivation from virus latency. J Virol. 1999;73(8):6618–25. doi: 10.1128/jvi.73.8.6618-6625.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Thomas SK, Lilley CE, Latchman DS, Coffin RS. A Protein Encoded by the Herpes Simplex Virus (HSV) Type 1 2-Kilobase Latency-Associated Transcript Is Phosphorylated, Localized to the Nucleus, and Overcomes the Repression of Expression from Exogenous Promoters When Inserted into the Quiescent HSV Genome. J Virol. 2002;76(8):4056–67. doi: 10.1128/JVI.76.8.4056-4067.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Perng GC, Esmail D, Slanina S, et al. Three herpes simplex virus type 1 latency-associated transcipt mutants with distinct and assymetric effects on virulence in mice compared with rabbits. J Virol. 2001;75(19):9018–28. doi: 10.1128/JVI.75.19.9018-9028.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Henderson G, Jaber T, Carpenter D, Wechsler SL, Jones C. Identification of herpes simplex virus type 1 (HSV-1) proteins encoded within the first 1.5 kb of the latency-associated transcript (LAT) J Neurovirology. 2009;15:5–6. 479–88. doi: 10.3109/13550280903296353. [DOI] [PubMed] [Google Scholar]
- 157.Rock DL, Beam SL, Mayfield JE. Mapping bovine herpesvirus type 1 latency-related RNA in trigeminal ganglia of latently infected rabbits. J Virol. 1987;61(12):3827–31. doi: 10.1128/jvi.61.12.3827-3831.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Devireddy L, Zhang Y, Jones C. Cloning and initial characterization of an alternatively spliced transcript encoded by the bovine herpes virus 1 latency related (LR) gene. J Neurovirology. 2003;9(6):612–22. doi: 10.1080/13550280390247542. [DOI] [PubMed] [Google Scholar]
- 159.Devireddy LR, Jones C. Alternative splicing of the latency-related transcript of bovine herpesvirus 1 yields RNAs containing unique open reading frames. J Virol. 1998;72(9):7294–301. doi: 10.1128/jvi.72.9.7294-7301.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Meyer F, Perez S, Geiser V, Sintek M, Inman M, Jones C. A protein encoded by the bovine herpes virus 1 (BHV-1) latency related gene interacts with specific cellular regulatory proteins, including the CCAAT enhancer binding protein alpha (C/EBP-a) J Virol. 2007;81(1):59–67. doi: 10.1128/JVI.01171-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Meyer F, Jones C. C/EBP-alpha cooperates with bTIF to activate the bovine herpesvirus 1 immediate early transcription unit 1 promoter. J Neuro Virology. 2008;2:1–8. [Google Scholar]
- 162.Workman A, Sinani D, Pittayakhajonwut D, Jones C. A Protein (ORF2) Encoded by the Latency Related Gene of Bovine Herpesvirus 1 Interacts with Notch1 and Notch3. J Virol. 2011;85:2536–46. doi: 10.1128/JVI.01937-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Bray SJ. Notch signalling: a simple pathway becomes complex. Nat Rev Mol Cell Biol. 2006;7(9):678–89. doi: 10.1038/nrm2009. [DOI] [PubMed] [Google Scholar]
- 164.Ehebauer M, Penelope P, Arias AM. Notch, a universal arbiter of cell fate decisions. Science. 2006;314(5804):1414–5. doi: 10.1126/science.1134042. [DOI] [PubMed] [Google Scholar]
- 165.Berezovska O, McLean P, Knowles R, et al. Notch1 inhibits neurite outgrowth in postmitotic primary neurons. Neuroscience. 1999;93(2):433–9. doi: 10.1016/s0306-4522(99)00157-8. [DOI] [PubMed] [Google Scholar]
- 166.Cornell R, Eisen JS. Notch in the pathway: the roles of Notch signalling in neural crest development. Semin Cell Dev Biol. 2005;16(6):663–72. doi: 10.1016/j.semcdb.2005.06.009. [DOI] [PubMed] [Google Scholar]
