

Abstract
Studies in animal models have shown that repeated episodes of alcohol dependence and withdrawal promote escalation of drinking that is presumably associated with alterations in the addiction neurocircuitry. Using a lithium chloride-ethanol pairing procedure to devalue the reinforcing properties of ethanol, the present study determined whether multiple cycles of chronic intermittent ethanol (CIE) exposure by vapor inhalation also alters the sensitivity of drinking behavior to the devaluation of ethanol's reinforcing effects. The effect of devaluation on operant ethanol self-administration and extinction was examined in mice prior to initiation of CIE (short drinking history) and after repeated cycles of CIE or air control exposure (long drinking history). Devaluation significantly attenuated the recovery of baseline ethanol self-administration when tested either prior to CIE or in the air-exposed controls that had experienced repeated bouts of drinking but no CIE. In contrast, in mice that had undergone repeated cycles of CIE exposure that promoted escalation of ethanol drinking, self-administration was completely resistant to the effect of devaluation. Devaluation had no effect on the time course of extinction training in either pre-CIE or post-CIE mice. Taken together, these results are consistent with the suggestion that repeated cycles of ethanol dependence and withdrawal produce escalation of ethanol self-administration that is associated with a change in sensitivity to devaluation of the reinforcing properties of ethanol.
Keywords: operant conditioning, ethanol self-administration, ethanol dependence, Pavlovian conditioning, mice
Introduction
Alcoholism and alcohol abuse are a major problem worldwide with devastating health and societal consequences. While most individuals who consume alcohol can control their intake, a significant proportion of individuals cannot control their consumption and subsequently develop an addiction to alcohol that is characterized by uncontrollable and compulsive consumption. While studies have demonstrated that both positive and negative reinforcing properties of ethanol contribute to and promote self-administration (Becker, 2013; Heilig, Egli, Crabbe, & Becker, 2010; Koob, 2003; Meish, 1982, 1984), accumulating evidence in rodents indicates that prolonged self-administration of ethanol is also associated with the development of resistance to aversive stimuli associated with ethanol. For example, a study conducted in C57BL/6J mice showed that addition of quinine (an aversive tastant) to the ethanol solution lowered ethanol intake in mice with a short (2-week) history of voluntary ethanol intake. In contrast, when mice were allowed to self-administer ethanol for a longer period of time (8 weeks), they were no longer affected by this quinine-induced adulteration of ethanol, and continued to self-administer it despite the aversive taste (Lesscher, van Kerkhof, & Vanderschuren, 2010). Similar results were obtained in rats that had been drinking ethanol adulterated with quinine after a short (1.5 months) versus a long (3–4 months) history of ethanol self-administration (Hopf, Chang, Sparta, Bowers, & Bonci, 2010).
In addition to the differences in drinking behaviors observed in animals with a short versus long history of drinking, recent evidence has demonstrated that models of ethanol dependence that involve multiple episodes of withdrawal also promote changes in drinking behavior. For example, it was reported that subjecting rats to repeated cycles of chronic ethanol exposure and withdrawal resulted in resistance to the effect of adulteration of ethanol with quinine (Vendruscolo et al., 2012). In addition, studies in mice have shown that repeated cycles of chronic intermittent ethanol (CIE) exposure promote increases in voluntary ethanol consumption when tested using various 2-bottle choice drinking procedures (Becker & Lopez, 2004; Finn et al., 2007; Griffin, Lopez, & Becker, 2009; Griffin, Lopez, Yanke, Middaugh, & Becker, 2009; Lopez & Becker, 2005; Lopez, Griffin, Melendez, & Becker, 2012). Importantly, this heightened ethanol intake after CIE exposure is associated with increased blood (Becker & Lopez, 2004) and brain (Griffin, Lopez, Yanke, et al., 2009) ethanol concentrations compared to that measured in non-dependent mice that consume more moderate amounts of ethanol. In a recent study, we further observed that mice trained to lever press for ethanol using a fixed-ratio (FR) schedule exhibited significant increases in ethanol self-administration after CIE exposure (Lopez & Becker, under review). This CIE-induced elevation of operant oral ethanol self-administration engendered responding that was resistant to extinction and more sensitive to cue-induced reinstatement. While CIE-exposure models have clearly demonstrated escalated ethanol self-administration, it is not clear whether this augmented consumption reflects a change in the reinforcing value of ethanol. To begin to address this question, the present study examined whether the CIE-induced escalation of operant self-administration is also associated with changes in response to devaluation of the reinforcing properties of ethanol.
Methods
Subjects
Adult male C57BL/6 mice purchased from Jackson Laboratories (Bar Harbor, ME) were individually housed with free access to food (Harland Teklad, Madison, WI) and water throughout all phases of the experiments. Body weights were recorded weekly during ethanol self-administration or daily during chronic intermittent ethanol (CIE) or air exposure (detailed below). Mice were housed in a temperature- and humidity-controlled animal facility under a reversed 12-h light/dark cycle (lights on at 1800 h). All procedures were approved by the Institutional Animal Care and Use Committee and followed the NIH Guide for the Care and Use of Laboratory Animals (8th edition, National Research Council, 2011).
Study design
The general study design (Fig. 1) involved examination of the effects of devaluation on operant ethanol self-administration and extinction before or after repeated cycles of CIE or air control exposure. Mice tested before CIE exposure had 6 weeks of ethanol self-administration and are referred to as short-drinking history mice, and those tested after repeated CIE or air exposure as long-drinking history mice (10 weeks of ethanol self-administration). The study involved first establishing stable baseline levels of operant ethanol responding and intake, as described below. Mice were then separated into 2 groups to be exposed to either weekly cycles of CIE exposure (CIE group) or air control (CTL group) exposure. Seventy-two hours after each cycle of CIE (or air control) exposure, mice resumed ethanol self-administration for 5 consecutive days. This pattern of exposure followed by 5 days of ethanol self-administration was repeated for 4 cycles until the mice were used in studies to evaluate the effect of devaluation of ethanol on drinking behavior. Separate groups of mice received devaluation treatment after either baseline or a fifth cycle of CIE (or air control) exposure (see schema in Fig. 1). Mice were first evaluated for ethanol self-administration or extinction responding the day before initiation of the devaluation procedure. Seventy-two hours after the last devaluation session, mice were tested under regular self-administration or extinction conditions.
