

Abstract
To illuminate the evolution and mechanisms of actinobacterial complexity, we evaluate the distribution and origins of known Streptomyces developmental genes and the developmental significance of actinobacteria-specific genes. As an aid, we developed the Actinoblast database of reciprocal blastp best hits between the Streptomyces coelicolor genome and more than 100 other actinobacterial genomes (http://streptomyces.org.uk/actinoblast/). We suggest that the emergence of morphological complexity was underpinned by special features of early actinobacteria, such as polar growth and the coupled participation of regulatory Wbl proteins and the redox-protecting thiol mycothiol in transducing a transient nitric oxide signal generated during physiologically stressful growth transitions. It seems that some cell growth and division proteins of early actinobacteria have acquired greater importance for sporulation of complex actinobacteria than for mycelial growth, in which septa are infrequent and not associated with complete cell separation. The acquisition of extracellular proteins with structural roles, a highly regulated extracellular protease cascade, and additional regulatory genes allowed early actinobacterial stationary phase processes to be redeployed in the emergence of aerial hyphae from mycelial mats and in the formation of spore chains. These extracellular proteins may have contributed to speciation. Simpler members of morphologically diverse clades have lost some developmental genes.
Keywords: mycelial growth, polar growth, cell division, sporulation, nitric oxide, mycothiol
Introduction
Bacteria in the ancient phylum Actinobacteria have extraordinary diversity of function and form. They include pathogens of humans and other mammals (the agents of tuberculosis, leprosy, mycetomas, diphtheria, Whipple's disease, and skin, oral and vaginal infections of humans) and plants (potato scab, ratoon stunting disease of sugarcane); major agents of symbiotic nitrogen fixation (Frankia); industrially important producers of amino acids (Corynebacterium glutamicum); genera such as Streptomyces, Micromonospora, Saccharopolyspora and Actinoplanes that are the richest natural source of antibiotics and other secondary metabolites; probiotic bifidobacteria; and agents of bioremediation, notably rhodococci (Ventura et al., 2007). There is also growing interest in their frequent occurrence as plant endophytes and arthropod exosymbionts (Seipke et al., 2012).
Actinobacteria are Gram-positive bacteria that typically have a high G + C content in their DNA. They range from simple cocci to the various complex mycelial forms found in some of the Actinomycetales order (Fig. 1). This morphological diversity is spectacularly illustrated in the ‘Atlas of Actinomycetes’ (Miyadoh, 1997). Mycelial organisms present particular problems for growth and development: their hyphae are intrinsically nonsymmetrical; special mechanisms must be needed to permit and control branching; and they must have some phase of fragmentation that permits dispersal. Often, the fragmentation of actinomycete hyphae leads to the formation of dessication-resistant spores, of a general type distinct from the endospores formed inside ‘mother cells’ of Bacillus spp. and other firmicute bacteria: they are formed directly by cell division from multigenomic hyphal compartments, followed by changes in the cell wall to permit rounding and thickening of the spore wall and the acquisition of resistance properties. These ‘exospores’ appear in or on a considerable variety of specialised morphological structures, including short hyphal side branches, large sporangia and specialised aerial hyphae that turn into long spore chains. In some genera, although not in Streptomyces, spores may be motile.
Fig. 1.

Morphological diversity of actinobacteria. (a) Colonies of Streptomyces coelicolor A3(2) wild type (upper) and bldA mutant (lower). (b) Phase contrast image of sporulating mycelium of S. coelicolor. (c) Micrococcus luteus. (d) Actinosynemma mirum. (e) Microbispora rosea. Images [c–e] are scanning electron micrographs taken from Miyadoh (1997), with permission.
Streptomycetes, the central subject of this article, are the most extensively studied mycelial actinobacteria. They are sporulating organisms whose considerable morphological complexity is interlinked with an extraordinary ability to make diverse secondary metabolites (Chater, 2011; Liu et al., 2013a). Two or three days after a spore germinates on agar media, the biomass-accumulating vegetative or substrate mycelium of the colony becomes covered with a fuzzy white aerial mycelium. The individual aerial hyphae grow to give rise to long unbranched tip cells often containing more than 50 copies of the genome. The tip cells are then divided into multiple prespore compartments by sporulation septation, during which synchronously assembled and regularly spaced FtsZ rings lead septal ingrowth. During sporulation septation, the uncondensed mass of chromosomes partitions into nucleoids, so that each prespore compartment contains a single copy of the genome. The change of cylindrical prespore compartments into ovoid spores involves remodelling and thickening of the cell wall, while inside the developing spore further changes contribute to the onset of dormancy, including chromosome condensation.
Three model species have provided nearly all the available experimental information about the molecular basis of the morphological development of streptomycetes. The most widely studied of these is the genetically amenable S. coelicolor A3(2) (Hopwood, 2007), while S. griseus (one of the first streptomycetes to be used as the source of a major antibiotic, streptomycin) has been particularly intensively studied for its production of, and responsiveness to, a hormone-like developmental signalling molecule, A-factor (Horinouchi, 2002, 2007). The third model species, S. venezuelae, an early industrial producer of chloramphenicol, has recently been taken up as a developmental model, because it sporulates rapidly, synchronously and comprehensively in submerged culture, in contrast to most streptomycetes, which sporulate gradually and nonsynchronously and do not form spores in submerged culture (Flärdh & Buttner, 2009; Bibb et al., 2012). This makes S. venezuelae especially suitable for sensitive biochemical, cytological and molecular studies of consecutive developmental states. The genomes of all three species have been sequenced (Bentley et al., 2002; Ohnishi et al., 2008; FR845719: for annotated presentation, see http://strepdb.streptomyces.org.uk), along with those of numerous other members of the genus.
A previous comparative genomic survey of actinobacteria (Ventura et al., 2007) was based on 21 sequences, encompassing 10 genera, and with many gaps in its phylogenetic coverage. When we began the analysis leading to this article in April 2011, about 100 further actinobacterial genomes had been sequenced and annotated to a level that made productive comparative analysis possible. The number had increased to 157 complete sequences and 474 in progress, in a recent and comprehensive review on the genome-based phylogeny of actinobacteria (Gao & Gupta, 2012). That review extended earlier work in which 28 ‘signature proteins’ peculiar to, and near-universal among, actinobacteria were identified, along with a further 48 peculiar to, and near-universal among, actinomycetes (Gao et al., 2006). These proteins form an important background to this review, and we summarise them in Table 1, using Streptomyces coelicolor gene designations (SCO numbers) as the key instead of those originally used (mainly from Mycobacterium leprae).
Table 1.
Conserved actinobacterial signature proteins/genes identified by Gao et al. (2006) and Gao & Gupta (2012)†. (A) The 26 most frequent actinobacterial signature proteins include at least six with likely developmental roles (asterisks). (B) Seven actinomycete signature proteins referred to in the text‡
| SCO number | ML number | Comments such as gene or protein name, function, conserved linkage, references, etc. |
|---|---|---|
| (A) | ||
| 5199 | 0642 | Often next to conserved gene for ‘epimerase/dehydratase’. Similar to SCO3407 (25% identity over 336 aa overlap), which is also very widespread and actinospecific, but is not listed in Gao et al. (2006). SCO3407 is neighboured by SCO3408 (= ML00211, actinospecific, widely conserved, predicted D-ala, D-ala carboxypeptidase, PBP4 class, similar to dacB of E. coli) and by a cluster conserved even in B. subtilis (SCO3406, possible MesJ-like cell division-associated ATPase; SCO3405, probable hypoxanthine phosphoribosyl transferase; SCO3405, FtsH2, ATP-dependent protease) |
| 1997* | 1009 | Closely similar to ParJ. Function unknown, but structure established (Gao et al., 2009). May perhaps interact with ParA or the ubiquitous ParA2 (= SCO1772). Absent from nonactinomycete actinobacteria and from two actinomycetes, Trophyrema whipplei and Saccharopolyspora erythraea. |
| 5869 | 1029 | DUF3710 domain; probably cotranscribed with SCO5868 (3nt in between; dut, probable deoxyuridine 5′-triphosphate nucleotidohydrolase) and SCO5867 (phenylacetic acid thioesterase, Paa1). Linkage with dut conserved throughout actinomycetes |
| 1662* | 1306 | parJ. ParJ interacts with ParA (Ditkowski et al., 2010) |
| 3034* | 0760 | whiB, developmental regulatory gene (Fowler-Goldsworthy et al., 2011) |
| 5240* | 0804 | wblE, encodes WhiB-like protein of uncertain role, possibly essential in streptomycetes but not in C. glutamicum (Kim et al., 2005; Fowler-Goldsworthy et al., 2011) |
| 2196 | 0857 | 234 aa, probable integral membrane protein |
| 2169 | 0869 | 251 aa, DUF3034, probable integral membrane protein |
| 2947 | 1016 | 97 aa, DUF3039 |
| 5864 | 1026 | 98 aa; note conserved linkage of SCO5864 and 5869, and ML1026 and 1029 |
| 1381 | 2073 | 228 aa; present in all actinobacteria except Acidimicrobium ferrooxidans and Coriobacteriales. Removed by Gao & Gupta (2012) |
| 5855 | 2137 | 252 aa, DUF3071 |
| 4088 | 2204 | 84 aa, DUF3073 |
| 3854* | 0013 | crgA (whiP in S. avermitilis), 84-aa membrane protein, septation inhibitor, absent from nonactinomycete actinobacteria (Del Sol et al., 2003, 2006; Plocinski et al., 2011, 2012) |
| 3872 | 0007 | 185 aa, DUF3566, invariably very close to oriC |
| 1938 | 0580 | opcA; assembly of glucose-6-phosphate dehydrogenase (also in cyanobacteria: Hagen & Meeks, 2001; and next to the zwf2 gene); a less widely occurring paralogue, SCO6660, is downstream of zwf1 in S. coelicolor |
| 2078 | 0921 | 94 aa, possible transmembrane protein, invariably next to divIVA |
| 1421 | 1439 | rpbA, RNA polymerase-binding protein (Tabib-Salazar et al., 2013; Bortoluzzi et al., 2013; note that ML1439 was listed twice by Gao et al. (2006)) |
| 5601 | 1610 | 102aa, DUF2469, conserved linkage with SCO5602 |
| 4084 | 2207 | 437 aa. Note conserved linkage of SCO4084 and 4088, and ML2207 and 2204 |
| 3095* | 0256 | divIC; part of cell division apparatus, interacts with FtsL (Bennett et al., 2007; sequence divergence of DivIC orthologues in other bacteria took them beyond the threshold adopted by Gao et al., 2006; but they were removed by Gao & Gupta, 2012) |
| 3011 | 0775 | lpqB/Putative lipoprotein |
| 3031 | 0761 | 117 aa, DUF1025. Note conserved linkage of SCO3031 and another signature gene, SCO3034 (whiB) |
| 5169 | 0814 | 94 aa, DUF3107, possible ATP-binding protein |
| 2370 | 1649 | 159 aa, DUF3052, invariably next to gene for possible thiol-specific antioxidant protein |
| 4330 | 2142 | 308 aa, DUF3027 |
| (B) | ||
| 3375 | 0234 | lsr2/HNS-like DNA-bridging protein, iron-regulated in M. tuberculosis (Gordon et al., 2010) |
| 2097 | 0904 | 135 aa, DUF3040, part of spore wall-synthesising complex (Kleinschnitz et al., 2011) |
| 4179 | 2200 | 191 aa, cd07821, likely nitrobindin. NO or fatty acid-binding protein domain, structure known for M. tuberculosis (Shepard et al., 2007), conserved synteny with adjacent fur homologue |
| 1480 | 0540 | 107 aa, nucleoid-binding protein sIHF (Yang et al., 2012; Swiercz et al., 2013) |
| 1664 | 1300 | 265 aa, generally very close to mshC gene for mycothiol biosynthesis |
| 3097 | 2030 | rpfC/RPF, secreted protein, peptidoglycan binding, several paralogues |
| 4205 | 2442 | 168 aa, DUF2596, downstream of and overlapping mshA |
The gene identifiers listed by Gao et al. (2006) were for the Mycobacterium leprae genome. Here, we have listed S. coelicolor orthologues as defined by reciprocal best-hit BLASTP analysis. The function descriptions are based on the cited papers where given, but where no reference is given, the commentary is derived from synteny and conserved domain analysis carried out for this review, using StrepDB (http://strepdb.streptomyces.org.uk).
The remaining 39 actinomycete signature genes identified by Gao et al. (2006) were as follows (M. leprae, L. xyli or T. fusca designations given in brackets after SCO equivalent): SCO numbers: 0908 (Tfu_0365), 1372 (Lxx16410), 1383 (ML2075), 1437 (ML0561), 1653 (ML1312), 1665 (ML1299), 1929 (ML0589), 2105 (ML0898), 2153 (ML2446), 2154 (ML0876), 2197 (Lxx10090), 2391 (ML1781), 2460 (Tfu_1340), 2557 (Lxx08190), 2643 (ML1485), 2893 (ML0169), 2915 (ML1166), 2916 (ML1165), 3016 (Tfu_2498), 3030 (ML0762), 3576 (Lxx03620), 3647 (ML0284), 3822 (ML0115), 3902 (ML2687), 4043 (Tfu_0030), 4287 (ML1927), 4579 (ML2064), 4590 (Tfu_1240), 5145 (ML1067), 5167 (Tfu_0515), 5173 (ML0816), 5414 (ML1176), 5493 (ML1706), 5697 (Tfu_0751), 5766 (ML0986), 5866 (ML1027), 6030 (ML1041). One (ML2428A) was similar to SCO3327, but did not give a reciprocal blastp best hit, and another (ML0899) was absent from S. coelicolor, but present in S. avermitilis (SAV1313) and many other streptomycetes.
Our aim in this article is to combine comparative genomics, knowledge about Streptomyces development and growing information about gene function gleaned from other actinobacteria, particularly from the intensive focus of many researchers on the globally important pathogen Mycobacterium tuberculosis, to address several questions: What are the evolutionary origins of genes important for Streptomyces sporulation? Are the mechanisms leading to sporulation widely homologous in phylogenetically diverse actinobacteria, or did they evolve independently? Does the developmental process contribute to speciation? Are today's simple actinobacterial species primitive, or are they degenerate descendants of morphologically much more complex ancestors? What gave the ancestral ur-actinobacterium the potential for such morphological complexity in its modern descendants? And can studies of the development of complex actinomycetes assist our understanding of the cell biology of their simpler cousins?
Our analysis was aided by tabulating reciprocal blastp best hits of the translated products of each S. coelicolor gene with those of more than 100 actinobacterial genomes (http://streptomyces.org.uk/actinoblast/). We further analysed these tabulations using different approaches to identify proteins widespread among actinobacteria, but absent from other bacteria (as represented by E. coli and B. subtilis), in an extension of the work of Gao et al. (2006) and Gao & Gupta (2012). These approaches, which we do not describe in detail, included listing proteins in order of their frequency of representation in all actinobacteria analysed and analysing proteins present in both S. coelicolor and Micrococcus luteus, two morphologically and phylogenetically distinct organisms. Proteins of interest were further investigated using the NCBI Conserved Domain Database, which in several cases proved illuminating in relation to possible function. Throughout the article, the SCO identifiers used in the S coelicolor genome are used to designate genes and their protein products interchangeably. The rich genome sequence database used in this survey has caused us to modify some of the conclusions of an earlier exploration of this theme (Chater & Chandra, 2006) and to put forward some new ideas.
Taxonomy and phylogeny of actinobacteria, in relation to developmental complexity
The taxonomy of actinobacteria has been through several phases. Initially, the phylum consisted of mycelial bacteria termed actinomycetes. Genera were named in accordance with their different modes of sporulation (e.g. Micromonospora, Streptosporangium, etc.). Subsequently, the use of chemotaxonomy and numerical taxonomy led to the inclusion of some nonmycelial organisms in the phylum. Eventually, the sequencing of 16S ribosomal RNA began to provide a clearer phylogenetic basis for the taxonomy, and further genera of simple bacteria, such as Bifidobacterium, were shown to be related to the Actinomycetales, leading to the recognition of a more inclusive phylum, Actinobacteria. Using such a 16S RNA-based scheme, Zhi et al. (2009) divided Actinobacteria into five orders, one of which, the Actinomycetales, contained the great majority of families, the other four orders being made up of very few families: Rubrobacterales, including Rubrobacter and Conexibacter as genera with sequenced representatives; Acidimicrobiales, comprising only Acidimicrobium; Bifidobacteriales, including the genera Bifidobacterium and Gardnerella; and Coriobacteriales, including sequenced representatives in the genera Coriobacterium, Atopobium, Cryptobacterium, Eggerthella, Olsenella and Slackia. To maximise the ease of relating this article to the existing literature, we are using this taxonomic scheme. However, in the last few years, genome-level information has been employed in various ways to increase the resolution of actinobacterial phylogeny. Alam et al. (2010) combined several approaches, including gene order, to arrive at a well-resolved phylogeny, the main limitation of which was the lack of genome sequences representing the deepest branches. A key element in their analysis was the use of catenated sequences of 155 conserved proteins. A catenated set of 21 conserved protein sequences was used by Penn & Jensen (2012) to generate a tree from 186 actinobacterial genome sequences, while Gao & Gupta (2012) used 35 catenated conserved proteins to generate a very well-resolved tree from 98 actinobacterial genomes chosen to give comprehensive coverage of the phylum. Gao & Gupta (2012) went on to show that the distribution of taxon-specific signature indels (small insertions or deletions) and signature proteins fully supported the branch order of their tree, which we have therefore taken as the scaffolding for the rest of this article, but without adopting their revised taxonomic scheme (because it can be confusing for nonspecialists in relation to the pre-existing literature). It was pointed out by Gao & Gupta (2012) that the Coriobacteriales, previously included in Actinobacteria, lacked all the actinobacterial signature proteins and indels and should therefore be excluded from the phylum (a suggestion re-examined by Gupta et al., 2013). Likewise, we found that all the actinobacteria-specific genes that we discuss in this article were absent from Coriobacteriales. The term Actinobacteria is therefore taken to exclude Coriobacteriales throughout this article, although they are included in the reciprocal blastp best-hit analyses shown in some of the figures.
Because of our underlying emphasis on Streptomyces, we needed to re-present this phylogeny from the perspective of Streptomyces. To do this, we reorganised the tree of Gao & Gupta (2012) to show clearly the nodes at which various taxa shared a last common ancestor with Streptomyces, and aligned it with an estimated timescale derived from Battistuzzi et al. (2004) (Fig. 2). A complication of this scheme is the division of one of the Actinomycetales suborders, Micrococcineae, into three suborders, one of which has a more ancient origin than Bifidobacteriales (Gao & Gupta, 2012) and is indicated in Fig. 2 by node 3A. For the purposes of this article, we consider that node 4 of Fig. 2 represents the origin of Actinomycetales (i.e. actinomycetes).
Fig. 2.

The evolutionary path leading to Streptomyces. The diagram was derived from the phylogenetic tree in Fig. 2 of Gao & Gupta (2012), and the boxes correspond in colour to those used in Fig 3. Nodes 1 to 9 are reference points for the main text. Arrow lengths are not proportional to phylogenetic difference. Micrococcales genera showing a simple mycelium are indicated by asterisks, and sporulating mycelial genera are given in bold type. The approximate evolutionary timescale is based on Battistuzzi et al. (2004).
It can be seen in Fig. 2 that the actinobacteria originating from nodes 1 to 3B on the path to Streptomyces all show variations on a simple rod/coccus morphology and do not sporulate. These organisms include most of the obligately anaerobic genera, consistent with the earliest actinobacteria having preceded the major oxygenation of the atmosphere, around 2.3 Gya (Battistuzzi et al., 2004). The earliest branch of the order Actinomycetales (node 4) also leads almost exclusively to rod/coccus organisms (suborders Micrococcineae and Kineosporineae), with (rudimentary) mycelial growth being found only in Rothia and Intrasporangium. Extensive mycelium formation and sporulation occur in organisms originating at or after node 5, although some genera such as Corynebacterium and Mycobacterium arising from a later node do not show obvious developmental complexity (readers interested in mycobacterial dormancy or the controversial suggestion that mycobacteria can sporulate are referred to Gengenbacher & Kaufmann, 2012; Lamont et al., 2012). The simplest explanation for this discontinuity is that extensive and obligatory mycelial growth arose once and was closely associated with the evolution of sporulation, but such developmental complexity was lost from some lines later in evolution. We show later that loss of complexity is associated with the loss of several developmental regulatory genes.
The Streptomyces sporulation regulatory cascade is built on ancient roots
Focused genetic studies of model streptomycetes have revealed several tens of key developmental genes (Flärdh & Buttner, 2009; Chater, 2011; McCormick & Flärdh, 2012). Mutations in some of these genes result in the loss of aerial mycelium formation, at least under most culture conditions (Merrick, 1976; Champness, 1988). Because of the bald appearance of the colonies, such genes are mostly designated bld. Another major phenotypic class of developmental mutants – those that form aerial hyphae but do not sporulate efficiently – identified the whi genes, so-called because the mutants fail to accumulate spore pigment in their aerial mycelium, which remains white on prolonged incubation (Hopwood et al., 1970; Chater, 1972). Here, we evaluate the phylogenetic distribution of many of these genes and interpret the results in terms of the evolution and mechanisms of Streptomyces development (Fig. 3: the legend to Fig. 3 includes information about the methods used to generate the data and how to access the full tables). Unless stated otherwise, orthologues of these genes are absent from B. subtilis and E. coli, so they may well be confined to actinobacteria. It appears from this that the key developmental regulatory roots of Streptomyces sporulation described in this section lie in some of the Whi proteins, while actions of the Bld proteins (also mostly regulatory) have come to be overlaid on the initiation of the whi gene cascade. These genes are discussed in the inferred order of their appearance during the c. 2.7 G years since the emergence of the first actinobacteria (Battistuzzi et al., 2004). We also identify some potentially interesting, but sometimes little-studied, genes whose patterns of occurrence across the actinobacteria are congruent with those of certain well-known developmental genes, and speculate on the significance of this congruence.
Fig. 3.