- 167.Justice NJ, Jan YN. Variations on the Notch pathway in neural development. Curr Op Neurobiol. 2002;12(1):64–70. doi: 10.1016/s0959-4388(02)00291-x. [DOI] [PubMed] [Google Scholar]
- 168.Wang T, Holt CM, Xu C, et al. Notch 3 activation modulates growth behavior and cross talks to Wnt/TCF signalling pathway. Cell Signal. 2007;19(12):2458–67. doi: 10.1016/j.cellsig.2007.07.019. [DOI] [PubMed] [Google Scholar]
- 169.Nair P, Somasundaram K, Krishna S. Activated Notch1 inhibits p53-induced apoptosis and sustains transformation by human papilloma virus type 16 E6 and E7 oncogenes through a PI3K-PKB/Akt-dependent pathway. J Virol. 2003;77(12):7106–12. doi: 10.1128/JVI.77.12.7106-7112.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Sade H, Krishna S, Sarin A. The anti-apoptotic effect of Notch-1 requires p56lck-dependent, AKT/PKB-mediated signaling in T cells. J Biol Chem. 2004;279(4):2937–44. doi: 10.1074/jbc.M309924200. [DOI] [PubMed] [Google Scholar]
- 171.Franklin JL, Berechid BE, Cutting FB, et al. Autonomous and non-autonomous regulation of mammalian neurite development by Notch1 and Delta1. Curr Biol. 1999;9(24):1448–57. doi: 10.1016/s0960-9822(00)80114-1. [DOI] [PubMed] [Google Scholar]
- 172.Levy OA, Lah JJ, Levy AI. Notch signaling inhibits PC12 cell neurite outgrowth via RBP-J-dependent and -independent mechanisms. Dev Neurosci. 2002;24:79–88. doi: 10.1159/000064948. [DOI] [PubMed] [Google Scholar]
- 173.Levy OA, Lah JJ, Levey AI. Notch signaling inhibits PC12 cell neurite outgrowth via RBP-J-dependent and -independent mechanisms. Dev Neurosc. 2002;24:79–88. doi: 10.1159/000064948. [DOI] [PubMed] [Google Scholar]
- 174.Sestan N, Artavanis-Tsakonas S, Rakic P. Contact-dependent inhibition of cortical neurite growth mediated by Notch signaling. Science. 1999;286(5440):741–6. doi: 10.1126/science.286.5440.741. [DOI] [PubMed] [Google Scholar]
- 175.Coleman MP, Freeman MR. Wallerian degeneration, wld(S), and nmnat. Annu Rev Neurosci. 2010;33:245–67. doi: 10.1146/annurev-neuro-060909-153248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Raff MC, Whitmore AV, Finn JT. Axonal self-destruction and neurodegeneration. Science. 2002;296(5569):868–71. doi: 10.1126/science.1068613. [DOI] [PubMed] [Google Scholar]
- 177.Yang Y, Klein R, Tian X, Cheng HT, Kopan R, Shen J. Notch activation induces apoptosis in neural progenitor cells through a p53-dependent pathway. Developmental Biol. 2004;269(1):81–04. doi: 10.1016/j.ydbio.2004.01.014. [DOI] [PubMed] [Google Scholar]
- 178.El Bejjani R Hammarlund M. Notch signaling inhibits axon regeneration. Neuron. 2012;73(2):268–78. doi: 10.1016/j.neuron.2011.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Sinani D, Frizzo da Silva L, Jones C. A bovine herpesvirus 1 protein expressed in latently infected neurons (ORF2) promotes neurite sprouting in the presence of activated Notch1 or Notch3. J Virol. 2013;87(2):1183–92. doi: 10.1128/JVI.02783-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Bratanich AC, Hanson ND, Jones C. The latency-related gene of bovine herpesvirus 1 inhibits the activity of immediate-early transcription unit 1. Virology. 1992;191(2):988–91. doi: 10.1016/0042-6822(92)90278-w. [DOI] [PubMed] [Google Scholar]
- 181.Geiser V, Inman M, Zhang Y, Jones C. The latency related (LR) gene of bovine herpes virus 1 (BHV-1) can inhibit the ability of bICP0 to activate productive infection. J Gen Virol. 2002;83:2965–71. doi: 10.1099/0022-1317-83-12-2965. [DOI] [PubMed] [Google Scholar]
