

Abstract
Recurrent respiratory papillomatosis (RRP) is a rare, chronic disease caused by human papillomaviruses (HPVs) types 6 and 11 that is characterized by the polarization of adaptive immune responses that support persistent HPV infection. Respiratory papillomas express elevated mRNA levels of IL-36γ, a proinflammatory cytokine in comparison to autologous clinically normal laryngeal tissues; however there is no evidence of inflammation in these lesions. Consistent with this, respiratory papillomas do not contain TH1-like CD4+ T-cells or cytotoxic CD8+ T-cells, but instead contain a predominance of TH2-like and T regulatory cells (Tregs). In addition, papillomas also are infiltrated with immature Langerhans cells (iLCs). In this study, we show that papilloma cells express IL-36γ protein, and that human keratinocytes transduced with HPV11 have reduced IL-36γ secretion. We now provide the first evidence that peripheral blood-derived iLCs respond to IL-36γ by expressing inflammatory cytokines and chemokines. When stimulated with IL-36γ, iLCs from patients with RRP had lower expression levels of the TH2-like chemokine CCL-20 as compared with controls. Patients’ iLCs also had decreased steady state levels of CCL-1, which is a proinflammatory chemokine. Moreover, CCL-1 levels in iLCs inversely correlated with the severity of RRP. The combined decrease of TH1- and a TH2-like chemokines by iLCs from patients could have consequences in the priming of IFN-γ expression by CD8+ T-cells. Taken together, our results suggest that, in RRP, there is a defect in the proinflammatory innate immune responses made by iLCs in response to IL-36γ. The consequence of this defect may lead to persistent HPV infection by failing to support an effective HPV-specific, TH1-like and/or Tc1-like adaptive response, thus resulting in the predominant TH2-like and/or Treg micromilieu present in papillomas.
INTRODUCTION
Recurrent respiratory papillomatosis (RRP) is a disease of the upper airway characterized by persistent human papillomavirus type 6/11 (HPV 6/11) infection that results in the chronic recurrence of benign tumors that can cause complete laryngeal occlusion. HPV infection of the airway is not uncommon, yet RRP is a rare disease. We have reported previously that patients with RRP have a biased TH2-like and/or Treg, antiinflammatory adaptive immune response to these HPVs (1–3). To determine the repertoire of immunologically relevant genes and pathways that are responsible for this skewed adaptive response, we previously performed a paired microarray analysis using papilloma tissues and clinically normal laryngeal tissues from the same patient to identify differential expression of genes and gene pathways in the papillomas (4). We observed that expression of the proinflammatory cytokine IL-36γ was markedly upregulated. In marked contrast to this observation, papilloma tissues fail to show signs of inflammation. To explore this paradox, and better understand why the immune system of RRP patients fails to control persistent laryngeal HPV infection, we have now addressed the innate immune responses made by Langerhans cells (LCs) in response to IL-36γ in patients with RRP.
IL-36γ is a member of the IL-1 superfamily that includes three closely related genes, IL-36α, β, and γ, formerly known as IL-1F6, F8, and F9 respectively. IL-36α has been detected in both neuronal and synovial tissue, whereas IL-36β and IL-36γ are expressed in both cutaneous and mucosal epithelial cells, including the respiratory tract. IL-36β and IL-36γ stimulate proliferation, maturation and/or cytokine expression by innate immune cells (such as keratinocytes and dendritic cells), and adaptive immune cells (neutrophils and T-cells) in both humans (5–8) and mice (9). A genetic defect in the regulation of IL-36γ stimulation results in the multisystemic inflammatory disease termed generalized pustular psoriasis (10), illustrating the importance of proper regulation of the proinflammatory signaling pathway mediated by IL-36 ligands in maintaining a normal epithelium (11).
The specific effect of IL-36γ stimulation of LCs, which are the resident professional antigen-presenting cells within the epithelium of the skin and mucosa, has not been described previously. Here we provide the first evidence that immature, monocyte-derived Langerhans cells (iLCs) respond to IL-36γ stimulation by increasing the expression of a specific subset of proinflammatory cytokines and chemokines. Monocyte-derived iLCs from patients with RRP show impaired upregulation of inflammatory mediators, especially CCL-20, following stimulation by IL-36γ, coupled with a lower basal expression of CCL-1 that correlates with disease severity. In addition, IL-36γ stimulation of both laryngeal and foreskin keratinocytes increases the expression of IL-36γ mRNA itself, similar to that observed for both IL-36β and IL-1β (12,13) indicating that a positive autocrine feedback loop exists in keratinocytes. However, we also find that HPVs can interfere with IL-36γ protein release from infected keratinocytes, which would further reduce the response by resident iLCs in papilloma tissues. Taken together, these results suggest that the normal innate immune function of LCs in response to IL-36γ is impaired in RRP.
MATERIALS AND METHODS
Sample Collection and Preparation
Respiratory papilloma tissue and clinically normal adjacent laryngeal biopsies were obtained during surgery to remove papillomas. Neonatal foreskin tissues were obtained following circumcision. Tissues were either snap frozen or cultured to generate primary cells as described previously (14). Airway keratinocytes were used at passage two or three. Foreskin keratinocytes were incubated in dispase at 4°C overnight, trypsinized and plated as single cells in keratinocyte culture media (3:1 DMEM:Ham’s F-12, 10% FetalClone II [Fisher Scientific, Pittsburgh, PA, USA {Thermo Fisher Scientific Inc., Waltham, MA, USA}], 0.4 μg/mL hydrocortisone, 0.1 nmol/L cholera toxin, 5 μg/mL transferrin, 2 nmol/L 3,3′-5-triiodo-l-thyronine, 5 ng/mL epidermal growth factor [EGF], 5 μg/mL insulin, 10 units/mL penicillin and streptomycin) in the presence of mitomycin C-treated J2-3T3 feeder layers, and used no later than passage five. Whole blood from patients was collected in heparin-coated tubes at the time of surgery. The disease severity score for each patient studied is a composite value of the rate of regrowth of papilloma mass divided by the time in days since the previous surgery (15,16). Any score over 0.06 is classified as severe disease. Control blood was collected from volunteers with no history of RRP. All patients and controls signed consent forms, and all studies were approved by the Institutional Review Board, Feinstein Institute for Medical Research, North Shore-LIJ Health System, Great Neck, NY, USA.
Western Blot
Pulverized frozen tissues and cultured cells were extracted in Tris/NP-40/EDTA (TNE) buffer (0.1 mol/L Tris pH 8.0, 1% NP-40, 5 mmol/L EDTA) supplemented with a protease inhibitor cocktail (Roche Diagnostics, Indianapolis, IN, USA) for 30 min with final protein concentrations determined as described previously (17). Proteins (20 μg per lane) were separated on 8%–16% SDS-PAGE gels, (Bio-Rad, Hercules, CA, USA) electro blotted onto Immobilon-FL membranes, (Millipore, Billerica, MA, USA) and imaged with the LI-COR Odyssey (LI-COR Biotechnology, Lincoln, NE, USA).