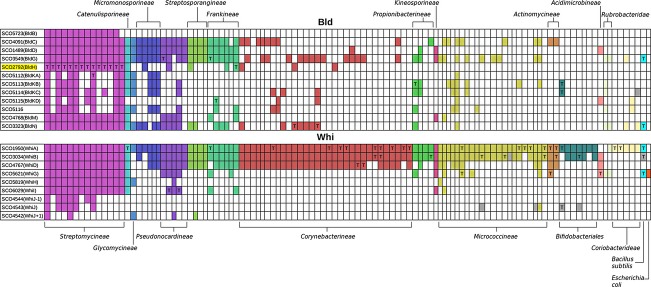
Distribution of probable orthologues of Bld and Whi proteins of Streptomyces coelicolor encoded in more than 100 actinobacterial genomes, as detected by reciprocal blastp best hits. Each column represents one genome, and the genomes are grouped and coloured to indicate subgroup relationships (e.g. Corynebacterineae columns, including Mycobacterium, Nocardia and Corynebacterium, were coloured Indian red). Grey boxes indicate reciprocal hits falling below the minimal criteria adopted for orthology. White boxes indicate the absence of a reciprocal hit. The yellow highlighted SCO genes contain a TTA codon, and the presence of TTA codons in apparent orthologues is indicated by a T in the coloured box. A similar display of reciprocal blastp analysis of the entire S. coelicolor genome against the 111 genomes, with links to StrepDB, is available at http://streptomyces.org.uk/actinoblast/. The tables at that site allow clicking onto any coloured box to show the gene identifier together with minimal annotation, as well as information about the length of the overlap and the percentage identity. The sources of genomes are listed in Table 1 of Gao & Gupta (2012). Organisms were as follows (in order across the tabulation). Magenta: Streptomycineae, S. lividans TK24, S. viridochromogenes DSM 40736, S. scabiei 87.22, S. sviceus ATCC 29083, S. avermitilis MA-4680, S. griseoflavus Tu4000, S. venezuelae ATCC 10712, S. griseus subsp. griseus NBRC 13350, S. hygroscopicus ATCC 53653, S. pristinaespiralis ATCC 25486, S. roseosporus NRRL 15998, S. albus G J1074, S. clavuligerus ATCC 27064, Kitasatospora setae KM-6054. Turquoise: Catenulispora acidiphila DSM 44928. Light blue: Stackebrandtia nassauensis DSM 44728. Dark blue: Salinispora, S. tropica CNB-440, S. arenicola CNS-205; Micromonospora, M. sp. L5, M. sp. ATCC39149, M. aurantiaca ATCC 27029. Purple: Saccharomonospora viridis DSM 43017; Saccharopolyspora erythraea NRRL 2338; Amycolatopsis mediterranei U32; Actinosynnema mirum DSM 43827; Thermobispora bispora DSM 43833. Yellow green: Streptosporangium roseum DSM 43021; Thermomonospora curvata DSM 43183; Thermobifida fusca YX; Nocardiopsis dassonvillei subsp. dassonvillei DSM 43111. Blue green: Acidothermus cellulolyticus 11B; Frankia, F. sp. EAN1pec, F. sp. CcI3, F. alni ACN14a; Geodermatophilus obscurus DSM 43160; Nakamurella multipartita DSM 44233. Rust red: Gordonia bronchialis DSM 43247; Nocardia farcinica IFM 10152; Segniliparus rotundus DSM 44985; Tsukamurella paurometabola DSM 20162; Rhodococcus, R. opacus B4, R. jostii RHA1, R. erythropolis PR4, R. equi 103S; Mycobacterium, M. vanbaalenii PYR-1, M. ulcerans Agy99, M. sp. Spyr1, M. sp. MCS, M. sp. KMS, M. sp. JLS, M. smegmatis str. MC2 155, M. marinum M, M. leprae Br4923, M. gilvum PYR-GCK, M. abscessus ATCC 19977, M. avium subsp. paratuberculosis K-10, M. avium 104, M. tuberculosis H37Rv, M. bovis AF2122/97; Corynebacterium, C. urealyticum DSM 7109, C. pseudotuberculosis FRC41, C. kroppenstedtii DSM 44385, C. jeikeium K411, C. glutamicum ATCC 13032 2, C. efficiens YS-314, C. diphtheriae NCTC 13129, C. aurimucosum ATCC 700975. Bright green: Nocardioides sp. JS614; Kribbella flavida DSM 17836; Propionibacterium, P. freudenreichii subsp. shermanii CIRM-BIA1, P. acnes KPA171202. Plum: Kineococcus radiotolerans SRS30216. Olive yellow: Beutenbergia cavernae DSM 12333; Cellulomonas flavigena DSM 20109; Brachybacterium faecium DSM 4810; Kytococcus sedentarius DSM 20547; Intrasporangium calvum DSM 43043; Jonesia denitrificans DSM 20603; Clavibacter michiganensis subsp. michiganensis NCPPB 382; Leifsonia xyli subsp. xyli str. CTCB07; Microbacterium testaceum StLB037; Arthrobacter, A. sp. FB24, A. phenanthrenivorans Sphe3, A. chlorophenolicus A6, A. aurescens TC1, A. arilaitensis Re117; Kocuria rhizophila DC2201; Micrococcus luteus NCTC 2665; Renibacterium salmoninarum ATCC 33209; Rothias, R. mucilaginosa DY-18, R. dentocariosa ATCC 17931; Xylanimonas cellulosilytica DSM 15894; Sanguibacter keddieii DSM 10542; Tropheryma whipplei str. Twist. Brown: Mobiluncus curtisii ATCC 43063; Arcanobacterium haemolyticum DSM 20595. Cyan: Gardnerella vaginalis ATCC 14019; Bifidobacterium, B. longum NCC2705, B. longum DJO10A, B. dentium Bd1, B. bifidum PRL2010, B. animalis subsp. lactis Bl-04, B. adolescentis ATCC 15703. Pink: Acidimicrobium ferrooxidans DSM10331. Pale grey green: Conexibacter woesii DSM14684; Rubrobacter xylanophilus DSM9941. Beige: Atopobium parvulum DSM 20469; Cryptobacterium curtum DSM 15641; Eggerthella lenta DSM 2243; Olsenella uli DSM 7084; Slackia heliotrinireducens DSM 20476.
WhiG, an orthologue of an ancient sigma factor, regulates more recently acquired regulatory genes specific to aerial sporulation
Considering likely orthologues of all the bld and whi genes studied, none is more widespread across the bacterial kingdom than whiG. WhiG protein is a sigma factor critically involved in the decision of aerial hyphae to sporulate, and in its absence, colonies develop long, thin aerial hyphae and entirely fail to sporulate (Chater, 1972). It is orthologous with the extensively studied FliA of E. coli and SigD of B. subtilis, which are involved in regulating genes important for motility and chemotaxis, adhesion and invasion, some aspect(s) of cell wall remodelling and cyclic di-AMP hydrolysis (Helmann, 1991; Claret et al., 2007; Luo & Helmann, 2012). It is possible to envisage connections between these functions and Streptomyces sporulation, as they are mostly associated with the transition from growth as a biofilm to dispersal as planktonic single cells. However, FliA in E. coli and SigD in B. subtilis are both regulated by an antisigma factor, FlgM, that has the extraordinary property of being exported via the flagellar basal body during flagellum assembly. This is clearly not feasible for nonmotile streptomycetes, so it is not surprising that no homologue of this antisigma factor has been found in streptomycetes.
WhiG orthologues are widely but intermittently present in diverse actinobacteria, including some that are morphologically simple (Fig. 3). In most cases, these simpler organisms have been recorded as motile, the exceptions being Acidithermus and Rubrobacter (but Acidithermus does have a set of flagellar genes: Barabote et al., 2009). The only node 6-branch organism possessing WhiG, Nocardioides, is the only mycelial, sporulating organism known in this branch, and it also has a set of flagellar genes (Barabote et al., 2009). Even if motility functions are regulated by WhiG orthologues in these actinobacteria, no FlgM-like protein is encoded in any of their genomes. The whiG-like genes all show some local synteny, part of which is even retained in B. subtilis, so whiG seems to have been lost independently from several actinobacterial lines, rather than having been absent from the last common ancestor and then reacquired later in actinobacterial evolution as we previously suggested (Chater & Chandra, 2006).
RNA polymerase containing WhiG sigma directly activates two regulatory genes involved in slightly later stages in sporulation (whiH, Ryding et al., 1998; whiI, Ainsa et al., 1999). WhiI protein resembles response regulators, many of which are part of two-component systems in which activity of the response regulator is determined by its phosphorylation by a partner sensor kinase. WhiI, however, does not have a known partner kinase, being one of 13 ‘orphan’ response regulators present in S. coelicolor (Hutchings, 2007), and lacks key residues normally required for phosphorylation (Tian et al., 2007). It occurs almost exclusively in developmentally complex WhiG-containing actinomycetes and is absent from WhiG-containing, morphologically simple, motile actinobacteria; but both WhiG and WhiI are absent from many mycelial actinomycetes whose sporulation does not involve the formation of chains of spores on long aerial hyphae (Frankia, Micromonospora, Salinispora, Thermobispora, Nocardiopsis, Thermobifida, Streptosporangium and Thermomonospora). The other WhiG target regulatory gene, whiH, encodes an autoregulating GntR-like protein (Ryding et al., 1998; Persson et al., 2013) confined to streptomycetes and their closest relatives (Catenulispora and Kitasatospora).
In summary, the WhiG-dependent part of the Streptomyces sporulation regulatory cascade (as known until recently, see below) appears to have evolved in a stepwise manner, in which an early role for WhiG may have been to facilitate planktonic dispersal from biofilms (but there is still no analysis of roles for WhiG in motility and chemotaxis of motile simple actinobacteria). This made it potentially appropriate for activating the analogous process of sporulation of mycelial mats. The subsequent acquisition (and WhiG dependence) of WhiI and WhiH may have permitted increased provision of components needed in large amounts for sporulation septation and spore maturation, as in WhiI-dependent upregulation of genes needed for phosphoinositides for membrane synthesis (Tian et al., 2007; Zhang et al., 2012) and an apparently WhiH-stimulated increase in the supply of FtsZ for sporulation septation (Flärdh et al., 1999, 2000).
whiA, part of a syntenous cluster of genes conserved across Gram-positive bacteria
Like WhiG (but no other sporulation regulator of S. coelicolor), WhiA orthologues are not confined to actinobacteria: one is present in most Gram-positive bacteria, including all actinobacteria except Acidimicrobium ferrooxidans. The structure and molecular function of WhiA have only been fruitfully studied recently. One of its two domains is an evolutionary relative of homing endonucleases, but lacks catalytic residues, and the other resembles the C-terminal domain of major sigma factors, which interacts with the -35 region of promoters (Knizewski & Ginalski, 2007; Kaiser et al., 2009). WhiA showed in vitro DNA binding to its own promoter and to a sporulation-activated promoter of the parAB operon (Kaiser & Stoddard, 2011), both of which are also WhiA-dependent in vivo (Jakimowicz et al., 2006). The whiA sporulation-specific promoter could be transcribed in vitro by WhiG-containing RNA polymerase (Kaiser & Stoddard, 2011), in contradiction of an earlier result (Ainsa et al., 2000). WhiA exerted a modest inhibitory effect on this transcription and showed some evidence of direct interaction with WhiG in a pull-down experiment involving the two purified proteins (Kaiser & Stoddard, 2011). These experiments, although not conclusive, provide the first suggestion of direct interplay between the WhiG- and WhiA-dependent parts of the sporulation regulatory cascade, previously thought to be separate (Chater, 1998; Flärdh et al., 1999).
whiA and the upstream three genes form a cluster that is highly conserved in actinobacteria and even in B. subtilis. This putative operon is probably responsible for a low level of whiA (SCO1950) expression during growth (Ainsa et al., 2000). The three upstream genes encode apparently unrelated deduced functions: the UvrC excinuclease (SCO1953); a NTPase that inactivates an sRNA (GlmZ) that regulates glucosamine-6-phosphate (GlcN6P) synthase production in E. coli (NCBI conserved domain PRK05416; SCO1952); and a protein of unknown function (SCO1951) that is related to an enzyme of cytochrome F420 biosynthesis, LPPG:Fo 2-phospho-L-lactate transferase (pfam01933). There is also conspicuous synteny on the other side of whiA in actinobacteria (but this does not extend to B. subtilis): three genes for steps in glycolysis/gluconeogenesis, glyceraldehyde-3-phosphate dehydrogenase, phosphoglycerate kinase and triose phosphate isomerase, are always found next to whiA (or separated from it by one or two genes in some streptomycetes), along with secG, encoding part of the protein secretion system. If the notion of ‘guilt by association’ is applied to whiA, we may guess that it operates in the context of a physiological transition resulting from nutritional limitation, such that assimilated nutrients are redirected via gluconeogenesis to generate glucose-6-phosphate, which may then be converted into N-acetyl glucosamine for cell wall synthesis during aerial growth (perhaps also feeding into mycothiol biosynthesis, see below). This model does not account for all the conserved genetic linkage of whiA, but it is consistent with the apparent inability of aerial hyphae of whiA mutants of streptomycetes to stop growing and switch to sporulation (Chater, 1972).
WhiB and its paralogues: ancient actinobacterial nitric oxide-binding proteins
A phenotype identical to that of whiA mutants results from mutations in whiB (SCO3034), which encodes one of the actinobacterial signature proteins (Table 1A, Fig. 3). Mutation of the whiB orthologue (whmD) of Mycobacterium smegmatis indicated a likely role in cell division that could represent its core activity (Gomez & Bishai, 2000). There are strong two-way transcriptional influences (not necessarily direct) between whiA and whiB (Jakimowicz et al., 2006), but little is known about other possible WhiB targets.
WhiB is the exemplar of a paralogous family of small proteins (Wbl for WhiB-like: Soliveri et al., 1993, 2000; Fowler-Goldsworthy et al., 2011) that all possess an oxygen-sensitive [4Fe, 4S] cluster coordinated by four conserved cysteinyl residues (Jakimowicz et al., 2005a,b; den Hengst & Buttner, 2008; Alam et al., 2009; Saini et al., 2011, 2012). Orthologues of four other Wbl proteins (WblA, WblC, WhiD and WblE) occur in most actinomycetes (Figs 3 and 4), even though WblA and WhiD have developmental roles in S. coelicolor: WblA plays a key part in the transition of aerial hyphal initial branches to a sporulation-directed fate (wblA mutants have thin aerial hyphae often embedded in an extracellular matrix, with only occasional spore chains: Fowler-Goldsworthy et al., 2011); and mutants lacking WhiD have defects at a later stage, having thin-walled spores and uncontrolled sporulation septation (McVittie, 1974; Molle et al., 2000). Limited information is available about the roles of these two proteins in simpler actinobacteria: in Corynebacterium glutamicum, the WblA orthologue WhcA negatively influences the oxidative stress response (Choi et al., 2009); and the WhiD orthologue of Mycobacterium tuberculosis (WhiB3) is required for virulence (Saini et al., 2011). Some nonactinomycete actinobacteria also contain Wbl proteins, notably Bifidobacteriales, which have orthologues of two S. coelicolor Wbls, WhiB and WblK, respectively, termed WhiB2 and WblE in a recent survey (Averina et al., 2012). Using WhiB as a probe in one-way blastp searches of all the translated actinobacterial genomes, paralogues were sometimes very abundant: in an extreme case, Rhodococcus jostii possessed 30 wbl genes, all but five of them being almost specific to this organism or genus. The frequent finding of wbl genes in actinophages (e.g. Dedrick et al., 2013) and plasmids (e.g. SCP1; Fig. 4 and Bentley et al., 2004) makes it plausible that the large family of Wbl paralogues evolved in these elements, which would also serve as agents for their genome-specific lateral acquisition by diverse actinobacteria (Saini et al., 2011).
Fig. 4.
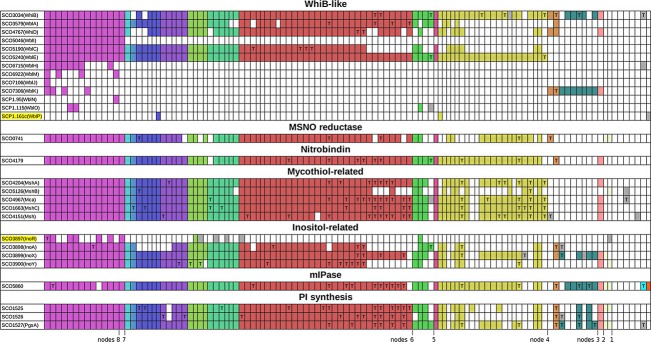
Distribution of WhiB-like (Wbl) proteins compared with proteins related to nitric oxide and mycothiol metabolism. The numbered nodes refer to Fig. 2. See Fig. 3 legend and text for further details.
Soliveri et al. (2000) suggested that WhiB and other Wbl proteins might interact with the major antioxidant thiol mycothiol (MSH), which is widespread among, and apparently confined to, actinobacteria (Fahey, 2012). Genomic searches confirmed that the MSH pathway is present in most Actinomycetales, but it is absent from nonactinomycete actinobacteria apart from Acidimicrobium ferrooxidans, even though wbl genes are present in many of these (Fig. 4). Thus, Wbl proteins can fulfil at least some function(s) in the absence of MSH. Nevertheless, another Wbl protein of M. tuberculosis, WhiB7 (=WblC), which is important in a global response to various antibiotics and other inhibitors and is widespread among actinobacteria, was found to control, directly or indirectly, the concentration of mycothiol (MSH+MSSM; Morris et al., 2005; Burian et al., 2012), and mycothiol-deficient mutants of Mycobacterium smegmatis (Rawat et al., 2002), Rhodococcus jostii (Dosanjh et al., 2008) and Corynebacterium glutamicum (Liu et al., 2013b) showed pleiotropic sensitivity to antibiotics similar to that of whiB7/wblC mutants of M. tuberculosis and Streptomyces lividans (Morris et al., 2005).
Several Wbl proteins have been shown to have specific DNA-binding activity (Rybniker et al., 2010; Smith et al., 2010; Stapleton et al., 2012). This can be enhanced by rapid, high-affinity interaction of the [4Fe, 4S] clusters with NO (Singh et al., 2007; Smith et al., 2010; Crack et al., 2011, 2013; Stapleton et al., 2012). Interestingly, we found two likely NO-related genes with a phylogenetic distribution similar (although not identical) to that of Wbl proteins (Fig. 4). One of these genes, SCO0741, encodes an orthologue of a mycobacterial protein that in vitro very rapidly reduces an NO conjugate (MSNO) of MSH to MSH sulphonamide, which in vivo is processed by M. smegmatis to MSSM (oxidised MSH) and nitrate (Vogt et al., 2003).
The second potentially NO-related protein with a very similar distribution, SCO4179, has clearcut similarity to nitrobindins of plants and animals (Bianchetti et al., 2010; Bianchetti et al., 2011). Nitrobindins are haem-containing proteins that bind NO in the absence of oxygen, and whose major structural features are conserved in the SCO4179 protein and its orthologues in other actinobacteria (Shepard et al., 2007; Bianchetti et al., 2010, 2011). A potential role for nitrobindin might be to transfer the NO groups of Wbl:NO complexes to MSH (Fig. 5). This would provide a means of recycling Wbl proteins to ensure that any burst of Wbl:NO-dependent transcription would be switched off once the Wbl-dependent gene expression cascade had been set in motion (Fig. 5). MSNO might in turn be denitrosylated by MSNO reductase. The MSSM formed in this process would then be reduced to MSH either by mycothiol reductase, which is present in most actinomycetes although apparently not in streptomycetes, or by some other, possibly less specific, thiol reductase. The MSNO reductase gene is immediately upstream of a gene (SCO0740) encoding a protein with homology to hydroxyacylglutathione hydrolases (Rawat & Av-Gay, 2007). This pairing is seen in almost all the actinobacteria that possess the MSNO reductase gene, with the genes usually overlapping by one nucleotide, indicating likely co-transcription and translational coupling. As glutathione is not present in most actinobacteria, an activity on a mycothiol derivative may be the real function of SCO0740 – perhaps in association with MSNO reductase.
Fig. 5.

Hypothetical scheme invoking the involvement of nitric oxide, mycothiol and Wbl proteins in major physiological or developmental decisions. It is supposed that early actinobacteria possessed the functions coloured grey green. They made phosphoinositol-containing phospholipids and used Wbl proteins to respond to nitrosative stress (the pink arrows indicate downstream regulatory events of different Wbl states). The putative nitrobindin may have aided the denitrosylation of Wbl:NO proteins. It is further suggested that the subsequent acquisition of mycothiol biosynthetic genes and MSNO reductase greatly increased the efficiency of NO removal and Wbl regeneration.
A further hint of a Wbl–NO connection has been found: in Corynebacterium glutamicum, the Wbl protein WhcA appears to interact with a protein showing very high similarity to nitronate monooxygenase (Park et al., 2011), an FMN-dependent fungal and bacterial enzyme that generates nitrite from alkyl nitronates (Gadda & Francis, 2010) and is found in nearly all actinobacteria (the S. coelicolor equivalent of this protein is SCO2553).
If MSNO was significant for early actinobacteria that emerged before the evolution of complex eukaryotes that produce NO as a defence and signalling molecule, NO may be an endogenous signal molecule in actinobacteria, which in Streptomyces fulfils roles in development (and in any other general physiological changes influenced by Wbl proteins). How might NO be generated, as the great majority of actinobacteria do not possess an obvious nitric oxide synthase? In plants, nitrate reductase has been implicated as a generator of endogenous NO that brings about the closure of stomata (Desikan et al., 2002), and nitrate and nitrite reductases generate NO in bacteria (Corker & Poole, 2003; Vine et al., 2011). Like the binding of NO by nitrobindin (Bianchetti et al., 2010), these reactions are anoxic. This may explain why the WhiB7 (=WblC)-dependent response of M. tuberculosis to antibiotics was surprisingly stimulated by reducing conditions (added dithiothreitol), but not by oxidative stress induced by the thiol oxidant diamide (Burian et al., 2012). It is interesting to note that in surveys of the thiol-oxidative stress responses mediated by the SigR system, only one of the genes discussed in this section (mshA, determining a step in MSH biosynthesis) was part of the SigR regulon (Paget et al., 2001; Kim et al., 2012). This is consistent with the idea of a partial separation of Wbl–NO–MSH physiology from responses to external oxidative stress (but does not exclude them from having some involvement).
The acquisition of MSH biosynthesis early in actinobacterial evolution seems to have been preceded by the means to generate an immediate precursor for MSH, myoinositol-1-phosphate (mIP), which is also absent from other bacteria. The relevant gene (SCO3899, inoA), is present in at least one nonactinomycete, Arcanobacterium haemolyticum, and is present in many actinomycetes, although surprisingly absent from corynebacteria and many Micrococcales. The source of inositol for MSH biosynthesis in these organisms is not known. When present, inoA is generally adjacent to its regulatory gene (inoR; Zhang et al., 2012). A similar distribution across actinobacteria was found for the biosynthetic genes of other inositol derivatives such as phosphoinositides (a three-gene cluster comprising SCO1527, putatively encoding phosphoinositide synthase, and SCO1525 and SCO1526, likely determinants of the further modification of phosphoinositides: Zhang et al., 2012; Fig. 4). From this, it seems likely that an early actinobacterial organism already possessed the ability to make phosphoinositides from glucose-6-phosphate and that the later acquisition of MSH biosynthesis, close to the time of emergence of the first actinomycetes, was made possible by the availability of the MSH precursor mIP (Fig. 5).