Figure 1.

Schematic depiction of the experimental design of the study. A. The effect of ethanol devaluation on ethanol self-administration and extinction was examined in mice with a short history of drinking prior to initiation of CIE and in mice with a long history of drinking after 5 cycles of CIE or air control exposure. B. In separate groups of mice, the effect of devaluation of the rewarding properties of ethanol was examined on operant ethanol self-administration (top) and extinction (bottom).
Operant Ethanol Self-Administration
Standard chambers for mouse operant conditioning were used (Med Associates, Inc., St. Albans, VT). The operant boxes were housed in sound-attenuating boxes with exhaust fans. Stimulus events and responses were controlled and monitored using Med PC, Version IV software (Med Associates, Inc., St. Albans, VT). Mice were not food- or water-deprived and were trained to self-administer ethanol using a modified sucrose fading procedure (Samson, 1986). All operant self-administration sessions were 30 min and were conducted 5 days per week (Mon-Fri) during the dark period of the light/dark cycle. Mice were placed in the operant box and the session began by introducing 2 levers into the box and turning on the house light. Once the subject pressed the active lever, an infusion pump was activated for 2 s to deliver the reinforcer into the well between the 2 retractable levers. For the duration of pump activation, ethanol delivery was cued by turning off the house light and presentation of a tone (80 dB) and yellow stimulus light. Responses on the inactive lever had no consequences. Initially, mice were trained to lever press under a Fixed Ratio 1 (FR1) schedule for the delivery of 20 μL of a 10% (v/v) ethanol solution combined with 5% (w/v) sucrose (10E/5S) during the daily 30-min sessions. Over successive days, the FR requirement was increased to 2 and finally to 4 responses on the active lever (FR4). After this initial period (4–5 days), the ethanol solution was increased and the sucrose solution reduced using 12E/5S, 12E/2S for 2 or 3 days each, and then 12E/1S was used for the remainder of the study. At the end of each session, any residual ethanol that was left in the well was measured and the amount was subtracted from the total self-administration volume. This corrected volume was used to calculate g/kg of ethanol intake during the session. Ethanol intake (g/kg) and number of responses on the active and inactive levers were recorded. After 2 weeks of sucrose fading, daily (Mon–Fri) ethanol self-administration sessions continued until mice reached stable ethanol self-administration (< 15% change over 3 consecutive days in the number of responses). The final week of FR4 responding was used as a baseline for data analysis purposes. Mice resumed ethanol self-administration under the same conditions for 5 consecutive days after each CIE exposure cycle. During extinction sessions, neither ethanol nor ethanol-associated cues (tone and lights) were presented after lever pressing.
CIE exposure
Mice in the CIE group were exposed to ethanol vapor in inhalation chambers for 16 h a day separated by an 8-h withdrawal period for 4 consecutive days. CTL mice were exposed to uncontaminated air in control inhalation chambers. The design of the inhalation chambers was similar to that previously described (Becker & Hale, 1993; Becker & Lopez, 2004). Ethanol (95%) was volatilized by passing air through an air stone submerged in ethanol in a flask and the ethanol vapor was delivered to the chambers at a rate of 10 L/min to maintain ethanol concentrations at 15–17 mg/L of air. Chamber ethanol concentrations were monitored daily and assayed as previously described (Lopez & Becker, 2005). The housing conditions in the chambers were identical to those in the colony room. At the beginning of each 16-h exposure bout, ethanol intoxication was initiated by administration of 1.6 g/kg ethanol (8% w/v; intra-peritoneally [i.p.]) and the alcohol dehydrogenase inhibitor pyrazole (1 mmol/kg; i.p.) to help stabilize blood ethanol concentrations (BEC). CTL mice were injected with pyrazole as well. All injections were given in a volume of 20 mL/kg body weight. During each CIE cycle, blood samples (~40 μL) were collected from the orbital sinus with heparinized capillary tubes. Blood was centrifuged and plasma was used to measure ethanol concentrations (mg/dL) using an Analox Instrument analyzer (Lunenburg, MA). In both experiments, blood ethanol concentrations during CIE cycles were maintained between 150 and 225 mg/dL.
Devaluation procedure
This procedure was based on previous studies conducted with rats in which the post-ingestive effects of ethanol were paired with an aversive stimulus in order to devalue the reinforcing effects of ethanol (Samson et al., 2004). Mice in the ethanol-devalued condition (DEV group) received 4 daily intragastric (i.g.) ethanol treatments (g/kg dose adjusted to that consumed during the last self-administration session) paired 10 min later with an injection of lithium chloride (LiCl) (0.4 M, i.p.). Mice in the non-devalued condition (Non-DEV group) received either water (i.g.) paired with LiCl (i.p.) or ethanol (i.g.) paired with saline (i.p.). Previous pilot experiments used to select the optimal parameters for this devaluation procedure indicated that neither association of water with LiCl or ethanol with saline changed ethanol intake level. As this was also observed in the experiments presented here, these groups were combined in the Non-DEV group (see supplemental figures 1–4). The volume administered of ethanol and water (i.g.), and saline and LiCl (i.p.) was 10 mL/kg in all instances. Mice were assigned to the DEV or Non-DEV group based on their self-administration or extinction level of responding evaluated in a single session to avoid differences between the groups before the devaluation procedure.
Data analysis
Ethanol intake (g/kg) and number of responses on the active lever served as the dependent variables. Responses on the inactive lever during self-administration and extinction sessions were very low and were not included in the analyses. Baseline ethanol responding and intake measured prior to devaluation were analyzed using t test. A mixed ANOVA with Devaluation (Dev, Non-DEV) as a between-subjects factor and Days as a repeated measure was used to analyze data after the devaluation procedure. Data for mice that completed 5 cycles of CIE exposure were analyzed by a mixed factorial ANOVA with Treatment (CIE, CTL) and Devaluation (DEV, Non-DEV) as between-subjects factors and Phase (Baseline, Drinking Test Cycles 1–4) as a repeated measure. Preliminary analyses indicated that there were no significant daily variations in ethanol self-administration and, therefore, data were averaged over the last 5 days of baseline and the 5 days of each testing cycle. Data collected after the devaluation procedure were analyzed with a mixed ANOVA with Treatment (CIE, CTL) and Devaluation (DEV, Non-DEV) as between-subjects factors and Days as a repeated measure. Further analyses of significant main effects and interactions were done by post hoc comparisons using Newman-Keuls test, and significance levels were set at p < 0.05.
