

Significance
The history of oxygen at Earth’s surface is intimately tied to its production by oxygenic photosynthesis, whereby plants, algae, and cyanobacteria release O2 as a waste product. Despite this metabolism’s profound importance, its evolutionary timing is poorly understood. Studies have increasingly revealed a temporal disconnect between evidence for the presence of O2 during weathering as early as 3.0 billion years ago and its atmospheric accumulation 500 million years later. We review this problem and numerically demonstrate that local O2 production and immediate consumption in surface-bound (benthic) microbial ecosystems at profound disequilibrium conditions is the most parsimonious explanation for this delay. Thus, emergence of continental landmass was likely a crucial factor in the earliest oxygenation of Earth’s surface environment.
Keywords: oxygenic photosynthesis, Great Oxidation Event, oxidative weathering, Precambrian
Abstract
The Great Oxidation Event (GOE) is currently viewed as a protracted process during which atmospheric oxygen increased above ∼10−5 times the present atmospheric level (PAL). This threshold represents an estimated upper limit for sulfur isotope mass-independent fractionation (S-MIF), an Archean signature of atmospheric anoxia that begins to disappear from the rock record at 2.45 Ga. However, an increasing number of papers have suggested that the timing for oxidative continental weathering, and by conventional thinking the onset of atmospheric oxygenation, was hundreds of million years earlier than previously thought despite the presence of S-MIF. We suggest that this apparent discrepancy can be resolved by the earliest oxidative-weathering reactions occurring in benthic and soil environments at profound redox disequilibrium with the atmosphere, such as biological soil crusts and freshwater microbial mats covering riverbed, lacustrine, and estuarine sediments. We calculate that oxygenic photosynthesis in these millimeter-thick ecosystems provides sufficient oxidizing equivalents to mobilize sulfate and redox-sensitive trace metals from land to the oceans while the atmosphere itself remained anoxic with its attendant S-MIF signature. As continental freeboard increased significantly between 3.0 and 2.5 Ga, the chemical and isotopic signatures of benthic oxidative weathering would have become more globally significant from a mass-balance perspective. These observations help reconcile evidence for pre-GOE oxidative weathering with the history of atmospheric chemistry, and support the plausible antiquity of a terrestrial biosphere populated by cyanobacteria well before the GOE.
A remarkably coherent ensemble of evidence points to a significant accumulation of atmospheric oxygen for the first time in Earth’s history beginning circa (ca.) 2.45 Ga, the so-called Great Oxidation Event (GOE). In brief, this includes the disappearance of detrital pyrite, uraninite and siderite from fluvial and deltaic deposits (1), an increase in the retention of iron in paleosols (2), an enrichment of Cr and U in iron formations (3, 4), and the disappearance of sedimentary sulfur isotope mass-independent anomalies indicative of atmospheric SO2 processing in the absence of appreciable ozone (5). These gross observations have been complemented by emerging trace element and isotopic proxies whose sensitivity to oxidative cycling now appears to extend evidence for such processes hundreds of millions of years prior. In 2.7–2.6 Gy old black shales, enrichment in Mo, Re, and Os, fractionation of Fe and Mo isotopes, and C isotope data indicating methanotrophy all suggest that O2-rich niches already existed at that time (6–10). Stüeken et al. (11) suggested that an increase in the total S and Mo supply to marginal marine sediments at 2.8 Ga is best explained by the biological oxidation of crustal sulfide minerals. Most recently, Mo and Cr isotope compositions, as well as U enrichment data, from a banded iron formation of the Pongola Supergroup appear to reflect some partial oxygenation at ca. 3.0 Ga (12, 13). Collectively these results imply that oxidative photosynthesis had evolved hundreds of millions of years before the GOE.
The superposition of pre-GOE signals for oxidative weathering with the abovementioned gross observations of a generally anoxic atmosphere represents a conundrum for which two broad and nonexclusive models have been proposed: (i) that pre-GOE oxidative weathering is the result of transient oxygenation events driven by “oxygen oases” in the marine realm (14–18); and (ii) that oxidative weathering was driven by atmospheric O2 present at concentrations below 10−5 present atmospheric level (PAL). Recent studies have bolstered the possibility of both operating on the pre-GEO Earth (12, 19); however, several conceptual difficulties remain. These include: the short lifetime of atmospheric O2 as a trace gas under a reducing atmosphere (20), the strong discrepancy between oxidative dissolution and burial timescales of redox-sensitive minerals in the marine realm (21), and a general lack of Fe retention in pre-GOE paleosols that argues against sustained diffusion of atmospheric O2 into the soil (2). We propose here an alternative model that appears to have been overlooked in many discussions of pre-GOE oxidative weathering, yet is highly attractive on mechanistic and quantitative grounds, that being intense O2 generation—and immediate consumption—at submeter scales by benthic oxygenic photosynthesis in the terrestrial realm, regardless of atmospheric O2 concentrations. (Benthic is used throughout the manuscript sensu amplo to refer to microorganisms living in or attached to sediments, rocks, soils, and other natural solid substrates.)
