

Abstract
When creating a public good, strategies or mechanisms are required to handle defectors. We first show mathematically and numerically that prior agreements with posterior compensations provide a strategic solution that leads to substantial levels of cooperation in the context of public goods games, results that are corroborated by available experimental data. Notwithstanding this success, one cannot, as with other approaches, fully exclude the presence of defectors, raising the question of how they can be dealt with to avoid the demise of the common good. We show that both avoiding creation of the common good, whenever full agreement is not reached, and limiting the benefit that disagreeing defectors can acquire, using costly restriction mechanisms, are relevant choices. Nonetheless, restriction mechanisms are found the more favourable, especially in larger group interactions. Given decreasing restriction costs, introducing restraining measures to cope with public goods free-riding issues is the ultimate advantageous solution for all participants, rather than avoiding its creation.
Keywords: evolutionary games, cooperation, commitment, public goods
1. Introduction
Arranging a prior commitment or agreement is an essential ingredient to encourage cooperative behaviour in a wide range of relationships, ranging from personal to political and religious ones [1–5]. Prior agreements clarify the intentions and preferences of other players. Hence, refusing to establish an agreement may be considered as intending or preferring not to cooperate (non-committers) [5–7]. Prior agreements may be highly rewarding in group situations, as in the case of public goods games (PGGs) [8], as it forces the other participants to signal their willingness to achieve a common goal. Especially for increasing group sizes, such prior agreements could be ultimately rewarding, as it becomes more and more difficult to assess the aspirations of all participants.
In a PGG, where players meet in groups of size N [9,10], all players can choose whether to cooperate and contribute an amount, c, to the public good or to defect and take advantage of the public good without contributing to it. The total contribution is multiplied by a constant public goods producing factor, r > 1, and the result is afterwards distributed equally among all players. With r smaller than the group size (r < N), non-contributing free-riders always gain more than contributors. Evolutionary game dynamics has shown that under those conditions cooperation disappears, which is famously known as the ‘tragedy of commons’ [10,11]. Various mechanisms, such as direct and indirect reciprocity, kin and group selections and costly punishment, have been proposed and evaluated both theoretically and experimentally, which explain the evolution of cooperation nevertheless [10,12,13], ranging from microbial systems to animals and humans societies [12,14–18].
Here, we examine a strategic solution based on prior agreements to address the problem of the evolution of cooperation in the PGG. Prior to the PGG, commitment proposing players ask their co-players to commit to contribute to the PGG, paying a personal proposer's cost to establish that agreement. If all the requested co-players accept the commitment, the proposers assume that everyone will contribute to the public good. Those individuals who commit, yet later do not contribute, receive a penalty and are forced to compensate the proposers at a cost [6,19,20]. As such, our model explicitly and novelly addresses the relevance of the commitment proposing behaviour regarding posterior compensations in group interactions, which has been suggested to be a major pathway to the emergence of cooperation [1,5].
As commitment proposers may encounter also non-committers, they require strategies that can deal with these kinds of individuals [1,6,21]. When dealing with non-committers, the simplest strategy is to not participate in the creation of the common good or, when the interaction is mandatory, to simply not contribute, i.e. defect [9,22], when not everyone commits. Yet, this avoidance strategy also removes the benefits for those who wished to establish the public good, hindering any advancements they could harness from this novel resource. Alternatively, one can try to establish boundaries on the common good so that only those who commit to make it work have access or that the benefit non-contributors can acquire from the common good is reduced, as is the case for food sharing, aid in social health and defence against predators [23–25]. An extreme case of exercising restriction is ostracism, which can be enforced through financial or social means [26,27]. Experimental studies with PGGs have shown that the threat of excluding or ostracizing non-cooperative members from the PGG can significantly increase contribution and cooperation [28,29]. As public goods are by definition non-excludable [8,30], ostracism may not be possible and non-committers may only be excluded to a certain degree. Moreover, a cost may be associated with this exclusion strategy, where the capacity to ostracize may be too costly. Evidence regarding restriction abounds in biological and social contexts: animals fence and defend territory and resources [25]. Trade restrictions against non-participating countries are widely implemented in international and environmental treaties [8,31], yet may be circumvented.
While showing the relevance of our prior conclusions on commitment obtained for the pairwise Prisoner's Dilemma (PD) [20] within the context of the more complex PGGs, we focus here on showing mathematically and numerically how best to deal with individuals that do not wish to make prior agreements and do not contribute to the common good. This issue is not only essential in the general discussion of the PGG, it is also fundamental in the case of the strategic commitment behaviour since we observed in the PD that the number of non-committers, together with those who free-ride on the investment of committers, increase markedly with the increase in the cost of setting up the commitment [20]. We will examine under which conditions avoidance, which is a generalization of the PD commitment behaviour towards the PGG, and restricting strategies are beneficial in the PGG, determining at the same time the conditions when the latter strategy is preferred over the former. The effect of the different parameters implicit to the strategies on their viability is carefully analysed. Interestingly, we will show that group size is an important factor in determining the conditions for which restriction may be better than simply avoiding non-committers.
2. Results
2.1. Commitment strategies in public goods game
Commitment strategies can propose a commitment deal to all members of the group before playing the PGG. The proposer(s) share the cost εP, while those who do not (but still can join the commitment) pay nothing. If all the requested co-players agree to the commitment, they are assumed to contribute to the public good. Those who commit though later do not contribute have to compensate the commitment proposers at their personal cost δ, and their compensation is shared among all proposers. Additionally, there may be group members who refuse the agreement, wishing to play the game without any prior commitment. As refusal may be conceived as a future defection (no contribution), proposers may wish to either avoid interacting with them or set up some mechanism that restricts their access to the public good. We define those strategies as follows:
— AVOID: refuses to play the game when there are non-committers in the group (hence, the PGG does not take place and each player receives 0 payoff).
