

Abstract
Inter-kingdom and interspecies interactions are ubiquitous in nature and are important for the survival of species and ecological balance. The investigation of microbe-microbe interactions is essential for understanding the in vivo activities of commensal and pathogenic microorganisms. Candida albicans, a polymorphic fungus, and Pseudomonas aeruginosa, a Gram-negative bacterium, are two opportunistic pathogens that interact in various polymicrobial infections in humans. To determine how P. aeruginosa affects the physiology of C. albicans and vice versa, we compared the proteomes of each species in mixed biofilms versus single-species biofilms. In addition, extracellular proteins were analyzed. We observed that, in mixed biofilms, both species showed differential expression of virulence proteins, multidrug resistance-associated proteins, proteases and cell defense, stress and iron-regulated proteins. Furthermore, in mixed biofilms, both species displayed an increase in mutability compared with monospecific biofilms. This characteristic was correlated with the downregulation of enzymes conferring protection against DNA oxidation. In mixed biofilms, P. aeruginosa regulates its production of various molecules involved in quorum sensing and induces the production of virulence factors (pyoverdine, rhamnolipids and pyocyanin), which are major contributors to the ability of this bacterium to cause disease. Overall, our results indicate that interspecies competition between these opportunistic pathogens enhances the production of virulence factors and increases mutability and thus can alter the course of host-pathogen interactions in polymicrobial infections.
Keywords: species interactions, mixed biofilm, virulence, antagonism, siderophores, iron
Introduction
In many natural environments and chronic human infections, microbes live in matrix-encased groups known as biofilms, rather than as free-living (planktonic) cells. In biofilms, cells are known to show distinct expression profiles that are not apparent in planktonic cells. For example, biofilm bacteria are more resistant to killing by antibiotics, biocides and host defenses than planktonic cells and may be less vulnerable to predators (Peleg et al., 2010). Microbial biofilms represent heterogeneous populations of species that form intimate contacts. Within these populations, species communicate, cooperate and compete (Davey and O'toole, 2000; Tyson et al., 2004; Hansen et al., 2007; Harrison et al., 2008; Garbeva et al., 2011). Some of these microbial interactions are beneficial, and some are detrimental from a human standpoint (Hughes and Kim, 1973; Lynch and Robertson, 2008).
The interactions between Pseudomonas aeruginosa and Candida albicans provide a model for many bacteria–eukaryote interactions. An in-depth understanding of these interactions could be exploited for the benefit of mankind. Pseudomonas and Candida are organisms that are commonly isolated from the sputum of cystic fibrosis patients (Hughes and Kim, 1973; Bauernfeind et al., 1987; Bakare et al., 2003). Both species are common non-pathogenic commensals of healthy individuals. In compromised individuals, they are able to initiate invasive growth that may result in serious disease and death (Naglik et al., 2004; Hube, 2006; Pfaller and Diekema, 2007). C. albicans is a dimorphic fungus that can live as a yeast or in a filamentous form. In response to certain stimuli (signaling molecules, temperature and host factors), it switches from its yeast to its hyphal form. Both morphological forms are important for virulence, and the ability to undergo morphological transformation is therefore an important virulence trait (Gow, 1997; Calderone and Fonzi, 2001; Gow et al., 2002; Liu, 2002; Whiteway and Oberholzer, 2004). Several studies have suggested that P. aeruginosa may inhibit C. albicans growth within the host (Bauernfeind et al., 1987; Kerr, 1994; Burns et al., 1999; Gupta et al., 2005; Kaleli et al., 2007). Hogan and Kolter (2002) demonstrated that P. aeruginosa is cytotoxic to the filamentous form of C. albicans but is unable to attach to or kill C. albicans yeast cells. In addition, it has been reported that 3-oxo-C12HSL, a signaling molecule produced by P. aeruginosa, and structurally related molecules can inhibit and even reverse the switch from yeast to hyphal growth in C. albicans (Hogan et al., 2004). Interestingly, in vitro studies have demonstrated that these signaling interactions between P. aeruginosa and C. albicans may in fact be bidirectional. It has been observed that the addition of farnesol leads to decreased Pseudomonas quinolone signal (PQS) production in P. aeruginosa and reduced levels of the PQS-regulated virulence factor pyocyanin (Cugini et al., 2007).
In a burned mouse model, Neely et al. (1986) demonstrated that Pseudomonas infections could predispose burned mice to fatal candidiasis and that the proteolytic activity generated by the bacteria was primarily responsible for the establishment of lethal fungal infections.
To understand the interactions between pathogens and the resident microbiome and how they affect gene expression patterns in the pathogens and contribute to bacterial diseases, Duan et al. (2003) investigated the interactions between pathogenic P. aeruginosa and avirulent oropharyngeal flora strains isolated from sputum samples of cystic fibrosis patients. Their results suggest that important contributions of the host microflora to P. aeruginosa infection occur via the modulation of gene expression through interspecies communications (autoinducer-2-mediated quorum sensing).
Using a Drosophila model of polymicrobial infections, Sibley et al. (2008) demonstrated that a large proportion of the organisms found in cystic fibrosis airways have the ability to act synergistically with P. aeruginosa. This synergy involves microbe–microbe interactions that result in modulation of P. aeruginosa virulence factor gene expression within infected Drosophila. Nevertheless, the mechanisms underlying this synergy were not identified.
Few studies have demonstrated that physiological processes are involved in the C. albicans–P. aeruginosa interaction and the impacts of this interaction on the ecology of the C. albicans–P. aeruginosa community. To determine how P. aeruginosa affects the physiology of C. albicans and vice versa, we compared the proteome of each species in mixed biofilms with single-species biofilms (Figure 1). In addition, we conducted an analysis of secreted proteins and small molecules involved in the interaction. To our knowledge, this is the first whole-cell proteome analysis of microbial species that coexist and form mixed biofilms.
Figure 1.

Experimental design for the proteomic analysis of C. albicans-P. aeruginosa interaction. Single- and mixed-species biofilms were cultured under hypoxic conditions in polystyrene Petri plates. To generate mixed biofilms, we inoculated with P. aeruginosa (Pa.) ∼1.5 × 108 cells ml−1 and ∼1 × 106 C. albicans (Ca.) cells ml−1 in Petri plates at the beginning of each experiment. For the single-species biofilms, the medium initially contained ∼1.5 × 108 bacterial cells ml−1 or ∼1 × 106 yeast cells ml−1. At 24 h postculture, the medium with planktonic cells was removed, and the biofilm at the bottom of the plate was scraped. The fungi and bacteria were separated from the mixed biofilms by centrifugation in a discontinuous Percoll gradient. The monospecific biofilms received the same treatment as the mixed biofilms. Whole-cell proteins of three independent cultures (biological replicates) were analyzed by 2D-PAGE. Spots whose relative expression changed in P. aeruginosa or C. albicans in single versus mixed biofilms were identified by MALDI-TOF mass spectrometry.
Here we show that interspecies competition for iron between these opportunistic pathogens causes encompasses changes in the expression/production of factors that have been previously identified as virulence factors in studies of disease. In consequence, iron competition has the potential to cause significant ‘collateral damage' to the host and determines the course and severity of a mixed infection.
Materials and methods
Microbial culture and growing conditions
P. aeruginosa PAO1 (Holloway, 1955) and C. albicans CAI4 (ura3::imm434/ura3::imm434) (Fonzi and Irwin, 1993) were cultured for 14 h at 30 °C in LB medium or YPD (1% yeast extract, 2% bactopeptone and 2% dextrose), respectively. The cells were then collected via centrifugation and washed twice with water. These cells were used for interaction experiments. Single- and mixed-species biofilms were cultured in 10-cm diameter polystyrene Petri plates containing 20 ml of RPMI 1640 medium plus L-glutamine (GIBCO no.11875, Grand Island, NY, USA), supplemented with 50 mM dextrose and 40 μg ml−1 uridine. This is a complete synthetic medium that contains vitamins, amino acids, inorganic salts and glutathione. It contains no proteins or growth-promoting agents. To generate mixed biofilms, we inoculated with ∼1.5 × 108 bacterial cells ml−1 and ∼1 × 106 fungal cells ml−1 in each Petri plates at the beginning of an experiment. For the single-species biofilms, the medium initially contained ∼1.5 × 108 bacterial cells ml−1 or ∼1 × 106 yeast cells ml−1. The cultures were incubated at 37 °C without shaking.
To obtain biofilms under hypoxic conditions, the plates were wrapped with Parafilm. In contrast, to obtain biofilms under normoxic conditions, the plates were not wrapped. At 24 h postculture, the medium with planktonic cells was removed, and 5 ml of cold stop buffer was added (20 mM sodium azide, 200 mM Tris-HCl pH 8, 20 mM EDTA, 300 μg ml−1 rifampicin and 500 μg ml−1 chloramphenicol), after which the biofilm that had formed at the bottom of the plate was scraped. The biofilms were collected via centrifugation at 10 000 g for 5 min at 4 °C, and the pellet was frozen with liquid nitrogen and stored at −70 °C.
Determination of populations dynamics
The total population (including planktonic and sessile cells) of each species in the cultures was determined by calculating the number of CFU (colony-forming units) ml−1. Samples of the cultures were diluted in Tween solution (0.01% Tween 80 and 10 mM MgSO4), sonicated for 30 s and plated on LB agar to determine the population densities of P. aeruginosa. To estimate the C. albicans population density, samples were centrifuged (4 min at 3000 g), washed and deflocculated using 300 mM EDTA before being diluted, sonicated for 30 s and plated on YPD agar, as previously described by Smukalla et al. (2008).
Iron and oxygen quantitation
The concentration of iron ions in the culture supernatant was determined separately for Fe2+ and Fe3+ through spectrophotometric assays using iron test kit no. 1.14761.0002 (Merck KGaA, Darmstadt, Germany). The concentration of dissolved oxygen in the culture at 24 h was determined using an OXEL-1 oxygen electrode (World Precision Instruments, Sarasota, FL, USA).
