

Abstract
Spectral and temporal features of human infant crying may detect neurobehavioral effects of prenatal cocaine exposure (PCE). Finding comparable measures of rodent ultrasonic vocalizations (USVs) would promote translational analyses by controlling the effects of correlated variables that confound human studies. To this end, two studies examined the sensitivity of similar acoustic structures in human infant and rat pup vocalizations to effects of PCE. In Study 1, cry sounds of 107 one month-old infants were spectrum analyzed to create a novel set of measures and to detect the presence of hyperphonation - a qualitative shift to an atypically high fundamental frequency (basic pitch) associated with neurobehavioral insult. Infants with PCE were compared to infants with prenatal polydrug-exposure (PPE) without cocaine and with infants in a standard comparison (SC) group with no prenatal drug exposure. In Study 2, USVs of 118 five day-old rat pups with either PCE, prenatal saline exposure or no prenatal exposures were spectrum analyzed to detect the presence of frequency shifts – acoustic features that have a frequency waveform similar to that of hyperphonation. Results of study 1 showed PCE had two sets of sex-dependent effects on human infants: PCE males had higher pitched cries with more dysphonation (turbulence); PCE females had longer pauses between fewer cry sounds that were of lower amplitude than comparison groups. PCE and PPE infants had more cries with hyperphonation than SC infants. In study 2, PCE pups had a greater percentage of USVs with shift in the acoustic structure than pups in the two control groups. As such, the novel measures of human infant crying and rat pup USVs were sensitive to effects of PCE. These studies provide the first known translational analysis of similar acoustic structures of vocalizations in two species to detect adverse effects of prenatal drug exposure.
Introduction
Maternal cocaine-use during pregnancy continues to be a significant public health concern [1]–[3] with subtle effects [4] having far-ranging implications for the health and development of exposed children [5]–[10]. Prenatal cocaine exposure (PCE) has adverse neurochemical, vasoconstrictive and dysregulatory effects [11] that may be evident in early neurobehavioral deficits [12]–[18] and problematic mother-infant interactions that may further contribute to poor developmental outcomes [19]–[22]. The independent role of PCE in the development of these neurobehavioral deficits, however, is less than clear due to the many confounding conditions, such as poverty and prenatal polydrug exposure (PPE), that are often a part of the larger context of maternal cocaine-use during pregnancy [23], [24]. Translational analyses of the effects of PCE that provide experimental control over such confounding conditions would be facilitated by finding sensitive measures of neurobehavioral function that are comparable across human and other species. The purpose of this paper is to examine the utility of a novel set of spectral characteristics of human infant cry sounds and measures of rat pup ultrasonic vocalizations (USVs) in a translational analysis of the effects of PCE on early neurobehavioral development.
Spectrum analysis of the acoustic and temporal features of human infant cry sounds has long been used to detect adverse effects on neurobehavioral integrity of a wide range of prenatal and perinatal conditions, extending from cases of obvious brain damage and genetic disorders [25], [26] to prenatal exposures to opiates [27], marijuana [28], tobacco [29], and alcohol [30]. Cries of infants with these and other nonoptimal pre- and perinatal conditions typically include a higher fundamental frequency (F0 or basic pitch), greater amounts of dysphonation (sonic turbulence), shorter initial expiratory sounds and variations in several other measures of the power spectrum. A hallmark of these distinctive cry sounds is the frequent presence of hyperphonation - a sudden, qualitative shift to a high-pitched acoustic structure with an F0 over 1000 Hz – that occurs more often in response to physically intense eliciting conditions [31]. Analyses of up to the first three cry expirations following a painful stimulus have shown these acoustic features to differentiate cry sounds of infants with PCE [12], [32], [33], but effects on specific measures have been inconsistent across studies and differences between PCE and comparison infants may no longer be evident when statistical analyses are adjusted for polydrug covariates [12]. Little is known, however, about the value of an analysis of cry sounds beyond the initial three cry expirations. For the present study, we developed a novel set of spectral and temporal measures designed to capture the presence of hyperphonation and dynamic characteristics in an extended period of infant crying.
Spectrum analyses of rodent USVs have also shown that variations in such measures as amplitude, F0, number, and repetition rate may reflect adverse effects of prenatal and postnatal drug exposures on early neurobehavioral development [34], [35]. Genotypic-dependent effects of PCE on the number and beginning pitch of USVs of infant mice have been described [36], but no effects of PCE have been found in rodent pup USVs using measures directly comparable to those used in studies of human infant cry sounds [37]. In contrast to measuring variations in specific acoustic features, another approach has been to examine categories of USV waveforms based on the pioneering taxonomy described by Brudzynski and colleagues [38]. Neonatal exposure to alcohol, for example, has been shown to selectively reduce the presence of specific waveform categories in mouse USVs [39], but no effects of cocaine have been found using this methodology. Interestingly, a rarely noted USV waveform, not included in the above taxonomy, has an acoustic structure similar to that of the hyperphonation found in human infant cry sounds. Described in the USVs of rat pups and other rodent species as steps or shifts [40], [41], this waveform is characterized by a qualitative shift in F0, in the absence of a vertical line in the spectrogram within the same USV. Anecdotal reports suggest that, like hyperphonation in human infant cries, steps or shifts occur more often in physically intense eliciting conditions [42] and may also reflect effects of drug exposure on neurobehavioral regulation [43]. Parallels between the acoustic structures of hyperphonation in human infant crying and shift in rat pup USVs may provide the basis for a translational analysis of vocalizations in the study of effects of PCE on neurobehavioral integrity. In the present study we systematically explored the presence of frequency shifts in the vocalizations of rat pups under intense thermal conditions and whether this acoustic waveform was differentially represented in the USVs of rat pups with PCE.
Methods
Study 1: Human infant crying
Subjects
Pregnant women and mothers of infants younger than 4 weeks of age were recruited via community advertisements, drug rehabilitation centers and social services as part of a larger study of the neurobehavioral and social effects of prenatal cocaine exposure. All human procedures were approved by the University of North Carolina's Office of Human Research Ethics Biomedical Institutional Review Board (IRB). An IRB-approved written Parental Permission for a Minor Child to Participate in a Research Study (Biomedical) was obtained for infant participation, and was signed by each infant's biological mother prior to testing. A signed written consent form approved by the University of North Carolina's Biomedical Institutional Review Board was also obtained from all mothers in order to collect maternal and prenatal characteristics prior to testing. Infants of these mothers (n = 131) participated in a cry analysis paradigm at approximately one month of age (days: M = 27.3, SD = 13.3) and were assigned to one of three groups for comparison. PCE infants (n = 31) had mothers who were currently involved in drug-treatment services, had self-report of cocaine-use during pregnancy on a Time Line Follow Back (TLFB) questionnaire, and/or had prenatal urine samples positive for cocaine use at the level of 0.300 ng/ml for cocaine and metabolites. PCE infants were also exposed prenatally to maternal use of antidepressants, alcohol, cigarettes, marijuana, and opiates during pregnancy. A standard comparison group (SC) was comprised of infants (n = 50) whose mothers did not use antidepressants and had a negative urine toxicology for cocaine, marijuana, methamphetamine, or opiates at laboratory visit, did not endorse drug-use on a phone screen, and/or did not test positive for drug use. A novel, second comparison group was comprised of infants (n = 38) with prenatal polydrug-exposure (PPE) if their mothers used tobacco, alcohol, marijuana, opiates and/or antidepressants in varying degrees, but not cocaine. Chi square analyses showed that PCE and PPE infants did not reliably differ in the distributions of prenatal exposure to antidepressants, alcohol, cigarettes, marijuana or opiates (all p's>.25).
