

Abstract
Nonalcoholic fatty liver disease (NAFLD) is a chronic liver disorder that is increasing in prevalence with the worldwide epidemic of obesity. NAFLD is the hepatic manifestation of the metabolic syndrome. The term NAFLD describes a spectrum of liver pathology ranges from simple steatosis to steatosis with inflammation nonalcoholic steatohepatitis and even cirrhosis. Metabolic syndrome and NAFLD also predict hepatocellular carcinoma. Many genetic and environmental factors have been suggested to contribute to the development of obesity and NAFLD, but the exact mechanisms are not known. Intestinal ecosystem contains trillions of microorganisms including bacteria, Archaea, yeasts and viruses. Several studies support the relationship between the intestinal microbial changes and obesity and also its complications, including insulin resistance and NAFLD. Given that the gut and liver are connected by the portal venous system, it makes the liver more vulnerable to translocation of bacteria, bacterial products, endotoxins or secreted cytokines. Altered intestinal microbiota (dysbiosis) may stimulate hepatic fat deposition through several mechanisms: regulation of gut permeability, increasing low-grade inflammation, modulation of dietary choline metabolism, regulation of bile acid metabolism and producing endogenous ethanol. Regulation of intestinal microbial ecosystem by diet modifications or by using probiotics and prebiotics as a treatment for obesity and its complications might be the issue of further investigations.
Keywords: Intestinal microbiota; Dysbiosis, Nonalcoholic fatty liver disease; Obesity
Core tip: There is increasing evidence for the relation between dietary habits, gut microbiota and obesity. Nonalcoholic fatty liver disease is a common complication of obesity. This manuscript summarizes the relationship between intestinal microbial dysregulation and fatty liver disease related with obesity, and their proposed mechanisms.
INTRODUCTION
Human gut consists of a large number of commensal microorganisms, collectively known as “intestinal microbiota”, which are essential for the preservation of the integrity of the mucosal barrier function, for the absorption of nutrients and energy homeostasis[1]. Recent evidence suggests that enteric microbiota may play a significant role in the development of obesity and its complications[2].
Nonalcoholic fatty liver disease (NAFLD) describes a condition caused by a deposition of fat within the liver cells in the absence of alcohol consumption, which is linked to being obese or overweight in most cases[3-5]. It encompasses a disease spectrum ranging from simple steatosis to nonalcoholic steatohepatitis (NASH), which is histologically characterized by hepatocyte injury, inflammation and variable degrees of fibrosis. Nonalcoholic steatohepatitis progress to advanced fibrosis and cirrhosis in 37% patients[6]. A “two hit” mechanism has been proposed; however, the complete pathogenesis remains incompletely understood. Fatty liver disease is dramatically increasing in childhood and adolescent obesity, and it has become the most common form of chronic liver disease in these age groups[7,8].
The liver is located on the first point of the body for bacteria and microbial components, as well as other endogenous and exogenous toxins present in the portal blood and it generates the initial immunological and hormonal response to these molecules[9]. Interactions between the gut and the liver are bidirectional; hormones, inflammatory mediators and the products of digestion and absorption all directly influence liver function. Changes prompted by spesific intestinal microbiota are characterized not only by a general obesogenic and dysmetabolic framework but also by a specific de novo hepatic lipogenesis[10]. This review will discuss the relationship between intestinal microbiota and obesity and also NAFLD, and their proposed mechanisms.
CHILDHOOD OBESITY AND NONALCOHOLIC FATTY LIVER DISEASE
Childhood obesity is a major health problem in all over the world because of its impact on the physical and psychological health of children, and also on the development of chronic diseases later in life such as atherosclerosis, NAFLD, hypertension, hyperlipidemia and diabetes[3,11-13]. Global prevalance of childhood overweight/obesity varies from 5.7% to 40% in different populations[14-17]. Several case reports, human and animal studies demonstrated that obesity is an important risk factor for carcinogenesis in many malignant neoplasms and also in hepatocellular carcinoma[18-21]. Moreover, childhood obesity was shown to be related with increased risk of primary liver cancer in later adulthood[22]. Obesity and related complications including fatty liver disease, cardiovascular disorders and hepatocellular carcinoma were found associated with the reduction in the life expectancy compared to general population[23]. Several genes contribute to weight gain by controlling feeding behavior, energy expenditure and metabolism, but can only partially account for the development of obesity. Thirty two loci of the human genome had been found associated with body mass index and increased body weight, but changes of these genes affect only 2% of the population[24]. On the other hand, exogenous obesity develops primarily due to energy intake that exceeds energy expenditure, and many environmental and host factors interact with this process in many ways.
Adipose tissue is not a passive site of energy storage. Although the major function of the adipocyte is to store and release energy in the form of triglyceride during excess food consumption and starved periods, respectively, it is also an endocrine organ producing several proteins (adipokines like adiponectin and leptin) and cytokine mediators interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-α) with many biological activities[25]. Development and complications of obesity consist of complex mechanisms; in which numerous adipokines, hormones and cytokines take place[26]. Although the liver participates in the systemic inflammation of obesity, the dominant controller organ is the adipose tissue. Adipokines when imbalanced, together orchestrate a proinflammatory and insulin-resistant state that further contributes to the pathogenesis of NAFLD and its progression to NASH[27]. A large body of emerging literature seems to suggest that intestinal microbiota is also involved in the development of obesity and its complications including obesity-related liver disease.
INTESTINAL MICROBIOTA
The human intestine contains a very crowded and heterogeneus microbial system, consisting at least 100 trillion (1014) microbial cells weighing about 1.5 kg and composed of more than 2000 species[28]. Luminal microbial cells contain genes 150 times more than our own host genomes[28]. This complex community contains taxa from bacteria, eukaryotes, viruses, and at least one archaeon, that interact with one another and with the host, involving regulation of local/systemic immunity, metabolic and trophic functions[29].
Microbial culture studies detect only a small number of the species of intestinal bacteria. Nowadays, composition and the diversity of intestinal microbiota is revealed by culture-independent genetic and metagenomic techniques[30]. Metagenomic analysis and 16S ribosomal RNA gene sequencing have shown that Firmicutes, Bacteriodetes, Actinobacteria, Proteobacteria, Fusobacteria, Spirochaetae and Verrucomicrobia are the predominant bacterial phyla among the intestinal bacteria in adults[31]. Firmicutes and Bacteriodetes constitute about 90% of all intestinal microbial cells. While the dominating phyla are relatively constant between individuals, diversity increases along the taxonomic line with each individual harboring over a hundred unique species.
Gut microbiota has evolved with humans as a mutualistic partner; however, changes in the composition of the gut microbiota (dysbiosis) have been found to be related with several clinical conditions such as obesity, diabetes, fatty liver disease, atherosclerosis, allergic diseases, gastrointestinal diseases, autoimmune diseases and cancer[30-35] (Figure 1).
Figure 1.
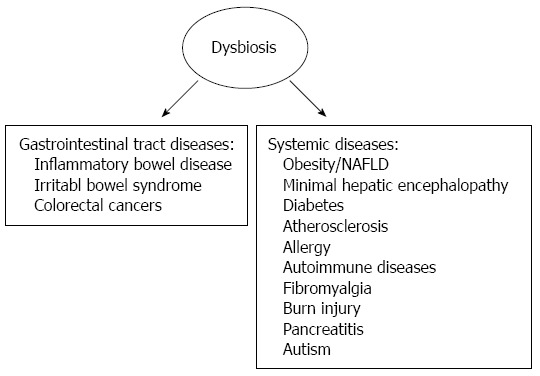
Some gastrointestinal and systemic conditions related with altered intestinal microbiota[31,32,35].
INTESTINAL MICROBIOTA AND OBESITY
There are many animal and human studies investigating the relationship between intestinal microbiota and obesity or body weight changes in the literature. In the study of Ley et al[36], which investigated over five thousands of bacterial gene sequences from the distal intestinal microbiota of genetically obese ob/ob mice, lean ob/+ and wild-type siblings, the investigators found that genetically obese mice had a 50% reduction in abundance of Bacteriodetes and a proportional increase in Firmicutes phyla compared to lean sibling mice although they were fed with the same polysaccharide-rich diet. Since both groups of animals had been fed with the same diet, it was suggested that obesity might affect the diversity of gut microbiota[36]. Similarly, several human studies showed that human obesity is associated with a low abundance of intestinal Bacteroidetes and high abundance of Firmicutes, and with reduced bacterial diversity[37-39]. Obese children was also shown to have different gut microbiota compared to lean peers, and their bacterial composition have been found similar to obese adults[40-45]. Although Bifidobacterium is not a predominating phylum in the gut, it seems to play an important role in host metabolism. Following a high-fat diet, reduced Bifidobacterium was observed, with a secondary increase of inflammatory activity, increased fat mass and insulin resistance in mice[46]. Interestingly, these changes in the gut microbes were shown to be reversed by low-calorie diet and consequent weight loss[34,42,47].