Lentiviral Transduction and Analysis of IL-36γ Release
The upstream regulatory region (URR) and early genes of HPV (nt7178-5963) were ligated into the lentiviral vector pSMPUW (Cell Biolabs, San Diego, CA, USA) to create a lentiviral vector in which all early genes of HPV11 are expressed under the control of their endogenous HPV promoters. Early passage human foreskin keratinocyte cultures (HFKs) derived from multiple specimens and pooled, were infected with 90 copies per cell of the HPV11-lentiviral vector, or with the empty vector as a control, and cultured for one week. Total RNA was isolated from transduced HFKs with the Roche High Pure RNA Isolation Kit (Roche Diagnostics); Q-PCR was performed using primers specific for HPV11E7 (forward primer nt133–nt157, reverse primer nt195–nt212, probe nt161–nt184); and the signal was compared with that for glyceraldehyde 3-phosphate dehydrogenase (GAPDH) to confirm transduction and viral expression. Transduced keratinocytes containing either HPV11 or empty vector (control), were treated with poly (I:C) (HMW) (InvivoGen, San Diego, CA, USA) for 96 h, after which supernatants were analyzed by IL-36γ enzyme-linked immunosorbent assay (ELISA) (Human IL-1F9 ELISA kit, Aviscera Bioscience, Santa Clara, CA, USA). The percentage of cell death was determined by trypan-blue staining of treated cells, and by measurement of lactic dehydrogenase in the supernatant as per the manufacturer (Cytotoxicity Detection Kit (LDH), Roche Diagnostics).
Isolation, Culture, and Flow Cytometry Characterization of Monocyte-Derived Langerhans Cells
Mononuclear cells were isolated from peripheral blood using Ficoll-Paque density gradient centrifugation (GE Healthcare, Fairfield, CT, USA). Monocytes were negatively selected using a commercially available magnetic isolation kit (Monocyte Isolation Kit II, Miltenyi Biotec, Bergisch Gladbach, Germany), which consistently yielded greater than 90% purity. Isolated monocytes were suspended at 1 × 106/mL in RPMI 1640 media (Gibco/Life Technologies [Thermo Fisher Scientific Inc.]) containing 10% fetal bovine serum (FBS) (Atlanta Biologicals, Flowery Branch, GA, USA), 100 ng/mL granulocyte-macrophage colony-stimulating factor (GMCSF), 20 ng/mL IL-4 and 10 ng/mL TGFβ1 (R&D Systems, Minneapolis, MN, USA) (18,19). Monocyte cell suspensions were cultured in 6-well poly-styrene trays (Falcon, Fisher Scientific [Thermo Fisher Scientific Inc.]) and supplemented with fresh medium without IL-4 on d 1 and d 4. On d 6 or 7, cells were harvested, stained for viability (Live/Dead Fixable Aqua Dead Cell Stain Kit [Invitrogen/Life Technologies {Thermo Fisher Scientific Inc.}]) and then surface stained with fluorochrome-labeled antibodies CD324 APC (BioLegend, San Diego, CA, USA), CD207 PE and CD1a PE (Beckman Coulter, Brea, CA, USA), CD83 APC and CD14 Pacific Blue, (BD Biosciences, San Jose, CA, USA). After washings, cells were resuspended in 1% formaldehyde and analyzed on a FACSCanto II. Forward scatter area versus forward scatter height gating was used to remove cell aggregates. Viable cells with large forward and side scatter properties were analyzed using FlowJo version 7.2.5 (TreeStar, Ashland, OR, USA). Positive fluorescence staining was established using isotype and fluorescence minus one control.
IL-36γ Stimulation and Analysis of Monocyte-Derived Immature Langerhans Cells
The iLCs (2.5 × 106; d 6 or 7) were resuspended in media without cytokines and stimulated for 4 h at 37°C with 10 ng/mL of recombinant active IL-36γ (aa 18–169) (R&D Systems), or with the recombinant inactive full length IL-36γ (aa 1–169) as a control. Cells were collected by centrifugation, and total RNA isolated, following DNase1 treatment, as per manufacturers instructions (Qiagen, Valencia, CA, USA). RNA integrity (RIN > 7.0) was measured using a Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). For microarray analysis, total RNA (300 ng) was reverse transcribed and biotin-labeled prior to hybridization to Human HT-12V4 expression arrays (Illumina, San Diego, CA, USA) as described previously (4). Data were processed using BeadStudio (Illumina), and tab-delimited text files were uploaded to GeneSifter software (Geospiza Inc., Seattle, WA, USA) and comparisons of various experimental conditions analyzed.
For quantitative reverse transcription polymerase chain reaction (RT-PCR) analysis, gene-specific intron-spanning primers were purchased from Applied Biosystems/Life Technologies (Thermo Fisher Scientific Inc.), or designed using a free bioinformatics program (Table 1) available on the Fisher website (Fisher Scientific, [Thermo Fisher Scientific Inc.]) and RT-Q-PCR performed on an Applied Biosystems 7900 HT (Applied Biosystems/Life Technologies) using the appropriate universal probes (Roche) together with the insert iScript One-Step RT-PCR kit (Bio-Rad). Ten nanograms of total RNA was reverse transcribed at 50°C for 10 min, followed by a 5 min hot start at 95°C followed by 40 cycles of 2-step PCR at 95°C for 15 sec and 60°C for 1 min. Individual RNA samples were run in duplicate and the average Ct values for the gene of interest compared with the average Ct values for a housekeeping gene (GAPDH) to calculate δδ Ct, where fold change = 2[δδCt].
Table 1.
RT-Q-PCR primers and probes.
| Forward primer | Reverse primer | Probe | Gene name |
|---|---|---|---|
| TACCTGTCCTGCGTGTTGAA | TCTAGGGTTTTTAATGGGTTTCT | AGCTGGAG | IL-1β NM_000576.2 |
| AAGTTTTACATGCCCAAGAAGG | AAGTGAAAGTTTTTGCTTTGAGCTA | CTGGAGGA | IL-2 ENST00000226730.3 |
| TGTTCATTTATGGAAGGGATGA | TTCTGCTTTTCTTTACGTTTTCATT | TGGGGAAG | IL-1R1 |
| CCCTGTGATCCATTCACCTAA | TGCGAGAATCCATCAGAAAAC | TCCTCTCC | IL-1RacP |
| GCTGGAGTGTCCACAGCATA | GCGATAAGCCCTCCTATCAA | CAGCTCCC | IL-1R6 |
| TGAGAGGCACTCCAGGAGAC | AGGGTCCACACTTGCTGATT | TGGTGGAG | IL-36γ |
| GATGAGTACAAAAGTCCTGATCC | CTGCAGCCACTGGTTCTGT | CAGCAGGC | IL-6 |
| TTGCTGCTAGCTGGGATGT | CTGGAGAAGGGTACCTGCAT | TGCTGTCC | CCL-1 |
| GTGGCTTTTCTGGAATGGAA | CAACCCCAGCAAGGTTCTT | UPL-H117 | CCL-20 |
RESULTS
IL-36γ Protein is Expressed in Respiratory Papillomas
We demonstrated previously that IL-36γ mRNA is elevated in respiratory papillomas and that expression correlated with disease severity (4). We now have confirmed that IL-36γ protein also was elevated in papilloma tissues in comparison to clinically normal (CN) autologous laryngeal epithelium (Figure 1). Shown are Western blots of two representative pairs of matched tissue sets. All papilloma tissues examined (n = 6) expressed abundant IL-36γ protein, which was not detectable at any appreciable levels in any of the matched CN tissues.