- 182.Geiser V, Jones C. The latency related gene encoded by bovine herpesvirus 1 encodes a small regulatory RNA that inhibits cell growth. J Neurovirol. 2005;11:563–70. doi: 10.1080/13550280500385286. [DOI] [PubMed] [Google Scholar]
- 183.Schang LM, Hossain A, Jones C. The latency-related gene of bovine herpesvirus 1 encodes a product which inhibits cell cycle progression. J Virol. 1996;70(6):3807–14. doi: 10.1128/jvi.70.6.3807-3814.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Jaber T, Workman A, Jones C. Small noncoding RNAs encoded within the bovine herpesvirus 1 latency-related gene can reduce steady-state levels of infected cell protein 0 (bICP0) J Virol. 2010;84(13):6297–307. doi: 10.1128/JVI.02639-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Inman M, Zhou J, Webb H, Jones C. Identification of a novel transcript containing a small open reading frame that is expressed during latency, and is antisense to the latency related gene of bovine herpes virus 1 (BHV-1) J Virol. 2004;78(10):5438–47. doi: 10.1128/JVI.78.10.5438-5447.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Bratanich AC, Jones CJ. Localization of cis-acting sequences in the latency-related promoter of bovine herpesvirus 1 which are regulated by neuronal cell type factors and immediate-early genes. J Virol. 1992;66(10):6099–106. doi: 10.1128/jvi.66.10.6099-6106.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Delhon G, Jones C. Identification of DNA sequences in the latency related promoter of bovine herpes virus type 1 which are bound by neuronal specific factors. Virus Res. 1997;51(1):93–103. doi: 10.1016/s0168-1702(97)00085-3. [DOI] [PubMed] [Google Scholar]
- 188.Jones C, Delhon G, Bratanich A, Kutish G, Rock D. Analysis of the transcriptional promoter which regulates the latency-related transcript of bovine herpesvirus 1. J Virol. 1990;64(3):1164–70. doi: 10.1128/jvi.64.3.1164-1170.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Perez S, Meyer F, Henderson G, et al. A protein encoded by the bovine herpesvirus 1 ORF E gene induces neurite-like morphological changes in mouse neuroblastoma cells and is expressed in trigeminal ganglionic neurons. J Neuro Virology. 2007;13(2):139–49. doi: 10.1080/13550280701191459. [DOI] [PubMed] [Google Scholar]
- 190.Ahmed M, Lock M, Miller CG, Fraser NW. Regions of the herpes simplex virus type 1 latency-associated transcript that protect cells from apoptosis in vitro and protect neuronal cells in vivo. J Virol. 2002;76(2):717–29. doi: 10.1128/JVI.76.2.717-729.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Inman M, Perng CG, Henderson G, et al. Region of herpes simplex virus type 1 latency-associated transcript sufficient for wild-type spontaneous reactivation promotes cell survival in tissue culture. J Virol. 2001;75(8):3636–46. doi: 10.1128/JVI.75.8.3636-3646.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Perng GC, Jones C, Ciacci-Zanella J, et al. Virus-induced neuronal apoptosis blocked by the herpes simplex virus latency-associated transcript (LAT) Science. 2000;287(5457):1500–3. doi: 10.1126/science.287.5457.1500. [DOI] [PubMed] [Google Scholar]