Table 1 shows maternal demographic and infant growth characteristics of this sample. Compared to mothers in both the PCE and PPE groups, mothers in the SC group were disproportionately Caucasian, as compared to African-American [X 2(2) = 8.07, p<.018], married [X 2(6) = 33.5, p<.001], and had a higher level of attained education [X 2(10) = 51.97, p<.001]. No group differences were found in methods of delivery (Vaginal vs. C-section) [X 2(2) = 3.85, p<.15]. One-way analyses of variance showed that the three groups of infants differed in birth weight [F(2,128) = 10.91, p<.001] and gestational age [F(2,128) = 6.63, p<.002]. LSD post-hoc tests showed that PCE infants had a lower birth weight and gestational age than infants in the two control groups. PPE and SC infant groups did not differ on these measures (all p's>.20). Infant birthweight was significantly correlated with gestational age (p<.001), maternal education (p<.001) and maternal ethnicity (p<.05). Infant groups also did not differ in post-conception age at the time cries were recorded [F(2,129) = .198, p<.82], or in the distribution of males and females (PCE: males = 20, females = 19; PPE: males = 20, females = 22; SC: males = 30, females = 20) [X 2(2) = 1.51, p<.48].
Table 1. Human Subject Demographics.
| Demographic | Comparison | Cocaine | Polydrug | |
| Number of subjects | 38 | 31 | 38 | |
| Maternal Ethnicity | Caucasian | 34 (68%) | 25 (64.1%) | 17 (40.5%) |
| African-American | 12 (24%) | 10 (25.6%) | 20 (47.6%) | |
| Hispanic | 1 (2%) | 2 (5.1%) | 20 (7.1%) | |
| Asian | 2 (4%) | 0 (0%) | 1 (2.4%) | |
| Other | 1 (2%) | 2 (5.1%) | 1 (2.4%) | |
| Maternal Marital Status | Divorced | 1 (2.3%) | 6 (17.2%) | 4 (11.8%) |
| Married | 24 (55.8%) | 1 (2.8%) | 5 (14.7%) | |
| Never Married | 17 (39.6%) | 24 (68.6%) | 21 (61.7%) | |
| Separated | 1 (2.3%) | 4 (11.4%) | 4 (11.8%) | |
| Maternal Education | ≤ High School | 10 (22.7%) | 16 (44.4%) | 13 (39.5%) |
| Some College/Trade School | 6 (13.7%) | 20 (55.6%) | 14 (42.4%) | |
| College Graduate | 14 (31.8%) | 0 (0%) | 4 (12.1%) | |
| Post Graduate Education | 14 (31.8%) | 0 (0%) | 2 (6.0%) | |
| Delivery Method | Vaginal | 40 (80%) | 33 (87%) | 29 (69%) |
| C-Section | 10 (20%) | 5 (13%) | 13 (31%) | |
| Infant Birth Weight (g) | 3506.8±52.4 | 3081.5±67.4 | 3388.9±78.2 | |
| Gestational Age (weeks) | 39.9±0.2 | 38.9±0.3 | 39.7±0.2 | |
| Age at Assessment (days) | 24.5±1.0 | 30.2±2.1 | 25.6±1.1 | |
Note: Values are raw numbers, with percentages in parentheses, and Mean ± SEM where appropriate.
Cry recording and analysis
Crying was elicited by placing infants on a cold metal scale maintained at 20°C and recorded using an Olympus DM-20 digital recorder (44.1 kHz sampling rate) held 20 centimeters vertically and mid-sternum horizontally from the infant's mouth. The initial 30 s of the crying bout was downloaded to a Multi-Speech Lab (Kay/Pentax) computer software program and down-sampled to 22,050 Hz for greater resolution (±21 Hz) of frequencies up to 11 kHz. Digital spectrograms were produced for each crying bout and examined by a research assistant blinded to infant group and sex. Twenty-four infants did not cry (PCE n = 8, SC n = 12, PPE n = 4), thus resulting in a total of 107 infants whose cries were analyzed (PCE n = 31: males = 15, females = 16; PPE n = 38: males = 18, females = 20; SC n = 38: males = 26, females = 12).
As described in previous work, measures of crying were derived from the temporal and spectral characteristics of each expiratory cry sound produced during the entire 30 s recording period [44]. Peak F0 (in Hz), and its amplitude (F0 Amp) in decibels (dB), were determined from the power spectrum of a Fast-Fourier Transform (FFT) conducted on the 25 ms sample at which F0 reached its highest point in each expiratory cry sound. The multiple assessments of Peak F0 and F0 Amp were summed and averaged for an overall mean value. F0SD was calculated as the standard deviation of the multiple assessments of Peak F0. Initial F0 was the highest Peak F0 found in any one of the first three cry expirations. Durations of temporal measures (seconds) were determined via cursor placement on the digital spectrogram and averaged across all cry expirations. Inter-Cry-Interval (ICI) was the mean duration from offset of one cry expiration to onset of the next; Expiration Duration was the mean duration from onset to offset of all cry expirations. Dysphonation was the mean amount of sonic turbulence in all cry expirations, as determined on a 0–4 point scale used in previous studies [45], averaged across all cry expirations. Table 2 shows the means, standard errors and results of statistical analyses for each of these measures, while Figure 1 shows an example of infant crying with hyperphonation in the latter segments of the crying bout.