Benthic Oxygenic Photosynthesis Past and Present
Nowhere on the modern Earth is free O2 found in greater excess than in benthic photosynthetic ecosystems and soils. A summary of ∼6,200 Eh–pH field measurements across diverse environments reveals that soils, mine waters, fresh waters and their associated sediments, and shallow ground waters dominate the oxidizing end of the Earth’s surface redox spectrum (22). These authors aptly related this, in part, to microbial activity and that of cyanobacteria in particular. Gross oxygenic photosynthetic activity is now understood to be 1,000–10,000 fold greater by volume in benthic communities relative to pelagic ones (23); in other words, 1 cm of microbial mat may produce oxygen at a rate comparable in the pelagic realm to a 100-m-deep water column (24). Similarly, because benthic microbes can cover all submerged surfaces and voids, they often exceed their pelagic counterparts in terms of biomass (25). Oxygenic photosynthesis by benthic and soil microbial communities is widespread across a vast range of terrestrial and marginal marine environments, with activity of the photosynthetic apparatus itself being largely governed by three key factors: temperature, CO2 concentration, and irradiation. Optimization of all three factors yields remarkably comparable maximal rates of benthic oxygenic photosynthetic across diverse natural environments (Fig. 1). From our literature survey of 84 in situ measurements of oxygenic photosynthesis in benthic and soil microbial ecosystems, depth-integrated net rates (O2 produced – O2 immediately consumed) are log-normally distributed and generally fall within two orders of magnitude (100 – 10−2 nmol cm−2 s−1) regardless of the benthic environment (Fig. 1). With this range in mind, we adopt the median net O2 production rate of our compilation (0.16 nmol cm−2 s−1) for further consideration below.
Fig. 1.

A survey of daytime in situ O2 production rates measured for natural benthic microbial biomass across diverse environments (n = 84). All production rates are net, representing the surplus O2 generated after competing O2-consuming processes (primarily aerobic respiration) are accounted for. See Table S1 for compilation data and references.
Despite the absence of a UV-protective ozone layer in the Archean, a terrestrial phototrophic biosphere may have existed in various sheltered environments. Before plant evolution in the Phanerozoic, and with fresh volcanic material to act as fertile soil, biological soil crusts (BSCs) likely colonized much of the exposed land surfaces (26, 27). Indeed, element mobility patterns in several paleosols suggest the presence of an ancient terrestrial biosphere where organic ligands chelated metals during weathering (28–30). Of particular interest are the 2.76-Ga Mount Roe paleosol in Western Australia and the 2.96-Ga Nsuze paleosol in South Africa. In the Mount Roe, δ13Corganic values as low as −51‰ Pee Dee belemnite (PDB) suggest that methanotrophic bacteria once inhabited these soils; their presence would seemingly indicate that some cyanobacterially produced O2 was locally available as these microbes either directly use O2 or they use substrates (e.g., NO3−, SO42−) that themselves required an oxidative process, i.e., NH3 + O2 → NO3− or H2S + O2 → SO42− (28). In the Nsuze paleosol, substantial enrichments in U relative to the typically cohosted but nonredox sensitive refractory elements Th and Zr suggest oxidative remobilization of U (12), presumably from more oxidizing upper parts of the paleosol that were not preserved. Importantly, it is generally accepted that deep soils could not have formed in the Archean abiotically because the absence of stabilizing BSCs would have fostered high rates of erosion, and thus, the presence of thick paleosols alone must signify the evolution of land microbiota (25). The age of these paleosols is consistent with molecular clock estimates for the colonization of land between 3.05 and 2.78 Ga based on the shared properties of pigment synthesis and resistance to dehydration exhibited by extant terrestrial microbiota (31), keeping in mind that such estimates are subject to considerable uncertainty (32).
In the absence of plant root systems, Archean surface terrains would also have been subject to rapid migration of riverbeds with a predisposition for wide, braided streams to transport and disperse the large volumes of supermature quartz-rich sands weathered from the source terrains (33). Tidal range may have been larger as well (34), with the net result being enhanced wetting of the emerged continental surface as a whole, conditions favoring colonization by freshwater microbial mats (FMM). Stromatolites in lacustrine sediments of the 2.72-Ga Tumbiana Formation, Western Australia are characterized by δ13Corganic values as low as −60‰ PDB, yet a narrow range in δ34Spyrite (35), a combination indicative of aerobic rather than anaerobic methanotrophy. The morphogenetic sequence of millimeter-scale clumps, reticulate ridge and laminated cones, has also been used implicate the ancient presence of O2-generating cyanobacteria in the formation of Tumbiana microbialites (36–38). Similar features exist in the 2.7-Ga Sodium Group, South Africa (39). The presence of microbially induced sedimentary structures (MISS) further supports the notion that FMM that likely hosted cyanobacteria were widespread within Neoarchean alluvial, interfluve, and lacustrine environments (40). Although those environments notoriously have poor preservation potential, their tidal flat counterparts are much more widely represented in the rock record, and in these settings, the MISS record extends as far back as ca. 3.2 Ga (41). Phylogenetic (42) and microfossil evidence (43) both may support the presence of benthic cyanobacteria in Mesoarchean FMM, in accordance with the geochemical evidence for oxidative weathering highlighted above. Before the evolution of grazers, BSC and FMM should have thrived under reduced selection pressure, a corollary that the Archean rock record certainly appears to support. Despite significant uncertainties surrounding the geochemical, morphological, and phylogenetic evidence for cyanobacteria before the GOE, as we demonstrate below the potential geochemical impact of their benthic expression, in strong disconnection from the atmosphere, is unquestionable.
