

Abstract
Purpose
This study was designed to examine the constrictive potential of C-Phycocyanin (C-PC) in regulating changes imposed on gene expression in the selenite-induced cataract model.
Methods
Wistar rat pups were divided into three groups of eight each. On P10, Group I received an intraperitoneal injection of normal saline. Groups II and III received a subcutaneous injection of sodium selenite (19 μmol/kg bodyweight); Group III also received an intraperitoneal injection of C-PC (200 mg/kg bodyweight) on P9–14. Total RNA was isolated on P16, and the relative abundance of mRNA of the crystallin structural genes, redox components, and apoptotic cascade were ascertained with real-time PCR with reference to the internal control β-actin.
Results
Real-time PCR analysis showed the crystallin genes (αA-, βB1-, γD-) and redox cycle components (Cat, SOD-1, Gpx) were downregulated, the apoptotic components were upregulated, and antiapoptotic Bcl-2 was downregulated in Group II. Treatment with 200 mg/kg bodyweight C-PC (Group III) transcriptionally regulated the instability of the expression of these genes, thus ensuring C-PC is a prospective anticataractogenic agent that probably delays the onset and progression of cataractogenesis induced by sodium selenite.
Conclusions
C-PC treatment possibly prevented cataractogenesis triggered by sodium selenite, by regulating the lens crystallin, redox genes, and apoptotic cascade mRNA expression and thus maintains lens transparency. C-PC may be developed as a potential antioxidant compound applied in the future to prevent and treat age-related cataract.
Introduction
Most human age-related metabolic diseases and disorders are due to oxidative damage, which plays a fundamental role in age-related metabolic changes such as cataract. Being an avascular organ, the lens is aided by an efficient glutathione redox cycle to resist oxidative stress [1]. The versatile regulation of intracellular redox components is the primary factor in lens homeostasis. Oxidative stress is a plethora of reactive oxygen species (ROS) such as superoxide anion, hydrogen peroxide, hydroxyl radical, and inducible nitric oxide species that exert a constellation of aberrant changes in the oxidation-reduction status of the cell. Redox-regulating elements such as superoxide dismutase (SOD), catalase (CAT), reduced glutathione (GSH), glutathione reductase (GR), glutathione-S-transferase (GST), and glutathione peroxidase (Gpx) dynamically neutralize the reactive free radicals generated during the physiological process [2,3].
Crystallins are water soluble and highly stable structural proteins synthesized prenatally in the lens epithelial cells classified as three major components such as α-, β- and γ- [4]. The higher solubility, stability, and chaperone function of α-crystallin contribute to the refractive function of this organ [5]. Being a member of the small heat shock family proteins, α-crystallin protects other native proteins from misfolding and aggregation thus ensuring conformational stability of the lenticular milieu [6]. One of the most striking changes in age-related cataract is the post-translational modifications such as racemization, phosphorylation, deamidation, truncation, methylation, and thiolation [7-9], which impose aberrant changes on the conformation and functionality of crystallins. β-crystallin occurs as five different polypeptide chains (βB1–4) as a heterogeneous partner of γ-crystallins [10]; also this apparent heterogeneity has a crucial role in maintaining lens transparency [11,12].
Taken at face value, senile cataract is intimately associated with the increased risk of oxidative stress, which levies irrevocable changes in the lenticular proteins that impel the lens epithelial cells to the apoptotic phase. Apoptotic signaling pathways are triggered by various molecular cues; one of the most common is ROS [13,14]. Apoptosis of lens epithelial cells initiates a cascade of events in senile cataract formation [15] followed by biochemical changes such as activation of calpain protease, loss of cytoskeleton integrity, and aggregation [16]. Sodium selenite–induced cataractogenesis in rodents is a rapid and convenient model for mimicking oxidative stress–related cataract and assessing the metabolic and molecular changes in the lens epithelium. In rat, selenite cataract is induced with a single subcutaneous injection of excess sodium selenite (Na2SeO3) in suckling rats on P10 resulting in severe bilateral, nuclear cataract [17,18] development on P16. These models have been applied by researchers to evaluate the efficacy of various natural products focused on biomedical applications [19].
Evaluating the anticataractogenic role of natural product C-Phycocyanin (C-PC) from cyanobacterial origin on gene expression is the primary objective that may reveal targets for therapeutic interventions and elucidate the molecular basis of the disease. In our earlier findings, the putative anticataract property of C-PC (200 mg/kg bodyweight) was established with biochemical, morphological [20], and histological analysis [21]. The present study aims to broaden the vision on molecular mechanism through which C-PC exerts the anticataractogenic potency by quantitative real time PCR analysis.
Methods
Extraction of C-PC from Spirulina platensis
Spirulina platensis (Kindly gifted by Dr. R. Sarada, Central Food and Technological Research Institute (CFTRI), Mysore, India) was cultured in Zarrouk’s media with a 12 h: 12 h light-dark photoperiod at pH 9.2 at room temperature (27±2 °C). C-PC was extracted by the standard method of freeze thawing and purified by two step column purification process as described in our previous study [20]. All chemicals used for this analysis were of analytical grade. For cDNA conversion, MMlv reverse transcriptase was purchased from Merck (Darmstadt, Germany). Primers specific to the respective full-length mRNA were designed with Net Primer and custom synthesized by Sigma-Aldrich (Bengaluru, India). Real-time PCR Ready Mix (SYBR Green, Bio-Rad, Hercules, CA) and DNA polymerase were purchased from KAPPA, Biosystem (Wilmington, MA).
Animal experiment regime
Neonatal rat pups of the Wistar strain (Procured from Tamil Nadu Veterinary and Animal Sciences University (TANUVAS), Chennai, India) initially weighing 10–12 g on P8 were used for this study. The study protocol was approved by the Institutional Animal Ethical Committee (Ref no. BDU/IAEC/33/2013/09.04.2013). All ethical guidelines were followed for conducting animal experiments in strict adherence with the Institutional Animal Ethical Committee and Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Government of India, and the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Research. The pups were housed along with their mother in polypropylene cages in rooms maintained with a constant 12 h:12 h light-dark cycle. They were fed rodent-pelleted chow and water ad libitum. The pups were randomly grouped into a control group and two experimental groups, comprising eight pups each. On P10, Group I (normal) received vehicle control saline intraperitoneally, and Groups II and III were injected with sodium selenite (19 μmol/kg bodyweight) subcutaneously. Group III received 200 mg/kg bodyweight of C-PC intraperitoneally (dosage was fixed based on our previous study) [20]. C-PC was administered from P9 (1 day before sodium selenite administration) and followed for 6 consecutive days thereafter. Sixteen days after birth (after the eyes opened), the rats were examined with the naked eye for the development of cataract and graded according to the degree of opacity that had developed. The animals were euthanized by cervical dislocation, and the eye lenses were dissected with the posterior approach.
