

Abstract
Coronaviruses (CoVs), enveloped positive-sense RNA viruses, are characterized by club-like spikes that project from their surface, an unusually large RNA genome, and a unique replication strategy. Coronaviruses cause a variety of diseases in mammals and birds ranging from enteritis in cows and pigs and upper respiratory disease in chickens to potentially lethal human respiratory infections. Here we provide a brief introduction to coronaviruses discussing their replication and pathogenicity, and current prevention and treatment strategies. We also discuss the outbreaks of the highly pathogenic Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) and the recently identified Middle Eastern Respiratory Syndrome Coronavirus (MERS-CoV).
Key words: Nidovirales, Coronavirus, Positive-sense RNA viruses, SARS-CoV, MERS-CoV
Classification
Coronaviruses (CoVs) are the largest group of viruses belonging to the Nidovirales order, which includes Coronaviridae, Arteriviridae, Mesoniviridae, and Roniviridae families. The Coronavirinae comprise one of two subfamilies in the Coronaviridae family, with the other being the Torovirinae. The Coronavirinae are further subdivided into four genera, the alpha, beta, gamma, and delta coronaviruses. The viruses were initially sorted into these genera based on serology but are now divided by phylogenetic clustering.
All viruses in the Nidovirales order are enveloped, non-segmented positive-sense RNA viruses. They all contain very large genomes for RNA viruses, with some viruses having the largest identified RNA genomes, containing up to 33.5 kilobase (kb) genomes. Other common features within the Nidovirales order include: (1) a highly conserved genomic organization, with a large replicase gene preceding structural and accessory genes; (2) expression of many non-structural genes by ribosomal frameshifting; (3) several unique or unusual enzymatic activities encoded within the large replicase–transcriptase polyprotein; and (4) expression of downstream genes by synthesis of 3′ nested sub-genomic mRNAs. In fact, the Nidovirales order name is derived from these nested 3′ mRNAs as nido is Latin for “nest.” The major differences within the Nidovirus families are in the number, type, and sizes of the structural proteins. These differences cause significant alterations in the structure and morphology of the nucleocapsids and virions.
Genomic Organization
Coronaviruses contain a non-segmented, positive-sense RNA genome of ~30 kb. The genome contains a 5′ cap structure along with a 3′ poly (A) tail, allowing it to act as an mRNA for translation of the replicase polyproteins. The replicase gene encoding the non-structural proteins (nsps) occupies two-thirds of the genome, about 20 kb, as opposed to the structural and accessory proteins, which make up only about 10 kb of the viral genome. The 5′ end of the genome contains a leader sequence and untranslated region (UTR) that contains multiple stem loop structures required for RNA replication and transcription. Additionally, at the beginning of each structural or accessory gene are transcriptional regulatory sequences (TRSs) that are required for expression of each of these genes (see Subheading 4.3 on RNA replication). The 3′ UTR also contains RNA structures required for replication and synthesis of viral RNA. The organization of the coronavirus genome is 5′-leader-UTR-replicase-S (Spike)-E (Envelope)-M (Membrane)-N (Nucleocapsid)-3′ UTR-poly (A) tail with accessory genes interspersed within the structural genes at the 3′ end of the genome (Fig. 1). The accessory proteins are almost exclusively nonessential for replication in tissue culture; however, some have been shown to have important roles in viral pathogenesis [1].
Fig. 1.
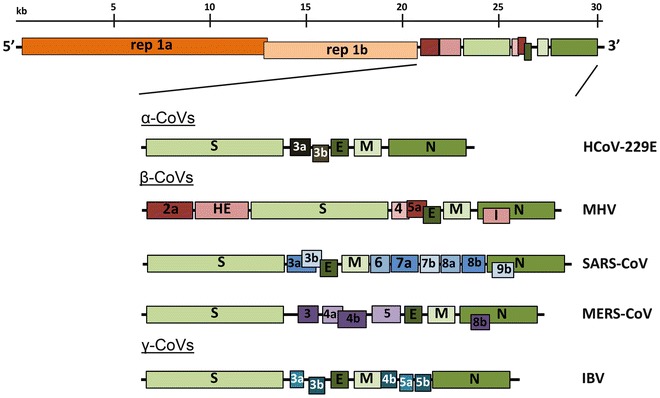
Genomic organization of representative α, β, and γ CoVs. An illustration of the MHV genome is depicted at the top. The expanded regions below show the structural and accessory proteins in the 3′ regions of the HCoV-229E, MHV, SARS-CoV, MERS-CoV and IBV. Size of the genome and individual genes are approximated using the legend at the top of the diagram but are not drawn to scale. HCoV-229E human coronavirus 229E, MHV mouse hepatitis virus, SARS-CoV severe acute respiratory syndrome coronavirus, MERS-CoV Middle East respiratory syndrome coronavirus, IBV infectious bronchitis virus
Virion Structure
Coronavirus virions are spherical with diameters of approximately 125 nm as depicted in recent studies by cryo-electron tomography and cryo-electron microscopy [2, 3]. The most prominent feature of coronaviruses is the club-shaped spike projections emanating from the surface of the virion. These spikes are a defining feature of the virion and give them the appearance of a solar corona, prompting the name, coronaviruses. Within the envelope of the virion is the nucleocapsid. Coronaviruses have helically symmetrical nucleocapsids, which is uncommon among positive-sense RNA viruses, but far more common for negative-sense RNA viruses.