Finally in this section, we draw attention to genes for two further signature proteins listed in Table 1B: SCO1664 and SCO4205 are invariably closely linked to MSH biosynthetic genes (mshC, SCO1663, and mshA, SCO4204), so it is possible that their functions may also be implicated in the network proposed in Fig. 5.
Evolution of the developmental roles of two ancient genes, bldC and bldD
In B. subtilis, sporulation is an extreme response to nutrient limitation usually taken only when all other solutions fail (Narula et al., 2012). Likewise, in streptomycetes, it seems that many (but not all) of the bld gene products, which are nearly all regulatory, feed in information relevant to this drastic decision and ensure that the whi gene cascade operates only under fully appropriate circumstances.
The most ubiquitous bld genes are bldC, encoding an apparently single-domain small protein with a helix-turn-helix of the MerR type (Hunt et al., 2005), and bldD, which encodes a protein distantly related to SinR, a transition state regulator of B. subtilis (Elliot et al., 1998). Orthologues of both are found in most of the morphologically complex, large-genome actinomycetes. Simpler organisms (relatively anciently diverged from the Streptomyces line) seldom have both, and often (as in the case of anaerobic actinobacteria) have neither (Fig. 3). bldD orthologues always show high conservation and local synteny, but bldC orthologues are somewhat less highly conserved and are located among less extensively conserved genes, although some evidence of bldC synteny could often be detected (Chater & Chandra, 2006) (except where the BldC reciprocal blastp best hits were to proteins showing well under 50% identity – such cases may well be laterally acquired paralogues). Importantly, there are convincing orthologues of bldC in Rubrobacteriales and bldD in Acidimicrobium. Thus, both genes were present in very early actinobacteria, but each gene has been lost many times in the later evolution of the phylum. These losses may conceivably have contributed to the evolution of branches such as Micrococcineae (Node 4 of Fig. 2) and Corynebacterineae (a sub-branch from node 6).
The BldD regulon has been subjected to detailed analysis by immunoprecipitation of in vivo BldD–DNA complexes, which showed that BldD directly targets about 147 transcription units in vegetative, liquid-grown S. coelicolor (den Hengst et al., 2010). These include 42 regulatory genes, several of which are developmental (bldA, bldC, bldD, bldH, bldM, bldN, whiB, whiG). These are all repressed by BldD. Based on a consensus sequence derived from these ChIP-chip data, BldD recognition sequences were found upstream of many of the same genes not only in other streptomycetes, but also in other sporulating actinobacteria (den Hengst et al., 2010). Such species included Saccharopolyspora erythraea, an organism in which a constructed bldD mutant had a bald colony phenotype (Chng et al., 2008). Thus, BldD orthologues appear to coordinate development in diverse sporulating actinomycetes, perhaps preventing the expression of genes for morphological differentiation and antibiotic production during vegetative growth and connecting the regulons of other regulators of these processes (den Hengst et al., 2010). BldD orthologues in simpler actinomycetes might well have roles both during growth, to repress functions associated with entry into stationary phase, and in stationary phase, in coordinating the expression of different stationary phase regulatory genes.
Despite the extensive characterisation of BldD and its regulon, it is not understood why, if BldD represses developmental functions, bldD mutants are bald rather than hypersporulating (but see the paragraphs on BldN below); and there is no information about any signals that BldD might respond to (an initial search for possible proteins interacting with BldD was reported to have had negative results: den Hengst et al., 2010). It has been suggested that an interaction of BldD with another sporulation regulatory protein, BldB, could determine the rate of turnover of BldD (McCormick & Flärdh, 2012); but BldB is confined to streptomycetes, so it could not fulfil such a role in other complex actinomycetes such as Sac. erythraea (Fig. 3).
Evolution of the BldD regulon
Some BldD-regulated bld genes of S. coelicolor belong to classes of genes that are widespread and often represented by multiple paralogues in any one genome. For such genes, it can be difficult to be confident that reciprocal blastp hits between genomes are meaningful, particularly when the extent of amino acid identity falls well below levels that are typically seen for conserved housekeeping genes. For example, the general kind of anti-anti-sigma factor to which the BldG protein belongs is almost universally found among both Gram-positive firmicutes and actinobacteria; so the presence in some actinobacteria of BldG reciprocal best hits with identities only in the 20–40% range is relatively uninformative (in fact, such low-scoring hits did not show the local synteny seen with those having higher identities). Evidently, anti-anti-sigmas (and corresponding antisigmas) of this general class were present in the ur-actinobacterium, giving rise to the possibility of subtle control of sigma factor activity by signals that might include morphological checkpoints (as in the case of the spoIIAA/spoIIAB genes of B. subtilis; Piggot & Hilbert, 2004) or stress (as in the case of sigB of B. subtilis; Price, 2000). Indeed, BldG influences the activity of the stress-responsive sigma factor SigH in S. coelicolor (Sevcikova et al., 2010; Takano et al., 2011), and the anti-anti-sigma/antisigma/sigma interactions of this general type have considerable potential for promiscuity in Streptomyces (Kim et al., 2008a,b; Sevcikova et al., 2010; Takano et al., 2011).
The problem of recognising orthologues among large families of paralogues is less severe with the phylogenetically distinct ECF class of sigmas and their antisigma partners, which are more diverse than the class regulated through BldG-like cascades, and usually show high partner specificity (Staron et al., 2009). bldN, a direct target of BldD, encodes one of about 50 S. coelicolor ECF sigma factors (Bibb et al., 2000; den Hengst et al., 2010). At least in S. venezuelae, BldN is a direct activator of the genes for chaplins (and their associated rodlins): amphipathic proteins that assemble at air–water interfaces and coat incipient aerial hyphae, facilitating their emergence into the air (Bibb et al., 2012; see below). This emergence into the air has been suggested as a trigger for the sporulation pathway controlled by the whi genes (Claessen et al., 2006). Convincing BldN reciprocal hits (at well over 50% identity and with local synteny) were found only among morphologically complex genera of actinomycetes (Fig. 3), suggesting a close connection of bldN with the emergence of complexity (reciprocal hits with other actinobacteria were all at well under 40% identity and lacked discernible synteny).
It has been demonstrated that, in S. venezuelae, an antisigma factor controlling BldN is encoded by the adjacent gene, termed rsbN (= SCO3324 in S.coelicolor; Bibb et al., 2012). In blastp analysis, a reciprocal best hit to rsbN is found next to nearly all bldN orthologues in actinomycete genomes; but, strikingly, the RsbN-like proteins are much more divergent than their BldN target or most other families of orthologous proteins of actinobacteria (Fig 6). We speculate that this may imply differences in the signal responsiveness of different RsbN proteins, thereby contributing to the differences between different organisms in the interplay of ecology and development: in other words, they may be potential agents of speciation.
Fig. 6.
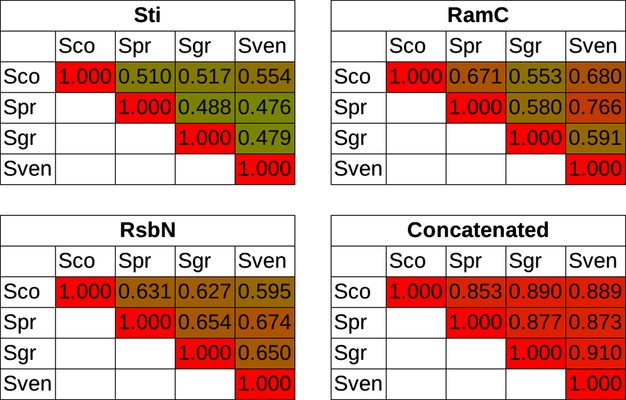
High rates of divergence of three developmentally significant proteins. Pairwise blastp comparisons are shown between three developmental proteins of four Streptomyces spp: S. coelicolor (Sco), S. pristinaespiralis (Spr), S. griseus (Sgr) and S. venezuelae (Sven). High percentage identity is indicated by the intensity of red, and low identity by the intensity of green. A control table (‘Concatenated’) shows the comparisons for a concatenated set of seven universal proteins (AtpD, DnaA, DnaG, DnaK, GyrB, RecA, RpoB).
The rsbN gene of S. venezuelae has its own promoter, which is BldN-dependent, and is also a BldD target (den Hengst et al., 2010; Bibb et al., 2012). As a bldD mutant might therefore be expected to overexpress rsbN, the resulting increase in anti-BldN activity might interfere with the expression of BldN-dependent genes and contribute significantly to the bald phenotype of bldD mutants.
The most well-studied target of BldN is bldM, which encodes an orphan response regulator (Molle & Buttner, 2000). The distribution of convincing reciprocal hits to bldM is closely similar to that of bldN hits, suggesting that the BldN to BldM regulatory step was established very early in the evolution of actinomycete complexity. The distribution of BldM was even more closely similar to that of orthologues of another developmental orphan response regulator, WhiI (Fig. 3).
The key developmental regulator AdpA emerged along with complex mycelial growth and is bldA-dependent only in Streptomycineae
BldD targets also include adpA, known as bldH in S. coelicolor (den Hengst et al., 2010). AdpA has been most comprehensively described in S. griseus, in which it is the agent of the effects of the hormone-like A-factor (Horinouchi, 2002). It comprises a structurally characterised C-terminal AraC/XylS-like DNA-binding domain (Yao et al., 2012) and an N-terminal domain that may sense adenine nucleotides (Wolanski et al., 2012; Liu et al., 2013a). It plays a central role in the decisions leading to colony differentiation, notably affecting extracellular functions such as protease cascades, extracellular morphogenetic peptides and secondary metabolism (Akanuma et al., 2009; Chater et al., 2011; Higo et al., 2012), but also contributing to the regulation of DnaA-mediated chromosome replication initiation (Wolanski et al., 2012). In S. griseus, many hundreds of direct targets for AdpA have been defined, and it is suspected that the unusually low DNA-binding specificity of AdpA may permit the ready recruitment of new targets, leading to species-specific differences in AdpA regulons (Higo et al., 2012). The phylogenetic distribution of adpA-like genes is similar to that of bldN-like genes (Fig. 3), but there is little evidence of direct regulatory interplay between the two genes. Possibly, then, AdpA evolved to regulate aspects of developmental physiology complementary to those regulated by BldN (if so, one might anticipate that some cross-checks between the two regulons will eventually be discovered).
The regulation of adpA in streptomycetes is remarkably complex (reviewed in detail in Liu et al., 2013a). It involves at least three levels of control: transcriptional [autorepression (Kato et al., 2005), repression by BldD (den Hengst et al., 2010), repression by gamma-butyrolactone-binding proteins (Horinouchi, 2007; Xu et al., 2009)]; mRNA processing by RNaseE (Xu et al., 2010); and mRNA translation (Nguyen et al., 2003; Takano et al., 2003). Translational regulation is via a very rare UUA codon in the adpA mRNA, falling between the segments encoding the two domains of AdpA. UUA is the only one of the six leucine codons to comprise only A and U residues, so the corresponding TTA codon is comparatively rare in GC-rich genomes – it occurs in only 147 chromosomal genes in S. coelicolor (Li et al., 2007). UUA codons have a special regulatory role in Streptomyces, as indicated by the finding that mutants (bldA) in the gene for the UUA-reading tRNA grow well, but fail to form aerial mycelium or some antibiotics (Merrick, 1976; Lawlor et al., 1987). adpA is the only gene that has a TTA codon in all the streptomycetes analysed (Fig. 3; Table 2; Chater & Chandra, 2008), a feature also found in the adpA orthologue in Kitasatospora setae. The TTA codon in adpA was shown by mutagenesis to be the main (but not entire) cause of the Bld phenotype of bldA mutants of S. coelicolor (Nguyen et al., 2003; Takano et al., 2003). A study of S. griseus and S. coelicolor has shown that the abundance of bldA tRNA is important in determining whether AdpA reaches levels sufficient to activate development and, remarkably, that there is a mutual feedforward mechanism in which AdpA activates bldA transcription (Higo et al., 2011). However, the adpA-like genes of other actinomycetes, including Catenulispora acidiphila (the closest genome-sequenced relative of Streptomyces and K. setae), are nearly all TTA-free (in the single exception, Nakamurella multipartita, the TTA codon is not located in the interdomain-coding region, but close to the 3′-end of the gene). Thus, bldA-adpA interplay was apparently established after node 7 (Fig. 2), branching to Catenulispora, but before the Streptomyces and Kitasatospora lines diverged (node 8). Indeed, the broader developmental significance of bldA may not extend beyond Streptomycineae, as in non-Streptomycineae genomes TTA codons do not show the positional bias towards the start of genes that is observed in streptomycetes, and sometimes occur in conserved growth-associated genes (Chater & Chandra, 2008). Interestingly, there is a strong target for BldD binding within bldA (den Hengst et al., 2010).
Table 2.
Streptomyces genes or gene clusters that frequently contain TTA codons*
| SCO number of gene *asterisk means TTA codons are absent from the S. coelicolor gene | Function | Fraction of TTA-containing orthologues among 14 Streptomyces genomes (see note) | K. setae orthologue, accepting > 40% identity (TTA present?) |
|---|---|---|---|
| 0381*;0382*;0383 | Enzymatic modifications | 6/12; 5/9; 2/10 | – |
| 0683*;0684*;0685* | Unknown | 2/12; 4/13; 2/13 |
67090 (–) *67100 (–) *67110 (–) |
| 1187(celB);1188* (celS2*) | Cellulose utilisation | 6/13; 7/14 | – *59340 (–) |
| 1242 | DNA-binding regulatory protein (WhiJ-like) | 9/11 | 58780 (–) |
| 1256* | Unknown | 6/10 | – |
| 1434 | Secreted AAA ATPase | 8/13 | 11520 (–) |
| 1592 | ADPribose pyrophosphatase | 8/12 | 41440 (–) |
| 1980 | Possible antisigma factor (AbaA-like) | 9/14 | – |
| 2426 | Regulatory | 8/14 | – |
| 2567*; 2568* | Competence operon | 3/11; 5/13 |
25860 (TTA) *25870 (–) |
| 2792 (adpA) | Major developmental regulator | 14/14 | 26930 (TTA) |
| 3195* | Unknown | 5/14 | – |
| 3423 | Possible antisigma factor (AbaA-like) | 12/14 | 17120 (TTA) |
| 3550* (widespread among actinobacteria) | Helicase | 6/14 | 35850 (–) |
| 3919* (abaB*) | LysR-like regulator of antibiotic biosynthesis | 7/13 | – |
| 3943* (rstP*) | LacI-like regulator | 5/14 | 40040 (–) |
| 4114 (widespread among actinobacteria) | Sporulation-associated protein (ankyrin-like repeats) | 9/14 | 43220 (–) |
| 4263 | LuxR-like regulator | 8/11 | – |
| 4395 | Hydroxylase | 12/14 | – |
| 5203 | 6/14 | 49090 | |
| 5460 | Possible antisigma factor (AbaA-like) | 6/14 | 50850 (TTA) |
| 5495 | Cyclic di-GMP cyclase/phosphodiesterase | 7/14 | 51100 (TTA) |
| 5970 | 6/13 | 26520 | |
| 6156* | Possible antisigma factor (AbaA-like) | 5/14 | 19600 (–) |
| 6245* | Unknown | 5/13 | – |
| 6476 | Unknown | 7/14 | – |
| 6681(amfC); 6685* (amfR*) | SapB biosynthetic enzyme; regulator of SapB biosynthesis | 2/12; 7/12 | – |
| 7251 | Possible phosphotransferase | 10/12 | 16190 |
| 7465 (cvnC13) | Component of conservon | 9/12 | 37840 (TTA) |
Throughout, examples included had a TTA codon in at least five genomes other than S. coelicolor (arbitrarily chosen as a level likely to indicate adaptive value). The table includes gene clusters likely to share a physiological role, selected because of the frequent occurrence of a TTA codon in one or another gene of the cluster.
Previously unnoticed aspects of the occurrence of conserved TTA codons
Earlier analyses had indicated that most of the S. coelicolor TTA-containing genes were absent from the few other Streptomyces genomes then available, and where the genes were conserved, the TTA codons often were not (Li et al., 2007; Chater & Chandra, 2008). With the availability of more genome sequences, it became possible to make a more sensitive search for orthologues of the 147 S. coelicolor TTA-containing genes (or in some cases gene clusters). We identified 19 that were widespread and frequently TTA-containing in 13 other Streptomyces genomes (Table 2). In addition, a further 10 genes or gene clusters frequently had TTA codons, even though their S. coelicolor orthologues were TTA-free (asterisks in Table 2). As 27/29 of the TTA-containing genes/clusters were found only in Streptomycineae, we infer that these genes and their TTA codons have adaptive value to streptomycetes and not to other actinobacteria. As shown in Table 2, about half of these genes encode proteins likely to be closely implicated in gene regulation or signal transduction, although their targets are mostly unknown. They include five conserved paralogues of genes found in the whiJ cluster (see below for further discussion) and gene sets for highly modified oligopeptides that contribute significantly to the ability of aerial hyphae to grow into the air (such as SapB, also discussed below).
Evolution of the multigene whiJ system, which represses development, and its abundant paralogues
The central feature of the complex whiJ locus of S. coelicolor is the whiJ gene (SCO4543) for a deduced DNA-binding protein (Gehring et al., 2000; Ainsa et al., 2010). Most of the 24 paralogues of whiJ in the S. coelicolor chromosome are associated with one or both of two kinds of immediately neighbouring genes, one kind encoding very small DNA-binding proteins (i.e. like SCO4542) and the other encoding proteins with features like antisigma factors (e.g. SCO4544) (Gehring et al., 2000; Ainsa et al., 2010). whiJ-like genes are widely present in complex actinobacteria, but they are absent from morphologically simple ones (corynebacteria, mycobacteria, rhodococci, propionibacteria and micrococci except Beutenbergia and Intrasporangium) and from nonactinobacterial bacteria. These genes are often clustered with one or both types of whiJ-associated genes. Most mycelial actinomycetes have two or three WhiJ paralogues, but K. setae has five, and all streptomycetes have more than 10, sometimes more than 20. Phylogenetic analysis of WhiJ paralogues from four well-studied streptomycetes is shown in Fig. 7. The branching pattern is consistent with underlying sequential gene duplication events in an early progenitor of the four streptomycetes, followed by lineage-specific further divergence and duplication events. Phylogenetic analyses of the two whiJ-associated gene families gave broadly similar patterns, supportive of the idea that the genes in each cluster co-evolved (results not shown). WhiJ paralogues in S. coelicolor vary considerably in their conservation in other organisms. One (SCO3421) was present in nearly all complex actinomycetes, and the adjacent gene encoding a likely antisigma factor nearly always contained a TTA codon in Streptomycineae (but in no other groups). Another present in all Streptomycineae (SCO4441) was also widespread among other complex actinomycetes. Four others were found in all or nearly all Streptomycineae but no other groups (SCO1242, 1979, 2513, 6236), among which a TTA was present in SCO1242 and in the antisigma factor gene next to SCO1979. Future studies might profitably focus on these six relatively long-established clusters. Twelve other S. coelicolor whiJ-like genes were represented in around half of streptomycetes (SCO2381*, 2865, 2869*, 3365*, 4176, 4301, 4678, 4998, 6129, 6537, 6629*, 7579) [asterisks indicate occurrence also in some other complex actinomycete(s)]; while seven others were found in four or fewer streptomycetes (SCO0704, 2246, 2253, 4543, 5125, 6003, 7615). TTA codons were not associated with any of the last 19 genes mentioned or their associated genes.
Fig. 7.
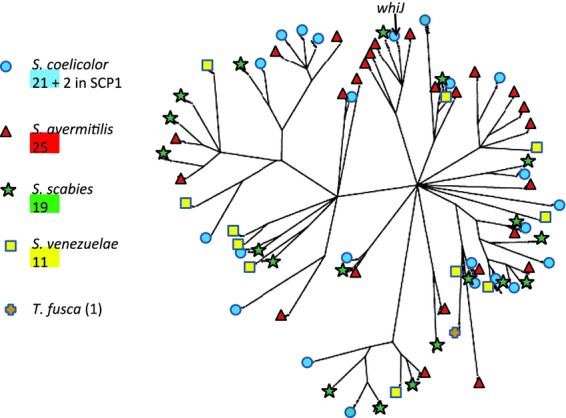
Phylogenetic analysis of WhiJ and its paralogues in four streptomycetes and another complex actinomycete. Genes encoding WhiJ paralogues were identified by probing translated gene products of the genomes of four streptomycetes [S. coelicolor A3(2) (blue circles); S. avermitilis (red triangles); S. scabies (green stars); S. venezuelae (yellow squares)] and Thermobifida fusca (brown crosses). The tree represents a phylogenetic analysis using PHYLIP (Felsenstein, 1989, 2005).
Certain mutations in whiJ gave rise to a white-colony appearance caused by a deficiency in sporulation, although the complete deletion of whiJ had no obvious phenotypic consequences (Ainsa et al., 2010). A mutant lacking the whiJ-neighbouring gene SCO4542, encoding a predicted small DNA-binding protein, had a bald colony phenotype and overproduced the pigmented antibiotic actinorhodin. This phenotype was entirely suppressed by the co-deletion of whiJ itself. Putting these observations together, it was suggested that WhiJ acts mainly to repress reproductive development until a suitable signal has been perceived via the SCO4542 DNA-binding protein, which would then directly interact with WhiJ to relieve repression (Ainsa et al., 2010). It is thought that WhiJ mediates its effects both on the emergence of aerial hyphae and, separately, on their further differentiation into spore chains. There is no information about the direct or indirect targets of WhiJ regulation or about the role of the antisigma-like protein (SCO4544).