Quantitative real-time PCR analysis
Total RNA was extracted from eye lenses using TRI reagent (Sigma Aldrich) [22]. The concentration and purity of total RNA were determined by absorbance at 260/280 nm in an ultraviolet (UV) spectrophotometer [23]. Starting with equal quantities of total RNA, first-strand cDNA (cDNA) was synthesized using MMlv reverse transcriptase (Merck, Darmstadt, Germany), and gene expression was quantitatively determined using real-time PCR with SYBR Green Supermix (Bio-Rad).
Ct (threshold cycle) is the cycle at which the fluorescence of the reaction mixture exceeds the baseline signal. The difference between the Ct value of the target transcripts and the internal control transcripts were taken into account for calculation. The following thermocycler program was used for real-time PCR: 10 min preincubation at 95 °C, followed by incubation at 95 °C for 30 s, varied annealing temperatures (Table 1) for each gene of interest for 45 s, and 72 °C for 1 min. The sense and antisense primers used for the study are shown in Table 1. Gene expression was normalized to β-actin (internal control) as described earlier [24]. The mRNA levels are reported as relative fold changes. Real-time PCR analysis was performed in triplicate for each gene along with the β-actin internal control to minimize inter-experimental variations. After amplification, a melt curve analysis was performed to confirm whether multiple specific products were amplified in these reactions. All products were melted at 95 °C, annealed at 55 °C, and subjected to increasing temperature changes. Fluorescent data were collected until the reaction reached 95 °C. The results were plotted as raw fluorescence data units versus temperature. The presence of single homogeneous melt peaks confirms specific amplification.
Table 1. Details of primers used for expression analysis.
| S.No | Name of the gene | NCBI Id | Sequence | Annealing temperature/cycles |
|---|---|---|---|---|
| 1. |
αA- cry |
NM_012534.2 |
Sense: TGGCAAACACAACGAGAGGCA
Antisense: GCCAGCATCCAAGCCAGACT |
58 °C/35 |
| 2. |
βB1 -cry |
NM_012936.2 |
Sense: CACCAACACAGCATCAGAAACCAT
Antisense: GTTTTCCTGCTCAAAGACGACCA |
60 °C/35 |
| 3. |
γD- cry |
NM_033095.1 |
Sense primer: CGACTCTGTCCGCTCCTGCC
Antisense: TACTCCCCGGGCCTCAGCAG |
59 °C/35 |
| 4. |
Cat |
NM_012520.1 |
Sense: ACTGGGACCTCGTGGGAAAC
Antisense: TCTGGAATCCCTCGGTCGCT |
57 °C/40 |
| 5. |
SOD-1 |
NM_017050.1 |
Sense: CGGTGCAGGGCGTCATTCACTT
Antisense: CTCTTCATCCGCTGGACCGCC |
58 °C/35 |
| 6. |
Gpx |
NM_030826.3 |
Sense: CACCACGACCCGGGACTACA
Antisense: AGGTAAAGAGCGGGTGAGCC |
58 °C/35 |
| 7. |
Fas |
NM_139194.2 |
Sense: TTCGAATGCAAGGGACTGAT
Antisense: TGGGCCCACTTGATATAACC |
56 °C/35 |
| 8. |
TNF-α |
NM_012675.2 |
Sense: TAGCAAACCACCAAGCAGAG
Antisense: CCTTGAAGAGAACCTGGGAG |
60 °C/40 |
| 9. |
Caspase-8 |
NM_022277.1 |
Sense: TGGGAAGGATCGACGATTAC
Antisense: CCACATGTCCTGCATTTTGA |
58 °C/35 |
| 10. |
Caspase-3 |
NM_012922.2 |
Sense: GGTTCATCCAGTCACTTTGC
Antisense: CCAGGGAGAAGGACTCAAAT |
60 °C/40 |
| 11. |
Bid |
NM_022684.1 |
Sense: CTAGGGACATGGAGAACGAC
Antisense: CCCTCACGTAGGAGAATAGG |
60 °C/40 |
| 12. |
Bad |
NM_022698.1 |
Sense: TCCGAAGAATGAGCGATGAA
Antisense: TTCCCCAAATTTCGATCCCA |
59 °C/40 |
| 13. |
Bax |
NM_017059.2 |
Sense: CGATGAACTGGACAACAACA
Antisense: AGCAAAGTAGAAAAGGGCAA |
60 °C/40 |
| 14. |
Bcl-2 |
NM_016993.1 |
Sense: GGGATGCCTTTGTGGAACTA
Antisense: CTCACTTGTGGCCCAGGTAT |
60 °C/35 |
| 15. |
Caspase-9 |
NM_031632.1 |
Sense: GAACGACCTGACTGCTAAGA
Antisense: GAACGACCTGACTGCTAAGA |
57 °C/35 |
| 16. |
Cytochrome- C |
JF919282.1 |
Sense: TTGTTCAAAAGTGTGCCCAG
Antisense: GTAAGAGAATCCAGCAGCCT |
61 °C/40 |
| 17. |
NFkB |
NM_001276711.1 |
Sense: ACACCTCTACACATAGCAGC
Antisense: GGCACTACTCCCTCATCTTC |
60 °C /40 |
| 18. | β-actin | NM_001109553.1 | Forward primer: AGTCCCTCACCCTCCCAAAA Reverse primer: AGCAATGCTGTCACCTTCCC | 60 °C /40 |
Data analysis
Results from real-time PCR were calculated from Ct values, indicating the threshold fluorescence reached the exponential phase of amplification. Gene expression was normalized to with respect to an invariant housekeeping gene. β-actin was used as reference gene, and the normalization representation of target gene (Ct1) to reference gene (Ct2) was calculated as ΔCt=(Ct1- Ct2). Using the 2-Δ(ΔCt) method, the data are presented as the fold change in gene expression normalized to the endogenous reference gene and relative to the untreated control. From this method, 2-Δ(ΔCt) is directly proportional to the ratio of target to the reference initial copy number for each sample [25].
Statistical analysis
Results are presented as mean ± standard error mean (SEM). Data were analyzed using one-way ANOVA (ANOVA) followed by Student’s Newman-Keul’s test (SNK) with GraphPad Prism Project 6 (GraphPad, San Diego, CA). In all cases, p<0.05 was considered statistically significant. A p value less than 0.05 were required to reject the null hypothesis that the group means were equivalent.
Results
In this study, the expression pattern of three structural genes, three redox genes, and 11 apoptotic pathway genes were studied with real-time PCR. Total RNA was isolated from three different groups and starting with equal quantities of total RNA (2 μg) with high purity and integrity. Dynamic changes in mRNA levels were expressed in fold changes and normalized with internal control β-actin (NM_031144.2). (The melt curve analysis for each gene expression is provided as supplementary data.)