Results
Devaluation before CIE exposure
The first set of studies examined the effects of devaluation on ethanol self-administration and extinction responding in mice that experienced a short history of drinking and no CIE exposure. Since mice were selected based on their baseline self-administration levels, there were no differences between DEV (n = 11) and Non-DEV (n = 10) mice in the number of active lever responses [t(19) = 0.01, p = 0.99] or the amount of ethanol (g/kg) consumed [t(19) = 0.18, p = 0.86] on the last self-administration session (data not shown). For these experiments, the mean ± SEM of the number of lever responses was 54.10 ± 10.0 and 54.10 ± 9.3 for the DEV and Non-DEV groups, respectively, and the mean ± SEM for ethanol intake was 0.59 ± 0.11 and 0.61 ± 0.10 g/kg for the DEV and Non-DEV groups, respectively.
Mice were subjected to the 4-day devaluation or sham procedure and then allowed to resume daily sessions of operant ethanol self-administration (Fig. 2). Analysis of the number of responses after devaluation indicated a significant effect of Day [F(4,76) = 15.16, p < 0.01] due to the increase in responding over the daily sessions. Importantly, there was a significant main effect of Devaluation [F(1,19) = 7.52, p < 0.025] such that mice in the DEV group responded significantly less for ethanol than mice in the Non-DEV group. The analysis of ethanol intake (g/kg) showed the same results (data not shown). There was a significant effect of Day [F(4,76) = 17.80, p < 0.01] due to lower intake (g/kg) on the early days after devaluation such that recovery to the pre-devaluation baseline intake was significantly delayed (Fig. 2) and a significant decrease in ethanol intake in the DEV group compared to Non-DEV group [F(1,19) = 5.09, p < 0.05] (data not shown). Taken together, these data demonstrate that mice with a short history of ethanol self-administration were sensitive to devaluation of ethanol reward.
Figure 2.
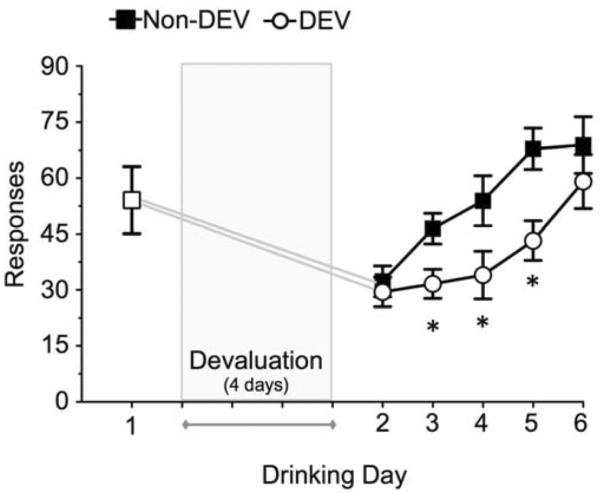
Mice with a short history of operant ethanol self-administration exhibited responding that was sensitive to ethanol devaluation. After obtaining stable baseline drinking, mice were subjected to the ethanol devaluation or sham procedure followed by resumption of ethanol self-administration. Baseline responding (mean ± SEM) for the 30-min self-administration sessions is shown for the day immediately prior to initiation of devaluation. Mice subjected to devaluation (DEV) exhibited a significant reduction in responding for ethanol compared to mice that underwent sham devaluation (Non-DEV). The * indicates significant difference between DEV and Non-DEV groups (ANOVA with Newman-Keuls post hoc test, p < 0.05).
As a complementary measure to the effects of devaluation on drinking, the next set of experiments examined whether devaluation affected extinction responding in a separate group of mice with a short history of drinking prior to CIE exposure (Fig. 3). As shown in Fig. 3, there was no significant difference between DEV (n = 9) and Non-DEV (n = 8) groups for ethanol responding [t(15) = −0.68, p = 0.50] and intake [t(15) = −0.65, p = 0.52] (average active lever responding: 59.00 ± 4.34 vs. 64.58 ± 7.12, and ethanol g/kg intake: 0.73 ± 0.07 vs. 0.81 ± 0.10 g/kg, for the DEV and Non-DEV groups, respectively). Mice were then subjected to a single 30-min extinction session on the day prior to initiation of the devaluation or sham procedure. The purpose of conducting the single extinction test session on the day prior to devaluation was to allow counter-balancing of the mice to ensure that initial extinction responding in the groups were equal. In this way, any subsequent difference in extinction responding could be directly attributed to an effect of devaluation. As shown in Fig. 3, the mean number of responses during this pre-devaluation extinction session did not differ between the DEV and Non-DEV groups [t(15) = −0.26, p = 0.80]. Following the devaluation or non-devaluation control procedure, mice were then subjected to extinction training for 5 consecutive days. Analysis of the number of responses during these sessions revealed a significant effect of Day [F(4,60) = 6.61, p < 0.01] that was due to a progressive decrease in responding over the successive extinction sessions. However, there was no significant effect of devaluation or interaction between Devaluation and Days. Therefore, in contrast to its effect on self-administration of ethanol, devaluation had no effect on the time course of extinction responding.
Figure 3.
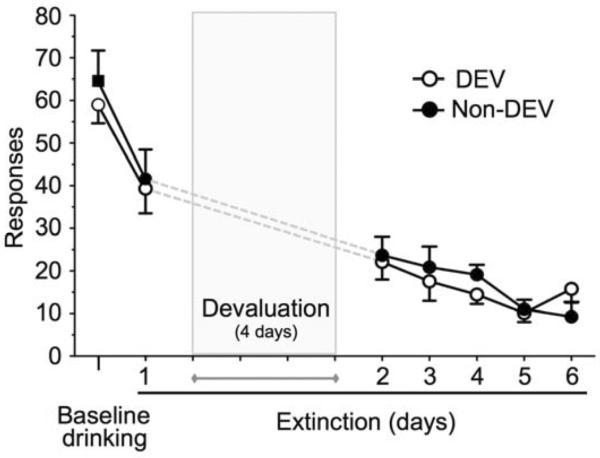
Devaluation of ethanol reward in mice with a short history of self-administration does not alter extinction responding. Following establishment of baseline drinking and a single day of extinction training, mice were subjected to ethanol or sham devaluation and then resumed extinction training. Shown are the mean ± SEM of baseline responding for ethanol and extinction of ethanol self-administration.