— RESTRICT: sets up, at an extra cost εR, a mechanism to restrict the access of the non-committers to the public good. This restriction is modelled through a factor, ψ < 1, representing the fraction of the common resource the non-committers receive compared to the committed players (i.e. the smaller ψ the greater the effect).
Next to the traditional unconditional contributors (C, who always commit when being proposed a commitment deal, contribute whenever the PGG is played, but do not propose commitment) and unconditional non-contributors (D, who do not accept commitment, defect when the PGG is played and do not propose commitment), we consider two commitment free-riding strategies, which we have shown to become dominant under certain conditions in the pairwise PD situation [20]: (i) fake committers (FAKE), who accept a commitment proposal yet do not contribute whenever the PGG is played. These players assume that they can exploit the commitment proposing players without suffering the consequences; (ii) commitment free-riders (FREE), who defect unless being proposed a commitment, which they then accept and cooperate subsequently in the PGG. In other words, these players are willing to contribute when a commitment is proposed but are not prepared to pay the cost of setting it up.
We consider a well-mixed, finite population of a constant size Z, potentially composed of those five strategies, i.e. AVOID (or RESTRICT), C, FREE, D and FAKE. To simplify the notations, the strategies are numerated 1, 2, 3, 4 and 5, respectively (where 1 can either indicate the AVOID or RESTRICT strategy). In each interaction, N individuals are randomly selected from the population for playing the PGG. Among N randomly selected players, the number of players in the group of size N using strategy i is denoted by Ni, i = 1, … , 5, such that N = N1 + N2 + N3 + N4 + N5.
We compute the payoffs of either the AVOID or RESTRICT strategy in the population in relation to the other strategies (see section ‘Material and methods’ and electronic supplementary material). In the case of the AVOID strategy, if there is a D player in the group, i.e. N4 ≥ 1, then the game is not played and every player in the group obtains 0 (the results will be unchanged if the game is not optional, as in that case AVOID players contribute nothing to the public good). In the case of the RESTRICT strategy, the game is always played and RESTRICT players share an additional cost, εR > 0, to restrict the benefits D players can obtain from the public good. The committing player (RESTRICT, C, FREE and FAKE) and non-committing player (D) in the PGG gain, respectively (r(N1 + N2 + N3)/(N1 + N2 + N3 + ψN4 + N5))c and (r(N1 + N2 + N3)ψ/(N1 + N2 + N3 + ψN4 + N5))c. The gain for a RESTRICT player is reduced by c + (1/N1)εP − (N5/N1)δ, if N4 = 0, and by c + (1/N1)(εP + εR) − (N5/N1)δ, otherwise. The payoff for a C and a FREE player is reduced by c. Finally, the payoff for a FAKE player is reduced by δ. The detailed calculation of the payoff for each strategy as well as the payoff matrix is provided in the electronic supplementary material.
2.2. Constraints for viability of AVOID and RESTRICT
We derive the conditions for which commitment strategies, AVOID and RESTRICT, are evolutionary viable in a PGG, showing when they are risk-dominant (see Material and methods) against all defectors and free-riders (i.e. FAKE, FREE and D players). Yet more importantly, we determine when RESTRICT becomes more advantageous than AVOID; that is, when it is worthwhile to pay extra cost to invest in restriction technologies and infrastructure that limit the benefits of non-committers (D).
Equation (4.5) (see Material and methods) allows one to determine when AVOID and RESTRICT are risk-dominant against FREE. This occurs when
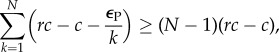 |
2.1 |
which can be simplified to
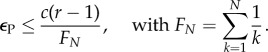 |
2.2 |
AVOID and RESTRICT are risk-dominant against FAKE if
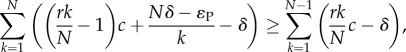 |
2.3 |
which can also be simplified to
| 2.4 |
Now, as we aim to examine when restriction works better than avoiding non-committers, we first examine independently when AVOID or RESTRICT are risk-dominant against D players. In the case of the AVOID strategy, this occurs when:
| 2.5 |
As equation (2.2) is more restrictive than equation (2.5), the two conditions in equations (2.2) and (2.4) define when AVOID is risk-dominant against all types of defectors and free-riders (see the electronic supplementary material for simplifications of these formulae using inequalities for FN). Both conditions can be understood intuitively. For a successful commitment, the cost of arranging the commitment needs to be justified with respect to the benefit of (mutual) cooperation (i.e. r(c − 1)), and a compensation needs to be arranged (see equation (2.4)) that is proportional to the player's contribution and the investment cost she paid for setting up the commitment.