The high degree of compartmentalization provided by BSCs and FMMs in isolating substrates from bulk surface conditions means that the weathering reactions that take place at the underlying rock surfaces, or within the mineral-bearing soil and sediment horizons, are out of equilibrium with the atmosphere. For instance, oxygen levels in BSCs may be orders of magnitude greater than in the overlying air. Garcia-Pichel and Belnap (44) demonstrated strong vertical gradients that resulted in O2 supersaturation within the top 200–400 μm of the crust surface, with low to negligible concentrations within just a few millimeters above and below the surface. In other words, as today, there is no reason to assume that the gas concentrations at weathering interfaces are directly reflective of atmospheric conditions. Indeed, 1D modeling with oxygen and sulfate boundary conditions encompassing those hypothesized from the Archean to present indicates that oxygen production and redox zonation within ancient microbial mats should have been independent of the chemistry of their environment (45). It is important to note that the attendant effects of BSCs and FMMs in the Archean would have been multiple, and would also have included enhanced mineral dissolution of underlying sediments (46) and increased flux of dissolved organic carbon to overlying waters, which in turn, facilitates transport of normally insoluble metals (e.g., Fe, Al) downstream (47).
Benthic Oxygen Oases before the GOE: Mechanistic and Numerical Considerations
We now turn to the O2 production potential of BSCs and FMMs, and implications for oxidative weathering before atmospheric oxidation. We first emphasize the crucial distinction between the quantities of oxygen tied up in major Earth surface reservoirs, for example the current atmospheric O2 inventory or the oxidizing equivalents currently contained in the upper continental crust, with the quantities of oxygen implicated in modern fluxes such as oxidative weathering, burial of reduced C, S, and Fe, and volcanic degassing (Fig. 2A). From this figure, it is clear how sustained imbalances in the latter can drive the former over timescales on the order of 105–108 y, and it is through this framework that the initial rise in atmospheric oxygen should be considered. Crucially, profound rearrangement of O2 sources and sinks, for example O2 production upon the evolution of oxygenic photosynthesis accompanied by immediate oxidative continental weathering and rereduction of the reaction products (see below), can be achieved with a net zero change in gross redox balance. Thus, such changes can operate on temporal and quantitative scales that should be considered independent of their role in the sustained redox imbalance (48–50) required by the GOE. In the specific case of benthic photosynthesis driving oxidative weathering, the O2 produced is confined to a diffusion-limited environment where even today, reaction with reductants below the uppermost zone of oxygen production is more important than O2 escape (i.e., net production << gross production) (51, 52). We focus on net production to conservatively reflect the amount of O2 that would be available to participate in reactions beyond those already consuming O2 in modern benthic ecosystems. This metric is additionally conservative in that aerobic microbial oxidation of sulfide in modern mats is not included in net production despite generating the same weathering signals under question, and in the sulfate-poor conditions of the Archean (53, 54), phototrophic/chemolithoautotrophic sulfur oxidation would have been inhibited, leading to enhanced net O2 production (45). Despite the term “net O2 production,” no gaseous O2 escape is necessarily implied in our model; rather we simply consider this net O2 as being available for further reaction with nearby reductants. Before atmospheric oxygenation, the fate of all biologically produced O2 should be rapid consumption by oxidative processes.
Fig. 2.
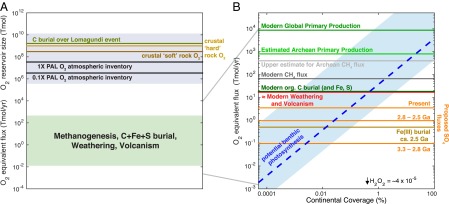
(A) Comparison of annual fluxes of principal O2 sources and sinks in the pre- and post-GOE Earth system (lower green box) with the size of major O2 reservoirs pertaining to the GOE itself (upper blue box). All values are in Tmol or Tmol/y O2 equivalents. This diagrammatically indicates the magnitude of the disconnect between the annual O2 fluxes associated with surficial redox processes (including modern oxidative continental weathering) and the amount of O2 equivalents implicated in the GOE itself. (B) Close up of A depicting annual O2 source and sink fluxes, in terms of O2 equivalents, contributing to Earth’s surface redox balance. Assuming the median diurnal benthic O2 production rate from the compilation presented in Fig. 2 (0.16 nmol cm−2 s−1), the potential net O2 production rate by terrestrial microbial ecosystems is displayed as a function of percent modern continental surface coverage as a blue dashed line (blue area encompasses 90% of compiled rates). See Table S2 for data compilation and sources; see also Claire et al. (48), Kasting (49), and Catling (50) for a detailed treatment. Only small degrees of benthic photosynthetic coverage are required to account for the earliest signals of oxidative weathering, highlighting a strong but previously unrecognized potential sensitivity in these signals to the evolution of terrestrial oxygenic photosynthesis. It is important to note that these signals may be generated with no net redox imbalance and thus independent of atmospheric oxygenation (see main text).