Purification of whole-cell proteins
Whole-cell proteins were isolated from three independent cultures (biological replicates). The fungi and bacteria were separated from the mixed biofilms via centrifugation in a discontinuous Percoll gradient, as described in the Supplementary Methods. The monospecific biofilms received the same treatment as the mixed biofilms. Protein isolation was performed through phenol extraction, as described in the Supplementary Methods.
Extracellular protein extraction
At 24 h after the initiation of the biofilm cultures, the medium was removed and centrifuged at 6000 g for 5 min. The supernatant was passed through a 0.45-μm pore-size membrane and stored at −20 °C. A 400-ml aliquot of the supernatant was subsequently lyophilized, and protein isolation was performed through phenol extraction (Hurkman and Tanaka, 1986).
Two-dimensional gel electrophoresis
The methods applied for sample preparation, preparative two-dimensional gel electrophoresis (2D PAGE) and image analysis have been previously described (Encarnación et al., 2003). Through isoelectric focusing, 600 μg of whole-cell protein and secreted protein was separated across a linear pH range of 3–10. All gel experiments were performed with proteins isolated from three independent cultures.
Analysis of differential protein expression
For this analysis, 2D images were captured by scanning Coomassie blue-stained gels using a GS-800 imaging densitometer (Bio-Rad, Hercules, CA, USA) and analyzed with the PDQuest 2-D Analysis software (Bio-Rad). Three gels obtained from three different assays were analyzed to guarantee representative results. Spot normalization, as a means of internal calibration to ensure the independence of the data from experimental variations between gels, was performed using relative volumes (%Vol) to quantify and compare the obtained gel spots. The %Vol corresponds to the volume of each spot divided by the total volume of all of the spots in the gel. Student's t-test was performed to assess the significance of differences between differentially expressed proteins. Based on the average spot volume ratio, spots whose relative expression changed at least 1.5-fold (increase or decrease) in P. aeruginosa or C. albicans in single versus mixed biofilms were considered significantly different, with P<0.05.
Determination of P. aeruginosa mutability
The mutability of P. aeruginosa in monocultures and cocultures was estimated by determining the frequency of spontaneous mutants resistant to rifampicin (RifR). Mutability was evaluated in heat-inactivated fetal bovine serum and RPMI medium under hypoxic conditions. Cultures in RPMI were performed in Petri plates as described above. Cultures in fetal bovine serum (Lonza, Cat. no. 14-502, Walkersville, MD, USA) supplemented with 10 mM dextrose and 20 μg ml−1 uridine were performed in 24-well plates containing 1 ml of fetal bovine serum per well. To generate monocultures, each well was inoculated with ∼5 × 106 bacterial cells ml−1. For the cocultures, the medium initially contained ∼5 × 106 bacterial cells ml−1 and ∼1 × 107 C. albicans cells ml−1. Plates were wrapped with Parafilm and incubated at 37 °C without shaking for 24 h. Three mililiters of culture were centrifuged, and the cells were spread on a LB plates containing rifampicin (100 μg ml−1) and incubated at 37 °C. Dilutions were also plated on LB plates without antibiotics to determine the total number of CFUs. The colonies were scored for Rif resistance 48 h after plating. The mutation frequencies were approximated as the mean number of RifR cells divided by the total number of CFUs.
Statistical analysis
All data were calculated as the mean±s.d. of at least three experiments. Statistical analysis was performed using the Student's t-test, and differences were considered statistically significant at P<0·05 compared with control experiments.
Results and discussion
Population dynamics during P. aeruginosa–C. albicans interaction
Studies have established that P. aeruginosa affects the growth of C. albicans in nutrient-rich environments and under normoxic conditions (Hogan and Kolter 2002; El-Azizi et al., 2004). Here we analyzed the development of the P. aeruginosa–C. albicans interaction in complete synthetic defined medium under environmental conditions similar to those observed in infections in which both species interact: hypoxic conditions (0.1% dissolved oxygen±0.08 s.d.) with low-iron concentrations (67 nM±12.5 s.d.). We observed that during the first 24 h the growth dynamics of C. albicans and P. aeruginosa were similar between cocultures and monocultures. In contrast, 24 h later, the presence of bacteria led to a rapid decrease in the fungal population (Figure 2). Microscopic analysis revealed that, after 48 h of coculture, the fungus cells had been killed by the bacterium (Figures 2c and d).
Figure 2.

Growth dynamics of C. albicans (squares) and P. aeruginose (circles) in (a) cocultures and (b) monocultures. Growth was based on colony-forming units (CFU). (c, d) Epifluorescence images from mixed C. albicans–P. aeruginosa biofilms stained with the LIVE/DEAD staining system are shown. A viability assay confirmed that C. albicans cells lost viability after 48 h in mixed biofilms. Red cells (stained with propidium iodide) are considered dead, while live cells remained green (stained with SYTO9). The panels present the means±s.e.m. (n=3).
Microbes competing with unrelated or distantly related species for limited resources in the same niche activate mechanisms such as secretion of proteins or small molecules to attack the competing strains or species (Czaran et al., 2002; Be'er et al., 2009). We hypothesized that during the first 24 h of coculture, the C. albicans and P. aeruginosa compete for nutrients, after which a parasitism is established in which the bacterium is the parasite, and the fungus is the host.
To understand the molecular mechanisms involved in this interaction, we compared the proteome of each species in mixed biofilms with single-species biofilms after 24 h of coculture.
Low oxygen levels influence the C. albicans–P. aeruginosa interaction
Previous in vitro studies have demonstrated that 3-oxo-C12 homoserine lactone, a cell–cell signaling molecule produced by P. aeruginosa, inhibits C. albicans filamentation, without affecting fungal growth rates in cocultures (Hogan et al., 2004). Interestingly, we observed that in cocultures under hypoxic conditions (0.1% dissolved oxygen±0.08 s.d.), P. aeruginosa was unable to completely inhibit the filamentation of C. albicans, in contrast to cocultures under normoxia (4% oxygen±0.18 s.d.) (Figures 3a and b). As acyl homoserine lactones (AHLs) are responsible for inhibiting fungal filamentation, we determined the concentrations of these molecules in cocultures and monocultures under normoxic and hypoxic conditions (Supplementary Materials and Methods). Figure 3b shows that the AHL concentration was lower under hypoxic conditions in the presence of C. albicans. The low concentrations of AHLs observed in hypoxic cocultures were not caused by a reduced population of bacteria, as the cell density of the cocultures was similar under hypoxia and normoxia (6.7 × 108±1.5 and 7.2 × 108±1.3 CFU ml−1). These results suggest that a low oxygen level and the interaction with the fungus are two factors that influence AHL production and therefore alter intraspecific and interspecific communication.
Figure 3.

P. aeruginosa inhibits the yeast-to-hyphal transition under normoxia but not under hypoxic conditions. (a) Percentage of hyphae observed in normoxic and hypoxic cultures after 24 h: CA; C. albicans. (b) Thin-layer chromatography (TLC) of acyl-HSLs produced by P. aeruginosa (PA) in monocultures and cocultures: Hypox, hypoxia and Nor, normoxia. The samples were obtained after 24 h of culture and processed as described in Supplementary Materials and Methods. Acyl-HSL standards: C12, 3-Oxo-C12-HSL, and C8-HSL. The asterisk indicates undetectable acyl-HSLs in a hypoxic coculture. The panels present the means±s.e.m. (n=3).
It has been shown that low oxygen enhances C. albicans filamentation (Dumitru et al., 2004) and that the fungal response to hypoxia overlaps the response to low iron (Synnott et al., 2010), indicating that competition for iron may be greater in hypoxic conditions and may be connected with hyphae formation. A genome-wide transcriptional analysis of morphology determination in C. albicans showed that a significant number of genes required for iron utilization were overexpressed during the yeast-hyphal transition (Carlisle and Kadosh, 2013). These data suggest a link between iron metabolism and yeast-hyphal transition; however, this has not been investigated. Taking together these results, we suggest that the low AHLs concentration, hypoxic condition and iron competition may be factors influencing the fungus filamentation in mixed biofilms under hypoxia.
Whole-cell proteome response of C. albicans and P. aeruginosa in coculture
Proteomics experiments were performed to identify P. aeruginosa and C. albicans proteins that were differentially regulated in mixed biofilms after 24 h of growth under hypoxic conditions. Bacteria and fungi were grown together in liquid medium under conditions that allowed physical interaction of the two species to form a mixed biofilm on a polystyrene surface (Figure 2c). Fungal and bacterial cells were separated from the mixed biofilms, and protein expression patterns were determined via 2D-PAGE. A total of ∼700 spots were detected and analyzed as described in the Materials and methods section (Supplementary Figures S1 and S2). The average protein levels in the spots were quantified, and those showing relative changes in abundance of>1.5-fold between conditions (increase or decrease) at the 95% confidence level (P<0.05) were considered significantly different.
The interaction with C. albicans induced changes in the proteome of P. aeruginosa, resulting in differential expression of 163 electrophoretic entities, 108 of which were identified (Supplementary Table S1). In addition, up to 126 electrophoretic entities obtained from C. albicans displayed altered expression in response to P. aeruginosa, 110 of which were identified (Supplementary Table S2).
