

Abstract
Optogenetics has become an emerging technique for neuroscience investigations owing to the great spatiotemporal precision and the target selectivity it provides. Here we extend the optogenetic strategy to GABAA receptors (GABAARs), the major mediators of inhibitory neurotransmission in the brain. We generated a light-regulated GABAA receptor (LiGABAR) by conjugating a photoswitchable tethered ligand (PTL) onto a mutant receptor containing the cysteine-substituted α1-subunit. The installed PTL can be advanced to or retracted from the GABA-binding pocket with 500 and 380 nm light, respectively, resulting in photoswitchable receptor antagonism. In hippocampal neurons, this LiGABAR enabled a robust photoregulation of inhibitory postsynaptic currents. Moreover, it allowed reversible photocontrol over neuron excitation in response to presynaptic stimulation. LiGABAR thus provides a powerful means for functional and mechanistic investigations of GABAAR-mediated neural inhibition.
Optogenetics, an integrated strategy that typically employs microbial opsins to overpower neuronal excitability, has become a revolutionary technique for neuroscience investigations.1 This technique enables light to remotely manipulate neuronal activity with high spatial and temporal precision. Moreover, it allows photocontrol over a defined neuron type within an intact tissue or even a behaving organism.1 However, microbial opsins are functionally distinct from neuronal signaling proteins (e.g., neurotransmitter receptors for synaptic transmission) and hence are constrained to decode neural circuits at the cellular level. To gain molecular insights into neural functions, methods that allow optogenetic control over a specific signaling mediator are highly desired.2 Here we present a chemical-genetic approach that enables photocontrol over inhibitory neurotransmission mediated by the ionotropic type-A GABA receptors (GABAARs). Our approach allows light to specifically modulate a defined subtype of GABAAR, which will help elucidate the unique functions of the receptor within a complex neural network.
The GABAARs, a group of neurotransmitter-gated chloride-permeable channels, are therapeutic targets in psychiatric disorders3 and epilepsy4 owing to their inhibitory control over neuronal excitation. They are also targets for many drugs of abuse, including alcohol, barbiturates, and benzodiazepines.3,5,6 The GABAARs are heteropentameric assemblies composed of two α, two β, and one tertiary subunit (usually γ or δ, Figure 1a).3,5 Among these components, the α-subunit is key in determining receptor localization and gating kinetics3,5,7,8 and, together with the β subunit, forms the GABA-binding site.3 There are six distinct α-isoforms expressed heterogeneously in different neuron types and brain regions.3,5,6 Adding to this complexity, a neuron can express multiple α-isoforms that are differentially distributed in subcellular compartments.7 These findings suggest that each α-isoform has unique roles in neuronal signaling, and understanding their individual functions will provide key insights into GABAAR-associated disorders and therapeutics.
Figure 1.

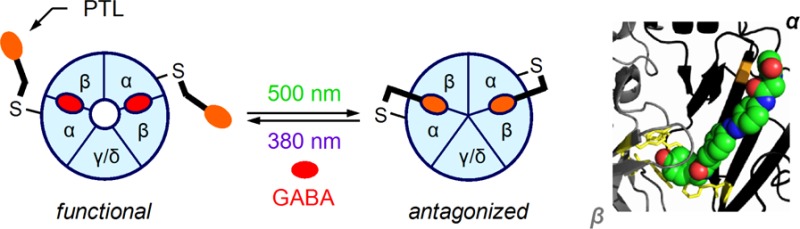
Engineering of the light-regulated GABAA receptor (LiGABAR). (a) A LiGABAR is generated by conjugating a photoswitchable tethered ligand (PTL) onto a receptor comprising the cysteine-substituted α-subunits (top). In the case of photoswitchable antagonism (bottom), the installed PTL reversibly isomerizes between two states in response to two different wavelengths of light, with one preventing and the other enabling GABA binding (and the subsequent gating of the transmembrane channel). (b) The structure and photochemistry of MAM-6 (the prototype PTL). (c–e) Identification of MAM-6 attachment sites in the α1 subunit. (c) Distribution of the tested cysteine-substituted residues (orange; side chain in sticks) in a homology model of α1β2.24 The GABA-binding site is indicated by a docked muscimol (red). (d) Representative traces showing reversible photoregulation of GABA-elicited currents by the tethered MAM-6. Mutant = α1(S68C). (e) Photoregulation of mutant receptors after MAM-6 conjugation. Each mutant was coexpressed with the wild-type β2 in Xenopus oocytes. The photoregulation index (mean ± SEM) was measured at 3 μM GABA, −80 mV. A ratio of 1 indicates no photosensitivity of the tested receptor.