Table 2. Spectral and Temporal Analysis of Human Infant Cry Sounds.
| Measure | Sex | Comparison | Cocaine | Polydrug |
| Initial F0 (Hz) | Male | 697.3±86.7 | 1010.0±115.3 | 851.9±102.3 |
| Female | 619.5±125.4 | 672.8±116.3 | 884.1±96.4 | |
| Peak F0 (Hz) | Male | 728.9±85.5 | 1204.9±113.7 | 952.6±100.9 |
| Female | 663.8±123.7 | 684.8±114.7 | 926.1±95.1 | |
| F0 SD (Hz) † | Male | 104.8±27.8 | 235.5±37.0 | 153.4±32.8 |
| Female | 70.0±40.2 | 81.0±37.3 | 125.2±30.9 | |
| F0 Amp (dB) | Male | 59.3±1.6 | 61.6±2.1 | 58.3±1.9 |
| Female | 58.1±2.3 | 51.2±2.2 | 58.9±1.8 | |
| Inter-Cry-Interval (s) | Male | 1.4±0.8 | 1.3±1.1 | 1.9±1.0 |
| Female | 1.1±1.2 | 5.9±1.1 | 2.5±0.9 | |
| Cry Expiration Duration (s) | Male | 1.3±0.1 | 1.2±0.1 | 1.4±0.1 |
| Female | 1.6±0.2 | 1.4±0.1 | 1.2±0.1 | |
| Number of Cry Expirations † | Male | 14.8±1.5 | 13.5±1.9 | 12.0±1.7 |
| Female | 12.4±2.1 | 7.9±2.0 | 13.7±1.6 | |
| Dysphonation | Male | 2.2±0.2 | 2.7±0.2 | 2.6±0.2 |
| Female | 2.6±0.2 | 2.1±0.2 | 2.1±0.2 |
Note: All values are Mean ± SEM. † denotes analyses conducted on log transformations.
Figure 1. Spectrographic Display of Human Infant Crying.

This figure shows the standard phonated harmonic structure of crying evident in the first 3 expiratory cry sounds, followed by expiratory cry sounds with hyperphonation, evident at the end of the fourth cry expiration and in cry expirations that occurred after 5 sec. A greater vertical distance between harmonic frequencies denoted by horizontal lines characterizes this acoustic structure. This figure is an example of finding the highest Peak F0 in the crying bout beyond the initial three cry expirations often used in previous studies of cry analysis.
Statistical analysis
A 3 (Exposure Group) ×2 (Sex) General Linear Model (GLM) analysis of variance (ANOVA) was conducted on each measure of infant crying using p<.05 for statistical significance. Log10 transforms were conducted on measures requiring normalization of their distributions, as denoted in Table 2. Analyses of covariance were conducted using birthweight as the covariate due to its biologically proximal and plausible role in neurobehavioral development and its significant correlation and covariance with the other demographic measures differentiating PCE infants. These analyses showed no reliable effects of birthweight on any cry measures, nor were any differences found in the pattern of results. As such, the presented analyses are based on the unadjusted data. LSD and t-test post-hoc comparisons (with alpha set at 0.05) were used to detect differences between cells in main-effects and interaction-effects, respectively. Chi Square analyses were used for all nonparametric analyses. Analyses were conducted using SPSS statistical software.
Study 2: Rat pup ultrasonic vocalizations
Subjects
Sprague-Dawley rat dams received twice-daily subcutaneous injections of 15 mg/kg cocaine hydrochloride (Sigma, St. Louis, MO) in normal saline (PCE group, n = 34) at approximately 9:00 AM and again at 4:00 PM throughout gestation (GD 1–20) and not thereafter. Saline-treated (SC group, n = 34) dams received twice-daily subcutaneous injections of 0.9% Saline solution (1 ml/kg/injection), and were food-yoked to cocaine-treated dams such that they were only given access to the amount of food consumed by cocaine dams each day. Pups in an untreated control group (UN group, n = 50) received no treatment. At birth, litters were culled to 8 pups per dam, and one male and one female pup were randomly selected for USV recording on postnatal day 5, resulting in a total of 118 pups. Pups were given a series of developmentally appropriate thermal isolation-challenges to induce vocalizations. In a temperature-controlled apparatus, pups were subjected to a 60-min period of habituation at 35.0°C, after which the apparatus temperature was reduced to 28°C (a moderate temperature challenge) for another 60-min. The temperature was then reduced to 21°C (a severe temperature challenge) for a final 60 min. USVs that occurred during two five-minute periods in the final hour were assessed: one after the change to a final temperature was stabilized (minutes 15–20) and one at the end of the hour (minutes 55–60). All animal procedures were approved by the University of North Carolina's Institutional Animal Care and Use Committee (NIH/PHS Animal Welfare Assurance Number: A3410-01) and conducted under federal guidelines for humane treatment of laboratory subjects.
Recording and acoustic analyses
Rat pup USVs were recorded and analyzed using previously described methods [44]. Ultrasonic recording equipment included model CM16/CMPA40-5V microphones (Avisoft Bioacoustics; Berlin, Germany) connected to a desktop computer through a National Instruments instrumentation recorder (PCI-6132) with microphones mounted 2cm above the subject platform. National Instruments software (LabView 2009) began acquisition of ultrasonic vocalizations at the session start and terminated at the session end. Digital audio files were down-sampled to 200,000 Hz from 1,000,000 Hz and viewed in a 3-second moving window in a digital spectrogram produced by Adobe Audition.
A research assistant, trained to criterion and blinded to exposure group and sex of the rat pup, examined all USVs emitted for the presence of a frequency shift. Shift usually occurred in USVs averaging 30 kHz–50 kHz and was defined as an “instantaneous and sudden qualitative shift change(s) in frequency in the absence of a vertical line in the spectrogram, within a single USV”. Figure 2 shows examples of USVs with the frequency shift waveform.
Figure 2. Spectrographic Display of Rat Pup Ultrasonic Vocalizations.
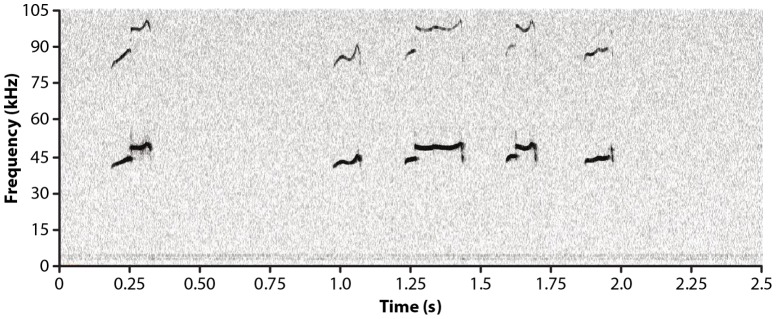
This figure shows the qualitative shift in frequency characteristics of shift in rat pup USVs that are similar to the harmonic structure of the hyperphonated cry sounds shown in Figure 4.
Although not the focus of the present investigation, a secondary set of variables were also compared in analyses to explore their sensitivity to PCE. All USVs were categorized as belonging to the nine categories described by Brudzynski et al. [38] and then consolidated for statistical analyses into five categories based on the complexity of the frequency modulation (change in the direction of the frequency): (1) Low Complexity (categories 0–3); (2) Moderate Complexity (categories 4 and 5); (3) High Complexity (categories 6 and 7); (4) Compound Complexity (category 8); (5) Disorganized (category 9). Further, all USVs in these two five-minute episodes were subjected to spectrum analysis to obtain standard measures of the minimum and maximum F0, maximum amplitude, duration, number of harmonics and inter-USV-interval used in previous studies [37]. Measures were obtained using Avisoft-SASLab Pro computer software that automatically detected and logged the selected acoustic features.