Devaluation after CIE exposure
The second set of studies examined the effects of devaluation of ethanol reward on ethanol self-administration and extinction responding in mice subjected to repeated cycles of CIE (or air control) exposure. Consistent with a recent study from our laboratory (Lopez & Becker, under review), repeated cycles of CIE exposure produced an escalation in ethanol self-administration using operant conditioning procedures. In mice that were used to determine the effects of devaluation on ethanol self-administration behavior (Fig. 4A), there was a significant Treatment × Phase interaction for ethanol responses [F(4,148) = 2.88; p < 0.05] and amount (g/kg) consumed [F(4,148) = 2.95; p < 0.05]. Post hoc comparisons revealed that mice in the CIE group (n = 22) responded and consumed significantly more ethanol during Test Cycle 4 compared to their baseline levels as well as in comparison to the CTL group (n = 19). In contrast, baseline ethanol self-administration in CTL mice remained similar across drinking Test Cycles. In a separate group of mice that were used to determine the effects of devaluation on extinction responding following a history of CIE exposure (Fig. 4B), ANOVA revealed significant Treatment × Phase interactions for active lever responding [F(4,136) = 4.50; p < 0.01] and ethanol consumed [F(4,136) = 5.78; p < 0.01]. Post hoc comparisons again indicated that mice in the CIE group (n = 18) self-administered significantly more ethanol during Test Cycle 4 compared to their own baseline self-administration level and to self-administration in CTL mice (n = 20).
Figure 4.

Repeated cycles of CIE exposure enhance operant self-administration. Shown are the mean ± SEM of responses (left graphs) and intake (g/kg) of ethanol (right graphs) for the experimental groups of mice subsequently used for examination of the effect of devaluation on self-administration (A) and extinction (B). For both groups of mice, baseline responding and intake are shown prior to initiation of CIE and after 4 cycles of CIE or air control exposure. The * indicates significant difference compared to respective control values (ANOVA with Newman-Keuls post hoc test; p < 0.05).
After the demonstration of CIE-induced escalation of drinking after 4 cycles of CIE (Fig. 4A), one set of mice was used to determine the effect of devaluation on ethanol self-administration following a subsequent (5th) CIE or air-control exposure cycle. Examination of self-administration behavior on the day prior to initiation of devaluation (after the 5th inhalation exposure cycle) revealed significantly greater ethanol responding [F(1,37) = 5.26, p < 0.05] (Fig. 5) and consumption [F(1,37) = 9.24, p < 0.01] (Fig. 6) in CIE-exposed mice compared to control mice. In addition, devaluation of ethanol reinforcement in the air control mice transiently reduced ethanol self-administration similar to that observed in mice with a short history of drinking. This was observed as a decrease in ethanol responding (Fig. 5) and ethanol intake (Fig. 6). Analysis of active lever responses revealed significant main effects of Treatment [F(1,37) = 6.19] and Day (Days 2–6) [F(4,148) = 5.05], and a significant Treatment × Devaluation × Day interaction [F(4,148) = 3.38] (all p < 0.05). Similarly, for ethanol consumption, there was a significant main effect of Treatment [F(1,37) = 7.61] and Day [F(4,148) = 6.54] and Treatment × Devaluation × Day interaction [F(4,148) = 3.17] (all p < 0.05). Post hoc analysis revealed significant reductions in ethanol responding (Fig. 5) and ethanol intake (Fig. 6) on test Drinking Day 2 and 3 in the CTL/DEV mice compared to the CTL/No-DEV mice (p < 0.05). In contrast, mice that were subjected to repeated cycles of CIE exposure were no longer sensitive to ethanol devaluation as reflected by the fact that the number of responses and amount of ethanol consumed was similar on the days immediately before and after devaluation. Taken together, these observations demonstrate that repeated cycles of CIE exposure altered sensitivity of mice to devaluation of ethanol reinforcement.
Figure 5.
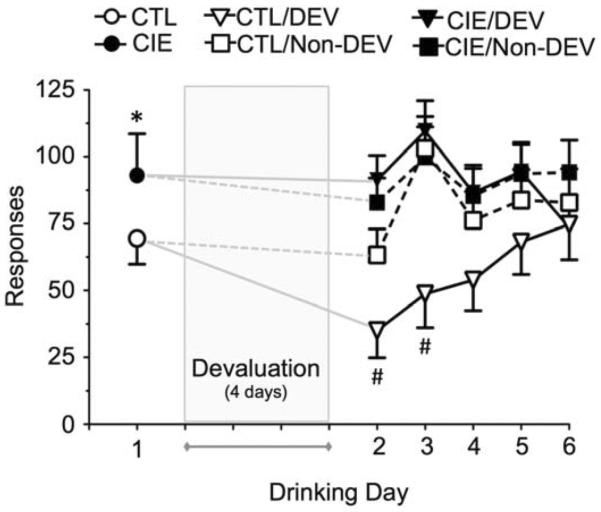
Repeated cycles of CIE promote insensitivity to devaluation of responding for ethanol. Prior to devaluation, mice subjected to 5 cycles of CIE exhibited enhanced responding for ethanol compared to the sham air-exposed controls. After ethanol or sham devaluation, responding in the air-exposed control mice was sensitive to devaluation. In contrast, CIE-exposed mice exhibited responding that was insensitive to devaluation. The * indicates significant increase of CIE versus CTL (t test, p < 0.05). The # indicates significant difference from respective non-devalued controls (ANOVA with Newman-Keuls post hoc test, p < 0.05).
Figure 6.