We have engineered a light-regulated GABAA receptor (LiGABAR) by covalently conjugating a photoswitchable tethered ligand (PTL) onto a receptor containing the cysteine-substituted α-subunits (Figure 1a). The PTL comprises a sulfhydryl-reactive maleimide group, a photoswitchable azobenzene core, and a ligand for the GABA-binding site (Figure 1b). It reversibly isomerizes between cis (twisted) and trans (extended) configurations upon the illumination of 380 and 500 nm light, respectively. Once installed near the GABA-binding pocket, the PTL can retract or advance its ligand via photoisomerization, leading to reversible photocontrol of the receptor activity. The wild-type GABAARs display very few extracellular cysteines which are either involved in disulfide formation or are distant from the GABA-binding site. Hence, the PTL will exert its function only on an engineered receptor with appropriately positioned cysteines, ensuring the target specificity of photocontrol within a heterogeneous population of GABAARs.
Our prototype PTL was designed based on previously published GABAAR probes in which the pharmacophore muscimol is conjugated to a fluorophore or biotin via a 6-aminohexanoyl spacer.9 The resulting PTL, named MAM-6, comprises a maleimide, an azobenzene, and a muscimol linked through a 6-carbon spacer (Figure 1b). Although muscimol can work as a photoaffinity probe for GABAARs,10 the wavelengths for MAM-6 photoisomerization are >100 nm longer than that for muscimol photolysis and are thus incapable of triggering this side reaction. We screened MAM-6 attachment sites for the α1-containing GABAAR, the most abundant α-isoform in the brain. The screening was carried out in Xenopus oocytes expressing the engineered receptors. Residues that are predicted to face the GABA-binding site were chosen for cysteine substitution (Figure 1c). Previous studies suggested that cysteine substitutions at these locations allow the mutant receptors to express normally and remain sensitive to GABA and allosteric modulators (e.g., pentobarbital and benzodiazepines).11,12 These mutants are thus ideal candidates for MAM-6 conjugation.
Although muscimol is ordinarily an agonist for GABAARs, the mutant receptors were not activated by either 380 or 500 nm light after MAM-6 conjugation. Instead, MAM-6 inhibited some of the mutant receptors in a light-dependent manner, with 500 nm (trans MAM-6) causing a greater decrement in GABA-elicited current than 380 nm (cis MAM-6) did (Figure 1d and e and Supplementary Figure S1). This phenomenon, i.e., photoswitchable antagonism mediated by an agonist-based PTL, is similar to an observation reported previously for the nicotinic acetylcholine receptors,13 possibly caused by the PTL disrupting conformational changes required for receptor activation.13,14 To quantitatively evaluate each attachment site, we indexed receptor photosensitivity as the ratio of GABA-elicited current in 380 nm vs in 500 nm (I380/I500). MAM-6 caused a strong photoregulation when attached at sites closer to the entrance of the GABA-binding pocket (Figure 1c and e). We selected α1(T125C) as optimal because it allows MAM-6 to substantially inhibit the receptor in 500 nm while allowing the receptor to function normally in 380 nm (Supplementary Table S1 and Figure S1). Moreover, cysteine substitution at this location does not alter the receptor’s GABA sensitivity (ref (11) and Supplementary Table S1). These properties make α1(T125C) and its resulting LiGABAR preferable for neurophysiological investigations.
We next focused on α1(T125C) and investigated the structural requirements for a PTL to operate at this conjugation site. We changed the ligand and spacer moieties of MAM-6 and indexed the photoregulation effects in HEK293T cells (at 10 μM GABA; Figure 2a). The negative charge of muscimol (at physiological pH) is involved in receptor binding. Consistent with this mechanism, replacing muscimol with a neutral analogue 4-hydroxybenzylamine reduced the photoregulation effect (I380/I500 = 2.5 ± 0.3 and 1.7 ± 0.1, n = 6 and 3 for MAM-6 and MAB-6, respectively; Figure 2a). Removing the 6-carbon spacer of MAM-6 suppressed photoregulation nearly completely (MAM-0; I380/I500 = 1.3 ± 0.1, n = 4; Figure 2a), suggesting that this spacer is important for delivering muscimol into the GABA-binding pocket. Surprisingly, substituting muscimol with 4-hydroxybenzylamine in MAM-0 boosted photoregulation, giving an effect greater than that by MAM-6 (MAB-0; I380/I500 = 3.4 ± 0.4, n = 7; Figure 2a).