Statistical analysis
The percentage of rat pup USVs with frequency shift was determined from the audio recordings. The recordings also produced measures of the mean Number of USVs, Duration (s), Inter-USV Interval (s), Maximum Frequency (kHz), Minimum Frequency (kHz), Maximum Amplitude (dBFS), and Number of Harmonics, as well as the percentage of each of the five waveform classifications. All measures were compared with 3 (Exposure Group) ×2 (Sex) Analyses of Variance (ANOVA) followed by LSD post-hoc tests. Alpha levels were set at p<0.05 for all analyses. One pair of male-female rat pups in the SC group was excluded from analysis due to both pups showing unusually significant outlying data (extreme number of vocalizations).
Results
Study 1: Human infant crying
Figure 3 shows sex-dependent effects of cocaine in the analyses of Peak F0 and Dysphonation. For the analysis of Peak F0, a significant Exposure Group × Sex interaction [F(2, 99) = 3.18, p<.046] and post-hoc analyses showed that PCE males had a higher Peak F0 than PCE females (p<.05). No differences were found between males and females in the PPE and SC groups. Additionally, PCE males had a higher Peak F0 than SC males (p<.05). No reliable differences were found in Peak F0 between males in the PCE and PPE groups, or among females across the three groups. A similar significant Exposure Group × Sex interaction was found for Dysphonation [F(2,99) = 3.66, p<.029]. Post-hoc analyses showed that both PCE and PPE males had cries with more Dysphonation than SC males and more Dysphonation than females in their respective PCE and PPE groups (p<.05) with no significant differences among females. These results indicate that males in both drug exposed groups (PCE and PPE) had similar high-pitched cries with more dysphonation than males in the standard control group, but only PCE males had a higher-pitched cry than PCE females, as well as a higher-pitched cry than SC males.
Figure 3. Means, standard errors and significant differences of (A) Peak F0 and (B) Dysphonation for male and female infants in the three exposure groups [Standard Comparison (Control), PCE (Cocaine) and PPE (Polydrug) groups].

The dysphonation score represents the mean rating on a 4-pt scale of the amount of dysphonation in each cry expiration (see text). These acoustic features comprised a cluster of cry measures that differentiated PCE males.
Figure 4 shows sex-dependent effects of cocaine in the analyses of Inter-Cry-Interval, Number of Expiratory Sounds and Peak Amplitude. The Exposure Group × Sex interaction for ICI [F(2,99) = 3.08, p<.05] and post-hoc analyses found that PCE females had longer ICIs than PCE males and females in both the SC and PPE groups (p<.05 all comparisons). The Number of Expiratory Sounds also showed a significant Exposure Group × Sex interaction [F(2,99) = 3.44, p<.036], with PCE females showing a lower Number of Expiratory Sounds than PCE males and females in both the SC and PPE groups (all p's<.05). Lastly, Peak Amplitude likewise showed an Exposure Group × Sex interaction [F(2,99) = 4.24, p<.017], with PCE females having a lower Peak Amplitude of the cry than PCE males and females in both the PPE and SC groups (all p's<.05). No significant effects were found on any of these cry measures for males (all p's>.25) or for the mean Duration of Expiratory Sounds (all p's>.16). Thus, PCE females had longer pauses between fewer cry expirations, which were lower in amplitude, than infants in all other groups.
Figure 4. Means, standard errors and significant differences of (A) Inter-Cry-Interval, (B) Number and (C) Peak Amplitude of cry expiratory sounds for male and female infants in the three exposure groups [Standard Comparison (Control), PCE (Cocaine) and PPE (Polydrug) groups].

These acoustic features comprised a cluster of cry measures that differentiated PCE females.
The multiple measurements of Peak F0 allowed us to examine the dynamic nature of the cry along with the distribution of hyperphonation throughout the full cry sound. First, a significant main-effect of Sex for F0SD [F(1,99) = 6.18, p<.015] showed that males had a greater amount of variability in Peak F0 over the duration of the cry sound than females. Second, 30.8% (n = 33) of all infants had at least one cry expiration with hyperphonation. This acoustic characteristic occurred more often in the cries of PCE (n = 11, 35.5%) and PPE (n = 15, 39.5%) infants than in the cries of SC infants (n = 7, 18.4%) [X2(2) = 4.26, p<.049] (see Figure 5). Additionally, males accounted for the majority of infants with cries containing hyperphonation: 9 of the 11 (82%) PCE infants, 8 of the 15 (53.3%) PPE infants and 5 of the 7 (71.4%) SC infants [X2(2) = 8.55, p<.01]. Third, hyperphonation occurred in expiratory sounds throughout the 30 s crying bout: (a) in only the initial three cry expirations for 12.1% of infants, (b) in only expirations after the initial three cry expirations for 8.4% of infants, and (c) in both the initial three and subsequent cry expirations for 10.2% of crying infants. The cry expiration with the highest Peak F0, independent of whether the acoustic structure was hyperphonated or not, occurred in the portion of crying that followed the initial three expirations for 28.7% of infants, equally distributed among the three exposure groups (PCE = 28%; PPE = 30%; SC = 27.5%). Finding Peak F0 in segments other than the initial three cry expirations may have contributed to finding no reliable differences among groups in the Initial Peak F0, [F(2,99) = 2.23, p>.113]. These data indicate that cry expirations throughout the crying bout contain important acoustic information that differentiates human infants with PCE.
Figure 5. Percentage of infants with hyperphonated cries.

This figure shows the increased presence of hyperphonation in PCE and PPE infants. Whereas a similar percentage of PPE male and female infants had hyperphonated cry sounds, PCE male infants showed a higher incidence of hyperphonated cries than PCE females. See text for statistical results.
Study 2: Rat pup ultrasonic vocalizations
The thermal-isolation challenge elicited USVs from pups in all groups (PCE: 23 of 34 (67%); UN: 28 of 50 (56%); SC: 18 of 34 (52.9%). Of those who vocalized, PCE pups averaged 46.6 USVs (range: 1–158); UN pups averaged 55.6 USVs (range: 1–410) and SC pups averaged 66.3 USVs (range: 1–360) during the 10-mins selected for study. The percentage of USVs for each pup that contained shift showed a significant main-effect for Exposure Group [F(2,112) = 3.72, p<.027], as shown in Figure 6. LSD post-hoc comparisons showed that PCE pups had a significantly higher percentage of USVs containing shift in their acoustic structure than pups in the UN group (p<.009) and marginally more than pups in the SC group (p<.06). No differences were found between pups in the two control groups in the amount of shift (p>.25) nor were there any differences found in the actual number of USVs produced by any of the groups (p‘s>.25).
Figure 6. Means, standard errors and significant differences of the percentage of USVs containing Shift for male and female rat pups in the three exposure groups [Untreated (Control), Cocaine and Saline (Control)].