The siderophore-mediated iron acquisition system has an essential and dominant role in C. albicans–P. aeruginosa mixed biofilms
Siderophores are small, high-affinity iron-chelating compounds secreted by microorganisms in response to iron limitation. Because the levels of free ferric iron in biological systems are always extremely low (10−8 M), siderophores have an important role in microbe–microbe and host-pathogen interactions (Ratledge and Dover, 2000; Harrison et al., 2008). For example, P. aeruginosa mutants incapable of either siderophore synthesis or siderophore transport are far less virulent than their wild-type counterparts (Takase et al, 2000). In the context of microbial interactions, iron competition in Pseudomonas has been studied, and the role of the pyoverdine siderophore produced by Pseudomonas species has been clearly demonstrated in competition with fungus and bacteria (Loper and Buyer, 1991; Harrison et al., 2008). For example, Purschke et al. (2012) demonstrated that P. aeruginosa increases pyoverdine production in response to iron competition with C. albicans in mixed biofilms. Consistent with these findings, we identified proteins with known roles in iron uptake mediated by siderophores that were upregulated in P. aeruginosa in mixed biofilms in response to the interaction with C. albicans. Among the most highly induced proteins were the group of siderophore receptors constituted by the ferric pyochelin receptor FptA, the hydroxamate-type ferri-siderophore receptor FiuA, the citrate hydroxamate siderophore receptor ChtA, the ferrienterobactin receptor PfeA, the ferripyoverdine receptor FpvA, the alternative type I ferripyoverdine receptor FpvB and the putative TonB-dependent receptor CirA. In addition, the expression of proteins involved in the biosynthesis of the siderophore pyoverdine (PvdH and PvdA) and two proteins (HasR and PhuR) required for heme and hemoglobin uptake was highly induced in mixed biofilms (Supplementary Table S1).
Supporting the above results, we observed that the pyoverdine production per cell was significantly increased in the presence of C. albicans (Figure 4a). We tested the ability of C. albicans-conditioned medium to induce pyoverdine production. We observed that a methanol extract of the culture supernatant increased the production of pyoverdine (Figure 4a). The extracts were fractionated through high-performance liquid chromatography (Figure 4b). A total of 12 fractions were evaluated, one of which, fraction 3 (F3), showed the capacity to induce the synthesis of pyoverdine (Figure 4a). Further experiments will be needed to determine the molecular structure of the active compound in this fraction. However, the preliminary results suggest that it contains phenolate molecules (Figure 4c). We propose that these molecules are produced by C. albicans to capture iron and may function as siderophores.
Figure 4.

The production of pyoverdine is increased in mixed biofilm. (a) Pyoverdine production under P. aeruginosa monoculture (Mon), coculture (Coc), P. aeruginosa monoculture+a methanolic extract from C. albicans supernatant (Ex) and P. aeruginosa monoculture+HPLC (high-performance liquid chromatographic) fractions of ethyl acetate extracts from C. albicans supernatants (F1-F4) and solvent control—ethyl acetate (−). All experiments were conducted in hypoxia. (b) Chromatogram of an ethyl acetate extract from the C. albicans supernatant. (c) TLC to detect siderophores. Ethyl acetate extracts were obtained from P. aeruginosa monocultures (Pa.), cocultures (Coc) and C. albicans monocultures (Ca.). C. albicans phenolates are indicated by asterisks. The panels present the means±s.e.m. (n=3).
We hypothesized that iron competition has an important role in the development of the interaction. To examine this hypothesis, we evaluated the effect of iron supplementation on the growth of the fungus and bacterium. We observed that iron supplementation significantly increased the growth of the bacterium in both mixed and pure cultures (Figure 5a). In contrast, in C. albicans monocultures, iron supplementation did not increase the growth of the fungus (Figure 5b), suggesting that the iron content in the unsupplemented culture is sufficient to support maximal growth of the fungus but not that of the bacterium. In the iron-supplemented medium, an early antagonistic interaction was established, in which the rapid growth of the bacterium eliminated the fungal population (Figure 5b).
Figure 5.

Effects of iron on P. aeruginosa (a) and C. albicans (b) growth. Monocultures (Mon) and cocultures (Coc). Growth was determined after 24 h of culture. To quantify the growth of C. albicans and P. aeruginosa in cocultures, cells (including planktonic and sessile cells) were separated on a Percoll gradient (Supplementary Materials and Methods) and were suspended in 20 ml water. Optical density (OD600) for each species was determined. The panels present the means±s.e.m. (n=3).
As yeast mutants deficient in iron utilization were unable to compete with P. aeruginosa in mixed biofilms (Supplementary Figure S3), we propose that the iron uptake ability of the fungus is a determining factor for the establishment of the interaction. Thus, these results indicate that, in mixed biofilms, the competence for iron uptake determines the nature and dynamics of the interaction.
Iron-regulated virulence factors of P. aeruginosa are overexpressed in mixed biofilms
Lamont et al. (2002) demonstrated that in addition to its role as an iron scavenger, the siderophore pyoverdine acts as a signaling molecule to control the production of at least three virulence factors (exotoxin A, an endoprotease and pyoverdine itself) that are major contributors to the ability of P. aeruginosa to cause disease. As pyoverdine production was increased in the mixed biofilms (Figure 4a), we hypothesized that the bacterium shows increased virulence when competing for iron with the fungus. To test this hypothesis, we analyzed the extracellular proteins present in the cocultures and monocultures.
In C. albicans monocultures, the concentration of total extracellular protein was significantly lower than the concentration detected in monocultures of P. aeruginosa (Supplementary Figure S4). In our analysis of coculture supernatants, we were not able to identify C. albicans proteins, which could be due to slow production of extracellular proteins by C. albicans and bacterial protease activity. Extracts of secreted proteins were analyzed via 2D-PAGE, and a total of ∼300 spots were detected, 75 of which were differentially expressed, and 55 of these were identified (Supplementary Table S3 and Supplementary Figure S4).
As previously reported by Purschke et al. (2012), we observed a significant increase in the production of iron-regulated proteases by P. aeruginosa in coculture supernatants, including PrpL, PasP, AprA and a hypothetical protein zinc-protease, PA0572 (Supplementary Table S3) (Wilderman et al., 2001; Marquart et al., 2005). These proteins are known for their strong proteolytic activities, which are responsible for rupturing tight junctions in the epithelium, leading to tissue invasion and bacterial spreading (Azghani et al., 1993). In contrast, the quorum-sensing-regulated proteases, including LasB and PepB, and a chitin-binding protein were downregulated in the cocultures, which is consistent with the low concentration of AHLs detected in cocultures under hypoxic conditions (Figure 3b).
Interestingly, we observed a significant increase in the production of PvdQ in the coculture supernatant (Supplementary Table S3). PvdQ is an iron-regulated AHL acylase that degrades long-acyl but not short-acyl AHLs (Sio et al., 2006; Nadal et al., 2010). This result may provide an explanation for the previous finding that the level of long-acyl AHLs is greatly reduced in cocultures as compared with P. aeruginosa pure cultures (Figure 3b).
Kirienko et al. (2013) demonstrated that P. aeruginosa does not require quorum-sensing pathways to kill Caenorhabditis elegans. Supporting a role for iron in P. aeruginosa pathogenesis, pyoverdine was found to be required to cause hypoxia and death in C. elegans.
The increased virulence was confirmed by our observation of an enhancement of the proteolytic activity of coculture supernatants against human serum proteins and against the adhesion of HeLa cells (Figures 6a and b). We propose that a significant increase of iron-regulated proteases could be a bacterial strategy for affecting the viability of C. albicans and causing release of iron from fungal proteins.
Figure 6.

Assays of cytotoxic activity of microbial culture supernatants and production of P. aeruginosa virulence factors. (a) Effect of supernatants on HeLa cells adherence. Cells were incubated with P. aeruginosa monoculture supernatants (Pa.), coculture supernatants (Coc.), C. albicans monoculture supernatants (Ca.) or phosphate-buffered saline (PBS) buffer (Control). The adhered cells were measured by crystal violet staining (DO570). (b) Degradation of serum proteins after exposure to microbial culture supernatants. Serum was incubated with P. aeruginosa monoculture supernatants, coculture supernatants, C. albicans monoculture supernatants or PBS buffer. The reactions were analyzed via sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The figure shows a representative image from three independent experiments. Albumin (asterisk) was totally degraded in the coculture supernatant. In contrast, the C. albicans supernatant showed no effect on albumin. (c, d) The production of rhamnolipids and pyocyanin is increased in mixed biofilms. Panel (c) shows rhamnolipid relative concentration in supernatants. Thin-layer chromatographic (TLC) analysis of P. aeruginosa rhamnolipid biosynthesis in P. aeruginosa monocultures (Pa.) and cocultures (Coc). Di-rhamnolipid production was measured by dividing the optical density (OD) of the spots in the TLC with the P. aeruginosa cell density (OD600). The bacteria cell density was determined as described in the legend of Figure 4. (d) Pyocyanin concentrations in culture supernatants.
Consistent with a role for pyoverdine as a signaling molecule that controls the production of virulence factors, we observed an increased concentration of Exotoxin A in coculture supernatants (Supplementary Table S3), which is one of the most powerful extracellular virulence factors produced by P. aeruginosa (Iglewski and Kabat, 1975).
To obtain iron in vivo, bacteria additionally or alternatively use other mechanisms. They obtain iron directly from host iron sources via specific outer membrane receptors that bind free heme (Ochsner et al., 2000). In mixed biofilms, we detected two secreted proteins from the heme-acquisition pathway, HasA and HasR. These proteins are considered to be important virulence factors that have a role in colonization, survival, tissue invasion and in the ultimate damage caused to the host.
Taking these results together, we suggest that competition for iron is the main mechanism that triggers the virulence of P. aeruginosa in mixed biofilms. In this manner, iron competition has the potential to cause significant ‘collateral damage' to the host and determines the course and severity of a mixed infection.
The production of small molecules regulated by the quinolone signaling system is increased in mixed biofilms
Rhamnolipids and pyocyanin (a phenazine pigment) are important P. aeruginosa virulence factors. These compounds are known for their toxicity in other bacteria (Baron and Rowe, 1981) and in eukaryotes ranging from nematodes to humans (Ran et al., 2003; Lau et al., 2004; Mavrodi et al., 2006; Morales et al., 2010; Morales et al., 2013).