Figure 1.
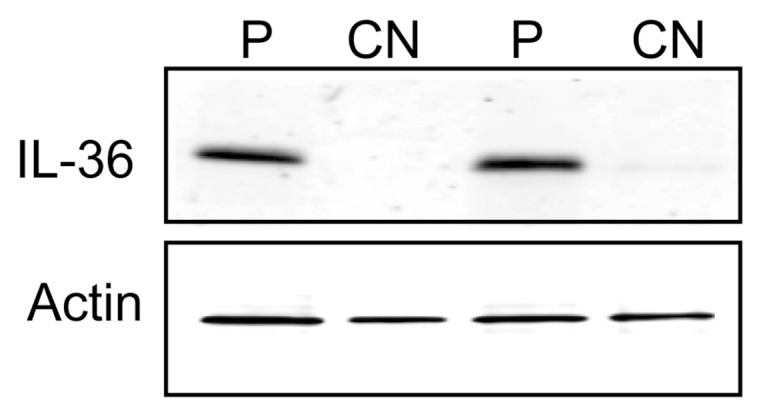
IL-36γ protein is overexpressed in papilloma tissues. Western blot showing IL-36γ expression levels in papilloma tissue (P) and autologous clinically normal laryngeal tissue (CN) from RRP patients. β-actin (lower panel) shown as a loading control. Representative of six matched pairs of tissues.
HPV-11 Suppresses IL-36γ Release from Keratinocytes
Since respiratory papillomas express increased IL-36γ mRNA and protein, we then asked whether HPV infection prevents the release of this proinflammatory cytokine. Others have shown that high concentrations (2.5–25 μg/mL) of the synthetic TLR-3 ligand poly (I:C), which is an analog of double-stranded RNA, can induce release of IL-36γ from human foreskin keratinocytes (HFKs) by inducing cell death (20). We have found that HFKs can be induced to secrete IL-36γ protein at much lower concentrations of poly (I:C) (0.25–0.75 μg/mL) without any detectable cell death. The amount of IL-36γ secreted in response to low dose poly (I:C) was markedly decreased in cells transduced by HPV-11, but was not completely blocked (Figure 2A). This difference in secretion was not caused by differences in protein expression, since intracellular levels of IL-36γ were comparable (Figure 2B).
Figure 2.

HPV-11 suppresses release of IL-36γ from keratinocytes. Human foreskin keratinocytes that had been transduced with either a lentivirus construct expressing all early HPV-11 genes or the empty vector were treated with poly(I:C) or solvent for 96 h. IL-36γ release was measured by ELISA (A), with results shown as the mean ± SD of three assays, *p < 0.05. Comparable levels of intracellular IL-36γ were confirmed by Western blot (B).
Respiratory Papillomas Contain Abundant iLCs
Since papillomas contain IL-36γ protein, and at least some IL-36γ can be released from keratinocytes infected with HPV-11, we asked whether the lack of inflammation in papillomas reflected the absence of LCs in the epithelium. We used immunohistochemistry to determine whether LCs were present in papillomas and found that they were easily identified and abundant in papilloma tissues (Figure 3A). These cells had the morphology of dendritic cells when visualized at high magnification (Figure 3B). They expressed both CD1a (Figure 3C) and langerin (CD207) (Figure 3D) but not the maturation marker CD83 (Figure 3E). Thus, the numerous LCs in papillomas are immature (iLCs). We were able to isolate iLCs from small papilloma biopsies (Figure 3G), and we determined that they were approximately half as frequent as the lymphocytes that we have previously characterized in respiratory papillomas (21,22). The absence of both CD14 and CD1a expression (Figure 3H) confirmed that they were indeed resident iLCs, and not simply monocytes from the blood vessels that permeate respiratory papillomas.
Figure 3.

Papillomas contain abundant iLCs. iLCs were CD1a+ (A,B) and langerin+ (C,D), but CD83− (E). A positive control (lymphoma) was CD83+ (F). (A–B) Paraffin sections, DAB stain. (C–D) frozen sections, AEC stain. (E–F) Paraffin sections, immunofluorescence with DAPI counterstain. (G, H) Flow cytometry of LCs isolated from papilloma biopsies. LCs (G, lower right) were approximately half as frequent as T-cells (G, upper left). LCs were CD14 negative (H, lower right), confirming that they were not monocytes and that there was no significant contamination by blood.
IL-36γ Induces iLCs to Express Proinflammatory Cytokines and Chemokines, but iLCs from Patients with RRP Show Reduced Responses Compared with Controls
Because iLCs were present in papillomas, we questioned whether they were able to respond to IL-36γ. However, nothing was known about the response of normal iLCs to this cytokine. We therefore first characterized the response of control iLCs to recombinant IL-36γ, using monocyte-derived iLCs because they are abundant and the number of resident iLCs that can be derived from tissue biopsies is limited. These blood-derived iLCs expressed CD1a and E-cadherin on their cell surfaces, and approximately half of the cells also expressed langerin (Figure 4A). The CD1a+ cells were CD83 negative (Figure 4B), consistent with our finding in papillomas (see Figure 3E) and reports that characterized resident LCs in the airway mucosa as immature (23). Furthermore, they were functional, since they could be induced to mature when stimulated with either IL-36γ or lipopolysaccharide (LPS) (Figures 4C–E).
Figure 4.

Characterization of LCs generated from peripheral blood monocytes. iLCs were generated from isolated peripheral blood monocytes by incubating blood monocytes with GMCSF, TGF-β1, and IL-4. After 6 d, they were analyzed by flow cytometry. The cells were E-cadherin+ and approximately 50% were langerin+ (A), but immature (CD83−) (B). Day 11 iLCs (C) could be induced to mature, as measured by surface expression of CD83 and HLADR, when stimulated with either IL-36γ (D) or LPS (E) for 48 h.
We initially performed dose response and time course experiments to determine the optimal conditions for IL-36γ stimulation. Monocytes on d 2, prior to differentiation into iLCs, did not respond to IL-36γ but responded to IL-1β and to lipopolysaccharide (LPS) as expected (data not shown). At d 6, after differentiation into iLCs, the cells responded to IL-36γ with statistically significant increases in mRNA expression of both inflammatory cytokines and chemokines (Figure 5). The response was detectable at 2–5 ng/mL and maximal at 10–20 ng/mL for all individuals tested. Comparison of 4, 6 and 8 h exposure to IL-36γ revealed that maximal responses were observed at 4 h and decreased thereafter.
Figure 5.

Response of iLCs to IL-36γ. Cells were stimulated with either recombinant inactive, full-length (aa 1–169) IL-36γ (control) or the active, shorter form (aa 18–169) of IL-36γ for 4 h. Induction of mRNA for specific proinflammatory cytokines (A) and chemokines (B) was measured by q-PCR. Results are the mean ± SD of at least six separate cultures, normalized to control cultures treated with solvent.