This observation becomes clearer when looking at the transition probabilities and stationary distribution in a population of AVOID players with the other four strategies, as shown in figure 1a. Note the cycles from C to defection strategies (FREE, D and FAKE) and back to AVOID strategists, showing that defection strategies cannot completely be avoided in the PGG context (see also the electronic supplementary material, figure S1): when the cost of arranging commitment, εP, is sufficiently small, the population spends most of the time in the homogeneous state with AVOID players, regardless of the initial composition of the population (figure 1b). For low εP, nearly homogeneous AVOID populations are almost always reached for sufficiently large δ. More interestingly, this high frequency is not affected by changes in the compensation δ, once a certain threshold is reached. Accordingly, as for the PD [20], the arrangement cost is the essential parameter for the emergence and survival of AVOID and mutual cooperation. Additionally, we observe that for a variety of group sizes N, the region of εP wherein AVOID is a viable strategy increases (see the electronic supplementary material, figure S3, considering also the fact that the right-hand side of equation (2.2) is an increasing function of N): AVOID can handle the commitment free-riding strategies for a wider range of arrangement costs. Yet as the groups size increases, the frequency of AVOID, for similar small values of εP, decreases, revealing that other strategies may be necessary to cope with the increasing number of defectors in the groups and the population (electronic supplementary material, figure S3). These results provide a novel insight, when moving from the PD to the PGG, which is that avoiding defectors by refusing to play the game when someone does not agree to commit might lead to cooperation at higher arrangement costs, yet may in turn be detrimental for the overall level of cooperation in the game.
Figure 1.

(a) Stationary distribution and fixation probabilities. The population spends most of the time in the homogeneous state of AVOID. The black arrows identify the advantageous transitions, where ρN = 1/Z denotes the neutral fixation probability. The dashed lines denote neutral transitions. Note the cyclic pattern from cooperation to defection to commitment strategies and back. (b) Contour plot of the frequency of AVOID as a function of εP and δ. For a small enough cost of arranging the commitment, AVOID is abundant whenever a sufficient compensation is associated with the commitment deal. Parameters: N = 5, Z = 100, r = 3; β = 0.1; in panel (a), εP = 0.25, δ = 2.
In turn, RESTRICT is risk-dominant against D when
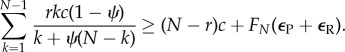 |
2.6 |
Because the left-hand side of equation (2.6) is a strictly decreasing function of ψ (see the electronic supplementary material), the necessary condition for RESTRICT to be risk-dominant against D is (i.e. when ψ = 0)
| 2.7 |
which is equivalent to
| 2.8 |
As the left-hand side of equation (2.6) is a continuous function of ψ, the satisfaction of equation (2.8) guarantees that, for any given εP and εR, there exists a threshold ψD such that RESTRICT is risk-dominant against D for any ψ below it. This restriction threshold could be interpreted as the organizational or technological advancement required to guarantee success against individuals that exploit the non-exclusive public good. Moreover, it specifies what the limit is on the cost, combining the restriction and the proposing costs, for this to work.
These observations are supported by figure 2a, in which we show the transition probabilities and stationary distribution in a population of RESTRICT players with other non-commitment proposing strategies. The main difference with figure 1a is the increase in the fixation probability from D to RESTRICT. This effect depends on the value of restriction factor ψ, as is shown in figure 2b. In general, the better the effect of restriction on non-committers (i.e. the smaller ψ), the higher the frequency of RESTRICT and cooperation in the long run, regardless of the initial composition of the population. These observations are robust against changes in the restriction εR, as seen in figure 2c, where we show the frequency of RESTRICT varying both ψ and εR. One can observe in both figures that ψ is the decisive parameter on the frequency of RESTRICT: to achieve significantly high frequency of RESTRICT, and as a consequence cooperation, a stringent restriction of non-committers must be possible. Even when costless restriction (i.e. εR = 0) is available, the frequency of RESTRICT decreases quickly when ψ approaches 1. Note that when ψ = 1, RESTRICT is never risk-dominant against D players, as can be seen from equation (2.6).
Figure 2.

(a) Transition probabilities and stationary distributions in case of RESTRICT. For an efficient restriction (εR = 0.5 and ψ = 0.25), the population spends most of the time in the homogeneous state of RESTRICT. Notations are the same as in figure 1a. (b) Frequencies of each strategy for varying ψ, in case of RESTRICT. For a given cost of restriction (εR = 0.5), in general the better the effect of restriction on non-committers (i.e. the smaller ψ), the greater the frequency of RESTRICT. (c) Frequency of RESTRICT as a function of εR and ψ, in a population with C, D, FREE and FAKE strategies. For a large range of cost for restricting the access of non-committers, εR, and the restriction, ψ, RESTRICT is highly frequent, having a higher frequency than AVOID. The double-stroke line corresponds to the part having the same frequency as AVOID (i.e. 0.64, with the same parameter values), and the area below this line identifies the area in which RESTRICT is more frequent than AVOID. In general, the larger εR, the smaller ψ is required for RESTRICT to be advantageous to AVOID. (d) Frequencies of each strategy as a function of the group size, N. RESTRICT becomes more frequent when the group size increases, even for a rather high cost of restriction (εR = 2.0). Parameters: in panels (a–c): N = 5; in all cases, Z = 100, r = 3; εP = 0.25, δ = 2; β = 0.1.
This notable success of the RESTRICT strategy in dealing with non-committers becomes even more significant when the group size increases (figure 2d). When the cost of restriction is extreme (e.g. εR = 2), D players dominate when the group size is small. But when the group size is sufficiently large, thereby reducing the individual cost of implementing the restriction, which is shared by the proposers, RESTRICT becomes dominant. The frequency of RESTRICT is even higher when εR is small (see already figure 3b). It is also interesting to note that the necessary condition for RESTRICT to be risk-dominant against D, as specified in equation (2.8), is simpler for larger N, since the right-hand side of the equation is an increasing function of N (see the electronic supplementary material).
Figure 3.