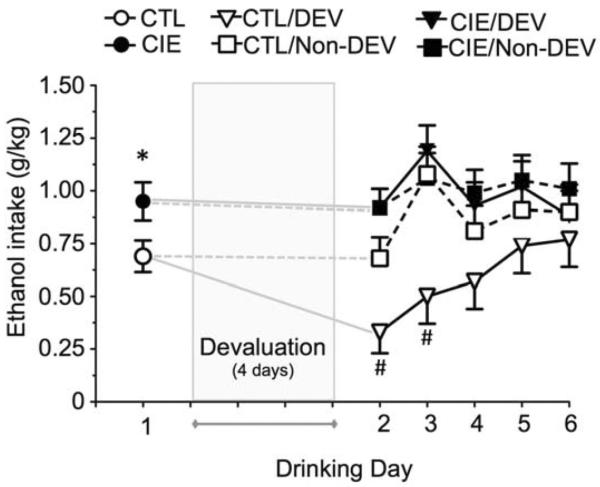
CIE-induced insensitivity to ethanol devaluation correlates with a reduction in ethanol intake. Shown is ethanol consumption (g/kg) for self-administration before and after ethanol or sham devaluation. Prior to devaluation, mice that had undergone 5 cycles of CIE showed enhanced ethanol intake compared to the air-exposed controls. After ethanol- or sham-devaluation, ethanol consumption in the air-exposed control group was sensitive to devaluation. In contrast, ethanol consumption in mice that had undergone CIE exhibited drinking that was not affected by devaluation. The * indicates significant increase of CIE versus CTL (t test, p < 0.05). The # indicates significant difference from respective non-devalued controls (ANOVA with Newman-Keuls post hoc test, p < 0.05)
In the final set of studies, we examined the effects of devaluation on extinction responding in mice with a long history of ethanol self-administration that included access to ethanol both before and after CIE or control air exposure (Fig. 4B). As indicated above, average ethanol responding was significantly greater in CIE-exposed mice compared to CTL mice during the Test Cycle 4. Responding during the single extinction session following the final (5th) exposure cycle – but prior to devaluation – was also significantly higher in the CIE group compared to responding in the control group [F(1,34) = 9.83, p < 0.01] (Fig. 7). Analysis of extinction responding following the devaluation (or sham) procedure revealed a significant main effect of Days [F(4,136) = 25.20, p < 0.01], which reflected a gradual decrease in ethanol responding over days. However, ANOVA did not reveal significant effects of Group or Devaluation, or significant interactions between these factors and Days. Therefore, in contrast to effect of devaluation on ethanol self-administration, repeated cycles of CIE exposure did not alter extinction responding following devaluation.
Figure 7.

Extinction responding in mice following repeated cycles of CIE was not altered by devaluation. Following the 5th cycle of CIE or air exposure, mice were subjected to ethanol devaluation or sham devaluation followed by daily extinction sessions. For ease of visual comparison, baseline responding for ethanol prior to the 5th cycle of CIE is shown. The values are the mean ± SEM lever responses for self-administration or extinction over the 30-min test sessions on each day. The * indicates significant increase of CIE versus CTL responding during pre-devaluation extinction test day (t test, p < 0.05).
Discussion
The present study demonstrated that mice with either a short or a long history of operant ethanol self-administration exhibited sensitivity to devaluation of the reinforcing properties of ethanol. In contrast, when mice were exposed to repeated cycles of CIE exposure, ethanol self-administration behavior transitioned to become insensitive to ethanol devaluation. This CIE-induced insensitivity to ethanol devaluation was also associated with an escalation in the amount of ethanol consumed. Taken together, these observations are consistent with the suggestion that CIE-induced escalation of drinking may involve, at least in part, changes in the role of the reinforcing properties of alcohol.
In the present study, mice were trained to respond for ethanol reinforcement using a fixed-ratio schedule that is known to involve goal-oriented responding. Consistent with this, we found that mice with either a short or a long history of ethanol self-administration were sensitive to the devaluation of the reinforcing effect of ethanol induced by pairing ethanol intoxication with the aversive effects of LiCl. Based on results from a recent study in rats (Corbit, Nie, & Janak, 2012), it was expected that mice with a longer history of ethanol self-administration should show greater resistance to ethanol devaluation. However, an important methodological consideration is that the present studies used a fixed-ratio schedule, while a random ratio schedule was used in the Corbit et al. (2012) study. This difference in the schedule of reinforcement may explain why mice with a short or long history of ethanol self-administration were sensitive to devaluation. In contrast, mice that experienced repeated cycles of CIE exposure exhibited a lack of sensitivity to devaluation of ethanol reward. It is important to note that studies conducted by Samson et al. (2004) demonstrated that the post-ingestive pharmacological effects of ethanol play an important role in modulating responding for ethanol. In the present study, ethanol was delivered intragastrically during the devaluation sessions to reduce the potential confound of ethanol taste. Similar to the effect described by Samson et al. (2004), the fact that mice that experienced CIE exposure continued to respond for and consume ethanol, indicates that devaluation using this procedure did not produce conditioned taste aversion. It could also be argued that chronic exposure to ethanol in the inhalation chamber could have prevented mice from learning the association between ethanol and the illness induced by LiCl during the devaluation procedure. We believe this is unlikely, as we have previously shown that CIE-exposed mice are capable of acquiring a 1-trial conditioned taste aversion involving either a novel taste associated with LiCl or a high dose of ethanol (Lopez et al., 2012). However, CIE exposure may have affected the association of ethanol intoxication that served as conditioned stimulus (CS) with LiCl (unconditioned stimulus, US) in the devaluation procedure through another mechanism. The considerable difference in the amount of experience with ethanol's interoceptive effects between CIE and control mice could have favored a latent inhibition effect (Lubow & Moore, 1959) in which association between the CS and US is weakened by previous exposure to the CS. The experiments presented here are also consistent with a report by Vendruscolo et al. (2012) in which it was observed that ethanol-dependent rats trained to lever respond for ethanol found ethanol more reinforcing (evidenced by higher breaking points in a progressive ratio test) and were more resistant to the adulteration of ethanol by quinine. Importantly, both the present results in mice and the observations in rats (Vendruscolo et al., 2012) suggest that the escalation of ethanol consumption associated with dependence may reflect reduced sensitivity to manipulations that devalue the reinforcing effects of ethanol.
Previous studies of drug taking behavior that utilized a random interval schedule of reinforcement have observed that devaluation results in continued responding for cocaine or alcohol under extinction conditions despite the devaluation of the reinforcer. This lack of sensitivity to devaluation has been interpreted to reflect a habit-like mode of drug-seeking behavior. In the current study, it was observed that devaluation of ethanol reward did not affect extinction responding, whether testing occurred prior to or after CIE exposure. However, this outcome is not surprising since our studies involved training mice to self-administer alcohol using a fixed-ratio schedule of reinforcement – a response-contingent reward paradigm that typically favors goal-oriented (response-outcome) motivated behavior (Samson et al., 2004; Yin & Knowlton, 2006). Another explanation for the lack of difference between groups tested under extinction conditions could be related to the duration of the testing itself. It could be that the extinction session per se was too long and that the testing conducted in separate mice in the 2 devaluation groups was enough to extinguish ethanol seeking.