Coronavirus particles contain four main structural proteins. These are the spike (S), membrane (M), envelope (E), and nucleocapsid (N) proteins, all of which are encoded within the 3′ end of the viral genome. The S protein (~150 kDa), utilizes an N-terminal signal sequence to gain access to the ER, and is heavily N-linked glycosylated. Homotrimers of the virus encoded S protein make up the distinctive spike structure on the surface of the virus [4, 5]. The trimeric S glycoprotein is a class I fusion protein [6] and mediates attachment to the host receptor [7]. In most, coronaviruses, S is cleaved by a host cell furin-like protease into two separate polypeptides noted S1 and S2 [8, 9]. S1 makes up the large receptor-binding domain of the S protein, while S2 forms the stalk of the spike molecule [10].
The M protein is the most abundant structural protein in the virion. It is a small (~25–30 kDa) protein with three transmembrane domains [11] and is thought to give the virion its shape. It has a small N-terminal glycosylated ectodomain and a much larger C-terminal endodomain that extends 6–8 nm into the viral particle [12]. Despite being co-translationally inserted in the ER membrane, most M proteins do not contain a signal sequence. Recent studies suggest the M protein exists as a dimer in the virion, and may adopt two different conformations, allowing it to promote membrane curvature as well as to bind to the nucleocapsid [13].
The E protein (~8–12 kDa) is found in small quantities within the virion. The coronavirus E proteins are highly divergent but have a common architecture [14]. The membrane topology of E protein is not completely resolved but most data suggest that it is a transmembrane protein. The E protein has an N-terminal ectodomain and a C-terminal endodomain and has ion channel activity. As opposed to other structural proteins, recombinant viruses lacking the E protein are not always lethal, although this is virus type dependent [15]. The E protein facilitates assembly and release of the virus (see Subheading 4.4), but also has other functions. For instance, the ion channel activity in SARS-CoV E protein is not required for viral replication but is required for pathogenesis [16].
The N protein constitutes the only protein present in the nucleocapsid. It is composed of two separate domains, an N-terminal domain (NTD) and a C-terminal domain (CTD), both capable of binding RNA in vitro, but each domain uses different mechanisms to bind RNA. It has been suggested that optimal RNA binding requires contributions from both domains [17, 18]. N protein is also heavily phosphorylated [19], and phosphorylation has been suggested to trigger a structural change enhancing the affinity for viral versus nonviral RNA. N protein binds the viral genome in a beads-on-a-string type conformation. Two specific RNA substrates have been identified for N protein; the TRSs [20] and the genomic packaging signal [21]. The genomic packaging signal has been found to bind specifically to the second, or C-terminal RNA binding domain [22]. N protein also binds nsp3 [18, 23], a key component of the replicase complex, and the M protein [24]. These protein interactions likely help tether the viral genome to the replicase–transcriptase complex (RTC), and subsequently package the encapsidated genome into viral particles.
A fifth structural protein, the hemagglutinin-esterase (HE), is present in a subset of β-coronaviruses. The protein acts as a hemagglutinin, binds sialic acids on surface glycoproteins, and contains acetyl-esterase activity [25]. These activities are thought to enhance S protein-mediated cell entry and virus spread through the mucosa [26]. Interestingly, HE enhances murine hepatitis virus (MHV) neurovirulence [27]; however, it is selected against in tissue culture for unknown reasons [28].
Coronavirus Life Cycle
Attachment and Entry
The initial attachment of the virion to the host cell is initiated by interactions between the S protein and its receptor. The sites of receptor binding domains (RBD) within the S1 region of a coronavirus S protein vary depending on the virus, with some having the RBD at the N-terminus of S1 (MHV), while others (SARS-CoV) have the RBD at the C-terminus of S1 [29, 30]. The S-protein–receptor interaction is the primary determinant for a coronavirus to infect a host species and also governs the tissue tropism of the virus. Many coronaviruses utilize peptidases as their cellular receptor. It is unclear why peptidases are used, as entry occurs even in the absence of the enzymatic domain of these proteins. Many α-coronaviruses utilize aminopeptidase N (APN) as their receptor, SARS-CoV and HCoV-NL63 use angiotensin-converting enzyme 2 (ACE2) as their receptor, MHV enters through CEACAM1, and the recently identified MERS-CoV binds to dipeptidyl-peptidase 4 (DPP4) to gain entry into human cells (see Table 1 for a list of known CoV receptors).
Table 1.