(a) Range of parameters ψ, εR and εP, generated from the analytical formula in equation (2.10), in which RESTRICT is better than AVOID. For a large range of cost for restricting, the access of non-committers, εR, and the effect of restriction, ψ, RESTRICT is better than AVOID. In general, the larger εR, the smaller ψ is required for RESTRICT to be advantageous to AVOID. (b) Group size is an important factor for making RESTRICT more viable than AVOID. We compute, as a function of the group size, N, the frequencies of RESTRICT for different values of restriction cost εR (the curves without markers), in comparison to the frequency of AVOID (the red curve with circled markers). In general, the lower the cost of restriction, the higher the frequency of RESTRICT. Also, the threshold of N above which RESTRICT is more frequent than AVOID is smaller. Parameters: in panel (b), Z = 100, εP = 0,25, ψ = 0.25, β = 0.1; in both panels, N = 5, r = 3.
2.3. What to do with the non-committers?
As there is no difference between AVOID and RESTRICT, except when playing D, one only needs to compare the stationary distribution of AVOID in a population with only D players against the stationary distribution of RESTRICT in a similar population. One can show that the frequency of RESTRICT is greater than AVOID if and only if the ratio of transition probabilities from D to RESTRICT and vice versa is greater than the ratio of transition probabilities from D to AVOID and vice versa, which can further be simplified, in the large population limit, to [32] (see the electronic supplementary material)
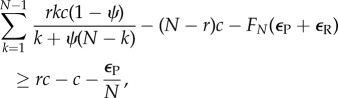 |
2.9 |
which is equivalent to
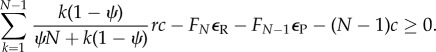 |
2.10 |
The left-hand side of the above equation is a decreasing function of ψ, εR and εP, but increasing in r (see proofs in the electronic supplementary material). Note that the cost εP still persists because RESTRICT players need to pay this cost when there are D players in the group, whereas AVOID players do not have to pay it as they refuse to play if D players are present in the group (thereby not arranging any commitment deal).
An observation that can be derived immediately from equation (2.10) is that, when ψ = 1, the equation is not satisfied. That is, unless restriction is possible, at least to some degree, it is better to refuse to play with those who explicitly do not agree to commit. Furthermore, with ψ = 0 we obtain the necessary condition for RESTRICT to be favoured over AVOID:
| 2.11 |
It means the cost of arranging the agreement and restricting the benefit of non-contributors needs to be justified with respect to the benefit of the PGG. Only in that case can restriction become justified over avoidance or non-participation. If this equation is not satisfied, AVOID is the better commitment strategy, however good the restriction of non-committers that can be brought about, including full exclusion or ostracism.
In addition, as the left-hand side of equation (2.10) is a continuous function of ψ, this equation guarantees that, for any given εP and εR, there exists a threshold ψAVOID which, for any ψ < ψAVOID, RESTRICT performs better than AVOID. This threshold moreover increases with r and can approach infinitely close to 1 when r tends to infinity, because, as mentioned above, the left-hand side of equation (2.10) is a decreasing function of ψ, but increasing in r (see the electronic supplementary material, figures S4 and S5). In addition, the equation provides the criteria for whether it is worthwhile to develop the restriction infrastructure or mechanism. As technological evolution may significantly (and unlimitedly) reduce the cost-to-impact ratio, for instance with respect to restriction mechanisms that require computing and communication power [33,34], one can postulate that under this condition RESTRICT will eventually be available and deployable, even when currently AVOID seems to be the best choice.
Figure 3a shows the region of parameters where RESTRICT is better than AVOID, generated from the analytical condition in equation (2.10). There is a large range of costs for restricting the access of non-committers, εR, and the effect of restriction, ψ, where RESTRICT is better than AVOID. Yet, if εR is too large, even a full exclusion (ostracism) does not lead to a better commitment strategy (as also can be seen from equation (2.11)). These analytical results are compatible with the numerical simulation results shown in figure 2c (see the electronic supplementary material, figure S6 for more comparisons).
In addition, because the right-hand side of equation (2.11) is a decreasing function of N, converging to 0 when N approaches infinity (see proofs in the electronic supplementary material), AVOID is less preferred for increasing N: as group size increases (figure 3b), the frequency of the strategy decreases. This indicates that avoiding non-committers in larger groups is less successful than avoiding them in smaller ones. Interestingly, RESTRICT seems to cope better with changes in group size: as group size increases, RESTRICT becomes more frequent than AVOID for a larger value of the restriction cost. When the cost of restriction is small (e.g. εR = 0.25), RESTRICT ensures a higher level of committers, and as a consequence contributors, in comparison to the AVOID strategy. Even when the restriction cost is high, there is a certain group size for which RESTRICT is more abundant than AVOID (figure 3b). Furthermore, as can be seen in electronic supplementary material, figures S4 and S5, when N increases, RESTRICT is the better strategy for strictly larger ranges of εP, εR and ψ, which becomes even more apparent for increasing values of the public good's multiplication factor r.
3. Discussion
We have shown that arranging prior commitment can lead to the evolution of cooperation in a PGG when the cost of arranging commitment is justified with respect to the benefit of cooperation. As such, this result generalizes the conclusions previously obtained for the PD [20], underlining again the evolutionary advantage of this capacity to make prior agreements in combination with the capacity to send signals and act accordingly [1,35]. Moreover, we show that even though the commitment strategies become viable for a wider range of the arrangement cost when moving from the PD to the PGG, and more generally, when the group size of the PGG increases, it might be detrimental for the overall level of cooperation when the cost of arrangement is low. Nonetheless, prior agreements remain more efficient in achieving cooperation when being compared with simple peer punishment [9,36] (see the electronic supplementary material, figure S7) in the PGG, an important result we observed also in case of the PD scenario [20].
