

Abstract
Drawing on an idea proposed by Darwin, it has recently been hypothesized that violent intergroup conflict might have played a substantial role in the evolution of human cooperativeness and altruism. The central notion of this argument, dubbed ‘parochial altruism’, is that the two genetic or cultural traits, aggressiveness against the out-groups and cooperativeness towards the in-group, including self-sacrificial altruistic behaviour, might have coevolved in humans. This review assesses the explanatory power of current theories of ‘parochial altruism’. After a brief synopsis of the existing literature, two pitfalls in the interpretation of the most widely used models are discussed: potential direct benefits and high relatedness between group members implicitly induced by assumptions about conflict structure and frequency. Then, a number of simplifying assumptions made in the construction of these models are pointed out which currently limit their explanatory power. Next, relevant empirical evidence from several disciplines which could guide future theoretical extensions is reviewed. Finally, selected alternative accounts of evolutionary links between intergroup conflict and intragroup cooperation are briefly discussed which could be integrated with parochial altruism in the future.
Keywords: intergroup conflict, altruism, parochialism, cooperation, warfare, coevolution
1. Introduction
Modern human (Homo sapiens) violent intergroup conflict, i.e. ‘war’, is unparalleled in scale and its functional differentiation. The problem of understanding how humans developed the abilities required to carry out such highly organized, deadly campaigns against conspecifics has long troubled researchers from various disciplines. The theory of ‘parochial altruism’ [1] is one of the most recent elaborations of Darwin's idea [2] that the readiness to behave altruistically to the benefit of in-group members (‘in-group love’) and to act hostilely toward out-groups (‘out-group hate’) have been closely linked in human evolution. Using complex game theoretical models and simulations, a number of recent studies have explored the conditions under which natural selection might favour the coevolution of these two traits [1,3–5] (see the electronic supplementary material, table S1, for an overview).
The aim of this review is threefold. First, a brief synopsis of the existing modelling approaches to parochial altruism is given in §2. Thereafter, two potential pitfalls in the interpretation of these models are addressed in §3. To assess their explanatory power and to highlight directions for their future extension, §4 points out a number of crucial limitations of the current models and §5 highlights relevant empirical evidence instructive for future work on parochial altruism. Finally, §6 presents selected alternative theories on motivations for in-group love and out-group hate which might be integrated with parochial altruism to form an overarching theory of the evolution of human intergroup conflict behaviour.
2. A brief overview of current theories of parochial altruism
The central aim of all theories on parochial altruism is to explain why we observe (i) such high levels of cooperation and altruism between (even unrelated) individuals [1] and (ii) such extreme scales of intergroup violence [4,6] in humans. While other theories concerned with human intergroup conflict focus on direct fitness benefits attainable through discrimination and cooperative aggression against out-groups (see §6), theories on parochial altruism start from the assumption that these behaviours might, at least to some degree, be truly altruistic, i.e. causing substantial and recurrent fitness costs for the parochial altruists [1,4], and explore scenarios in which these two traits could have coevolved.
To start with, all existing models assume a meta-population composed of multiple subgroups. Group membership is usually assumed to be heritable, which implicitly limits the dispersal of traits [7]. If modelled, within-group interactions are characterized as either inter-individual cooperation problems, for which the standard paradigm is the two-person Prisoner's Dilemma, or as collective action problems modelled as n-person Prisoner's Dilemmas or public goods games (PGG). With respect to behaviour towards in-group members, individuals generally are modelled as either ‘cooperators’ or ‘defectors’, or graded blends of these two types. Defectors never cooperate and free-ride on all public goods produced by cooperative in-group members. By doing so, they maximize their individual fitness compared with cooperative in-group members. Cooperators, on the other hand, do engage in in-group beneficial interactions, either by cooperating with in-group members in pairwise interactions or by contributing to the production of public goods, which are assumed to be shared equally among all in-group members. Consequently, cooperators incur a relative fitness disadvantage compared to defectors of their in-group because they pay the costs of cooperation, whereas defectors do not. Cooperators are selected against by differential, fitness proportional reproduction when the following conditions are met: (i) no additional mechanisms are present and (ii) the positive effect of limited dispersal on cooperators is balanced by sufficient levels of migration between groups [7]. Most models take measures to satisfy condition (ii). Models differ, however, in the details of the mechanisms of intergroup interaction that are added to this baseline set-up.
Before discussing these mechanisms in detail, however, a clarification of the terminology to be used is in order. It has proven fruitful to distinguish four basic categories of social behaviour based on their (direct) fitness consequences for the actors and for the individuals affected by their actions (‘recipients’ for short; [8]): mutually beneficial behaviour (+/+) increases the fitness of both actor and recipients, selfish behaviour (+/−) benefits the actor while harming the recipients, spiteful behaviour (−/−) harms both and altruistic behaviour (−/+) benefits the recipients while harming the actor. While selfish and mutually beneficial behaviours can evolve through natural selection on individual fitness, spite and altruism are conventionally assumed to evolve through selection on inclusive fitness [9–12]. Distinguishing these different evolutionary routes is crucial for a biologically meaningful interpretation of the models of parochial altruism which is conventionally characterized as a combination of ‘in-group altruism’ and ‘out-group spite’ [1,13]. It will be discussed later, in §3a, if this characterization is fully appropriate.
In the studies reviewed, intergroup interaction usually is modelled in one of two ways, although combinations of both can also be found. Either individuals of distinct groups are matched pairwise to interact or actual intergroup encounters are assumed. In both cases, however, the matching process is usually assumed to be strictly random to avoid additional assortment effects (but see [5]).
In pairwise matching, individuals are assumed to be able to condition their behaviour on their partner's group membership, resulting in four basic behavioural strategies: indiscriminate cooperation and defection, in-group favouritism, i.e. cooperation with in-group members plus defection against out-group members, and, vice versa, out-group favouritism. The latter, though, is often treated as a purely theoretical possibility (but see [14]).
The literature reviewed finds that in-group favouritism in pairwise interactions can evolve when: (i) groups are rather small while many groups coexist; (ii) migration and mutation rates are sufficiently low; and (iii) group membership is sufficiently reliably identifiable. It should be noted, however, that the two-person Prisoner's Dilemma paradigm does not capture spiteful ‘−/−’-behaviour towards out-group members [10]. In the Prisoner's Dilemma, individuals can only choose not to cooperate, thus forgoing potential benefits of mutual cooperation, but simultaneously protecting themselves from being exploited. Models exclusively using this paradigm thus, by definition, cannot speak to the question of why groups might try to harm each other. They can only shed light on the question of why individuals from different groups might hesitate to cooperate, i.e. they offer potential explanations for ‘in-group love’, but not for ‘out-group hate’ (also see §6a). For this reason, the rest of this review focuses on models which are not restricted to pairwise interactions, but implement some variant of between-group contest.