Coronavirus receptors
| Virus | Receptor | References |
|---|---|---|
| Alphacoronaviruses | ||
| HCoV-229E | APN | [115] |
| HCoV-NL63 | ACE2 | [116] |
| TGEV | APN | [117] |
| PEDV | APN | [118] |
| FIPV | APN | [119] |
| CCoV | APN | [120] |
| Betacoronaviruses | ||
| MHV | mCEACAM | [121, 122] |
| BCoV | N-acetyl-9-O-acetylneuraminic acid | [123] |
| SARS-CoV | ACE2 | [124] |
| MERS-CoV | DPP4 | [100] |
APN aminopeptidase N, ACE2 angiotensin-converting enzyme 2, mCEACAM murine carcinoembryonic antigen-related adhesion molecule 1, DPP4 dipeptidyl peptidase 4, HCoV human coronavirus, TGEV transmissible gastroenteritis virus, PEDV porcine epidemic diarrhea virus, FIPV feline infectious peritonitis virus, CCoV canine coronavirus, MHV murine hepatitis virus, BCoV bovine coronavirus, SARS-CoV severe acute respiratory syndrome coronavirus, MERS-CoV Middle East respiratory syndrome coronavirus
Following receptor binding, the virus must next gain access to the host cell cytosol. This is generally accomplished by acid-dependent proteolytic cleavage of S protein by a cathepsin, TMPRRS2 or another protease, followed by fusion of the viral and cellular membranes. S protein cleavage occurs at two sites within the S2 portion of the protein, with the first cleavage important for separating the RBD and fusion domains of the S protein [31] and the second for exposing the fusion peptide (cleavage at S2′). Fusion generally occurs within acidified endosomes, but some coronaviruses, such as MHV, can fuse at the plasma membrane. Cleavage at S2′ exposes a fusion peptide that inserts into the membrane, which is followed by joining of two heptad repeats in S2 forming an antiparallel six-helix bundle [6]. The formation of this bundle allows for the mixing of viral and cellular membranes, resulting in fusion and ultimately release of the viral genome into the cytoplasm.
Replicase Protein Expression
The next step in the coronavirus lifecycle is the translation of the replicase gene from the virion genomic RNA. The replicase gene encodes two large ORFs, rep1a and rep1b, which express two co-terminal polyproteins, pp1a and pp1ab (Fig. 1). In order to express both polyproteins, the virus utilizes a slippery sequence (5′-UUUAAAC-3′) and an RNA pseudoknot that cause ribosomal frameshifting from the rep1a reading frame into the rep1b ORF. In most cases, the ribosome unwinds the pseudoknot structure, and continues translation until it encounters the rep1a stop codon. Occasionally the pseudoknot blocks the ribosome from continuing elongation, causing it to pause on the slippery sequence, changing the reading frame by moving back one nucleotide, a -1 frameshift, before the ribosome is able to melt the pseudoknot structure and extend translation into rep1b, resulting in the translation of pp1ab [32, 33]. In vitro studies predict the incidence of ribosomal frameshifting to be as high as 25 %, but this has not been determined in the context of virus infection. It is unknown exactly why these viruses utilize frameshifting to control protein expression, but it is hypothesized to either control the precise ratio of rep1b and rep1a proteins or delay the production of rep1b products until the products of rep1a have created a suitable environment for RNA replication [34].
Polyproteins pp1a and pp1ab contain the nsps 1–11 and 1–16, respectively. In pp1ab, nsp11 from pp1a becomes nsp12 following extension of pp1a into pp1b. However, γ-coronaviruses do not contain a comparable nsp1. These polyproteins are subsequently cleaved into the individual nsps [35]. Coronaviruses encode either two or three proteases that cleave the replicase polyproteins. They are the papain-like proteases (PLpro), encoded within nsp3, and a serine type protease, the main protease, or Mpro, encoded by nsp5. Most coronaviruses encode two PLpros within nsp3, except the γ-coronaviruses, SARS-CoV and MERS-CoV, which only express one PLpro [36]. The PLpros cleave the nsp1/2, nsp2/3, and nsp3/4 boundaries, while the Mpro is responsible for the remaining 11 cleavage events.
Next, many of the nsps assemble into the replicase–transcriptase complex (RTC) to create an environment suitable for RNA synthesis, and ultimately are responsible for RNA replication and transcription of the sub-genomic RNAs. The nsps also contain other enzyme domains and functions, including those important for RNA replication, for example nsp12 encodes the RNA-dependent RNA polymerase (RdRp) domain; nsp13 encodes the RNA helicase domain and RNA 5′-triphosphatase activity; nsp14 encodes the exoribonuclease (ExoN) involved in replication fidelity and N7-methyltransferase activity; and nsp16 encodes 2′-O-methyltransferase activity. In addition to the replication functions other activities, such as blocking innate immune responses (nsp1; nsp16-2′-O-methyl transferase; nsp3-deubiquitinase) have been identified for some of the nsps, while others have largely unknown functions (nsp3-ADP-ribose-1″-phosphatase; nsp15-endoribo-nuclease (NendoU)). For a list of non-structural proteins and their proposed functions, see Table 2. Interestingly, ribonucleases nsp15-NendoU and nsp14-ExoN activities are unique to the Nidovirales order and are considered genetic markers for these viruses [37].