- 193.Jin L, Peng W, Perng GC, Nesburn AB, Jones C, Wechsler SL. Identification of herpes simplex virus type 1 (HSV-1) latency associated transcript (LAT) sequences that both inhibit apoptosis and enhance the spontaneous reactivation phenotype. J Virol. 2003;77(11):6556–61. doi: 10.1128/JVI.77.11.6556-6561.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Jin L, Carpenter D, Moerdyk-Schauwecker M, et al. Cellular FLIP can substitute for the herpes simplex virus type 1 LAT gene to support a wild type virus reactivation phenotype in mice. J Neurovirology. 2008;14(5):389–400. doi: 10.1080/13550280802216510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Jin L, Perng GC, Nesburn AB, Jones C, Wechsler SL. The baculovirus inhibitor of apoptosis gene (cpIAP) can restore reactivation of latency to a herpes simplex virus type 1 that does not express the latency associated transcript. J Virol. 2005;79(19):12286–95. doi: 10.1128/JVI.79.19.12286-12295.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Mott K, Osorio N, Jin L, et al. The bovine herpesvirus 1 LR ORF2 is crucial for this gene’s ability to restore the high reactivation phenotype to a Herpes simplex virus-1 LAT null mutant. J Gen Virol. 2003;84:2975–85. doi: 10.1099/vir.0.19421-0. [DOI] [PubMed] [Google Scholar]
- 197.Henderson G, Peng W, Jin L, et al. Regulation of caspase 8- and caspase 9-induced apoptosis by the herpes simplex virus latency-associated transcript. J Neurovirol. 2002;8(Suppl 2):103–11. doi: 10.1080/13550280290101085. [DOI] [PubMed] [Google Scholar]
- 198.Kruegger A, Baumann S, Krammer PH, Kirchhoff S. FLICE-Inhibitory proteins: regulators of death receptor-mediated apoptosis. Molec Cell Biol. 2001;21(24):8247–54. doi: 10.1128/MCB.21.24.8247-8254.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Schmitz I, Kirchhoff S, Krammer PH. Regulation of death receptor-mediated apoptosis pathways. Int J Biochem Cell Biol. 2000;32:11–12. 1123–36. doi: 10.1016/s1357-2725(00)00048-0. [DOI] [PubMed] [Google Scholar]
- 200.Wang X. The expanding role of mitochondria in apoptosis. Gen Devel. 2001;15(22):2922–33. [PubMed] [Google Scholar]
- 201.Carpenter D, Hsiang C, Jin L, et al. Stable cell lines expressing high levels of the herpes simplex virus type 1 LAT are refractory to caspase 3 activation and DNA laddering following cold shock induced apoptosis. Virology. 2007;369(1):12–8. doi: 10.1016/j.virol.2007.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Carpenter D, Henderson G, Hsiang C, et al. Introducing point mutations into the ATGs of the putative open reading frame of the HSV-1 gene encoding the latency associated transcript (LAT) reduces its anti-apoptotis activity. Microb Pathog. 2008;44(2):98–102. doi: 10.1016/j.micpath.2007.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Jiang X, Chentoufi A, Hsiang C, et al. The herpes simplex virus type 1 latency associated transcript (LAT) can protect cells from Granzyme B induced apoptosis and CD8 T-cell killing. J Virol. 2011;85(5):2325–32. doi: 10.1128/JVI.01791-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Alimonti JB, Shi L, Baijal PK, Greenberg AH. Granzyme B induces BID-mediated cytochrome c release and mitochondrial permeability transition. J Biol Chem. 2001;276(10):6974–82. doi: 10.1074/jbc.M008444200. [DOI] [PubMed] [Google Scholar]
- 205.Barry M, Heibein JA, Pinkoski MJ, et al. Granzyme B short-circuits the need for caspase 8 activity during granule-mediated cytotoxic T-lymphocyte killing by directly cleaving Bid. Mol Cell Biol. 2000;20(11):3781–94. doi: 10.1128/mcb.20.11.3781-3794.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Pinkoski MJ, Waterhouse NJ, Heibein JA, et al. Granzyme B-mediated apoptosis proceeds predominantly through a Bcl-2-inhibitable mitochondrial pathway. J Biol Chem. 2001;276(15):12060–7. doi: 10.1074/jbc.M009038200. [DOI] [PubMed] [Google Scholar]