The apparently repressing action of the whiJ locus raises the possibility that some or all of its paralogues may also act as developmental brakes. If so, it may be that during ‘normal’ colony development on laboratory media, these brakes are all off – in other words, all relevant checkpoints have been passed, and the WhiJ-like proteins are not repressing their target genes. The acquisition of additional clusters would presumably confer species-specific environmental adaptations. As different streptomycetes vary in their ability to develop normally on different media, it is possible that this (partly) reflects differences in the complement of WhiJ-like signal transduction cascades. The strikingly reduced number of paralogues in S. venezuelae (one of several streptomycetes that lack a cluster orthologous to whiJ itself) may underpin the ability of S. venezuelae to sporulate exceptionally readily and comprehensively even in submerged culture, which has led to its adoption as a model system for development (Flärdh & Buttner, 2009; Bibb et al., 2012). Like the wbl genes described earlier, whiJ-like clusters are also found in plasmids (Bentley et al., 2004), permitting horizontal transfer. Interestingly, one of the ‘classical’ bld genes, bldB, encodes a diverged member of the SCO4542 family, but is an ‘orphan’ lacking neighbouring whiJ- or SCO4544-like genes. It is curious that bldB is the only classical bld gene to be confined to, yet universal among, streptomycetes (Fig. 3). We speculate that the bald phenotype of bldB mutants could imply a promiscuous interaction of BldB with WhiJ-like proteins encoded elsewhere in the genome and that this may be connected with the large numbers of such proteins found in streptomycetes.
Special features of actinobacterial cell biology have contributed to the evolution of developmental complexity in Streptomyces
Cell growth and division in actinobacteria were recently thoughtfully reviewed by McCormick & Flärdh (2012) and Letek et al. (2012). Here, we consider the part played in these processes by conserved actinobacterial proteins, including some of the actinobacterial signature proteins in Table 1.
The origins of mycelial growth: actinobacteria are unusual in predominantly using polar growth
At least in streptomycetes, corynebacteria and mycobacteria, cells grow by the insertion of peptidoglycan precursors at cell poles, guided by large, pole-located complexes of a coiled-coil-containing protein, DivIVA (Flärdh, 2003; Flärdh, 2010; Letek et al., 2008). The actinobacterial divIVA gene is nearly always located immediately next to an actinobacterial signature gene encoding a small probable membrane protein (SCO2078, Table 1a). It is an interesting possibility that this protein plays a part in the adaptation of DivIVA to polar growth in actinobacteria.
In Bacillus subtilis and other rod-shaped firmicutes (nonactinobacterial Gram-positive bacteria), DivIVA has a different role: it is involved in selection of the division site midway between opposite cell poles. Nevertheless, in such firmicutes, DivIVA is located at both poles, at least partly because of an affinity for concave membrane surfaces (Strahl & Hamoen, 2012). In these organisms, DivIVA binds the cell-division-inhibitory MinJDC protein complex – a mechanism that ensures that the cell centre contains the lowest concentration of Min proteins, so that cell division is medial. At firmicute cell division, DivIVA accumulates at the nascent septum, and upon cell separation, the new and old poles have similar amounts of DivIVA (Bramkamp & van Baarle, 2009).
In contrast, DivIVA does not accumulate rapidly at nascent septa in rod-shaped actinobacteria such as corynebacteria or mycobacteria, so newborn cells have an intrinsic asymmetry with respect to polar DivIVA complexes. In M. tuberculosis, the lag in formation of a full-sized DivIVA complex at the new pole is in fact very long – it is comparable with the interval between cell divisions, as shown by fluorescence microscopy of DivIVA::GFP fusions (Kang et al., 2008). This may reflect the complexity of the tip-organising complex (‘polarisome’: Hempel et al., 2012) that assembles round DivIVA and includes cytoskeletal elements and the cell wall biosynthetic apparatus (Holmes et al., 2013). The asymmetry in DivIVA distribution underpins a striking asymmetry in cell division observed by live-cell imaging of mycobacterial cells confined to microfluidic chambers (Aldridge et al., 2012): cells grow at just one pole, which is inherited stably; the newborn cells that lack an active pole take significant time to form one, and cell division depends on time rather than cell dimensions. As a result, the population of progeny cells in a very young microcolony is physiologically heterogeneous, different cell types even differing in their patterns of sensitivity to antibiotics (Aldridge et al., 2012).
Comparative genomic analysis suggests that DivIVA-mediated apical growth is typical of actinobacteria, as, on one hand, divIVA is universally conserved (always located close to the cluster containing ftsZ and other genes concerned with division and cell wall biosynthesis); while, on the other hand, the mre gene cluster (which mediates lateral cell wall growth in nonactinobacterial rod-shaped bacteria including E. coli and B. subtilis) is absent from nearly all rod-shaped or coccal actinobacterial genera originating from nodes 3 and 4 of Fig. 2 (Fig. 8). Actinobacteria on the deepest branches (nodes 1 and 2) do have the mre cluster, so it is not possible to infer which growth mode they might use. Although coccal actinobacteria also possess DivIVA, they may possibly grow in a DivIVA/MreB-independent manner, as in staphylococci: there, peptidoglycan growth is confined to the septum, which becomes remodelled from a circular to a hemispherical form during cell separation (Touhami et al., 2004).
Fig. 8.
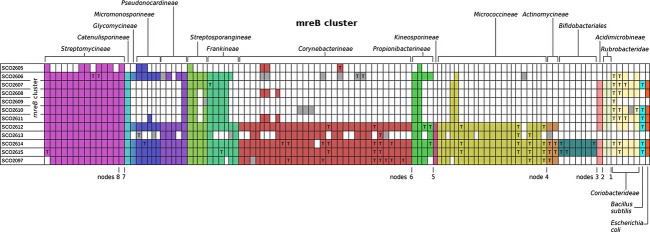
The mre gene cluster is absent from most simple actinobacteria. The reciprocal blastp best-hit tabulation includes the region from SCO2605 to SCO2615. The numbered nodes refer to Fig. 2. See Fig. 3 legend and text for further details. The mre gene (SCO2611) is part of a cluster (SCO2607-2611) present in all streptomycetes and morphologically complex actinomycetes, but absent from nearly all mycobacteria and corynebacteria (rust red), and from members of the Micrococcineae (olive yellow), Bifidobacteriales (dark green) and Rubrobacterideae (brown). Interestingly, the adjacent gene SCO2606 (encoding a likely radical SAM enzyme related to those involved in tRNA methylation) shows a very similar distribution. The Figure also shows the distribution of hits to the MreB-associated actinobacterial signature protein SCO2097 (Kleinschnitz et al., 2011).
What are the possible adaptive benefits of the two known growth modes of rod-shaped bacteria? The lateral wall growth of rod-shaped firmicutes may permit more efficient population growth, because the near-symmetry of growth and division allows both daughter cells to progress equally rapidly to subsequent divisions. In contrast, the asymmetry implicit in polar growth of rod-shaped actinobacteria has the potential to improve population resilience, because daughter cells have significantly different physiology, including different susceptibilities to some antibiotics (Aldridge et al., 2012). In the event of predivisional actinobacterial cells with three or more compartments (an apparent example of this may be seen in Fig. 5A of Singh et al., 2013), the tip-less compartments would have only nongrowing wall on their surface, with high levels of cross-linking. This might have enhanced survival value during exposure in the natural environment to physical stress, chemical or enzymatic attack of the wall itself, chemical or biochemical poisons (such as antibiotics produced by neighbouring organisms) and attack by bacteriophages. This increased resistance may have been a driving force for the evolution of mycelial growth.
A key requirement for the evolution of mycelial growth is a mechanism for cellular branching. This has become clarified by the discovery that, as a Streptomyces tip extends, it acquires increasing amounts of DivIVA (perhaps this is in some proportion to the number of genome copies in the tip compartment) and eventually splits, part remaining at the tip and part adhering to the lateral wall, which is thereby marked as a position of future branch emergence (Hempel et al., 2008; Flärdh et al., 2012). In a manner reminiscent of the situation already described for new poles in mycobacteria, branch emergence is not usually immediate, perhaps because the incipient polarisome has to be built up to some critical mass and/or organisation. However, new mycobacterial poles must be nucleated with DivIVA de novo, whereas mycelial branches are nucleated by the residue of a split polarisome, which may be a more avid target for DivIVA than septa.
Streptomyces polarisome splitting requires the activity of a serine/threonine protein kinase, AfsK, which phosphorylates DivIVA (Hempel et al., 2012). AfsK orthologues are present only in streptomycetes and K. setae, with a very weak hit also in Catenulospora acidiphila. How do other mycelial actinomycetes control branching? Most probably, by the action of other protein kinases on DivIVA – even in the nonbranching mycobacteria, DivIVA (=Wag31) is subject to phosphorylation during cell growth, but by serine/threonine protein kinases different from AfsK (Jani et al., 2010).
To achieve full mechanical strength, hyphae require FilP, a cytoskeletal coiled-coil protein that forms filaments along the hyphae (Bagchi et al., 2008). Orthologues of FilP appear to be very widespread among actinobacteria, including many morphologically simple organisms that diverged from the Streptomyces line early in evolution (e.g. bifidobacteria), but FilP is absent from coccal organisms, with the exception of Kineococcus radiodurans (Fig. 9). Thus, FilP is an ancient protein that may be important in generating resilient cylinders from the hemispherical nascent peptidoglycan emanating from poles. In streptomycetes, this involves a direct interaction with DivIVA (Fuchino et al., 2013). However, FilP is also apparently absent from corynebacteria and most mycobacteria. Possibly some other protein substitutes for it, or the sequence divergence of FilP in these organisms may be too great for identification by reciprocal blastp best-hit analysis.
Fig. 9.
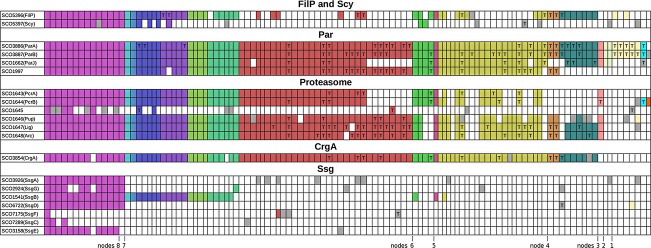
Distribution of some actinobacteria-specific cytoskeletal and related proteins. See Fig. 3 for further explanation of this reciprocal blastp best-hit analysis.
Ancient special features of actinobacterial cell division underpin the conversion of aerial hyphae into spores in Streptomyces
Surprisingly, little is understood about cell division in Streptomyces vegetative hyphae (Jakimowicz & van Wezel, 2012). Vegetative septa do not lead to cell separation, unlike their counterparts in nonmycelial actinobacteria. They are seldom found close to hyphal tips, and their positioning does not seem to conform to any recognisable pattern. A mycelium (albeit a relatively insubstantial one) can even be formed in the absence of septa, for example in an ftsZ mutant (McCormick et al., 1994; McCormick, 2009). Likewise, the mutational inactivation of chromosome partitioning functions ParA and ParB, or of the FtsK DNA translocase, has no effect on mycelial growth (Kim et al., 2000; Jakimowicz et al., 2005a; Jakimowicz et al., 2007; Wang et al., 2007; Ausmees et al., 2007; Dedrick et al., 2009). Effects of these mutations become apparent only during sporulation, when a kind of cell division more closely resembling that of unicellular organisms takes place. Here, we follow this parallel in a consideration of five partially overlapping aspects: (1) the transition from tip growth of an aerial hypha to sporulation septation; (2) the FtsZ-centred divisome; (3) partitioning of chromosomes into spores; (4) cell wall remodelling and thickening; and (5) spore germination.
(1) At the tips of aerial hyphae, a very long coiled-coil protein, Scy (Walshaw et al., 2010), interacts with two other coiled-coil proteins: DivIVA, and the FilP protein discussed in the preceding section. Scy is needed for properly organised sporulation septation (Holmes et al., 2013). Scy appears to be less widespread among actinobacteria than FilP, but its primary sequence is more prone to divergence (often a problem with comparative genomics of coiled-coil proteins), so reciprocal best-hit blastp analysis is less reliable. However, undoubted orthologues are present in some of the organisms emanating from node 4 onwards in Fig. 2 (Fig. 9). The authenticity of these hits is firm, because they are nearly all encoded by genes immediately next to filP. Most of the Scy-containing organisms show some propensity for mycelial growth. We tentatively suggest that, even in some simple actinobacteria, FilP may be involved not only in tip organisation, but also in communication between the tip and the establishment of septa, and that Scy modulates this communication in certain cell types of more complex actinobacteria, especially during reproductive fragmentation (including sporulation).
(2) In order for sporulation septation to take place, large amounts of the proteins making up the machinery of cell division must be made available. For ftsZ and parAB, this is achieved at least in part by the use of very strong sporulation-specific promoters, in addition to weaker ones used during vegetative growth. One of the most important roles of the Whi proteins discussed earlier is to cause such overexpression, in at least one case, directly (WhiA and the parAB promoter, as discussed earlier: Kaiser & Stoddard, 2011; Jakimowicz et al., 2006). Importantly, the activation of the sporulation-specific promoter of ftsZ is still not understood, although it is known that overexpression of ftsZ can partially suppress the phenotypes of all the whi mutants (Willemse et al., 2012) and that BldD represses the sporulation-specific promoter of ftsZ (den Hengst et al., 2010).
FtsZ of streptomycetes has special features dedicated to its role in sporulation, inasmuch as certain ftsZ mutations eliminate sporulation septation without eliminating vegetative septation (Grantcharova et al., 2003; Wasserstrom et al., 2013). The cell division apparatus (divisome) of actinobacteria also includes a form of DivIC so different from that of other bacteria that it was initially considered to be one of the actinobacterial signature proteins (Gao et al., 2006), until it was found by Bennett et al. (2007) to share 23/90 of the residues of conserved domain sequence 21 (pfam04977) of DivIC. (Gao & Gupta (2012) later removed this protein from their list of signature proteins because of its ‘presence in some other bacterial groups’.) The conserved domain 21 comprises a membrane-spanning segment and a C-terminal coiled-coil region located outside the cell membrane. The part of DivIC that is in the cytoplasm bears no apparent relatedness to DivIC in other bacteria. In other bacteria, DivIC cooperates with another membrane protein, FtsL, and the S. coelicolor protein likewise interacts with FtsL in a complex that contributes to both vegetative and sporulation septation. Mutants in divIC or/and ftsL are particularly affected in sporulation septation, forming many incomplete and asymmetrical septal ingrowths in aerial hyphal apical compartments of colonies grown on high osmolarity medium (Bennett et al., 2007). The primary amino acid sequence divergence between nonactinobacterial and actinobacterial DivIC proteins suggests co-evolution with some other cell division differences between organisms.
A remarkable feature of cell division in streptomycetes that appears specially significant for sporulation septation is the use of specialised actinobacteria-specific proteins (‘SALPs’, SsgA-like proteins) to determine the locations at which cell division and/or changes in cell wall structure will take place (Noens et al., 2005, 2007). Thus, SsgA is present in spores at the positions from which germ tubes will emerge, then at the growing tips of the apical compartments of aerial hyphae and then at the positions at which sporulation septation will take place (Noens et al., 2007), while the paralogous SsgB is more developmentally specific, appearing to be recruited by SsgA to the future sites of sporulation septation, where it forms a circumferential ring inside the cell membrane and directly recruits FtsZ (Willemse et al., 2011). SsgA and SsgB are present in all streptomycetes, as is SsgD, which is required for spore wall integrity (Fig. 9; Noens et al., 2005). Two other SALPs of S. coelicolor are present in some, but not all, other species: SsgE plays a part in the separation of spores, and SsgG is needed for the properly regular formation of sporulation septa (Noens et al., 2005). The other two S. coelicolor SALPs are absent from most species: SsgC, affecting the regularity of sporulation septation and partitioning of DNA into prespore compartments; and SsgF, affecting spore separation (Noens et al., 2005). It is not known whether other paralogues compensate for the absence of SsgC and SsgF in streptomycetes lacking them. Reciprocal blastp analysis with SsgB gave convincing hits with most developmentally complex actinomycetes and with Cellulomonas flavigena and Kineococcus radiodurans, but no other Ssg protein gave meaningful hits outside of Streptomycetaceae. It is likely that the acquisition of SsgB by an early actinomycete (before node 4 of Fig. 2) was a key to the later evolution of sporulation septation, but SALPs have been entirely lost independently from several subsequent branches.
The near-universal actinobacterial protein CrgA (Table 1, Fig. 9) has been studied in two streptomycetes (Del Sol et al., 2003, 2006) and in M. tuberculosis and M. smegmatis (Plocinski et al., 2011, 2012). It is a small protein with a C-terminal transmembrane domain. In M. tuberculosis, CrgA is abundant and interacts directly with cell division proteins (FtsZ, FtsQ, FtsI and PBPA) and with a newly characterised membrane protein, CwsA, that in turn interacts with the mycobacterial DivIVA orthologue Wag31 (Plocinski et al., 2012). It has been suggested that one role of CrgA in mycobacteria is to promote and/or stabilise FtsI localisation, facilitating septum formation (Plocinski et al., 2011) and helping to coordinate septal and polar peptidoglycan synthesis with FtsZ-ring assembly (Plocinski et al., 2012).
In streptomycetes, CrgA is considered to coordinate sporulation septation with hyphal growth, although it does not localise either to growing tips or to the sites of septation (Del Sol et al., 2006). This failure to co-localise with DivIVA is consistent with the absence of an obvious CwsA-like protein in S. coelicolor. The overexpression of CrgA inhibits septation (Del Sol et al., 2006), an effect that could be a secondary consequence of the over-occupation by CrgA of its various interaction partners. Interestingly, crgA mutants of streptomycetes do not always have the same phenotype: an S. coelicolor mutant shows premature production of spores that are slightly aberrant, but in the S. avermitilis mutant, sporulation septation does not take place (Del Sol et al., 2003).
(3) Actinobacteria all possess the ParABS partitioning system common to the majority of bacteria (but absent from E. coli). In this system, the ParB DNA-binding protein associates with newly replicated ParS sites located close to the chromosomal origin of replication (oriC), the two complexes being recognised by the ATPase motor protein ParA, which drives them apart (Toro & Shapiro 2010). The ParABS system has been shown to be important for the reliable segregation of chromosomes of corynebacteria and mycobacteria during normal growth, with ParB, ParA and Wag31 (= DivIVA) interacting in all pairwise combinations, presumably allowing coupling of polarisome activity to segregation (Donovan et al., 2010, 2012; Ginda et al., 2013). In these organisms, the Par proteins are predominantly associated with the cell pole (Donovan et al., 2010, 2012; Ginda et al., 2013), although transient localisation at the corynebacterial division septum is also seen for ParB, which appears to interact directly with FtsZ (Donovan et al., 2010). Thus, even in morphologically simple actinomycetes, the ParAB system provides a system of interplay between the growing pole, the septum and the chromosomal oriC. It appears that this has evolved to coordinate the more complex process of Streptomyces sporulation septation (Kim et al., 2000; Jakimowicz et al., 2005a, 2007).
Although the mechanism coordinating cessation of growth with the cessation of DNA replication and the initiation of chromosome partitioning in sporulating aerial hyphae of streptomycetes is not understood, a possible clue comes from vegetatively growing B. subtilis, in which there is interplay of the replication initiator protein DnaA with ParB (=Spo0J) and ParA (=Soj). DnaA is strongly inhibited by Soj monomers, but stimulated by ATP-dependent Soj dimers. The Soj monomers are generated from Soj/Spo0A complexes, simultaneously freeing Spo0A to form large partitioning complexes with the chromosomal oriC region (Scholefield et al., 2011). In these ParB/oriC complexes, ParB interferes with access of DnaA to oriC. During Streptomyces sporulation, ParB forms large complexes with the oriC region that probably have the same effect (Jakimowicz et al., 2002). Formation of these ParB complexes requires a shift in the behaviour of ParA, which is tip-associated while aerial hyphae are extending, but forms apparent helical filaments along the hyphae when growth stops (Jakimowicz et al., 2007). The ParA filaments dissociate immediately before sporulation septation and chromosome partitioning take place; and chromosome partitioning into prespore compartments is markedly irregular in the absence of ParA filaments or ParB (Jakimowicz et al., 2007). Dissociation involves interaction of ParA filaments with SCO1662 protein, one of two similar proteins (the other, SCO1997, shows near end-to-end alignment with SCO1662) that are among the ‘universally’ conserved actinobacterial proteins (Table 1A; Gao et al., 2006; Ditkowski et al., 2010). Like ParA, SCO1662 protein, renamed ParJ, is important for accurate chromosome partitioning into prespore compartments (Ditkowski et al., 2010). Actinobacteria lacking ParJ possess its SCO1997-like paralogue (Fig. 9). By analogy with B. subtilis, the action of ParJ in causing ParA polymers to dissociate may lead to inhibition of DnaA, reinforcing the repression of further DNA replication (but there is no published evidence of a ParA-DnaA interaction in any actinobacteria). As these proteins are all widespread in actinobacteria, such a mechanism could have ancient roots (Fig. 9).
Structures have been determined for the ParJ orthologue of S. avermitilis (Chang et al., 2010) and for SCO1997 protein (Gao et al., 2009) and its equivalents from Corynebacterium glutamicum and M. tuberculosis (Zhang et al., 2007; Graña et al., 2009). Although ParJ and SCO1997 proteins are only 28% identical to each other at the amino acid sequence level, both are structurally related to the PAC2 family of proteins (Gao & Gupta, 2012), which in eukaryotes are chaperones for the assembly of 20S proteasomes. Interestingly, although proteasomes are nearly universal among actinobacteria, they are absent from nearly all other bacteria (Striebel et al., 2013; Fig. 9) [proteasome-associated proteins were not listed among the actinobacterial signature proteins (Gao et al., 2006), probably because they happen to have been absent from two of the four organisms surveyed by those authors]. It therefore seems worth studying whether proteasomes also have an interface with this stage of the sporulation septation process. No genetic or cytological analysis of SCO1997 function has been reported, but all actinobacteria also possess a second parA-like gene (e.g. SCO1772, parA2: Fig. 9). In C. glutamicum, the product of this parA paralogue, named PldP, has been found to interact with ParB and to play a part in division site selection: PldP is predominantly found at developing septa (Donovan et al., 2010). Perhaps, in S. coelicolor, SCO1997 protein interacts with ParA2 to influence its aggregation state.
Although the Par system positions the oriC regions correctly, the parts of the sister chromosomes trailing behind the partitioning complex could potentially be guillotined by ingrowing septa, particularly in actinobacteria in which the septa are eccentrically located and no system of nucleoid occlusion of septation normally operates (although such a system may become operational in certain Streptomyces genotypes in which chromosome condensation is abnormal: Facey et al., 2009). However, as in most bacteria, FtsK-like DNA translocases (Reyes-Lamothe et al., 2012) are able to mobilise chromosome DNA through the septum as it closes, both in mycobacteria (Singh et al., 2013) and in sporulating aerial hyphae of S. coelicolor (Ausmees et al., 2007; Wang et al., 2007; Dedrick et al., 2009). FtsK orthologues were found in 97% of the actinobacteria considered in this article. In addition, a second weakly FtsK-like protein (SCO1416, SffA) is present in all streptomycetes, Kitasataspora setae and Catenulospora acidiphila, along with a small membrane protein SmeA encoded by the adjacent gene, SCO1415. SmeA and SffA are absent from all other actinobacteria, and both are specifically targeted to sporulation septa (Ausmees et al., 2007). Their roles are not known, but mutation of smeA causes aberrant sporulation septation (Ausmees et al., 2007).