Table 2.
Functions of coronavirus non-structural proteins (nsps)
| Protein | Function | References |
|---|---|---|
| nsp1 | Promotes cellular mRNA degradation and blocks host cell translation, results in blocking innate immune response | [125–128] |
| nsp2 | No known function, binds to prohibitin proteins | [129, 130] |
| nsp3 |
Large, multi-domain transmembrane protein, activities include: • Ubl1 and Ac domains, interact with N protein • ADRP activity, promotes cytokine expression • PLPro/Deubiquitinase domain, cleaves viral polyprotein and blocks host innate immune response • Ubl2, NAB, G2M, SUD, Y domains, unknown functions |
[131–138] |
| nsp4 | Potential transmembrane scaffold protein, important for proper structure of DMVs | [139, 140] |
| nsp5 | Mpro, cleaves viral polyprotein | [141] |
| nsp6 | Potential transmembrane scaffold protein | [142] |
| nsp7 | Forms hexadecameric complex with nsp8, may act as processivity clamp for RNA polymerase | [143] |
| nsp8 | Forms hexadecameric complex with nsp7, may act as processivity clamp for RNA polymerase; may act as primase | [143, 144] |
| nsp9 | RNA binding protein | [145] |
| nsp10 | Cofactor for nsp16 and nsp14, forms heterodimer with both and stimulates ExoN and 2-O-MT activity | [146, 147] |
| nsp12 | RdRp | [148] |
| nsp13 | RNA helicase, 5′ triphosphatase | [149, 150] |
| nsp14 | N7 MTase and 3′-5′ exoribonuclease, ExoN; N7 MTase adds 5′ cap to viral RNAs, ExoN activity is important for proofreading of viral genome | [151–154] |
| nsp15 | Viral endoribonuclease, NendoU | [155, 156] |
| nsp16 | 2′-O-MT; shields viral RNA from MDA5 recognition | [157, 158] |
Ubl ubiquitin-like, Ac acidic, ADRP ADP-ribose-1′-phosphatase, PLPro papain-like protease, NAB nucleic acid binding, SUD SARS-unique domain, DMVs double-membrane vesicles, Mpro main protease, RdRp RNA-dependent RNA polymerase, MTase methyltransferase, Exo N viral exoribonuclease, Nendo U viral endoribonuclease, 2′-O-MT 2′-O-methyltransferase, MDA5 melanoma differentiation associated protein 5
Replication and Transcription
Viral RNA synthesis follows the translation and assembly of the viral replicase complexes. Viral RNA synthesis produces both genomic and sub-genomic RNAs. Sub-genomic RNAs serve as mRNAs for the structural and accessory genes which reside downstream of the replicase polyproteins. All positive-sense sub-genomic RNAs are 3′ co-terminal with the full-length viral genome and thus form a set of nested RNAs, a distinctive property of the order Nidovirales. Both genomic and sub-genomic RNAs are produced through negative-strand intermediates. These negative-strand intermediates are only about 1 % as abundant as their positive-sense counterparts and contain both poly-uridylate and anti-leader sequences [38].
Many cis-acting sequences are important for the replication of viral RNAs. Within the 5′ UTR of the genome are seven stem-loop structures that may extend into the replicase 1a gene [39–42]. The 3′ UTR contains a bulged stem-loop, a pseudoknot, and a hypervariable region [43–46]. Interestingly, the stem-loop and the pseudoknot at the 3′ end overlap, and thus cannot form simultaneously [44, 47]. Therefore, these different structures are proposed to regulate alternate stages of RNA synthesis, although exactly which stages are regulated and their precise mechanism of action are still unknown.
Perhaps the most novel aspect of coronavirus replication is how the leader and body TRS segments fuse during production of sub-genomic RNAs. This was originally thought to occur during positive-strand synthesis, but now it is largely believed to occur during the discontinuous extension of negative-strand RNA [48]. The current model proposes that the RdRp pauses at any one of the body TRS sequences (TRS-B); following this pause the RdRp either continues elongation to the next TRS or it switches to amplifying the leader sequence at the 5′ end of the genome guided by complementarity of the TRS-B to the leader TRS (TRS-L). Many pieces of evidence currently support this model, including the presence of anti-leader sequence at the 3′ end of the negative-strand sub-genomic RNAs [38]. However, many questions remain to fully define the model. For instance, how does the RdRp bypass all of the TRS-B sequences to produce full-length negative-strand genomic RNA? Also, how are the TRS-B sequences directed to the TRS-L and how much complementarity is necessary [49]? Answers to these questions and others will be necessary to gain a full perspective of how RNA replication occurs in coronaviruses.