IL-36γ signals through a single heterodimer receptor comprised of IL-1R6 and the receptor accessory protein IL-1RacP (IL-1R3). This receptor accessory protein is common to both the IL-36 receptor and the IL-1 receptor (IL-1R1) (24). We therefore asked if the expression of the unique receptor subunit for the IL-36 receptor changed from d 2 to d 6 of culture. While IL-1R6 mRNA levels were four- to eight-fold lower than IL-1R1 and IL-1RacP, they were still expressed at relatively high levels in the d 2 cells and the mRNA levels for each of the unique receptor subunits (IL-1R1, IL-1R6), as well as the common accessory protein IL-1RacP, were unchanged between d 2 and d 6 (Figure 6). This result suggested that the change in ability to respond to IL-36γ is not determined by increased expression of either subunit.
Figure 6.
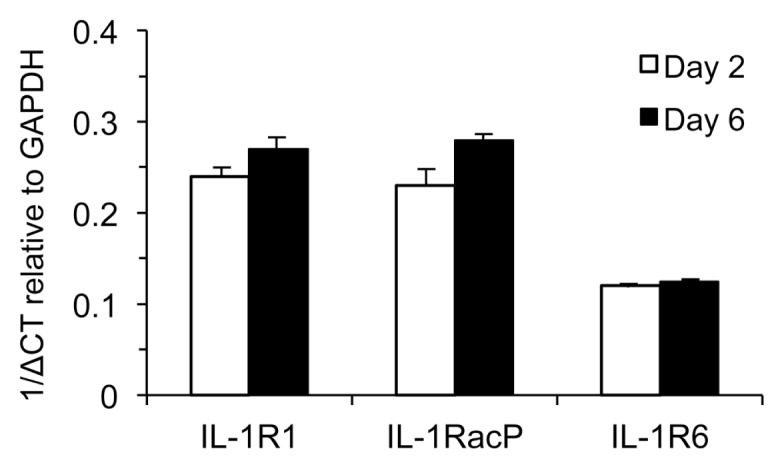
Differentiation of LCs from monocytes does not alter IL-1 receptor and IL-36 receptor expression. Blood-derived monocytes (d 2) and iLCs derived from monoctyes (d 6) were assayed for mRNA expression of IL-1R1, IL-1R3, and IL-1R6 and compared with GAPDH mRNA expression (δCT).
Six cultures of iLCs (three from patients with RRP and three controls) were studied by gene microarray to identify the broad repertoire of genes that were upregulated by IL-36γ stimulation. Among the top 20 genes upregulated at least three-fold above baseline (Table 2), seven encoded either cytokines or chemokines. These genes are shown in descending order of abundance: CCL-1, IL-1β, LTα, CCL-20, LTβ, CXCL-1, and TNF-α. There was no qualitative difference in the transcriptional profiles of iLCs induced by IL-36γ stimulation between the patients with RRP and the controls.
Table 2.
Top 20 genes upregulated by IL36γ in iLCs.
| Rank order | p Value | Identifier | Gene name/protein name |
|---|---|---|---|
| 1 | 0.00258 | NM_002981 | Chemokine (C-C motif) ligand 1/CCL1 |
| 2 | 7.85e-07 | XM_938742 | PREDICTED: sphingosine-1-phosphate phosphotase 2 (SGPP2), mRNA. |
| 3 | 0.00065 | NM_000930 | Plasminogen activator, tissue |
| 4 | 2.15e-06 | NM_007115 | Tumor necrosis factor, α-induced protein 6 |
| 5 | 1.36e-05 | NM_000576 | Interleukin 1, beta/IL-1β |
| 6 | 3.19e-07 | NM_003327 | Tumor necrosis factor receptor superfamily, member 4 |
| 7 | 5.76e-06 | NM_153259 | Mucolipin 2 |
| 8 | 1.54e-05 | NM_001024465 | Superoxide dismutase 2, mitochondrial |
| 9 | 0.00166 | NM_000595 | Lymphotoxin α (TNF superfamily, member 1) |
| 10 | 0.00012 | NM_014398 | Lysosomal-associated membrane protein 3 |
| 11 | 0.00031 | NM_004591 | Chemokine (C-C motif) ligand 20/CCL20 |
| 12 | 2.34e-05 | NM_001024466 | Superoxide dismutase 2, mitochondrial |
| 13 | 3.39e-05 | NM_003037 | Signaling lymphocytic activation molecule family member 1 |
| 14 | 0.00035 | NM_002341 | Lymphotoxin β (TNF superfamily, member 3) |
| 15 | 4.19e-05 | NM_003088 | Fascin homolog 1, actin-bundling protein (Strongylocentrotus purpuratus) |
| 16 | 0.00113 | NM_172374 | Nucleoporin 62kDa |
| 17 | 0.00027 | NM_001511 | Chemokine (C-X-C motif) ligand 1 (melanoma growth stimulating activity, alpha)/CXCL1 |
| 18 | 0.00085 | NM_001165 | Baculoviral IAP repeat-containing 3 |
| 19 | 0.00026 | NM_178452 | Leucine rich repeat containing 50 |
| 20 | 0.00041 | NM_000594 | Tumor necrosis factor (TNF superfamily, member 2)/TNF-α |
We then used qPCR to quantify the enhanced expression of select cytokines and chemokines using a larger cohort from controls and patients. As seen in Figure 7, induction of several proinflammatory mediators was different between the two groups. iLCs from controls clearly responded to IL-36γ by markedly upregulating the expression of the proinflammatory mediators CCL-20, and IL-1β and TNF-α (see Figure 7), and IL-6 which can be either pro- or antiinflammatory (25–28). The response levels varied somewhat between individuals, but they were increased in all samples. The responses to IL-36γ were very specific, given that not all cytokines/chemokines were induced. Expression of IL-2, IL-4, IL-8, IL-10, IL-12a, and IL-23 were all unchanged (data not shown), as predicted by the microarray analysis.
Figure 7.
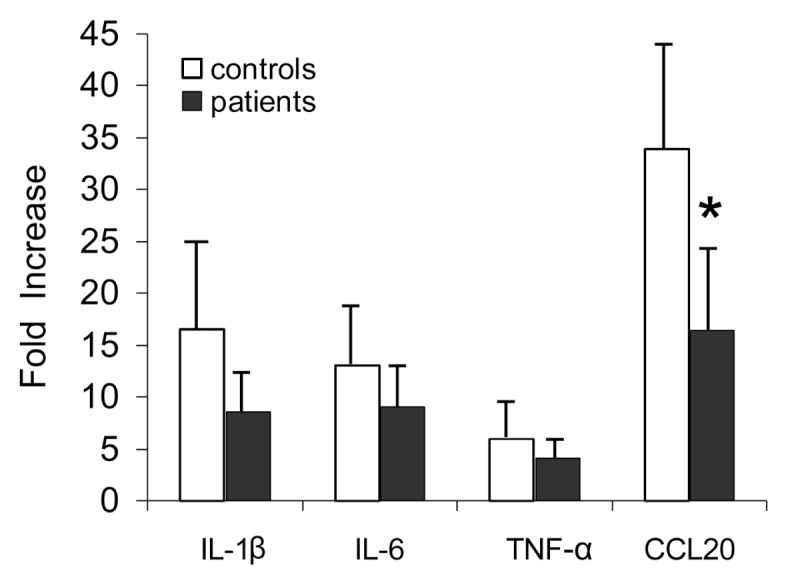
iLCs from patients are less responsive to IL-36γ. Cells from both healthy controls and RRP patients were stimulated with active IL-36γ for 4 h, and increased expression of proinflammatory cytokines and chemokines measured by q-PCR. A consistent pattern of blunted expression by the patient-derived LCs was evident, with the difference in CCL20 significant and the difference in IL-1β just missing significance. Results are the mean ± SD of at least six separate iLC cultures from controls and seven isolates from patients. *p < 0.05.