Cocaine-exposed pups had significantly more frequency shifts in their USVs than pups in the Untreated and Saline control groups. No sex differences were found.
ANOVAs of the secondary set of variables showed no reliable effects for sex, cocaine or cocaine by sex interactions in the percentage of other waveform categories (all p's>.25, see Table 3), nor for any of the spectral and temporal characteristics of the USVs (all p's>.19, see Table 4).
Table 3. Percentages of USVs Varying in Waveform Complexity.
| Complexity | Sex | Untreated | Cocaine | Saline |
| Low | Male | 17.6±6.1 | 10.4±6.7 | 16.5±6.1 |
| Female | 21.0±5.3 | 20.4±5.9 | 13.0±10.6 | |
| Moderate | Male | 30.8±6.9 | 28.2±7.6 | 34.9±6.9 |
| Female | 35.3±6.0 | 24.6±6.6 | 23.2±12.0 | |
| High | Male | 31.3±6.5 | 31.7±7.1 | 26.0±6.5 |
| Female | 27.4±5.7 | 28.1±6.3 | 26.2±11.3 | |
| Compound | Male | 3.4±2.4 | 2.6±2.6 | 8.9±2.4 |
| Female | 3.4±2.1 | 2.1±2.3 | 1.5±4.2 | |
| Disorganized | Male | 1.33±0.95 | 0.00±1.04 | 1.08±0.95 |
| Female | 1.06±0.82 | 1.42±0.91 | 4.75±1.64 |
Note: All values are Mean ± SEM.
Table 4. Spectral Measures of Rat Pup USVs.
| Measure | Sex | Untreated | Cocaine | Saline |
| Number of USVs | Male | 72.5±19.9 | 47.2±20.8 | 25.7±19.0 |
| Female | 36.9±17.0 | 38.1±19.0 | 35.8±32.9 | |
| Duration (s) | Male | 0.066±0.010 | 0.080±0.011 | 0.069±0.010 |
| Female | 0.072±0.009 | 0.070±0.010 | 0.060±0.017 | |
| Inter-USV Interval (s) | Male | 2.69±6.03 | 15.45±6.03 | 18.61±5.75 |
| Female | 9.22±5.29 | 10.31±5.75 | 6.33±9.54 | |
| Maximum Frequency (kHz) | Male | 51.32±3.1 | 55.47±3.3 | 49.86±3.0 |
| Female | 57.73±2.7 | 52.26±3.0 | 52.41±5.1 | |
| Minimum Frequency (kHz) | Male | 49.25±2.9 | 54.16±2.6 | 46.97±5.0 |
| Female | 51.21±3.2 | 47.45±3.0 | 46.72±2.9 | |
| Maximum Amplitude (dBFS) | Male | 58.8±1.9 | 59.1±2.0 | 53.1±1.8 |
| Female | 55.2±1.7 | 54.8±1.8 | 56.8±3.2 | |
| Number of Harmonics | Male | 1.11±0.12 | 1.10±0.13 | 1.08±0.12 |
| Female | 0.86±0.11 | 1.12±0.12 | 1.22±0.21 |
Note: All values are Mean ± SEM.
Discussion
Across species, crying can be conceptualized as a biological siren that reflects changes in arousal and neurobehavioral integrity and then communicates those changes to the social environment [46], [35]. As a biological signal reflecting infant neurobehavioral integrity, we found PCE to be associated with two sex-dependent clusters of measures of human infant crying. The first cluster of cry measures differentiated PCE males and was comprised of two acoustic features derived from the harmonic structure of the power spectrum: Peak F0, Hyperphonation, and Dysphonation. The second cluster of cry measures differentiated PCE females and was comprised of three characteristics of the production of crying: the Number and Amplitude of cry expirations and the Inter-Cry-Interval between them. As would be expected from previous studies of licit and illicit prenatal drug exposure [27]–[30], we also found measures of the harmonic structure of infant crying to be sensitive to effects of PPE in the absence of cocaine. A disproportionately higher number of PPE infants (both males and females) had cries with a hyperphonated acoustic structure and PPE males had cries with greater amounts of dysphonation than SC males. Further, experimental evidence of the effects of PCE were found on a measure of rat pup USVs that has a similar acoustic structure to that of hyperphonation in human infant cry sounds. These studies provide the first known direct translational examination of similar acoustic structures of vocalizations of two species in the detection of effects of a similar adverse prenatal condition.
Findings of the present study contribute to a growing literature documenting sex differences in behavioral and physiological reactivity following prenatal substance exposure, in general [47], and prenatal cocaine exposure, in particular [48]–[50]. PCE males are typically described as showing high reactivity to stress-eliciting conditions, such as a higher number of cry cycles in response to heel-lance procedures [48] and greater cortisol reactivity in a paradigm designed to induce stress at 7 months of age [49]. PCE males in the present study showed a cluster of cry features indicative of heightened or excitatory reactivity – cries with a higher pitch and more dysphonation. Importantly, PCE females also showed a distinct and different cluster of cry features depicting a diminished or inhibitory pattern of reactivity - cry expirations that were fewer in number, lower in amplitude and slower in repetition rate. Thus, PCE had perhaps opposite effects on reactivity patterns on male and female infants, as evidenced in the analysis of their cry sounds. Similar excitatory and inhibitory patterns of reactivity in PCE males and females, respectively, have also been found at 13 months of age in a measure of respiratory-sinus arrhythmia [50].
Interestingly, the two clusters of cry measures that differentiated male and female infants in the present study have also been associated with two orthogonal dimensions of neurobehavioral activity in newborn infants [31]. Whereas measures of the fundamental frequency loaded on a neurobehavioral dimension signaling a poorer capacity of the infant to orient to its social environment, measures of cry production loaded on a dimension indicating a poorer ability to regulate behavioral state. As such, the present study provides evidence of sex-specific effects of PCE on physiological reactivity that may be associated with different dimensions of behavior and indicates that these effects are detectable by measures of infant crying at one month of age.
In a direct benefit of the translational analysis, we found independent effects of PCE on an acoustic feature of rat pup USVs that is similar to the acoustic structure of hyperphonation found in human infant cry sounds. Consistent with suggestions that shift may reflect effects of drug exposure on neurobehavioral regulation [43], we found PCE pups to have nearly twice the number of USVs containing that waveform than the two control groups. Also consistent with previous work suggesting that steps or shifts occur more often in a physically intense thermal challenge [42], we found pups in both the UN and SC control groups had 5% to 10% of their USVs with a frequency shift in the acoustic structure as well. To the extent that PCE increases pup sensitivity to environment stressors [51], the increased presence of shift found in USVs of PCE pups may reflect an increased stress response to the physically intense thermal challenge. Whereas sex-dependent effects of PCE were found in the analysis of human infant cries, no such effects were found in rat pup USVs. However, recent work has found sex- and age-dependent effects of PCE in the number, amplitude, harmonics and duration of USVs of older rats [52]. Lastly, no significant effects of PCE or sex were found for any of the other USV waveform categories or the standard spectral and temporal measures analyzed in the present study. Thus, the presence of shift in rat pup USVs may uniquely provide a sensitive translational measure of neurobehavioral integrity following prenatal cocaine exposure.