We observed a significant increase in the concentration of rhamnolipids and pyocyanin in the coculture supernatants (Figures 6c and d). The production of rhamnolipids in P. aeruginosa is controlled by both the LasR/3-oxo-C12-HSL quorum-sensing signaling pathway and the MvfR/PQS/PqsE quinolone signaling system, which uses 2-heptyl-3-hydroxy-4(1H)-quinolone (PQS) (Diggle et al., 2003). Is known that iron limitation promote the production of rhamnolipids, because the expression of genes for rhamnolipids biosynthesis and their transcriptional regulator rhlR, are enhanced by iron limitation (Bredenbruch et al., 2006).
Similar to AHLs, PQS regulates the production of virulence determinants, including rhamnolipids, pyocyanin, elastase and the galactophilic lectin LecA, in addition to influencing biofilm development (Pesci et al., 1999; Diggle et al., 2003).
As we observed that there was a reduced concentration of AHLs in the cocultures (probably due to upregulation of PvdQ triggered by iron competition), we propose that, in mixed biofilms, the increased production of rhamnolipids and pyocyanin is due to the activation of the PQS signaling pathway rather than the LasR/3-oxo-C12-HSL pathways. PQS has been demonstrated to induce the expression of genes associated with the iron-deprivation pathway, in which the iron-chelating activity of PQS induces pyoverdine production (Bredenbruch et al., 2006; Diggle et al., 2007).
Consistent with the observed increase in the production of pyocyanin and rhamnolipids, we observed an increase in the concentration of PQS in the coculture supernatants (Supplementary Figure S5), which supports our hypothesis that the PQS signaling pathway is active in mixed biofilms. The finding that C. albicans increases production of PQS in mixed biofilms, even in the absence of increased AHLs, is consistent with previous reports (Cugini et al., 2010). We propose that the accumulation of PQS in the cocultures occurs because iron competition induces the expression of PrrF (a small regulatory RNA expressed under low-iron conditions), which represses gene-encoding enzymes involved in the degradation of anthranilate, a precursor of PQS (Oglesby et al., 2008). Another proposal is that in mixed biofilms C. albicans-produced farnesol stimulates PQS production in P. aeruginosa (Cugini et al., 2010).
Drug-resistance proteins and other outer membrane proteins are overexpressed in mixed biofilms
In mixed biofilms, P. aeruginosa overexpress the porin OprC, OprE, OprF, OprQ and outer membrane proteins, such as OprL, Opr86 and OpmH (Supplementary Table S1). These outer membrane proteins are key molecules that participate in the interaction between the cell and its environment. In addition, it has been demonstrated that these proteins are intrinsic participants in antibiotic resistance (Hancock and Brinkman, 2002). OprC, OprL and GroEL are necessary for biofilm formation, drug resistance and for adaptation to anaerobic conditions (Yoneyama et al., 1995). It is possible that the observed overexpression of outer membrane proteins involved in drug resistance and stress is a response to environmental changes generated by C. albicans.
P. aeruginosa shows increased mutability during the interaction with C. albicans
P. aeruginosa possesses three catalases, KatA, KatB and KatC. KatA is the catalase that shows primary activity in all growth phases. KatB and KatC are induced when cells are exposed to hydrogen peroxide and other components that generate superoxide radicals (Brown et al., 1995). Ma et al. (1999) demonstrated that, under decreased concentrations of iron, the expression and activity of KatA are strongly reduced. We observed that, in mixed biofilms, KatA was downregulated, which could be a consequence of the iron-deficient conditions. Furthermore, we observed that an alkyl hydroperoxidase (AhpC) and a putative peroxidase (PA3529) were downregulated in mixed biofilms. To validate these results, we measured the total catalase activity of P. aeruginosa in mixed and monospecific biofilms. We observed that the specific activity of these enzymes was 49.66%±15.69 s.d. lower in mixed biofilms with respect to monospecific biofilms (Supplementary Figure S6). Driffield et al. (2008) observed that the expression of KatA, AhpC and PA3529 were reduced in biofilms, and the frequency of ciprofloxacin-resistant mutants was increased in sessile cells with respect to planktonic cells. These authors suggested that the downregulation of antioxidant enzymes in P. aeruginosa biofilms may enhance the rate of mutagenic events owing to accumulation of DNA damage.
To know the effects of interspecific interaction in P. aeruginosa mutability, we used the rifampicin-resistance method (Oliver et al., 2000; Weigand and Sundin, 2012). Mutations responsible for RifR map exclusively to rpoB and produce amino-acid substitutions in three primary regions of the β subunit of RNA polymerase (Garibyan et al., 2003; Jatsenko et al., 2010). The 38 diverse mutational possibilities in rpoB available to achieve resistance include all transition and transversion substitutions, providing a suitable system for analyzing mutability. We observed a significant increase in the frequency of RifR mutants in cocultures with respect to monocultures in two different culture media (Figure 7a). To confirm that the RifR mutants have an independent genotype and arise by an increase in mutation rate, we analyzed phenotypic traits and genotype (sequencing rpoB) of 30 RifR mutant isolates of 24-h-old cocultures. Our results show that 27 mutants differ in at least one trait (Supplementary Table S4). In addition, by competition experiments between wild and mutant strain, we found that the RifR mutants do not have a growth advantage in cocultures (Supplementary Figure S7). These results demonstrate that the increased incidence of RifR mutants arise by an increase in mutation rate and not by having a growth advantage in cocultures. Consistent with these results, we observed a significant increase in the frequency of P. aeruginosa mutants resistant to streptomycin and tetracycline in cocultures (Figures 7b and c). We propose that iron deficiency and the increased production of oxidants (for example, pyocyanin) cause a decrease in total catalase activity in mixed biofilms and thereby increase oxidative stress, which, in turn, increase mutation events, enhancing the mutability of P. aeruginosa. Therefore, interespecific competition can promote the rapid emergence of antibiotic-resistant mutants among bacterial populations during infections.
Figure 7.
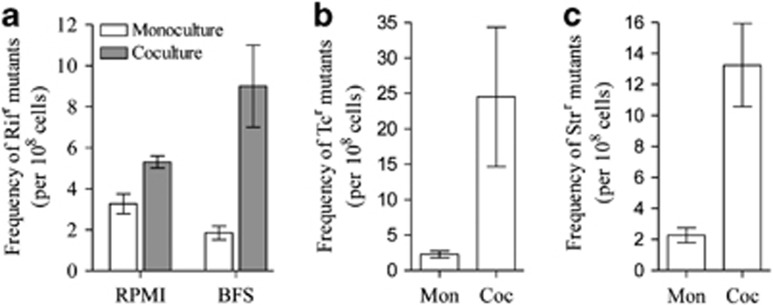
Frequency of spontaneous mutants resistant to antibiotics in P. aeruginosa cells collected from monoculture (Mon) and coculture (Coc) growth in hypoxia. (a) Frequency of mutants resistant to rifampicin (RifR), (b) tetracycline (TcR) and (c) streptomycin (StrR). Frequency of RifR mutants was estimated to bacterial population growth in RPMI medium and fetal bovine serum. Frequency of TcR and StrR mutants was estimated to bacterial population growth in RPMI. Dilutions of 24 h old cultures were spread on LB plates containing 100 μg ml−1 Rif, 20 μg ml−1 tetracycline or 20 μg ml−1 streptomycin. The plates were incubated at 37 °C, and resistant colonies were scored 48 h after plating. The dilutions were also plated on LB plates without antibiotics to determine the total number of CFUs. The mutation frequencies were approximated as the mean number of antibiotic-resistant cells divided by the total number of CFUs. Results shown represent the average of three independent experiments±s.e.m.
Ecological mechanisms maintaining biodiversity seem to be diverse themselves. We suggest that competition between species is an ecological process that can influence the evolution (Czaran et al., 2002) of microbial virulence, and consequently, it is expected that the evolutionary response (rapid evolution) alters the dynamics of microbial-microbial–host interactions (Andrade-Domínguez et al., 2014).
Differential expression of C. albicans iron-regulated proteins indicate iron competence in mixed biofilms
Consistent with our hypothesis that in mixed biofilms fungus and bacteria compete for iron, we observed in mixed biofilms, relative to the monospecific biofilms, downregulation of C. albicans iron-regulated proteins. These included proteins involved in a variety of iron-dependent processes, such as aerobic respiration (Aco1p, Idh2p), the respiratory electron transport chain (Qcr2p) heme biosynthesis (Hem13p and Hem15p) and haem-containing proteins (Cat1p). A similar iron-regulated pattern of ACO1, IDH2, HEM13 and CAT1 gene expression has been observed in C. albicans when iron is limited (Lan et al., 2004; Chen et al., 2011).
Interestingly, we found that aconitase and catalase were also downregulated in P. aeruginosa during interaction with the fungus, showing a conserved pattern in the regulation of these proteins in response to iron competition.
C. albicans Hap43p is a repressor that is induced under low-iron conditions and is essential for iron-responsive transcriptional regulation and virulence (Hsu et al., 2011). Consistent with downregulation of C. albicans iron-regulated proteins in mixed biofilms, we found that 19.71% (16) of differentially expressed proteins in mixed biofilms are Hap43p regulated (Supplementary Table S2). This suggests important roles for Hap43p transcription factor in the regulation of iron homeostasis in C. albicans during the interaction with P. aeruginosa.
C. albicans proteins associated with virulence and drug resistance are upregulated in mixed biofilms
In C. albicans, many secreted hydrolytic enzymes are potential virulence factors that are located at the cell surface before their secretion and are thought to be involved in the adhesion to/invasion of host tissues (Naglik et al., 2003). We detected a neutral arginine, alanine, leucine-specific metallo-aminopeptidase (Ape2p) that was upregulated in mixed biofilms. Klinke et al. (2008) suggest that this aminopeptidase has the potential to contribute to the pathogenicity of C. albicans. In addition, a 1,3-beta-glucosyltransferase (Bgl2p), which has a virulence role in mouse systemic infection (Sarthy et al., 1997), was also overexpressed (Supplementary Table S3). Upregulation of these proteins in mixed biofilms may have a synergistic interaction with the virulence factors of the bacterium, resulting in enhanced pathogenesis.