Finally, coronaviruses are also known for their ability to recombine using both homologous and nonhomologous recombination [50, 51]. The ability of these viruses to recombine is tied to the strand switching ability of the RdRp. Recombination likely plays a prominent role in viral evolution and is the basis for targeted RNA recombination, a reverse genetics tool used to engineer viral recombinants at the 3′ end of the genome.
Assembly and Release
Following replication and sub-genomic RNA synthesis, the viral structural proteins, S, E, and M are translated and inserted into the endoplasmic reticulum (ER). These proteins move along the secretory pathway into the endoplasmic reticulum–Golgi intermediate compartment (ERGIC) [52, 53]. There, viral genomes encapsidated by N protein bud into membranes of the ERGIC containing viral structural proteins, forming mature virions [54].
The M protein directs most protein–protein interactions required for assembly of coronaviruses. However, M protein is not sufficient for virion formation, as virus-like particles (VLPs) cannot be formed by M protein expression alone. When M protein is expressed along with E protein VLPs are formed, suggesting these two proteins function together to produce coronavirus envelopes [55]. N protein enhances VLP formation, suggesting that fusion of encapsidated genomes into the ERGIC enhances viral envelopment [56]. The S protein is incorporated into virions at this step, but is not required for assembly. The ability of the S protein to traffic to the ERGIC and interact with the M protein is critical for its incorporation into virions.
While the M protein is relatively abundant, the E protein is only present in small quantities in the virion. Thus, it is likely that M protein interactions provide the impetus for envelope maturation. It is unknown how E protein assists M protein in assembly of the virion, and several possibilities have been suggested. Some work has indicated a role for the E protein in inducing membrane curvature [57–59], although others have suggested that E protein prevents the aggregation of M protein [60]. The E protein may also have a separate role in promoting viral release by altering the host secretory pathway [61].
The M protein also binds to the nucleocapsid, and this interaction promotes the completion of virion assembly. These interactions have been mapped to the C-terminus of the endodomain of M with CTD of the N-protein [62]. However, it is unclear exactly how the nucleocapsid complexed with virion RNA traffics to the ERGIC to interact with M protein and become incorporated into the viral envelope. Another outstanding question is how the N protein selectively packages only positive-sense full-length genomes among the many different RNA species produced during infection. A packaging signal for MHV has been identified in the nsp15 coding sequence, but mutation of this signal does not appear to affect virus production, and a mechanism for how this packaging signal works has not been determined [22]. Furthermore, most coronaviruses do not contain similar sequences at this locus, indicating that packaging may be virus specific.
Following assembly, virions are transported to the cell surface in vesicles and released by exocytosis. It is not known if the virions use the traditional pathway for transport of large cargo from the Golgi or if the virus has diverted a separate, unique pathway for its own exit. In several coronaviruses, S protein that does not get assembled into virions transits to the cell surface where it mediates cell–cell fusion between infected cells and adjacent, uninfected cells. This leads to the formation of giant, multinucleated cells, which allows the virus to spread within an infected organism without being detected or neutralized by virus-specific antibodies.
Pathogenesis
Animal Coronaviruses
Coronaviruses cause a large variety of diseases in animals, and their ability to cause severe disease in livestock and companion animals such as pigs, cows, chickens, dogs, and cats led to significant research on these viruses in the last half of the twentieth century. For instance, Transmissible Gastroenteritis Virus (TGEV) and Porcine Epidemic Diarrhea Virus (PEDV) cause severe gastroenteritis in young piglets, leading to significant morbidity, mortality, and ultimately economic losses. PEDV recently emerged in North America for the first time, causing significant losses of young piglets. Porcine hemagglutinating encephalomyelitis virus (PHEV) mostly leads to enteric infection but has the ability to infect the nervous system, causing encephalitis, vomiting, and wasting in pigs. Feline enteric coronavirus (FCoV) causes a mild or asymptomatic infection in domestic cats, but during persistent infection, mutation transforms the virus into a highly virulent strain of FCoV, Feline Infectious Peritonitis Virus (FIPV), that leads to development of a lethal disease called feline infectious peritonitis (FIP). FIP has wet and dry forms, with similarities to the human disease, sarcoidosis. FIPV is macrophage tropic and it is believed that it causes aberrant cytokine and/or chemokine expression and lymphocyte depletion, resulting in lethal disease [63]. However, additional research is needed to confirm this hypothesis. Bovine CoV, Rat CoV, and Infectious Bronchitis Virus (IBV) cause mild to severe respiratory tract infections in cattle, rats, and chickens, respectively. Bovine CoV causes significant losses in the cattle industry and also has spread to infect a variety of ruminants, including elk, deer, and camels. In addition to severe respiratory disease, the virus causes diarrhea (“winter dysentery” and “shipping fever”), all leading to weight loss, dehydration, decreased milk production, and depression [63]. Some strains of IBV, a γ-coronavirus, also affect the urogenital tract of chickens causing renal disease. Infection of the reproductive tract with IBV significantly diminishes egg production, causing substantial losses in the egg-production industry each year [63]. More recently, a novel coronavirus named SW1 has been identified in a deceased Beluga whale [64]. Large numbers of virus particles were identified in the liver of the deceased whale with respiratory disease and acute liver failure. Although, electron microscopic images were not sufficient to identify the virus as a coronavirus, sequencing of the liver tissue clearly identified the virus as a coronavirus. It was subsequently determined to be a γ-coronavirus based on phylogenetic analysis but it has not yet been verified experimentally that this virus is actually a causative agent of disease in whales. In addition, there has been intense interest in identifying novel bat CoVs, since these are the likely ancestors for SARS-CoV and MERS-CoV, and hundreds of novel bat coronaviruses have been identified over the past decade [65]. Finally, another novel family of nidoviruses, Mesoniviridae, has been recently identified as the first nidoviruses to exclusively infect insect hosts [66, 67]. These viruses are highly divergent from other nidoviruses but are most closely related to the roniviruses. In size, they are ~20 kb, falling in between large and small nidoviruses. Interestingly, these viruses do not encode for an endoribonuclease, which is present in all other nidoviruses. These attributes suggest these viruses are the prototype of a new nidovirus family and may be a missing link in the transition from small to large nidoviruses.