MECHANISMS LINKING THE INTESTINAL MICROBIOTA AND OBESITY
Western type fat- and energy-rich diet changes intestinal microbiota. On the other hand, altered microbiota also affects the host metabolism and causes inflammation and increased fat deposition of the body. Numerous animal models consistently demonstrated that gut microbiota can modulate host energy homeostasis and adiposity through different mechanisms, for example energy harvest from the diet, lipopolysaccharides (LPSs)-induced chronic inflammation, and modulation of tissue fatty acid composition, host gene expression and gut-derived peptide secretion[48].
Intestinal microbiota and energy harvest from the diet (caloric salvage)
Mice with gut microbiota were shown to have an increased capability to harvest energy from the gut contents compared with germ-free rats[49]. Metagenomic analyses of the microbiota performed in obese mice and humans revealed an increased capacity for the degradation (fermentation) of carbohydrates[50,51]. This microbial fermentation increases the amount of short-chain fatty acids (SCFAs), such as acetate, propionate, butyrate, and L-lactate. These SCFAs have important roles in the reduction of intestinal pH, in the regulation of energy metabolism, immunity, and adipose tissue expansion and in modulating cancer cell development[52]. Butyrate is used as an energy substrate for colonocytes, acetate is potentially used as a cholesterol or fatty acid precursor and propionate is used as a gluconeogenic substrate in the liver[52-54]. In addition to these functions, SCFAs are also physiological ligands of G-protein coupled receptors GPR43 and 41 (also called free fatty acid receptor 2 and 3, respectively), which are expressed in several cell types (immune cells, endocrine cells, and adipocytes) of host tissues[55,56] (Table 1). Activation of GPR43 by the SCFAs contributes to the inhibition of lipolysis and to adipocyte differentiation, thereby increases the adipose tissue in high-fat-diet-fed mice[57,58]. Monosaccharides produced by microbial fermentation, absorbed and transferred to the liver via portal vein, activate the hepatic carbohydrate response element binding protein (ChREBP) that increases the transcription of several proteins involved in hepatic lipogenesis thus contributing to hepatic fat accumulation[55]. The effect of intestinal microbiota on obtaining energy from the gut contents had also been tested in human studies. Jumpertz et al[59] reported that the amount of stool energy in proportion of ingested calories was positively and negatively correlated with the abundance of phylum Bacteroidetes and phylum Firmicutes in the feces, respectively. Approximately a-150 kcal difference could be achieved with a change of 20% relative increases of Firmicutes and decreases of Bacteroidetes in the stool of lean individuals. Thus, excessive calories taken in the form of SCFAs from microbial fermentation of luminal contents may be a contributing factor in the obese state.
Table 1.
Some key host proteins and factors those their expressions were changed by intestinal microbial changes and those play role in the development of obesity
| Host protein/factor | Function |
| Fiaf (fasting-induced adipocyte factor) | A protein that inhibits lipoprotein lipase activity |
| ChREBP (carbohydrate response element-binding protein) | A transcription factor that recognizes monosaccharides in the portal vein and plays a key role in the hepatic carbohydrate metabolism |
| SREBP-1 (liver sterol response element-binding protein type-1) | A transcription factor family that controls the lipid synthesis in the liver and other tissues |
| G-protein coupled receptors (GPR43 and GPR41) | Proteins expressed in enteroendocrine L-cells those recognize luminal SCFAs and mediate SCFA-induced GLP-1 release. They also present in adipocytes and promote adipogenesis by increasing lipid accumulation and inhibiting lipolysis and stimulate leptin production in response to SCFAs |
| Toll like receptors | Transmembrane molecules those recognize bacterial breakdown products |
| GLP 1 (Glucagon-like peptide 1) | A protein produced by intestinal epithelial endocrine L-cells that stimulates insulin secretion, inhibits gastrointestinal motility, regulates appetite and food intake |
| Peptide YY | A peptide hormone produced by intestinal epithelial endocrine L-cells that inhibits intestinal motility |
| Farnesoid X receptor | A receptor expressed in liver and intestine that regulates bile acid synthesis,transport and detoxification |
GLP-1: Glucagon-like peptide 1; SCFAs: Short-chain fatty acids.
Fiaf (fasting-induced adipocyte factor)
In addition to extract calories from otherwise indigestible dietary polysaccharides, the presence of the intestinal microbiota also stimulates changes in the expression of genes coding for peptides in host tissues, which control energy homeostasis and nutrient availability (Table 1). Bäckhed et al[60] firstly demonstrated that colonizing germ-free mice with gut microbiota had led to a decrease in the intestinal expression of angiopoietin-like factor IV [ANGPTL4, also called fasting-induced adipose factor (Fiaf)], thereby blunting the inhibition of lipoprotein lipase in the adipose tissue. In the study of Mandrad et al[61] inhibition of lipoprotein lipase blocked the dissociation of fatty acids from triglycerides for uptake into tissues and upregulated fatty acid oxidation and uncoupling proteins, and potentially reduced the amount of fat storage. These results may explain why conventionalized mice are more sensitive than germ-free mice to fat storage when fed with a high-fat diet and supports the role of the gut microbiota in the development of obesity[50,52,62].
Increased intestinal permeability and inflammation
Obesity, diabetes and insulin resistance are associated with a low grade systemic inflammation[63-65]. Several studies have been conducted on the effect of systemic inflammation on glucose and lipid metabolism, but little is known about its triggers.
Intestinal microorganisms have highly conserved microbial molecules, called “pathogen- associated molecular patterns” (PAMPs) and endogeneous products called “damage-associated molecular patterns” (DAMPs), which are recognized by pattern recognition receptors. These receptors include membranous toll-like receptors (TLRs) and intracellular NOD-like receptors (NLRs). TLRs recognise the potential pathogens in the intestinal lumen and induce the immune response. Microbial products such as LPSs, lipopeptides, DNA and RNA have potentially hepatotoxic effects as they are potent inducers of inflammation. Among 13 known TLRs, TLR2, TLR4 and TLR9 have been shown to play a role in the development of NAFLD. Stimulation of TLRs results in the activation of several different intracellular signaling cascades including stres-activated and mitogen activated protein (MAP) kinases, Jun N-terminal kinases, p38 MAP kinase, interferon regulatory factor 3 and nuclear factor kappa B (NF-κB) pathways[9,66]. NF-κB, is an important transcription factor in the cell, translocates to the nucleus and induces the transcription of a variety of inflammatory cytokines and chemokines such as TNF-α and IL-1β[67,68]. de la Serre et al[69] showed that rats prone to weight gain exhibited an increase in TLR4 activation associated with ileal inflammation, decreased intestinal alkaline phosphatase activity, a luminal enzyme that detoxifies the bacterial component (LPS), known to cause inflammation, and increased innate immune system activation in the luminal wall when compared to the rats resistant to obesity. In another study, Cani et al[46] demonstrated knockout rats of an immunoprotein (CD14), which was necessary to cause an inflammatory reaction to LPS, were resistant to weight gain. Vijay-Kumar et al[70] showed that TLR5 knockout mice exhibited hyperphagia and developed hallmark features of metabolic syndrome, including hyperlipidemia, hypertension, insulin resistance, and increased visceral fat deposition. These metabolic changes correlated with changes in the composition of the gut microbiota, and transfer of the gut microbiota from TLR5-deficient mice to wild-type germ-free mice conferred many features of metabolic syndrome to the recipients[70]. Lam et al[71] showed that mice fed with high-fat diet had reduced zona occludens-1 mRNA expression (40%) and increased permeability in proximal colon, and increased levels of TNF-α and IL-1 in mesenteric fat compared to mice fed with the control diet. In human studies, low grade endotoxemia (metabolic endotoxemia) was found associated with high-fat meal, obesity, NAFLD and diabetes[72-75]. All these data shows that the inflammatory mileu is the key component of the development of obesity and its complications.
Releasing of gut hormones
Intestinal microbial system regulates entero-endocrine cells and promotes the release of several gut hormones. Peptide YY (PYY) is an enteroendocrine cell-derived hormone normally inhibits gut motility. Samuel et al[76] showed that PYY expression was lower in both Gpr41 -/- germ-free and conventionalized mice compared to Gpr41 +/+ mice. Reduced PYY expression resulted in increased intestinal transit time and reduced harvest of dietary energy[72-76]. In other studies, dietary fructo-oligo-saccharides increased the abundance of Bifidobacterium in the distal intestine, which led to increased colonic fermentation and glucagon-like peptide 1 (GLP-1) levels, decreased serum orexigenic peptide ghrelin and decreased food intake, fat mass, and hepatic steatosis in rats[77,78]. Based on these results, it can be concluded that dietary inulin-type fructans could play a role in the management of obesity and diabetes through their capacity to promote secretion of endogenous gastrointestinal peptides involved in appetite regulation[77,78].