Notwithstanding this efficiency with respect to punishment, individuals that do not accept an agreement (D) or individuals that free-ride on the investment that commitment proposers make (FREE) may increase in frequency within both the PD and PGG contexts as the cost of setting up the commitment increases. We examined here in detail how to deal with the former: either the commitment proposers can decide to avoid interacting with those non-committers (AVOID) or implement an infrastructure that allows them to restrict their access (RESTRICT), which corresponds to a reduction in the benefit they can obtain from the public good. For both strategies, it is assumed that an institution is present in order to enforce the compensation when someone in the commitment does not honour it. As such, both strategies are closely related to pool punishment models in terms of the presence of a third party required for the execution of the process [9,22]. Yet, AVOID and RESTRICT are also different from pool punishment as in the latter system no prior agreement on the posterior compensations is made. Additionally, the main difference between AVOID and RESTRICT, in terms of execution, is that, AVOID does not pay to set up the agreement when someone does not accept the agreement, whereas RESTRICT always initiates the agreement to play and pays an additional fee to the institution to reduce the access of the (explicit) non-committers.
We show here that for both strategies one can identify intuitive conditions, defined by costs and compensations, that lead to increased likelihood in receiving contributions for the common good. Furthermore, we have compared the AVOID and RESTRICT strategies and provided analytical conditions for when one mechanism is better than the other. Our results show that RESTRICT is better than AVOID if non-committers can be restricted to a certain degree, with a small enough cost. Otherwise, it is better to rely on AVOID, i.e. simply refuse to interact if there are non-committers in the group (or, if the interaction is mandatory, to not contribute when playing the PGG). The restriction effect ψ, which is defined as the reduction of the benefit of the non-committer, was shown to be a decisive factor. Interestingly, its threshold ψAVOID (i.e. for all ψ < ψAVOID, RESTRICT is better than AVOID) increases with r and can be infinitely close to 1. This indicates that for given costs and compensations of commitment, any restriction mechanism can be more advantageous than AVOID when the PGG is sufficiently beneficial.
Moving from pairwise to multi-player interactions requires an analysis of the role of the group size in relation to the viability of the commitment strategies. Indeed, as group size increases, RESTRICT becomes more viable than AVOID even when it is costlier to implement the restriction measure. Furthermore, it is so for a strictly larger range of the commitment costs and restriction factors, especially when the public good's producing factor (r) is high. As such, these results indicate that larger public goods with higher levels of contribution can be established once restriction becomes possible. The amount of restriction depends on the gain each participant gets from the public good: the lower the gain, the tougher the restriction needs to be.
These results differ from other observations related to the impact of group size on the level of cooperation in PGGs [37,38]: Lehmann et al. [37] showed that if the group size can be expanded stochastically, for instance, as a result of an increase in fecundity and/or a decrease in mortality, the kin-competition pressure induced by the limited dispersal in their networked model, can be significantly reduced, thereby favouring the evolution of cooperative behaviour. In a similar manner, Alizon & Taylor [38] showed that if the group size and compositions can be adapted over time, in a way that reduces the competition among relatives in a structured population (by allowing groups or patches with high fecundity rates to grow faster), the cooperation level is increased.
By contrast, the strategic mechanisms we examine here do not consider any forms of relatedness between group members or structured populations. The essential message is that for smaller groups one is better off to avoid individuals that do not wish to accept an agreement, prior to the game, when the cost of restriction is too large. Yet as group size increases, restriction mechanisms are more efficient in achieving cooperation, depending on the association with the restriction. As such, these results provide a completely new perspective on the role of group size on the level of cooperation. We envisage that AVOID and RESTRICT may also be more efficient in a structured population, because the free-riders can be avoided and excluded permanently by removing links with them. Furthermore, an introduction of relatedness among individuals may reduce the need for arranging commitments as it provides additional incentive to not free-ride. Both issues may be explored in subsequent papers.
The results presented here are in accordance with the outcomes of different behavioural commitment experiments [6,19,28,29]. High levels of cooperation were observed in a PGG experiment where a binding agreement, which was enabled through a prior communication stage among the members of the group, could be arranged before the PGG interaction occurred [6]. The experiment showed that whenever a commitment deal is not binding or not enforced, corresponding to a low compensation cost δ in our commitment models, defectors are widespread and the contribution level is low. Commitment can also take the form of a deposit–refund scheme [19], where those who agree to commit have to deposit an amount which will be refunded only if they fulfill the commitment and contribute to the public good. Similar to our results for the AVOID strategy, the most successful commitment strategy in that work was shown to be the one that refuses to set up the public good whenever there is a non-committer in the group (RESTRICT was not considered). The outcome of this deposit–refund experiment showed that when the deposit amount, corresponding to δ in our model, is sufficiently high, the contribution level is significant [19]. Note, however, that in both these experiments [6,19], the cost of setting up the commitment is always set to 0, thereby leading to effortless and effective commitment strategies. But as we have shown, this cost is the decisive factor for the viability of commitments strategies. This said, despite the fact that commitment has been shown experimentally to be a successful strategy for promoting cooperation in the PGG, our results further the understanding of the mechanism by identifying under which region of the parameters' values the mechanism works (as in the experiments) and when it does not. As a result, the outcome of our analysis suggests the need to study how varying the parameters would affect the outcomes of those commitment experiments.