The inclusion of a complete chromosome in each spore compartment is also aided by proteins that bring about nucleoid compaction. Facey et al. (2009) showed that S. coelicolor has three Dps proteins, which are all involved in sporulation-associated nucleoid partitioning and compaction as well as in the osmotic stress response. DpsB (SCO5756) is represented in most streptomycetes and many other actinobacteria, but DpsA (SCO0596) is less universally present, and DpsC (SCO1050) is absent from most. An intriguing interplay between the Dps proteins of S. coelicolor, indicated by surprising differences in the nucleoid compaction phenotypes of various single and double mutants (Facey et al., 2009), therefore does not seem to be generalisable among streptomycetes; and an extended analysis of the evolution of the three proteins (Facey et al., 2013) does not clarify this interplay. Contributions to spore nucleoid partitioning and compaction are also made by the Smc protein and its partner proteins ScpA and ScpB (Dedrick et al., 2009; Kois et al., 2009), and by sIHF (SCO1480: Yang et al., 2012; Swiercz et al., 2013). These proteins are also present in nearly all actinobacteria, SCO1480 being one of the actinobacteria-specific proteins listed by Gao et al. (2006) (Table 1). Another of the actinobacteria-specific proteins in Table 1, Lsr2 (SCO3375), is also a nucleoid-associated protein, functionally equivalent to H-NS of E. coli (Gordon et al., 2010), but it is not known whether Lsr2 has a developmental role in streptomycetes [all streptomycetes, and a few other actinomycetes, have a second Lsr2-like protein (e.g. SCO4076)]. The apparent absence of any effects of elimination of Dps, Smc or Scp proteins, and the relatively slight effect of sIHF elimination, on vegetative growth of S. coelicolor, are consistent with the idea that sporulation is a specialised version of a process that takes place even in unicellular actinobacteria, perhaps upon entry into stationary phase. The sIHF orthologues in M. tuberculosis and M. smegmatis appear to be essential (mIHF: Pedulla & Hatfull, 1998; Sassetti et al., 2003).
Another type of DNA-packaging protein, HupS, contributes to the final packaging of DNA in spores, although apparently not to the completion of partitioning (Salerno et al., 2009). HupS resembles HU proteins found throughout bacteria, but contains an extra domain peculiar to actinomycetes. HupS orthologues are absent from morphologically simple actinobacteria, including Corynebacterium spp. and most Micrococcineae, but they are present in nearly all mycelial actinobacteria and Mycobacterium spp. Interestingly, the M. tuberculosis orthologue (Hlp) is upregulated in anaerobically induced dormancy (Lee et al., 1998). As HupS contributes to resistance of spores to heat (Salerno et al., 2009), the first acquisition of its progenitor may have provided positive selection for the evolution of sporulation in actinobacteria.
(4) The remodelling and thickening of the cell wall of cylindrical, thin-walled prespore compartments, and their separation to generate near-spherical thick-walled spores, are carried out by a multiprotein complex organised by MreB and its paralogue Mbl (Heichlinger et al., 2011; Kleinschnitz et al., 2011). One of the proteins in this complex is the product of the actinomycete signature gene SCO2097 (Kleinschnitz et al., 2011; Fig. 8). Thus, SCO2097 may be involved in the formation or rounding off of sporulation septa prior to separation, or the maturation of peptidoglycan at cell poles (which, during growth and cell division, contain nascent peptidoglycan that is not fully cross-linked), or the thickening of spore walls. These processes may also be needed in the formation of resting cells even of simple actinomycetes (and presumably also the ancestral ur-actinomycete): at least in M. tuberculosis, anaerobically grown resting cells not only have increased levels of the DNA-packaging protein Hpl (see preceding section) but also have thicker walls than aerobically grown cells (Cunningham & Spreadbury, 1998).
(5) Part of the biomass of incipent stationary phase cells is sacrificed to adaptations that improve survival against the ravages of time and environmental insults. These adaptations are most pronounced in spores, with their thickened cell walls, accumulation of stored reserves such as trehalose, and special packaging of nonreplicating DNA. Some of these features may act as barriers to growth, predicating the evolution of germination mechanisms, for example to permit breaking out from the thickened wall, and the casting off of DNA packaging. A widely conserved actinobacteria-specific protein family, represented in Gao et al. (2006) by the product of the Mycobacterium leprae gene ML2030, is made up of ‘resuscitation-promoting factors’ (RPF). The first example of an RPF was isolated from stationary phase culture fluids of Micrococcus luteus, where it was shown to greatly increase the number of colonies generated by plating out suspensions of stationary phase cells (Mukamolova et al., 1998). The closest relative of ML2030 in S. coelicolor is SCO3097, although the absence of local synteny makes it unclear whether they are true orthologues. The 70-aa Rpf domain of SCO3097 is also found in four other secreted proteins in S. coelicolor. Proteins possessing this domain are present in nearly all actinomycetes, often as a small series of paralogues. They are generally predicted to be secreted. The Rpf domain, which was predicted to be muralytic on bioinformatic grounds (Ravagnani et al., 2005), possesses a lysozyme-like fold (Cohen-Gonsaud et al., 2005; Ruggiero et al., 2009) and, at least in two RPFs, has demonstrable peptidoglycan hydrolase activity (Mukamolova et al., 2006; Haiser et al., 2009). Mutational analysis indicates considerable functional redundancy among RPFs in mycobacteria (reviewed by Kana & Mizrahi, 2010), but phenotypic effects were observed in S. coelicolor when SCO3097 was disrupted: the mutant produced thin-walled, heat-sensitive, irregular spores in chains that tended not to separate as readily as wild-type spore chains and showed a modest delay in germination (Haiser et al., 2009).
Interestingly, RpfB and RpfE (but not RfpA, C or D) proteins of M. tuberculosis are found in a synergistically acting complex with another kind of peptidoglycan hydrolase (RipA; Hett et al., 2007, 2008) and appear to play dual roles in cell wall growth and septation on one hand, and emergence from dormancy on the other. Clearly, such hydrolytic activity must be highly controlled if cell wall integrity is to be retained. One mechanism for this control may involve the peptidoglycan biosynthetic enzyme PBP1, which interacts with the RpfB/RipA complex and inhibits its hydrolase activity, most likely by a RipA partner-switching mechanism, as RipA uses the same C-terminal 25 aa to interact with PBP1 and RpfB (Hett et al., 2010). In organisms exhibiting polar growth, septation requires the nongrowing (and presumably most extensively cross-linked) part of the cell wall to be re-established as a template for renewed synthesis, so Rpf proteins may be involved in this. RpfB and RipA localise to the septa of mycobacterial cells (Hett et al., 2007), while PBP1 localises mainly to cell poles but also to some septa (Hett et al., 2010).
Re-establishment of cell wall growth is also needed for the emergence of hyphal side branches. Could Rpf proteins and RipA homologues influence branching, including aerial branches, in Streptomyces? Streptomycetes have proteins modestly resembling RipA – the most similar, SCO4793, has near end-to-end c. 30% identity with RipA, but has an additional central segment. The evolutionary origin of the RPF domain and its interaction with RipA-like proteins may be hinted at by the finding that the ‘tape-measure’ proteins of many actinophages include both domains and, in at least one instance, can degrade the cell walls of stationary phase host bacteria (Piuri & Hatfull, 2006).
Extracellular functions important for Streptomyces sporulation mostly entered the actinobacterial lineage at the time when morphological complexity emerged
Streptomyces aerial growth and sporulation depend on both nutritional and mechanical support. Some of the elements involved are extracellular and have been discussed in some detail (Chater et al., 2010). Here, we show that the acquisition of some of these elements coincided with the last node preceding the emergence of complex actinomycetes (Fig. 2), raising the possibility of causal connections.
An extracellular protease cascade
One important source of nutrients for aerial growth is provided by lysis (sometimes referred to as ‘programmed cell death’: Manteca et al., 2007) of the part of the mycelium from which sporulating aerial hyphae emerge. Studies in S. coelicolor and several other species indicate that this lytic process involves an extracellular protease cascade (Chater et al., 2010). The cascade is held inactive by an extracellular protease inhibitor protein, such as Sti of S. coelicolor (Kim et al., 2008a). Using blastp reciprocal best hits as a guide, most Streptomyces genomes contain a sti-like gene, and such genes are also present in most developmentally complex actinomycetes (Catenulosporineae, Glycomycineae, Micromonosporineae, Pseudonocardineae and Streptosporangineae, but not Frankineae: Table 3). This suggests that extracellular protease cascades may have been significant in the evolution of actinomycete complexity. Among streptomycetes, the sequences of the protease inhibitors diverge more than most other conserved gene products, even though there is detectable synteny of the determinants (Table 3; see also Fig. 6). It is therefore possible that divergence of the different inhibitors may have contributed to speciation, with the inhibitors being species-specific. A similar argument was used earlier, in considering the antisigma factor RsbN.
Table 3.
Occurrence of Sti-like protease inhibitor genes (SCO0762-like) in complex actinomycetes
| Organism | Amino acid identity in blastp | Length of overlap | Local synteny |
|---|---|---|---|
| Streptomyces lividans | 99% | 100% | Yes |
| Streptomyces avermitilis, griseoflavus, sviceus, viridochromogenes | 72–77% | c. 100% | Yes |
| Streptomyces clavuligerus, griseus, hygroscopicus, pristinaespiralis, roseosporus, scabies, venezuelae | 50–66% | > 75% | SCO0767 orthologue in most cases |
| Streptomyces albus, Kitasatospora setae | No reciprocal hit | ||
| Catenulispora | 37% | 47% | No |
| Stackebrandtia | 49% | 60% | No |
| Micromonospora aurantica, M. sp. L5 | 45% | 73% | No |
| Actinosynnemma, Amycolatopsis mediterranea, Saccharomonospora viridis, Saccharopolyspora erythraea | 36–41% | 61–80% | No |
Sti has two activities – a general one against serine proteases and a specific one mediated via specialised C-terminal interaction domains on target proteases (P-domains: Kim et al., 2008a; Chater et al., 2010). Two P-domain proteases, SCO1355 (serine peptidase) and SCO5447 (neutral zinc metalloprotease), are present in S. coelicolor. A survey of the genomes in StrepDB (http://strepdb.streptomyces.org.uk) shows P-domains to be present typically in up to five proteins in any one streptomycete (Table 4). Although these proteins were mostly serine proteases, two examples of P-domain aminopeptidases were found. The six genomes surveyed all encoded other proteases closely similar to the P-domain-containing proteases, but lacking the C-terminal P-domain. This complicates consideration of the significance of the Sti-like protease inhibitors, as the proteases without P-domains would presumable bypass inhibition. Possibly, there are important differences in the regulation or substrate specificities of the proteases with and without P-domains. Future studies may address the question of how P-domains have been acquired or lost over evolutionary time.
Table 4.
P-domain-containing proteins in six streptomycetes
| Streptomyces species | P-domain-containing neutral zinc metalloendoproteases (orthologue) | P-domain-containing serine endo- /exopeptidases (orthologue) | P-domain-containing putative aminopeptidases |
|---|---|---|---|
| S. coelicolor | SCO5447* | SCO1355 | – |
| S. avermitilis | SAV_2794*, 1037 | – | – |
| S. clavuligerus | SCLAV_4359 | – | – |
| S. griseus | SGR_2095 (= sgmA) | SGR_6802, 2549, 918 (3 P-domains) | SGR_5809 |
| S. scabies | SCAB27921* | 20151, 89701 | – |
| S. venezuelae | SVEN_5109 | SVEN_4741, 6538, 6996, | SVEN_4288 |
Adjacent gene encodes close paralogue without a P-domain.
Sti-mediated inhibition of the S. coelicolor protease cascade is released by specific cleavage of Sti by another protease, SCO5913 (Kim et al., 2008a,b). blastp analysis showed that orthologues of the Sti-degrading protease SCO5913 were absent from all species examined except S. lividans, a very close relative of S. coelicolor that has a nearly identical Sti determinant. Thus, the set of proteases involved in developmental proteolysis may differ in different streptomycetes.
Analogies with eukaryotic P-domain proteases suggest that, on their release from inhibition, Streptomyces P-domain proteases cleave, and thereby activate, pro-enzymes that then go on to take part in development. One such protein may be transglutaminase (TGase), which may be involved in cross-linking surface-located proteins (Zotzel et al., 2003a,b; Zhang et al., 2008b), although TGase is absent from some species including S. coelicolor. Among the substrates for Streptomyces morbaraensis TGase are three protease-inhibiting proteins, each having specificity for a different protease involved in TGase activation (Schmidt et al., 2008; Sarafeddinov et al., 2011). Not all Sti-like proteins from other streptomycetes are substrates for the TGase (Taguchi et al., 2000), perhaps providing a window into the unusually highly diverged phylogeny of Sti orthologues referred to above. In a further twist, it appears that in Streptomyces hygroscopicus, the action of TGase results in the incorporation of the surfactant Sti-like protein into the surface of aerial hyphae (Zhang et al., 2008a), which may contribute in some way to the exoskeleton that we describe next.
Exoskeletal proteins
Physical support for aerial growth of streptomycetes is provided by various secreted amphipathic proteins (chaplins and rodlins) and peptides (SapB-like) that can assemble on the hyphal surface (Claessen et al., 2003, 2004; Elliot et al., 2003; Willey et al., 2006; Capstick et al., 2007, 2011). These may solve two problems – the breaching of the surface tension at an air–water interface when aerial hyphae emerge from a hydrated environment, and the provision of a hydrated extracellular compartment to aerial hyphae, permitting ‘normal’ cell physiology such as the generation of membrane potential to be sustained, as well as potentially providing a route for nutrients to reach the apical compartment (Wosten & Willey, 2000; Chater et al., 2010; Chater, 2011).
Chaplins typically occur in long and short forms, the long chaplins containing a C-terminal domain that is a substrate for attachment to the cell wall by the action of sortase enzymes. Chaplins assemble at air–water interfaces and on the surface of hyphae growing into the air (Claessen et al., 2003; Elliot et al., 2003; Elliot & Talbot, 2004). All Streptomycetes have several chaplins. Streptomyces coelicolor has eight, but none of these is represented in every streptomycetes: four (ChpB, C, D, E) are found in more than half of other species (Di Berardo et al., 2008); and ChpB and ChpD reciprocal blastp hits are present in the nearest known relatives of streptomycetes, K. setae and Catenulispora acidiphila. Further blastp analysis revealed that K. setae had four other short chaplins. When the K. setae chaplin complement was used in blastp reciprocal best-hit analysis, further genes for chaplin-like proteins were found in a few developmentally complex organisms that are more remotely related to streptomycetes (Stackebrandtia nassauensis, Thermobifida fusca, Streptosporangium roseum and Nocardiopsis dassonvillei) and even in the nonsporulating rod-coccus Arthrobacter chlorophenolicus. It has been shown that some chaplins also play a significant role in mycelial attachment to surfaces (De Jong et al., 2009b), a function that might conceivably have preceded their role in aerial growth and might explain their presence in organisms not known to exhibit aerial growth.
Chaplins can assemble into paired rodlet structures under the influence of rodlin proteins, of which there are two in S. coelicolor. Obvious rodlins were found only in streptomycetes, although very low-scoring reciprocal blastp best hits were present in a few other morphologically complex actinomycetes (Thermomonospora curvata and Stackebrandtia nassauensis). They appear to be lost readily, as S. avermitilis, S. griseoflavus and S. hygroscopicus lack rodlin genes.
The S.coelicolor modified oligopeptide SapB is a post-translationally processed product of the small gene ramS (SCO6682; Kodani et al., 2004). AmfS, the equivalent oligopeptide of S. griseus, has also been studied extensively (Ueda et al., 2002). Processing is carried out by the product of the adjacent gene ramC (SCO6681) and includes the generation of lanthionine bridges like those found in lantibiotics. Two adjacent transporter genes (SCO6683, SCO6684) are thought to be responsible for SapB export, and the cluster depends on the regulatory gene ramR (SCO6685). We found similar clusters in most streptomycetes (S. hygroscopicus was an exception) and sporadically among other morphologically complex genera (but ramS orthologues are sometimes missed in the reciprocal blastp analysis because these genes are very small). As already mentioned, most Streptomyces amf-like clusters contain a TTA codon either in the ramR-like gene or in the ramC orthologue. We found one occurrence of a ramS-like gene among simpler actinomycetes, in Kribbella flavida, but in this case, the orthologues of the rest of the amf cluster were scattered. The regulatory gene ramR was found only in streptomycetes.
As in the cases of Sti-like protease inhibitors and RsbN antisigma factors already discussed, apparent orthologues of the genes in the amf cluster are unusually diverged, which might suggest a role in speciation (Fig. 6). However, species specificity in the action of SapB-like proteins has not been reported, and a SapB-deficient mutant of S. coelicolor could be induced to undergo full aerial growth and sporulation by adding SapT, isolated from Streptomyces tendae, or a fungal hydrophobin, SC3 (Kodani et al., 2005).
Pathways related to that for a Streptomyces spore wall pigment are present in other complex actinomycetes
At a late stage of sporulation, the spore wall is modified by the attachment of an aromatic spore pigment, which may be specified either by a type II polyketide biosynthetic gene cluster (the whiE cluster of S. coelicolor: Davis & Chater, 1990) or a type III polyketide synthase (e.g. in S. griseus: Funa et al., 1999, 2005). The process may be equivalent to lignification (although such speculation is untested). In the case of whiE, a cluster of eight genes SCO5314-5321 is required (Davis & Chater, 1990). Paralogues of the core whiE polyketide synthase genes are frequently present in gene clusters for type II polyketide antibiotics, which might complicate the interpretation of reciprocal blastp analysis; but despite this, the occurrence in some non-Streptomyces genomes of several adjacent reciprocal hits to whiE genes makes a strong case for the presence of a whiE pathway. On this basis, a whiE-like pathway is present in species of Catenulispora, Frankia, Micromonospora, Salinispora, Amycolatopsis, Saccharomonospora, Kitasatospora, Nocardiopsis and Streptosporangium – all complex sporulating species. Phylogenetic trees of WhiE proteins were broadly congruent with the actinobacterial phylogeny (not shown). Most likely, therefore, the WhiE pathway was present in the last common ancestor of these organisms (node 6 of Fig. 2). Phylogenetic analysis showed that the polyketide synthases for aromatic polyketides such as actinorhodin and tetracyclines did not emerge from within the WhiE lineage (Metsa-Ketela et al., 2002), so the common ancestral synthase presumably pre-dated the evolution of complex actinomycetes.
Conclusion – how development develops during evolution
At the start of this article, we raised several questions that might be addressed in the light of the information reviewed. Here, we return to these questions.
What are the evolutionary origins of genes specifically important for Streptomyces development?
A gene is considered specifically important for development if the relevant mutation has detectable phenotypic effects on development, but not on vegetative growth. Orthologues of many such genes were evidently present in early simple actinobacteria including some of the major developmental regulatory genes (though there are important exceptions). This suggests (in answer to another of the questions) that research focused on Streptomyces developmental biology is likely to provide clues about the cell biology of simpler actinobacteria, including pathogenic organisms, and vice versa. We have tried to summarise some of these reflections across taxa and time in Fig. 10.
Fig. 10.

Acquisition and roles of actinobacteria-specific genes and processes during evolution. It is postulated that a difference in the location of cell wall growth, associated with the use of different central organising proteins (MreB or DivIVA) for peptidoglycan synthesis, was a key element in the early separation of the lines leading to firmicutes and actinobacteria. Polar growth may have been facilitated by the early acquisition of certain conserved cell-biological proteins (FilP, ParJ, CrgA) and proteasomes. Wbl proteins may have sensed the sudden difference in cell physiology consequent on asymmetrical division into a larger rapidly growing daughter cell and a smaller virtually nongrowing one. Entry of primitive Actinobacteria into stationary phase probably involved regulation by orthologues of BldD and BldC (as well as of actinobacteria-non-specific proteins such as WhiA and WhiG that are not shown). Tip growth potentiated the emergence of mycelial growth, which in turn predicated the need for some kind of fragmentation, which takes the form of sporulating aerial mycelium in Streptomyces. The stepwise acquisition of additional functions playing important roles in Streptomyces development is indicated. Proteins with structural or enzymatic roles are shown in coloured boxes, and their locations (or the locations of their products) are shown in corresponding colours. Regulatory proteins are unboxed. Wbl proteins are shown in pale green ovals.
Are the mechanisms leading to sporulation widely homologous in phylogenetically diverse actinobacteria or did they evolve independently?
There is not a simple answer to this, but it does appear that there is a single underlying shared evolutionary pathway, involving both regulatory and cell-biological elements originating from the last common ancestor some 2.6 billion years ago (Battistuzzi et al., 2004), followed by the sequential addition of further elements. Only a few of the latter are specific to streptomycetes and/or their very closest relatives: in one such case, the role of AfsK in controlling DivIVA-mediated branch formation in Streptomyces is likely to be taken by different protein kinases in other mycelial organisms; and in another case, WhiH may provide little more than a modulating influence on the balance of components needed to maximise the efficiency of sporulation in the particular context of Streptomyces aerial hyphae. A very small number of developmental genes have been acquired so recently that they are specific to particular streptomycetes – the whiJ cluster is a good example of this.
Does the developmental process contribute to speciation?
In this review, we have noted two kinds of species-associated diversity that might imply an interplay of development with speciation. One involves the richness and diversity of multiple paralogues of ‘whiJ-like’ clusters, which may imply differences in the sensitivity of development to environmental input (but we also note that the three types of protein encoded in these clusters are all predicted to be cytoplasmically located, so any such sensory role would probably require the independent uptake of soluble small molecules). On the other hand, the second kind of species-associated diversity involves unusually high divergence between orthologous extracellular proteins or proteins with a likely extracellular face. The examples highlighted are Sti-like protease inhibitors, SapB-like aerial growth-facilitating proteins and their biosynthetic enzymes, and the anti-sigma-BldN protein RsbN. These are all candidates for further evaluation as possible agents of speciation, although other explanations such as selection only for broad structural conservation may apply in some cases.
Are today's simple actinobacterial species primitive or are they degenerate descendants of morphologically much more complex ancestors?
It is clear from our analysis that ancestral developmental genes have been lost in some phylogenetic branches (for example, FilP is absent from corynebacteria, and WhiG has been lost many times), showing that evolution does not always proceed in the direction ‘simple to complex’. Nevertheless, the actinobacteria diverging at very early nodes from the line leading to Streptomyces (Fig. 2) do have simple morphology and do lack some important developmental genes.