The conventional contest mechanism used in the existing models randomly matches groups pairwise and compares them by the amount of within-group welfare they produce, i.e. the sums of each group's members' pay-offs earned in within-group interaction. The group with lower welfare is then replaced with a newly created group consisting of fitness proportionally procreated offspring of the individuals of the superior group. The probability with which groups are selected to compete in this way, i.e. the ‘frequency of wars’, often is varied systematically, consistently indicating that higher frequencies of conflict favour selection for within-group cooperation. More elaborate models allow for a calibration of the fraction of individuals killed in the defeated group, i.e. the ‘brutality’ of the victors. The majority of the models, however, simply assume the total annihilation of the losing group (see the electronic supplementary material, table S1).
To summarize, the existing models show that Darwin's idea that cooperative groups might be favoured by selection when intergroup conflicts are frequent is consistent, given that certain assumptions about population and interaction structure hold. Particularly, when in-group favouritism and aggression against out-groups coincide, evolutionary dynamics become viable in which these potentially individual costly traits coevolve and spread in the meta-population. Before discussing crucial limitations of the existing models in §4, however, two pitfalls in their interpretation will be highlighted.
3. Interpreting the existing models on parochial altruism
Two basic requirements for a meaningful evolutionary interpretation of theoretical models of animal behaviour are: (i) information on the net direct fitness consequences of a particular behavioural trait for both actor and recipients, and (ii) information on genetic relatedness of the interacting parties, i.e. information on net indirect fitness consequences. The existing studies on parochial altruism, however, have not yet explicitly analysed these factors (see [13,15–17] for relevant analogous discussions).
(a). Direct fitness consequences of parochial altruism
A simple example can illustrate the intricacies of determining the direct fitness consequences of behaviour in a situation with intergroup conflict present. Assume there are two groups of equal size n and equal initial individual resource endowments e. Also assume that individuals are not related at all. Within each group, a one-shot linear PGG is played, characterized by the pay-off function πi(ci) = e + bGk − ci, where 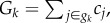 i.e. the (real-valued) contributions ci ∈ [0, e] of all individuals i in a given group gk are summed, multiplied with a constant m > 1 and then redistributed equally among all group members; let b ≡ m/n for simplicity. Wherever b < 1, this represents an n-person Prisoner's Dilemma situation in which non-contribution, ci = 0, is the fitness maximizing strategy for all individuals and in which values of ci > 0 thus represent ‘−/+’-behaviour. The resulting equilibrium of full defection is inefficient, though. Now, add a simple intergroup conflict mechanism to this set-up, e.g. let the group with the lower value of Gk be annihilated completely after the PGG has been played. In case of a draw, G1 = G2, let the surviving group be chosen randomly with a 50% chance. Even without a full analysis, it is obvious that this modification fundamentally changes the incentive structure: assume all individuals are fully defective. Any given individual would then receive an expected pay-off of e/2. Every individual now has an incentive to raise its contribution, ci, by a marginal unit ɛ, because this marginal unilateral increase would remove the threat of being annihilated and raise the individual's pay-off to e − (1−b)ɛ ≈ e. Thus, full defection, ci = 0, no longer represents the individually fitness maximizing strategy, i.e. positive contributions turn into mutually beneficial ‘+/+’-behaviour in this case. Rather, full contribution, ci = e, becomes an equilibrium strategy in this simple example, because unilateral deviation from full contribution would mean certain annihilation.
i.e. the (real-valued) contributions ci ∈ [0, e] of all individuals i in a given group gk are summed, multiplied with a constant m > 1 and then redistributed equally among all group members; let b ≡ m/n for simplicity. Wherever b < 1, this represents an n-person Prisoner's Dilemma situation in which non-contribution, ci = 0, is the fitness maximizing strategy for all individuals and in which values of ci > 0 thus represent ‘−/+’-behaviour. The resulting equilibrium of full defection is inefficient, though. Now, add a simple intergroup conflict mechanism to this set-up, e.g. let the group with the lower value of Gk be annihilated completely after the PGG has been played. In case of a draw, G1 = G2, let the surviving group be chosen randomly with a 50% chance. Even without a full analysis, it is obvious that this modification fundamentally changes the incentive structure: assume all individuals are fully defective. Any given individual would then receive an expected pay-off of e/2. Every individual now has an incentive to raise its contribution, ci, by a marginal unit ɛ, because this marginal unilateral increase would remove the threat of being annihilated and raise the individual's pay-off to e − (1−b)ɛ ≈ e. Thus, full defection, ci = 0, no longer represents the individually fitness maximizing strategy, i.e. positive contributions turn into mutually beneficial ‘+/+’-behaviour in this case. Rather, full contribution, ci = e, becomes an equilibrium strategy in this simple example, because unilateral deviation from full contribution would mean certain annihilation.
In more complicated cases, equilibrium strategies will depend on the underlying model assumptions about individual endowments, frequency and brutality of conflicts, etc. Obviously, however, intergroup conflict can introduce a threshold value θ for individual contributions which marks a tipping point: as long as ci < θ, positive contributions represent ‘+/+’-behaviour and only where ci > θ we have actual ‘−/+’-behaviour, i.e. (parochial) altruism. It is an important task for future research to analyse the interplay of the characteristics of models including intergroup conflict, the resulting equilibria and evolutionary dynamics more closely (see also [13,15–17]).
(b). Indirect fitness consequences: relatedness within and between groups
As has just been argued, it is not always the case that full defection maximizes individual fitness in scenarios of intergroup conflict even when individuals are unrelated. Conversely, it has already been analysed in some detail [18,19], how relatedness changes the incentives in PGGs played by groups including kin when no intergroup conflict is present. These analyses analogously show that positive contributions to the production of public goods can be an optimal strategy when accounting for inclusive fitness benefits; i.e. while positive contributions represent altruistic ‘−/+’-behaviour on the individual level, when sufficient numbers of relatives are present in the target individual's group this behaviour can be promoted through kin selection.
The existing models of parochial altruism frequently include some migration between groups and assume that this migration is strictly random to counterbalance the positive effect of limited dispersal, i.e. high induced relatedness, on cooperative individuals [7]. Nevertheless, how closely related the individuals in these models actually are must be analysed in more detail in future studies (see [16] for an advance in this direction). If the assumptions of frequent, very brutal conflicts and limited migration between groups induce sufficiently high levels of relatedness between group members in these models, parochially altruistic behaviour could be explained as a strategy maximizing the inclusive fitness of the bearers of this trait. However, it currently seems that the literature on parochial altruism is in danger of sparking yet another reiteration of the group selection debate [20]. This could be amended by including thorough analyses of the kinship structures implicitly assumed in these models.