INTESTINAL MICROBIOTA AND FATTY LIVER DISEASE
Nonalcoholic fatty liver disease is a multifactorial disease and the underlying mechanisms are incompletely understood. Various genetic, metabolic, inflammatory, nutritional and environmental factors are thought to contribute to its pathogenesis[9]. Nutrition is the most important environmental factor; its role may be more complex than inducing fat acumulation in the liver and may involve interactions with the microbiota[9]. The potential role of the intestinal microbiota on the liver diseases had been known since 1921[79]. Several animal and human studies have investigated possible relationships between the intestinal microbiota and NAFLD[46,60,75,79-85]. Bäckhed et al[60] firstly showed that microbiota stimulated monosaccharide absorption from the intestinal lumen, promoted de novo fatty acid synthesis and triglyceride production, as confirmed by increased activity of acetyl-CoA carboxylase and fatty acid synthase. The relation between altered microbiota and liver was shown even in pediatric age group studies. Karlsson et al[45] showed that obese/overweight preschool children had an increased amount of the gram-negative family Enterobacteriaceae and had inverse correlation between Bifidobacterium concentration and alanine aminotransferase levels.
MECHANISMS LINKING THE INTESTINAL MICROBIOTA AND NAFLD
Studies suggest that intestinal microbiota may stimulate liver steatosis though several mechanisms[86]: (1) induction of obesity by harvesting energy from otherwise indigestible dietary polysaccharides (above mentioned); (2) regulation of gut permeability and stimulation of low grade inflammation; (3) modulation of dietary choline metabolism; (4) regulation of bile acid metabolism; and (5) stimulation of endogenous ethanol production by enteric bacteria.
Small intestinal bacterial overgrowth and increased intestinal permeability
Intestinal epithelial cells separate the intestinal microbial environment from the host immune system. Epithelial cells are linked to each other with tight junctions, which play a central role in maintaining intestinal barrier integrity[87]. In human studies NAFLD has been associated with increased LPS plasma levels, through mechanisms involving increased intestinal permeability, small intestinal bacterial overgrowth (SIBO), tight junction alteration and bacterial translocation[83,84,88-90].
SIBO is defined as an increase in the number and/or alteration in the composition of bacteria in the proximal gastrointestinal tract. Association of increased intestinal permeability and fatty liver was first demonstrated by Miele et al[84]. They reported that patients with fatty liver disease had increased gut permeability related to SIBO and disrupted tight junctions compared to healthy adults[84]. They also showed that gut permeability and SIBO were correlated with the severity of the liver steatosis. In the study of Gäbele et al[91], mice were fed with high-fat diet to stimulate NASH and then exposed to dextran sulphate sodium, an agent that causes intestinal epithelial injury. They showed that combined administration of high-fat diet and dextran sulphate sodium induced fibrosis in the liver[91]. All these data support the hypothesis that altered homeostasis between host and intestinal microbial system at the intestinal epithelial barrier level promotes bacterial translocation from the gut into the portal circulation and induces the liver damage[86,92,93].
Low grade inflammation and fatty liver
Several studies demonstrated that the altered microbiota caused low-grade inflammation, which had a pivotal role for the development of obesity and its complications including NAFLD. Association of inflammation and fatty liver was firstly demonstrated by Cani et al[46]. They showed that continuous LPS infusion in mice increased the insulin resistance, liver triglyceride content and adipose tissue inflammation. After this study, other animal studies conducted in TLR4 knockout mice confirmed that TLR4 was essential for hepatic steatosis and NASH development[94-96]. Kupffer cells, which express the highest levels of TLR4 in the liver, are the primary cells in liver inflammation that respond to LPSs in order to produce inflammatory cytokines, chemokines and reactive oxygen species (ROS)[97,98]. In genetically obese mice, the administration of LPS induces changes in Kupffer cells function and increases liver parenchymal sensitivity to TNF-α[99]. TNF-α, which is the most important of LPS-TLR4 induced cytokines in these cells, is recognized as a mediator of hepatotoxicity, inflammation an NASH development in mice[100]. Besides, hepatic stellate cells might also have a substantial role in constituting the inflammatory cascade of the liver consequently associated with metabolic endotoxemia. Indeed, these cells are the major fibrogenic cell type in injured liver were shown to be the target through which TLR4 promoted fibrogenesis via enhancement of transforming growth factor-β (TGF-β) signalling[101-103].
The “second hit” mechanism of the NAFLD/NASH pathogenesis include enhanced lipid peroxidation and increased generation of ROS[104]. Inflammasomes, major contributors of inflammation, are cytoplasmic multiprotein complexes, which include nucleotide-binding domain (NLRPSs). NLRPSs are sensors of the bacterial PAMPs and DAMPs[105,106]. They manipulate the cleavage of pro-inflammatory cytokines such as pro-IL-1β and pro-IL-18. Most DAMPs induce the production of ROS, which is known to activate NLRP3 inflammasome[107,108]. Henao-Mejia et al[108] reported that inflammasome alterations or IL-18 deficiency cause intestinal microbial changes by enhancing portal influx of TLR4 and TLR9 ligands, which in turn increase hepatic TNF-α production in mice. Human studies also demonstrated that endotoxin levels were increased in both adult and pediatric obese patients with fatty liver disease[74,109,110]. Moreover, endotoxin levels were found to be correlated with the severity of the disease[75,111]. All these data support that chronic low-grade inflammation caused by obesogenic microbial ecosystem is a real “hepatotoxin” and has a key role in the pathogenesis of obesity related fatty liver disease.
Altered choline metabolism
Choline is a water-soluble essential nutrient. It is an important phospholipid component of the cell membrane and is the precursor molecule for the neurotransmitter acetylcholine. Choline has important roles in fat metabolism in the liver and a very-low-lipoprotein assembly, and also it promotes lipid transport from the liver[86,112]. Exogenous sources of choline are meat, dairy products, fish, soybeans, nuts and whole grains. Endogenous sources of choline, in the form of phosphatidylcholine, are biliary lipids, exfoliated epithelial cells and intestinal bacteria[113]. Buchman et al[114] showed that choline-deficient nutrition stimulated the liver steatosis. Furthermore, they demonstrated that a 6-wk choline supplementation reversed this pathology in patients. Gut microbiota secrete enzymes that cleave the dietary choline to its toxic metabolites (dimethylamine and trimethylamine). Liver uptakes these toxic methylamines and converts them to trimethylamine-N-oxide which induce inflammation in the liver[115,116]. Spencer et al[117] showed that the compositions of the gastrointestinal microbial communities changed with dietary choline content and especially Gammaproteobacteria and Erysipelotrichi levels were directly associated with changes liver steatosis in each subject during choline depletion. The role of dietary choline in NAFLD can be explained by the bioavailibility of free choline to for lipoproteins in the liver (especially very- low-density-lipoprotein-VLDL), which allows the export of free fatty acids from this organ[113]. If the gut microbiota converts excessive amounts of dietary choline into trimethylamine, this leads to reduced choline bioavailibility and consequent fatty liver disease[118].
Altered bile acid metabolism
Bile acids modulate lipid absorption and cholesterol homeostasis. The nuclear bile acid receptor, called farnesoid X receptor (FXR), is strongly expressed at bile acid excretion (liver) and absorption (intestine) regions. Bile acids also act as signaling molecules and activate FXR and the G-protein coupled receptor TGR5. Through activation of downstream signaling pathways of these key receptors, bile acids regulate not only their own synthesis and enterohepatic circulation, but also impact on hepatic lipid, glucose, and energy homeostasis[119]. FXR plays a key role in the control of hepatic de novo lipogenesis, VLDL triglyceride export and plasma triglyceride turnover[120]. TGR5 binds secondary bile acids and promotes glucose homeostasis, by stimulating secretion of GLP-1[121]. Besides, bile acids have a bacteriostatic activity. Gut microbiota can modulate bile acid metabolism. Swann et al[82] showed that gut microbiota can indirectly promote hepatic steatosis and lipid peroxidation through FXR stimulation changes in bile acid secretion. On the other hand, high-fat diet changes the bile acid composition, which influences the conditions for gut microbial environment and causes dysbiosis[122,123].