Our understanding of the physiological mechanisms underlying effects of PCE on vocalizations varies between the two species examined in this study. For human infants, pathophysiological pathways by which PCE affects neurobehavioral function [11] and the physioacoustic basis for variations in human infant cry sounds [53], [54] have been well-described. Variations in F0, for example, have been suggested to result from activity of lower brainstem mechanisms that control tension of laryngeal muscles through the vagal complex and phrenic and thoracic nerves. Instability in these neural mechanisms is thought to produce hyperphonation, while dysphonation, amplitude and temporal morphology cry reflect inhibitory and excitatory aspects of respiratory function. In other species, the specific effects of PCE on nervous system development, including the mesolimbic dopamine system [55], have been more fully described, but less is known about how PCE specifically affects production of rodent USVs in general, and waveform shifts in particular. While the vocal anatomy of the rat pup is similar to other mammals who have vibrating vocal folds at the top of the trachea, a current consensus is that ultrasonic tones result from a still unknown whistle-type mechanism [40], [56]. Some suggest effects of PCE on USVs may result from stimulation of endogenous opioid systems [57], activation of “pleasure” systems [43], and effects of PCE on maturation of the central medial hypothalamus and central amygdala [53]. Future work that examines the bases of shift and other variations in acoustic features of USVs will contribute to our understanding of the translational value of these analyses.
As a biological siren, variations in the sound of crying are also salient social signals [46] that may contribute to problem mother-infant interactions that are often a part of the context that guides developmental outcome in cocaine-exposed infants [19]–[22]. For example, the number of expiratory cry sounds, slower cry repetition rate [58] and variability in Peak F0 [59] found in the present study have been shown to create specific perceptual responses in potential caregivers. Hyperphonation, in particular, is a powerfully salient vocal signal. In many mammals, vocalizations that induce attention and arousal often have sharp onsets, dramatic fluctuations in frequency and amplitude and either shorter or longer, upward sweeps in frequency [60]. These vocalization patterns are characteristic of both hyperphonation and shift. The attention and arousal elicited by hyperphonated cry sounds are mediated by two orthogonal perceptual dimensions that may provide the basis for ameliorative care from some caregivers and heightened distress and withdrawal from others [46], [61]. Typical of the latter response set, cocaine-using mothers report they would give less nurturing responses than comparison mothers to infant cries, especially as the sound of crying increases in fundamental frequency [62]. In this case, hyperphonated cries may contribute to maternal withdrawal and other behaviors that put the infant with PCE at further risk for detrimental social interactions, including physical child abuse and neglect [63], [64]. Similarly, variations in acoustic and temporal features of rat pup USVs are part of the sensory environment that elicits differential caregiving responses from the dam [65], yet no known work has yet examined the social significance of the shift waveform examined in this study. Future work would benefit from the exploration of the functional significance of this unusual acoustic characteristic.
Several limitations of the methods should be considered when interpreting the results of these studies. The Time Line Follow Back and urine analyses do not provide a full understanding of the timing and amount of human maternal cocaine-use during pregnancy. The two species differed in the vehicle by which cocaine-exposure was delivered, the methods by which vocalizations were elicited and the developmental age at which vocalizations were recorded. Although there was no evidence of such, variations in spectral and temporal characteristics of the cries of infants with PCE could also be possibly due to unknown adverse conditions that occurred during the first postnatal month. The effect of the long observation period on the rate of rat pup vocalizations in this study is unknown. Last, exploratory analyses of the secondary set of measures of rat pup USVs were conducted to “rule out” their sensitivity to the effects of PCE. These analyses increased the number of statistical comparisons and possibility of a Type I error in which significant findings may have occurred by chance. While none of these secondary, exploratory analyses were statistically significant, most of the primary measures that were the focus of our study were statistically reliable.
In conclusion, these two studies provide the first known direct translational examination of similar acoustic structures of vocalizations of two species in the detection of effects of a similar adverse prenatal condition. A novel set of measures in an analysis of an extended period of human infant crying uncovered different effects of PCE on patterns of reactivity in male and female infants. The analysis of shift in rat pup USVs distinguished PCE pups from controls – in the absence of variables that often confound results in studies of human infants. While the context and mechanisms producing vocalizations differ between human infants and rat pups, there is an intriguing similarity of acoustic structures following prenatal cocaine-exposure that differentiated their vocalizations. Further, while many challenges preclude making direct homologous comparisons between the distress vocalizations of human infants and rat pups, a recent review concludes that there is ample evidence for employing apparent similarities in research on the biological effects of PCE on animals and humans, as well as contributing effects of the rearing environment [55]. Within this context, the variations in the sounds of infant crying and rat pup USVs may also have important functional value, impacting the rearing environment. As such, the analysis of cry vocalizations may have significant translational value by affording a window into both the neurobehavioral and social complexities underlying the development of infants with prenatal cocaine and other drug exposures. The methods and findings presented here may provide the basis for future research questions and a foundation for future translational analyses into a possibly wide range of developmental disorders.
Supporting Information
Raw data from the experiment.