What gave the ancestral ur-actinobacterium the potential for mycelial growth and aerial sporulation in its more modern descendants?
We have provided some circumstantial evidence and speculative discussion in support of the idea that DivIVA-mediated polar growth could have provided the platform for the evolution of mycelial growth and that actinobacteria-specific features of cell division may have contributed to the specialised processes by which aerial hyphae form chains of spores. However, some innovations, presumably acquired through horizontal gene transfer, were apparently very significant in providing the physical scaffolding for aerial growth (notably the chaplins and SapB-like proteins) and the specialised control of a proteolytic cascade that allows reuse of the substrate mycelium biomass to support aerial growth (Sti-like protease inhibitors), both of which appeared in the actinobacterial lineage at the earliest node giving rise to aerial mycelium.
Can studies of the development of complex actinomycetes assist our understanding of the cell biology of their simpler cousins?
Orthologues of some of the proteins involved in hyphal growth and sporulation of streptomycetes, including several that were previously ‘function unknown’, are found in simple organisms that diverged early in the actinobacterial lineage. The Streptomyces-led breakthroughs in understanding how some such proteins influence hyphal polarity, and others sporulation septation or spore morphogenesis, may provide fertile testing ground for understanding the early evolutionary divergence of proto-actinobacteria from firmicutes and other simple bacteria.
A new question: what is the meaning of disparities in the distribution of developmental genes?
A surprise emerging from this analysis was the occurrence of genes associated with developmental complexity in Kineococcus radiodurans, an apparently simple organism belonging to a phylogenetic group originating from a node that preceded those leading to well-defined complex groups. This raises two perplexing questions, and we have no answers to either of them: what functions do these genes fulfil in a simple coccus? And how is one to account for their phylogenetically inappropriate occurrence?
Concluding comment
This review was stimulated by Gao and Gupta's search for actinobacterial signature proteins. We believe that many aspects of microbial behaviour and physiology can be illuminated by paying close attention both to ancient and less ancient taxon-specific proteins. Reciprocal blastp best-hit analysis such as is shown in Figs 3, 4, 8 and 9 has proved a considerable aid to thinking about the evolution and mechanisms of actinobacterial cell biology and developmental complexity, and access to the full tables at http://streptomyces.org.uk/actinoblast/ may prove useful in investigating other aspects of actinobacterial biology.
References
- Ainsa JA, Parry HD, Chater KF. A response regulator-like protein that functions at an intermediate stage of sporulation in Streptomyces coelicolor A3(2) Mol Microbiol. 1999;34:607–619. doi: 10.1046/j.1365-2958.1999.01630.x. [DOI] [PubMed] [Google Scholar]
- Ainsa JA, Ryding NJ, Hartley N, Findlay KC, Bruton CJ, Chater KF. WhiA, a protein of unknown function conserved among gram-positive bacteria, is essential for sporulation in Streptomyces coelicolor A3(2) J Bacteriol. 2000;182:5470–5478. doi: 10.1128/jb.182.19.5470-5478.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ainsa JA, Bird N, Ryding NJ, Findlay KC, Chater KF. The complex whiJ locus mediates environmentally sensitive repression of development of Streptomyces coelicolor A3(2) Antonie Van Leeuwenhoek. 2010;98:225–236. doi: 10.1007/s10482-010-9443-3. [DOI] [PubMed] [Google Scholar]
- Akanuma G, Hara H, Ohnishi Y, Horinouchi S. Dynamic changes in the extracellular proteome caused by absence of a pleiotropic regulator AdpA in Streptomyces griseus. Mol Microbiol. 2009;73:898–912. doi: 10.1111/j.1365-2958.2009.06814.x. [DOI] [PubMed] [Google Scholar]
- Alam MS, Garg SK, Agrawal P. Studies on structural and functional divergence among seven WhiB proteins of Mycobacterium tuberculosis H37Rv. FEBS J. 2009;276:76–93. doi: 10.1111/j.1742-4658.2008.06755.x. [DOI] [PubMed] [Google Scholar]
- Alam MT, Merlo ME, Takano E, Breitling R. Genome-based phylogenetic analysis of Streptomyces and its relatives. Mol Phylogenet Evol. 2010;54:763–772. doi: 10.1016/j.ympev.2009.11.019. [DOI] [PubMed] [Google Scholar]
- Aldridge BB, Fernandez-Suarez M, Heller D, Ambravaneswaran V, Irimia D, Toner M, Fortune SM. Asymmetry and aging of mycobacterial cells lead to variable growth and antibiotic susceptibility. Science. 2012;335:100–104. doi: 10.1126/science.1216166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ausmees N, Wahlstedt H, Bagchi S, Elliot MA, Buttner MJ, Flardh K. SmeA, a small membrane protein with multiple functions in Streptomyces sporulation including tatgeting of a SpoIIIE/FtsK-like protein to cell division septa. Mol Microbiol. 2007;65:1458–1473. doi: 10.1111/j.1365-2958.2007.05877.x. [DOI] [PubMed] [Google Scholar]
- Averina OA, Zakharevich NV, Danilenko VN. Identification and characterization of WhiB-like family proteins of the Bifidobacterium genus. Anaerobe. 2012;18:421–429. doi: 10.1016/j.anaerobe.2012.04.011. [DOI] [PubMed] [Google Scholar]
- Bagchi S, Tomenius H, Belova LM, Ausmees N. Intermediate filament-like proteins in bacteria and a cytoskeletal function in Streptomyces. Mol Microbiol. 2008;70:1037–1050. doi: 10.1111/j.1365-2958.2008.06473.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barabote RD, Xie G, Leu DH, et al. Complete genome of the cellulolytic thermophile Acidothermus cellulolyticus 11B provides insights into its ecophysiological and evolutionary adaptations. Genome Res. 2009;19:1033–1043. doi: 10.1101/gr.084848.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Battistuzzi FU, Feijao A, Hedges SB. A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land. BMC Evol Biol. 2004;4:44. doi: 10.1186/1471-2148-4-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett JA, Aimino RM, McCormick JR. Streptomyces coelicolor genes ftsL and divIC play a role in cell division but are dispensable for colony formation. J Bacteriol. 2007;189:8982–8992. doi: 10.1128/JB.01303-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bentley SD, Chater KF, Cerdeno-Tarraga AM, et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2) Nature. 2002;417:141–147. doi: 10.1038/417141a. [DOI] [PubMed] [Google Scholar]
- Bentley SD, Brown S, Murphy LD, et al. SCP1, a 356 023 bp linear plasmid adapted to the ecology and developmental biology of its host, Streptomyces coelicolor A3(2) Mol Microbiol. 2004;51:1615–1628. doi: 10.1111/j.1365-2958.2003.03949.x. [DOI] [PubMed] [Google Scholar]
- Bianchetti CM, Blouin GC, Bitto E, Olson JS, Phillips GN., Jr The structure and NO binding properties of the nitrophorin-like hemebinding protein from Arabidopsis thaliana gene locus At1g79260.1. Proteins. 2010;78:917–931. doi: 10.1002/prot.22617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bianchetti CM, Bingman CA, Phillips GN., Jr Structure of the C-terminal heme-binding domain of THAP domain containing protein 4 from Homo sapiens. Proteins. 2011;9:1337–1341. doi: 10.1002/prot.22944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bibb MJ, Molle V, Buttner MJ. Sigma(BldN), an extracytoplasmic function RNA polymerase sigma factor required for aerial mycelium formation in Streptomyces coelicolor A3(2) J Bacteriol. 2000;182:4606–4616. doi: 10.1128/jb.182.16.4606-4616.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bibb MJ, Domonkos A, Chandra G, Buttner MJ. Expression of the chaplin and rodlin hydrophobic sheath proteins in Streptomyces venezuelae is controlled by σ(BldN) and a cognate anti-sigma factor, RsbN. Mol Microbiol. 2012;84:1033–1049. doi: 10.1111/j.1365-2958.2012.08070.x. [DOI] [PubMed] [Google Scholar]
- Bortoluzzi A, Muskett FW, Waters LC, et al. Mycobacterium tuberculosis RNA polymerase binding protein A (RbpA) and its interactions with sigma factors. J Biol Chem. 2013;288:14438–14450. doi: 10.1074/jbc.M113.459883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bramkamp M, van Baarle S. Division site selection in rod-shaped bacteria. Curr Opin Microbiol. 2009;12:683–688. doi: 10.1016/j.mib.2009.10.002. [DOI] [PubMed] [Google Scholar]
- Burian J, Ramón-García S, Sweet G, Gómez-Velasco Av-Gay Y, Thompson CJ. The mycobacterial transcriptional regulator whiB7 links redox homeostasis and intrinsic antibiotic resistance. J Biol Chem. 2012;287:299–310. doi: 10.1074/jbc.M111.302588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Capstick DS, Willey JM, Buttner MJ, Elliot MA. SapB and the chaplins: connections between morphogenetic proteins in Streptomyces coelicolor. Mol Microbiol. 2007;64:602–613. doi: 10.1111/j.1365-2958.2007.05674.x. [DOI] [PubMed] [Google Scholar]
- Capstick DS, Jomaa A, Hanke C, Ortega J, Elliot MA. Dual amyloid domains promote differential functioning of the chaplin proteins during Streptomyces aerial morphogenesis. P Natl Acad Sci USA. 2011;108:9821–9826. doi: 10.1073/pnas.1018715108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champness WC. New loci required for Streptomyces coelicolor morphological and physiological differentiation. J Bacteriol. 1988;170:1168–1174. doi: 10.1128/jb.170.3.1168-1174.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang C, Hatzos C, Morgan T, Bearden J, Joachimiak A. 2010. & Crystal structure of PAC2 family protein from Streptomyces avermitilis MA. MMDB ID: 81922 DOI: 10.2210/pdb3mnf/pdb.
- Chater KF. A morphological and genetic mapping study of white colony mutants of Streptomyces coelicolor. J Gen Microbiol. 1972;72:9–28. doi: 10.1099/00221287-72-1-9. [DOI] [PubMed] [Google Scholar]
- Chater KF. Taking a genetic scalpel to the Streptomyces colony. Microbiology. 1998;144:1465–1478. doi: 10.1099/00221287-144-6-1465. [DOI] [PubMed] [Google Scholar]
- Chater KF. Differentiation in Streptomyces: the properties and programming of diverse cell-types. In: Dyson P, editor. Streptomyces: Molecular Biology and Biotechnology. Norfolk, UK: Caister Academic Press; 2011. pp. 43–86. [Google Scholar]
- Chater KF, Chandra G. The evolution of development in Streptomyces analysed by genome comparisons. FEMS Microbiol Rev. 2006;30:651–672. doi: 10.1111/j.1574-6976.2006.00033.x. [DOI] [PubMed] [Google Scholar]
- Chater KF, Chandra G. The use of the rare UUA codon to define ‘expression space’ for genes involved in secondary metabolism, development and environmental adaptation in Streptomyces. J Microbiol. 2008;46:1–11. doi: 10.1007/s12275-007-0233-1. [DOI] [PubMed] [Google Scholar]
- Chater KF, Biro S, Lee KJ, Palmer T, Schrempf H. The complex extracellular biology of Streptomyces. FEMS Microbiol Rev. 2010;34:171–198. doi: 10.1111/j.1574-6976.2009.00206.x. [DOI] [PubMed] [Google Scholar]
- Chng C, Lum AM, Vroom JA, Kao CM. A key developmental regulator controls the synthesis of the antibiotic erythromycin in Saccharopolyspora erythraea. P Natl Acad Sci USA. 2008;105:11346–11351. doi: 10.1073/pnas.0803622105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi W-W, Park S-D, Lee S-M, Kim H-B, Kim Y, Lee H-S. The whcA gene plays a negative role in oxidative stress response of Corynebacterium glutamicum. FEMS Microbiol Lett. 2009;290:32–38. doi: 10.1111/j.1574-6968.2008.01398.x. [DOI] [PubMed] [Google Scholar]
- Claessen D, Rink R, de Jong W, Siebring J, de Vreugd P, Boersma FG, Dijkhuizen L, Wosten HA. A novel class of secreted hydrophobic proteins is involved in aerial hyphae formation in Streptomyces coelicolor by forming amyloid-like fibrils. Genes Dev. 2003;17:1714–1726. doi: 10.1101/gad.264303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claessen D, Stokroos I, Deelstra HJ, et al. The formation of the rodlet layer of Streptomycetes is the result of the interplay between rodlins and chaplins. Mol Microbiol. 2004;53:433–443. doi: 10.1111/j.1365-2958.2004.04143.x. [DOI] [PubMed] [Google Scholar]
- Claessen D, de Jong W, Dijkhuizen L, Wosten HA. Regulation of Streptomyces development: reach for the sky! Trends Microbiol. 2006;14:313–319. doi: 10.1016/j.tim.2006.05.008. [DOI] [PubMed] [Google Scholar]
- Claret L, Miquel S, Vieille N, Ryjenkov DA, Gomelsky M, Darfeuille-Michaud A. The flagellar sigma factor FliA regulates adhesion and invasion of Crohn disease-associated Escherichia coli via a cyclic dimeric GMP-dependent pathway. J Biol Chem. 2007;282:33275–33283. doi: 10.1074/jbc.M702800200. [DOI] [PubMed] [Google Scholar]
- Cohen-Gonsaud M, Barthe P, Bagneris C, Henderson B, Ward J, Roumestand C, Keep NH. The structure of a resuscitation-promoting factor domain from Mycobacterium tuberculosis shows homology to lysozymes. Nat Struct Mol Biol. 2005;12:270–273. doi: 10.1038/nsmb905. [DOI] [PubMed] [Google Scholar]
- Corker H, Poole RK. Nitric oxide formation by Escherichia coli - Dependence on nitrite reductase, the NO-sensing regulator FNR, and flavohemoglobin Hmp. J Biol Chem. 2003;278:31584–31592. doi: 10.1074/jbc.M303282200. [DOI] [PubMed] [Google Scholar]
- Crack JC, Smith LJ, Stapleton MR, et al. Mechanistic insights into the nitrosylation of the [4Fe-4S] cluster of WhiB-like proteins. J Am Chem Soc. 2011;133:1112–1121. doi: 10.1021/ja109581t. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crack JC, Stapleton MR, Green J, Thomson AJ, Le Brun NE. Mechanism of [4Fe-4S](Cys)4 cluster nitrosylation is conserved amongst NO-responsive regulators. J Biol Chem. 2013;288:11492–11502. doi: 10.1074/jbc.M112.439901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham AF, Spreadbury CL. Mycobacterial stationary phase induced by low oxygen tension: cell wall thickening and localization of the 16-kilodalton alpha- crystallin homolog. J Bacteriol. 1998;180:801–808. doi: 10.1128/jb.180.4.801-808.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis NK, Chater KF. Spore colour in Streptomyces coelicolor A3(2) involves the developmentally regulated synthesis of a compound biosynthetically related to polyketide antibiotics. Mol Microbiol. 1990;4:1679–1691. doi: 10.1111/j.1365-2958.1990.tb00545.x. [DOI] [PubMed] [Google Scholar]
- De Jong W, Wosten HAB, Dijkhuisen L, Claessen D. Attachment of Streptomyces coelicolor is mediated by amyloidal fimbriae that are anchored to the cell surface via cellulose. Mol Microbiol. 2009b;73:1128–1140. doi: 10.1111/j.1365-2958.2009.06838.x. [DOI] [PubMed] [Google Scholar]
- Dedrick RM, Wildschutte H, McCormick JR. Genetic interactions of smc ftsK, and parB genes in Streptomyces coelicolor and their developmental genome segregation phenotypes. J Bacteriol. 2009;191:320–332. doi: 10.1128/JB.00858-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dedrick RM, Marinelli LJ, Newton GL, Pogliano K, Pogliano J, Hatfull GF. Functional requirements for bacteriophage growth: gene essentiality and expression in mycobacteriophage Giles. Mol Microbiol. 2013;88:577–589. doi: 10.1111/mmi.12210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Sol R, Pitman A, Herron P, Dyson P. The product of a developmental gene, crgA, that coordinates reproductive growth in Streptomyces, belongs to a novel family of actinomycete-specific proteins. J Bacteriol. 2003;185:6678–6685. doi: 10.1128/JB.185.22.6678-6685.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Sol R, Mullins JG, Grantcharova N, Flärdh K, Dyson P. Influence of CrgA on assembly of the cell division protein FtsZ during development of Streptomyces coelicolor. J Bacteriol. 2006;188:1540–1550. doi: 10.1128/JB.188.4.1540-1550.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- den Hengst CD, Buttner MJ. Redox control in actinobacteria. Biochim Biophys Acta. 2008;1780:1201–1216. doi: 10.1016/j.bbagen.2008.01.008. [DOI] [PubMed] [Google Scholar]
- den Hengst CD, Tran NT, Bibb MJ, Chandra G, Leskiw BK, Buttner MJ. Genes essential for morphological development and antibiotic production in Streptomyces coelicolor are targets of BldD during vegetative growth. Mol Microbiol. 2010;78:361–379. doi: 10.1111/j.1365-2958.2010.07338.x. [DOI] [PubMed] [Google Scholar]
- Desikan R, Griffiths R, Hancock J, Neill S. A new role for an old enzyme: nitrate reductase-mediated nitric oxide generation is required for abscisic acid-induced stomatal closure in Arabidopsis thaliana. P Natl Acad Sci USA. 2002;99:16314–16318. doi: 10.1073/pnas.252461999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Berardo C, Capstick DS, Bibb MJ, Findlay K, Buttner MJ, Elliot MA. Function and redundancy of the chaplin cell- surface proteins in aerial hyphae formation, rodlet assembly, and viability in Streptomyces coelicolor. J Bacteriol. 2008;190:5879–5889. doi: 10.1128/JB.00685-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ditkowski B, Troc P, Ginda K, Donczew M, Chater KF, Zakrzewska-Czerwinska J, Jakimowicz D. The actinobacterial signature protein ParJ (SCO1662) regulates ParA polymerization and affects chromosome segregation and cell division during Streptomyces sporulation. Mol Microbiol. 2010;78:1403–1415. doi: 10.1111/j.1365-2958.2010.07409.x. [DOI] [PubMed] [Google Scholar]
- Donovan C, Schwaiger A, Kramer R, Bramkamp M. Subcellular localization and characterization of the ParAB system from Corynebacterium glutamicum. J Bacteriol. 2010;192:3441–3451. doi: 10.1128/JB.00214-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donovan C, Sieger B, Kramer R, Bramkamp M. A synthetic Escherichia coli system identifies a conserved origin tethering factor in Actinobacteria. Mol Microbiol. 2012;84:105–116. doi: 10.1111/j.1365-2958.2012.08011.x. [DOI] [PubMed] [Google Scholar]
- Dosanjh M, Newton GL, Davies J. Characterization of a mycothiol ligase mutant of Rhodococcus jostii RHA1. Res Microbiol. 2008;159:643–650. doi: 10.1016/j.resmic.2008.08.006. [DOI] [PubMed] [Google Scholar]
- Elliot M, Talbot NJ. Building filaments in the air: aerial morphogenesis in bacteria and fungi. Curr Opin Microbiol. 2004;7:594–601. doi: 10.1016/j.mib.2004.10.013. [DOI] [PubMed] [Google Scholar]
- Elliot M, Damji F, Passantino R, Chater K, Leskiw B. The bldD gene of Streptomyces coelicolor A3(2): a regulatory gene involved in morphogenesis and antibiotic production. J Bacteriol. 1998;180:1549–1555. doi: 10.1128/jb.180.6.1549-1555.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliot MA, Karoonuthaisiri N, Huang J, Bibb MJ, Cohen SN, Kao CM, Buttner MJ. The chaplins: a family of hydrophobic cell-surface proteins involved in aerial mycelium formation in Streptomyces coelicolor. Genes Dev. 2003;17:1727–1740. doi: 10.1101/gad.264403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Facey PD, Hitchings MD, Saavedra-Garcia P, Fernandez-Martinez L, Dyson PJ, Del Sol R. Streptomyces coelicolor Dps-like proteins: differential dual roles in response to stress during vegetative growth and in nucleoid condensation during reproductive cell division. Mol Microbiol. 2009;73:1186–1202. doi: 10.1111/j.1365-2958.2009.06848.x. [DOI] [PubMed] [Google Scholar]
- Facey PD, Hitchings MD, Williams JS, Skibinski DO, Dyson PJ, Sol RD. The evolution of an osmotically inducible Dps in the genus Streptomyces. PLoS ONE. 2013;8:e60772. doi: 10.1371/journal.pone.0060772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fahey RC. Glutathione analogs in prokaryotes. Biochim Biophys Acta. 2012;1830:3182–3198. doi: 10.1016/j.bbagen.2012.10.006. [DOI] [PubMed] [Google Scholar]
- Felsenstein J. PHYLIP – Phylogeny Inference Package (Version 3.2) Cladistics. 1989;5:164–166. [Google Scholar]
- Felsenstein J. PHYLIP (Phylogeny Inference Package) Version 3.6. Seattle, WA: Department of Genome Sciences, University of Washington; 2005. Distributed by the Author. [Google Scholar]
- Flärdh K. Essential role of DivIVA in polar growth and morphogenesis in Streptomyces coelicolor A3(2) Mol Microbiol. 2003;49:1523–1536. doi: 10.1046/j.1365-2958.2003.03660.x. [DOI] [PubMed] [Google Scholar]
- Flärdh K. Cell polarity and the control of apical growth in Streptomyces. Curr Opin Microbiol. 2010;13:758–765. doi: 10.1016/j.mib.2010.10.002. [DOI] [PubMed] [Google Scholar]
- Flärdh K, Buttner MJ. Streptomyces morphogenetics: dissecting differentiation in a filamentous bacterium. Nat Rev Microbiol. 2009;7:36–49. doi: 10.1038/nrmicro1968. [DOI] [PubMed] [Google Scholar]
- Flärdh K, Findlay KC, Chater KF. Association of early sporulation genes with suggested developmental decision points in Streptomyces coelicolor A3(2) Microbiology. 1999;145:2229–2243. doi: 10.1099/00221287-145-9-2229. [DOI] [PubMed] [Google Scholar]