Stimulation of endogenous ethanol production by enteric bacteria
Intestinal microbiota produces a number of potentially hepatotoxic compounds such as ethanol, phenols, ammonia and they are transported to liver by portal system. These toxins stimulate hepatic Kupffer cells for production of nitric oxide and cytokines such as TNF-α[9,124]. Acetaldehyde and acetate are two major metabolites of ethanol. Ethanol can increase acetate production via inhibition of the Krebs cycle. Acetate is a substrate for fatty acid synthesis. On the other hand, actaldehyde and its metabolites may lead to the formation of reactive oxygen species. ROS production could be involved in liver injury by contributing to the disruption of intestinal barrier function and to the two hit mechanisms of steatohepatitis[80,86,125]. Ethanol and LPS also stimulate the production of ROS by parenchymal and nonparenchymal liver cells. Gustot et al[124] showed that enteral ethanol exposure induced steatosis and increased liver weight, aminotransferase levels, and TLR1, 2, 4, 6, 7, 8, and 9 liver mRNA expressions in mice. They concluded that ethanol-fed mice exhibited an oxidative stress dependent on upregulation of multiple TLRs in the liver and were sensitive to liver inflammation induced by multiple bacterial products recognized by TLRs[124].
In a human study, Nair et al[126] demonstrated that obese women with NASH had higher breath ethanol concentrations than healthy controls detected by gas chromatography. Similarly, Zhu et al[127] showed in their pediatric age group study that Proteobacteria, Enterobacteriaceae, and Escherichia (is a well-known ethanol producer bacteria) were the only phylum, family and genus types exhibiting significant difference between the patients with and without NASH microbiomes. Similar blood-ethanol concentrations were observed between healthy subjects and obese non-NASH patients; however, NASH patients exhibited significantly elevated blood ethanol levels[127]. Ethanol contributes to iNOS-mediated intestinal hyperpermeability, and therefore enhances the passage of endotoxins from the intestinal lumen into the portal system[128].
CONCLUSION
High energy diets alter intestinal microbiata, induce gut dysfunction, which subsequently result in visceral fat inflammation and systemic metabolic dysregulation. An obesogenic microbiota can alternate liver function by stimulating hepatic triglyceride and by modulating systemic lipid metabolism that indirectly impact the storage of fatty acids in the liver (Figure 2). Several studies suggested that intestinal microbiota might also play an important part in progression of NAFLD to NASH. Modulation of gut microbiota by diet modifications or by using probiotics, prebiotics and synbiotics as a treatment for obesity and fatty liver disease might be the issue of further investigations (Figure 3).
Figure 2.
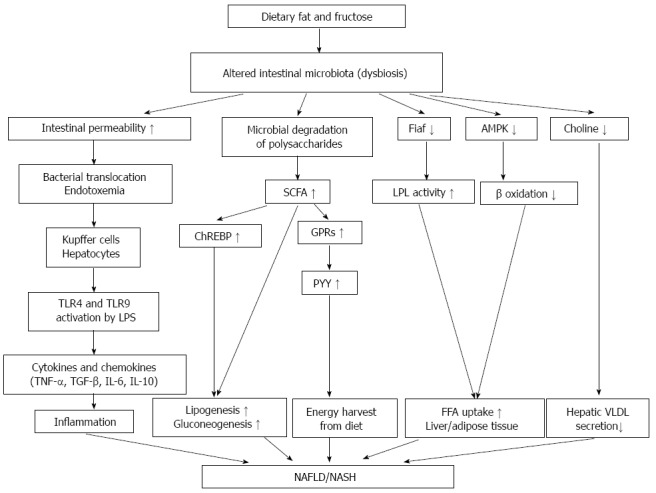
Effects of gut microbiota on the development of nonalcoholic fatty liver disease and nonalcoholic steatohepatitis through the gut-liver axis. Altered intestinal bacterial composition (dysbiosis) results in degradation of carbohydrates in the intestinal lumen and produces short-chain fatty acids (SCFAs), which are substrates for hepatic lipogenesis and gluconeogenesis. The interaction of SCFAs with G-protein coupled receptors (GPRs) releases the peptide YY (PYY), which modulates gut motility and nutrient absorption. SCFAs stimulate hepatic carbohydrate response element binding protein (ChREBP) and increase lipogenesis. Bacterial translocation to the portal circulation causes interaction of bacterial endotoxins (lipopolysaccharides, LPS) with hepatic toll-like receptors (TLR4 and TLR9) and results in the release of cytokines and chemokines. Decreased Fiaf (fasting-induced adipose factor) levels enhance lipoprotein lipase (LPL) activity and cause fat accumulation. Choline deficiency causes liver steatosis via decreased secretion of very- low-density-lipoprotein-(VLDL) from the liver.
Figure 3.
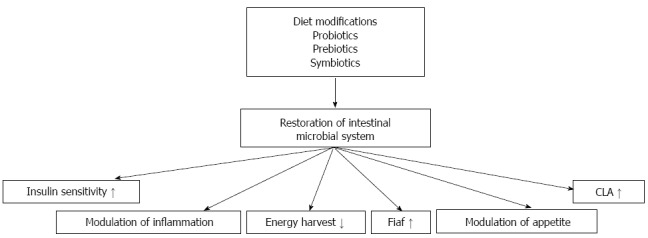
Probable mechanisms of action of the antiobesity effects of modulated intestinal microbiota. CLA: Conjugated linoleic acid; Fiaf: Fasting-induced adipocyte factor.
ACKNOWLEDGMENTS
I would like to thank Professor Erbil Unsal for editing and revising the language of the manuscript.
Footnotes
P- Reviewer: Marinho RT, Nseir W S- Editor: Qi Y L- Editor: A E- Editor: Zhang DN
References
- 1.Ley RE, Peterson DA, Gordon JI. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell. 2006;124:837–848. doi: 10.1016/j.cell.2006.02.017. [DOI] [PubMed] [Google Scholar]
- 2.Ridaura VK, Faith JJ, Rey FE, Cheng J, Duncan AE, Kau AL, Griffin NW, Lombard V, Henrissat B, Bain JR, et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science. 2013;341:1241214. doi: 10.1126/science.1241214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Arslan N, Büyükgebiz B, Oztürk Y, Cakmakçi H. Fatty liver in obese children: prevalence and correlation with anthropometric measurements and hyperlipidemia. Turk J Pediatr. 2005;47:23–27. [PubMed] [Google Scholar]
- 4.Sinatra FR. Nonalcoholic fatty liver disease in pediatric patients. JPEN J Parenter Enteral Nutr. 2012;36:43S–48S. doi: 10.1177/0148607111421610. [DOI] [PubMed] [Google Scholar]
- 5.Alisi A, Feldstein AE, Villani A, Raponi M, Nobili V. Pediatric nonalcoholic fatty liver disease: a multidisciplinary approach. Nat Rev Gastroenterol Hepatol. 2012;9:152–161. doi: 10.1038/nrgastro.2011.273. [DOI] [PubMed] [Google Scholar]
- 6.Argo CK, Northup PG, Al-Osaimi AM, Caldwell SH. Systematic review of risk factors for fibrosis progression in non-alcoholic steatohepatitis. J Hepatol. 2009;51:371–379. doi: 10.1016/j.jhep.2009.03.019. [DOI] [PubMed] [Google Scholar]
- 7.Berardis S, Sokal E. Pediatric non-alcoholic fatty liver disease: an increasing public health issue. Eur J Pediatr. 2014;173:131–139. doi: 10.1007/s00431-013-2157-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Widhalm K, Ghods E. Nonalcoholic fatty liver disease: a challenge for pediatricians. Int J Obes (Lond) 2010;34:1451–1467. doi: 10.1038/ijo.2010.185. [DOI] [PubMed] [Google Scholar]
- 9.Abu-Shanab A, Quigley EM. The role of the gut microbiota in nonalcoholic fatty liver disease. Nat Rev Gastroenterol Hepatol. 2010;7:691–701. doi: 10.1038/nrgastro.2010.172. [DOI] [PubMed] [Google Scholar]
- 10.Vajro P, Paolella G, Fasano A. Microbiota and gut-liver axis: their influences on obesity and obesity-related liver disease. J Pediatr Gastroenterol Nutr. 2013;56:461–468. doi: 10.1097/MPG.0b013e318284abb5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Demircioğlu F, Koçyiğit A, Arslan N, Cakmakçi H, Hizli S, Sedat AT. Intima-media thickness of carotid artery and susceptibility to atherosclerosis in obese children with nonalcoholic fatty liver disease. J Pediatr Gastroenterol Nutr. 2008;47:68–75. doi: 10.1097/MPG.0b013e31816232c9. [DOI] [PubMed] [Google Scholar]
- 12.Iughetti L, De Simone M, Verrotti A, Iezzi ML, Predieri B, Bruzzi P, Bernasconi S, Balli F, Bedogni G. Thirty-year persistence of obesity after presentation to a pediatric obesity clinic. Ann Hum Biol. 2008;35:439–448. doi: 10.1080/03014460802232714. [DOI] [PubMed] [Google Scholar]