(ZIP)
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the National Institute on Drug Abuse [grant numbers DA022446 to JMJ, DA030060 to ETC and DA026251 to MSM]. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Lambert BL, Bauer CR (2012) Developmental and behavioral consequences of prenatal cocaine exposure: a review. J Perinatol 11: 819–828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Cain MA, Bornick P, Whiteman V (2013) The maternal, fetal, and neonatal effects of cocaine exposure in pregnancy. Clin Obstet Gynecol 56: 124–132. [DOI] [PubMed] [Google Scholar]
- 3. Behnke M, Smith VC (2013) Prenatal substance abuse: Short- and long-term effects on the exposed fetus. Pediatrics 131: 1009–1024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Lester BM, LaGasse LL, Seifer R (1998) Cocaine exposure and children: The meaning of subtle effects. Science 282: 633–634. [DOI] [PubMed] [Google Scholar]
- 5. Eiden RD, Coles CD, Schuetze P, Colder CR (2013) Externalizing behavior problems among polydrug cocaine-exposed children: Indirect pathways via maternal harshness and self-regulation in early childhood. Psych Addictive Behav 28(1): 139–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Schuetze P, Molnar DS, Eiden RD (2012) Profiles of reactivity in cocaine-exposed children,. J Applied Dev Psych 33: 282–293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Lester BM, LaGasse LL, Shankaran S, Bada H, Bauer CR, et al. (2010) Prenatal cocaine exposure alters cortisol stress reactivity in 11 year old children. J Pediatr 157: 288–295.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Bridgett DJ, Mayes LC (2011) Development of inhibitory control among prenatally cocaine exposed and non-cocaine exposed youths from late childhood to early adolescence: The effects of gender and risk and subsequent aggressive behavior. Neurotoxicol Teratol 33: 47–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Rando K, Chaplin TM, Potenza MN, Mayes L, Sinha R (2013) Prenatal cocaine exposure and gray matter volume in adolescent boys and girls: Relationship to substance use initiation. Biol Psychiatry 74: 482–489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Lester BM, Lin H, DeGarmo DS, Fisher P, LaGasse LL, et al. (2012) Neurobehavioral disinhibition predicts initiation of substance use in children with prenatal cocaine exposure. Drug and Alc Dependence 126: 80–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Lester BM, Padbury JF (2009) The third pathophysiology of prenatal cocaine exposure. Dev Neurosci 31: 23–35. [DOI] [PubMed] [Google Scholar]
- 12. Lester BM, Tronick EZ, LaGasse L, Seifer R, Bauer CR, et al. (2002) The maternal lifestyle study (MLS): effects of substance exposure during pregnancy on neurodevelopmental outcome in 1-month-old infants. Pediatrics 110: 1182–1192. [DOI] [PubMed] [Google Scholar]
- 13. Eyler FD, Behnke M, Conlon M, Woods NS, Wobie K (1998) Birth outcome from a prospective, matched study of prenatal crack/cocaine use: II. Interactive and dose effects on neurobehavioral assessment. Pediatrics 101: 237–241. [DOI] [PubMed] [Google Scholar]
- 14. DiPietro JA, Suess PE, Wheeler JS, Smouse PH, Newlin DB (1995) Reactivity and regulation in cocaine-exposed neonates. Inf Beh Devel 18: 407–414. [Google Scholar]
- 15. Coles CD, Platzman KA, Smith I, James ME (1992) Effects of cocaine and alcohol use in pregnancy on neonatal growth and neurobehavioral status. Neurotox Teratol 14: 23–33. [DOI] [PubMed] [Google Scholar]
- 16. Schuetze P, Eiden RD, Danielwicz S (2009) The association between prenatal cocaine exposure and physiological regulation at 13 months of age. J Child Psychol Psychiatry 50: 1401–1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Mayes LC, Bornstein MH, Chawarski K, Haynes OM (1996) Impaired regulation of arousal in 3-month old infants exposed prenatally to cocaine and other drugs. Devel and Psychopathol 8: 29–42. [Google Scholar]
- 18. Behnke M, Eyler FD, Garvan CW, Wobie K, Hou W (2002) Cocaine exposure and developmental outcome from birth to six months. Neurotoxicol Teratol 24: 283–295. [DOI] [PubMed] [Google Scholar]
- 19. Strathearn L, Mayes LC (2010) Cocaine addiction in mothers. Ann NY Acad Sci 1187: 172–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Eiden RD, Peterson M, Coleman T (1999) Maternal substance use and the caregiving environment during early childhood. Psych of Add Beh 13: 293–302. [Google Scholar]
- 21. Nephew BC, Febo M (2012) Effects of Cocaine on Maternal Behavior and Neurochemistry. Cur Neuropharmacol 10: 53–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Arendt RE, Short E, Singer LT, Minnes S, Hewitt J, et al. (2004) Children prenatally exposed to cocaine: Developmental outcomes and environmental risks at seven years of age. Devel Behav Pediatr 25: 83–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Messinger DS, Bauer CR, Das A, Seifer R, Lester BM, et al. (2004) The maternal lifestyle study: Cognitive, motor, and behavioral outcomes of cocaine-exposed and opiate-exposed infants through three years of age. Pediatrics 113: 1677–1685. [DOI] [PubMed] [Google Scholar]
- 24.LaGasse LL, Seifer R, Lester BM (1999) Interpreting research on prenatal substance exposure in the context of multiple confounding factors. In: Lester BM, editor. Clinics in Perinatology: Prenatal Drug Exposure and Child Outcome. Philadelphia: WB Saunders. pp. 9–54. [PubMed] [Google Scholar]
- 25. LaGasse LL, Neal AR, Lester BM (2005) Assessment of infant cry: Acoustic cry analysis and parental perception. Mental Retar and Dev Dis Res Rev 11: 83–93. [DOI] [PubMed] [Google Scholar]
- 26.Zeskind PS, Lester BM (2001) The diagnostic utility of the assessment of infant cry sounds. In: Singer LT, Zeskind PS, editors. Biobehavioral assessment of the newborn and young infant. New York: Guilford Publications. [Google Scholar]
- 27. Huntington L, Hans SL, Zeskind PS (1990) The relations among cry characteristics, demographic variables, and developmental test scores in infants prenatally exposed to methadone. Inf Beh Devel 13: 533–538. [Google Scholar]
- 28. Lester BM, Dreher M (1989) Effects of marijuana use during pregnancy on newborn cry. Child Devel 60: 765–771. [PubMed] [Google Scholar]
- 29. Nugent JK, Lester BM, Greene SM, Wieczorek Deering D, O'Mahony P (1996) The effects of maternal alcohol consumption and cigarette smoking during pregnancy on acoustic cry analysis. Child Dev 67: 1806–1815. [PubMed] [Google Scholar]
- 30. Zeskind PS, Platzman K, Coles CD, Schuetze P (1996) Cry analysis detects subclinical effects of prenatal alcohol exposure. Inf Beh Devel 19: 497–500. [Google Scholar]
- 31.Zeskind PS (1983) Production and spectral analysis of neonatal crying and its relation to other biobehavioral systems in the infant at risk. In: Field T, Sostek A editors. Infants Born At Risk: Physiological, Perceptual, and Cognitive Processes. Philadelphia: WB Saunders. pp. 23–43. [Google Scholar]
- 32. Corwin MJ, Lester BM, Sepkoski C, McLaughlin S, Kayne H, et al. (1992) Effects of in utero cocaine exposure on newborn acoustical cry characteristics. Pediatrics 89: 1199–1203. [PubMed] [Google Scholar]