Real-time expression of the crystallin
The status of expression of the candidate genes of the crystallin family αA- (Figure 1A), βB1- (Figure 1B), and γD-crystallin (Figure 1C) were examined. Deleterious downregulation of all crystallin gene expression was observed in cataract-induced Group II. A 0.5-fold downregulation of αA-crystallin (NM_012534.2), 0.75-fold downregulation of βB1-crystallin (NM_012936.2), and 0.5-fold downregulation of γD-crystallin (NM_033095.1) recorded in Group II compared to Groups I and III. However, on treatment with C-PC the mRNA transcripts were maintained significantly (p<0.05) at normal levels.
Figure 1.

Changes in the expression pattern of αA- (A), βB1- (B), and γD- crystallin (C) mRNA analyzed with comparative real-time PCR normalized to β-actin and expressed as folds of increases or decreases relative to controls. Data are the means of three comparative real-time PCR experiments, each performed in triplicate. Values were significantly (p<0.05) decreased when the animals were exposed to sodium selenite and were significantly restored in Group III induced with sodium selenite and treated with C-Phycocyanin (C-PC) 200 mg/kg bodyweight. The significance was considered at the level of p<0.05. a represents the statistical significance between the Group I and Group II, and b represents the statistical significance of Group II versus Group III.
Changes in redox enzyme expression
A concomitant decrease in the mRNA expression of the redox enzymes such as Cat (NM_012520.1; Figure 2A), SOD-1 (NM_017050.1; Figure 2B), and Gpx (NM_030826.3; Figure 2C) was ascertained in selenite-induced cataractogenic group (Group II). Interestingly, 0.9-fold downregulation of CAT gene, 0.5-fold downregulation of SOD-1 gene, and 0.5-fold downregulation of Gpx gene were observed in Group II compared to Group I. The aberrant change on the redox mechanism was counteracted by the C-PC treatment, which may be due to the scavenging of free radicals by C-PC, which thus conserved the expression of the antioxidant enzymes significantly (p<0.05) in Group III.
Figure 2.
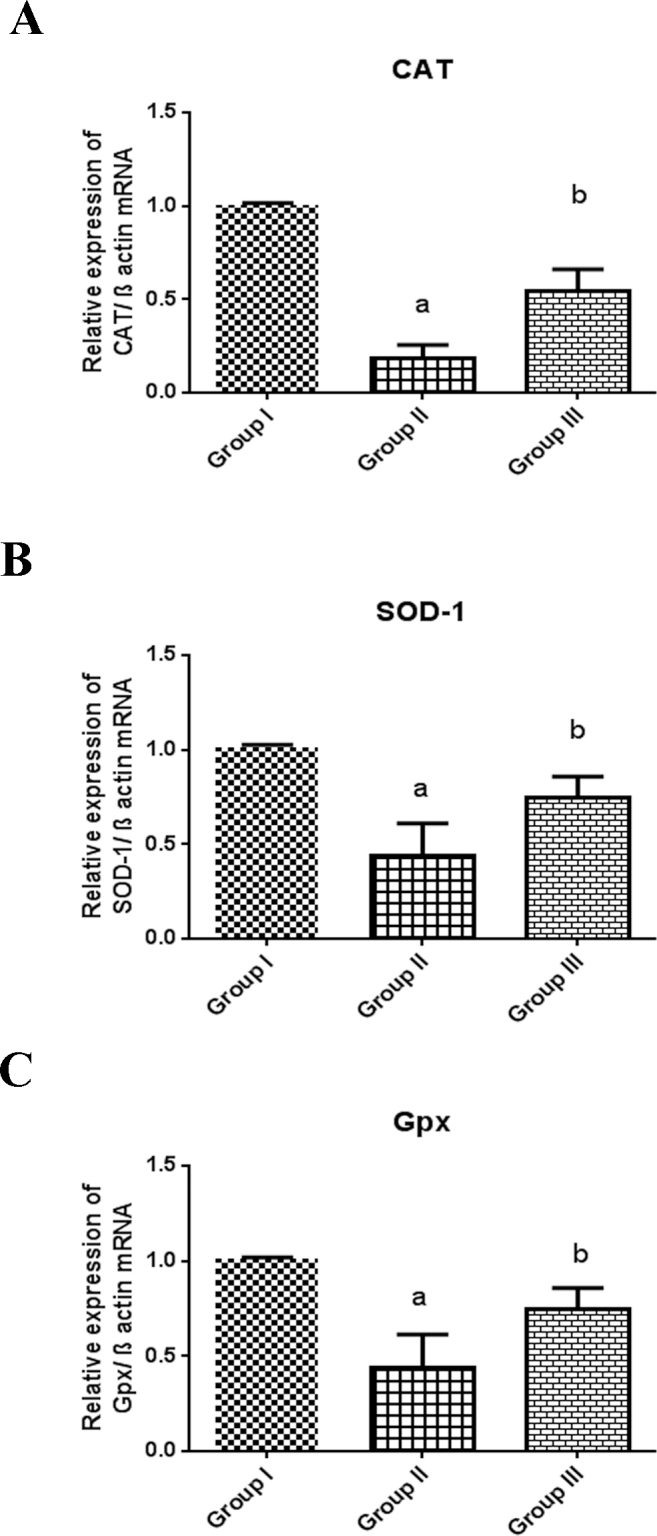
mRNA expression of Cat (A), SOD-1 (B), and Gpx (C) from Group I, II, and III rat lenses. Total RNA was extracted from rat lens tissues, and mRNA levels were measured using real-time PCR normalized to β-actin and expressed as folds of increases or decreases relative to controls. The data are expressed as mean ± standard error of the mean (SEM) of three independent observations. Statistical significance between the control group versus the other groups was considered at the level of p<0.05 using SNK test. a represents the statistical significance Group I versus Group II, and b represents the statistical significance between Group II and Group III.
Regulation of mRNA expression of extrinsic signaling molecules
The extrinsic pathway is activated by the death domain which further recruits downstream signaling molecules such as caspase-8 and caspase-3 that triggers apoptosis. The results confirmed increased mRNA expression levels of Fas (NM_139194.2; Figure 3A; 7-fold increase), TNFα (NM_012675.2; Figure 3B; 12-fold increase), caspase-8 (NM_022277.1; Figure 3C; 0.5-fold increase), caspase-3 (NM_012922.2; Figure 3D; 1.5-fold increase), and Bid (NM_022684.1; Figure 3E; 2.1-fold increase) genes in Group II. However, Group III treated with C-PC strikingly conserved the signaling factors at placebo levels.
Figure 3.