To summarize, our understanding of the existing theoretical models on parochial altruism is currently incomplete in two domains: it has not been fully illuminated to what extent positive contributions to the production of public goods represent: (i) behaviour that is mutually beneficial, and thus directly fostered by natural selection, given the modified incentive structure induced by intergroup conflicts; and (ii) behaviour that is altruistic but benefits sufficiently many relatives of the altruists, thus spreading through kin selection.
While the clarification of these issues is important for future research, the existing models of parochial altruism have already been used frequently to motivate and explain the results of empirical research on human intergroup conflict behaviour. The following section critically assesses the explanatory power of the current models by highlighting their limitations. Future research will hopefully transform this criticism directly into extensions of the current models in order to further our understanding of the evolutionary interaction of in-group love and out-group hate.
4. Limitations of existing models on parochial altruism
As the theory of parochial altruism is relatively new, most existing studies focus on developing elementary models. To this end, they use a number of simplifying assumptions. By their nature, simplifications limit applicability. Now that parochial altruism has become a consistent theory, however, it is time to evaluate its current explanatory power by pointing out real-world phenomena which it is not yet able to describe. After a discussion of five such limitations in this section, relevant empirical evidence is reviewed in §5.
(a). Missing strategies for peaceful interactions
Most current models of parochial altruism do not distinguish peacetime within-group cooperation from the contributions to ‘war efforts’ (but see [1]). While it is certainly plausible that groups prospering because of working in-group cooperation might possess more resources to invest in fighting, it seems unrealistic that they will unconditionally do so. Cooperative groups could also refrain from conflict, invest their resources in peacekeeping efforts or even pay danegeld to aggressors. As long as the costs of appeasing are smaller than the loss entailed by fighting, there is a yet unexplored incentive for groups to ‘buy peace’. Most current studies on parochial altruism, however, do not discuss the conditions favouring behaviours other than intergroup hostility (but see [1]). Furthermore, by identifying peacetime cooperation with war efforts, the conventional modelling approach excludes parochial altruists from the benefits of peace, leaving to them only defection or out-group aggression as behavioural options.
(b). Equal distribution of the benefits of war and individual differences
Another simplifying assumption, inherent in the PGG paradigm of in-group cooperation implemented by most studies, is equal resource distribution among victors combined with identical investible endowments. It certainly makes some sense to assume that the costs of defeat are distributed equally, or randomly, among the defeated. It can be questioned, though, if the subgroup of those individuals in the victorious group who bear the risk of dying in order to bring about victory will be willing to share the ‘spoils of war’ equally with non-contributors of their in-group.
It largely depends on what kinds of goods these spoils of war are and on their respective fitness relevance for individual group members if it is apt to represent them as non-excludable and equally shared. Territorial gains and deterrence are frequently mentioned as examples of public goods produced through conflict. It is an important question, though, how relevant these goods are compared to excludable loot, like food, livestock, slaves and, in particular, women (also see §5c). If we assume that among the spoils of war there potentially are also private goods and/or that the distribution of at least some loot might not be independent of individual effort, this could substantially change the incentives for participation in intergroup conflict, at least for some individuals. Their participation would then be better described as mutually beneficial or even selfish behaviour rather than as altruism (also see §6). Deeper analyses of more realistic incentive structures which acknowledge that individuals of the same group differ—by sex, physical strength, relatedness, etc.—and might be affected by the outcomes of intergroup conflicts in quite distinct ways are indispensable. Two instructive analyses of this kind can be found in the literature [21,22] (also see §6c).
(c). Differences between attacks and defences
Most models of parochial altruism implicitly presume that attacks and defences have a symmetrical structure regarding risks and benefits (but see [1]). This assumption, however, is unlikely to hold when applied to many species, including humans. In territorial species, an attack requires that a subgroup of the aggressors leaves the home range, travels into enemy territory, strikes and returns, transporting back loot, if applicable. The most important points to account for are: (i) attackers only have their own lives to lose if the attack fails—at least in the short run; (ii) aggressors can choose to retreat prior to an outbreak of violence, e.g. when risk seems high or prospect of loot low; and (iii) attackers usually have the advantage of surprise on their side. These factors substantially reduce risks for individual attackers [23]. For defenders, the situation presumably is much worse. (i) Defenders likely are in disarray initially. (ii) They are probably outnumbered. (iii) Not only defenders' own lives but also those of their spouses, offspring and other relatives are in imminent danger, probably precluding fight. Defenders, however, are likely to have a home advantage [24]. Two recent studies provide further analyses and a first empirical test of behavioural adaptations potentially resulting from this asymmetry of attacks and defences [25,26].
(d). The assumption of total war between groups
The majority of models assume that of two competing groups, only the more in-group cooperative one will survive and take the place of the less cooperative one. Some authors acknowledge that this is unlikely to be true for individual ancestral skirmishes. Rather, they argue, this should be regarded as the long-term outcome of intergroup conflict [3]. A deeper analysis of at least two points seems promising here.
(i) Before being able to repopulate the ‘cleansed’ habitats, many members of the ultimately victorious group will probably also have lost their lives in battle. None of the existing models allows for less cooperative third parties to seize the opportunity of attacking more cooperative groups while they are weakened by conflict. It has also not been analysed, yet, how the temporary, but substantial, decreases in growth that fighting groups suffer during times of conflict relate to growth rates of less cooperative groups engaging in fewer conflicts. Both these limitations are due to the modelling convention of letting all conflicts be fought pairwise and simultaneously, so that repopulation is always complete before the next round of conflicts begins.
(ii) It can be questioned if every war will be fought to its bitter end. It would be very interesting to incorporate mechanisms of surrender and peace-making into the models in order to be able to analyse conditions favouring peaceful solutions of conflicts (see §4a).
(e). Sex differences
Current models frequently abstract away from sexual selection and reproduction. It is important to introduce these factors into future models in order to understand sex differences in intergroup conflict psychology better (an advance in this direction has been made, e.g. by [22]). Currently, however, parochial altruism neither offers an explanation for these repeatedly observed sex differences (see §§5 and 6) nor for the repeatedly observed lack of such differences regarding altruism and prosociality, in general [27].