Figure 2.
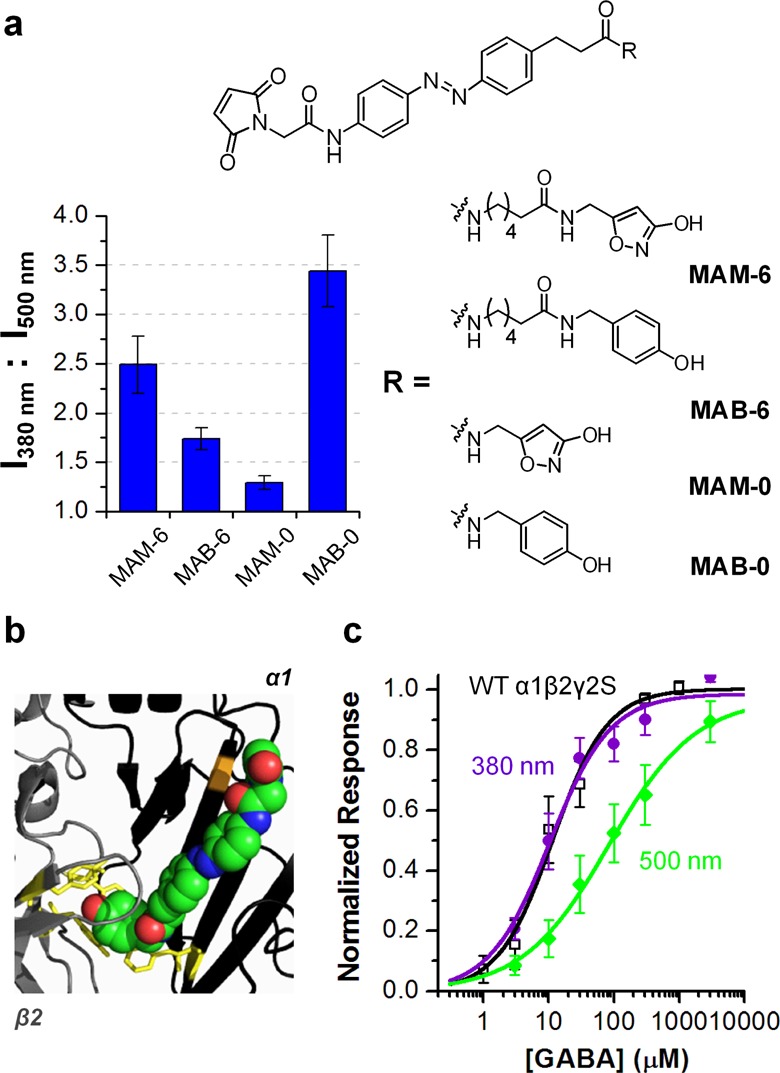
Characterizations for the α1(T125C)-based LiGABAR. (a) Structure-activity investigation of the PTL modules. Photosensitivity of each conjugated α1(T125C)β2γ2S was indexed at 10 μM GABA. n = 3–7. (b) A representative docking pose of trans MAB-0 (spheres) in a homology model of α1(T125C)β2 complex. A positional constraint was applied to mimic the tethering of trans MAB-0 at α1(T125C) (orange). Residues of the aromatic box (α1Phe64, β2Tyr97, β2Tyr157, and β2Tyr205) are shown as yellow sticks. (c) Dose-response curves for the wild-type α1β2γ2S (black) and MAB-0 conjugated α1(T125C)β2γ2S under 380 nm (purple) and 500 nm (green) illumination. n = 3 for the wild-type and 4 for the conjugated receptor. Data are presented as mean ± SEM. Recordings were carried out in HEK293T cells held at −70 mV.
The strong photoregulation by MAB-0 suggests a favorable interaction between its ligand moiety and the GABA-binding pocket that is specific for the trans configuration. The GABA-binding pocket is lined by several aromatic residues (i.e., the aromatic box),14,15 and these residues may interact with 4-hydroxybenzylamide, enabling it to compete with GABA. This hypothesis was further supported by the docking of trans MAB-0 in a homology model. The result suggested that trans MAB-0 spans the distance between the attachment site and the binding pocket, delivering its terminal phenol group to the aromatic box (Figure 2b). Trans-to-cis isomerization allows the PTL to twist, which withdraws the ligand and subsequently relieves receptor antagonism. Consistent with this working model, the EC50 of this LiGABAR increased 8-fold when the light was switched from 380 to 500 nm (10.2 ± 2.6 μM and 84.0 ± 19.3 μM, respectively, n = 4 for each; Figure 2c). In addition, the EC50 of the conjugated receptor in 380 nm light was the same as that of the wild-type (11.4 ± 2.2 μM, n = 3; Figure 2c). This LiGABAR thus provides an ideal system in which the receptor can be reversibly switched between normal and inhibited states by two different wavelengths of light. These findings also suggest that a typical agonist/antagonist is not mandatory for an effective PTL. When the scaffold is tethered at an appropriate position, a molecule that potentially interacts with the GABA-binding pocket could be a potent ligand, which opens a door for future PTL design. Moreover, because the GABA-binding site is highly homologous among the GABAAR family, this strategy may be applicable to other α-isoforms to expand the toolkit.