Another interesting observation in the present study was that mice with a long history of ethanol self-administration along with CIE exposure showed significantly higher levels of responding for ethanol under extinction conditions, suggesting that greater ethanol seeking in dependent (CIE-exposed) mice may reflect greater reinforcing efficacy for ethanol. This is consistent with recent observations in our lab in dependent mice and previous studies in rats showing significantly higher breaking points in a progressive ratio test after chronic intermittent exposure to ethanol vapor (Vendruscolo et al., 2012). As indicated above, mice were trained under a fixed ratio schedule that is known to engender goal-directed behavior. We observed that mice with a short history of ethanol self-administration showed a significant decrease in the number of responses during the first session of extinction training, whereas mice with a long history of ethanol self-administration, and those that experienced CIE exposure, showed an increase in responding during the first extinction session before initiation of the devaluation procedure. It is possible that this single session was sufficient to extinguish ethanol seeking. Therefore, when mice were evaluated after devaluation conditioning was completed, the level of extinction responding was very low and may have obviated the ability to observe an effect of devaluation. In contrast, when mice were allowed to respond for ethanol, re-exposure to the reinforcer (ethanol) allowed for recovery of previous levels of self-administration only in the non-devalued mice.
Studies utilizing various cognitive models have supported the idea that drug use in dependent subjects is governed by automatic (non-cognitive) processes under certain conditions that can be triggered by drug-associated stimuli (e.g., Tiffany, 1990). These conditions include responding when the reinforcing properties of the drug have been devalued (by satiety or outcome devaluation), when responding does not result in delivery of the drug (extinction), or when responding is no longer required to obtain the drug (contingency devaluation). These and other observations provide valuable insight into the mechanisms that may underlie responding for ethanol. It is important to consider that ethanol is typically consumed initially for its positive reinforcing effects, but continued consumption of ethanol over a prolonged period of time can lead to sensitization of these positive effects accompanied by tolerance to ethanol's aversive effects. Furthermore, with the development of dependence, drinking can also be driven by the capacity of ethanol to serve as a negative reinforcer (i.e., drinking to avoid the unpleasant consequences of withdrawal) (Becker, 2013; Koob, 2003). Therefore, the results of the present study add to this literature by demonstrating that repeated cycles of dependence and withdrawal may decrease sensitivity to the devaluation of ethanol's reinforcing properties.
Supplementary Material
Mice with a short history of operant ethanol self-administration exhibited responding that was sensitive to ethanol devaluation. After obtaining stable baseline drinking, mice were subjected to the ethanol devaluation or sham procedure followed by resumption of ethanol self-administration. Baseline responding (mean ± SEM) for the 30-min self-administration sessions is shown for the day immediately prior to initiation of devaluation. Mice subjected to devaluation (DEV) exhibited a significant reduction in responding for ethanol compared to mice that underwent sham devaluation (Non-DEV). The * indicates significant difference between DEV and Non-DEV groups combined (ANOVA with Newman-Keuls post hoc test, p < 0.05). For the Non-DEV group, half of the mice received EtOH (i.g.) followed by a saline injection (i.p.) while the other half received water (i.g.) followed by a LiCl injection (i.p.). A separate ANOVA conducted to evaluate these groups (Non-DEV EtOH-Sal, Non-DEV Water-LiCl) over the 5 drinking days (repeated measure) after devaluation indicated a significant main effect of days [F(4,32) = 5.98, p < .01] due to the gradual increase on responding over days. However, the ANOVA failed to indicate any significant difference between the two Non-DEV groups or interactions with days [F(1,32) = 0.04, and F(4,32) = 0.31, respectively; both p = NS].
Devaluation of ethanol reward in mice with a short history of self-administration does not alter extinction responding. Following establishment of baseline drinking and a single day of extinction training, mice were subjected to ethanol or sham devaluation after which extinction training was resumed. Shown are the mean ± SEM of baseline responding for ethanol and extinction of ethanol self-administration. Half of the mice in the Non-DEV group received EtOH (i.g.) followed by a saline injection (i.p.) while the other half received water (i.g.) followed by a LiCl injection (i.p.). A separate ANOVA conducted to evaluate these groups (Non-DEV EtOH-Sal, Non-DEV Water-LiCl) over the 5 extinction testing days (repeated measure) after devaluation indicated a significant main effect of days [F(4,28) = 6.16, p <. 01] due to the gradual increase on responding over days. However, the ANOVA indicated there was no significant difference between the 2 Non-DEV groups or interactions with days [F(1,28) = 2.28, and F(4,28) = 1.15, respectively; both p = NS].
Repeated cycles of CIE promote insensitivity to devaluation of responding for ethanol. Prior to devaluation, mice subjected to 5 cycles of CIE exhibited enhanced responding for ethanol compared to the sham air-exposed controls. After ethanol or sham devaluation, responding in the air-exposed control mice was sensitive to devaluation. In contrast, CIE-exposed mice exhibited responding that was insensitive to devaluation. The * indicates significant increase of CIE versus CTL (t test, p < 0.05). The # indicates significant difference from respective non-devalued combined controls (ANOVA with Newman Keuls post hoc test, p < 0.05). Half of the mice in the Non-DEV group received EtOH (i.g.) followed by a saline injection (i.p.) while the other half received water (i.g.) followed by a LiCl injection (i.p.). A separate ANOVA was conducted using Group (CIE, CTL), Non-DEV conditions (Non-DEV EtOH-Sal, Non-DEV Water-LiCl) over the five drinking days (repeated measure) after devaluation. The ANOVA revealed a significant main effect of days [F(4,68) = 5.03, p <. 01] due to the gradual increase on responding over days. However, the ANOVA indicated there was no significant difference between the two Non-DEV groups or interactions with days [F(1,68) = 0.31, p = NS]. Interactions between this factor and Group (CIE, CTL), as well as the three-way interaction, were also not significant.