- 13.Balakrishnan PL. Identification of obesity and cardiovascular risk factors in childhood and adolescence. Pediatr Clin North Am. 2014;61:153–171. doi: 10.1016/j.pcl.2013.09.013. [DOI] [PubMed] [Google Scholar]
- 14.Gulati S, Misra A, Colles SL, Kondal D, Gupta N, Goel K, Bansal S, Mishra M, Madkaikar V, Bhardwaj S. Dietary intakes and familial correlates of overweight/obesity: a four-cities study in India. Ann Nutr Metab. 2013;62:279–290. doi: 10.1159/000346554. [DOI] [PubMed] [Google Scholar]
- 15.Jafar TH, Qadri Z, Islam M, Hatcher J, Bhutta ZA, Chaturvedi N. Rise in childhood obesity with persistently high rates of undernutrition among urban school-aged Indo-Asian children. Arch Dis Child. 2008;93:373–378. doi: 10.1136/adc.2007.125641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Moraes SA, Beltrán Rosas J, Mondini L, Freitas IC. [Prevalence of overweight and obesity, and associated factors in school children from urban area in Chilpancingo, Guerrero, Mexico, 2004] Cad Saude Publica. 2006;22:1289–1301. doi: 10.1590/s0102-311x2006000600018. [DOI] [PubMed] [Google Scholar]
- 17.Ogden CL, Carroll MD, Kit BK, Flegal KM. Prevalence of childhood and adult obesity in the United States, 2011-2012. JAMA. 2014;311:806–814. doi: 10.1001/jama.2014.732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bugianesi E. Non-alcoholic steatohepatitis and cancer. Clin Liver Dis. 2007;11:191–207, x-xi. doi: 10.1016/j.cld.2007.02.006. [DOI] [PubMed] [Google Scholar]
- 19.Kikuchi L, Oliveira CP, Carrilho FJ. Nonalcoholic fatty liver disease and hepatocellular carcinoma. Biomed Res Int. 2014;2014:106247. doi: 10.1155/2014/106247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Calle EE, Rodriguez C, Walker-Thurmond K, Thun MJ. Overweight, obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N Engl J Med. 2003;348:1625–1638. doi: 10.1056/NEJMoa021423. [DOI] [PubMed] [Google Scholar]
- 21.Nair S, Mason A, Eason J, Loss G, Perrillo RP. Is obesity an independent risk factor for hepatocellular carcinoma in cirrhosis? Hepatology. 2002;36:150–155. doi: 10.1053/jhep.2002.33713. [DOI] [PubMed] [Google Scholar]
- 22.Berentzen TL, Gamborg M, Holst C, Sørensen TI, Baker JL. Body mass index in childhood and adult risk of primary liver cancer. J Hepatol. 2014;60:325–330. doi: 10.1016/j.jhep.2013.09.015. [DOI] [PubMed] [Google Scholar]
- 23.Adams LA, Lymp JF, St Sauver J, Sanderson SO, Lindor KD, Feldstein A, Angulo P. The natural history of nonalcoholic fatty liver disease: a population-based cohort study. Gastroenterology. 2005;129:113–121. doi: 10.1053/j.gastro.2005.04.014. [DOI] [PubMed] [Google Scholar]
- 24.Speliotes EK, Willer CJ, Berndt SI, Monda KL, Thorleifsson G, Jackson AU, Lango Allen H, Lindgren CM, Luan J, Mägi R, et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat Genet. 2010;42:937–948. doi: 10.1038/ng.686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gregor MF, Hotamisligil GS. Thematic review series: Adipocyte Biology. Adipocyte stress: the endoplasmic reticulum and metabolic disease. J Lipid Res. 2007;48:1905–1914. doi: 10.1194/jlr.R700007-JLR200. [DOI] [PubMed] [Google Scholar]
- 26.Arslan N, Erdur B, Aydin A. Hormones and cytokines in childhood obesity. Indian Pediatr. 2010;47:829–839. doi: 10.1007/s13312-010-0142-y. [DOI] [PubMed] [Google Scholar]
- 27.Machado MV, Coutinho J, Carepa F, Costa A, Proença H, Cortez-Pinto H. How adiponectin, leptin, and ghrelin orchestrate together and correlate with the severity of nonalcoholic fatty liver disease. Eur J Gastroenterol Hepatol. 2012;24:1166–1172. doi: 10.1097/MEG.0b013e32835609b0. [DOI] [PubMed] [Google Scholar]
- 28.Neish AS. Microbes in gastrointestinal health and disease. Gastroenterology. 2009;136:65–80. doi: 10.1053/j.gastro.2008.10.080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hooper LV, Gordon JI. Commensal host-bacterial relationships in the gut. Science. 2001;292:1115–1118. doi: 10.1126/science.1058709. [DOI] [PubMed] [Google Scholar]
- 30.Zoetendal EG, Vaughan EE, de Vos WM. A microbial world within us. Mol Microbiol. 2006;59:1639–1650. doi: 10.1111/j.1365-2958.2006.05056.x. [DOI] [PubMed] [Google Scholar]
- 31.Chan YK, Estaki M, Gibson DL. Clinical consequences of diet-induced dysbiosis. Ann Nutr Metab. 2013;63 Suppl 2:28–40. doi: 10.1159/000354902. [DOI] [PubMed] [Google Scholar]
- 32.Sekirov I, Russell SL, Antunes LC, Finlay BB. Gut microbiota in health and disease. Physiol Rev. 2010;90:859–904. doi: 10.1152/physrev.00045.2009. [DOI] [PubMed] [Google Scholar]
- 33.Clemente JC, Ursell LK, Parfrey LW, Knight R. The impact of the gut microbiota on human health: an integrative view. Cell. 2012;148:1258–1270. doi: 10.1016/j.cell.2012.01.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444:1022–1023. doi: 10.1038/4441022a. [DOI] [PubMed] [Google Scholar]
- 35.Knights D, Lassen KG, Xavier RJ. Advances in inflammatory bowel disease pathogenesis: linking host genetics and the microbiome. Gut. 2013;62:1505–1510. doi: 10.1136/gutjnl-2012-303954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci USA. 2005;102:11070–11075. doi: 10.1073/pnas.0504978102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Angelakis E, Armougom F, Million M, Raoult D. The relationship between gut microbiota and weight gain in humans. Future Microbiol. 2012;7:91–109. doi: 10.2217/fmb.11.142. [DOI] [PubMed] [Google Scholar]
- 38.Sweeney TE, Morton JM. The human gut microbiome: a review of the effect of obesity and surgically induced weight loss. JAMA Surg. 2013;148:563–569. doi: 10.1001/jamasurg.2013.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Verdam FJ, Fuentes S, de Jonge C, Zoetendal EG, Erbil R, Greve JW, Buurman WA, de Vos WM, Rensen SS. Human intestinal microbiota composition is associated with local and systemic inflammation in obesity. Obesity (Silver Spring) 2013;21:E607–E615. doi: 10.1002/oby.20466. [DOI] [PubMed] [Google Scholar]
- 40.Bervoets L, Van Hoorenbeeck K, Kortleven I, Van Noten C, Hens N, Vael C, Goossens H, Desager KN, Vankerckhoven V. Differences in gut microbiota composition between obese and lean children: a cross-sectional study. Gut Pathog. 2013;5:10. doi: 10.1186/1757-4749-5-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Xu P, Li M, Zhang J, Zhang T. Correlation of intestinal microbiota with overweight and obesity in Kazakh school children. BMC Microbiol. 2012;12:283. doi: 10.1186/1471-2180-12-283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Santacruz A, Marcos A, Wärnberg J, Martí A, Martin-Matillas M, Campoy C, Moreno LA, Veiga O, Redondo-Figuero C, Garagorri JM, et al. Interplay between weight loss and gut microbiota composition in overweight adolescents. Obesity (Silver Spring) 2009;17:1906–1915. doi: 10.1038/oby.2009.112. [DOI] [PubMed] [Google Scholar]
- 43.Abdallah Ismail N, Ragab SH, Abd Elbaky A, Shoeib AR, Alhosary Y, Fekry D. Frequency of Firmicutes and Bacteroidetes in gut microbiota in obese and normal weight Egyptian children and adults. Arch Med Sci. 2011;7:501–507. doi: 10.5114/aoms.2011.23418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kalliomäki M, Collado MC, Salminen S, Isolauri E. Early differences in fecal microbiota composition in children may predict overweight. Am J Clin Nutr. 2008;87:534–538. doi: 10.1093/ajcn/87.3.534. [DOI] [PubMed] [Google Scholar]