- 33. Lester BM1, Corwin MJ, Sepkoski C, Seifer R, Peucker M, et al. (1991) Neurobehavioral syndromes in cocaine-exposed newborn infants. Child Dev 62(4): 694–705. [DOI] [PubMed] [Google Scholar]
- 34.Brudzynski SM (2010) Handbook of Mammalian Vocalization: An Integrative Neuroscience Approach. London: Academic Press. [Google Scholar]
- 35.Altenmuller E, Schmidt S, Zimmerman E (2012) Evolution of Emotional Communication: From Sounds in Nonhuman Mammals to Speech and Music in Man. Oxford: Oxford University Press. [Google Scholar]
- 36. Hahn ME, Benno RH, Schanz N, Phadia E (2000) The effects of prenatal cocaine exposure and genotype on the ultrasonic calls of infant mice. Pharm Biochem and Beh 67: 729–738. [DOI] [PubMed] [Google Scholar]
- 37. McMurray MS, Zeskind PS, Meiners SM, Garber KA, Tien H, et al. (2013) Effects of prenatal cocaine on early postnatal thermoregulation and ultrasonic vocalization production. Front Psychol 4: 882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Brudzynski SM, Kehoe P, Callahan M (1999) Sonographic structure of isolation-induced ultrasonic calls of rat pups. Dev Psychobiol 34: 195–204. [DOI] [PubMed] [Google Scholar]
- 39. Barron S, Gilbertson R (2005) Neonatal ethanol exposure but not neonatal cocaine selectively reduces specific isolation-induced vocalization waveforms in rats. Beh Gen 35: 93–102. [DOI] [PubMed] [Google Scholar]
- 40. Roberts LH (1975) Evidence for the laryngeal source of ultrasonic and audible cries of rodents. J Zool 175: 243–257. [Google Scholar]
- 41. Sales GD, Smith JC (1978) Comparative studies of the ultrasonic calls of infant murid rodents. Dev Psychobiol 11: 595–619. [DOI] [PubMed] [Google Scholar]
- 42. Branchi I, Santucci D, Vitale A, Alleva E (1998) Ultrasonic vocalizations by infant laboratory mice: a preliminary spectrographic characterization under different conditions. Dev Psychobiol 33: 249–256. [DOI] [PubMed] [Google Scholar]
- 43.Burgdorf J, Moskal JR (2010) Frequency modulated 50 kHz ultrasonic vocalizations reflect a positive emotional state in the rat: neural substrates and therapeutic implications. In: Brudzynski, SM editor. Handbook of Mammalian Vocalization: An Integrative Neuroscience Approach. London: Academic Press. pp. 209–214. [Google Scholar]
- 44. Zeskind PS, McMurray MS, Garber KA, Neuspiel JM, Cox ET, et al. (2011) Development of translational methods in spectral analysis of human infant crying and rat pup ultrasonic vocalizations for early neurobehavioral assessment. Front Psychiatry 2: 1–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Tutag Lehr V, Zeskind PS, Frattarelli DAC, Ofenstein JP, Cepeda E, et al. (2007) Neonatal facial coding scale scores and spectral characteristics of infant crying during newborn circumcision. Clin J Pain 23: 417–424. [DOI] [PubMed] [Google Scholar]
- 46.Zeskind PS (2012) Infant crying and the synchrony of arousal. In: Altenmuller E, Schmidt S, Zimmerman E, editors. Evolution of Emotional Communication: From Sounds in Nonhuman Mammals to Speech and Music in Man. Oxford: Oxford University Press. pp. 155–174. [Google Scholar]
- 47.Lewis M, Kestler L (2012) Gender Differences in Prenatal Substance Exposure. Washington DC: American Psychological Association. [Google Scholar]
- 48. Grunau RV, Craig KD (1987) Pain expression in neonates: facial action and cry. Pain 28: 395–410. [DOI] [PubMed] [Google Scholar]
- 49. Eiden RD, Veira Y, Granger D (2009) Prenatal cocaine exposure and infant cortisol reactivity. Child Dev 80: 528–543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Schuetze P, Eiden RD, Danielwicz S (2009) The association between prenatal cocaine exposure and physiological regulation at 13 months of age. J Child Psych and Psychiatry 50(11): 1401–1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Spear LP, Campbell J, Snyder K, Silveri M, Katovic N (1998) Animal behavior models: Increased sensitivity to stressors and other environmental experiences after prenatal cocaine exposure. In: Harvey JA, Kosofsky BE, editors. Cocaine: Effects on the Developing Brain. New York: New York Academy of Sciences. pp. 76–88. [PubMed] [Google Scholar]
- 52. Cox ET, Hodge CW, Sheikh MJ, Abramowitz AC, Jones GF, et al. (2012) Delayed developmental changes in neonatal vocalizations correlates with variations in ventral medial hypothalamus and central amygdala development in the rodent infant: effects of prenatal cocaine. Behav Brain Res 235(2): 166–75.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Golub HL, Corwin MJ (1985) A physioacoustic model of the infant cry. In: Lester BM, Boukydis CFZ, editors. Infant Crying: Theoretical and Research Perspectives. New York: Plenum. pp. 59–82. [Google Scholar]
- 54.Lester BM (1984) A biosocial model of infant crying. In: Lipsitt L, editor. Advances in Infant Research. Norwood: Ablex. pp. 167–212. [Google Scholar]
- 55. Dow-Edwards D (2011) Translational issues for prenatal cocaine studies and the role of environment. Neurotoxicol Teratol 33: 9–16. [DOI] [PubMed] [Google Scholar]
- 56.Brudzynski SM, Fletcher NH (2010) Rat ultrasonic vocalization: short-range communication. In: Brudzynski SM, editor. Handbook of Mammalian Vocalization: An Integrative Neuroscience Approach. London: Academic Presspp. 69–76. [Google Scholar]
- 57. Kehoe P, Boylan C (1992) Cocaine-induced effects on isolation stress in neonatal rats. Behav Neurosci 106: 374–379. [DOI] [PubMed] [Google Scholar]
- 58. Zeskind PS, Klein L, Marshall TR (1992) Experimental modification of relative durations of pauses and expiratory sounds in infant cries alters adults' perceptions. Dev Psychol 28: 1153–1162. [Google Scholar]
- 59. Zeskind PS, Marshall TR (1988) The relation between mothers' perceptions and pitch of infant crying. Child Dev 59: 193–196. [Google Scholar]
- 60. Owren MJ, Rendell D (2001) Sound on the rebound: bringing form and function back to the forefront in understanding nonhuman primate vocal signaling. Evol Anthro 10: 58–71. [Google Scholar]
- 61. Zeskind PS, Lester BM (1978) Acoustic features and auditory perceptions of the cries of newborns with prenatal and perinatal complications. Child Dev 49: 580–589. [PubMed] [Google Scholar]
- 62. Schuetze P, Zeskind PS, Eiden RD (2003) The perceptions of infant distress signals varying in pitch by cocaine-using mothers. Infancy 4: 25–34. [DOI] [PubMed] [Google Scholar]
- 63. Zeskind PS, Ramey CT (1981) Prevention of intellectual and interactional sequelae of fetal malnutrition: a longitudinal, transactional and synergistic approach to development. Child Dev 52: 214–218. [PubMed] [Google Scholar]
- 64. Crowe HP, Zeskind PS (1992) Psychophysiological and perceptual responses to infant cries varying in pitch: comparison of adults with low and high scores on the Child Abuse Potential Inventory. Child Abuse and Negl 16: 19–29. [DOI] [PubMed] [Google Scholar]
- 65. Zimmerberg B, Kim JH, Davidson AN, Rosenthal AJ (2003) Early deprivation alters the vocalization behavior of neonates directing maternal attention in a rat model of child neglect. Ann N Y Acad Sci 1008: 308–13. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Raw data from the experiment.
(ZIP)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.