The soluble oxidation products of such reactions (e.g., sulfate, nitrate, metal oxyanions) are themselves most likely to be rereduced in short order, but they may also escape their immediate environment and eventually be transported to a marine basin and reduced there instead. In either case, this rereduction step yields reduced weathering products [e.g., S(-II)] that balance the O2 originally produced and immediately consumed by oxidative weathering close to the site of oxygen production. This is equivalent to the immediate rereaction of O2 and organic matter produced by oxygenic photosynthesis, but with aqueous intermediates (oxidative weathering products) shuttling oxidizing power from localized O2 sources to fuel the respiration of organic matter produced elsewhere—both entail no net redox effect. Regardless of whether the transport of oxidative weathering products, in either oxidized or rereduced form, occurs over only millimeters or over catchment scales to marine basins, the elemental or isotopic fractionation signals testifying to their oxidative history are nonetheless generated. In the case of chalcophile trace elements (e.g., Mo, Re), their sourcing to marine basins has been attributed to the oxidation of crustal sulfides (8, 17), a process shown to be likely insignificant in a marine photic zone “oxygen oasis” scenario considering the timescales of burial vs. oxidation of detrital crustal sulfide grains, but feasible in contact with benthic ecosystems in continental settings due to longer transport and exposure times (21). Considering the short lifetimes gaseous O2 likely experienced under CH4-rich atmospheric conditions (20), highly localized oxygen production by benthic photosynthesis represents the logical explanation for their initial liberation in surface continental settings, concomitant with the SO42- from the crustal sulfides that hosted them.
Although reconstruction of oxidative sulfide weathering from sedimentary trace element enrichments is in its infancy, significantly more data exists for the SO42- that would have accompanied them. We use the recent reconstruction of evolving continental SO42- fluxes from Stüeken et al. (11) to evaluate from a quantitative perspective to what extent benthic oxygenic photosynthesis is capable of supplying the necessary oxidative power to account for those fluxes (Fig. 2B). Shown for comparison are modern global primary production (close to 103 times the carbon buried annually), of which ∼50% is terrestrial (55), estimated Archean global primary production (∼1/10 the modern value), the modern methane flux, and an upper estimate for the Archean methane flux (see Table S2 for values and data sources)—the latter two contributing to global redox by their positive effect on hydrogen escape (48–50, 56). Using the median areal benthic net O2 production rate from our compilation (Fig. 1), we calculate the potential generation of oxidative equivalents by benthic oxygenic photosynthesis for the modern continental surface area, as a function of percent surface coverage (Fig. 2B, blue dashed line, where potential O2 production is calculated as the median areal production rate times continental coverage; light blue area encompasses 90% of compiled rates). It is immediately apparent that only small degrees of continental coverage are required to account for evolving pre-GOE continental sulfur fluxes. In fact, we calculate that the modern continental oxidative weathering and volcanic O2 sink (red line) is balanced by benthic oxygenic photosynthesis at a modern continental surface coverage of only 0.5%. The sensitivity of sulfur-related redox proxies to benthic oxidative photosynthesis on land thus appears to be quite high, with only small degrees of coverage required to account for the entirety of modern continental oxidative weathering. As long as the SO42- generated by oxidative weathering after benthic photosynthesis is ultimately rereduced by respiration of organic matter, no net redox change in the Earth system is implied. We emphasize that oxidative weathering driven by pre-GOE benthic photosynthesis can be considered independently, on both mass balance and temporal bases, from the gross redox imbalances implicated in the oxygenation of the ocean–atmosphere system ca. 2.5–2.3 Ga, whatever they may have been (cf. refs. 48–50).
From this perspective, and in the absence of robust biomarker evidence (57), the timing of the evolution of oxygenic photosynthesis may be constrained by when the threshold of oxidants consumed (e.g., as indicated by S fluxes) surpassed the amount plausibly supplied by alternative abiotic mechanisms. These mechanisms include UV photooxidation of Fe(II) and photochemical H2O2 formation. The upper limit of the former may approximated by Fe(III) burial ca. 2.5 Ga, and the latter appears effectively negligible (58, 59) (brown line and black text; Fig. 2B). As both anoxygenic photosynthetic (60) and abiotic (61) Fe(II) oxidation mechanisms remain plausible, additional constraints on Fe(II) oxidation pathways in the Archean are required to pursue this question further. Nonetheless, our sensitivity analysis indicates that as soon as cyanobacteria evolved and were able to colonize significant continental landmass, sedimentary geochemical signals of their activity above the background of inorganic oxidants should have been immediate. Considering inferred Archean continental sulfur fluxes (11), this occurred before 2.8 Ga, and with a degree of surface coverage as little as 0.01% of the modern continental landmass. A global shift in multiple sulfur isotope systematics ca. 2.7 Ga indicate that biological oxygen fluxes to the atmosphere may have began outpacing the methane flux around this time (62).