- 207.Yang X, Stennicke HR, Wang B, et al. Granzyme B mimics apical caspases. Description of a unified pathway for trans-activation of executioner cas-pase-3 and -7. J Biol Chem. 1998;273:34278–83. doi: 10.1074/jbc.273.51.34278. [DOI] [PubMed] [Google Scholar]
- 208.Nash AA, Jayasuriya A, Phelan J, Cobbold SP, Waldmann H, Prospero T. Different roles for L3T4+ and Lyt 2+ T cell subsets in the control of an acute herpes simplex virus infection of the skin and nervous system. J Gen Virol. 1987;68(Pt 3):825–33. doi: 10.1099/0022-1317-68-3-825. [DOI] [PubMed] [Google Scholar]
- 209.Simmons A, Tscharke D, Speck P. The role of immune mechanisms in control of herpes simplex virus infection of the peripheral nervous system. Curr Top Microbiol Immunol. 1992;179:31–56. doi: 10.1007/978-3-642-77247-4_3. [DOI] [PubMed] [Google Scholar]
- 210.Meyer F, Perez S, Jiang Y, Zhou Y, Henderson G, Jones C. Identification of a novel protein encoded by the latency-related gene of bovine herpesvirus 1. J Neuro Virology. 2007;13(6):569–78. doi: 10.1080/13550280701620754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Inman M, Lovato L, Doster A, Jones C. A mutation in the latency related gene of bovine herpesvirus 1 interferes with the latency-reactivation cycle of latency in calves. J Virol. 2002;76(13):6771–9. doi: 10.1128/JVI.76.13.6771-6779.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Inman M, Lovato L, Doster A, Jones C. A mutation in the latency-related gene of bovine herpesvirus 1 leads to impaired ocular shedding in acutely infected calves. J Virol. 2001;75(18):8507–15. doi: 10.1128/JVI.75.18.8507-8515.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Lovato L, Inman M, Henderson G, Doster A, Jones C. Infection of cattle with a bovine herpesvirus 1 (BHV-1) strain that contains a mutation in the latency related gene leads to increased apoptosis in trigeminal ganglia during the transition from acute infection to latency. J Virol. 2003;77(8):4848–57. doi: 10.1128/JVI.77.8.4848-4857.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Ciacci-Zanella J, Stone M, Henderson G, Jones C. The latency-related gene of bovine herpesvirus 1 inhibits programmed cell death. J Virol. 1999;73(12):9734–40. doi: 10.1128/jvi.73.12.9734-9740.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Henderson G, Perng GC, Nesburn A, Wechsler S, Jones C. The latency related gene of bovine herpesvirus 1 can suppress caspase 3 and caspase 9 during productive infection. J Neurovirol. 2004;10(1):64–70. doi: 10.1080/13550280490261716. [DOI] [PubMed] [Google Scholar]
- 216.Shen W, Jones C. Open reading frame 2 encoded by the latency related gene of bovine herpesvirus 1 has anti-apoptosis activity in transiently transfected neuroblastoma cells. J Virol. 2008;82:10940–5. doi: 10.1128/JVI.01289-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Sinani D, Jones C. Localization of sequences in a protein encoded by the latency related gene of bovine herpesvirus 1 (ORF2) that inhibits apoptosis and interferes with Notch1 mediated trans-activation of the bICP0 promoter. J Virol. 2011;85(23):12124–33. doi: 10.1128/JVI.05478-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Silva LF, Jones C. Two microRNAs encoded within the BHV-1 latency related (LR) gene promote cell survival by interacting with RIG-I and stimulating nuclear factor-kappa B (NF-kB) dependent transcription and beta-interferon signaling pathways. J Virol. 2012;86(3):1670–82. doi: 10.1128/JVI.06550-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Shen W, Jones C. Open reading frame 2, encoded by the latency-related gene of bovine herpesvirus 1, has antiapoptotic activity in transiently transfected neuroblastoma cells. J Virol. 2008;82(21):10940–5. doi: 10.1128/JVI.01289-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Antwerp DJ, Martin SJ, Kafri T, Green DR, Verma IM. Suppression of TNF-alpha-induced apoptosis by NF-kB. Science. 1996;274(5288):787–9. doi: 10.1126/science.274.5288.787. [DOI] [PubMed] [Google Scholar]