Extinction responding in mice following repeated cycles of CIE was not altered by devaluation. Following the 5th cycle of CIE or air exposure, mice were subjected to ethanol devaluation or sham devaluation followed by daily extinction sessions. For ease of visual comparison, baseline responding for ethanol prior to the 5th cycle of CIE is presented. The values are the mean ± SEM of lever responses for self-administration or extinction over the 30-min test sessions on each day. The * indicates significant increase of CIE versus CTL responding during pre-devaluation extinction test day (t test, p < 0.05). For the Non-DEV group, half of the mice received EtOH (i.g.) followed by a saline injection (i.p.) while the other half received water (i.g.) followed by a LiCl injection (i.p.). A separate ANOVA was conducted using Group (CIE, CTL), Non-DEV conditions (Non-DEV EtOH-Sal, Non-DEV Water-LiCl) over the five extinction testing days (repeated measure) after devaluation. The ANOVA indicated a significant main effect of days [F(4,60) = 10.94, p < .01] due to the gradual decrease on responding over days. However, there was no significant difference between the two Non-DEV groups or interactions with days [F(1,60) = 0.15, p = NS]. Interactions between this factor and Group (CIE, CTL), as well as the three-way interaction, were also not significant.
Acknowledgments
This work was funded by NIAAA grants P50 AA010761, U01 AA014095, U01 AA020929, R01 AA010983, U01 AA019967, and the Department of Veterans Affairs Medical Research. The authors thank Laura Ralston for her excellent technical assistance in conducting this work and Jacqueline Barker for helpful comments on the manuscript.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Becker HC. Animal models of excessive alcohol consumption in rodents. Current Topics in Behavioral Neurosciences. 2013;13:355–377. doi: 10.1007/7854_2012_203. [DOI] [PubMed] [Google Scholar]
- Becker HC, Hale RL. Repeated episodes of ethanol withdrawal potentiate the severity of subsequent withdrawal seizures: an animal model of alcohol withdrawal “kindling”. Alcoholism: Clinical & Experimental Research. 1993;17:94–98. doi: 10.1111/j.1530-0277.1993.tb00731.x. [DOI] [PubMed] [Google Scholar]
- Becker HC, Lopez MF. Increased ethanol drinking after repeated chronic ethanol exposure and withdrawal experience in C57BL/6 mice. Alcoholism: Clinical and Experimental Research. 2004;28:1829–1838. doi: 10.1097/01.alc.0000149977.95306.3a. [DOI] [PubMed] [Google Scholar]
- Corbit LH, Nie H, Janak PH. Habitual alcohol seeking: time course and the contribution of subregions of the dorsal striatum. Biological Psychiatry. 2012;72:389–395. doi: 10.1016/j.biopsych.2012.02.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finn DA, Snelling C, Fretwell AM, Tanchuck MA, Underwood L, Cole M, et al. Increased drinking during withdrawal from intermittent ethanol exposure is blocked by the CRF receptor antagonist D-Phe-CRF(12–41) Alcoholism: Clinical and Experimental Research. 2007;31:939–949. doi: 10.1111/j.1530-0277.2007.00379.x. [DOI] [PubMed] [Google Scholar]
- Griffin WC, 3rd, Lopez MF, Becker HC. Intensity and duration of chronic ethanol exposure is critical for subsequent escalation of voluntary ethanol drinking in mice. Alcoholism: Clinical and Experimental Research. 2009;33:1893–1900. doi: 10.1111/j.1530-0277.2009.01027.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffin WC, 3rd, Lopez MF, Yanke AB, Middaugh LD, Becker HC. Repeated cycles of chronic intermittent ethanol exposure in mice increases voluntary ethanol drinking and ethanol concentrations in the nucleus accumbens. Psychopharmacology. 2009;201:569–580. doi: 10.1007/s00213-008-1324-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heilig M, Egli M, Crabbe JC, Becker HC. Acute withdrawal, protracted abstinence and negative affect in alcoholism: are they linked? Addiction Biology. 2010;15:169–184. doi: 10.1111/j.1369-1600.2009.00194.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hopf FW, Chang SJ, Sparta DR, Bowers MS, Bonci A. Motivation for alcohol becomes resistant to quinine adulteration after 3 to 4 months of intermittent alcohol self-administration. Alcoholism: Clinical and Experimental Research. 2010;34:1565–1573. doi: 10.1111/j.1530-0277.2010.01241.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koob GF. Alcoholism: allostasis and beyond. Alcoholism: Clinical and Experimental Research. 2003;27:232–243. doi: 10.1097/01.ALC.0000057122.36127.C2. [DOI] [PubMed] [Google Scholar]
- Lesscher HM, van Kerkhof LW, Vanderschuren LJ. Inflexible and indifferent alcohol drinking in male mice. Alcoholism: Clinical and Experimental Research. 2010;34:1219–1225. doi: 10.1111/j.1530-0277.2010.01199.x. [DOI] [PubMed] [Google Scholar]
- Lopez MF, Becker HC. Effect of pattern and number of chronic ethanol exposures on subsequent voluntary ethanol intake in C57BL/6J mice. Psychopharmacology. 2005;181:688–696. doi: 10.1007/s00213-005-0026-3. [DOI] [PubMed] [Google Scholar]
- Lopez MF, Becker HC. Increased Reinforcing Efficacy and Reinstatement of Ethanol-Seeking Behavior Following Repeated Cycles of Chronic Intermittent Ethanol Exposure in C57BL/6J Mice. Addiction Biology. under review. [Google Scholar]
- Lopez MF, Griffin WC, 3rd, Melendez RI, Becker HC. Repeated cycles of chronic intermittent ethanol exposure leads to the development of tolerance to aversive effects of ethanol in C57BL/6J mice. Alcoholism: Clinical and Experimental Research. 2012;36:1180–1187. doi: 10.1111/j.1530-0277.2011.01717.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lubow RE, Moore AU. Latent inhibition: the effect of nonreinforced pre-exposure to the conditional stimulus. Journal of Comparative and Physiological Psychology. 1959;52:415–419. doi: 10.1037/h0046700. [DOI] [PubMed] [Google Scholar]
- Meisch RA. Animal studies of alcohol intake. The British Journal of Psychiatry: The Journal of Mental Science. 1982;141:113–120. doi: 10.1192/bjp.141.2.113. [DOI] [PubMed] [Google Scholar]
- Meisch RA. Alcohol self-administration by experimental animals. In: Smart RG, Cappell HD, Glaser FB, Israel Y, Kalant H, Popham RE, et al., editors. Research Advances in Alcohol and Drug Problems. Plenum Press; New York: 1984. pp. 23–45. [Google Scholar]