Oxidative Weathering in the Context of Archean Earth System Change
It is apparent that localized O2 production within the confines of ancient biological communities on land could have generated sufficient oxidizing power to mobilize trace metals and SO42- from sulfide-bearing minerals in underlying substrata without affecting the redox balance of the ocean–atmosphere system before the GOE. Although intuitive in the face of the hyperoxic state of modern benthic photosynthetic ecosystems, this finding is at significant odds with the contention that Earth’s earliest free O2 was generated in open marine oxygen oases and transported in gas phase over considerable distances (Fig. 3A). If cyanobacteria were conceivably metabolizing at modern rates on land by perhaps 3.0 Ga, an intriguing question that follows is what happened in the hundreds of million years between the first, rare signals of oxidative weathering and the first accumulation of atmospheric oxygen?
Fig. 3.

Schematic of (A) pre-GOE oxygen oases occurring primarily in the open marine realm, implying atmospheric transport of O2 (blue whiffs), versus (B) localized oxygen production and oxidative weathering by benthic oxygenic photosynthesis on the continents before the GOE. Although atmospheric O2 “whiffs” stemming from marine oxygen oases may have been important during periods of depressed atmospheric sinks for O2 (19), such as during the GOE itself, benthic pre-GOE oxygen oases would have operated efficiently independent of atmospheric conditions.
The evolution of emerged landmass and its microbial colonization likely played a strong role in the frequency and amplitude of signals for oxidative weathering in the run-up to the GOE. Kump and Barley (63) previously suggested that the timing of the GOE was due to a shift from submarine volcanism to subaerial volcanism coincident with continental crust stabilization, which led to a reduced sink for oxygen. Ancient cratons preserving signals of pre-GOE oxidative weathering appear to have stabilized ca. 3.0–2.5 Ga: (i) the Pilbara and Kaapvaal cratons in Western Australia and South Africa, respectively, achieved tectonic stability during a period of mantle plume activity between 3.0–2.9 Ga, and (ii) the majority of Archaean cratons achieved long-term tectonic stability as elevated continental land masses between 2.7 and 2.5 Ga, after a period of mantle plume activity (2.72–2.66 Ga) (64, 65). Although diverse models exist for the cumulative history of continental crust (see review by ref. 66, their figure 1), it is continental freeboard that would have determined the potential extent of benthic cyanobacterial colonization as well as the composition of volcanic gases in the Kump and Barley (63) model. Continental freeboard depends more on mantle temperature and continental hypsometry (also temperature dependent) than on the growth of continental crust; recent models suggest a continental landmass that was effectively flooded before 3.5 Ga, emerged to cover ∼1–2% of Earth’s surface by ca. 3.0 Ga, and then expanded to ∼5–8% ca. 2.5–2.0 Ga (67). Tracing this trajectory in Fig. 2B shows that benthic O2 production readily accounts for evolving pre-GOE sulfur fluxes, even if both areal coverage and photosynthetic efficiency were severely depressed. It has long been recognized that several factors may have depressed areal coverage or photosynthetic efficiency of cyanobacteria, and thus masked their potential presence, before the GOE (68), including higher clastic fluxes to shallow water environments before the establishment of extensive carbonate platforms, the absence of an UV-attenuating ozone layer, and a lack of genetic diversity that would enable niche expansion. Although the exact confluence of factors controlling the success of Earth’s earliest oxygenic phototrophs remains an open question, it is clear that Earth’s earliest signals for the presence of free O2 may not only have been strongly decoupled from atmospheric oxygenation itself, but were also likely less tied to the consumption of Archean reductants (i.e., gases, solutes) than to the simple problem of having available and colonizable surface area for oxidative weathering. Future efforts coupling a freeboard-tuned mass balance approach with additional constraints on sulfur and redox-sensitive trace metal fluxes before the GOE should help reconcile the role of emerging continents and their microbial colonizers in Earth’s earliest oxidative weathering.
Materials and Methods
Literature sources used for the compilation of benthic microbial net oxidative photosynthesis rates are tabulated with Table S1. For the reservoirs and fluxes of oxidizing equivalents presented in Fig. 2, values and literatures sources are tabulated in Table S2; see Claire et al. (48), Kasting (49), and Catling (50) for detailed treatments.
Supplementary Material
Acknowledgments
S.V.L. and K.O.K. acknowledge financial support from LabexMER and National Sciences and Engineering Research Council of Canada, respectively, as well as stimulating exchanges with Christopher Reinhard, Noah Planavsky, Ernesto Pecoits, and Pierre Sans Jofre.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1415718112/-/DCSupplemental.