The most heavily studied animal coronavirus is murine hepatitis virus (MHV), which causes a variety of outcomes in mice, including respiratory, enteric, hepatic, and neurologic infections. These infections often serve as highly useful models of disease. For instance, MHV-1 causes severe respiratory disease in susceptible A/J and C3H/HeJ mice, A59 and MHV-3 induce severe hepatitis, while JHMV causes severe encephalitis. Interestingly, MHV-3 induces cellular injury through the activation of the coagulation cascade [68]. Most notably, A59 and attenuated versions of JHMV cause a chronic demyelinating disease that bears similarities to multiple sclerosis (MS), making MHV infection one of the best models for this debilitating human disease. Early studies suggested that demyelination was dependent on viral replication in oligodendrocytes in the brain and spinal cord [69, 70]; however, more recent reports clearly demonstrate that the disease is immune-mediated. Irradiated mice or immunodeficient (lacking T and B cells) mice do not develop demyelination, but addition of virus-specific T cells restores the development of demyelination [71, 72]. Additionally, demyelination is accompanied by a large influx of macrophages and microglia that can phagocytose infected myelin [73], although it is unknown what the signals are that direct immune cells to destroy myelin. Finally, MHV can be studied under BSL2 laboratory conditions, unlike SARS-CoV or MERS-CoV, which require a BSL3 laboratory, and provides a large number of suitable animal models. These factors make MHV an ideal model for studying the basics of viral replication in tissue culture cells as well as for studying the pathogenesis and immune response to coronaviruses.
Human Coronaviruses
Prior to the SARS-CoV outbreak, coronaviruses were only thought to cause mild, self-limiting respiratory infections in humans. Two of these human coronaviruses are α-coronaviruses, HCoV-229E and HCoV-NL63, while the other two are β-coronaviruses, HCoV-OC43 and HCoV-HKU1. HCoV-229E and HCoV-OC43 were isolated nearly 50 years ago [74–76], while HCoV-NL63 and HCoV-HKU1 have only recently been identified following the SARS-CoV outbreak [77, 78]. These viruses are endemic in the human populations, causing 15–30 % of respiratory tract infections each year. They cause more severe disease in neonates, the elderly, and in individuals with underlying illnesses, with a greater incidence of lower respiratory tract infection in these populations. HCoV-NL63 is also associated with acute laryngotracheitis (croup) [79]. One interesting aspect of these viruses is their differences in tolerance to genetic variability. HCoV-229E isolates from around the world have only minimal sequence divergence [80], while HCoV-OC43 isolates from the same location but isolated in different years show significant genetic variability [81]. This likely explains the inability of HCoV-229E to cross the species barrier to infect mice while HCoV-OC43 and the closely related bovine coronavirus, BCoV, are capable of infecting mice and several ruminant species. Based on the ability of MHV to cause demyelinating disease, it has been suggested that human CoVs may be involved in the development of multiple sclerosis (MS). However, no evidence to date suggests that human CoVs play a significant role in MS.