The quantitative responses by iLCs from patients with RRP were reduced compared with iLCs from controls, with reduced expression of IL-1β, IL-6, TNF-α and CCL20 (see Figure 7). Of note, the reduction in CCL-20 was significantly different (p < 0.05), reduction of IL-1β expression approached significance (p = 0.06), and a trend to reduced expression was clearly seen for IL-6 and TNF-α. It is possible that analysis of a larger cohort of patients would show a significant reduction in the levels for all four immune mediators.
Constitutive CCL-1 Expression by iLCs from Patients’ with RRP Correlates with Disease Severity
CCL-1 was the most elevated immune mediator following IL-36γ stimulation, increasing more than 100-fold (p < 0.05) in controls, but mean levels of induction did not differ between RRP patients and controls. However, there was considerable variability in the baseline levels of CCL-1 expressed by the patients’ cells. We therefore asked whether there was a correlation between CCL-1 mRNA levels in unstimulated iLCs from patients and the severity of their disease (Figure 8). The highest constitutive expression was detected in patients with mild disease, at levels comparable to the controls without RRP. In contrast, there was significantly lower baseline expression in patients with moderate or severe disease, with the most severe patients having the lowest baseline levels. Thus, low constitutive CCL-1 mRNA expression by iLCs from patients predicted disease severity.
Figure 8.
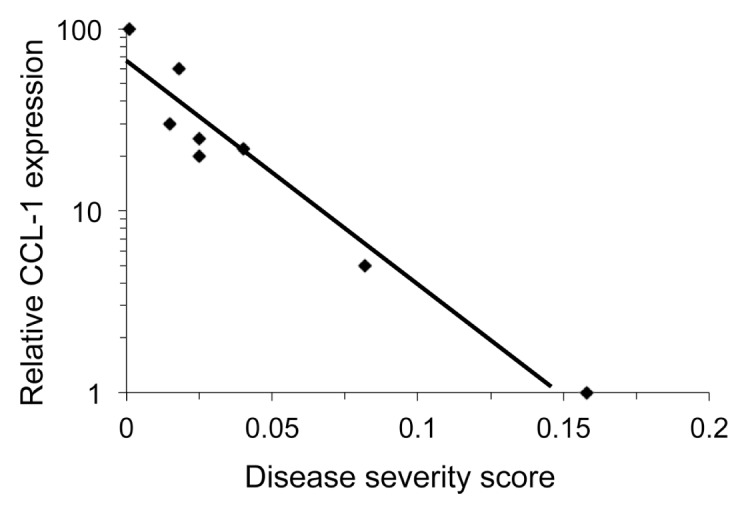
Expression of CCL1 in iLCs correlates with severity of RRP. The level of constitutive CCL1 expression in iLCs from patients with RRP was determined by q-PCR. Expression levels in iLCs from eight controls were defined as 100%; and individual samples clustered tightly around the 100% value (not shown); there was nearly a two-log reduction in expression as severity increased in the patients, p < 0.0001. Disease severity score is a composite value of the rate of regrowth of papilloma mass divided by the time in days since the previous surgery. A score over 0.06 is classified as severe disease.
DISCUSSION
Infection of laryngeal keratinocytes with HPV6/11 induces the production of a number of inflammatory cytokines, including IL-36γ, which should facilitate a proinflammatory adaptive immune response that ultimately results in the clearance or control of this infection. The chronic nature of RRP suggests that, especially for those with severe disease, an effective adaptive immune response to these viruses is not achieved. We previously showed that the adaptive immune responses made by RRP patients are polarized toward immune tolerance of HPVs, with a strong TH2-like and/or Treg bias (29–32). However, the innate responses that orchestrate the polarization of the adaptive immune responses to these viruses had not yet been characterized, and were the focus of this study.
Airway epithelial cells constitute the primary antimicrobial defense against aerosolized pathogens by providing a physical barrier that is enhanced by the production of mucus and the expression of proinflammatory cytokines, chemoattractants, and defensins which we have previously shown to be upregulated in RRP (4). We also reported that a single member of the IL-1 superfamily, the proinflammatory cytokine IL-36γ, was markedly and consistently upregulated in respiratory papillomas, yet there was no evidence of inflammation in these tissues (4). Select members of the IL-36 family of cytokines are expressed by both mouse and human bronchial epithelial cells (33,34), and by laryngeal keratinocytes (4). Unopposed signaling from the IL-36 receptor, secondary to the functional loss of the IL-36 receptor antagonist (IL-36Ra) has been shown to lead to the severe dermal inflammatory disease generalized pustular psoriasis (10), illustrating the strong proinflammatory effects of unregulated IL-36 signaling in the epidermis.
Although it has been shown previously that myeloid dendritic cells can respond to IL-36γ (35), those authors used the full length, inactive form of the protein, which precluded an appreciation of how this cytokine triggers a proinflammatory response (9). They also suggested that LC might respond to IL-36γ, but did not demonstrate this. LCS are a migratory subset of dendritic cells that specialize in uptake, transport, processing and presentation of antigens to T-cells (36–39), and are essential in generating Tregs that induce tolerance in vitro and in vivo (40). LCs mature during antigen encounter and migrate to lymph nodes (41) where they present peptides to T-cells. The balance of pro-and antiinflammatory cytokines made by keratinocytes (42–44), with contributions from LCs themselves, influence LC migration, maturation, and function.
There is considerable plasticity in LCs that skews T-helper cell responses, dependent upon the antigen and the cytokine milieu present during this encounter (45). LCs can induce TH1 responses but also can be converted to support TH2 responses when IL-10 is present (41,46–52). Thus, LCs can function as either pro- or antiinflammatory (53–57) mediators of adaptive immunity. Others have reported that IL-36β, which binds to the same receptor (IL-1R6) as IL-36γ, can increase expression of IL-12p70 and IL-18 by monocyte-derived DCs, and thereby induce IFN-γ expression by human TH1-like T-cells(6). They also reported that IL-36γ can mature these DCs, manifested by increasing their expression of HLA-DR and CD83 and reducing CD1a (6). We previously showed that both IL-4 and IL-10 (1) are expressed in papillomas with little if any IFN-γ, IL-12, or IL-18 (58,59).