- 221.Foehr ED, Lin X, O’Mahony A, Geleziunas R, Bradshaw RA, Greene WC. NF-kB signaling promotes both cell survival and neurite process formation in nerver growth factor-stimulated PC12 cells. J Neuroscience. 2000;20(20):7556–63. doi: 10.1523/JNEUROSCI.20-20-07556.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Mattson MP, Meffert MK. Roles for NF-kB in nerve cell survival, plasticity, and disease. Cell Death Differentiation. 2006;13(5):852–60. doi: 10.1038/sj.cdd.4401837. [DOI] [PubMed] [Google Scholar]
- 223.Yang CH, Murti A, Pfeffer SR, Basu L, Kim JG, Pfeffer LM. IFNalpha/beta promotes cell survival by activating NF-kappa B. Proc Natl Acad Sci U S A. 2000;97(25):13631–6. doi: 10.1073/pnas.250477397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Ozaki N, Sugiura Y, Yamamoto M, Yokoya S, Wanaka A, Nishiyama Y. Apoptosis induced in the spinal cord and dorsal root ganglion by infection of herpes simplex virus type 2 in the mouse. Neurosci Lett. 1997;228:99–102. doi: 10.1016/s0304-3940(97)00364-9. [DOI] [PubMed] [Google Scholar]
- 225.Knotts FB, Cook ML, Stevens JG. Pathogenesis of herpetic encephalitis in mice after ophthalmic inoculation. J Infect Dis. 1974;130(1):16–27. doi: 10.1093/infdis/130.1.16. [DOI] [PubMed] [Google Scholar]
- 226.Kramer MF, Chen SH, Knipe DM, Coen DM. Accumulation of viral transcripts and DNA during establishment of latency by herpes simplex virus. J Virol. 1998;72(2):1177–85. doi: 10.1128/jvi.72.2.1177-1185.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Speck PG, Simmons A. Divergent molecular pathways of productive and latent infection with a virulent strain of herpes simplex virus type 1. J Virol. 1991;65(8):4001–5. doi: 10.1128/jvi.65.8.4001-4005.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Schang L, Jones C. Analysis of bovine herpesvirus 1 transcripts during a primary infection of trigeminal ganglia of cattle. J Virol. 1997;71(9):6786–95. doi: 10.1128/jvi.71.9.6786-6795.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Li S, Carpenter D, Hsiang C, Wechsler SL, Jones C. The herpes simplex virus type 1 latency-associated transcript (LAT) locus inhibits apoptosis and promotes neurite sprouting in neuroblastoma cells following serum starvation by maintaining active AKT (protein kinase B) J Gen Virol. 2010;91:858–66. doi: 10.1099/vir.0.015719-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Branco FJ, Fraser NW. Herpes simplex virus type 1 latency-associated transcript expression protects trigeminal ganglion neurons from apoptosis. J Virol. 1993;79(14):9019–25. doi: 10.1128/JVI.79.14.9019-9025.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Jones C, Chowdhury S. A review of the biology of bovine herpesvirus type 1 (BHV-1), its role as a cofactor in the bovine respiratory disease complex, and development of improved vaccines. Adv Anim Health. 2007;8(2):187–205. doi: 10.1017/S146625230700134X. [DOI] [PubMed] [Google Scholar]
- 232.Perng GC, Jones C. Towards an understanding of the Herpes Simplex Virus Type 1 latency-reactivation cycle. Interdiscip Perspect Infect Dis. 2010;2010:262415. doi: 10.1155/2010/262415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Joels M, de Kloet ER. Control of neuronal excitability by corticosteroid hormones. Trends Neurosci. 1992;15(1):25–30. doi: 10.1016/0166-2236(92)90345-9. [DOI] [PubMed] [Google Scholar]