SARS-CoV, a group 2b β-coronavirus, was identified as the causative agent of the Severe Acute Respiratory Syndrome (SARS) outbreak that occurred in 2002–2003 in the Guangdong Province of China. It is the most severe human disease caused by any coronavirus. During the 2002–2003 outbreak approximately 8,098 cases occurred with 774 deaths, resulting in a mortality rate of 9 %. This rate was much higher in elderly individuals, with mortality rates approaching 50 % in individuals over 60 years of age. Furthermore, the outbreak resulted in the loss of nearly $40 billion dollars in economic activity, as the virus nearly shut down many activities in Southeast Asia and Toronto, Canada for several months. The outbreak began in a hotel in Hong Kong and ultimately spread to more than two dozen countries. During the epidemic, closely related viruses were isolated from several exotic animals including Himalayan palm civets and raccoon dogs [82]. However, it is widely accepted that SARS-CoV originated in bats as a large number of Chinese horseshoe bats contain sequences of SARS-related CoVs and contain serologic evidence for a prior infection with a related CoV [83, 84]. In fact, two novel bat SARS-related CoVs have been recently identified that are more similar to SARS-CoV than any other virus identified to date [85]. They were also found to use the same receptor as the human virus, angiotensin converting enzyme 2 (ACE2), providing further evidence that SARS-CoV originated in bats. Although some human individuals within wet animal markets had serologic evidence of SARS-CoV infection prior to the outbreak, these individuals had no apparent symptoms [82]. Thus, it is likely that a closely related virus circulated in the wet animal markets for several years before a series of factors facilitated its spread into the larger population.
Transmission of SARS-CoV was relatively inefficient, as it only spread through direct contact with infected individuals after the onset of illness. Thus, the outbreak was largely contained within households and healthcare settings [86], except in a few cases of superspreading events where one individual was able to infect multiple contacts due to an enhanced development of high viral burdens or ability to aerosolize virus. As a result of the relatively inefficient transmission of SARS-CoV, the outbreak was controllable through the use of quarantining. Only a small number of SARS cases occurred after the outbreak was controlled in June 2003.
SARS-CoV primarily infects epithelial cells within the lung. The virus is capable of entering macrophages and dendritic cells but only leads to an abortive infection [87, 88]. Despite this, infection of these cell types may be important in inducing pro-inflammatory cytokines that may contribute to disease [89]. In fact, many cytokines and chemokines are produced by these cell types and are elevated in the serum of SARS-CoV infected patients [90]. The exact mechanism of lung injury and cause of severe disease in humans remains undetermined. Viral titers seem to diminish when severe disease develops in both humans and in several animal models of the disease. Furthermore, animals infected with rodent-adapted SARS-CoV strains show similar clinical features to the human disease, including an age-dependent increase in disease severity [91]. These animals also show increased levels of proinflammatory cytokines and reduced T-cell responses, suggesting a possible immunopathological mechanism of disease [92, 93].
While the SARS-CoV epidemic was controlled in 2003 and the virus has not since returned, a novel human CoV emerged in the Middle East in 2012. This virus, named Middle East Respiratory Syndrome-CoV (MERS-CoV), was found to be the causative agent in a series of highly pathogenic respiratory tract infections in Saudi Arabia and other countries in the Middle East [94]. Based on the high mortality rate of ~50 % in the early stages of the outbreak, it was feared the virus would lead to a very serious outbreak. However, the outbreak did not accelerate in 2013, although sporadic cases continued throughout the rest of the year. In April 2014, a spike of over 200 cases and almost 40 deaths occurred, prompting fears that the virus had mutated and was more capable of human-to-human transmission. More likely, the increased number of cases resulted from improved detection and reporting methods combined with a seasonal increase in birthing camels. As of August 27th, 2014 there have been a total of 855 cases of MERS-CoV, with 333 deaths and a case fatality rate of nearly 40 %, according to the European Center for Disease Prevention and Control.
MERS-CoV is a group 2c β-coronavirus highly related to two previously identified bat coronaviruses, HKU4 and HKU5 [95]. It is believed that the virus originated from bats, but likely had an intermediate host as humans rarely come in contact with bat secreta. Serological studies have identified MERS-CoV antibodies in dromedary camels in the Middle East [96], and cell lines from camels have been found to be permissive for MERS-CoV replication [97] providing evidence that dromedary camels may be the natural host. More convincing evidence for this comes from recent studies identifying nearly identical MERS-CoVs in both camels and human cases in nearby proximities in Saudi Arabia [98, 99]. In one of these studies the human case had direct contact with an infected camel and the virus isolated from this patient was identical to the virus isolated from the camel [99]. At the present time it remains to be determined how many MERS-CoV cases can be attributed to an intermediate host as opposed to human-to-human transmission. It has also been postulated that human-to-camel spread contributed to the outbreak.
MERS-CoV utilizes Dipeptidyl peptidase 4 (DPP4) as its receptor [100]. The virus is only able to use the receptor from certain species such as bats, humans, camels, rabbits, and horses to establish infection. Unfortunately for researchers, the virus is unable to infect mouse cells due to differences in the structure of DPP4, making it difficult to evaluate potential vaccines or antivirals. Recently, a small animal model for MERS-CoV has been developed using an Adenoviral vector to introduce the human DPP4 gene into mouse lungs [101]. This unique system makes it possible to test therapeutic interventions and novel vaccines for MERS-CoV in any animal sensitive to adenoviral transductions.