Alterations in glucose metabolism in C. albicans during the interaction with P. aeruginosa
Glycolytic enzymes are transcriptionally regulated in response to environmental conditions, such as oxygen levels and carbon sources and availability, and in response to cellular demands, such as metabolite concentrations and energy needs (Chambers et al., 1995). We observed that, in mixed biofilms, C. albicans overexpressed hexokinase isoenzyme 2, Hxk2p, which has been demonstrated to be induced in the presence of glucose (Askew et al., 2009). In contrast, other proteins involved in glycolysis/gluconeogenesis (Tdh3p and Pgk1p) and pyruvate metabolism (Pdc11p, Pda1p and Lpd1p) were downregulated, suggesting that glucose metabolism was altered by the interaction with P. aeruginosa (Figure 8). Consistent with these results, the glucose concentration at 24 h in cocultures was 98% (±1.2 s.d.) of the total supplied, whereas that detected in C. albicans monocultures was 1% (±0.07 s.d.) of the total supplied. Although glucose consumption was limited in the cocultures, fungal growth was still observed (Figure 5b), which suggests that C. albicans used other carbon sources for growth. In contrast, amino acids such as glutamine, aspartate and arginine were depleted in the cocultures (Supplementary Table S5). We found that supplementation with glutamine significantly increased the growth of C. albicans in mixed biofilms, but not in monospecific biofilms (Supplementary Figure S8), indicating that C. albicans used amino acids as an alternative carbon source in mixed biofilms. These results indicate that the glucose metabolism of C. albicans was affected by the interaction with P. aeruginosa (Figure 8). Because we observed that, in mixed biofilms, C. albicans was exposed to pyocyanin, we hypothesized that this metabolite secreted by the bacterium was responsible for the inhibition of glucose consumption by the fungus. To verify this hypothesis, we exposed C. albicans cells to concentrations of commercial pyocyanin similar to those detected in the mixed biofilms (10 μg ml−1) and assayed the resultant glucose consumption. We observed that pyocyanin inhibited 90% (±5 s.d.) of glucose consumption and affected the growth of C. albicans (Supplementary Figure S9). Price-whelan et al., (2007) demonstrated that pyocyanin alters the redox balance and the flow of carbon through the central metabolic routes in P. aeruginosa. These authors proposed that the superoxide radicals generated by pyocyanin can reduce the sulfhydro groups of the lipoamine cofactor from the E2 subunit of pyruvate dehydrogenase, resulting in inhibition of enzyme activity.
Figure 8.

There was a widespread repression of glycolysis, pyruvate metabolism and the TCA cycle of C. albicans in mixed biofilms. In contrast, proteins involved in the pentose phosphate pathway were upregulated. A schematic representation of the pathways is presented.
C. albicans respiration proteins are downregulated in mixed biofilms
In mixed biofilms, we observed downregulation of important proteins required for the tricarboxylic acid cycle in C. albicans (the aconitase Aco1p, isocitrate dehydrogenase Idh2 and malate dehydrogenase Mdh1p). A previous study on C. albicans demonstrated that these enzymes are repressed when cells are cultured under low-iron conditions (Lan et al., 2004). Aconitate dehydrogenase contains an Fe–S (iron–sulfur) group and shows activity that is dependent on the concentration of this metal; therefore, there is a reduction of the flow of carbon from the Krebs cycle under increased iron conditions (Shakoury-Elizeh et al., 2010). We also detected downregulation of the ubiquinol–cytochrome c–reductase Qcr2p. This protein is essential in the mitochondrial respiratory chain and is repressed under iron-deficient conditions and when there is a lack of heme groups (Dorsman and Grivell, 1990). In addition to the repression of these enzymes, it is possible that the Krebs cycles of C. albicans in mixed biofilms is affected by pyocyanin (Figure 8), which has been demonstrated to block respiration by inhibiting the activity of aconitase and affect the membrane potential of mitochondria from human cells (O'Malley et al., 2003).
Formate metabolism
We observed that the formate dehydrogenase Fdh1p was overexpressed in mixed biofilms but undetectable in C. albicans monocultures. It has been proposed that Fdh1p participates in the generation of energy through the glyoxylate cycle and beta oxidation of lipids (Prigneau et al., 2003 and Kusch et al., 2008). In plants and in bacteria, the formate pathway is induced by iron deficiency or anaerobiosis, and it has been proposed that this pathway provides a source of NADPH for the cell (Francs-Small et al., 1993). We propose that (A) the overexpression of Fdh1p observed in mixed biofilms suggests that C. albicans is obtaining energy through alternative metabolic routes, because it is unable to utilize the available glucose in the medium, and (B) the low levels of iron detected in mixed biofilms are potentially factors that induce the overexpression of Fdh1p in C. albicans.
Response of C. albicans to oxidative stress and to alterations in the redox equilibrium
The pentose phosphate pathway is of great importance for the generation of NADPH and substrates for the biosynthesis of nucleic acids and amino acids (Miosga and Zimmermann, 1996). It is also important in the protection of cells against oxidative stress, as NADPH is a cofactor of antioxidant enzymes (Minard and McAlister-Henn, 2001). We identified two proteins from the non-oxidative branch of the pentose phosphate pathway overexpressed in mixed biofilms: the transaldolase Tal1p and the transketolase Tkl1p (Supplementary Table S2 and Figure 8). Both enzymes are overexpressed under conditions of oxidative stress in C. albicans (Wang et al., 2006). As phenazines, such as pyocyanin, inhibit normal fungal respiratory activity (Morales et al., 2013), it is likely that activation of the pentose phosphate pathway contributes to maintaining the metabolic activity and redox equilibrium of C. albicans (Figure 8).
Other proteins related to oxidative stress were found to be overexpressed by C. albicans in mixed biofilms, including a chaperone from the HSP70p family, the heat-shock protein Hsp78p, alkyl hydroperoxide peroxidase Tsa1p and the putative alkyl hydroperoxidase Ahp1. In contrast, the catalase Cat1p and chaperones Kar2p, Ssa2p, Ssb1p and Ssz1p were downregulated in mixed biofilms (Supplementary Table S2). This downregulation could be caused by the reduced iron levels, as it has been demonstrated that these proteins are downregulated in low-iron conditions (Lan et al., 2004). Based on the observed reduction of the activity of the most important enzyme for detoxifying oxygen radicals and the oxidative damage generated by pyocyanin, it is expected that in mixed biofilms, the fungus undergoes oxidative stress, which may increase mutation events and thus the emergence of hypermutator cells. This hypothesis is supported by our observation of a significant increase in the frequency of amphotericin-resistant cells in mixed biofilms (Figure 9).
Figure 9.
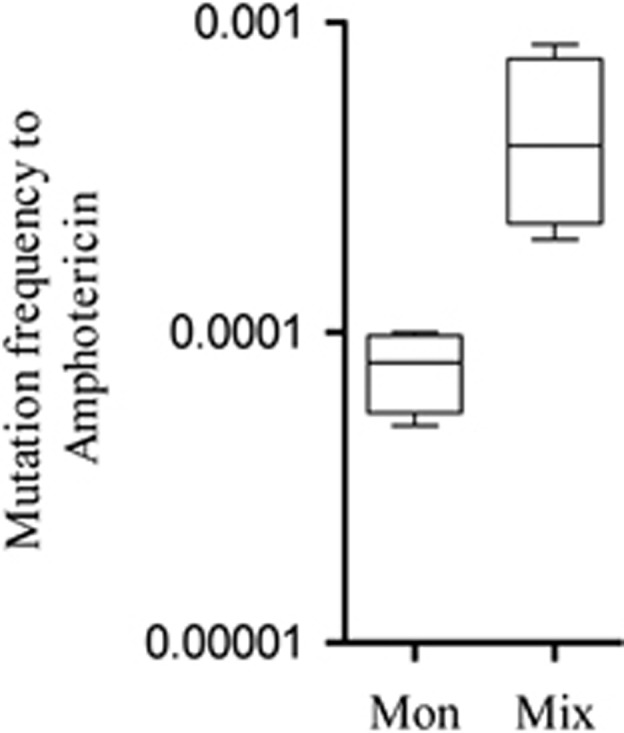
C. albicans mutation frequency following exposure to 100 μg ml−1 Amphotericin. Cells were collected from monospecific biofilms (Mon) and mixed biofilms (Mix). Mutation frequencies were expressed as the number of Amphotericin-resistant mutants recovered as a fraction of the viable count (Supplementary Materials and Methods). The panel presents the means±s.e.m. (n=4).
The P. aeruginosa–C. albicans interaction resulted in a strong synergistic effect on mouse mortality
Mixed-species infections can have consequences that differ from those associated with single-species infections, increasing the virulence of the organisms involved in the co-infection. To verify this hypothesis for interaction studied here, immunodeficient Nu/Nu nude mice were co-infected intraperitoneally with sublethal inocula of P. aeruginosa PAO1 (∼2 × 107 cells ml−1) and C. albicans CAI-4 (∼5 × 106 cells ml−1) as described in Supplementary Materials and Methods. The co-infected mice showed a high mortality rate after 12 h with respect to the groups inoculated with P. aeruginosa (2 × 107 cells ml−1) or C. albicans (∼5 × 106 cells ml−1) (Supplementary Figure S9). Doubling the dose of C. albicans (107cells ml−1) did not result in mortality of the mice after 2 weeks of observation. In contrast, doubling the dose of P. aeruginosa (4 × 107cells ml−1) caused 100% mortality within 12 h (Supplementary Figure S10). These results suggest that P. aeruginosa was the cause of death in the co-infected mice.