- Flärdh K, Leibovitz E, Buttner MJ, Chater KF. Generation of a non-sporulating strain of Streptomyces coelicolor A3(2) by the manipulation of a developmentally controlled ftsZ promoter. Mol Microbiol. 2000;38:737–749. doi: 10.1046/j.1365-2958.2000.02177.x. [DOI] [PubMed] [Google Scholar]
- Flärdh K, Richards DM, Hempel AM, Howard M, Buttner MJ. Regulation of apical growth and hyphal branching in Streptomyces. Curr Opin Microbiol. 2012;15:737–743. doi: 10.1016/j.mib.2012.10.012. [DOI] [PubMed] [Google Scholar]
- Fowler-Goldsworthy K, Gust B, Mouz S, Chandra G, Findlay KC, Chater KF. The actinobacteria-specific gene wblA controls major developmental transitions in Streptomyces coelicolor A3(2) Microbiology. 2011;157:1312–1328. doi: 10.1099/mic.0.047555-0. [DOI] [PubMed] [Google Scholar]
- Fuchino K, Bagchi S, Cantlay S, Sandblad L, Wu D, Bergman J, Kamali-Moghaddam M, Flärdh K, Ausmees N. Dynamic gradients of an intermediate filament-like cytoskeleton are recruited by a polarity landmark during apical growth. P Natl Acad Sci USA. 2013;110:E1889–E1897. doi: 10.1073/pnas.1305358110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Funa N, Ohnishi Y, Fujii I, Shibuya M, Ebizuka Y, Horinouchi S. A new pathway for polyketide synthesis in microorganisms. Nature. 1999;400:897–899. doi: 10.1038/23748. [DOI] [PubMed] [Google Scholar]
- Funa N, Funabashi M, Ohnishi Y, Horinouchi S. Biosynthesis of hexahydroxyperylenequinone melanin via oxidative aryl coupling by cytochrome P-450 in Streptomyces griseus. J Bacteriol. 2005;187:8149–8155. doi: 10.1128/JB.187.23.8149-8155.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gadda G, Francis K. Nitronate monooxygenase, a model for anionic flavin semiquinone intermediates in oxidative catalysis. Arch Biochem Biophys. 2010;493:53–61. doi: 10.1016/j.abb.2009.06.018. [DOI] [PubMed] [Google Scholar]
- Gao B, Gupta RS. Phylogenetic framework and molecular signatures for the main clades of the phylum Actinobacteria. Microbiol Mol Biol Rev. 2012;76:66–112. doi: 10.1128/MMBR.05011-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao B, Parmanathan R, Gupta RS. Signature proteins that are distinctive characteristics of Actinobacteria and their subgroups. Antonie Van Leeuwenhoek. 2006;90:69–91. doi: 10.1007/s10482-006-9061-2. [DOI] [PubMed] [Google Scholar]
- Gao B, Sugiman-Marangos S, Junop MS, Gupta RS. Structural and phylogenetic analysis of a conserved actinobacteria-specific protein (ASP1; SCO1997) from Streptomyces coelicolor. BMC Struct Biol. 2009;9:40. doi: 10.1186/1472-6807-9-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gehring A, Nodwell J, Beverley S, Losick R. Genomewide insertional mutagenesis in Streptomyces coelicolor reveals additional genes involved in morphological differentiation. P Natl Acad Sci USA. 2000;97:9642–9647. doi: 10.1073/pnas.170059797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gengenbacher M, Kaufmann SH. Mycobacterium tuberculosis: success through dormancy. FEMS Microbiol Rev. 2012;36:514–532. doi: 10.1111/j.1574-6976.2012.00331.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ginda K, Bezulska M, Ziółkiewicz M, Dziadek J, Zakrzewska-Czerwińska J, Jakimowicz D. ParA of Mycobacterium smegmatis co-ordinates chromosome segregation with the cell cycle and interacts with the polar growth determinant DivIVA. Mol Microbiol. 2013;87:998–1012. doi: 10.1111/mmi.12146. [DOI] [PubMed] [Google Scholar]
- Gomez JE, Bishai WR. whmD is an essential mycobacterial gene required for proper septation and cell division. P Natl Acad Sci USA. 2000;97:8554–8559. doi: 10.1073/pnas.140225297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon BR, Li Y, Wang L, Sintsova A, van Bakel H, Tian S, Navarre WW, Xia B, Liu J. Lsr2 is a nucleoid-associated protein that targets AT-rich sequences and virulence genes in Mycobacterium tuberculosis. P Natl Acad Sci USA. 2010;107:5154–5159. doi: 10.1073/pnas.0913551107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graña M, Bellinzoni M, Miras I, Fiez-Vandal C, Haouz A, Shepard W, Buschiazzo A, Alzari PM. Structure of Mycobacterium tuberculosis Rv2714, a representative of a duplicated gene family in Actinobacteria. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2009;65:972–977. doi: 10.1107/S1744309109035027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grantcharova N, Ubhayasekera W, Mowbray SL, McCormick JR, Flärdh K. A missense mutation in ftsZ differentially affects vegetative and developmentally controlled cell division in Streptomyces coelicolor A3(2) Mol Microbiol. 2003;47:645–656. doi: 10.1046/j.1365-2958.2003.03334.x. [DOI] [PubMed] [Google Scholar]
- Gupta RS, Chen WJ, Adeolu M, Chai Y. Molecular signatures for the class Coriobacteriia and its different clades; Proposal for division of the class Coriobacteriia into the emended order Coriobacteriales, containing the emended family Coriobacteriaceae and Atopobiaceae fam. nov., and Eggerthellales ord. nov., containing the family Eggerthellaceae fam. nov. Int J Syst Evol Microbiol. 2013;63:3379–3397. doi: 10.1099/ijs.0.048371-0. [DOI] [PubMed] [Google Scholar]
- Hagen KD, Meeks JC. The unique cyanobacterial protein OpcA is an allosteric effector of glucose-6-phosphate dehydrogenase in Nostoc punctiforme ATCC 29133. J Biol Chem. 2001;276:11477–11486. doi: 10.1074/jbc.M010472200. [DOI] [PubMed] [Google Scholar]
- Haiser HJ, Yousef MR, Elliot MA. Cell wall hydrolases affect germination, vegetative growth, and sporulation in Streptomyces coelicolor. J Bacteriol. 2009;191:6501–6512. doi: 10.1128/JB.00767-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heichlinger A, Ammelburg M, Kleinschnitz EM, Latus A, Maldener I, Flärdh K, Wohlleben W, Muth G. The MreB-like protein Mbl of S. coelicolor A3(2) depends on MreB for proper localization and contributes to spore wall synthesis. J Bacteriol. 2011;193:1533–1542. doi: 10.1128/JB.01100-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helmann JD. Alternative sigma factors and the regulation of flagellar gene expression. Mol Microbiol. 1991;5:2875–2882. doi: 10.1111/j.1365-2958.1991.tb01847.x. [DOI] [PubMed] [Google Scholar]
- Hempel AM, Wang SB, Letek M, Gil JA, Flardh K. Assemblies of DivIVA mark sites for hyphal branching and can establish new zones of cell wall growth in Streptomyces coelicolor. J Bacteriol. 2008;190:7579–7583. doi: 10.1128/JB.00839-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hempel AM, Cantlay S, Molle V, Wang SB, Naldrett MJ, Parker JL, Richards DM, Jung YG, Buttner MJ, Flärdh K. The Ser/Thr protein kinase AfsK regulates polar growth and hyphal branching in the filamentous bacteria Streptomyces. P Natl Acad Sci USA. 2012;109:E2371–E2379. doi: 10.1073/pnas.1207409109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hett EC, Chao MC, Steyn AJ, Fortune SM, Deng LL, Rubin EJ. A partner for the resuscitation-promoting factors of Mycobacterium tuberculosis. Mol Microbiol. 2007;66:658–668. doi: 10.1111/j.1365-2958.2007.05945.x. [DOI] [PubMed] [Google Scholar]
- Hett EC, Chao MC, Deng LL, Rubin EJ. A mycobacterial enzyme essential for cell division synergizes with resuscitation-promoting factor. PLoS Pathog. 2008;4:e1000001. doi: 10.1371/journal.ppat.1000001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hett EC, Chao MC, Rubin EJ. Interaction and modulation of two antagonistic cell wall enzymes of mycobacteria. PLoS Pathog. 2010;6:e1001020. doi: 10.1371/journal.ppat.1001020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higo A, Horinouchi S, Ohnishi Y. Strict regulation of morphological differentiation and secondary metabolism by a positive feedback loop between two global regulators AdpA and BldA in Streptomyces griseus. Mol Microbiol. 2011;81:1607–1622. doi: 10.1111/j.1365-2958.2011.07795.x. [DOI] [PubMed] [Google Scholar]
- Higo A, Hara H, Horinouchi S, Ohnishi Y. Genome-wide distribution of AdpA, a global regulator for secondary metabolism and morphological differentiation in Streptomyces, revealed the extent and complexity of the AdpA regulatory network. DNA Res. 2012;19:259–273. doi: 10.1093/dnares/dss010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes NA, Walshaw J, Leggett RM, Thibessard A, Dalton KA, Gillespie MD, Hemmings AM, Gust B, Kelemen GH. Coiled-coil protein Scy is a key component of a multiprotein assembly controlling polarized growth in Streptomyces. P Natl Acad Sci USA. 2013;110:E397–E406. doi: 10.1073/pnas.1210657110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hopwood DA. Streptomyces in Nature and Medicine: The Antibiotic Makers. New York, NY: Oxford University Press; 2007. [Google Scholar]
- Hopwood DA, Wildermuth H, Palmer HM. Mutants of Streptomyces coelicolor defective in sporulation. J Gen Microbiol. 1970;61:397–408. doi: 10.1099/00221287-61-3-397. [DOI] [PubMed] [Google Scholar]
- Horinouchi S. A microbial hormone, A-factor, as a master switch for morphological differentiation and secondary metabolism in Streptomyces griseus. Front Biosci. 2002;7:D2045–D2057. doi: 10.2741/A897. [DOI] [PubMed] [Google Scholar]
- Horinouchi S. Mining and polishing of the treasure trove in the bacterial genus Streptomyces. Biosci Biotechnol Biochem. 2007;71:283–299. doi: 10.1271/bbb.60627. [DOI] [PubMed] [Google Scholar]
- Hunt AC, Servin-Gonzalez L, Kelemen GH, Buttner MJ. The bldC developmental locus of Streptomyces coelicolor encodes a member of a family of small DNA-binding proteins related to the DNA-binding domains of the MerR family. J Bacteriol. 2005;187:716–728. doi: 10.1128/JB.187.2.716-728.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchings MI. Unusual two-component signal transduction pathways in the Actinobacteria. Adv Appl Microbiol. 2007;61:1–26. doi: 10.1016/S0065-2164(06)61001-0. [DOI] [PubMed] [Google Scholar]
- Jakimowicz D, van Wezel GP. Cell division and DNA segregation in Streptomyces: how to build a septum in the middle of nowhere? Mol Microbiol. 2012;85:393–404. doi: 10.1111/j.1365-2958.2012.08107.x. [DOI] [PubMed] [Google Scholar]
- Jakimowicz D, Chater K, Zakrzewska-Czerwinska J. The ParB protein of Streptomyces coelicolor A3(2) recognizes a cluster of parS sequences within the origin-proximal region of the linear chromosome. Mol Microbiol. 2002;45:1365–1377. doi: 10.1046/j.1365-2958.2002.03102.x. [DOI] [PubMed] [Google Scholar]
- Jakimowicz D, Gust B, Zakrzewska-Czerwinska J, Chater KF. Developmental-stage-specific assembly of ParB complexes in Streptomyces coelicolor hyphae. J Bacteriol. 2005a;187:3572–3580. doi: 10.1128/JB.187.10.3572-3580.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakimowicz P, Cheesman MR, Bishai WR, Chater KF, Thomson AJ, Buttner MJ. Evidence that the Streptomyces developmental protein WhiD, a member of the WhiB family, binds a 4Fe-4S cluster. J Biol Chem. 2005b;280:8309–8315. doi: 10.1074/jbc.M412622200. [DOI] [PubMed] [Google Scholar]
- Jakimowicz D, Mouz S, Zakrzewska-Czerwinska Z, Chater KF. Regulation of developmentally controlled parAB promoter leading to formation of ParB complexes in Streptomyces coelicolor aerial hyphae. J Bacteriol. 2006;188:1710–1720. doi: 10.1128/JB.188.5.1710-1720.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakimowicz D, Zydek P, Kois A, Zakrzewska-Czerwińska J, Chater KF. Alignment of multiple chromosomes along helical ParA scaffolding in sporulating Streptomyces hyphae. Mol Microbiol. 2007;65:625–641. doi: 10.1111/j.1365-2958.2007.05815.x. [DOI] [PubMed] [Google Scholar]
- Jani C, Eoh H, Lee JJ, et al. Regulation of polar peptidoglycan biosynthesis by Wag31 phosphorylation in mycobacteria. BMC Microbiol. 2010;10:327. doi: 10.1186/1471-2180-10-327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaiser BK, Stoddard BL. DNA recognition and transcriptional regulation by the WhiA sporulation factor. Sci Rep. 2011;1:156. doi: 10.1038/srep00156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaiser BK, Clifton MC, Shen BW, Stoddard BL. The structure of a bacterial DUF199/WhiA protein: domestication of an invasive endonuclease. Structure. 2009;17:1368–1376. doi: 10.1016/j.str.2009.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kana BD, Mizrahi V. Resuscitation-promoting factors as lytic enzymes for bacterial growth and signaling. FEMS Immunol Med Microbiol. 2010;58:39–50. doi: 10.1111/j.1574-695X.2009.00606.x. [DOI] [PubMed] [Google Scholar]
- Kang CM, Nyayapathy S, Lee JY, Suh JW, Husson RN. Wag31, a homologue of the cell division protein DivIVA, regulates growth, morphology and polar cell wall synthesis in mycobacteria. Microbiology. 2008;154:725–735. doi: 10.1099/mic.0.2007/014076-0. (Erratum in: Microbiology 2008 154:2168.) [DOI] [PubMed] [Google Scholar]
- Kato JY, Ohnishi Y, Horinouchi S. Autorepression of AdpA of the AraC/XylS family, a key transcriptional activator in the A-factor regulatory cascade in Streptomyces griseus. J Mol Biol. 2005;350:12–26. doi: 10.1016/j.jmb.2005.04.058. [DOI] [PubMed] [Google Scholar]
- Kim HJ, Calcutt MJ, Scmidt FJ, Chater KF. Partitioning of the linear chromosome during sporulation of Streptomyces coelicolor A3(2) involves an oriC-linked parAB locus. J Bacteriol. 2000;182:1313–1320. doi: 10.1128/jb.182.5.1313-1320.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim T-H, Park J-S, Kim H-J, Kim Y, Kim P, Lee H-S. The whcE gene of Corynebacterium glutamicum is important for survival following heat and oxidative stress. Biochem Biophys Res Commun. 2005;337:757–764. doi: 10.1016/j.bbrc.2005.09.115. [DOI] [PubMed] [Google Scholar]
- Kim DW, Hesketh A, Kim ES, Song JY, Lee DH, Kim IS, Chater KF, Lee KJ. Complex extracellular interactions of proteases and a protease inhibitor influence multicellular development of Streptomyces coelicolor. Mol Microbiol. 2008a;70:1180–1193. doi: 10.1111/j.1365-2958.2008.06471.x. [DOI] [PubMed] [Google Scholar]
- Kim ES, Song JY, Kim DW, Chater KF, Lee KJ. A possible extended family of regulators of sigma factor activity in Streptomyces coelicolor. J Bacteriol. 2008b;190:7559–7566. doi: 10.1128/JB.00470-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim MS, Dufour YS, Yoo JS, Cho YB, Park JH, Nam GB, Kim HM, Lee KL, Donohue TJ, Roe JH. Conservation of thiol-oxidative stress responses regulated by SigR orthologues in actinomycetes. Mol Microbiol. 2012;85:326–344. doi: 10.1111/j.1365-2958.2012.08115.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleinschnitz EM, Heichlinger A, Schirner K, Winkler J, Latus A, Maldener I, Wohlleben W, Muth G. Proteins encoded by the mre gene cluster in Streptomyces coelicolor A3(2) cooperate in spore wall synthesis. Mol Microbiol. 2011;79:1367–1379. doi: 10.1111/j.1365-2958.2010.07529.x. [DOI] [PubMed] [Google Scholar]
- Knizewski L, Ginalski K. Bacterial DUF199/COG1481 proteins including sporulation regulator WhiA are distant homologs of LAGLIDADG homing endonucleases that retained only DNA binding. Cell Cycle. 2007;6:1666–1670. doi: 10.4161/cc.6.13.4471. [DOI] [PubMed] [Google Scholar]
- Kodani S, Hudson E, Durrant MC, Buttner MJ, Nodwell JR, Willey JM. The SapB morphogen is a lantibiotic-like peptide derived from the product of the developmental gene ramS in Streptomyces coelicolor. P Natl Acad Sci USA. 2004;101:11448–11453. doi: 10.1073/pnas.0404220101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kodani S, Lodato MA, Durrant MC, Picart F, Willey JM. SapT, a lanthionine-containing peptide involved in aerial hyphae formation in the Streptomycetes. Mol Microbiol. 2005;58:1368–1380. doi: 10.1111/j.1365-2958.2005.04921.x. [DOI] [PubMed] [Google Scholar]
- Kois A, Swiatek M, Jakimowicz D, Zakrzewska-Czerwinska J. SMC protein-dependent chromosome condensation during aerial hyphal development in Streptomyces. J Bacteriol. 2009;191:310–319. doi: 10.1128/JB.00513-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamont EA, Bannantine JP, Armién A, Ariyakumar DS, Sreevatsan S. Identification and characterization of a spore-like morphotype in chronically starved Mycobacterium avium subsp. paratuberculosis cultures. PLoS ONE. 2012;7:e30648. doi: 10.1371/journal.pone.0030648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawlor EJ, Baylis HA, Chater KF. Pleiotropic morphological and antibiotic deficiencies result from mutations in a gene encoding a tRNA-like product in Streptomyces coelicolor A3(2) Genes Dev. 1987;1:1305–1310. doi: 10.1101/gad.1.10.1305. [DOI] [PubMed] [Google Scholar]
- Lee BH, Murugasu-Oei B, Dick T. Upregulation of a histonelike protein in dormant Mycobacterium smegmatis. Mol Gen Genet. 1998;260:475–479. doi: 10.1007/s004380050919. [DOI] [PubMed] [Google Scholar]
- Letek M, Ordóñez E, Vaquera J, Margolin W, Flärdh K, Mateos LM, Gil JA. DivIVA is required for polar growth in the MreB-lacking rod-shaped actinomycete Corynebacterium glutamicum. J Bacteriol. 2008;190:3283–3292. doi: 10.1128/JB.01934-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Letek M, Fiuza M, Villadangos AF, Mateos LM, Gil JA. Cytoskeletal proteins of actinobacteria. Int J Cell Biol2012. 2012 doi: 10.1155/2012/905832. Article ID 905832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li W, Wu J, Tao W, et al. A genetic and bioinformatic analysis of Streptomyces coelicolor genes containing TTA codons, possible targets for regulation by a developmentally significant tRNA. FEMS Microbiol Lett. 2007;266:20–28. doi: 10.1111/j.1574-6968.2006.00494.x. [DOI] [PubMed] [Google Scholar]
- Liu G, Chater KF, Chandra G, Niu G, Tan H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol Mol Biol Rev. 2013a;77:112–143. doi: 10.1128/MMBR.00054-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu YB, Long MX, Yin YJ, Si MR, Zhang L, Lu ZQ, Wang Y, Shen XH. Physiological roles of mycothiol in detoxification and tolerance to multiple poisonous chemicals in Corynebacterium glutamicum. Arch Microbiol. 2013b;195:419–429. doi: 10.1007/s00203-013-0889-3. [DOI] [PubMed] [Google Scholar]
- Luo Y, Helmann JD. A σD-dependent antisense transcript modulates expression of the cyclic-di-AMP hydrolase GdpP in Bacillus subtilis. Microbiology. 2012;158:2732–2741. doi: 10.1099/mic.0.062174-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madej T, Addess KJ, Fong JH, et al. MMDB: 3D structures and macromolecular interactions. Nucleic Acids Res. 2012;40:D461–D464. doi: 10.1093/nar/gkr1162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manteca A, Claessen D, Lopez-Iglesias C, Sanchez J. Aerial hyphae in surface cultures of Streptomyces lividans and Streptomyces coelicolor originate from viable segments surviving an early programmed cell death event. FEMS Microbiol Lett. 2007;274:118–125. doi: 10.1111/j.1574-6968.2007.00825.x. [DOI] [PubMed] [Google Scholar]
- McCormick JR. Cell division is dispensable but not irrelevant in Streptomyces. Curr Opin Microbiol. 2009;12:689–698. doi: 10.1016/j.mib.2009.10.004. [DOI] [PubMed] [Google Scholar]
- McCormick JR, Flärdh K. Signals and regulators that govern Streptomyces development. FEMS Microbiol Rev. 2012;36:206–231. doi: 10.1111/j.1574-6976.2011.00317.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCormick JR, Su EP, Driks A, Losick R. Growth and viability of Streptomyces coelicolor mutant for the celldivision gene ftsZ. Mol Microbiol. 1994;14:243–254. doi: 10.1111/j.1365-2958.1994.tb01285.x. [DOI] [PubMed] [Google Scholar]
- McVittie A. Ultrastructural studies on wild-type and white colony mutants of Streptomyces coelicolor. J Gen Microbiol. 1974;81:291–302. doi: 10.1099/00221287-81-2-291. [DOI] [PubMed] [Google Scholar]
- Merrick M. A morphological and genetic mapping study of bald colony mutants of Streptomyces coelicolor. J Gen Microbiol. 1976;96:299–315. doi: 10.1099/00221287-96-2-299. [DOI] [PubMed] [Google Scholar]
- Metsa-Ketela M, Halo L, Munukka E, Mantsala P, Ylihonko K. Molecular evolution of aromatic polyketides and comparative sequence analysis of polyketide ketosynthase and 16S ribosomal DNA genes from various Streptomyces species. Appl Environ Microbiol. 2002;68:4472–4479. doi: 10.1128/AEM.68.9.4472-4479.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyadoh S. Atlas of Actinomycetes. Japan, Tokyo, Japan: The Society for Actinomycetes; 1997. [Google Scholar]
- Molle V, Buttner MJ. Different alleles of the response regulator gene bldM arrest Streptomyces coelicolor development at distinct stages. Mol Microbiol. 2000;36:1265–1278. doi: 10.1046/j.1365-2958.2000.01977.x. [DOI] [PubMed] [Google Scholar]