mRNA expression of extrinsic apoptotic signaling molecules (A) Fas, (B) TNFα, (C) Caspase-8, (D) Caspase-3, and (E) Bid in selenite-induced cataractogenesis. The mRNA expression of apoptotic genes were analyzed with real-time PCR using SYBR Green dye. Target gene expression was normalized to β-actin mRNA expression, and the results are expressed as fold change from control. a represents statistical significance of Group I versus Group II, and b represents the statistical significance of Group II versus Group III at the the level of p<0.05 using SNK test.
Expression of Bcl-2 family members
Bcl-2 family proteins are intracellular signaling molecules that consist of proapoptotic factors such as Bad (NM_022698.1; Figure 4A) and Bax (NM_017059.2; Figure 4B) and antiapoptotic Bcl-2 (NM_016993.1; Figure 4C) that regulate mitochondrial membrane integrity, cytochrome-C release, and caspase activation [26,27]. A dynamic overexpression of the proapoptotic Bad (24-fold increase) and Bax (1.9-fold increase) genes demonstrated in the Group II samples accompanied by concerted downregulation of the antiapoptotic gene Bcl-2 was addressed in the selenite-induced cataractogenic group. The ratio of Bax/Bcl-2 is an important factor that determines the fate of a cell entering apoptosis. Pertaining to the results, the antiapoptotic Bcl-2 was downregulated 0.9 fold during cataractogenesis pushing the epithelial cells toward death. This imbalance in gene expression was counteracted and stabilized by C-PC treatment in Group III animals.
Figure 4.
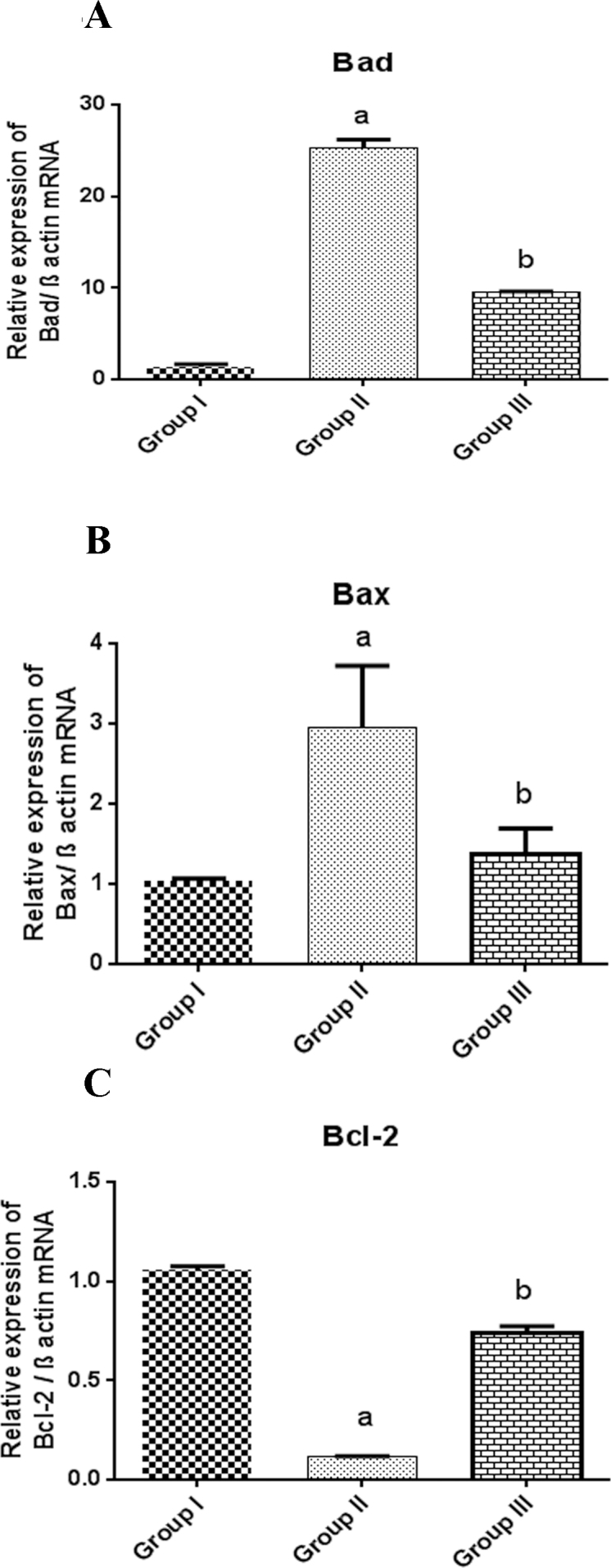
mRNA expression of Bad (A), Bax (B), and Bcl-2 (C) in the eye lens of sodium selenite induced and C-PC treated animals. β-actin was used as an internal marker, and the fluorescent intensity was normalized by the intensity of β-actin. Each bar represents mean ± standard error of the mean (SEM) of three independent observations. Statistical significance was considered at p<0.05. a Group I versus Group II; b represents Group II versus Group III.
Expression of intrinsic apoptotic signaling molecules
To deduce the protective role of C-PC on the mitochondria-driven signals, the expression of intrinsic apoptotic signaling genes were examined. The gene expression of caspase-9 (NM_031632.1; Figure 5A; 0.8-fold increase) and cytochrome-C (JF919282.1; Figure 5B; 1.7-fold increase) were upregulated in the selenite-induced cataractogenic Group II suggesting the monolithic apoptotic load on the intrinsic signals. The mRNA transcripts were conserved significantly (p<0.05) at nearly normal levels on treatment with 200 mg/kg bodyweight of C-PC in Group III.
Figure 5.