5. Relevant empirical evidence from humans and other animals
Much relevant empirical evidence regarding intergroup conflict behaviour is available not only from the anthropological and psychological literature but also from studies on chimpanzees (Pan troglodytes) and on species more distantly related to humans. The empirical studies listed in this section directly relate to the model limitations discussed above and provide instructive starting points for their future extension.
(a). Non-human animals
(i). Conditional strategies in intergroup conflict
Two recent studies on green wood hoopoes (Phoeniculus purpureus) found that these cooperatively breeding territorial birds seem to strategically use allopreening to increase in-group affiliation which then translates into a greater ability to defend territories [28,29]. While at first this might look like a simple form of parochial altruism, a closer analysis revealed that the observed increase in allopreening results from increased efforts of the dominant breeding pair, i.e. those birds in the group which suffer the greatest fitness costs in case of a failed defence, indicating that they use prosocial behaviour as a means to reduce individual risk of fitness loss.
In a similar vein, a recent study on red wood ants (Formica rufa) established that these aggressive territorial colonial ants condition their fighting effort on the perceived resource holding potential of their own compared with the enemy group [30]. Whenever the in-group is superior, individual ants reduce their fighting effort. Consequently, small groups of ants were found to fight hardest, showing that these ants strategically adjust their fighting efforts in a way that minimizes individual mortality risks, but not individual effort (see §3.1). Particularly, fighting effort is greatest in groups facing an imminent threat of being killed, also supporting the notion that individuals in defending groups face different incentives than those attacking.
Analogously, a recent study on meerkats (Suricata suricatta) established that behaviour in defences can be explained via individual cost/benefit analyses to a good extent in this cooperative breeder [31]. Again, those individuals who were likely to suffer the greatest costs in terms of status and reproductive access should have a defence fail, in this case high-ranking reproductive males invested most in group defence.
A recent review of 42 studies on non-human primates made similar observations [32]. It found that individual readiness to engage in violent conflicts with other groups largely depends on the necessity of food- and mate-defence as between-sex factors and on rank and reproductive access as within-sex factors.
In summary, these studies show that in many non-human animals participation in intergroup conflict depends strongly on a number of factors that influence direct fitness, indicating that selection for direct fitness benefits plays an important role in this context.
(ii). Behavioural differences between attacks and defences
Unambiguous evidence can also be found regarding the existence of differences in behavioural strategies for attacks and defences. Chimpanzees, for example, have been found to condition their participation in aggressive encounters with out-group members on their numerical superiority [33]. They only attack when they outnumber the enemy to a sufficient degree, a tactic that substantially reduces risks for individual attackers. A similar strategy was found in ants (Formica xerophila; [34]). Like F. rufa, F. xerophila assess their group's resource holding potential and adjust their efforts accordingly. Moreover, their fighting potential was found to increase nonlinearly with group size because they use a strategy of singling out and collectively attacking lone enemies.
For defences, on the other hand, a comprehensive analysis of aggregate data on 135 primate species recently established an illuminating relationship between residence patterns and the ability to hold a territory [35]. In species where effective territorial defence plays an important role for survival and reproductive success, protection of the home range is achieved better by species whose dominant sex is philopatric, like in many pre-modern human societies [36]. Complementing observations are reported in a recent experimental study with capuchins (Cebus capucinus, [37]). The likelihood that an individual capuchin would flee from a conflict with intruding out-group members was found to increase with the size of the in-group, indicating an individual tendency to free-ride on collective defensive efforts. However, control for the location of the conflict showed that this likelihood was substantially reduced (by 91%) when the conflict took place close to the centre of the home range compared to its more distant border zone. Thus, in this study, individual readiness to incur risks during defence seems to increase with the relevance of the resources at stake.
(b). Comparative studies on chimpanzees and humans
Two recent comparative studies provide further instructive insights into aggression patterns in humans and chimpanzees. The first study compared death rates caused by intergroup aggression in five chimpanzee and 12 hunter–gatherer communities and of intragroup killing in two chimpanzee and one hunter–gatherer group [38]. Although chimpanzees were found to show much higher levels of intragroup aggression, intergroup violence resulted in comparable rates of casualties in both species. This study's results are limited because of small sample size and high variance within the sample. However, they suggest the interesting question of why intergroup conflicts might have influenced the evolution of human cooperativeness while not leading to a comparable dynamic in chimpanzees.
The second comparative study focused on genetic instead of behavioural patterns and analysed between-group genetic differences in a large sample of contemporary human (hunter–gatherers and pre-industrialized societies) and chimpanzee groups [39]. It found that genetic between-group differentiation has about the same, small, magnitude in both species. Combined with the observation that intergroup conflict might have comparable dimensions in both species while in-group cooperativeness differs significantly, this study thus adds to the urgency of the question of why our species differ so remarkably in this respect.
(c). Anthropological evidence
Although most anthropologists now agree that violent conflict has been frequent in hunter–gatherer societies [6,36,40], its forms, causes and demographic impacts are subject to debate [4,41,42]. Comprehensive overviews of the different forms, motivations and outcomes of violent intergroup conflicts present in human (pre-)history can be found in the literature [6,43–45]. Notable are Keeley's descriptions of societies whose different subgroups constantly engage in violent conflicts, but do not aim at their enemies' total annihilation [6]. Also astonishing are cases of ‘tolerated intergroup theft’ [26]. In some historical cases, inferior groups actually ‘bought peace’ through ritualized danegeld payments to superior neighbours, mostly in the form of nubile women. In addition, the anthropological record is full of examples of between-group peace-making mechanisms [6,41,46]. The patterns of intergroup relations described in the cited surveys thus yield a highly differentiated picture. In addition, a number of instructive attempts at quantifying and analysing certain aspects of these patterns exist in the literature.
(i). Spoils of war
A recent paper [24] presents arguments and evidence in favour of the view that territorial expansion by violent means can actually result in a net loss of exploitable territory, challenging the idea that territory is one of the important public goods obtained through conflict. The logic is this: when two neighbouring groups of comparable fighting strength quarrel, this creates perilous buffer-zones between their territories which cannot be used safely for foraging until the conflict is settled or one group ultimately defeated. As it is unlikely that conflicts between two groups of comparable resource holding potential are decided quickly, accessible territory is thus decreased for quite some time. If the two groups are capable of foreseeing such a mutually disadvantageous stalemate, they might opt for peace in the first place.
Another study surveys the annual mortality rates owing to warfare in 20 simple societies [47], disentangling group-level benefits of raiding (territorial gains and increased deterrence) from direct benefits for the individual warriors (increased reproductive access, status and prestige). It finds that the prospect of direct benefits, explicitly excluding territorial gains, is positively correlated with mortality rates owing to warfare. This finding raises the suspicion that deterrence and territorial gains might be side effects, rather than primary goals, of intergroup aggression, at least in some cases. Complementary evidence is presented in a study which found that in some societies even peacetime meat distribution by hunters is not egalitarian but used strategically to raise coalitional support [48].