- 45.Karlsson CL, Onnerfält J, Xu J, Molin G, Ahrné S, Thorngren-Jerneck K. The microbiota of the gut in preschool children with normal and excessive body weight. Obesity (Silver Spring) 2012;20:2257–2261. doi: 10.1038/oby.2012.110. [DOI] [PubMed] [Google Scholar]
- 46.Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, Neyrinck AM, Fava F, Tuohy KM, Chabo C, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761–1772. doi: 10.2337/db06-1491. [DOI] [PubMed] [Google Scholar]
- 47.Nadal I, Santacruz A, Marcos A, Warnberg J, Garagorri JM, Moreno LA, Martin-Matillas M, Campoy C, Martí A, Moleres A, et al. Shifts in clostridia, bacteroides and immunoglobulin-coating fecal bacteria associated with weight loss in obese adolescents. Int J Obes (Lond) 2009;33:758–767. doi: 10.1038/ijo.2008.260. [DOI] [PubMed] [Google Scholar]
- 48.Esteve E, Ricart W, Fernández-Real JM. Gut microbiota interactions with obesity, insulin resistance and type 2 diabetes: did gut microbiote co-evolve with insulin resistance? Curr Opin Clin Nutr Metab Care. 2011;14:483–490. doi: 10.1097/MCO.0b013e328348c06d. [DOI] [PubMed] [Google Scholar]
- 49.Wostmann BS, Larkin C, Moriarty A, Bruckner-Kardoss E. Dietary intake, energy metabolism, and excretory losses of adult male germfree Wistar rats. Lab Anim Sci. 1983;33:46–50. [PubMed] [Google Scholar]
- 50.Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444:1027–1031. doi: 10.1038/nature05414. [DOI] [PubMed] [Google Scholar]
- 51.Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, Sogin ML, Jones WJ, Roe BA, Affourtit JP, et al. A core gut microbiome in obese and lean twins. Nature. 2009;457:480–484. doi: 10.1038/nature07540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Delzenne NM, Cani PD. Interaction between obesity and the gut microbiota: relevance in nutrition. Annu Rev Nutr. 2011;31:15–31. doi: 10.1146/annurev-nutr-072610-145146. [DOI] [PubMed] [Google Scholar]
- 53.Al-Lahham SH, Peppelenbosch MP, Roelofsen H, Vonk RJ, Venema K. Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim Biophys Acta. 2010;1801:1175–1183. doi: 10.1016/j.bbalip.2010.07.007. [DOI] [PubMed] [Google Scholar]
- 54.Delzenne NM, Williams CM. Prebiotics and lipid metabolism. Curr Opin Lipidol. 2002;13:61–67. doi: 10.1097/00041433-200202000-00009. [DOI] [PubMed] [Google Scholar]
- 55.Le Poul E, Loison C, Struyf S, Springael JY, Lannoy V, Decobecq ME, Brezillon S, Dupriez V, Vassart G, Van Damme J, et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J Biol Chem. 2003;278:25481–25489. doi: 10.1074/jbc.M301403200. [DOI] [PubMed] [Google Scholar]
- 56.Stoddart LA, Smith NJ, Milligan G. International Union of Pharmacology. LXXI. Free fatty acid receptors FFA1, -2, and -3: pharmacology and pathophysiological functions. Pharmacol Rev. 2008;60:405–417. doi: 10.1124/pr.108.00802. [DOI] [PubMed] [Google Scholar]
- 57.Bjursell M, Admyre T, Göransson M, Marley AE, Smith DM, Oscarsson J, Bohlooly-Y M. Improved glucose control and reduced body fat mass in free fatty acid receptor 2-deficient mice fed a high-fat diet. Am J Physiol Endocrinol Metab. 2011;300:E211–E220. doi: 10.1152/ajpendo.00229.2010. [DOI] [PubMed] [Google Scholar]
- 58.Ge H, Li X, Weiszmann J, Wang P, Baribault H, Chen JL, Tian H, Li Y. Activation of G protein-coupled receptor 43 in adipocytes leads to inhibition of lipolysis and suppression of plasma free fatty acids. Endocrinology. 2008;149:4519–4526. doi: 10.1210/en.2008-0059. [DOI] [PubMed] [Google Scholar]
- 59.Jumpertz R, Le DS, Turnbaugh PJ, Trinidad C, Bogardus C, Gordon JI, Krakoff J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am J Clin Nutr. 2011;94:58–65. doi: 10.3945/ajcn.110.010132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, Semenkovich CF, Gordon JI. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA. 2004;101:15718–15723. doi: 10.1073/pnas.0407076101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Mandard S, Zandbergen F, Tan NS, Escher P, Patsouris D, Koenig W, Kleemann R, Bakker A, Veenman F, Wahli W, et al. The direct peroxisome proliferator-activated receptor target fasting-induced adipose factor (FIAF/PGAR/ANGPTL4) is present in blood plasma as a truncated protein that is increased by fenofibrate treatment. J Biol Chem. 2004;279:34411–34420. doi: 10.1074/jbc.M403058200. [DOI] [PubMed] [Google Scholar]
- 62.Harris K, Kassis A, Major G, Chou CJ. Is the gut microbiota a new factor contributing to obesity and its metabolic disorders? J Obes. 2012;2012:879151. doi: 10.1155/2012/879151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Arslan N, Tokgoz Y, Kume T, Bulbul M, Sayın O, Harmancı D, Akdogan GG. Evaluation of serum neopterin levels and its relationship with adipokines in pediatric obesity-related nonalcoholic fatty liver disease and healthy adolescents. J Pediatr Endocrinol Metab. 2013;26:1141–1147. doi: 10.1515/jpem-2013-0029. [DOI] [PubMed] [Google Scholar]
- 64.Wang XA, Zhang R, She ZG, Zhang XF, Jiang DS, Wang T, Gao L, Deng W, Zhang SM, Zhu LH, et al. Interferon regulatory factor 3 constrains IKKβ/NF-κB signaling to alleviate hepatic steatosis and insulin resistance. Hepatology. 2014;59:870–885. doi: 10.1002/hep.26751. [DOI] [PubMed] [Google Scholar]
- 65.Leon-Cabrera S, Solís-Lozano L, Suárez-Álvarez K, González-Chávez A, Béjar YL, Robles-Díaz G, Escobedo G. Hyperleptinemia is associated with parameters of low-grade systemic inflammation and metabolic dysfunction in obese human beings. Front Integr Neurosci. 2013;7:62. doi: 10.3389/fnint.2013.00062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Machado MV, Cortez-Pinto H. Gut microbiota and nonalcoholic fatty liver disease. Ann Hepatol. 2012;11:440–449. [PubMed] [Google Scholar]
- 67.Miele L, Marrone G, Lauritano C, Cefalo C, Gasbarrini A, Day C, Grieco A. Gut-liver axis and microbiota in NAFLD: insight pathophysiology for novel therapeutic target. Curr Pharm Des. 2013;19:5314–5324. [PubMed] [Google Scholar]
- 68.Ilan Y. Leaky gut and the liver: a role for bacterial translocation in nonalcoholic steatohepatitis. World J Gastroenterol. 2012;18:2609–2618. doi: 10.3748/wjg.v18.i21.2609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.de La Serre CB, Ellis CL, Lee J, Hartman AL, Rutledge JC, Raybould HE. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am J Physiol Gastrointest Liver Physiol. 2010;299:G440–G448. doi: 10.1152/ajpgi.00098.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S, Srinivasan S, Sitaraman SV, Knight R, Ley RE, Gewirtz AT. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science. 2010;328:228–231. doi: 10.1126/science.1179721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Lam YY, Ha CW, Campbell CR, Mitchell AJ, Dinudom A, Oscarsson J, Cook DI, Hunt NH, Caterson ID, Holmes AJ, et al. Increased gut permeability and microbiota change associate with mesenteric fat inflammation and metabolic dysfunction in diet-induced obese mice. PLoS One. 2012;7:e34233. doi: 10.1371/journal.pone.0034233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Erridge C, Attina T, Spickett CM, Webb DJ. A high-fat meal induces low-grade endotoxemia: evidence of a novel mechanism of postprandial inflammation. Am J Clin Nutr. 2007;86:1286–1292. doi: 10.1093/ajcn/86.5.1286. [DOI] [PubMed] [Google Scholar]