References
- 1.Rasmussen B, Buick R. Redox state of the Archean atmosphere: Evidence from detrital heavy minerals in ca. 3250–2750 Ma sandstones from the Pilbara Craton, Australia. Geology. 1999;27:115–118. [Google Scholar]
- 2.Rye R, Holland HD. Paleosols and the evolution of atmospheric oxygen: A critical review. Am J Sci. 1998;298(8):621–672. doi: 10.2475/ajs.298.8.621. [DOI] [PubMed] [Google Scholar]
- 3.Konhauser KO, et al. Aerobic bacterial pyrite oxidation and acid rock drainage during the Great Oxidation Event. Nature. 2011;478(7369):369–373. doi: 10.1038/nature10511. [DOI] [PubMed] [Google Scholar]
- 4.Partin CA, et al. Uranium in iron formations and the rise of atmospheric oxygen. Chem Geol. 2013;362:82–90. [Google Scholar]
- 5.Farquhar J, Bao H, Thiemens M. Atmospheric influence of Earth’s earliest sulfur cycle. Science. 2000;289(5480):756–759. doi: 10.1126/science.289.5480.756. [DOI] [PubMed] [Google Scholar]
- 6.Siebert C, Kramers JD, Meisel T, Morel P, Nägler TF. PGE, Re-Os, and Mo isotope systematics in Archean and early Proterozoic sedimentary systems as proxies for redox conditions of the early Earth. Geochim Cosmochim Acta. 2005;69:1787–1801. [Google Scholar]
- 7.Wille M, et al. Evidence for a gradual rise of oxygen between 2.6 and 2.5 Ga from Mo isotopes and Re-PGE signatures in shales. Geochim Cosmochim Acta. 2007;71:2417–2435. [Google Scholar]
- 8.Kendall B, et al. Pervasive oxygenation along late Archaean ocean margins. Nat Geosci. 2010;3:647–652. [Google Scholar]
- 9.Czaja AD, et al. Evidence for free oxygen in the Neoarchean ocean based on coupled iron–molybdenum isotope fractionation. Geochim Cosmochim Acta. 2012;86:118–137. [Google Scholar]
- 10.Thomazo C, Nisbet EG, Grassineau NV, Peters M, Strauss H. Multiple sulfur and carbon isotope composition of sediments from the Belingwe Greenstone Belt (Zimbabwe): A biogenic methane regulation on mass independent fractionation of sulfur during the Neoarchean? Geochim Cosmochim Acta. 2013;121:120–138. [Google Scholar]
- 11.Stüeken EE, Catling DC, Buick R. Contributions to late Archaean sulphur cycling by life on land. Nat Geosci. 2012;5:722–725. [Google Scholar]
- 12.Crowe SA, et al. Atmospheric oxygenation three billion years ago. Nature. 2013;501(7468):535–538. doi: 10.1038/nature12426. [DOI] [PubMed] [Google Scholar]
- 13.Planavsky NJ, et al. Evidence for oxygenic photosynthesis half a billion years before the Great Oxidation Event. Nat Geosci. 2014;7:283–286. [Google Scholar]
- 14.Macgregor AM. The problem of the pre-Cambrian atmosphere. S Afr J Sci. 1927;24:155–172. [Google Scholar]
- 15.Cloud PE., Jr Significance of the Gunflint (Precambrian) Microflora: Photosynthetic oxygen may have had important local effects before becoming a major atmospheric gas. Science. 1965;148(3666):27–35. doi: 10.1126/science.148.3666.27. [DOI] [PubMed] [Google Scholar]
- 16.Fischer AG. NAS symposium on the evolution of the Earth's atmosphere: Fossils, early life, and atmospheric history. Proc Natl Acad Sci USA. 1965;53:1205–1215. [Google Scholar]
- 17.Anbar AD, et al. A whiff of oxygen before the great oxidation event? Science. 2007;317(5846):1903–1906. doi: 10.1126/science.1140325. [DOI] [PubMed] [Google Scholar]
- 18.Kaufman AJ, et al. Late Archean biospheric oxygenation and atmospheric evolution. Science. 2007;317(5846):1900–1903. doi: 10.1126/science.1138700. [DOI] [PubMed] [Google Scholar]
- 19.Olson SL, Kump LR, Kasting JF. Quantifying the areal extent and dissolved oxygen concentrations of Archean oxygen oases. Chem Geol. 2013;362:35–43. [Google Scholar]
- 20.Zahnle K, Claire M, Catling D. The loss of mass-independent fractionation in sulfur due to a Palaeoproterozoic collapse of atmospheric methane. Geobiology. 2006;4:271–283. [Google Scholar]
- 21.Reinhard CT, Lalonde SV, Lyons TW. Oxidative sulfide dissolution on the early Earth. Chem Geol. 2013;362:44–55. [Google Scholar]
- 22.Baas Becking LGM, Kaplan IR, Moore D. Limits of the natural environment in terms of pH and oxidation-reduction potentials. J Geol. 1960;68:243–284. [Google Scholar]
- 23.Kirk JTO. Light and Photosynthesis in Aquatic Ecosystems. Cambridge Univ Press; New York: 1994. [Google Scholar]