- Samson HH. Initiation of ethanol reinforcement using a sucrose-substitution procedure in food- and water-sated rats. Alcoholism: Clinical and Experimental Research. 1986;10:436–442. doi: 10.1111/j.1530-0277.1986.tb05120.x. [DOI] [PubMed] [Google Scholar]
- Samson HH, Cunningham CL, Czachowski CL, Chappell A, Legg B, Shannon E. Devaluation of ethanol reinforcement. Alcohol. 2004;32:203–212. doi: 10.1016/j.alcohol.2004.02.002. [DOI] [PubMed] [Google Scholar]
- Tiffany ST. A cognitive model of drug urges and drug-use behavior: role of automatic and nonautomatic processes. Psychological Review. 1990;97:147–168. doi: 10.1037/0033-295x.97.2.147. [DOI] [PubMed] [Google Scholar]
- Vendruscolo LF, Barbier E, Schlosburg JE, Misra KK, Whitfield TW, Jr., Logrip ML, et al. Corticosteroid-dependent plasticity mediates compulsive alcohol drinking in rats. The Journal of Neuroscience. 2012;32:7563–7571. doi: 10.1523/JNEUROSCI.0069-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin HH, Knowlton BJ. The role of the basal ganglia in habit formation. Nature Reviews. Neuroscience. 2006;7:464–476. doi: 10.1038/nrn1919. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Mice with a short history of operant ethanol self-administration exhibited responding that was sensitive to ethanol devaluation. After obtaining stable baseline drinking, mice were subjected to the ethanol devaluation or sham procedure followed by resumption of ethanol self-administration. Baseline responding (mean ± SEM) for the 30-min self-administration sessions is shown for the day immediately prior to initiation of devaluation. Mice subjected to devaluation (DEV) exhibited a significant reduction in responding for ethanol compared to mice that underwent sham devaluation (Non-DEV). The * indicates significant difference between DEV and Non-DEV groups combined (ANOVA with Newman-Keuls post hoc test, p < 0.05). For the Non-DEV group, half of the mice received EtOH (i.g.) followed by a saline injection (i.p.) while the other half received water (i.g.) followed by a LiCl injection (i.p.). A separate ANOVA conducted to evaluate these groups (Non-DEV EtOH-Sal, Non-DEV Water-LiCl) over the 5 drinking days (repeated measure) after devaluation indicated a significant main effect of days [F(4,32) = 5.98, p < .01] due to the gradual increase on responding over days. However, the ANOVA failed to indicate any significant difference between the two Non-DEV groups or interactions with days [F(1,32) = 0.04, and F(4,32) = 0.31, respectively; both p = NS].
Devaluation of ethanol reward in mice with a short history of self-administration does not alter extinction responding. Following establishment of baseline drinking and a single day of extinction training, mice were subjected to ethanol or sham devaluation after which extinction training was resumed. Shown are the mean ± SEM of baseline responding for ethanol and extinction of ethanol self-administration. Half of the mice in the Non-DEV group received EtOH (i.g.) followed by a saline injection (i.p.) while the other half received water (i.g.) followed by a LiCl injection (i.p.). A separate ANOVA conducted to evaluate these groups (Non-DEV EtOH-Sal, Non-DEV Water-LiCl) over the 5 extinction testing days (repeated measure) after devaluation indicated a significant main effect of days [F(4,28) = 6.16, p <. 01] due to the gradual increase on responding over days. However, the ANOVA indicated there was no significant difference between the 2 Non-DEV groups or interactions with days [F(1,28) = 2.28, and F(4,28) = 1.15, respectively; both p = NS].
Repeated cycles of CIE promote insensitivity to devaluation of responding for ethanol. Prior to devaluation, mice subjected to 5 cycles of CIE exhibited enhanced responding for ethanol compared to the sham air-exposed controls. After ethanol or sham devaluation, responding in the air-exposed control mice was sensitive to devaluation. In contrast, CIE-exposed mice exhibited responding that was insensitive to devaluation. The * indicates significant increase of CIE versus CTL (t test, p < 0.05). The # indicates significant difference from respective non-devalued combined controls (ANOVA with Newman Keuls post hoc test, p < 0.05). Half of the mice in the Non-DEV group received EtOH (i.g.) followed by a saline injection (i.p.) while the other half received water (i.g.) followed by a LiCl injection (i.p.). A separate ANOVA was conducted using Group (CIE, CTL), Non-DEV conditions (Non-DEV EtOH-Sal, Non-DEV Water-LiCl) over the five drinking days (repeated measure) after devaluation. The ANOVA revealed a significant main effect of days [F(4,68) = 5.03, p <. 01] due to the gradual increase on responding over days. However, the ANOVA indicated there was no significant difference between the two Non-DEV groups or interactions with days [F(1,68) = 0.31, p = NS]. Interactions between this factor and Group (CIE, CTL), as well as the three-way interaction, were also not significant.
Extinction responding in mice following repeated cycles of CIE was not altered by devaluation. Following the 5th cycle of CIE or air exposure, mice were subjected to ethanol devaluation or sham devaluation followed by daily extinction sessions. For ease of visual comparison, baseline responding for ethanol prior to the 5th cycle of CIE is presented. The values are the mean ± SEM of lever responses for self-administration or extinction over the 30-min test sessions on each day. The * indicates significant increase of CIE versus CTL responding during pre-devaluation extinction test day (t test, p < 0.05). For the Non-DEV group, half of the mice received EtOH (i.g.) followed by a saline injection (i.p.) while the other half received water (i.g.) followed by a LiCl injection (i.p.). A separate ANOVA was conducted using Group (CIE, CTL), Non-DEV conditions (Non-DEV EtOH-Sal, Non-DEV Water-LiCl) over the five extinction testing days (repeated measure) after devaluation. The ANOVA indicated a significant main effect of days [F(4,60) = 10.94, p < .01] due to the gradual decrease on responding over days. However, there was no significant difference between the two Non-DEV groups or interactions with days [F(1,60) = 0.15, p = NS]. Interactions between this factor and Group (CIE, CTL), as well as the three-way interaction, were also not significant.