Also instructive are the results of two studies explicitly analysing the importance of women as potential objectives of raiding. One study comprising data from 44 South American societies [49] found that women represented the second most important reason for raids (retaliation was number one), with an average of approximately 0.6 women being captured per incidence of between-group violence (n = 187). Another study investigating the Chaco pueblo peoples of the late pre-Hispanic US southwest even finds evidence that the impact of ‘raiding for women’ was strong enough to result in biased sex ratios between neighbouring communities [50].
(ii). Status and reproductive access
Apart from being an important objective of raids, women have also been found to exert a more indirect influence. As known from the classic study by Chagnon [51], men who have earned the status of a well-proven warrior through killing enemies in combat enjoy increased reproductive success in the South American Yanomami culture (but see [52] for a counterexample). There is quite consistent evidence that male competition over reproductive access to females, not only of out-groups but also of the in-group, is linked with intergroup conflict. Residence patterns have been found to be associated with the frequency of between-group violence, with matrilocally residing societies experiencing lower rates of conflict than patrilocal ones [53]. The same qualitative observation has been made in a comparative study of humans and other primates [54]. Complementing these findings, a recent study on perceived attractiveness and status of males in another small-scale Amazonian society found that warriorship was strongly correlated with perceived attractiveness [55]. All these findings add to the impression that engaging in intergroup conflict might be a means for men to attain status which then translates into the direct fitness benefit of increased reproductive success [56].
(d). Studies on the psychology of human intergroup relations
Most laboratory and field studies on parochial altruism and in-group favouritism use monetarily incentivized decision experiments (see the electronic supplementary material, table S2, for a comprehensive list of relevant references). The strand of literature investigating in-group favouritism frequently analyses decision behaviour in pairwise interactions when subjects are informed about each other's group membership. These studies quite reliably find that subjects condition their decisions on group membership and usually allocate (positive) ‘goods’ mostly to in-group members and ‘bads’, i.e. negative goods, mostly to out-group members. Refined experimental designs, however, show that this pattern is not explained well by a simple notion of unconditional in-group love and out-group hate [57]. Two points are particularly noteworthy. First, most studies using this paradigm fail to establish actual out-group hate. While subjects do allocate more goods to in-group members, they still share significant amounts with out-group members. Second, when the experimental design assures that the recipients of allocations do not know the group membership of the allocator while the allocator does know that he/she is playing with an in-group member, in-group favouritism can disappear [58,59]. Revealing results were also obtained with respect to third-party punishment of allocators [60,61].
Another strand of literature uses games played by groups of subjects [62]. Various experimental designs can be found, including some that very closely resemble the conventional intergroup contest mechanism described above. The general finding is that, when comparing single groups who play variants of the n-person Prisoner's Dilemma in the absence of an out-group to paired groups in competition with each other, competing groups show elevated levels of in-group cooperation and, when available, use costly punishment to coerce in-group members into cooperating [63,64]. While this general finding seems directly in line with parochial altruism theories at first glance, again, elaborated experimental designs show that this pattern might be caused by more subtle motivations. In two recent studies, it was found that contributions to in-group cooperation were almost completely explained by the expectation of in-group reciprocity [65,66]. After control for this influence, no significant effect of intergroup competition remained. Furthermore, a number of studies found that when given the opportunity of in-group cooperation with no negative effect on the out-group or of between-group communication, subjects frequently chose to avoid conflict and even cooperated with out-groups [67–70]. Noteworthy differences in the decision behaviour of men and women in reaction to between-group competition have also been found [71–73].
6. Theoretical alternatives to parochial altruism
The theory of parochial altruism starts from a combination of the observed tendencies of humans to discriminate and cooperatively engage in violent aggression against out-group members and finds that there are conceivable evolutionary scenarios in which these two traits might coevolve. However, it leaves open the question of how these traits might have come into existence in the first place. This section briefly reviews answers proposed by three complementary theories. An overarching future theory of human intergroup conflict psychology will certainly benefit from trying to integrate these with parochial altruism.
(a). Group reciprocity as a heuristic
While there are numerous theoretical approaches to the question of why humans so readily condition behaviour on group membership, only the ‘group heuristic model’ [74] will be highlighted here, because it is an explicitly evolutionary account. Put simply, the group heuristic model states that in-group bias is not the result of an unconditional strategy of group discrimination. Rather, in-group membership is taken to represent an indicator of a higher probability of repeated encounter and a higher probability of the availability of reputational mechanisms. Thus, it is argued that interactions with in-group members are more likely to be perceived as being repeated with high probability, triggering strategies of direct reciprocity, and that individuals will be more sensitive to reputational concerns in these interactions, because indirect reciprocity works much better in small groups [75]. Yamagishi and co-workers [58,59] were able to show by experimental manipulation that when reputational concerns in one-shot games with in-group members are credibly removed, in-group bias can disappear. It is a matter of further research, however, to establish whether the group heuristic model, in comparison to other accounts proposed, captures all relevant motivations for in-group favouritism.
(b). The chimpanzee model
The second complementary approach focuses particularly on groupwise interactions, analysing them with respect to potential direct fitness benefits. Based on observations that chimpanzees and other species use tactics which minimize individual risks for attackers, Wrangham et al. [23,38] have proposed and tested an ‘imbalance-of-power hypothesis’, later labelled ‘the chimpanzee model’. It states that offensive coalitionary violence can be favoured by natural selection whenever the risks encountered by attackers are outweighed by sufficient prospects of individual benefits. Two points are noteworthy. First, the chimpanzee model takes into account that incentives to participate can differ substantially between individuals and is thus able to explain why some individuals will be more likely to engage in intergroup aggression than others, yielding testable predictions about individual behaviour and even differences between species. Second, the chimpanzee model does not require any form of altruism to explain why we observe groups fighting for a shared goal, because this common goal is argued to be pursued as a consequence of individual fitness maximization (see also [76]). However, this second point reveals a blind spot of the chimpanzee model: we do observe altruistic self-sacrifice in war—at least in humans [25]. This remains unexplained by the chimpanzee model.