Cis-to-trans isomerization of azobenzenes can be driven by 500 nm light or will occur spontaneously in darkness through thermal relaxation (Figure 1b and ref (16)). The rate of thermal relaxation for a cis azobenzene is dependent on its chemical structure and the local environment.16 Thermal relaxation of MAB-0 (tethered to α1(T125C)) was measured by a functional assay illustrated in Figure 3. GABA-elicited current was measured in HEK293T cells every 2 min in darkness after an initial response under 5 s of 380 nm illumination. The current decreased slowly during the dark period but was fully restored by a subsequent illumination of 380 nm light, suggesting that the observed current reduction arose from thermal relaxation of cis MAB-0 (Figure 3a). We plotted the normalized light-sensitive current over time and fitted the data with a single-exponential decay (Figure 3b). The half-life of cis MAB-0 was calculated to be 23.6 ± 2.7 min (n = 5) in darkness. The slow thermal relaxation of cis PTL makes LiGABAR bistable, allowing the receptor to stay in the uninhibited state for minutes after a brief pulse of 380 nm light. This feature provides a way to minimize possible adverse effects of light during the course of an experiment.
Figure 3.
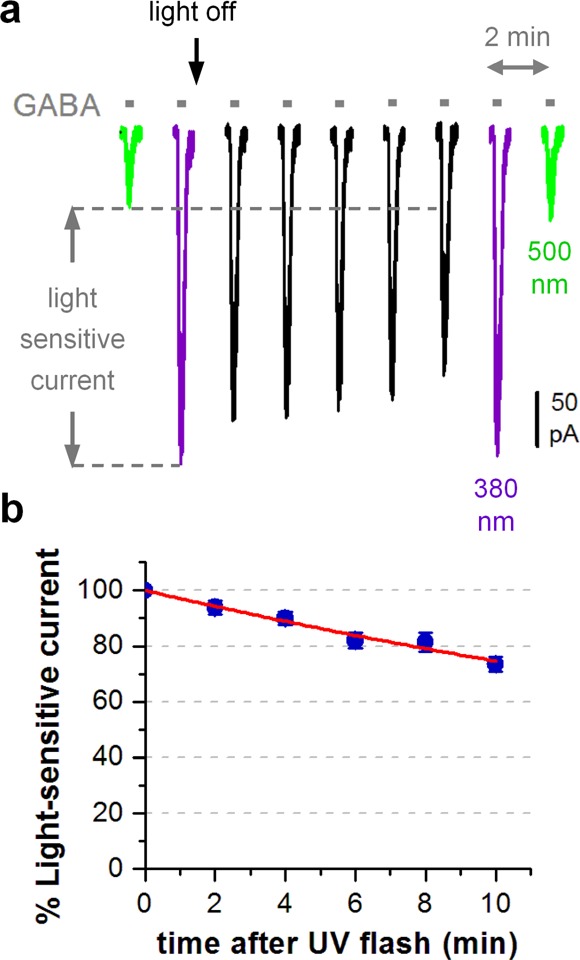
Thermal relaxation of the tethered MAB-0. (a) A representative cell (with MAB-0 conjugated α1(T125C)β2γ2S) showing the slow current reduction in darkness after an initial response measured in 380 nm. [GABA] = 10 μM. (b) Group data (mean ± SEM, n = 5) showing the time course of thermal relaxation, plotted as changes in the normalized light-sensitive current component (defined in panel a) and fitted with a single-exponential decay (red curve). Recordings were carried out in HEK293T cells held at −70 mV.