CONCLUSION
In this study, we have shown for the first time that iLCs generated from peripheral blood monocytes respond to IL-36γ by upregulation of additional specific proinflammatory cytokine and chemokine mRNAs, with high levels of IL-1β and CCL1 mRNA, and moderate levels of TNF-α, demonstrating that they clearly could support adaptive TH1-like T-cell responses (see Figures 5,7). Their expression of IL-6 and CCL20 showed that they also could support TH2-like responses, which has recently been suggested to be the principle role of LCs in the epidermis (60,61). Acquisition of the ability to respond to IL-36γ required differentiation to iLCs, since monocytes did not respond. The mechanism of this difference has not been elucidated, although we have ruled out a difference in IL-36γ receptor mRNA expression (see Figure 6). We have been unable to measure receptor protein expression or surface localization using commercially available antibodies. Future studies will determine whether there are differences in activation of signal transduction pathway intermediates downstream of the receptor.
The cytokine/chemokine responses of iLCs from patients stimulated with IL-36γ were qualitatively similar to controls, but the responses were generally less robust (see Figure 7). The lower baseline expression of the proinflammatory chemokine CCL-1 in iLCs from patients with the most severe disease could provide significant insight into the mechanism underlying our earlier studies of the adaptive response in RRP which showed that the more extreme the TH2-like/Treg cytokine polarization of adaptive immune responses made by patients with RRP, the more severe the disease (4,29,31,32,62). CCL-1 links adaptive and innate immune function by recruiting LC-like dendritic cells and cytotoxic CD8+ T-cells (63). CCL-1 is required along with CCL-17 for the IL-12-induced coupling of dendritic cells and CD8+ T-cells to generate IFN-γ (64). IL-12 expression and the presence of functional cytotoxic T-cells are both reduced in RRP, and IFN-γ expression is extremely low, while CCL-17 is not reduced (1,31), suggesting that deficient IL-12, linked with disproportional CCL-1/CCL17 expression, would reduced dendritic cell-naïve CD8+ T-cell priming leading to reduced IFN-γ production which is characteristic of adaptive response in RRP (1,32).
We were surprised by the reduced expression of CCL20 by patients’ iLCs in response to IL-36γ, since this chemokine is important in recruiting Tregs and LCs into tissues (23,65,66) and both are abundant in papillomas (22). However, newly recruited LCs are required for CD8+ T-cell priming to microbial proteins and adjuvants such as poly(I:C) and flagellin, and for their subsequent expression of IFN-γ(23). Thus, reduced expression of CCL20 by iLCs from RRP patients would alter the balance between resident and newly recruited LCs. This, in conjunction with low baseline levels of CCL-1, may explain the failure of CD8+CD28− T-cells that are enriched in respiratory papillomas (29) to support an inflammatory response.
In summary, we demonstrate, for the first time, that iLCs respond to IL-36 agonists by increasing their expression of both pro- and antiinflammatory interleukins and chemokines, but that patients with RRP have a lower level of response. Our findings also suggest that suppressed IL-36γ release by HPV-infected keratinocytes, coupled with the systemic alterations in iLCs, results in suppressed innate signaling that contribute to the ineffective adaptive immune responses to HPV by patients with RRP. Further characterization of the response of resident iLC in respiratory papillomas to IL-36γ and other cytokines, together with illuminating the mechanism(s) by which HPVs alter IL-36γ release, should provide critical insight into how these viruses evade the normal mucosal immune responses and why patients with RRP are unable to control their disease.
ACKNOWLEDGMENTS
The authors would like to acknowledge the helpful discussion and encouragement of Isaac Rodriguez-Chavez. Research reported in this publication was supported by the National Institute of Dental and Craniofacial Research of the National Institutes of Health under Award Number R01DE017227, and the National Institute of Allergy and Infectious Diseases of the National Institutes of Health under Award Number R21AI105987, and by the Feinstein Institute for Medical Research, North Shore-LIJ Health System. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Footnotes
Online address: http://www.molmed.org
DISCLOSURE
The authors declare they have no competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
REFERENCES
- 1.DeVoti JA, et al. Failure of gamma interferon but not interleukin-10 expression in response to human papillomavirus type 11 E6 protein in respiratory papillomatosis. Clin. Diagn. Lab. Immunol. 2004;11:538–47. doi: 10.1128/CDLI.11.3.538-547.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bonagura VR, et al. KIR3DS1, KIR2DS1, and KIR2DS5 protect against the development of severe recurrent respiratory papillomatosis (RRP) in HPV-6/11-Infected patients. J. Allergy Clin. Immunol. 2009;123:S165. [Google Scholar]
- 3.Bonagura VR, et al. Recurrent respiratory papillomatosis: a complex defect in immune responsiveness to human papillomavirus-6 and -11. APMIS. 2010;118:455–70. doi: 10.1111/j.1600-0463.2010.02617.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.DeVoti JA, et al. Immune dysregulation and tumor-associated gene changes in recurrent respiratory papillomatosis: A paired microarray analysis. Mol Med. 2008;14:608–17. doi: 10.2119/2008-00060.DeVoti. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vigne S, et al. IL-36 signaling amplifies Th1 responses by enhancing proliferation and Th1 polarization of naive CD4+ T cells. Blood. 2012;120:3478–87. doi: 10.1182/blood-2012-06-439026. [DOI] [PubMed] [Google Scholar]
- 6.Mutamba S, Allison A, Mahida Y, Barrow P, Foster N. Expression of IL-1Rrp2 by human myelomonocytic cells is unique to DCs and facilitates DC maturation by IL-1F8 and IL-1F9. Eur. J. Immunol. 2011;42:607–17. doi: 10.1002/eji.201142035. [DOI] [PubMed] [Google Scholar]
- 7.Blumberg H, et al. Opposing activities of two novel members of the IL-1 ligand family regulate skin inflammation. J. Exp. Med. 2007;204:2603–14. doi: 10.1084/jem.20070157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Johnston A, et al. IL-1F5, -F6, -F8, and -F9: a novel IL-1 family signaling system that is active in psoriasis and promotes keratinocyte antimicrobial peptide expression. J Immunol. 2011;186:2613–22. doi: 10.4049/jimmunol.1003162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Vigne S, et al. IL-36R ligands are potent regulators of dendritic and T cells. Blood. 2011;118:5813–23. doi: 10.1182/blood-2011-05-356873. [DOI] [PubMed] [Google Scholar]