(c). The male warrior hypothesis
A third alternative perspective on individual motivations for engaging in intergroup aggression is the ‘male warrior hypothesis’ proposed by van Vugt & co-workers [71,72]. It builds on the observations that it is mostly men who engage in violent intergroup conflicts, that proven warriors seem to be perceived as more attractive and might have increased reproductive success, and that in laboratory experiments men frequently react differently to the presence of intergroup contest than women. It assumes that intergroup conflict is an arena of male reproductive interests to a large extent. Men might not use war only to gain additional reproductive access to women of the out-group but also as an opportunity to display their physical, tactical and leadership qualities to women of the in-group. Women, accordingly, might condition their mate choice on merits earned by in-group males through aggressive behaviour in conflicts. A recent theoretical study has demonstrated that this logic is consistent [22], showing that even in the absence of public goods created through raiding female selection for male heroism and bravery in war can result in stable incentives for males to risk their individual well-being by engaging in warfare.
While the male warrior hypothesis is supported by a number of archive, field and experimental studies, it is still subject to debate. Open questions include: (i) why are sex differences found in some laboratory experiments and field studies but absent in others? (ii) do females prefer warriorship independent of its results? and (iii) is female selection for male peaceableness also conceivable?
7. Conclusion
The aim of this review was to provide a first evaluation of the explanatory power of the theory of parochial altruism and to point out urgent questions yet unanswered. It has been argued that parochial altruism theory in its current state is consistent but limited in a number of important respects. Most importantly, future work needs to analyse which selective mechanisms are at work in the standard models of parochial altruism. Candidates include individual selection for (i) mutually beneficial behaviour like group defence and (ii) selfish behaviour like status competition and mate acquisition, and furthermore, (iii) kin selection for discriminatory altruism and spite. Second, we urgently need a better understanding of (i) how individual incentives are affected by theoretical assumptions about the structure of intergroup conflicts and (ii) the nature of the actual incentives different individuals face in the various forms in which intergroup conflict manifests. Table 1 presents a systematic overview of the literature cited in this reivew; also see tables S1 and S2 in the electronic supplementary material for a more comprehensive overview of relevant references.
Table 1.
Systematic overview of literature cited; also see the electronic supplementary material, table S2.
| review section | relevant references |
|---|---|
| theoretical studies on parochial altruism | [1,3–5,14]; also see the electronic supplementary material, table S1 |
| interpretation of the models | [7,8,10–13,15–21] |
| extension 1: peaceful conflict solutions | [1,6,41,42,67–70] |
| extension 2: individual benefits and differences | [6,21–23,28–32,35,47,48,57–61,65,66,74] |
| extension 3: attacks and defences | [23–26,33,34,37] |
| extension 4: scale of warfare | [3,6,38–40,43–45,47,53] |
| extension 5: sex differences | [22,27,36,49–56,71–73,76] |
Supplementary Material
Supplementary Material
Acknowledgements
I thank Max Albert, Robert Böhm, Matthias Greiff, Carl Salk, Eckart Voland and two anonymous reviewers for valuable comments on earlier versions of this review. Additionally, I thank Robin I.M. Dunbar, Nir Halevy, Shaun Hargreaves Heap and Mikael Puurtinen for helpful answers to my queries about their work.
References
- 1.Choi J-K, Bowles S. 2007. The coevolution of parochial altruism and war. Science 318, 636–640. ( 10.1126/science.1144237) [DOI] [PubMed] [Google Scholar]
- 2.Darwin C. 1871. The descent of man and selection in relation to sex. London, UK: John Murray. [Google Scholar]
- 3.Bowles S. 2006. Group competition, reproductive leveling, and the evolution of human altruism. Science 314, 1569–1572. ( 10.1126/science.1134829) [DOI] [PubMed] [Google Scholar]
- 4.Bowles S. 2009. Did warfare among ancestral hunter-gatherers affect the evolution of human social behaviors? Science 324, 1293–1298. ( 10.1126/science.1168112) [DOI] [PubMed] [Google Scholar]
- 5.García J, van den Bergh JC. 2011. Evolution of parochial altruism by multilevel selection. Evol. Hum. Behav. 32, 277–287. ( 10.1016/j.evolhumbehav.2010.07.007) [DOI] [Google Scholar]
- 6.Keeley LH. 1996. War before civilization. The myth of the peaceful savage. New York, NY: Oxford University Press. [Google Scholar]
- 7.West SA, Griffin AS, Gardner A. 2007. Evolutionary explanations for cooperation. Curr. Biol. 17, R661–R672. ( 10.1016/j.cub.2007.06.004) [DOI] [PubMed] [Google Scholar]
- 8.West SA, Griffin AS, Gardner A. 2007. Social semantics: altruism, cooperation, mutualism, strong reciprocity and group selection. J. Evol. Biol. 20, 415–432. ( 10.1111/j.1420-9101.2006.01258.x) [DOI] [PubMed] [Google Scholar]
- 9.Hamilton WD. 1964. The genetical evolution of social behaviour. I&II. J. Theor. Biol. 7, 1–52. ( 10.1016/0022-5193(64)90038-4) [DOI] [PubMed] [Google Scholar]
- 10.Gardner A, West SA. 2004. Spite and the scale of competition. J. Evol. Biol. 17, 1195–1203. () [DOI] [PubMed] [Google Scholar]
- 11.Leimar O, Hammerstein P. 2010. Cooperation for direct fitness benefits. Phil. Trans. R. Soc. B 365, 2619–2626. ( 10.1098/rstb.2010.0116) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gardner A, West SA. 2014. Inclusive fitness: 50 years on. Phil. Trans. R. Soc. B 369, 20130356 ( 10.1098/rstb.2013.0356) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Konrad KA, Morath F. 2012. Evolutionarily stable in-group favoritism and out-group spite in intergroup conflict. J. Theor. Biol. 306, 61–67. ( 10.1016/j.jtbi.2012.04.013) [DOI] [PubMed] [Google Scholar]
- 14.Fu F, Tarnita CE, Christakis NA, Wang L, Rand DG, Nowak MA. 2012. Evolution of in-group favoritism. Sci. Rep. 2, 460 ( 10.1038/srep00460) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kerr B, Godfrey-Smith P. 2002. Individualist and multi-level perspectives on selection in structured populations. Biol. Phil. 17, 477–517. ( 10.1023/A:1020504900646) [DOI] [Google Scholar]
- 16.Lehmann L, Keller L, West SA, Roze D. 2007. Group selection and kin selection: two concepts but one process. Proc. Natl Acad. Sci. USA 104, 6736–6739. ( 10.1073/pnas.0700662104) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.van Veelen M, García J, Sabelis MW, Egas M. 2012. Group selection and inclusive fitness are not equivalent; the Price equation vs. models and statistics. J. Theor. Biol. 299, 64–80. ( 10.1016/j.jtbi.2011.07.025) [DOI] [PubMed] [Google Scholar]
- 18.Rusch H. 2014. A threshold for biological altruism in public goods games played in groups including kin. MAGKS Jt. Discuss. Pap. Ser. Econ. 29–2014.