Furthermore, PGG experiments where exclusion of disapproved members (for example, through voting) is allowed, exhibit a high level of contribution and commitment [28,29]. But therein exclusion is carried out after the PGG takes place, towards the observed non-contributors, as in the model of Sasaki & Uchida [39], which is different from our model where restriction occurs before the game takes place. This suggests that social exclusion or ostracism, even when it requires an additional cost and/or has a reduced effect in terms of the restriction, is an important mechanism for promoting group cooperation [26,29]. However, we envisage that exclusion imposed through arranging prior commitment as in this work may be more suitable in the case where there is rivalry in the game, as is the case for common-pool resource games [8], for in that case posterior restriction would not hinder the participating players in collecting their benefit.
Various extensions to the current model can be addressed. First, one can consider to move beyond the symmetric commitments, where the cost for arranging and managing the agreement is equally shared among the proposers. Asymmetric commitments, where the contribution to manage the agreement may depend on the wealth and the potential benefits of each member as in inequality models [40,41], may further increase the realism of the conclusions one can draw from these models. Moreover, when extending to the repeated interaction scenario [42], it is also natural to consider that those who benefited more from the previous interactions should contribute more to the management of the commitments. We envisage that these seemingly fairer ways of sharing the benefit and cost of commitment can elevate the willingness to commit and contribute. In this repeated interaction context, commitments can also be made incrementally, conditional on behaviours of others in the previous round of interaction; this option has been shown to promote a higher level of contribution in a repeated PGG experiment [43].
In summary, our results have demonstrated that arranging prior commitments provides an important pathway for the emerge of cooperation in the one-shot PGG, suggesting that good agreements make good friends [20] also in group interactions. Furthermore, always avoiding playing with those unwilling to commit is detrimental for the overall level of contribution, especially when interacting in large groups, and restriction towards those players might provide a better path to enhance the contribution level.
4. Material and methods
4.1. Population set-up and evolutionary dynamics
Both the analytical and numerical results obtained here use evolutionary game theory methods for finite populations [10,44,45]. In such a setting, individuals' payoff represents their fitness or social success, and evolutionary dynamics is shaped by social learning [10,46,47], whereby the most successful individuals will tend to be imitated more often by the others. In the current work, social learning is modelled using the so-called pairwise comparison rule [48], assuming that an individual A with fitness fA adopts the strategy of another individual B with fitness fB with probability given by the Fermi function, 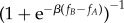 . The parameter β represents the ‘imitation strength’ or ‘intensity of selection’, i.e. how strongly the individuals base their decision to imitate on fitness comparison. For β = 0, we obtain the limit of neutral drift—the imitation decision is random. For large β, imitation becomes increasingly deterministic.
. The parameter β represents the ‘imitation strength’ or ‘intensity of selection’, i.e. how strongly the individuals base their decision to imitate on fitness comparison. For β = 0, we obtain the limit of neutral drift—the imitation decision is random. For large β, imitation becomes increasingly deterministic.
In the absence of mutations or exploration, the end states of evolution are inevitably monomorphic: once such a state is reached, it cannot be escaped through imitation. We thus further assume that, with a certain mutation probability, an individual switches randomly to a different strategy without imitating another individual. In the limit of small mutation rates, the behavioural dynamics can be conveniently described by a Markov chain, where each state represents a monomorphic population, whereas the transition probabilities are given by the fixation probability of a single mutant [9,45,49]. The resulting Markov chain has a stationary distribution, which characterizes the average time the population spends in each of these monomorphic end states.
In finite populations, the groups engaging in PGGs are given by multivariate hypergeometric sampling. For transition between two pure states (small mutation rate), this reduces to sampling (without replacement) from a hypergeometric distribution [9]. Denote
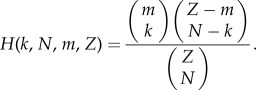 |
Let Πij(k) and Πji(k) denote the payoff of a strategists of type i and j, respectively, when the random sampling consists of k players of type i and N − k players of type j (as given in the payoff matrix in equation (1) in the electronic supplementary material). Hence, in a population of x i-strategists and (Z − x) j-strategists, the average payoffs to i- and j-strategists are [9,10]
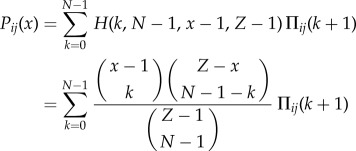 |
4.1 |
and
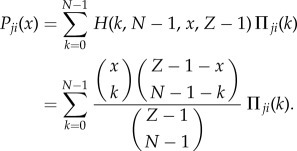 |
4.2 |
Note that several Pij(x) can be further simplified (see the electronic supplementary material). Now, the probability to change the number k of individuals using strategy i by ±1 in each time step can be written as [48]
| 4.3 |
The fixation probability of a single mutant with a strategy i in a population of (Z − 1) individuals using j is given by [45,48–50]
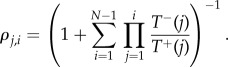 |
4.4 |
In the limit of neutral selection (i.e. β = 0), ρB,A equals the inverse of population size, 1/Z.
Considering a set {1, … , q} of different strategies, these fixation probabilities determine a transition matrix 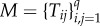 , with Tij,j≠i = ρji/(q − 1) and
, with Tij,j≠i = ρji/(q − 1) and 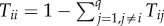 , of a Markov chain. The normalized eigenvector associated with the eigenvalue 1 of the transposed of M provides the stationary distribution described above [45,49,50], describing the relative time the population spends adopting each of the strategies.
, of a Markov chain. The normalized eigenvector associated with the eigenvalue 1 of the transposed of M provides the stationary distribution described above [45,49,50], describing the relative time the population spends adopting each of the strategies.