- 24.Kühl M, Glud RN, Borum J, Roberts R, Rysgaard S. Photosynthetic performance of surface-associated algae below sea ice as measured with a pulse-amplitude-modulated (PAM) fluorometer and O2 microsensors. Mar Ecol Prog Ser. 2001;223:1–14. [Google Scholar]
- 25.Findlay S. Stream microbial ecology. J N Am Benthol Soc. 2010;29:170–181. [Google Scholar]
- 26.Schwartzman DW, Volk T. Biotic enhancement of weathering and the habitability of Earth. Nature. 1989;340:457–460. [Google Scholar]
- 27.Simpson EL, et al. Evidence for 2.0 Ga continental microbial mats in a paleodesert setting. Precambrian Res. 2013;237:36–50. [Google Scholar]
- 28.Rye R, Holland HD. Life associated with a 2.76 Ga ephemeral pond?: Evidence from Mount Roe #2 paleosol. Geology. 2000;28(6):483–486. doi: 10.1130/0091-7613(2000)28<483:lawage>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 29.Neaman A, Chorover J, Brantley SL. Element mobility patterns record organic ligands in soils on early Earth. Geology. 2005;33:117–120. [Google Scholar]
- 30.Driese SG, et al. Neoarchean paleoweathering of tonalite and metabasalt: Implications for reconstructions of 2.69 Ga early terrestrial ecosystems and paleoatmospheric chemistry. Precambrian Res. 2011;189:1–17. [Google Scholar]
- 31.Battistuzzi FU, Feijao A, Hedges SB. A genomic timescale of prokaryote evolution: Insights into the origin of methanogenesis, phototrophy, and the colonization of land. BMC Evol Biol. 2004;4:44. doi: 10.1186/1471-2148-4-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pulquério MJ, Nichols RA. Dates from the molecular clock: how wrong can we be? Trends Ecol Evol. 2007;22(4):180–184. doi: 10.1016/j.tree.2006.11.013. [DOI] [PubMed] [Google Scholar]
- 33.Donaldson JA, de Kemp EA. Archaean quartz arenites in the Canadian Shield: Examples from the Superior and Churchill Provinces. Sediment Geol. 1998;120:153–176. [Google Scholar]
- 34.Eriksson KA, Simpson EL, Mueller W. An unusual fluvial to tidal transition in the mesoarchean Moodies Group, South Africa: A response to high tidal range and active tectonics. Sediment Geol. 2006;190:13–24. [Google Scholar]
- 35.Thomazo C, Ader M, Farquhar J, Philippot P. Methanotrophs regulated atmospheric sulfur isotope anomalies during the Mesoarchean (Tumbiana Formation, Western Australia) Earth Planet Sci Lett. 2009;279:65–75. [Google Scholar]
- 36.Buick R. The antiquity of oxygenic photosynthesis: Evidence from stromatolites in sulphate-deficient Archaean lakes. Science. 1992;255(5040):74–77. doi: 10.1126/science.11536492. [DOI] [PubMed] [Google Scholar]
- 37.Sim MS, et al. Oxygen-dependent morphogenesis of modern clumped photosynthetic mats and implications for the Archean stromatolite record. Geosciences. 2012;2:235–259. [Google Scholar]
- 38.Coffey JM, Flannery DT, George SC. Sedimentology, stratigraphy and geochemistry of a stromatolite biofacies in the 2.72 Ga Tumbiana Formation, Fortescue Group, Western Australia. Precambrian Res. 2013;236:282–296. [Google Scholar]
- 39.Altermann W, Lenhardt N. The volcano-sedimentary succession of the Archean Sodium Group, Ventersdorp Supergroup, South Africa: Volcanology, sedimentology and geochemistry. Precambrian Res. 2012;214:60–81. [Google Scholar]
- 40.Rasmussen B, et al. Evidence for microbial life in synsedimentary cavities from 2.75 Ga terrestrial environments. Geology. 2009;37:423–426. [Google Scholar]
- 41.Heubeck C. An early ecosystem of Archean tidal microbial mats (Moodies Group, South Africa, ca. 3.2 Ga) Geology. 2009;3:931–934. [Google Scholar]
- 42.Schirrmeister BE, de Vos JM, Antonelli A, Bagheri HC. Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event. Proc Natl Acad Sci USA. 2013;110(5):1791–1796. doi: 10.1073/pnas.1209927110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Javaux EJ, Marshall CP, Bekker A. Organic-walled microfossils in 3.2-billion-year-old shallow-marine siliciclastic deposits. Nature. 2010;463(7283):934–938. doi: 10.1038/nature08793. [DOI] [PubMed] [Google Scholar]
- 44.Garcia-Pichel F, Belnap J. Microenvironments and microscale productivity of cyanobacterial desert crusts. J Phycol. 1996;32:774–782. [Google Scholar]