Roux et al. (2009) demonstrated that C. albicans impairs macrophage function and facilitates P. aeruginosa pneumonia in rats. Overall, these results suggest that C. albicans promotes the pathogenicity of P. aeruginosa in mixed infections. Previously, Roux et al. (2009) reported that the pseudomonal proteolytic enzyme elastase (LasB; also known as pseudolysin) is responsible for the increased virulence of C. albicans, but the details of this mechanism remain unclear. In a similar study using a rat pneumonia model, the same group observed that rats administered a subclinical dose of P. aeruginosa developed pneumonia only in the presence of viable C. albicans (Roux et al., 2009).
Microbes competing with unrelated or distantly related species for limited resources in the same niche activate mechanisms, such as secretion of proteins or small molecules, to attack the competing strains or species (Czaran et al., 2002; Be'er et al., 2009). Because we found that in mixed biofilms C. albicans was attacked by P. aeruginosa with mechanisms similar to those used to cause harm to their mammalian hosts, we propose that the synergism observed in coinoculated mice was caused by the increased virulence of the bacteria as a result of competition with the fungus for iron (Figure 10).
Figure 10.
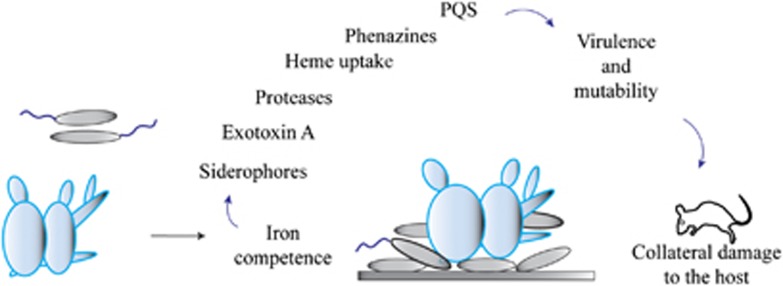
An abstract model depicting how interspecies competition triggers P. aeruginosa virulence and mutability in mixed biofilms. We propose that in mixed biofilms, the fungus and bacterium compete for iron, which triggers the expression of iron-regulated virulence factors in P. aeruginosa, for example, the siderophores pyoverdine and pyochelin, exotoxin A, proteases and heme-binding proteins. In addition, in mixed biofilms, P. aeruginosa shows increased production of the cytotoxic molecules pyocyanin, rhamnolipids and PQS. The decrease of the activity of antioxidant proteins caused by iron deficiency in both species may be the cause of the increase in the frequency of mutant cells in mixed biofilms.
Conclusions
Microbes involved in polymicrobial infections often display synergistic interactions that result in enhanced pathogenesis. However, the molecular mechanisms governing these interactions are not well understood. Our results provide compelling mechanistic evidence that interspecies competition between opportunistic pathogens can alter the course of pathogenesis in polymicrobial communities and enhance the selection of mutator cells.
Acknowledgments
We thank the Biomedical Sciences PhD Program of the Universidad Nacional Autónoma de México. AT was a recipient of a PhD Studentship from the CONACYT. We also thank Miguel Elizalde Contreras for technical assistance and Dr Veronica Domínguez-Martínez for provide some ragents for the analysis of small molecules. C. albicans strain was privide by Dr Irene Castaño-Navarro. Part of this work was supported by a DGAPA-PAPIIT Grant IN-206113.
The authors declare no conflict of interest.
Footnotes
Supplementary Information accompanies this paper on The ISME Journal website (http://www.nature.com/ismej)
Supplementary Material
References
- Andrade-Domínguez A, Salazar E, Vargas-Lagunas M, Kolter R, Encarnación S.2013Eco-evolutionary feedbacks drive species interactions ISME Je-pub ahead of print 5 December 2013 doi: 10.1038/ismej.2013.208 [DOI] [PMC free article] [PubMed]
- Askew C, Sellam A, Epp E, Hogues H, Mullick A, Nantel A, et al. Transcriptional regulation of carbohydrate metabolism in the human pathogen Candida albicans. PLoS Pathogens. 2009;5:e1000612. doi: 10.1371/journal.ppat.1000612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azghani AO, Gray LD, Johnson AR. A bacterial protease perturbs the paracellular barrier function of transporting epithelial monolayers in culture. Infect Immun. 1993;61:2681–2686. doi: 10.1128/iai.61.6.2681-2686.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bakare N, Rickerts V, Bargon J, Just-Nubling G. Prevalence of Aspergillus fumigatus and other fungal species in the sputum of adult patients with cystic fibrosis. Mycoses. 2003;46:19–23. doi: 10.1046/j.1439-0507.2003.00830.x. [DOI] [PubMed] [Google Scholar]
- Baron SS, Rowe JJ. Antibiotic action of pyocyanin. Antimicrob Agents Chemother. 1981;20:814–820. doi: 10.1128/aac.20.6.814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bauernfeind A, Bertele RM, Harms K, Horl G, Jungwirth R, Petermuller C, et al. Qualitative and quantitative microbiological analysis of sputa of 102 patients with cystic fibrosis. Infection. 1987;15:270–277. doi: 10.1007/BF01644137. [DOI] [PubMed] [Google Scholar]
- Be'er A, Zhang HP, Florin E-L, Payne SM, Ben-Jacob E, Swinney HL. Deadly competition between sibling bacterial colonies. Proc Natl Acad Sci USA. 2009;106:428–433. doi: 10.1073/pnas.0811816106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bredenbruch F, Geffers R, Nimtz M, Buer J, Häussler S. The Pseudomonas aeruginosa quinolone signal (PQS) has an iron-chelating activity. Environ Microbiol. 2006;8:1318–1329. doi: 10.1111/j.1462-2920.2006.01025.x. [DOI] [PubMed] [Google Scholar]
- Brown SM, Howell ML, Vasil ML, Anderson A, Hassett DJ. Cloning and characterization of the katB gene of Pseudomonas aeruginosa encoding a hydrogen peroxide-inducible catalase: purification of KatB, cellular localization, and demonstration that it is essential for optimal resistance to hydrogen peroxide. J Bacteriol. 1995;177:6536–6544. doi: 10.1128/jb.177.22.6536-6544.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns JL, Van Dalfsen JM, Shawar RM, Otto KL, Garber RL, Quan JM, et al. Effect of chronic intermittent administration of inhaled tobramycin on respiratory microbial flora in patients with cystic fibrosis. J Infect Dis. 1999;179:1190–1196. doi: 10.1086/314727. [DOI] [PubMed] [Google Scholar]
- Calderone RA, Fonzi WA. Virulence factors of Candida albicans. Trends Microbiol. 2001;9:327–335. doi: 10.1016/s0966-842x(01)02094-7. [DOI] [PubMed] [Google Scholar]
- Carlisle PL, Kadosh D. A genome-wide transcriptional analysis of morphology determination in Candida albicans. Mol Biol Cell. 2013;24:246–260. doi: 10.1091/mbc.E12-01-0065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers A, Packham EA, Graham IR. Control of glycolytic gene expression in the budding yeast (Saccharomyces cerevisiae) Curr Genet. 1995;29:1–9. doi: 10.1007/BF00313187. [DOI] [PubMed] [Google Scholar]
- Chen C, Pande K, French SD, Tuch BB, Noble SM. An iron homeostasis regulatory circuit with reciprocal roles in Candida albicans commensalism and pathogenesis. Cell Host Microbe. 2011;10:118–135. doi: 10.1016/j.chom.2011.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cugini C, Calfee MW, Farrow JM, Morales DK, Pesci EC, Hogan Da. Farnesol, a common sesquiterpene, inhibits PQS production in Pseudomonas aeruginosa. Mol Microbiol. 2007;65:896–906. doi: 10.1111/j.1365-2958.2007.05840.x. [DOI] [PubMed] [Google Scholar]
- Cugini C, Morales DK, Hogan Da. Candida albicans-produced farnesol stimulates Pseudomonas quinolone signal production in LasR-defective Pseudomonas aeruginosa strains. Microbiology. 2010;156:3096–3107. doi: 10.1099/mic.0.037911-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Czaran TL, Hoekstra RF, Pagie L. Chemical warfare between microbes promotes biodiversity. Proc Natl Acad Sci USA. 2002;99:786–790. doi: 10.1073/pnas.012399899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davey ME, O'toole GA. Microbial biofilms: from ecology to molecular genetics. Microbiol Molecular Biol Rev. 2000;64:847–867. doi: 10.1128/mmbr.64.4.847-867.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diggle SP, Matthijs S, Wright VJ, Fletcher MP, Chhabra SR, Lamont IL, et al. The Pseudomonas aeruginosa 4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum sensing and iron entrapment. Chem Biol. 2007;14:87–96. doi: 10.1016/j.chembiol.2006.11.014. [DOI] [PubMed] [Google Scholar]
- Diggle SP, Winzer K, Chhabra SR, Worrall KE, Cámara M, Williams P. The Pseudomonas aeruginosa quinolone signal molecule overcomes the cell density-dependency of the quorum sensing hierarchy, regulates rhl-dependent genes at the onset of stationary phase and can be produced in the absence of LasR. Mol Microbiol. 2003;50:29–43. doi: 10.1046/j.1365-2958.2003.03672.x. [DOI] [PubMed] [Google Scholar]
- Dorsman JC, Grivell LA. Expression of the gene encoding subunit II of yeast QH2: cytochrome c oxidoreductase is regulated by multiple factors. Curr Genet. 1990;17:459–464. doi: 10.1007/BF00313072. [DOI] [PubMed] [Google Scholar]
- Driffield K, Miller K, Bostock JM, O'Neill aJ, Chopra I. Increased mutability of Pseudomonas aeruginosa in biofilms. J Antimicrob Chemother. 2008;61:1053–1056. doi: 10.1093/jac/dkn044. [DOI] [PubMed] [Google Scholar]