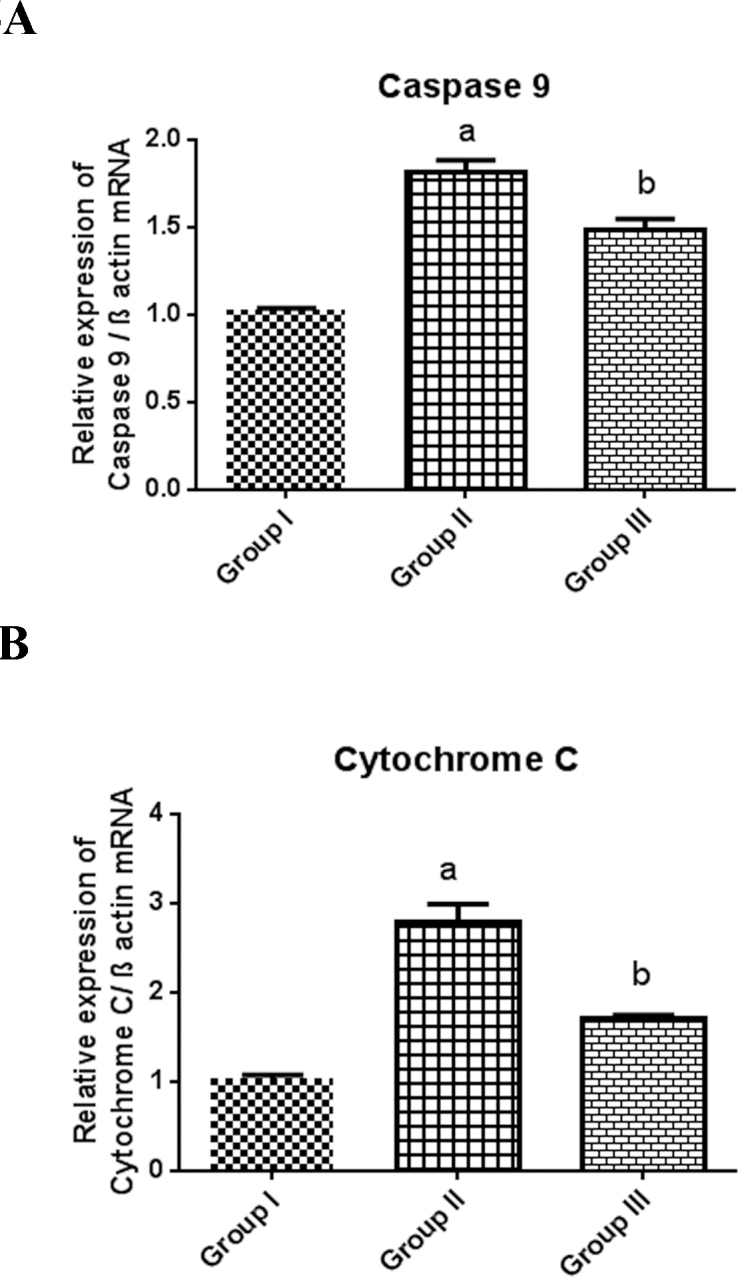
mRNA expression of intrinsic apoptotic factors such as caspase-9 (A) and cytochrome-C (B). β-actin was used as an internal marker. Statistical significance between the control group versus other groups was considered at the level of p<0.05 using SNK test. a represents the statistical significance Group I versus Group II, and b represents the statistical significance of Group II versus Group III.
Regulation of apoptosis by transcriptional factor NFκB
NFκB [NM_001276711.1] is a class of inducible transcriptional factors that directly intervene with and regulate the apoptotic factors. Strikingly, the expression of this transcriptional factor was upregulated significantly (p<0.05) in Group II (Figure 6; 0.2-fold increase) during cataractogenesis, thus eliciting the apoptotic cascade. The results elegantly demonstrate that C-PC treatment conserved the expression of the transcriptional factor and supported the notion that C-PC averts an oxidative stress–induced mimic of cataract in rodents by suppressing the activation of NFκB during the cataractogenic process in Group III.
Figure 6.
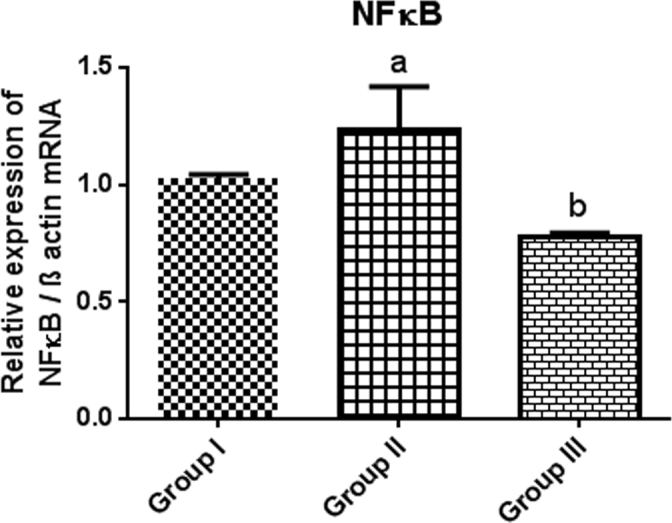
The relative mRNA level of NFκB was determined with real-time PCR and normalized to β-actin. Statistical significance was considered at p<0.05. a denotes the statistical significance of Group I versus Group II; b refers to the statistical significance of Group II versus Group III.
Discussion
The entire lens tissue depends on epithelial cells for nutrition and survival; endurance of the epithelium during adverse conditions is essential for retaining its integrity and proceeding with vital organ functions [28]. Therefore, the factors that play a key role in epithelial cell survival must be deduced. Classical changes that precede opacification in the entire lens are the depletion of lens epithelial cells, modification in the metabolism of the epithelium probably due to DNA damage, inhibition of DNA synthesis, endoplasmic stressors inducing the unfolded protein response pathway, and the apparent drop in the oxidative defense mechanism [29]. It is clear that with the impairment of epithelial cells and influx of calcium the cysteine proteinase Calpain is activated, which degrades the cytoskeletal compartments resulting in lens protein modification. Anomalous changes in the crystallin structure and aggregation state are striking features of cortical and nuclear cataractogenesis [4]. With increased age, the α-, β-, and γ-crystallins undergo extensive conformational changes in the secondary and tertiary structures, including loss in the chaperone activity of α-crystallin [30,31]. In addition, at the onset of cataract, the solubility and stability of crystallin proteins are compromised [12]. Comprehensive research on crystallins has revealed that these proteins are constantly formed throughout an animal’s lifetime; however, major modifications during the onset of cataract are imposed on the solubility and stability of the protein. In this study, a systemic analysis of lens crystallin mRNA was conducted that revealed a profound loss of transcripts in the selenite cataractous group (Group II), suggesting decreased expression of these genes. The results were consistent with earlier reports that αA-, βB1-, and γD-crystallin transcripts were degraded in dexamethasone-induced cataractogenesis [6]. Restoration of the mRNA levels in the C-PC-treated group may be attributed to the antioxidant mechanism of C-PC that evidently conserves the crystallin transcripts in selenite-induced cataract.
Extrapolating epidemiological data revealed that cataractogenesis elicits an unfathomable relationship between the loss of antioxidant enzyme activity and the progression of opacification [29,32]. A rapid increase in the loss of enzyme activity ought to be the foremost impingement on the epithelial cells [33]. The mRNA of the major antioxidant enzymes such as Cat, SOD-1, and Gpx were dramatically decreased in the cataractogenic group of animals (Group II), which compared favorably to the challenged redox machinery. This is the significant result of the study because the development of cataract in part is bound to the decrease in antioxidant enzyme activity [20]. The deleterious oxidative stress burden imposed on antioxidant enzymes were thwarted in Group III on treatment with C-PC. The positive influence observed on treatment with the natural antioxidant underscores the magnitude of anticataractogenic potential of C-PC on selenite-induced cataractogenesis. Earlier, our biochemical studies [20] showed that C-PC modulates the antioxidant enzyme activity status in selenite-induced cataractogenesis, and thus obviated any pertinacious changes that impede the redox state of the lens. The results of the current study corroborate the earlier findings [20] and suggest that the tetrapyrrole ring, a core component of phycobiliproteins (an active constituent), is linked to the scavenging of pestilent free radicals generated during the cataractogenic process, and conserves the antioxidant status of the lens tissue.