The α1-GABAAR is known to cluster at the inhibitory synapse,7 where it detects GABA released from the presynaptic axon terminal and mediates transient postsynaptic responses.17,18 GABA is released into the synaptic cleft through vesicle exocytosis either spontaneously (which elicits miniature inhibitory postsynaptic current, mIPSC) or driven by action potential.17 To test whether LiGABAR enables photocontrol over synaptic inhibition, we measured the photosensitivity of mIPSCs in LiGABAR-containing neurons. Cultured hippocampal neurons were transfected with a bicistronic construct encoding α1(T125C) and an expression marker (eGFP). The cells were subsequently treated with MAB-0 to generate LiGABARs in situ. As shown in Figure 4a, the amplitude of mIPSCs in a LiGABAR-containing neuron increased within 5 s of exposure to 380 nm light and remained elevated in darkness for at least 2 min, consistent with the slow thermal relaxation of cis MAB-0 measured in Figure 3. The amplitude of mIPSCs decreased again upon exposure to 500 nm light. To quantify photosensitivity, we calculated the average mIPSC when MAB-0 was in either trans or cis configuration (Figure 4b). Cis-to-trans photoisomerization caused a 38 ± 2% decrease in the peak amplitude and a 57 ± 4% decrease in the total charge transfer (n = 6; Figure 4b and c). The robust photocontrol over mIPSCs validates the applicability of LiGABAR for neurophysiological investigations.
Figure 4.
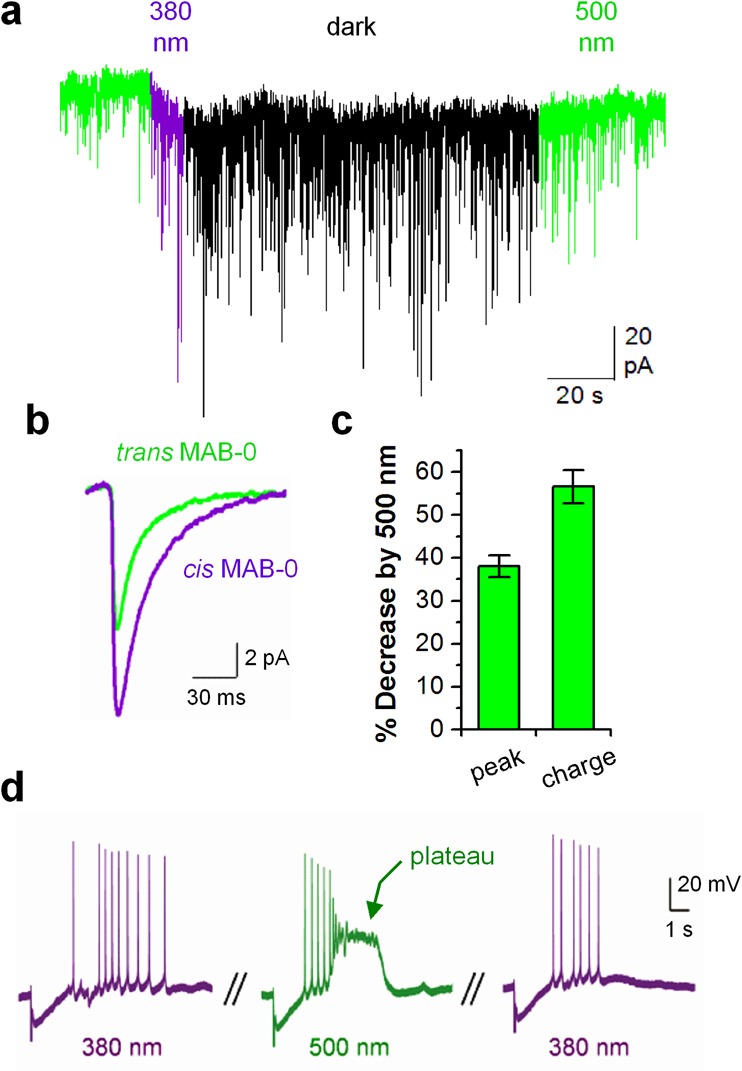
α1-LiGABAR enables photocontrol over miniature inhibitory postsynaptic currents (mIPSCs) and epileptiform formation in hippocampal neurons. (a) A representative continuous trace from a cultured hippocampal neuron containing α1-LiGABAR. The cell was held at −60 mV and was treated with inhibitors of voltage-gated sodium channels and ionotropic glutamate receptors. (b) Average mIPSC traces from the same cell shown in panel a. The green and purple traces represent average mIPSCs when MAB-0 was in the trans (500 nm) and cis (380 nm + dark) configuration, respectively. (c) Quantification of mIPSC photoregulation (mean ± SEM) as the percent decrease when MAB-0 was switched from cis to trans. The total charge transfer was measured by integrating the area of average mIPSC.17 (d) Photocontrol over neuronal excitation in a hippocampal slice. Current-clamp recording was carried out in a LiGABAR-containing CA1 pyramidal neuron. Illumination by 500 nm light resulted in an “epileptic” plateau potential that was subsequently eliminated by 380 nm light.