4.2. Risk-dominance condition
An important analytical criteria to determine the viability of a given strategy is whether it is risk-dominant with respect to other strategies [13,32]. Namely, one considers which selection direction is more probable: an i mutant fixating in a homogeneous population of individuals playing j or a j mutant fixating in a homogeneous population of individuals playing i. When the first is more likely than the latter, i is said to be risk-dominant against j [32], which holds for any intensity of selection and in the limit of large Z when
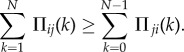 |
4.5 |
Supplementary Material
Funding statement
T.A.H. and T.L. acknowledges the support from the FWO Vlaanderen. T.L. also acknowledges the support of the Fonds de la Recherche Scientifique—FNRS.
References
- 1.Nesse RM. 2001. Evolution and the capacity for commitment. Russell Sage Foundation series on trust New York, NY: Russell Sage. [Google Scholar]
- 2.Frank RH. 2001. Cooperation through emotional commitment. In Evolution and the capacity for commitment (ed. Nesse RM.), pp. 55–76. New York, NY: Russell Sage. [Google Scholar]
- 3.Sosis R. 2000. Religion and intra-group cooperation: preliminary results of a comparative analysis of utopian communities. Cross Cult. Res. 34, 70–87. ( 10.1177/106939710003400105) [DOI] [Google Scholar]
- 4.Irons W. 2001. Religion as a hard-to-fake sign of commitment. In Evolution and the capacity for commitment (ed. Nesse RM.), pp. 292–309. New York, NY: Russell Sage. [Google Scholar]
- 5.Sterelny K. 2012. The evolved apprentice. Cambridge, MA: MIT Press. [Google Scholar]
- 6.Chen XP, Komorita SS. 1994. The effects of communication and commitment in a public goods social dilemma. Organ. Behav. Hum. Decis. Process. 60, 367–386. ( 10.1006/obhd.1994.1090) [DOI] [Google Scholar]
- 7.Han TA, Pereira LM, Santos FC. 2011. Intention recognition promotes the emergence of cooperation. Adapt. Behav. 19, 264–279. ( 10.1177/1059712311410896) [DOI] [Google Scholar]
- 8.Ostrom E. 1990. Governing the commons: the evolution of institutions for collective action. Cambridge, UK: Cambridge university press. [Google Scholar]
- 9.Hauert C, Traulsen A, Brandt H, Nowak MA, Sigmund K. 2007. Via freedom to coercion: the emergence of costly punishment. Science 316, 1905–1907. ( 10.1126/science.1141588) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sigmund K. 2010. The calculus of selfishness. Princeton, NJ: Princeton University Press. [Google Scholar]
- 11.Hardin G. 1968. The tragedy of the commons. Science 162, 1243–1248. ( 10.1126/science.162.3859.1243) [DOI] [PubMed] [Google Scholar]
- 12.West SA, Griffin AA, Gardner A. 2007. Evolutionary explanations for cooperation. Curr. Biol. 17, R661–R672. ( 10.1016/j.cub.2007.06.004) [DOI] [PubMed] [Google Scholar]
- 13.Nowak MA. 2006. Five rules for the evolution of cooperation. Science 314, 1560–1563. ( 10.1126/science.1133755) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hamilton WD. 1964. The genetical evolution of social behaviour. I. J. Theoret. Biol. 7, 1–16. ( 10.1016/0022-5193(64)90038-4) [DOI] [PubMed] [Google Scholar]
- 15.Axelrod R, Hamilton WD. 1981. The evolution of cooperation. Science 211, 1390–1396. ( 10.1126/science.7466396) [DOI] [PubMed] [Google Scholar]
- 16.Chuang JS, Rivoire O, Leibler S. 2009. Simpson's paradox in a synthetic microbial system. Science 323, 272–275. ( 10.1126/science.1166739) [DOI] [PubMed] [Google Scholar]
- 17.Xavier JB, Kim W, Foster KR. 2011. A molecular mechanism that stabilizes cooperative secretions in Pseudomonas aeruginosa. Mol. Microbiol. 79, 166–179. ( 10.1111/j.1365-2958.2010.07436.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.de Vargas Roditi L, Boyle KE, Xavier JB. 2013. Multilevel selection analysis of a microbial social trait. Mol. Syst. Biol. 9, 684 ( 10.1038/msb.2013.42) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cherry TL, McEvoy DM. 2013. Enforcing compliance with environmental agreements in the absence of strong institutions: an experimental analysis. Environ. Resour. Econ. 54, 63–77. ( 10.1007/s10640-012-9581-3) [DOI] [Google Scholar]
- 20.Han TA, Pereira LM, Santos FC, Lenaerts T. 2013. Good agreements make good friends. Sci. Rep. 3, 2695 ( 10.1038/srep02695) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Han TA. 2013. Intention recognition, commitments and their roles in the evolution of cooperation: from artificial intelligence techniques to evolutionary game theory models, vol. 9 SAPERE series Berlin, Germany: Springer. [Google Scholar]
- 22.Sigmund K, Silva HD, Traulsen A, Hauert C. 2010. Social learning promotes institutions for governing the commons. Nature 466, 7308 ( 10.1038/nature09203) [DOI] [PubMed] [Google Scholar]
- 23.Woodburn J. 1982. Egalitarian societies. Man 17, 431–451. ( 10.2307/2801707) [DOI] [Google Scholar]
- 24.Sugiyama LS. 2004. Illness, injury, and disability among Shiwiar forager–horticulturalists: implications of health-risk buffering for the evolution of human life history. Am. J. Phys. Anthropol. 123, 371–389. ( 10.1002/ajpa.10325) [DOI] [PubMed] [Google Scholar]