Clinical samples from human cataractous lenses showed a steady rate of apoptotic death in the capsular epithelial cells, which is one of the striking perturbations of the cataractogenic process [34]. Apoptosis is an early event in selenite cataract culminating in loss of epithelial cell metabolism [35]. Okamura et al. [36] reported a high level of Fas mRNA expression, indicating the greater chance of epithelial cells heading toward apoptosis induced by the death receptor pathway. The external stress stimuli were sensitized by the Fas system that reconciles the activation of caspase-3 and caspase-8. In the present study, treatment with C-PC suppressed the extrinsic signaling molecules of mRNA expression (Fas and TNF-α). Further activation of caspase-3, caspase-8, and Bid in Group II affirms the hypothesis that the cataractogenic process was potentiated in the extrinsic and intrinsic pathways of apoptosis. Induction with selenite resulted in a significant increase in caspase-3 expression, and activation was observed in Group II, while C-PC treatment significantly (p<0.05) inhibited the change in vivo. These results are consistent with previous studies on the protective effect of C-PC on D-galactose-induced apoptosis in SRA01/04 cells [37]. Although the extrinsic apoptotic trigger accounts for majority of the cell death, an equal apoptotic load is imposed on the intrinsic pathway. The downregulation of mitochondrial genes in Group II suggests that mitochondrial integrity is lost after induction with selenite thus contributing to the intrinsic pathway. The relative equipoise of proapoptotic (Bax and Bad) and antiapoptotic factor (Bcl-2) are crucial determinants of cellular homeostasis [38]. Accumulating evidence from real-time PCR results shows that C-PC subdues apoptosis in lens epithelial cells by tightly regulating intrinsic apoptotic regulators such as Bcl-2 family proteins and by preventing the activation of caspases. We observed that C-PC suppressed selenite-induced upregulation of Bad and Bax and downregulation of Bcl-2 expression. The findings of the present study substantiate the earlier findings of Ou et al. [37,39] and our biochemical studies [20] that link to the anticataractogenic attributes of C-PC. Consequently, the release of apoptogenic proteins such as cytochrome-c from the mitochondria culminates in caspase-9 activation and caspase-dependent cell death. Treatment with C-PC resulted in suppression of caspase-9 gene expression, and this may explain, at least in part, that C-PC concealed apoptosis and membrane permeabilization of cytochrome-c. Belusko [40] reported the loss of integrity of lens epithelial cells by selenite caused by preferential downregulation of mitochondrial RNAs accompanied by the release of cytochrome-c and impaired mitochondrial function. These findings provide insights into the diverse caveats of the molecular mechanism of C-PC against apoptosis and in line, averting cataract. A previous study by Ou et al. [37] showed that cotreatment with C-PC markedly diminished the activation of D-galactose-triggered unfolded protein response (UPR) pathway, a hallmark of age-related cataract.
NFκB is a crucial redox sensitive transcription factor that controls the expression of stress inducible genes [41]. ROS-induced stimuli can release inhibitor kappa B (IκB) and facilitate the translocation of NFκB into the nucleus that binds to DNA response elements and manipulates the transcription of specific genes that determine cellular fate [42]. Roy et al. [43,44] studied the role of C-PC in ROS-mediated activation of NFκB and stated that C-PC treated RAW 264.7 cells showed reduced translocation of NFκB even after ROS were induced with 2-acetylaminofluorene. TNF-α is an excitatory cytokine of human lens epithelial cells that activates NFκB. The biologic response to oxidative stress during cataractogenesis upregulates the inflammatory cytokine TNF-α in Group II (an approximate 12-fold increase), which is paired with the concerted upregulation of NFκB compared to the concordant expression in the lens tissue of Group I. The observed decrease in NFκB expression in the Group III rat lenses treated with C-PC shows a concrete relationship with the earlier findings that NFκB plays a critical role in regulating the apoptotic machinery of lens epithelial cells in a temporal sequence. Due to diverse mechanism of action and its potent antioxidant capacity, C-PC is a more effective anticataractogenic agent of cyanobacterial origin.
Hayashi et al. [24] investigated the effects of antioxidant supplementation on the mRNA expression of antioxidative enzymes in the lens. The authors also reported that assessing the synthesis of antioxidant enzymes are important criterion in verifying the effectiveness of antioxidant supplementation in preventing cataract. Since the first stage of protein synthesis is initiated by mRNA transcription, evaluating changes in mRNA expression may point toward changes in the synthesis of proteins, such as antioxidant enzymes. Therefore, measuring changes in the mRNA expression levels of antioxidant enzymes in the lens tissue may be important for substantiating the effects of antioxidant supplementation on preventing cataracts.
The current study was designed to evaluate the effectiveness of C-PC in preventing changes in the gene expression pattern of vital antioxidant enzymes such as SOD-1, Cat, and Gpx. Real-time quantification of mRNA transcripts confirmed that the synthesis of the transcripts were challenged on selenite-induced cataractogenesis. Owing to free radical scavenging property, C-PC acts directly on the transcriptional modification process and might subsequently modulate other cellular proteins and thus slow cataractogenesis. Previously, Ontawong et al. [45] showed that Spirogyra neglecta extract modulated NFκB activation and the subsequent translocation that is directly reflected in mRNA expression of phase II antioxidant enzymes genes such as Cat, SOD, and Gpx. Our results also corroborate with the earlier findings of Ontawong et al. [45], which is directly reflected in the mRNA expression of the antioxidant enzymes.
Pathogenic stimuli causes the release of IκB facilitating the translocation of NFκB into the nucleus that binds to the DNA control elements and thus influences the transcription of specific genes. In our study, NFκB expression was upregulated in Group II thus indirectly contributing to the suppression of gene expression of antioxidant enzymes. Gene expression was stabilized in Group III on treatment with 200 mg/kg bodyweight of C-PC. C-PC, an effective antioxidant, manipulates the expression of NFκB, maintains full control of oxidative stress apoptosis, and provides indirect protection against free radical injury by stimulating antioxidant enzymes.
This study shows C-PC can withstand oxidative stress and apoptotic negotiator that can be applied in future drug discovery approaches against cataractogenesis. The tight regulation of extrinsic and intrinsic cascade signaling that subsequently decreases lens epithelial cell death by the ensuing changes in NFκB dynamics poses an encouraging and reasonable target against cataractogenesis. Since apoptosis of lens epithelial cells is closely tied to cataract incidence, C-PC may be a promising candidate for preventing age-related cataract.