- 19.Ohtsuki H. 2014. Evolutionary dynamics of n-player games played by relatives. Phil. Trans. R. Soc. B 369, 20130359 ( 10.1098/rstb.2013.0359) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Leigh EJJ. 2010. The group selection controversy. J. Evol. Biol. 23, 6–19. () [DOI] [PubMed] [Google Scholar]
- 21.Gavrilets S, Fortunato L. 2014. A solution to the collective action problem in between-group conflict with within-group inequality. Nat. Commun. 5, 3526 ( 10.1038/ncomms4526) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lehmann L, Feldman MW. 2008. War and the evolution of belligerence and bravery. Proc. R. Soc. B 275, 2877–2885. ( 10.1098/rspb.2008.0842) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wrangham RW, Glowacki L. 2012. Intergroup aggression in chimpanzees and war in nomadic hunter-gatherers. Hum. Nat. 23, 5–29. ( 10.1007/s12110-012-9132-1) [DOI] [PubMed] [Google Scholar]
- 24.Kelly RC. 2005. The evolution of lethal intergroup violence. Proc. Natl Acad. Sci. USA 102, 15 294–15 298. ( 10.1073/pnas.0505955102) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rusch H. 2013. Asymmetries in altruistic behavior during violent intergroup conflict. Evol. Psychol. 11, 973–993. [PubMed] [Google Scholar]
- 26.Rusch H. 2014. The two sides of warfare. Hum. Nat. 25, 359–377. ( 10.1007/s12110-014-9199-y) [DOI] [PubMed] [Google Scholar]
- 27.Balliet D, Li NP, Macfarlan SJ, van Vugt M. 2011. Sex differences in cooperation: a meta-analytic review of social dilemmas. Psychol. Bull. 137, 881–909. ( 10.1037/a0025354) [DOI] [PubMed] [Google Scholar]
- 28.Radford AN. 2008. Duration and outcome of intergroup conflict influences intragroup affiliative behaviour. Proc. R. Soc. B 275, 2787–2791. ( 10.1098/rspb.2008.0787) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Radford AN. 2011. Preparing for battle? Potential intergroup conflict promotes current intragroup affiliation. Biol. Lett. 7, 26–29. ( 10.1098/rsbl.2010.0507) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Batchelor TP, Briffa M. 2011. Fight tactics in wood ants: individuals in smaller groups fight harder but die faster. Proc. R. Soc. B 278, 3243–3250. ( 10.1098/rspb.2011.0062) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mares R, Young AJ, Clutton-Brock TH. 2012. Individual contributions to territory defence in a cooperative breeder: weighing up the benefits and costs. Proc. R. Soc. B 279, 3989–3995. ( 10.1098/rspb.2012.1071) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kitchen D, Beehner J. 2007. Factors affecting individual participation in group-level aggression among non-human primates. Behaviour 144, 1551–1581. ( 10.1163/156853907782512074) [DOI] [Google Scholar]
- 33.Wilson ML, Britton NF, Franks NR. 2002. Chimpanzees and the mathematics of battle. Proc. R. Soc. Lond. B 269, 1107–1112. ( 10.1098/rspb.2001.1926) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tanner CJ. 2006. Numerical assessment affects aggression and competitive ability: a team-fighting strategy for the ant Formica xerophila. Proc. R. Soc. B 273, 2737–2742. ( 10.1098/rspb.2006.3626) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Willems EP, Hellriegel B, van Schaik CP. 2013. The collective action problem in primate territory economics. Proc. R. Soc. B 280, 20130081 ( 10.1098/rspb.2013.0081) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ember CR. 1978. Myths about hunter-gatherers. Ethnology 17, 439–448. ( 10.2307/3773193) [DOI] [Google Scholar]
- 37.Crofoot MC, Gilby IC. 2012. Cheating monkeys undermine group strength in enemy territory. Proc. Natl Acad. Sci. USA 109, 501–505. ( 10.1073/pnas.1115937109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wrangham RW, Wilson ML, Muller MN. 2006. Comparative rates of violence in chimpanzees and humans. Primates 47, 14–26. ( 10.1007/s10329-005-0140-1) [DOI] [PubMed] [Google Scholar]
- 39.Langergraber K, Schubert G, Rowney C, Wrangham RW, Zommers Z, Vigilant L. 2011. Genetic differentiation and the evolution of cooperation in chimpanzees and humans. Proc. R. Soc. B 278, 2546–2552. ( 10.1098/rspb.2010.2592) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ember CR, Ember M. 1992. Resource unpredictability, mistrust, and war: a cross-cultural study. J. Confl. Resolut. 36, 242–262. ( 10.1177/0022002792036002002) [DOI] [Google Scholar]
- 41.Boehm C. 2012. Ancestral hierarchy and conflict. Science 336, 844–847. ( 10.1126/science.1219961) [DOI] [PubMed] [Google Scholar]
- 42.Fry DP, Söderberg P. 2013. Lethal aggression in mobile forager bands and implications for the origins of war. Science 341, 270–273. ( 10.1126/science.1235675) [DOI] [PubMed] [Google Scholar]
- 43.van der Dennen JM, Johan MG. 2002. (Evolutionary) theories of warfare in preindustrial (foraging) societies. Endocrinol. Lett. 23, 55–65. [PubMed] [Google Scholar]
- 44.Liddle JR, Shackelford TK, Weekes-Shackelford VA. 2012. Why can't we all just get along? Rev. Gen. Psychol. 16, 24–36. ( 10.1037/a0026610) [DOI] [Google Scholar]
- 45.Tooby J, Cosmides L. 2010. Groups in mind. In Human morality and sociality. Evolutionary and comparative perspectives (ed. Høgh-Olesen H.), pp. 191–234. Basingstoke, UK: Palgrave Macmillan. [Google Scholar]
- 46.Fry DP. 2012. Life without war. Science 336, 879–884. ( 10.1126/science.1217987) [DOI] [PubMed] [Google Scholar]
- 47.Glowacki L, Wrangham RW. 2013. The role of rewards in motivating participation in simple warfare. Hum. Nat. 24, 444–460. ( 10.1007/s12110-013-9178-8) [DOI] [PubMed] [Google Scholar]