- 10.Marrakchi S, et al. Interleukin-36-receptor antagonist deficiency and generalized pustular psoriasis. N. Engl. J. Med. 2011;365:620–8. doi: 10.1056/NEJMoa1013068. [DOI] [PubMed] [Google Scholar]
- 11.Towne JE, Sims JE. IL-36 in psoriasis. Curr. Opin. Pharmacol. 2012;12:486–90. doi: 10.1016/j.coph.2012.02.009. [DOI] [PubMed] [Google Scholar]
- 12.Dinarello CA. Overview of the interleukin-1 family of ligands and receptors. Semin Immunol. 2013;25:389–93. doi: 10.1016/j.smim.2013.10.001. [DOI] [PubMed] [Google Scholar]
- 13.Carrier Y, et al. Inter-regulation of Th17 cytokines and the IL-36 cytokines in vitro and in vivo: implications in psoriasis pathogenesis. J. Invest. Dermatol. 2011;131:2428–37. doi: 10.1038/jid.2011.234. [DOI] [PubMed] [Google Scholar]
- 14.Steinberg BM, Abramson AL, Meade RP. Culture of human laryngeal papilloma cells in vitro. Otolaryngol Head Neck Surg. 1982;90:728–35. doi: 10.1177/019459988209000610. [DOI] [PubMed] [Google Scholar]
- 15.Kashima HB, Leventhal B, Mounts P Papilloma Study Group. In: Howley PM, Broker TR, editors. Scoring system to assess severity and course in recurrent respiratory papillomatosis; Papillomavirus: molecular and clinical aspects: proceedings of the Burroughs-Wellcome-UCLA Symposium held in Steamboat Springs; Colorado. April 8–14, 1985; New York: Alan R. Liss; 1985. [Google Scholar]
- 16.Abramson AL, et al. Clinical effects of photodynamic therapy on recurrent laryngeal papillomas. Arch. Otolaryngol. Head Neck Surg. 1992;118:25–29. doi: 10.1001/archotol.1992.01880010029011. [DOI] [PubMed] [Google Scholar]
- 17.Wu R, Coniglio SJ, Chan A, Symons MH, Steinberg BM. Up-regulation of Rac1 by epidermal growth factor mediates COX-2 expression in recurrent respiratory papillomas. Mol Med. 2007;13:143–50. doi: 10.2119/2007-00005.Wu. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kis-Toth K, et al. Monocyte-derived dendritic cell subpopulations use different types of matrix metalloproteinases inhibited by GM6001. Immunobiology. 2013;218:1361–9. doi: 10.1016/j.imbio.2013.06.012. [DOI] [PubMed] [Google Scholar]
- 19.Chung DJ, et al. Langerhans-type and monocyte-derived human dendritic cells have different susceptibilities to mRNA electroporation with distinct effects on maturation and activation: implications for immunogenicity in dendritic cell-based immunotherapy. J. Transl. Med. 2013;11:166. doi: 10.1186/1479-5876-11-166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lian LH, Milora KA, Manupipatpong KK, Jensen LE. The double-stranded RNA analogue polyinosinic-polycytidylic acid induces keratinocyte pyroptosis and release of IL-36gamma. J. Invest. Dermatol. 2012;132:1346–53. doi: 10.1038/jid.2011.482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hatam LJ, et al. CD4(+)Foxp3(+)CD127(+low) T-Regulatory cells are increased in HPV infected papillomas in patients with recurrent respiratory papillomatosis (RRP) J. Allergy Clin. Immunol. 2008;121:S211. [Google Scholar]
- 22.Hatam LJ, et al. Immune suppression in pre-malignant respiratory papillomas: enriched functional CD4+Foxp3+ regulatory T cells and PD-1/PD-L1/L2 expression. Clin Cancer Res. 2012;18:1925–35. doi: 10.1158/1078-0432.CCR-11-2941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Geissmann F, et al. Accumulation of immature Langerhans cells in human lymph nodes draining chronically inflamed skin. J. Exp. Med. 2002;196:417–30. doi: 10.1084/jem.20020018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Towne JE, Garka KE, Renshaw BR, Virca GD, Sims JE. Interleukin (IL)-1F6, IL-1F8, and IL-1F9 signal through IL-1Rrp2 and IL-1RAcP to activate the pathway leading to NF-kappaB and MAPKs. J. Biol. Chem. 2004;279:13677–88. doi: 10.1074/jbc.M400117200. [DOI] [PubMed] [Google Scholar]
- 25.Zhang C, Zhang X, Chen XH. Inhibition of the interleukin-6 signaling pathway: a strategy to induce immune tolerance. Clin. Rev. Allergy Immunol. 2014 2014 Mar 20; doi: 10.1007/s12016-014-8413-3. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 26.Weng Z, Patel AB, Vasiadi M, Therianou A, Theoharides TC. Luteolin inhibits human keratinocyte activation and decreases NF-κB induction that is increased in psoriatic skin. PLoS One. 2014;9:e90739. doi: 10.1371/journal.pone.0090739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Junk DJ, Bryson BL, Jackson MW. HiJAK’d signaling; the STAT3 paradox in senescence and cancer progression. Cancers (Basel) 2014;6:741–55. doi: 10.3390/cancers6020741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Guggino G, et al. Targeting IL-6 signalling in early rheumatoid arthritis is followed by Th1 and Th17 suppression and Th2 expansion. Clin. Exp. Rheumatol. 2014;32:77–81. [PubMed] [Google Scholar]
- 29.Bonagura VR, Hatam L, DeVoti J, Zeng FF, Steinberg BM. Recurrent respiratory papillomatosis: Altered CD8(+) T-cell subsets and T(H)1/T(H)2 cytokine imbalance. Clin Immunol. 1999;93:302–11. doi: 10.1006/clim.1999.4784. [DOI] [PubMed] [Google Scholar]
- 30.Rosenthal DW, et al. Recurrent respiratory papillomatosis (RRP): Increased T(H)2-like chemokine expression. J. Allergy Clin. Immunol. 2006;117:S104. [Google Scholar]
- 31.Rosenthal DW, Devoti JA, Steinberg BM, Abramson AL, Bonagura VR. TH2-like chemokine patterns correlate with disease severity in patients with recurrent respiratory papillomatosis. Mol Med. 2012;18:1338–45. doi: 10.2119/molmed.2012.00284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bonagura VR, et al. HLA alleles, IFN-gamma responses to HPV-11 E6, and disease severity in patients with recurrent respiratory papillomatosis. Human Immunol. 2004;65:773–82. doi: 10.1016/j.humimm.2004.05.014. [DOI] [PubMed] [Google Scholar]
- 33.Ramadas RA, Ewart SL, Medoff BD, LeVine AM. Interleukin-1 family member 9 stimulates chemokine production and neutrophil influx in mouse lungs. Am. J. Respir. Cell Mol. Biol. 2011;44:134–45. doi: 10.1165/rcmb.2009-0315OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chustz RT, et al. Regulation and function of the IL-1 family Cytokine IL-1F9 in human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 2011;45:145–53. doi: 10.1165/rcmb.2010-0075OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Towne JE, et al. Interleukin-36 (IL-36) ligands require processing for full agonist (IL-36alpha, IL-36beta, and IL-36gamma) or antagonist (IL-36Ra) activity. J. Biol. Chem. 2011;286:42594–602. doi: 10.1074/jbc.M111.267922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Steinman RM. The dendritic cell system and its role in immunogenicity. Annu. Rev. Immunol. 1991;9:271–96. doi: 10.1146/annurev.iy.09.040191.001415. [DOI] [PubMed] [Google Scholar]