Acknowledgments
Financial assistance from DST-PURSE (SR/S9/Z-23/2010/17(C) & 17(G)2011), DST-INSPIRE-SRF (2010/129B) to the first author and third authors are greatly acknowledged. The authors also thank the Department of Science and Technology (DST), New Delhi for the DST-YS project (SR/FT/LS-153/2008/ (2010–2013)) sanctioned to the corresponding author. The authors thank Dr. M. Michael Aruldhas, Professor and Head, Department of Endocrinology, Dr. A. L. M. PG IBMS, University of Madras for supporting with the instrumentation facility.
References
- 1.Giblin FJ. Glutathione: a vital lens antioxidant. J Ocul Pharmacol Ther. 2000;16:121–35. doi: 10.1089/jop.2000.16.121. [DOI] [PubMed] [Google Scholar]
- 2.Halliwell B. Biochemistry of oxidative stress. Biochem Soc Trans. 2007;35:1147–50. doi: 10.1042/BST0351147. [DOI] [PubMed] [Google Scholar]
- 3.Makri OE, Ferlemi AV, Lamari FN, Georgakopoulos CD. Saffron administration prevents selenite-induced cataractogenesis. Mol Vis. 2013;19:1188–97. [PMC free article] [PubMed] [Google Scholar]
- 4.Ramkumar S, Fujii N, Fujii N, Thankappan B, Sakaue H, Ingu K, Natarajaseenivasan K, Anbarasu K. Comparison of effect of gamma ray irradiation on wild-type and N-terminal mutants of αA-crystallin. Mol Vis. 2014;20:1002–16. [PMC free article] [PubMed] [Google Scholar]
- 5.Bloemendal H. The vertebrate eye lens. Science. 1977;197:127–38. doi: 10.1126/science.877544. [DOI] [PubMed] [Google Scholar]
- 6.Wang L, Zhao WC, Yin XL, Ge JY, Bu ZG, Ge HY, Meng QF, Liu P. Lens proteomics: analysis of rat crystallins when lenses are exposed to dexamethasone. Mol Biosyst. 2012;8:888–901. doi: 10.1039/c2mb05463a. [DOI] [PubMed] [Google Scholar]
- 7.Fujii N, Kawaguchi T, Sasaki H. Simultaneous stereoinversion and isomerization at the Asp-4 residue in betaB2-crystallin from the aged human eye lenses. Biochemistry. 2011;50:8628–35. doi: 10.1021/bi200983g. [DOI] [PubMed] [Google Scholar]
- 8.Abraham EC, Huaqian J, Aziz A, Kumarasamy A, Datta P. Role of the specifically targeted lysine residues in the glycation dependent loss of chaperone activity of alpha A- and alpha B-crystallins. Mol Cell Biochem. 2008;310:235–9. doi: 10.1007/s11010-007-9685-1. [DOI] [PubMed] [Google Scholar]
- 9.Kumarasamy A, Abraham EC. Interaction of C-terminal truncated human alphaA-crystallins with target proteins. PLoS ONE. 2008;3:e3175. doi: 10.1371/journal.pone.0003175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gangalum RK, Horwitz J, Kohan SA, Bhat SP. Alphaa-crystallin and alphaB-crystallin reside in separate subcellular compartments in the developing ocular lens. J Biol Chem. 2012;287:42407–16. doi: 10.1074/jbc.M112.414854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Horwitz J. Alpha-crystallin. Exp Eye Res. 2003;76:145–53. doi: 10.1016/s0014-4835(02)00278-6. [DOI] [PubMed] [Google Scholar]
- 12.Harrington V, Srivastava OP, Kirk M. Proteomic analysis of water insoluble proteins from normal and cataractous human lenses. Mol Vis. 2007;13:1680–94. [PubMed] [Google Scholar]
- 13.Nakajima T, Nakajima E, Fukiage C, Azuma M, Shearer TR. Differential gene expression in the lens epithelial cells from selenite injected rats. Exp Eye Res. 2002;74:231–6. doi: 10.1006/exer.2001.1131. [DOI] [PubMed] [Google Scholar]
- 14.Bai J, Dong L, Song Z, Ge H, Cai X, Wang G, Liu P. The role of melatonin as an antioxidant in human lens epithelial cells. Free Radic Res. 2013;47:635–42. doi: 10.3109/10715762.2013.808743. [DOI] [PubMed] [Google Scholar]
- 15.Long AC, Colitz CM, Bomser JA. Apoptotic and necrotic mechanisms of stress-induced human lens epithelial cell death. Exp Biol Med (Maywood) 2004;229:1072–80. doi: 10.1177/153537020422901012. [DOI] [PubMed] [Google Scholar]
- 16.Li W-C, Kuszak JR, Wang G-M, Wu Z-Q, Abraham S. Calcimycin-induced lens epithelial cell apoptosis contributes to cataract formation. Exp Eye Res. 1995;61:91–8. doi: 10.1016/s0014-4835(95)80062-x. [DOI] [PubMed] [Google Scholar]
- 17.Shearer TR, Ma H, Fukiage C, Azuma M. Selenite nuclear cataract: review of the model. Mol Vis. 1997;3:8. [PubMed] [Google Scholar]
- 18.David LL, Shearer TR. Calcium-activated proteolysis in the lens nucleus during selenite cataractogenesis. Invest Ophthalmol Vis Sci. 1984;25:1275–83. [PubMed] [Google Scholar]
- 19.Thiagarajan R, Manikandan R. Antioxidants and cataract. Free Radic Res. 2013;47:337–45. doi: 10.3109/10715762.2013.777155. [DOI] [PubMed] [Google Scholar]
- 20.Kumari RP, Sivakumar J, Thankappan B, Anbarasu K. C-Phycocyanin modulates selenite-induced cataractogenesis in rats. Biol Trace Elem Res. 2013;151:59–67. doi: 10.1007/s12011-012-9526-2. [DOI] [PubMed] [Google Scholar]
- 21.Kumari RP, Anbarasu K. Protective role of C-Phycocyanin against secondary changes during sodium selenite mediated cataractogenesis. Nat Prod Bioprospect. 2014;4:81–9. doi: 10.1007/s13659-014-0008-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–9. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- 23.Sambrook J, Russell DW. Molecular Cloning: A Laboratory Manual: Cold Spring Harbor Laboratory Press; 2001; "p.7.4-7.9. [Google Scholar]
- 24.Hayashi R, Hayashi S, Arai K, Chikuda M, Obara Y. Effects of antioxidant supplementation on mRNA expression of glucose-6-phosphate dehydrogenase, beta-actin and 18S rRNA in the anterior capsule of the lens in cataract patients. Exp Eye Res. 2012;96:48–54. doi: 10.1016/j.exer.2012.01.001. [DOI] [PubMed] [Google Scholar]