- 48.Patton JQ. 2005. Meat sharing for coalitional support. Evol. Hum. Behav. 26, 137–157. ( 10.1016/j.evolhumbehav.2004.08.008) [DOI] [Google Scholar]
- 49.Walker RS, Bailey DH. 2013. Body counts in lowland South American violence. Evol. Hum. Behav. 34, 29–34. ( 10.1016/j.evolhumbehav.2012.08.003) [DOI] [Google Scholar]
- 50.Kohler TA, Kramer Turner K. 2006. Raiding for women in the pre-Hispanic northern pueblo southwest? Curr. Anthropol. 47, 1035–1045. ( 10.1086/508697) [DOI] [Google Scholar]
- 51.Chagnon NA. 1988. Life histories, blood revenge, and warfare in a tribal population. Science 239, 985–992. ( 10.1126/science.239.4843.985) [DOI] [PubMed] [Google Scholar]
- 52.Beckerman S, Erickson PI, Yost J, Regalado J, Jaramillo L, Sparks C, Iromenga M, Long K. 2009. Life histories, blood revenge, and reproductive success among the Waorani of Ecuador. Proc. Natl Acad. Sci. USA 106, 8134–8139. ( 10.1073/pnas.0901431106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Adams DB. 1983. Why there are so few women warriors. Cross-Cult. Res. 18, 196–212. ( 10.1177/106939718301800302) [DOI] [Google Scholar]
- 54.Manson JH, Wrangham RW. 1991. Intergroup aggression in chimpanzees and humans. Curr. Anthropol. 32, 369–390. ( 10.1086/203974) [DOI] [Google Scholar]
- 55.Escasa M, Gray PB, Patton JQ. 2010. Male traits associated with attractiveness in Conambo, Ecuador. Evol. Hum. Behav. 31, 193–200. ( 10.1016/j.evolhumbehav.2009.09.008) [DOI] [Google Scholar]
- 56.von Rueden C, Gurven M, Kaplan H. 2011. Why do men seek status? Fitness payoffs to dominance and prestige. Proc. R. Soc. B 278, 2223–2232. ( 10.1098/rspb.2010.2145) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Buttelmann D, Böhm R. 2014. The ontogeny of the motivation that underlies in-group bias. Psychol. Sci. 25, 921–927. ( 10.1177/0956797613516802) [DOI] [PubMed] [Google Scholar]
- 58.Mifune N, Hashimoto H, Yamagishi T. 2010. Altruism toward in-group members as a reputation mechanism. Evol. Hum. Behav. 31, 109–117. ( 10.1016/j.evolhumbehav.2009.09.004) [DOI] [Google Scholar]
- 59.Yamagishi T, Mifune N. 2009. Social exchange and solidarity: in-group love or out-group hate? Evol. Hum. Behav. 30, 229–237. ( 10.1016/j.evolhumbehav.2009.02.004) [DOI] [Google Scholar]
- 60.Shinada M, Yamagishi T, Ohmura Y. 2004. False friends are worse than bitter enemies. Evol. Hum. Behav. 25, 379–393. ( 10.1016/j.evolhumbehav.2004.08.001) [DOI] [Google Scholar]
- 61.Bernhard H, Fischbacher U, Fehr E. 2006. Parochial altruism in humans. Nature 442, 912–915. ( 10.1038/nature04981) [DOI] [PubMed] [Google Scholar]
- 62.Bornstein G. 2003. Intergroup conflict: individual, group, and collective interests. Pers. Soc. Psychol. Rev. 7, 129–145. ( 10.1207/S15327957PSPR0702_129-145) [DOI] [PubMed] [Google Scholar]
- 63.Gneezy A, Fessler DMT. 2011. Conflict, sticks and carrots: war increases prosocial punishments and rewards. Proc. R. Soc. B 279, 219–223. ( 10.1098/rspb.2011.0805) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sääksvuori L, Mappes T, Puurtinen M. 2011. Costly punishment prevails in intergroup conflict. Proc. R. Soc. B 278, 3428–3436. ( 10.1098/rspb.2011.0252) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Koopmans R, Rebers S. 2009. Collective action in culturally similar and dissimilar groups: an experiment on parochialism, conditional cooperation, and their linkages. Evol. Hum. Behav. 30, 201–211. ( 10.1016/j.evolhumbehav.2009.01.003) [DOI] [Google Scholar]
- 66.Rebers S, Koopmans R. 2012. Altruistic punishment and between-group competition. Hum. Nat. 23, 173–190. ( 10.1007/s12110-012-9136-x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Halevy N, Bornstein G, Sagiv L. 2008. ‘In-group love’ and ‘out-group hate’ as motives for individual participation in intergroup conflict. Psychol. Sci. 19, 405–411. ( 10.1111/j.1467-9280.2008.02100.x) [DOI] [PubMed] [Google Scholar]
- 68.Halevy N, Weisel O, Bornstein G. 2012. ‘In-group love’ and ‘out-group hate’ in repeated interaction between groups. J. Behav. Decis. Making 25, 188–195. ( 10.1002/bdm.726) [DOI] [Google Scholar]
- 69.Pan XS, Houser D. 2013. Cooperation during cultural group formation promotes trust towards members of out-groups. Proc. R. Soc. B 280, 20130606 ( 10.1098/rspb.2013.0606) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Leibbrandt A, Sääksvuori L. 2012. Communication in intergroup conflicts. Eur. Econ. Rev. 56, 1136–1147. ( 10.1016/j.euroecorev.2012.05.003) [DOI] [Google Scholar]
- 71.van Vugt M, de Cremer D, Janssen DP. 2007. Gender differences in cooperation and competition. Psychol. Sci. 18, 19–23. ( 10.1111/j.1467-9280.2007.01842.x) [DOI] [PubMed] [Google Scholar]
- 72.McDonald MM, Navarrete CD, van Vugt M. 2012. Evolution and the psychology of intergroup conflict. Phil. Trans. R. Soc. B 367, 670–679. ( 10.1098/rstb.2011.0301) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Pan XS, Houser D, Rankin D. 2011. Competition for trophies triggers male generosity. PLoS ONE 6, e18050 ( 10.1371/journal.pone.0018050) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Yamagishi T, Kiyonari T. 2000. The group as the container of generalized reciprocity. Soc. Psychol. Q. 63, 116–132. ( 10.2307/2695887) [DOI] [Google Scholar]
- 75.Roberts G. 2008. Evolution of direct and indirect reciprocity. Proc. R. Soc. B 275, 173–179. ( 10.1098/rspb.2007.1134) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Díaz-Muñoz SL, DuVal EH, Krakauer AH, Lacey EA. 2014. Cooperating to compete. Anim. Behav. 88, 67–78. ( 10.1016/j.anbehav.2013.11.008) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.