- 37.Hart DN. Dendritic cells: unique leukocyte populations which control the primary immune response. Blood. 1997;90:3245–87. [PubMed] [Google Scholar]
- 38.Matzinger P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994;12:991–1045. doi: 10.1146/annurev.iy.12.040194.005015. [DOI] [PubMed] [Google Scholar]
- 39.Noel W, et al. Alternatively activated macrophages during parasite infections. Trends Parasitol. 2004;20:126–33. doi: 10.1016/j.pt.2004.01.004. [DOI] [PubMed] [Google Scholar]
- 40.McGuirk P, McCann C, Mills KH. Pathogen-specific T regulatory 1 cells induced in the respiratory tract by a bacterial molecule that stimulates interleukin 10 production by dendritic cells: a novel strategy for evasion of protective T helper type 1 responses by Bordetella pertussis. J. Exp. Med. 2002;195:221–31. doi: 10.1084/jem.20011288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shortman K, Liu YJ. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2002;2:151–61. doi: 10.1038/nri746. [DOI] [PubMed] [Google Scholar]
- 42.Shirakata Y. Regulation of epidermal keratinocytes by growth factors. J. Dermatol. Sci. 2010;59:73–80. doi: 10.1016/j.jdermsci.2010.05.002. [DOI] [PubMed] [Google Scholar]
- 43.Werner S, Krieg T, Smola H. Keratinocyte-fibroblast interactions in wound healing. J. Invest. Dermatol. 2007;127:998–1008. doi: 10.1038/sj.jid.5700786. [DOI] [PubMed] [Google Scholar]
- 44.Pastore S, Mascia F, Girolomoni G. The contribution of keratinocytes to the pathogenesis of atopic dermatitis. Eur. J. Dermatol. 2006;16:125–31. [PubMed] [Google Scholar]
- 45.Bergmann C, et al. T regulatory type 1 cells in squamous cell carcinoma of the head and neck: mechanisms of suppression and expansion in advanced disease. Clin Cancer Res. 2008;14:3706–15. doi: 10.1158/1078-0432.CCR-07-5126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Banchereau J, et al. Immunobiology of dendritic cells. Annu. Rev. Immunol. 2000;18:767–811. doi: 10.1146/annurev.immunol.18.1.767. [DOI] [PubMed] [Google Scholar]
- 47.Banchereau J, et al. Dendritic cells: controllers of the immune system and a new promise for immunotherapy. Ann. N. Y. Acad. Sci. 2003;987:180–7. doi: 10.1111/j.1749-6632.2003.tb06047.x. [DOI] [PubMed] [Google Scholar]
- 48.Caux C, et al. CD34+ hematopoietic progenitors from human cord blood differentiate along two independent dendritic cell pathways in response to granulocyte-macrophage colony-stimulating factor plus tumor necrosis factor alpha: II. Functional analysis. Blood. 1997;90:1458–70. [PubMed] [Google Scholar]
- 49.Steinman RM, Turley S, Mellman I, Inaba K. The induction of tolerance by dendritic cells that have captured apoptotic cells. J. Exp. Med. 2000;191:411–6. doi: 10.1084/jem.191.3.411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Iwasaki A, Kelsall BL. Freshly isolated Peyer’s patch, but not spleen, dendritic cells produce interleukin 10 and induce the differentiation of T helper type 2 cells. J. Exp. Med. 1999;190:229–39. doi: 10.1084/jem.190.2.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Enk AH, Angeloni VL, Udey MC, Katz SI. Inhibition of Langerhans cell antigen-presenting function by IL-10. A role for IL-10 in induction of tolerance. J Immunol. 1993;151:2390–8. [PubMed] [Google Scholar]
- 52.Beissert S, Hosoi J, Grabbe S, Asahina A, Granstein RD. IL-10 inhibits tumor antigen presentation by epidermal antigen-presenting cells. J Immunol. 1995;154:1280–6. [PubMed] [Google Scholar]
- 53.Romani N, et al. Epidermal Langerhans cells—changing views on their function in vivo. Immunol Lett. 2006;106:119–25. doi: 10.1016/j.imlet.2006.05.010. [DOI] [PubMed] [Google Scholar]
- 54.Elias AN, Nanda VS, Barr RJ. CD1a expression in psoriatic skin following treatment with propylthiouracil, an antithyroid thioureylene. BMC Dermatol. 2003;3:3. doi: 10.1186/1471-5945-3-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kaplan DH, Jenison MC, Saeland S, Shlomchik WD, Shlomchik MJ. Epidermal Langerhans cell-deficient mice develop enhanced contact hypersensitivity. Immunity. 2005;23:611–20. doi: 10.1016/j.immuni.2005.10.008. [DOI] [PubMed] [Google Scholar]
- 56.Klechevsky E, et al. Functional specializations of human epidermal Langerhans cells and CD14+ dermal dendritic cells. Immunity. 2008;29:497–510. doi: 10.1016/j.immuni.2008.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Bennett CL, et al. Langerhans cells regulate cutaneous injury by licensing CD8 effector cells recruited to the skin. Blood. 2011;117:7063–9. doi: 10.1182/blood-2011-01-329185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.James E, et al. Human papillomavirus (HPV)-specific T-cells recognizing dominant E2/E6 epitopes elicit reduced IFN-γ in patients with recurrent respiratory papillomatosis (RRP) J. Allergy Clin. Immunol. 2010;125 S. [Google Scholar]
- 59.James EA, et al. Papillomavirus-specific CD4(+) T cells exhibit reduced STAT-5 signaling and altered cytokine profiles in patients with recurrent respiratory papillomatosis. J Immunol. 2011;186:6633–40. doi: 10.4049/jimmunol.1004181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Chopin M, et al. Langerhans cells are generated by two distinct PU.1-dependent transcriptional networks. J. Exp. Med. 2013;210:2967–80. doi: 10.1084/jem.20130930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Chopin M, Nutt SL. Establishing and maintaining the Langerhans cell network. Semin. Cell. Dev. Biol. 2014 2014 Feb 7; doi: 10.1016/j.semcdb.2014.02.001. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 62.Bonagura VR, et al. Activating killer cell immunoglobulin-like receptors 3DS1 and 2DS1 protect against developing the severe form of recurrent respiratory papillomatosis. Human Immunol. 2010;71:212–9. doi: 10.1016/j.humimm.2009.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gombert M, et al. CCL1-CCR8 interactions: an axis mediating the recruitment of T cells and Langerhans-type dendritic cells to sites of atopic skin inflammation. J Immunol. 2005;174:5082–91. doi: 10.4049/jimmunol.174.8.5082. [DOI] [PubMed] [Google Scholar]
- 64.Henry E, et al. Dendritic cells genetically engineered to express IL-10 induce long-lasting antigen-specific tolerance in experimental asthma. J Immunol. 2008;181:7230–42. doi: 10.4049/jimmunol.181.10.7230. [DOI] [PubMed] [Google Scholar]
- 65.Dieu-Nosjean MC, et al. Macrophage inflammatory protein 3alpha is expressed at inflamed epithelial surfaces and is the most potent chemokine known in attracting Langerhans cell precursors. J. Exp. Med. 2000;192:705–18. doi: 10.1084/jem.192.5.705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Merad M, et al. Langerhans cells renew in the skin throughout life under steady-state conditions. Nat Immunol. 2002;3:1135–41. doi: 10.1038/ni852. [DOI] [PMC free article] [PubMed] [Google Scholar]