LiGABAR is developed for specifically probing a defined GABAAR subtype within a complex neural system. Efficacious PTL performance should occur only when the receptor possesses the engineered subunit. Consistent with our design principle, MAB-0 treatment did not confer detectable photosensitivity onto wild-type GABAARs (Supplementary Figure S2). Likewise, MAB-0 treatment did not photosensitize ionotropic glutamate receptors or voltage-gated channels in neurons (Supplementary Figure S2). All together, our results suggest that light can specifically modulate the inhibitory responses mediated by the engineered GABAAR with no apparent off-target effects.
Finally, we examined how the manipulation of LiGABAR alters neuronal excitation. Native GABAARs play a crucial role in preventing runaway excitation in the brain. Pharmacological blockade of GABAARs19,20 or mutations that alter GABAAR functions4 can result in epilepsy. Accordingly, we tested whether epileptiform activity can be induced by photoantagonizing LiGABAR in an intact, ex vivo preparation. To express the mutant subunit, we infected a hippocampal slice with an adeno-associated virus encoding α1(T125C) and an eGFP marker. After treating the slice with MAB-0, we applied a train of electrical pulses to stimulate presynaptic inputs and recorded the postsynaptic potential in 380 or 500 nm light. As shown in Figure 4d, the presynaptic stimulation triggered a series of postsynaptic action potentials in 380 nm light. When LiGABAR was antagonized with 500 nm light, the spike train culminated in a plateau potential that is characteristic of epileptiform activity.21 Switching light back to 380 nm eliminated the effect, demonstrating the reversibility of LiGABAR modulation. This result suggests that optical perturbation of LiGABAR function is sufficient to cause a profound impact on neuronal excitation.
In summary, our approach enables light to manipulate the inhibitory events mediated by a designated subtype of GABAAR. LiGABAR inherits two renowned benefits from opsin-based optogenetics, namely, the fine spatiotemporal precision of optical control and the capability of targeted gene expression using cell-type specific promoters.1,2 PTL photoisomerization can alter neuronal activity within milliseconds reversibly and reliably,22 suitable for accurate neurophysiological manipulations. Aside from these advantages, LiGABAR differs from microbial opsins in that it allows direct photocontrol over a genuine mediator in neuronal signaling. GABAARs mediate inhibitory neurotransmission in the nervous system, with different subtypes exerting distinct biophysical properties and cellular/subcellular distributions.3,5,7,8 The biochemical specificity of photocontrol makes LiGABAR favorable for probing the function/distribution of a GABAAR subtype within a neuron or a neural circuit. Compared to diffusible caged agonists or photochromic modulators (i.e., optopharmacology),2 the use of LiGABAR is more technically demanding due to the requirement of two exogenous components (mutant subunit and PTL). However, LiGABAR enables advanced investigations that aim to explore a specific receptor and/or neuron subtype in a complex system, which cannot be achieved with existing optopharmacology agents because of their lack of target specificity.2 LiGABAR thus presents a unique opportunity for understanding the mechanisms of neural inhibition, and will ultimately provide new insights into brain function and dysfunction.
Methods
Buffer formulations are available in Supporting Information. Animal care and experimental protocols were approved by the University of California Berkeley Animal Care and Use Committee.
Receptor Expression and PTL Treatment
Xenopus Oocytes
RNA was prepared with the mMessage mMachine T7 kit (Ambion). Defolliculated Xenopus oocytes were injected with 2.5–20 ng of mRNA in 50 nL of H2O in a ratio of 1:1 (α1:β2). The injected oocytes were incubated at 18 °C (in ORI buffer) and subjected to two-electrode voltage-clamp recording 1–3 days after RNA injection. To screen MAM-6 attachment sites, oocytes expressing each mutant receptor were incubated with 50 μM MAM-6 for 0.5–1.5 h prior to recording.
HEK293T Cells
Cells were maintained in Dulbecco’s Minimum Essential Medium (Gibco) supplemented with 10% fetal bovine serum (Gibco) at 37 °C and 5% CO2. Cells were plated at 18–24 × 103 cells/cm2 on poly-l-lysine-coated coverslips and were transfected via calcium phosphate precipitation. A total of 1.1 μg of DNA per coverslip (α:β:γ:eGFP = 0.15:0.15:0.75:0.05) was used. Recordings were carried out 36–48 h after transfection. Prior to recording, cells (in extracellular recording solution) were treated with dithiothreitol (DTT; 3 mM, 5 min), incubated in fresh extracellular recording solution (5 min), and then treated with PTL (25 μM, 20 min) at RT in the dark.