- 25.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Methods. 2001;25:402–8. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 26.Swanton E, Savory P, Cosulich S, Clarke P, Woodman P. Bcl-2 regulates a caspase-3/caspase-2 apoptotic cascade in cytosolic extracts. Oncogene. 1999;18:1781–7. doi: 10.1038/sj.onc.1202490. [DOI] [PubMed] [Google Scholar]
- 27.Donovan M, Cotter TG. Control of mitochondrial integrity by Bcl-2 family members and caspase-independent cell death. Biochim Biophys Acta. 2004;1644:133–47. doi: 10.1016/j.bbamcr.2003.08.011. [DOI] [PubMed] [Google Scholar]
- 28.Yang Y, Sharma R, Cheng JZ, Saini MK, Ansari NH, Andley UP, Awasthi S, Awasthi YC. Protection of HLE B-3 cells against hydrogen peroxide- and naphthalene-induced lipid peroxidation and apoptosis by transfection with hGSTA1 and hGSTA2. Invest Ophthalmol Vis Sci. 2002;43:434–45. [PubMed] [Google Scholar]
- 29.Shinohara T, White H, Mulhern ML, Maisel H. Cataract: window for systemic disorders. Med Hypotheses. 2007;69:669–77. doi: 10.1016/j.mehy.2006.11.051. [DOI] [PubMed] [Google Scholar]
- 30.Michael R, Bron AJ. The ageing lens and cataract: a model of normal and pathological ageing. Philos Trans R Soc Lond B Biol Sci. 2011;366:1278–92. doi: 10.1098/rstb.2010.0300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Song S, Hanson MJ, Liu BF, Chylack LT, Liang JJ. Protein-protein interactions between lens vimentin and alphaB-crystallin using FRET acceptor photobleaching. Mol Vis. 2008;14:1282–7. [PMC free article] [PubMed] [Google Scholar]
- 32.Shichi H. Cataract formation and prevention. Expert Opin Investig Drugs. 2004;13:691–701. doi: 10.1517/13543784.13.6.691. [DOI] [PubMed] [Google Scholar]
- 33.Kisic B, Miric D, Zoric L, Ilic A, Dragojevic I. Antioxidant capacity of lenses with age-related cataract. Oxid Med Cell Longev. 2012;2012:467130. doi: 10.1155/2012/467130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Li DW, Liu JP, Mao YW, Xiang H, Wang J, Ma WY, Dong Z, Pike HM, Brown RE, Reed JC. Calcium-activated RAF/MEK/ERK signaling pathway mediates p53-dependent apoptosis and is abrogated by alpha B-crystallin through inhibition of RAS activation. Mol Biol Cell. 2005;16:4437–53. doi: 10.1091/mbc.E05-01-0010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tamada Y, Fukiage C, Nakamura Y, Azuma M, Kim YH, Shearer TR. Evidence for apoptosis in the selenite rat model of cataract. Biochem Biophys Res Commun. 2000;275:300–6. doi: 10.1006/bbrc.2000.3298. [DOI] [PubMed] [Google Scholar]
- 36.Okamura N, Ito Y, Shibata MA, Ikeda T, Otsuki Y. Fas-mediated apoptosis in human lens epithelial cells of cataracts associated with diabetic retinopathy. Med Electron Microsc. 2002;35:234–41. doi: 10.1007/s007950200027. [DOI] [PubMed] [Google Scholar]
- 37.Ou Y, Yuan Z, Li K, Yang X. Phycocyanin may suppress D-galactose-induced human lens epithelial cell apoptosis through mitochondrial and unfolded protein response pathways. Toxicol Lett. 2012;215:25–30. doi: 10.1016/j.toxlet.2012.09.017. [DOI] [PubMed] [Google Scholar]
- 38.Lee DH, Cho KS, Park SG, Kim EK, Joo CK. Cellular death mediated by nuclear factor kappa B (NF-kappaB) translocation in cultured human lens epithelial cells after ultraviolet-B irradiation. J Cataract Refract Surg. 2005;31:614–9. doi: 10.1016/j.jcrs.2004.05.053. [DOI] [PubMed] [Google Scholar]
- 39.Ou Y, Geng P, Liao GY, Zhou Z, Wu WT. Intracellular GSH and ROS levels may be related to galactose-mediated human lens epithelial cell apoptosis: role of recombinant hirudin variant III. Chem Biol Interact. 2009;179:103–9. doi: 10.1016/j.cbi.2008.10.039. [DOI] [PubMed] [Google Scholar]
- 40.Belusko PB, Nakajima T, Azuma M, Shearer TR. Expression changes in mRNAs and mitochondrial damage in lens epithelial cells with selenite. Biochim Biophys Acta. 2003;1623:135–42. doi: 10.1016/j.bbagen.2003.08.008. [DOI] [PubMed] [Google Scholar]
- 41.D'Angio CT, Finkelstein JN. Oxygen regulation of gene expression: a study in opposites. Mol Genet Metab. 2000;71:371–80. doi: 10.1006/mgme.2000.3074. [DOI] [PubMed] [Google Scholar]
- 42.Jin XH, Ohgami K, Shiratori K, Koyama Y, Yoshida K, Kase S, Ohno S. Inhibition of nuclear factor-kappa B activation attenuates hydrogen peroxide-induced cytotoxicity in human lens epithelial cells. Br J Ophthalmol. 2007;91:369–71. doi: 10.1136/bjo.2006.107037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Roy KR, Arunasree KM, Dhoot A, Aparna R, Reddy GV, Vali S, Reddanna P. C-Phycocyanin inhibits 2-acetylaminofluorene-induced expression of MDR1 in mouse macrophage cells: ROS mediated pathway determined via combination of experimental and In silico analysis. Arch Biochem Biophys. 2007;459:169–77. doi: 10.1016/j.abb.2007.01.006. [DOI] [PubMed] [Google Scholar]
- 44.Roy KR, Arunasree KM, Reddy NP, Dheeraj B, Reddy GV, Reddanna P. Alteration of mitochondrial membrane potential by Spirulina platensis C-phycocyanin induces apoptosis in the doxorubicinresistant human hepatocellular-carcinoma cell line HepG2. Biotechnol Appl Biochem. 2007;47:159–67. doi: 10.1042/BA20060206. [DOI] [PubMed] [Google Scholar]
- 45.Ontawong A, Saowakon N, Vivithanaporn P, Pongchaidecha A, Lailerd N, Amornlerdpison D, Lungkaphin A, Srimaroeng C. Antioxidant and renoprotective effects of Spirogyra neglecta (Hassall) Kützing extract in experimental type 2 diabetic rats. Biomed Res Int. 2013;2013:820786. doi: 10.1155/2013/820786. [DOI] [PMC free article] [PubMed] [Google Scholar]