- 234.McEwen BS. Non-genomic and genomic effects of steroids on neural activity. Trends Pharmacol Sci. 1991;12(4):141–7. doi: 10.1016/0165-6147(91)90531-v. [DOI] [PubMed] [Google Scholar]
- 235.Halford WP, Gebhardt BM, Carr DJ. Mechanisms of herpes simplex virus type 1 reactivation. J Virol. 1996;70(8):5051–60. doi: 10.1128/jvi.70.8.5051-5060.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Collett JW, Steele RE. Alternative splicing of a neural-specific Src mRNA (Src+) is a rapid and protein synthesis-independent response to neural induction in Xenopus laevis. Dev Biol. 1993;158(2):487–95. doi: 10.1006/dbio.1993.1206. [DOI] [PubMed] [Google Scholar]
- 237.Du T, Zhou G, Roizman B. Induction of apoptosis accelerates reactivation from latent HSV-1 in ganglionic organ cultures and replication in cell cultures. Proc Natl Acad Sci U S A. 2012;109(36):14616–21. doi: 10.1073/pnas.1212661109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Hunsperger EA, Wilcox CL. Caspase-3-dependent reactivation of latent herpes simplex virus type 1 in sensory neuronal cultures. J Neurovirology. 2003;9(3):390–8. doi: 10.1080/13550280390201678. [DOI] [PubMed] [Google Scholar]
- 239.Dieken ES, Miesfeld RL. Transcriptional transactivation functions localized to the glucocorticoid receptor N terminus are necessary for steroid induction of lymphocyte apoptosis. Mol Cell Biol. 1992;12(2):589–97. doi: 10.1128/mcb.12.2.589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Shimeld C, Easty DL, Hill TJ. Reactivation of herpes simplex virus type 1 in the mouse trigeminal ganglion: an in vivo study of virus antigen and cytokines. J Virol. 1999;73(3):1767–73. doi: 10.1128/jvi.73.3.1767-1773.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Shimeld C, Hill TJ, Blyth WA, Easty DL. Reactivation of latent infection and induction of recurrent herpetic eye disease in mice. J Gen Virol. 1990;71(2):397–404. doi: 10.1099/0022-1317-71-2-397. [DOI] [PubMed] [Google Scholar]
- 242.Shimeld C, Whiteland JL, Williams NA, Easty DL, Hill TJ. Reactivation of herpes simplex virus type 1 in the mouse trigeminal ganglion: an in vivo study of virus antigen and immune cell infiltration. J Gen Virol. 1996;77(3):2583–90. doi: 10.1099/0022-1317-77-10-2583. [DOI] [PubMed] [Google Scholar]
- 243.Feldman LT, Ellison AR, Voytek CC, Yang L, Krause P, Margolis TP. Spontaneous molecular reactivation of herpes simplex virus type 1 latency in mice. Proc Natl Acad Sci U S A. 2002;99(2):978–83. doi: 10.1073/pnas.022301899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Rock D, Lokensgard J, Lewis T, Kutish G. Characterization of dexamethasone-induced reactivation of latent bovine herpesvirus 1. J Virol. 1992;66(4):2484–90. doi: 10.1128/jvi.66.4.2484-2490.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Khanna KM, Bonneau RH, Kinchington PR, Hendricks RL. Herpes simplex virus-specific memory CD8+ T cells are selectively activated and retained in latently infected sensory ganglia. Immunity. 2003;18(5):593–603. doi: 10.1016/s1074-7613(03)00112-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Liu T, Khanna KM, Carriere BN, Hendricks RL. Gamma interferon can prevent herpes simplex virus type 1 reactivation from latency in sensory neurons. J Virol. 2001;75(22):11178–84. doi: 10.1128/JVI.75.22.11178-11184.