Cultured Hippocampal Neurons
Primary cultures of neonatal rat hippocampal neurons were prepared and maintained following previously described procedures.23 Neurons were transfected via calcium phosphate precipitation (1.2 μg of the bicistronic pAAV construct per well) 6–8 days after preparation. Recordings were carried out 1–2 weeks after transfection. Prior to recording, neurons (in extracellular recording solution) were treated with tris(2-carboxyethyl)phosphine (TCEP; 5 mM, 5 min), incubated in fresh extracellular recording solution (5 min), and then treated with MAB-0 (25 μM, 20 min) at RT in the dark.
Organotypic Slice Cultures
Sprague–Dawley rat pups (postnatal day 8) were anaesthetized and decapitated. Hippocampi were removed and sliced into 350-μm-thick sections. Slices were maintained at 34 °C on cell culture inserts in Neurobasal-A medium (Invitrogen) supplemented with 20% horse serum (Thermo Scientific), 0.03 units/mL insulin (Sigma), 0.5 mM ascorbic acid (Sigma), 1X Gluta-Max (Life Technologies), 80 units/mL penicillin (Life Technologies), 80 μg/mL streptomycin (Life Technologies), and 25 mM HEPES. One day after preparation, slices were injected with AAV9 (7.5 × 1012 vg/mL) encoding the bicistronic construct of eGFP-2A-α1(T125C). The CA1 pyramidal cell body layer was injected at 3–7 sites/slice with 100 nL of virus. Recordings were carried out 5–14 days postinjection. Prior to recording, slices were incubated for 2 min with 1 mM TCEP in artificial cerebrospinal fluid (ACSF), washed, and incubated with 25 μM MAB-0 in ACSF for 30 min at RT.
Acknowledgments
We thank Prof. C. Czajkowski (University of Wisconsin-Madison) for sharing GABAAR cDNAs and Dr. K. Durkin (University of California, Berkeley) for helpful assistance in molecular modeling. This work was supported by grants from the National Institute of Health (R01 EY018957, P30 EY003176, and PN2 EY018241 to R.H.K.) and the University of Massachusetts, Amherst (to J.J.C.).
Supporting Information Available
Details of PTL synthesis, preparation of plasmids and virus, electrophysiology, and molecular modeling. This material is available free of charge via the Internet at http://pubs.acs.org.
Author Present Address
¶ CNRS UMR 8246, INSERM UMR-S 1130, Institut Neuroscience Paris Seine, Université Pierre et Marie Curie, F-75005, Paris, France.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
Supplementary Material
References
- Yizhar O.; Fenno L. E.; Davidson T. J.; Mogri M.; Deisseroth K. (2011) Optogenetics in neural systems. Neuron 71, 9–34. [DOI] [PubMed] [Google Scholar]
- Kramer R. H.; Mourot A.; Adesnik H. (2013) Optogenetic pharmacology for control of native neuronal signaling proteins. Nat. Neurosci. 16, 816–823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudolph U.; Knoflach F. (2011) Beyond classical benzodiazepines: novel therapeutic potential of GABAA receptor subtypes. Nat. Rev. Drug Discovery 10, 685–697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones-Davis D. M.; Macdonald R. L. (2003) GABAA receptor function and pharmacology in epilepsy and status epilepticus. Curr. Opin. Pharmacol. 3, 12–18. [DOI] [PubMed] [Google Scholar]
- Möhler H. (2006) GABAA receptor diversity and pharmacology. Cell Tissue Res. 326, 505–516. [DOI] [PubMed] [Google Scholar]
- Korpi E. R.; Gründer G.; Lüddens H. (2002) Drug interactions at GABAA receptors. Prog. Neurobiol. 67, 113–159. [DOI] [PubMed] [Google Scholar]
- Brünig I.; Scotti E.; Sidler C.; Fritschy J. M. (2002) Intact sorting, targeting, and clustering of gamma-aminobutyric acid A receptor subtypes in hippocampal neurons in vitro. J. Comp. Neurol. 443, 43–55. [DOI] [PubMed] [Google Scholar]