- Duan K, Dammel C, Stein J, Rabin H, Surette MG. Modulation of Pseudomonas aeruginosa gene expression by host microflora through interspecies communication. Mol Microbiol. 2003;50:1477–1491. doi: 10.1046/j.1365-2958.2003.03803.x. [DOI] [PubMed] [Google Scholar]
- Dumitru R, Hornby JM, Nickerson KW. Defined anaerobic growth medium for studying Candida albicans basic biology and resistance to eight antifungal drugs. Antimicrob Agents Chemother. 2004;48:2350–2354. doi: 10.1128/AAC.48.7.2350-2354.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Azizi MA, Starks SE, Khardori N. Interactions of Candida albicans with other Candida spp. and bacteria in the biofilms. J Appl Microbiol. 2004;96:1067–1073. doi: 10.1111/j.1365-2672.2004.02213.x. [DOI] [PubMed] [Google Scholar]
- Encarnación S, Guzmán Y, Dunn MF, Hernández M, Vargas MC, Mora J. Proteome analysis of aerobic and fermentative metabolism in Rhizobium etli CE3. Proteomics. 2003;3:1077–1085. doi: 10.1002/pmic.200300427. [DOI] [PubMed] [Google Scholar]
- Fonzi WA, Irwin MY. Isogenic strain construction and gene mapping in Candida albicans. Genetics. 1993;134:717–728. doi: 10.1093/genetics/134.3.717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francs-Small C, Ambard-Bretteville F, Small ID, Rémy R. Identification of a major soluble protein in mitochondria from nonphotosynthetic tissues as NAD-dependent formate dehydrogenase. Plant Physiol. 1993;102:1171–1177. doi: 10.1104/pp.102.4.1171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garbeva P, Silby MW, Raaijmakers JM, Levy SB, Boer WDe. Transcriptional and antagonistic responses of Pseudomonas fluorescens Pf0-1 to phylogenetically different bacterial competitors. ISME J. 2011;5:973–985. doi: 10.1038/ismej.2010.196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garibyan L, et al. Use of the rpoB gene to determine the specificity of base substitution mutations on the Escherichia coli chromosome. DNA Repair. 2003;2:593–608. doi: 10.1016/s1568-7864(03)00024-7. [DOI] [PubMed] [Google Scholar]
- Gow NA. Germ tube growth of Candida albicans. Curr Top Med Mycol. 1997;8:43–55. [PubMed] [Google Scholar]
- Gow NA, Brown AJ, Odds FC. Fungal morphogenesis and host invasion. Curr Opin Microbiol. 2002;5:366–371. doi: 10.1016/s1369-5274(02)00338-7. [DOI] [PubMed] [Google Scholar]
- Gupta N, Haque A, Mukhopadhyay G, Narayan RP, Prasad R. Interactions between bacteria and Candida in the burn wound. Burns. 2005;31:375–378. doi: 10.1016/j.burns.2004.11.012. [DOI] [PubMed] [Google Scholar]
- Hancock REW, Brinkman FSL. Function of pseudomonas porins in uptake and efflux. Annu Rev Microbiol. 2002;56:17–38. doi: 10.1146/annurev.micro.56.012302.160310. [DOI] [PubMed] [Google Scholar]
- Hansen SK, Rainey PB, Haagensen JAJ, Molin S. Evolution of species interactions in a biofilm community. Nature. 2007;445:533–536. doi: 10.1038/nature05514. [DOI] [PubMed] [Google Scholar]
- Harrison F, Paul J, Massey RC, Buckling A. Interspecific competition and siderophore-mediated cooperation in Pseudomonas aeruginosa. ISME J. 2008;2:49–55. doi: 10.1038/ismej.2007.96. [DOI] [PubMed] [Google Scholar]
- Hogan DA, Kolter R. Pseudomonas-Candida interactions: an ecological role for virulence factors. Science. 2002;296:2229–2232. doi: 10.1126/science.1070784. [DOI] [PubMed] [Google Scholar]
- Hogan DA, Vik A, Kolter R. A Pseudomonas aeruginosa quorum-sensing molecule influences Candida albicans morphology. Mol Microbiol. 2004;54:1212–1223. doi: 10.1111/j.1365-2958.2004.04349.x. [DOI] [PubMed] [Google Scholar]
- Holloway BW. Genetic recombination in Pseudomonas aeruginosa. J Gen Microbiol. 1955;13:572–581. doi: 10.1099/00221287-13-3-572. [DOI] [PubMed] [Google Scholar]
- Hsu P, Yang C, Lan C. Candida albicans Hap43 is a repressor induced under low-iron conditions and is essential for iron-responsive transcriptional regulation and virulence. Eukaryot Cell. 2011;10:207–225. doi: 10.1128/EC.00158-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hube B. Infection-associated genes of Candida albicans. Future Microbiol. 2006;1:209–218. doi: 10.2217/17460913.1.2.209. [DOI] [PubMed] [Google Scholar]
- Hughes WT, Kim HK. Mycoflora in cystic fibrosis: some ecologic aspects of Pseudomonas aeruginosa and Candida albicans. Mycopathol Mycol Appl. 1973;50:261–269. doi: 10.1007/BF02053377. [DOI] [PubMed] [Google Scholar]
- Hurkman WJ, Tanaka CK. Solubilization of plant membrane proteins for analysis by two-dimensional gel electrophoresis. Plant Physiol. 1986;81:802–806. doi: 10.1104/pp.81.3.802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iglewski BH, Kabat D. NAD-dependent inhibition of protein synthesis by Pseudomonas aeruginosa toxin. Proc Natl Acad Sci USA. 1975;72:2284–2288. doi: 10.1073/pnas.72.6.2284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jatsenko T, Tover A, Tegova R, Kivisaar M. Molecular characterization of Rif(r) mutations in Pseudomonas aeruginosa and Pseudomonas putida. Mutat Res Fund Mol Mech Mut. 2010;683:106–114. doi: 10.1016/j.mrfmmm.2009.10.015. [DOI] [PubMed] [Google Scholar]
- Kaleli I, Cevahir N, Demir M, Yildirim U, Sahin R. Anticandidal activity of Pseudomonas aeruginosa strains isolated from clinical specimens. Mycoses. 2007;50:74–78. doi: 10.1111/j.1439-0507.2006.01322.x. [DOI] [PubMed] [Google Scholar]
- Kerr J. Inhibition of fungal growth by Pseudomonas aeruginosa and Pseudomonas cepacia isolated from patients with cystic fibrosis. J Infect. 1994;28:305–310. doi: 10.1016/s0163-4453(94)91943-7. [DOI] [PubMed] [Google Scholar]
- Kirienko NV, Kirienko DR, Larkins-Ford J, Wählby C, Ruvkun G, Ausubel FM. Pseudomonas aeruginosa disrupts Caenorhabditis elegans iron homeostasis, causing a hypoxic response and death. Cell Host Microbe. 2013;13:406–416. doi: 10.1016/j.chom.2013.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klinke T, Rump A, Pönisch R, Schellenberger W, Müller E-C, Otto A, et al. Identification and characterization of CaApe2-a neutral arginine/alanine/leucine-specific metallo-aminopeptidase from Candida albicans. FEMS Yeast Res. 2008;8:858–869. doi: 10.1111/j.1567-1364.2008.00411.x. [DOI] [PubMed] [Google Scholar]
- Kusch H, Engelmann S, Bode R, Albrecht D, Morschhäuser J, Hecker M. A proteomic view of Candida albicans yeast cell metabolism in exponential and stationary growth phases. Int J Med Microbiol. 2008;298:291–318. doi: 10.1016/j.ijmm.2007.03.020. [DOI] [PubMed] [Google Scholar]
- Lamont IL, Beare PA, Ochsner U, Vasil AI, Vasil ML. Siderophore-mediated signaling regulates virulence factor production in Pseudomonas aeruginosa. Proc Natl Acad Sci USA. 2002;99:7072–7077. doi: 10.1073/pnas.092016999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lan C-Y, Rodarte G, Murillo LA, Jones T, Davis RW, Dungan J, et al. Regulatory networks affected by iron availability in Candida albicans. Mol Microbiol. 2004;53:1451–1469. doi: 10.1111/j.1365-2958.2004.04214.x. [DOI] [PubMed] [Google Scholar]
- Lau GW, Hassett DJ, Ran H, Kong F. The role of pyocyanin in Pseudomonas aeruginosa infection. Trends Mol Med. 2004;10:599–606. doi: 10.1016/j.molmed.2004.10.002. [DOI] [PubMed] [Google Scholar]
- Liu H. Co-regulation of pathogenesis with dimorphism and phenotypic switching in Candida albicans, a commensal and a pathogen. Int J Med Microbiol. 2002;292:299–311. doi: 10.1078/1438-4221-00215. [DOI] [PubMed] [Google Scholar]
- Loper JE, Buyer JS. Siderophores in microbial interactions on plant surfaces. Mol Plant Microbe Interact. 1991;4:5–13. [Google Scholar]
- Lynch AS, Robertson GT. Bacterial and fungal biofilm infections. Annu Rev Med. 2008;59:415–428. doi: 10.1146/annurev.med.59.110106.132000. [DOI] [PubMed] [Google Scholar]
- Ma J, Ochsner UA, Klotz MG, Vagira K, Howell ML, Johnson Z, et al. Bacterioferritin A modulates catalase A (KatA) activity and resistance to hydrogen peroxide in Pseudomonas aeruginosa. J Bacteriol. 1999;181:3730–3742. doi: 10.1128/jb.181.12.3730-3742.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marquart ME, Caballero AR, Chomnawang M, Thibodeaux Ba, Twining SS, O'Callaghan RJ. Identification of a novel secreted protease from Pseudomonas aeruginosa that causes corneal erosions. Invest Ophthalmol Vis Sci. 2005;46:3761–3768. doi: 10.1167/iovs.04-1483. [DOI] [PubMed] [Google Scholar]
- Mavrodi DV, Blankenfeldt W, Thomashow LS. Phenazine compounds in fluorescent Pseudomonas spp. biosynthesis and regulation. Annu Rev Phytopathol. 2006;44:417–445. doi: 10.1146/annurev.phyto.44.013106.145710. [DOI] [PubMed] [Google Scholar]
- Minard KI, McAlister-Henn L. Antioxidant function of cytosolic sources of NADPH in yeast. Free Radic Biol Med. 2001;31:832–843. doi: 10.1016/s0891-5849(01)00666-9. [DOI] [PubMed] [Google Scholar]