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Liu T, Khanna KM, Chen X, Fink DJ, Hendricks RL. CD8(+) T cells can block herpes simplex virus type 1 (HSV-1) reactivation from latency in sensory neurons. J Exp Med. 2000;191(9):1459–66. doi: 10.1084/jem.191.9.1459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Liu T, Tang Q, Hendricks RL. Inflammatory infiltration of the trigeminal ganglion after herpes simplex virus type 1 corneal infection. J Virol. 1996;70(1):264–71. doi: 10.1128/jvi.70.1.264-271.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Prbhakaran K, Sheridan BS, Kinchington PR, et al. Sensory neurons regulate the effector functions of CD8+ T cells in controlling HSV-1 latency ex vivo. Immunity. 2005;23(5):515–23. doi: 10.1016/j.immuni.2005.09.017. [DOI] [PubMed] [Google Scholar]
- 250.Simmons A, Tscharke DC. Anti-CD8 impairs clearance of herpes simplex virus from the nervous system: implications for the fate of virally infected neurons. J Exp Med. 1992;175(5):1337–44. doi: 10.1084/jem.175.5.1337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Knickelbein JE, Khanna KM, Yee MB, Baty CJ, Kinchington PR, Hendricks RL. Noncytotoxic lytic granule-mediated CD8+ T cell inhibition of HSV-1 reactivation from neuronal latency. Science. 2008;322(5899):268–72. doi: 10.1126/science.1164164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Orr MT, Mathis MA, Lagunoff M, Sacks JA, Wilson CB. CD8 T cell control og HSV reactivation from latency is abrogated by viral inhibition of MHC Class I. Cell Host Microbe. 2007;2(3):172–80. doi: 10.1016/j.chom.2007.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Chou J, Kern ER, Whitley RJ, Roizman B. Mapping of herpes simplex virus-1 neurovirulence to gamma 134.5, a gene nonessential for growth in culture. Science. 1990;250(4985):1262–6. doi: 10.1126/science.2173860. [DOI] [PubMed] [Google Scholar]
- 254.Chou J, Roizman B. The herpes simplex virus 1 gene for ICP34.5, which maps in inverted repeats, is conserved in several limited-passage isolates but not in strain 17 syn+ J Virol. 1990;64(3):1014–20. doi: 10.1128/jvi.64.3.1014-1020.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Yeh L, Schaffer PA. A novel class of transcripts expressed with late kinetics in the absence of ICP4 spans the junction between the long and short segments of the herpes simplex virus type 1 genome. J Virol. 1993;67(12):7373–82. doi: 10.1128/jvi.67.12.7373-7382.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Bohenzky RA, Lagunoff M, Roizman B, Wagner EK, Silverstein S. Two overlapping transcription units which extend across the L-S junction of herpes simplex virus type 1. J Virol. 1995;69(5):2889–97. doi: 10.1128/jvi.69.5.2889-2897.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Bohenzky RA, Papavassiliou AG, Gelman IH, Silverstein S. Identification of a promoter mapping within the reiterated sequences that flank the herpes simplex virus type 1 UL region. J Virol. 1993;67(2):632–42. doi: 10.1128/jvi.67.2.632-642.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Peng W, Vitvitskaia O, Carpenter D, Wechsler SL, Jones C. Identification of two small RNAs within the first 1.5-kb of the herpes simplex virus type 1 (HSV-1) encoded latency-associated transcript (LAT) J Neuro Virology. 2008;14(1):41–52. doi: 10.1080/13550280701793957. [DOI] [PubMed] [Google Scholar]
- 259.Hossain A, Schang LM, Jones C. Identification of gene products encoded by the latency-related gene of bovine herpesvirus 1. J Virol. 1995;69(9):5345–52. doi: 10.1128/jvi.69.9.5345-5352.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]