- Picton A. J.; Fisher J. L. (2007) Effect of the alpha subunit subtype on the macroscopic kinetic properties of recombinant GABAA receptors. Brain Res. 1165, 40–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vu T. Q.; Chowdhury S.; Muni N. J.; Qian H.; Standaert R. F.; Pepperberg D. R. (2005) Activation of membrane receptors by a neurotransmitter conjugate designed for surface attachment. Biomaterials 26, 1895–1903. [DOI] [PubMed] [Google Scholar]
- Cavalla D.; Neff N. H. (1985) Photoaffinity labeling of the GABAA receptor with [3H]muscimol. J. Neurochem. 44, 916–921. [DOI] [PubMed] [Google Scholar]
- Kloda J. H.; Czajkowski C. (2007) Agonist-, antagonist-, and benzodiazepine-induced structural changes in the α1 Met113-Leu132 region of the GABAA receptor. Mol. Pharmacol. 71, 483–493. [DOI] [PubMed] [Google Scholar]
- Sancar F.; Czajkowski C. (2011) Allosteric modulators induce distinct movements at the GABA-binding site interface of the GABAA receptor. Neuropharmacology 60, 520–528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tochitsky I.; Banghart M. R.; Mourot A.; Yao J. Z.; Gaub B.; Kramer R. H.; Trauner D. (2012) Optochemical control of genetically engineered neuronal nicotinic acetylcholine receptors. Nat. Chem. 4, 105–111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller P. S.; Smart T. G. (2010) Binding, activation and modulation of Cys-loop receptors. Trends Pharmacol. Sci. 31, 161–174. [DOI] [PubMed] [Google Scholar]
- Padgett C. L.; Hanek A. P.; Lester H. A.; Dougherty D. A.; Lummis S. C. R. (2007) Unnatural amino acid mutagenesis of the GABAA receptor binding site residues reveals a novel cation-pi interaction between GABA and β2 Tyr97. J. Neurosci. 27, 886–892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura N.; Sueyoshi T.; Yamanaka H.; Imai E.; Yamamoto S.; Hasegawa S. (1976) Thermal cis-to-trans isomerization of substituted azobenzenes. 2. Substituent and solvent effects. Bull. Chem. Soc. Jpn. 49, 1381–1387. [Google Scholar]
- Farrant M.; Nusser Z. (2005) Variations on an inhibitory theme: phasic and tonic activation of GABAA receptors. Nat. Rev. Neurosci. 6, 215–229. [DOI] [PubMed] [Google Scholar]
- Goldstein P. A.; Elsen F. P.; Ying S. W.; Ferguson C.; Homanics G. E.; Harrison N. L. (2002) Prolongation of hippocampal miniature inhibitory postsynaptic currents in mice lacking the GABAA receptor alpha1 subunit. J. Neurophysiol. 88, 3208–3217. [DOI] [PubMed] [Google Scholar]
- Matthews W. D.; McCafferty G. P. (1979) Anticonvulsant activity of muscimol against seizures induced by impairment of GABA-mediated neurotransmission. Neuropharmacology 18, 885–889. [DOI] [PubMed] [Google Scholar]
- Gibbs J. W. 3rd; Shumate M. D.; Coulter D. A. (1997) Differential epilepsy-associated alterations in postsynaptic GABAA receptor function in dentate granule and CA1 neurons. J. Neurophysiol. 77, 1924–1938. [DOI] [PubMed] [Google Scholar]
- Dingledine R.; Hynes M. A.; King G. L. (1986) Involvement of N-methyl-D-aspartate receptors in epileptiform bursting in the rat hippocampal slice. J. Physiol. 380, 175–189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szobota S.; Gorostiza P.; Del Bene F.; Wyart C.; Fortin D. L.; Kolstad K. D.; Tulyathan O.; Volgraf M.; Numano R.; Aaron H. L.; Scott E. K.; Kramer R. H.; Flannery J.; Baier H.; Trauner D.; Isacoff E. Y. (2007) Remote control of neuronal activity with a light-gated glutamate receptor. Neuron 54, 535–545. [DOI] [PubMed] [Google Scholar]
- Fortin D. L.; Banghart M. R.; Dunn T. W.; Borges K.; Wagenaar D. A.; Gaudry Q.; Karakossian M. H.; Otis T. S.; Kristan W. B.; Trauner D.; Kramer R. H. (2008) Photochemical control of endogenous ion channels and cellular excitability. Nat. Methods 5, 331–338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Mara M.; Cromer B.; Parker M.; Chung S. H. (2005) Homology model of the GABAA receptor examined using Brownian dynamics. Biophys. J. 88, 3286–3299. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.