- 73.Hawkesworth S, Moore SE, Fulford AJ, Barclay GR, Darboe AA, Mark H, Nyan OA, Prentice AM. Evidence for metabolic endotoxemia in obese and diabetic Gambian women. Nutr Diabetes. 2013;3:e83. doi: 10.1038/nutd.2013.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Harte AL, da Silva NF, Creely SJ, McGee KC, Billyard T, Youssef-Elabd EM, Tripathi G, Ashour E, Abdalla MS, Sharada HM, et al. Elevated endotoxin levels in non-alcoholic fatty liver disease. J Inflamm (Lond) 2010;7:15. doi: 10.1186/1476-9255-7-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Verdam FJ, Rensen SS, Driessen A, Greve JW, Buurman WA. Novel evidence for chronic exposure to endotoxin in human nonalcoholic steatohepatitis. J Clin Gastroenterol. 2011;45:149–152. doi: 10.1097/MCG.0b013e3181e12c24. [DOI] [PubMed] [Google Scholar]
- 76.Samuel BS, Shaito A, Motoike T, Rey FE, Backhed F, Manchester JK, Hammer RE, Williams SC, Crowley J, Yanagisawa M, et al. Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor, Gpr41. Proc Natl Acad Sci USA. 2008;105:16767–16772. doi: 10.1073/pnas.0808567105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Delzenne NM, Cani PD, Daubioul C, Neyrinck AM. Impact of inulin and oligofructose on gastrointestinal peptides. Br J Nutr. 2005;93 Suppl 1:S157–S161. doi: 10.1079/bjn20041342. [DOI] [PubMed] [Google Scholar]
- 78.Daubioul CA, Taper HS, De Wispelaere LD, Delzenne NM. Dietary oligofructose lessens hepatic steatosis, but does not prevent hypertriglyceridemia in obese zucker rats. J Nutr. 2000;130:1314–1319. doi: 10.1093/jn/130.5.1314. [DOI] [PubMed] [Google Scholar]
- 79.Hoefert B. Uber die bakterienbefunde im duodenalsaft von gesunden und kraken. Zschr Klin Med. 1921;92:221–235. [Google Scholar]
- 80.Cope K, Risby T, Diehl AM. Increased gastrointestinal ethanol production in obese mice: implications for fatty liver disease pathogenesis. Gastroenterology. 2000;119:1340–1347. doi: 10.1053/gast.2000.19267. [DOI] [PubMed] [Google Scholar]
- 81.Rivera CA, Adegboyega P, van Rooijen N, Tagalicud A, Allman M, Wallace M. Toll-like receptor-4 signaling and Kupffer cells play pivotal roles in the pathogenesis of non-alcoholic steatohepatitis. J Hepatol. 2007;47:571–579. doi: 10.1016/j.jhep.2007.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Swann JR, Want EJ, Geier FM, Spagou K, Wilson ID, Sidaway JE, Nicholson JK, Holmes E. Systemic gut microbial modulation of bile acid metabolism in host tissue compartments. Proc Natl Acad Sci USA. 2011;108 Suppl 1:4523–4530. doi: 10.1073/pnas.1006734107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Sabaté JM, Jouët P, Harnois F, Mechler C, Msika S, Grossin M, Coffin B. High prevalence of small intestinal bacterial overgrowth in patients with morbid obesity: a contributor to severe hepatic steatosis. Obes Surg. 2008;18:371–377. doi: 10.1007/s11695-007-9398-2. [DOI] [PubMed] [Google Scholar]
- 84.Miele L, Valenza V, La Torre G, Montalto M, Cammarota G, Ricci R, Mascianà R, Forgione A, Gabrieli ML, Perotti G, et al. Increased intestinal permeability and tight junction alterations in nonalcoholic fatty liver disease. Hepatology. 2009;49:1877–1887. doi: 10.1002/hep.22848. [DOI] [PubMed] [Google Scholar]
- 85.Mouzaki M, Comelli EM, Arendt BM, Bonengel J, Fung SK, Fischer SE, McGilvray ID, Allard JP. Intestinal microbiota in patients with nonalcoholic fatty liver disease. Hepatology. 2013;58:120–127. doi: 10.1002/hep.26319. [DOI] [PubMed] [Google Scholar]
- 86.Aron-Wisnewsky J, Gaborit B, Dutour A, Clement K. Gut microbiota and non-alcoholic fatty liver disease: new insights. Clin Microbiol Infect. 2013;19:338–348. doi: 10.1111/1469-0691.12140. [DOI] [PubMed] [Google Scholar]
- 87.Artis D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat Rev Immunol. 2008;8:411–420. doi: 10.1038/nri2316. [DOI] [PubMed] [Google Scholar]
- 88.Lichtman SN, Keku J, Schwab JH, Sartor RB. Hepatic injury associated with small bowel bacterial overgrowth in rats is prevented by metronidazole and tetracycline. Gastroenterology. 1991;100:513–519. doi: 10.1016/0016-5085(91)90224-9. [DOI] [PubMed] [Google Scholar]
- 89.Wigg AJ, Roberts-Thomson IC, Dymock RB, McCarthy PJ, Grose RH, Cummins AG. The role of small intestinal bacterial overgrowth, intestinal permeability, endotoxaemia, and tumour necrosis factor alpha in the pathogenesis of non-alcoholic steatohepatitis. Gut. 2001;48:206–211. doi: 10.1136/gut.48.2.206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Shanab AA, Scully P, Crosbie O, Buckley M, O’Mahony L, Shanahan F, Gazareen S, Murphy E, Quigley EM. Small intestinal bacterial overgrowth in nonalcoholic steatohepatitis: association with toll-like receptor 4 expression and plasma levels of interleukin 8. Dig Dis Sci. 2011;56:1524–1534. doi: 10.1007/s10620-010-1447-3. [DOI] [PubMed] [Google Scholar]
- 91.Gäbele E, Dostert K, Hofmann C, Wiest R, Schölmerich J, Hellerbrand C, Obermeier F. DSS induced colitis increases portal LPS levels and enhances hepatic inflammation and fibrogenesis in experimental NASH. J Hepatol. 2011;55:1391–1399. doi: 10.1016/j.jhep.2011.02.035. [DOI] [PubMed] [Google Scholar]
- 92.Alisi A, Ceccarelli S, Panera N, Nobili V. Causative role of gut microbiota in non-alcoholic fatty liver disease pathogenesis. Front Cell Infect Microbiol. 2012;2:132. doi: 10.3389/fcimb.2012.00132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.De Gottardi A, McCoy KD. Evaluation of the gut barrier to intestinal bacteria in non-alcoholic fatty liver disease. J Hepatol. 2011;55:1181–1183. doi: 10.1016/j.jhep.2011.05.003. [DOI] [PubMed] [Google Scholar]
- 94.Ye D, Li FY, Lam KS, Li H, Jia W, Wang Y, Man K, Lo CM, Li X, Xu A. Toll-like receptor-4 mediates obesity-induced non-alcoholic steatohepatitis through activation of X-box binding protein-1 in mice. Gut. 2012;61:1058–1067. doi: 10.1136/gutjnl-2011-300269. [DOI] [PubMed] [Google Scholar]
- 95.Poggi M, Bastelica D, Gual P, Iglesias MA, Gremeaux T, Knauf C, Peiretti F, Verdier M, Juhan-Vague I, Tanti JF, et al. C3H/HeJ mice carrying a toll-like receptor 4 mutation are protected against the development of insulin resistance in white adipose tissue in response to a high-fat diet. Diabetologia. 2007;50:1267–1276. doi: 10.1007/s00125-007-0654-8. [DOI] [PubMed] [Google Scholar]
- 96.Saberi M, Woods NB, de Luca C, Schenk S, Lu JC, Bandyopadhyay G, Verma IM, Olefsky JM. Hematopoietic cell-specific deletion of toll-like receptor 4 ameliorates hepatic and adipose tissue insulin resistance in high-fat-fed mice. Cell Metab. 2009;10:419–429. doi: 10.1016/j.cmet.2009.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Baffy G. Kupffer cells in non-alcoholic fatty liver disease: the emerging view. J Hepatol. 2009;51:212–223. doi: 10.1016/j.jhep.2009.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Su GL, Klein RD, Aminlari A, Zhang HY, Steinstraesser L, Alarcon WH, Remick DG, Wang SC. Kupffer cell activation by lipopolysaccharide in rats: role for lipopolysaccharide binding protein and toll-like receptor 4. Hepatology. 2000;31:932–936. doi: 10.1053/he.2000.5634. [DOI] [PubMed] [Google Scholar]
- 99.Yang SQ, Lin HZ, Lane MD, Clemens M, Diehl AM. Obesity increases sensitivity to endotoxin liver injury: implications for the pathogenesis of steatohepatitis. Proc Natl Acad Sci USA. 1997;94:2557–2562. doi: 10.1073/pnas.94.6.2557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Rabelo F, Oliveira CP, Faintuch J, Mazo DF, Lima VM, Stefano JT, Barbeiro HV, Soriano FG, Alves VA, Carrilho FJ. Pro- and anti-inflammatory cytokines in steatosis and steatohepatitis. Obes Surg. 2010;20:906–912. doi: 10.1007/s11695-010-0181-4. [DOI] [PubMed] [Google Scholar]