- Molle V, Palframan WJ, Findlay KC, Buttner MJ. WhiD and WhiB, homologous proteins required for different stages of sporulation in Streptomyces coelicolor A3(2) J Bacteriol. 2000;182:1286–1295. doi: 10.1128/jb.182.5.1286-1295.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris R, Nguyen L, Gatfield J, et al. Ancestral antibiotic resistance in Mycobacterium tuberculosis. P Natl Acad Sci USA. 2005;102:12200–12205. doi: 10.1073/pnas.0505446102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mukamolova GV, Kaprelyants AS, Young DI, Young M, Kell DB. A bacterial cytokine. P Natl Acad Sci USA. 1998;95:8916–8921. doi: 10.1073/pnas.95.15.8916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mukamolova GV, Murzin AG, Salina EG, Demina GR, Kell DB, Kaprelyants AS, Young M. Muralytic activity of Micrococcus luteus Rpf and its relationship to physiological activity in promoting bacterial growth and resuscitation. Mol Microbiol. 2006;59:84–98. doi: 10.1111/j.1365-2958.2005.04930.x. [DOI] [PubMed] [Google Scholar]
- Narula J, Devi SN, Fujita M, Igoshin OA. Ultrasensitivity of the Bacillus subtilis sporulation decision. P Natl Acad Sci USA. 2012;109:E3513–E3522. doi: 10.1073/pnas.1213974109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen KT, Tenor J, Stettler H, Nguyen LT, Nguyen LD, Thompson CJ. Colonial differentiation in Streptomyces coelicolor depends on translation of a specific codon within the adpA gene. J Bacteriol. 2003;185:7291–7296. doi: 10.1128/JB.185.24.7291-7296.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noens EE, Mersinias V, Traag BA, Smith CP, Koerten HK, vanWezel GP. SsgA-like proteins determine the fate of peptidoglycan during sporulation of Streptomyces coelicolor. Mol Microbiol. 2005;58:929–944. doi: 10.1111/j.1365-2958.2005.04883.x. [DOI] [PubMed] [Google Scholar]
- Noens EE, Mersinias V, Willemse J, Traag BA, Laing E, Chater KF, Smith CP, Koerten HK, van Wezel GP. Loss of the controlled localization of growth stage-specific cell-wall synthesis pleiotropically affects developmental gene expression in an ssgA mutant of Streptomyces coelicolor. Mol Microbiol. 2007;64:1244–1259. doi: 10.1111/j.1365-2958.2007.05732.x. [DOI] [PubMed] [Google Scholar]
- Ohnishi Y, Ishikawa J, Hara H, Suzuki H, Ikenoya M, Ikeda H, Yamashita A, Hattori M, Horinouchi S. Genome sequence of the streptomycin-producing microorganism Streptomyces griseus IFO 13350. J Bacteriol. 2008;190:4050–4060. doi: 10.1128/JB.00204-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paget MSB, Molle V, Cohen G, Aharonowitz Y, Buttner MJ. Defining the disulphide stress response in Streptomyces coelicolor A3(2): identification of the σR regulon. Mol Microbiol. 2001;42:1007–1020. doi: 10.1046/j.1365-2958.2001.02675.x. [DOI] [PubMed] [Google Scholar]
- Park JS, Shin S, Kim ES, Kim P, Kim Y, Lee HS. Identification of SpiA that interacts with Corynebacterium glutamicum WhcA using a two-hybrid system. FEMS Microbiol Lett. 2011;322:8–14. doi: 10.1111/j.1574-6968.2011.02318.x. [DOI] [PubMed] [Google Scholar]
- Pedulla ML, Hatfull GF. Characterization of the mIHF gene of Mycobacterium smegmatis. J Bacteriol. 1998;180:5473–5477. doi: 10.1128/jb.180.20.5473-5477.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penn K, Jensen PR. Comparative genomics reveals evidence of marine adaptation in Salinispora species. BMC Genomics. 2012;13:86. doi: 10.1186/1471-2164-13-86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Persson J, Chater KF, Flärdh K. Molecular and cytological analysis of the expression of Streptomyces sporulation regulatory gene whiH. FEMS Microbiol Lett. 2013;341:96–105. doi: 10.1111/1574-6968.12099. [DOI] [PubMed] [Google Scholar]
- Piggot PJ, Hilbert DW. Sporulation of Bacillus subtilis. Curr Opin Microbiol. 2004;7:579–586. doi: 10.1016/j.mib.2004.10.001. [DOI] [PubMed] [Google Scholar]
- Piuri M, Hatfull GF. A peptidoglycan hydrolase motif within the mycobacteriophage TM4 tape measure protein promotes efficient infection of stationary phase cells. Mol Microbiol. 2006;62:1569–1585. doi: 10.1111/j.1365-2958.2006.05473.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plocinski P, Ziolkiewicz M, Kiran M, et al. Characterization of CrgA, a new partner of the Mycobacterium tuberculosis peptidoglycan polymerization complexes. J Bacteriol. 2011;193:3246–3256. doi: 10.1128/JB.00188-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plocinski P, Arora N, Sarva K, et al. Mycobacterium tuberculosis CwsA Interacts with CrgA and Wag31, and the CrgA-CwsA complex is involved in peptidoglycan synthesis and cell shape determination. J Bacteriol. 2012;194:6398–6409. doi: 10.1128/JB.01005-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price CW. Protective function and regulation of the general stress response in Bacillus subtilis and related bacteria. In: Hengge-Aronis R, Storz G, editors. Bacterial Stress responses. Washington, DC: American Society for Microbiology; 2000. pp. 179–197. [Google Scholar]
- Ravagnani A, Finan CL, Young M. A novel firmicute protein family related to the actinobacterial resuscitation-promoting factors by non-orthologous domain displacement. BMC Genomics. 2005;6:39. doi: 10.1186/1471-2164-6-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawat M, Av-Gay Y. Mycothiol-dependent proteins in actinomycetes. FEMS Microbiol Rev. 2007;31:278–292. doi: 10.1111/j.1574-6976.2006.00062.x. [DOI] [PubMed] [Google Scholar]
- Rawat M, Newton GL, Ko M, Martinez GJ, Fahey RC, Av-Gay Y. Mycothiol-deficient Mycobacterium smegmatis mutants are hypersensitive to alkylating agents, free radicals and antibiotics. Antimicrob Agents Chemother. 2002;46:3348–3355. doi: 10.1128/AAC.46.11.3348-3355.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reyes-Lamothe R, Nicolas E, Sherratt D. Chromosome replication and segregation in bacteria. Annu Rev Genet. 2012;46:121–143. doi: 10.1146/annurev-genet-110711-155421. [DOI] [PubMed] [Google Scholar]
- Ruggiero A, Tizzano B, Pedone E, Pedone C, Wilmanns M, Berisio R. Crystal structure of the resuscitation-promoting factor (DeltaDUF)RpfB from M. tuberculosis. J Mol Biol. 2009;385:153–162. doi: 10.1016/j.jmb.2008.10.042. [DOI] [PubMed] [Google Scholar]
- Rybniker J, Nowag A, van Gumpel E, Nissen N, Robinson N, Plum G, Hartmann P. Insights into the function of the WhiB-like protein of mycobacteriophage TM4–a transcriptional inhibitor of WhiB2. Mol Microbiol. 2010;77:642–657. doi: 10.1111/j.1365-2958.2010.07235.x. [DOI] [PubMed] [Google Scholar]
- Ryding NJ, Kelemen GH, Whatling CA, Flardh K, Buttner MJ, Chater KF. A developmentally regulated gene encoding a repressor-like protein is essential for sporulation in Streptomyces coelicolor A3(2) Mol Microbiol. 1998;29:343–357. doi: 10.1046/j.1365-2958.1998.00939.x. [DOI] [PubMed] [Google Scholar]
- Saini V, Farhana A, Steyn AJC. Mycobacterium tuberculosis WhiB3: a novel iron-sulfur cluster protein that regulates redox homeostasis and virulence. Antioxid Redox Signal. 2011;16:687–697. doi: 10.1089/ars.2011.4341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saini V, Farhana A, Steyn AJC. Iron sulfur cluster proteins and microbial regulation: implications for understanding tuberculosis. Curr Opin Chem Biol. 2012;16:45–53. doi: 10.1016/j.cbpa.2012.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salerno P, Larsson J, Bucca G, Laing E, Smith CP, Flärdh K. One of the two genes encoding nucleoid-associated HU proteins in Streptomyces coelicolor is developmentally regulated and specifically involved in spore maturation. J Bacteriol. 2009;191:6489–6500. doi: 10.1128/JB.00709-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarafeddinov A, Arif A, Peters A, Fuchsbauer HL. A novel transglutaminase substrate from Streptomyces mobaraensis inhibiting papain-like cysteine proteases. J Microbiol Biotechnol. 2011;21:617–626. [PubMed] [Google Scholar]
- Sassetti CM, Boyd DH, Rubin EJ. Genes required for mycobacterial growth defined by high density mutagenesis. Mol Microbiol. 2003;48:77–84. doi: 10.1046/j.1365-2958.2003.03425.x. [DOI] [PubMed] [Google Scholar]
- Schmidt S, Adolf F, Fuchsbauer HL. The transglutaminase activating metalloprotease inhibitor from Streptomyces mobaraensis is a glutamine and lysine donor substrate of the intrinsic transglutaminase. FEBS Lett. 2008;582:3132–3138. doi: 10.1016/j.febslet.2008.07.049. [DOI] [PubMed] [Google Scholar]
- Scholefield G, Whiting R, Errington J, Murray H. Spo0J regulates the oligomeric state of Soj to trigger its switch from an activator to an inhibitor of DNA replication initiation. Mol Microbiol. 2011;79:1089–1100. doi: 10.1111/j.1365-2958.2010.07507.x. [DOI] [PubMed] [Google Scholar]
- Seipke RF, Kaltenpoth M, Hutchings MI. Streptomyces as symbionts: an emerging and widespread theme? FEMS Microbiol Rev. 2012;36:862–876. doi: 10.1111/j.1574-6976.2011.00313.x. [DOI] [PubMed] [Google Scholar]
- Sevcikova B, Rezuchova B, Homerova D, Kormanec J. The anti-anti sigma factor BldG is involved in activation of stress-response sigma factor σH in Streptomyces coelicolor A3(2) J Bacteriol. 2010;192:5674–5681. doi: 10.1128/JB.00828-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepard W, Haouz A, Grana M, Buschiazzo A, Betton JM, Cole ST, Alzari PM. The crystal structure of Rv0813c from Mycobacterium tuberculosis reveals a new family of fatty acid-binding protein-like proteins in bacteria. J Bacteriol. 2007;189:1899–1904. doi: 10.1128/JB.01435-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh A, Guidry L, Narasimhulu KV, Mai D, Trombley J, Redding KE, Giles GI, Lancaster JR, Jr, Steyn AJ. Mycobacterium tuberculosis WhiB3 responds to O2 and nitric oxide via its [4Fe–4S] cluster and is essential for nutrient starvation survival. P Natl Acad Sci USA. 2007;104:11562–11567. doi: 10.1073/pnas.0700490104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh B, Nitharwal RG, Ramesh M, Pettersson BM, Kirsebom LA, Dasgupta S. Asymmetric growth and division in Mycobacterium spp.: Compensatory mechanisms for non-medial septa. Mol Microbiol. 2013;88:64–76. doi: 10.1111/mmi.12169. [DOI] [PubMed] [Google Scholar]
- Smith LJ, Stapleton MR, Fullstone GJ, et al. Mycobacterium tuberculosis WhiB1 is an essential DNA-binding protein with a nitric oxide-sensitive iron sulfur cluster. Biochem J. 2010;432:417–427. doi: 10.1042/BJ20101440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soliveri J, Vijgenboom E, Granozzi C, Plaskitt KA, Chater KF. Functional and evolutionary implications of a survey of various actinomycetes for homologues of two Streptomyces coelicolor sporulation genes. J Gen Microbiol. 1993;139:2569–2578. doi: 10.1099/00221287-139-11-2569. [DOI] [PubMed] [Google Scholar]
- Soliveri JA, Gomez J, Bishai WR, Chater KF. Multiple paralogous genes related to the Streptomyces coelicolor developmental regulatory gene whiB are present in Streptomyces and other actinomycetes. Microbiology. 2000;146:333–343. doi: 10.1099/00221287-146-2-333. [DOI] [PubMed] [Google Scholar]
- Stapleton MR, Smith LJ, Hunt DM, Buxton RS, Green J. Mycobacterium tuberculosis WhiB1 represses transcription of the essential chaperonin GroEL2. Tuberculosis. 2012;92:328–332. doi: 10.1016/j.tube.2012.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staron A, Sofia HJ, Dietrich S, Ulrich LE, Liesegang H, Mascher T. The third pillar of bacterial signal transduction: classification of the extracytoplasmic function (ECF) sigma factor protein family. Mol Microbiol. 2009;74:557–558. doi: 10.1111/j.1365-2958.2009.06870.x. [DOI] [PubMed] [Google Scholar]
- Strahl H, Hamoen LW. Finding the corners in a cell. Curr Opin Microbiol. 2012;15:731–736. doi: 10.1016/j.mib.2012.10.006. [DOI] [PubMed] [Google Scholar]
- Striebel F, Imkamp F, Ozcelik D, Weber-Ban E. Pupylation as a signal for proteasomal degradation in bacteria. Biochim Biophys Acta. 2013 doi: 10.1016/j.bbamcr.2013.03.022. pii: S0167-4889(13)00123-7. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Swiercz JP, Nanji T, Gloyd M, Guarné A, Elliot MA. A novel nucleoid-associated protein specific to the actinobacteria. Nucleic Acids Res. 2013;41:4171–4184. doi: 10.1093/nar/gkt095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabib-Salazar A, Liu B, Doughty P, Lewis RA, Ghosh S, Parsy ML, Simpson PJ, O'Dwyer K, Matthews SJ, Paget MS. The actinobacterial transcription factor RbpA binds to the principal sigma subunit of RNA polymerase. Nucleic Acids Res. 2013;41:5679–5691. doi: 10.1093/nar/gkt277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taguchi S, Nishihama K, Igi K, Ito K, Taira H, Motoki M, Momose H. Substrate specificity analysis of microbial transglutaminase using proteinaceous protease inhibitors as natural model substrates. J Biochem. 2000;128:415–425. doi: 10.1093/oxfordjournals.jbchem.a022769. [DOI] [PubMed] [Google Scholar]
- Takano E, Tao M, Long F, Bibb MJ, Wang L, Li W, Buttner MJ, Bibb MJ, Deng ZX, Chater KF. A rare leucine codon in adpA is implicated in the morphological defect of bldA mutants of Streptomyces coelicolor. Mol Microbiol. 2003;50:475–486. doi: 10.1046/j.1365-2958.2003.03728.x. [DOI] [PubMed] [Google Scholar]
- Takano H, Fujimoto M, Urano H, Beppu T, Ueda K. Cross-interaction of anti-σH factor RshA with BldG, an anti-sigma factor antagonist in Streptomyces griseus. FEMS Microbiol Lett. 2011;314:158–163. doi: 10.1111/j.1574-6968.2010.02155.x. [DOI] [PubMed] [Google Scholar]
- Tian Y, Fowler K, Findlay K, Tan H, Chater KF. An unusual response regulator influences sporulation at early and late stages in Streptomyces coelicolor. J Bacteriol. 2007;189:2873–2885. doi: 10.1128/JB.01615-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toro E, Shapiro L. Bacterial chromosome organization and segregation. Cold Spring Harb Perspect Biol. 2010;2:a000349. doi: 10.1101/cshperspect.a000349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Touhami A, Jericho MH, Beveridge TJ. Atomic force microscopy of cell growth and division in Staphylococcus aureus. J Bacteriol. 2004;186:3286–3295. doi: 10.1128/JB.186.11.3286-3295.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ueda K, Oinuma K, Ikeda G, Hosono K, Ohnishi Y, Horinouchi S, Beppu T. AmfS, an extracellular peptidic morphogen in Streptomyces griseus. J Bacteriol. 2002;184:1488–1492. doi: 10.1128/JB.184.5.1488-1492.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ventura M, Canchaya C, Tauch A, Chandra G, Fitzgerald GF, Chater KF, Van Sinderen D. Genomics of Actinobacteria: tracing the evolutionary history of an ancient phylum. Microbiol Mol Biol Rev. 2007;71:495–548. doi: 10.1128/MMBR.00005-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vine CE, Purewal SK, Cole JA. NsrR-dependent method for detecting nitric oxide accumulation in the Escherichia coli cytoplasm and enzymes involved in NO production. FEMS Microbiol Lett. 2011;325:108–114. doi: 10.1111/j.1574-6968.2011.02385.x. [DOI] [PubMed] [Google Scholar]
- Vogt RN, Steenkamp DJ, Zheng R, Blanchard JS. The metabolism of nitrosothiols in the mycobacteria: identification and characterization of S-nitrosomycothiol reductase. Biochem J. 2003;374:657–666. doi: 10.1042/BJ20030642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walshaw J, Gillespie MD, Kelemen GH. A novel coiled-coil repeat variant in a class of bacterial cytoskeletal proteins. J Struct Biol. 2010;170:202–215. doi: 10.1016/j.jsb.2010.02.008. [DOI] [PubMed] [Google Scholar]
- Wang L, Yu Y, He X, Zhou X, Deng Z, Chater KF, Tao M. Role of an FtsK-like protein in genetic stability in Streptomyces coelicolor A3(2) J Bacteriol. 2007;189:2310–2318. doi: 10.1128/JB.01660-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wasserstrom S, Grantcharova N, Ubhayasekera W, Ausmees N, Sandblad L, Flardh K. Non-sporulating ftsZ mutants in Streptomyces coelicolor reveal amino acid residues critical for FtsZ polymerization dynamics. Microbiology. 2013;159:890–901. doi: 10.1099/mic.0.066480-0. [DOI] [PubMed] [Google Scholar]
- Willemse J, Borst JW, de Waal E, Bisseling T, van Wezel GP. Positive control of cell division: FtsZ is recruited by SsgB during sporulation of Streptomyces. Genes Dev. 2011;25:89–99. doi: 10.1101/gad.600211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willemse J, Mommaas AM, van Wezel GP. Constitutive expression of ftsZ overrides the whi developmental genes to initiate sporulation of Streptomyces coelicolor. Antonie Van Leeuwenhoek. 2012;101:619–632. doi: 10.1007/s10482-011-9678-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willey J, Willems A, Kodani S, Nodwell JR. Morphogenetic surfactants and their role in the formation of aerial hyphae in Streptomyces coelicolor. Mol Microbiol. 2006;59:731–742. doi: 10.1111/j.1365-2958.2005.05018.x. [DOI] [PubMed] [Google Scholar]
- Wolanski M, Jakimowicz D, Zakrzewska-Czerwinska J. AdpA, key regulator for morphological differentiation regulates bacterial chromosome replication. Open Biol. 2012;2:120097. doi: 10.1098/rsob.120097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wosten HA, Willey JM. Surface-active proteins enable microbial aerial hyphae to grow into the air. Microbiology. 2000;146:767–773. doi: 10.1099/00221287-146-4-767. [DOI] [PubMed] [Google Scholar]
- Xu D, Kim TJ, Park ZY, Lee SK, Yang SH, Kwon HJ, Suh JW. A DNA-binding factor, ArfA, interacts with the bldH promoter and affects undecylprodigiosin production in Streptomyces lividans. Biochem Biophys Res Commun. 2009;379:319–323. doi: 10.1016/j.bbrc.2008.12.052. [DOI] [PubMed] [Google Scholar]
- Xu W, Huang J, Lin R, Shi J, Cohen SN. Regulation of morphological differentiation in S. coelicolor by RNase III (AbsB) cleavage of mRNA encoding the AdpA transcription factor. Mol Microbiol. 2010;75:781–791. doi: 10.1111/j.1365-2958.2009.07023.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang YH, Song E, Willemse J, Park SH, Kim WS, Kim EJ, Lee BR, Kim JN, van Wezel GP, Kim BG. A novel function of Streptomyces integration host factor (sIHF) in the control of antibiotic production and sporulation in Streptomyces coelicolor. Antonie Van Leeuwenhoek. 2012;101:479–492. doi: 10.1007/s10482-011-9657-z. [DOI] [PubMed] [Google Scholar]
- Yao MD, Miyazono KI, Ohtsuka J, Hirano S, Nagata K, Horinouchi S, Ohnishi Y, Tanokura M. Purification, crystallization and preliminary X-ray analysis of the DNA-binding domain of AdpA, the central transcription factor in the A-factor regulatory cascade in the filamentous bacterium Streptomyces griseus, in complex with a duplex DNA. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2012;68:946–949. doi: 10.1107/S1744309112026899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang R, Duggan E, Gu M, Joachimiak A, et al. 2007. The crystal structure of a protein of unknown function from Corynebacterium glutamicum ATCC 13032. MMDB 46531 (See Madej., 2012)
- Zhang D, Wang M, Du G, Zhao Q, Wu J, Chen J. Surfactant protein of the Streptomyces subtilisin inhibitor family inhibits transglutaminase activation in Streptomyces hygroscopicus. J Agric Food Chem. 2008a;56:3403–3408. doi: 10.1021/jf703567t. [DOI] [PubMed] [Google Scholar]
- Zhang D, Wang M, Du G, Zhao Q, Wu J, Chen J. Two different proteases from Streptomyces hygroscopicus are involved in transglutaminase activation. J Agric Food Chem. 2008b;56:10261–10264. doi: 10.1021/jf8008519. [DOI] [PubMed] [Google Scholar]
- Zhang G, Tian Y, Hu K, Zhu Y, Chater KF, Feng C, Liu G, Tan H. Importance and regulation of inositol biosynthesis during growth and differentiation of Streptomyces. Mol Microbiol. 2012;83:1178–1194. doi: 10.1111/j.1365-2958.2012.08000.x. [DOI] [PubMed] [Google Scholar]
- Zhi XY, Li WJ, Stackebrandt E. An update of the structure and 16S rRNA gene sequence-based definition of higher ranks of the class Actinobacteria, with the proposal of two new suborders and four new families and emended descriptions of the existing higher taxa. Int J Syst Evol Microbiol. 2009;59:589–608. doi: 10.1099/ijs.0.65780-0. [DOI] [PubMed] [Google Scholar]
- Zotzel J, Keller P, Fuchsbauer HL. Transglutaminase from Streptomyces mobaraensis is activated by an endogenous metalloprotease. Eur J Biochem. 2003a;270:3214–3222. doi: 10.1046/j.1432-1033.2003.03703.x. [DOI] [PubMed] [Google Scholar]
- Zotzel J, Pasternack R, Pelzer C, Ziegert D, Mainusch M, Fuchsbauer HL. Activated transglutaminase from Streptomyces mobaraensis is processed by tripeptidyl aminopeptidase in the final step. Eur J Biochem. 2003b;270:4149–4155. doi: 10.1046/j.1432-1033.2003.03809.x. [DOI] [PubMed] [Google Scholar]