- 45.Herman EK, Kump LR. Biogeochemistry of microbial mats under Precambrian environmental conditions: A modelling study. Geobiology. 2005;3:77–92. [Google Scholar]
- 46.Maurice PA, McKnight DM, Leff L, Fulghulm JE, Gooseff M. Direct observations of aluminosilicate weathering in the hyporheic zone of an Antarctic Dry Valley stream. Geochim Cosmochim Acta. 2002;66:1335–1347. [Google Scholar]
- 47.Aiken G, McKnight D, Harnish R, Wershaw R. Geochemistry of aquatic humic substances in the Lake Fryxell Basin, Antarctica. Biogeochemistry. 1996;34:157–188. [Google Scholar]
- 48.Claire MW, Catling DC, Zahnle KJ. Biogeochemical modelling of the rise in atmospheric oxygen. Geobiology. 2006;4:239–269. [Google Scholar]
- 49.Kasting JF. What caused the rise of atmospheric O2? Chem Geol. 2013;362:13–25. [Google Scholar]
- 50.Catling DC. The Great Oxidation Event transition. In: Turekian KK, Holland HD, editors. Treatise on Geochemistry. 2nd Ed. Elsevier; Amsterdam: 2014. pp. 177–195. [Google Scholar]
- 51.Revsbech NP, Jorgensen BB, Blackburn TH, Cohen Y. Microelectrode studies of the photosynthesis and O2, H, S, and pH profiles of a microbial mat. Limnol Oceanogr. 1983;28:1062–1074. [Google Scholar]
- 52.Woelfel J, et al. Oxygen evolution in a hypersaline crust: In situ photosynthesis quantification by microelectrode profiling and use of planar optode spots in incubation chambers. Aquat Microb Ecol. 2009;56:263–273. [Google Scholar]
- 53.Crowe SA, et al. Sulfate was a trace constituent of Archean seawater. Science. 2014;346(6210):735–739. doi: 10.1126/science.1258966. [DOI] [PubMed] [Google Scholar]
- 54.Zhelezinskaia I, Kaufman AJ, Farquhar J, Cliff J. Large sulfur isotope fractionations associated with Neoarchean microbial sulfate reduction. Science. 2014;346(6210):742–744. doi: 10.1126/science.1256211. [DOI] [PubMed] [Google Scholar]
- 55.Field CB, Behrenfeld MJ, Randerson JT, Falkowski P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science. 1998;281(5374):237–240. doi: 10.1126/science.281.5374.237. [DOI] [PubMed] [Google Scholar]
- 56.Zahnle KJ, Catling DC, Claire MW. The rise of oxygen and the hydrogen hourglass. Chem Geol. 2013;362:26–34. [Google Scholar]
- 57.Rasmussen B, Fletcher IR, Brocks JJ, Kilburn MR. Reassessing the first appearance of eukaryotes and cyanobacteria. Nature. 2008;455(7216):1101–1104. doi: 10.1038/nature07381. [DOI] [PubMed] [Google Scholar]
- 58.Haqq-Misra J, Kasting JF, Lee S. Availability of O(2) and H(2)O(2) on pre-photosynthetic Earth. Astrobiology. 2011;11(4):293–302. doi: 10.1089/ast.2010.0572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pecoits E, et al. Atmospheric hydrogen peroxide and Eoarchean iron formations. Geobiology. 2015;13(1):1–14. doi: 10.1111/gbi.12116. [DOI] [PubMed] [Google Scholar]
- 60.Konhauser KO, et al. Could bacteria have formed the Precambrian banded iron formations? Geology. 2002;30:1079–1082. [Google Scholar]
- 61.Konhauser KO, et al. Decoupling photochemical Fe (II) oxidation from shallow-water BIF deposition. Earth Planet Sci Lett. 2007;258:87–100. [Google Scholar]
- 62.Kurzweil F, et al. Atmospheric sulfur rearrangement 2.7 billion years ago: Evidence for oxygenic photosynthesis. Earth Planet Sci Lett. 2013;366:17–26. [Google Scholar]
- 63.Kump LR, Barley ME. Increased subaerial volcanism and the rise of atmospheric oxygen 2.5 billion years ago. Nature. 2007;448(7157):1033–1036. doi: 10.1038/nature06058. [DOI] [PubMed] [Google Scholar]
- 64.Condie K. Episodic continental growth and supercontinents: a mantle avalanche connection? Earth Planet Sci Lett. 1998;163:97–108. [Google Scholar]
- 65.Bleeker W. The late Archean record: A puzzle in ca. 35 pieces. Lithos. 2003;71:99–134. [Google Scholar]
- 66.Hawkesworth CJ, et al. The generation and evolution of the continental crust. J Geol Soc London. 2010;167:229–248. [Google Scholar]
- 67.Flament N, Coltice N, Rey PF. The evolution of the 87Sr/86Sr of marine carbonates does not constrain continental growth. Precambrian Res. 2013;229:177–188. [Google Scholar]
- 68.Knoll AH. Archean photoautotrophy: Some alternatives and limits. Orig Life. 1979;9(4):313–327. doi: 10.1007/BF00926824. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.