- 101.Aoyama T, Paik YH, Seki E. Toll-like receptor signaling and liver fibrosis. Gastroenterol Res Pract. 2010;2010 doi: 10.1155/2010/192543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Paik YH, Schwabe RF, Bataller R, Russo MP, Jobin C, Brenner DA. Toll-like receptor 4 mediates inflammatory signaling by bacterial lipopolysaccharide in human hepatic stellate cells. Hepatology. 2003;37:1043–1055. doi: 10.1053/jhep.2003.50182. [DOI] [PubMed] [Google Scholar]
- 103.Guo J, Friedman SL. Toll-like receptor 4 signaling in liver injury and hepatic fibrogenesis. Fibrogenesis Tissue Repair. 2010;3:21. doi: 10.1186/1755-1536-3-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Sanyal AJ, Campbell-Sargent C, Mirshahi F, Rizzo WB, Contos MJ, Sterling RK, Luketic VA, Shiffman ML, Clore JN. Nonalcoholic steatohepatitis: association of insulin resistance and mitochondrial abnormalities. Gastroenterology. 2001;120:1183–1192. doi: 10.1053/gast.2001.23256. [DOI] [PubMed] [Google Scholar]
- 105.Franchi L, Muñoz-Planillo R, Núñez G. Sensing and reacting to microbes through the inflammasomes. Nat Immunol. 2012;13:325–332. doi: 10.1038/ni.2231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Martinon F, Mayor A, Tschopp J. The inflammasomes: guardians of the body. Annu Rev Immunol. 2009;27:229–265. doi: 10.1146/annurev.immunol.021908.132715. [DOI] [PubMed] [Google Scholar]
- 107.Zhou R, Yazdi AS, Menu P, Tschopp J. A role for mitochondria in NLRP3 inflammasome activation. Nature. 2011;469:221–225. doi: 10.1038/nature09663. [DOI] [PubMed] [Google Scholar]
- 108.Henao-Mejia J, Elinav E, Jin C, Hao L, Mehal WZ, Strowig T, Thaiss CA, Kau AL, Eisenbarth SC, Jurczak MJ, et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature. 2012;482:179–185. doi: 10.1038/nature10809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Thuy S, Ladurner R, Volynets V, Wagner S, Strahl S, Königsrainer A, Maier KP, Bischoff SC, Bergheim I. Nonalcoholic fatty liver disease in humans is associated with increased plasma endotoxin and plasminogen activator inhibitor 1 concentrations and with fructose intake. J Nutr. 2008;138:1452–1455. doi: 10.1093/jn/138.8.1452. [DOI] [PubMed] [Google Scholar]
- 110.Alisi A, Manco M, Devito R, Piemonte F, Nobili V. Endotoxin and plasminogen activator inhibitor-1 serum levels associated with nonalcoholic steatohepatitis in children. J Pediatr Gastroenterol Nutr. 2010;50:645–649. doi: 10.1097/MPG.0b013e3181c7bdf1. [DOI] [PubMed] [Google Scholar]
- 111.Ruiz AG, Casafont F, Crespo J, Cayón A, Mayorga M, Estebanez A, Fernadez-Escalante JC, Pons-Romero F. Lipopolysaccharide-binding protein plasma levels and liver TNF-alpha gene expression in obese patients: evidence for the potential role of endotoxin in the pathogenesis of non-alcoholic steatohepatitis. Obes Surg. 2007;17:1374–1380. doi: 10.1007/s11695-007-9243-7. [DOI] [PubMed] [Google Scholar]
- 112.Vance DE. Role of phosphatidylcholine biosynthesis in the regulation of lipoprotein homeostasis. Curr Opin Lipidol. 2008;19:229–234. doi: 10.1097/MOL.0b013e3282fee935. [DOI] [PubMed] [Google Scholar]
- 113.Russell WR, Hoyles L, Flint HJ, Dumas ME. Colonic bacterial metabolites and human health. Curr Opin Microbiol. 2013;16:246–254. doi: 10.1016/j.mib.2013.07.002. [DOI] [PubMed] [Google Scholar]
- 114.Buchman AL, Dubin MD, Moukarzel AA, Jenden DJ, Roch M, Rice KM, Gornbein J, Ament ME. Choline deficiency: a cause of hepatic steatosis during parenteral nutrition that can be reversed with intravenous choline supplementation. Hepatology. 1995;22:1399–1403. [PubMed] [Google Scholar]
- 115.Zeisel SH, Wishnok JS, Blusztajn JK. Formation of methylamines from ingested choline and lecithin. J Pharmacol Exp Ther. 1983;225:320–324. [PubMed] [Google Scholar]
- 116.Wang Z, Klipfell E, Bennett BJ, Koeth R, Levison BS, Dugar B, Feldstein AE, Britt EB, Fu X, Chung YM, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472:57–63. doi: 10.1038/nature09922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Spencer MD, Hamp TJ, Reid RW, Fischer LM, Zeisel SH, Fodor AA. Association between composition of the human gastrointestinal microbiome and development of fatty liver with choline deficiency. Gastroenterology. 2011;140:976–986. doi: 10.1053/j.gastro.2010.11.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Dumas ME, Barton RH, Toye A, Cloarec O, Blancher C, Rothwell A, Fearnside J, Tatoud R, Blanc V, Lindon JC, et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc Natl Acad Sci USA. 2006;103:12511–12516. doi: 10.1073/pnas.0601056103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Fuchs M. Non-alcoholic Fatty liver disease: the bile Acid-activated farnesoid x receptor as an emerging treatment target. J Lipids. 2012;2012:934396. doi: 10.1155/2012/934396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Trauner M, Claudel T, Fickert P, Moustafa T, Wagner M. Bile acids as regulators of hepatic lipid and glucose metabolism. Dig Dis. 2010;28:220–224. doi: 10.1159/000282091. [DOI] [PubMed] [Google Scholar]
- 121.Thomas C, Gioiello A, Noriega L, Strehle A, Oury J, Rizzo G, Macchiarulo A, Yamamoto H, Mataki C, Pruzanski M, et al. TGR5-mediated bile acid sensing controls glucose homeostasis. Cell Metab. 2009;10:167–177. doi: 10.1016/j.cmet.2009.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Devkota S, Wang Y, Musch MW, Leone V, Fehlner-Peach H, Nadimpalli A, Antonopoulos DA, Jabri B, Chang EB. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature. 2012;487:104–108. doi: 10.1038/nature11225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Turnbaugh PJ. Microbiology: fat, bile and gut microbes. Nature. 2012;487:47–48. doi: 10.1038/487047a. [DOI] [PubMed] [Google Scholar]
- 124.Gustot T, Lemmers A, Moreno C, Nagy N, Quertinmont E, Nicaise C, Franchimont D, Louis H, Devière J, Le Moine O. Differential liver sensitization to toll-like receptor pathways in mice with alcoholic fatty liver. Hepatology. 2006;43:989–1000. doi: 10.1002/hep.21138. [DOI] [PubMed] [Google Scholar]
- 125.Volynets V, Küper MA, Strahl S, Maier IB, Spruss A, Wagnerberger S, Königsrainer A, Bischoff SC, Bergheim I. Nutrition, intestinal permeability, and blood ethanol levels are altered in patients with nonalcoholic fatty liver disease (NAFLD) Dig Dis Sci. 2012;57:1932–1941. doi: 10.1007/s10620-012-2112-9. [DOI] [PubMed] [Google Scholar]
- 126.Nair S, Cope K, Risby TH, Diehl AM. Obesity and female gender increase breath ethanol concentration: potential implications for the pathogenesis of nonalcoholic steatohepatitis. Am J Gastroenterol. 2001;96:1200–1204. doi: 10.1111/j.1572-0241.2001.03702.x. [DOI] [PubMed] [Google Scholar]
- 127.Zhu L, Baker SS, Gill C, Liu W, Alkhouri R, Baker RD, Gill SR. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: a connection between endogenous alcohol and NASH. Hepatology. 2013;57:601–609. doi: 10.1002/hep.26093. [DOI] [PubMed] [Google Scholar]
- 128.Forsyth CB, Tang Y, Shaikh M, Zhang L, Keshavarzian A. Role of snail activation in alcohol-induced iNOS-mediated disruption of intestinal epithelial cell permeability. Alcohol Clin Exp Res. 2011;35:1635–1643. doi: 10.1111/j.1530-0277.2011.01510.x. [DOI] [PMC free article] [PubMed] [Google Scholar]