- Miosga T, Zimmermann FK. Cloning and characterization of the first two genes of the non-oxidative part of the Saccharomyces cerevisiae pentose-phosphate pathway. Curr Genet. 1996;30:404–409. doi: 10.1007/s002940050149. [DOI] [PubMed] [Google Scholar]
- Morales DK, Grahl N, Okegbe C. Control of Candida albicans metabolism and biofilm formation by Pseudomonas aeruginosa phenazines. M Bio. 2013;4:e00526-12. doi: 10.1128/mBio.00526-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morales DK, Jacobs NJ, Rajamani S, Krishnamurthy M, Cubillos-Ruiz JR, Hogan DA. Antifungal mechanisms by which a novel Pseudomonas aeruginosa phenazine toxin kills Candida albicans in biofilms. Mol Microbiol. 2010;78:1379–1392. doi: 10.1111/j.1365-2958.2010.07414.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadal Jimenez P, Koch G, Papaioannou E, Wahjudi M, Krzeslak J, Coenye T, et al. Role of PvdQ in Pseudomonas aeruginosa virulence under iron-limiting conditions. Microbiology. 2010;156:49–59. doi: 10.1099/mic.0.030973-0. [DOI] [PubMed] [Google Scholar]
- Naglik JR, Challacombe SJ, Naglik JR, Challacombe SJ, Hube B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiol Mol Biol Rev. 2003;67:400–428. doi: 10.1128/MMBR.67.3.400-428.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naglik J, Albrecht A, Bader O, Hube B. Candida albicans proteinases and host/pathogen interactions. Cell Microbiol. 2004;6:915–926. doi: 10.1111/j.1462-5822.2004.00439.x. [DOI] [PubMed] [Google Scholar]
- Neely AN, Law EJ, Holder IA. Increased susceptibility to lethal Candida infections in burned mice preinfected with Pseudomonas aeruginosa or pretreated with proteolytic enzymes. Infect Immun. 1986;52:200–204. doi: 10.1128/iai.52.1.200-204.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Malley YQ, Abdalla MY, McCormick ML, Reszka KJ, Denning GM, Britigan BE. Subcellular localization of Pseudomonas pyocyanin cytotoxicity in human lung epithelial cells. Am J Physiol Lung Cell Mol Physiol. 2003;284:L420–L430. doi: 10.1152/ajplung.00316.2002. [DOI] [PubMed] [Google Scholar]
- Ochsner UA, Johnson Z, Vasil ML. Genetics and regulation of two distinct haem-uptake systems, phu and has, in Pseudomonas aeruginosa. Microbiology. 2000;146:185–198. doi: 10.1099/00221287-146-1-185. [DOI] [PubMed] [Google Scholar]
- Oglesby AG, Farrow JM, Lee J-H, Tomaras AP, Greenberg EP, Pesci EC, et al. The influence of iron on Pseudomonas aeruginosa physiology: a regulatory link between iron and quorum sensing. J Biol Chem. 2008;283:15558–15567. doi: 10.1074/jbc.M707840200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oliver A, Canton R, Campo P, Baquero F, Blazquez J. High frequency of hypermutable Pseudomonas aeruginosa in cystic fibrosis lung infection. Science. 2000;288:1251–1253. doi: 10.1126/science.288.5469.1251. [DOI] [PubMed] [Google Scholar]
- Peleg AY, Hogan DA, Mylonakis E. Medically important bacterial-fungal interactions. Nat Rev Microbiol. 2010;8:340–349. doi: 10.1038/nrmicro2313. [DOI] [PubMed] [Google Scholar]
- Pesci EC, Milbank JBJ, Pearson JP, McKnight S, Kende AS, Greenberg EP, et al. Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc Natl Acad Sci USA. 1999;96:11229–11234. doi: 10.1073/pnas.96.20.11229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfaller MA, Diekema DJ. Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev. 2007;20:133–163. doi: 10.1128/CMR.00029-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price-whelan A, Dietrich LEP, Newman DK. Pyocyanin alters redox homeostasis and carbon flux through central metabolic pathways in Pseudomonas aeruginosa PA14. J Bacteriol. 2007;189:6372–6381. doi: 10.1128/JB.00505-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prigneau O, Porta A, Poudrier JA, Colonna-Romano S, Noël T, Maresca B. Genes involved in beta-oxidation, energy metabolism and glyoxylate cycle are induced by Candida albicans during macrophage infection. Yeast. 2003;20:723–730. doi: 10.1002/yea.998. [DOI] [PubMed] [Google Scholar]
- Purschke FG, Hiller E, Trick I, Rupp S. Flexible survival strategies of Pseudomonas aeruginosa in biofilms result in increased fitness compared with Candida albicans. Mol Cell Proteomics. 2012;11:1652–1669. doi: 10.1074/mcp.M112.017673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ran H, Hassett DJ, Lau GW. Human targets of Pseudomonas aeruginosa pyocyanin. Proc Natl Acad Sci USA. 2003;100:14315–14320. doi: 10.1073/pnas.2332354100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratledge C, Dover LG. Metabolism in pathogenic bacteria. Annu Rev Microbiol. 2000;54:881–941. doi: 10.1146/annurev.micro.54.1.881. [DOI] [PubMed] [Google Scholar]
- Roux D, Gaudry S, Dreyfuss D, El-Benna J, De Prost N, Denamur E, et al. Candida albicans impairs macrophage function and facilitates Pseudomonas aeruginosa pneumonia in rat. Crit Care Med. 2009;37:1062–1067. doi: 10.1097/CCM.0b013e31819629d2. [DOI] [PubMed] [Google Scholar]
- Sarthy AV, Mcgonigal T, Coen M, Frost DJ, Meulbroek JA, Goldman RC, et al. Phenotype in Candida albicans of a disruption of the BGL2 gene encoding a 1,3-beta-glucosyltransferase. Microbiology. 1997;143 (Pt 2:367–376. doi: 10.1099/00221287-143-2-367. [DOI] [PubMed] [Google Scholar]
- Shakoury-Elizeh M, et al. Metabolic response to iron deficiency in Saccharomyces cerevisiae. J Biol Chem. 2010;285:14823–14833. doi: 10.1074/jbc.M109.091710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shakoury-Elizeh M, Protchenko O, Berger A, Cox J, Gable K, Dunn TM, et al. Metabolic response to iron deficiency in Saccharomyces cerevisiae. J Biol Chem. 2010;285:14823–14833. doi: 10.1074/jbc.M109.091710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sibley CD, Duan K, Fischer C, Parkins MD, Storey DG, Rabin HR, et al. Discerning the complexity of community interactions using a Drosophila model of polymicrobial infections. PLoS Pathogens. 2008;4:e1000184. doi: 10.1371/journal.ppat.1000184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sio CF, Otten LG, Cool RH, Stephen P, Braun PG, Bos R, et al. Quorum quenching by an N-acyl-homoserine lactone acylase from Pseudomonas quorum quenching by an N-acyl-homoserine lactone acylase from Pseudomonas aeruginosa PAO1. Infect Immun. 2006;74:1673–1682. doi: 10.1128/IAI.74.3.1673-1682.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Synnott JM, Guida A, Mulhern-Haughey S, Higgins DG, Butler G. Regulation of the hypoxic response in Candida albicans. Eukaryot Cell. 2010;9:1734–1746. doi: 10.1128/EC.00159-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smukalla S, Caldara M, Pochet N, Beauvais A, Guadagnini S, Yan C, et al. FLO1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast. Cell. 2008;135:726–737. doi: 10.1016/j.cell.2008.09.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takase H, Nitanai H, Hoshino K, Otani T. Impact of siderophore production on Pseudomonas aeruginosa infections in immunosuppressed mice. Infect Immun. 2000;68:1834–1839. doi: 10.1128/iai.68.4.1834-1839.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tyson GW, Chapman J, Hugenholtz P, Allen EE, Ram RJ, Richardson PM, et al. Community structure and metabolism through reconstruction of microbial genomes from the environment. Nature. 2004;428:37–43. doi: 10.1038/nature02340. [DOI] [PubMed] [Google Scholar]
- Wang Y, Cao YY, Jia XM, Cao YB, Gao PH, Fu XP, et al. Cap1p is involved in multiple pathways of oxidative stress response in Candida albicans. Free Radic Biol Med. 2006;40:1201–1209. doi: 10.1016/j.freeradbiomed.2005.11.019. [DOI] [PubMed] [Google Scholar]
- Weigand MR, Sundin GW. General and inducible hypermutation facilitate parallel adaptation in Pseudomonas aeruginosa despite divergent mutation spectra. Proc Natl Acad Sci USA. 2012;109:13680–13685. doi: 10.1073/pnas.1205357109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whiteway M, Oberholzer U. Candida morphogenesis and host-pathogen interactions. Curr Opin Microbiol. 2004;7:350–357. doi: 10.1016/j.mib.2004.06.005. [DOI] [PubMed] [Google Scholar]
- Wilderman PJ, Vasil AI, Johnson Z, Wilson MJ, Cunliffe HE, Lamont IL, et al. Characterization of an endoprotease (PrpL) encoded by a PvdS-regulated gene in Pseudomonas aeruginosa. Infect Immun. 2001;69:5385–5394. doi: 10.1128/IAI.69.9.5385-5394.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoneyama H, Yamano Y, Nakae T. Role of porins in the antibiotic susceptibility of Pseudomonas aeruginosa: construction of mutants with deletions in the multiple porin genes. Biochem Biophys Res Commun. 1995;213:88–95. doi: 10.1006/bbrc.1995.2102. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.