- 25.Hofmann HA, Schildberger K. 2001. Assessment of strength and willingness to fight during aggressive encounters in crickets. Anim. Behav. 62, 337–348. ( 10.1006/anbe.2001.1746) [DOI] [Google Scholar]
- 26.Williams KD. 2002. Ostracism: the power of silence. Emotions and social behavior. New York, NY: Guilford Publication. [Google Scholar]
- 27.Robertson TE, Delton AW, Klein SB, Cosmides L, Tooby J. 2014. Keeping the benefits of group cooperation: domain-specific responses to distinct causes of social exclusion. Evol. Hum. Behav. 35, 472–480. ( 10.1016/j.evolhumbehav.2014.06.006) [DOI] [Google Scholar]
- 28.Cinyabuguma M, Page T, Putterman L. 2005. Cooperation under the threat of expulsion in a public goods experiment. J. Public Econ. 89, 1421–1435. ( 10.1016/j.jpubeco.2004.05.011) [DOI] [Google Scholar]
- 29.Maier-Rigaud FP, Martinsson P, Staffiero G. 2010. Ostracism and the provision of a public good: experimental evidence. J. Econ. Behav. Organ. 73, 387–395. ( 10.1016/j.jebo.2009.11.001) [DOI] [Google Scholar]
- 30.Hummel JR, Lavoie D. 2007. National defense and the public-goods problem. In Anarchy and the law: the political economy of choice (ed. Stringham EP.), pp. 127–148. Oakland, CA: Independent Institute. [Google Scholar]
- 31.Barrett S. 2003. Environment and statecraft: the strategy of environmental treaty-making. Oxford, UK: Oxford University Press. [Google Scholar]
- 32.Gokhale CS, Traulsen A. 2010. Evolutionary games in the multiverse. Proc. Natl Acad. Sci. USA 107, 5500–5504. ( 10.1073/pnas.0912214107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Moore GE. 1965. Cramming more components onto integrated circuits. Electronics 38, 114–117. [Google Scholar]
- 34.Hilbert M, López P. 2011. The world's technological capacity to store, communicate, and compute information. Science 332, 60–65. ( 10.1126/science.1200970) [DOI] [PubMed] [Google Scholar]
- 35.Skyrms B. 2010. Signals: evolution, learning, and information. Oxford, UK: Oxford University Press. [Google Scholar]
- 36.Sigmund K, Hauert C, Nowak M. 2001. Reward and punishment. Proc. Natl Acad. Sci. USA 98, 10 757–10 762. ( 10.1073/pnas.161155698) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lehmann L, Perrin N, Rousset F. 2006. Population demography and the evolution of helping behaviors. Evolution 60, 1137–1151. ( 10.1111/j.0014-3820.2006.tb01193.x) [DOI] [PubMed] [Google Scholar]
- 38.Alizon S, Taylor P. 2008. Empty sites can promote altruistic behavior. Evolution 62, 1335–1344. ( 10.1111/j.1558-5646.2008.00369.x) [DOI] [PubMed] [Google Scholar]
- 39.Sasaki T, Uchida S. 2013. The evolution of cooperation by social exclusion. Proc. R. Soc. B 280, 20122498 ( 10.1098/rspb.2012.2498) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tavoni A, Dannenberg A, Kallis G, Löschel A. 2011. Inequality, communication and the avoidance of disastrous climate change in a public goods game. Proc. Natl Acad. Sci. USA 108, 11 825–11 829. ( 10.1073/pnas.1102493108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Vasconcelos VV, Santos FC, Pacheco JM, Levin SA. 2014. Climate policies under wealth inequality. Proc. Natl Acad. Sci. USA 111, 2212–2216. ( 10.1073/pnas.1323479111) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Van Segbroeck S, Pacheco JM, Lenaerts T, Santos FC. 2012. Emergence of fairness in repeated group interactions. Phys. Rev. Lett. 108, 158104 ( 10.1103/PhysRevLett.108.158104) [DOI] [PubMed] [Google Scholar]
- 43.Kurzban R, McCabe K, Smith VL, Wilson BJ. 2001. Incremental commitment and reciprocity in a real-time public goods game. Pers. Soc. Psychol. Bull. 27, 1662–1673. ( 10.1177/01461672012712009) [DOI] [Google Scholar]
- 44.Nowak MA, Sasaki A, Taylor C, Fudenberg D. 2004. Emergence of cooperation and evolutionary stability in finite populations. Nature 428, 646–650. ( 10.1038/nature02414) [DOI] [PubMed] [Google Scholar]
- 45.Imhof LA, Fudenberg D, Nowak MA. 2005. Evolutionary cycles of cooperation and defection. Proc. Natl Acad. Sci. USA 102, 10 797–10 800. ( 10.1073/pnas.0502589102) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hofbauer J, Sigmund K. 1998. Evolutionary games and population dynamics. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 47.Rendell L, et al. 2010. Why copy others? Insights from the social learning strategies tournament. Science 328, 208–213. ( 10.1126/science.1184719) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Traulsen A, Nowak MA, Pacheco JM. 2006. Stochastic dynamics of invasion and fixation. Phys. Rev. E 74, 11909 ( 10.1103/PhysRevE.74.011909) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fudenberg D, Imhof LA. 2005. Imitation processes with small mutations. J. Econ. Theory 131, 251–262. ( 10.1016/j.jet.2005.04.006) [DOI] [Google Scholar]
- 50.Karlin S, Taylor HE. 1975. A first course in stochastic processes. New York, NY: Academic Press. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.