Diagnosis, Treatment, and Prevention
In most cases of self-limited infection, diagnosis of coronaviruses is unnecessary, as the disease will naturally run its course. However, it may be important in certain clinical and veterinary settings or in epidemiological studies to identify an etiological agent. Diagnosis is also important in locations where a severe CoV outbreak is occurring, such as, at present, in the Middle East, where MERS-CoV continues to circulate. The identification of cases will guide the development of public health measures to control outbreaks. It is also important to diagnose cases of severe veterinary CoV-induced disease, such as PEDV and IBV, to control these pathogens and protect food supplies. RT-PCR has become the method of choice for diagnosis of human CoV, as multiplex real-time RT-PCR assays have been developed, are able to detect all four respiratory HCoVs and could be further adapted to novel CoVs [102, 103]. Serologic assays are important in cases where RNA is difficult to isolate or is no longer present, and for epidemiological studies.
To date, there are no antiviral therapeutics that specifically target human coronaviruses, so treatments are only supportive. In vitro, interferons (IFNs) are only partially effective against coronaviruses [104]. IFNs in combination with ribavirin may have increased activity in vitro when compared to IFNs alone against some coronaviruses; however, the effectiveness of this combination in vivo requires further evaluation [105]. The SARS and MERS outbreaks have stimulated research on these viruses and this research has identified a large number of suitable antiviral targets, such as viral proteases, polymerases, and entry proteins. Significant work remains, however, to develop drugs that target these processes and are able to inhibit viral replication.
Only limited options are available to prevent coronavirus infections. Vaccines have only been approved for IBV, TGEV, and Canine CoV, but these vaccines are not always used because they are either not very effective, or in some cases have been reported to be involved in the selection of novel pathogenic CoVs via recombination of circulating strains. Vaccines for veterinary pathogens, such as PEDV, may be useful in such cases where spread of the virus to a new location could lead to severe losses of veterinary animals. In the case of SARS-CoV, several potential vaccines have been developed but none are yet approved for use. These vaccines include recombinant attenuated viruses, live virus vectors, or individual viral proteins expressed from DNA plasmids. Therapeutic SARS-CoV neutralizing antibodies have been generated and could be retrieved and used again in the event of another SARS-CoV outbreak. Such antibodies would be most useful for protecting healthcare workers. In general, it is thought that live attenuated vaccines would be the most efficacious in targeting coronaviruses. This was illustrated in the case of TGEV, where an attenuated variant, PRCV, appeared in Europe in the 1980s. This variant only caused mild disease and completely protected swine from TGEV. Thus, this attenuated virus has naturally prevented the reoccurrence of severe TGEV in Europe and the U.S. over the past 30 years [106]. Despite this success, vaccine development for coronaviruses faces many challenges [107]. First, for mucosal infections, natural infection does not prevent subsequent infection, and so vaccines must either induce better immunity than the original virus or must at least lessen the disease incurred during a secondary infection. Second, the propensity of the viruses to recombine may pose a problem by rendering the vaccine useless and potentially increasing the evolution and diversity of the virus in the wild [108]. Finally, it has been shown in FIPV that vaccination with S protein leads to enhanced disease [109]. Despite this, several strategies are being developed for vaccine development to reduce the likelihood of recombination, for instance by making large deletions in the nsp1 [110] or E proteins [111], rearranging the 3′ end of the genome [112], modifying the TRS sequences [113], or using mutant viruses with abnormally high mutation rates that significantly attenuate the virus [114].
Owing to the lack of effective therapeutics or vaccines, the best measures to control human coronaviruses remain a strong public health surveillance system coupled with rapid diagnostic testing and quarantine when necessary. For international outbreaks, cooperation of governmental entities, public health authorities, and health care providers is critical. During veterinary outbreaks that are readily transmitted, such as PEDV, more drastic measures such as destruction of entire herds of pigs may be necessary to prevent transmission of these deadly viruses.
Conclusion
Over the past 50 years the emergence of many different coronaviruses that cause a wide variety of human and veterinary diseases has occurred. It is likely that these viruses will continue to emerge and to evolve and cause both human and veterinary outbreaks owing to their ability to recombine, mutate, and infect multiple species and cell types.
Future research on coronaviruses will continue to investigate many aspects of viral replication and pathogenesis. First, understanding the propensity of these viruses to jump between species, to establish infection in a new host, and to identify significant reservoirs of coronaviruses will dramatically aid in our ability to predict when and where potential epidemics may occur. As bats seem to be a significant reservoir for these viruses, it will be interesting to determine how they seem to avoid clinically evident disease and become persistently infected. Second, many of the non-structural and accessory proteins encoded by these viruses remain uncharacterized with no known function, and it will be important to identify mechanisms of action for these proteins as well as defining their role in viral replication and pathogenesis. These studies should lead to a large increase in the number of suitable therapeutic targets to combat infections. Furthermore, many of the unique enzymes encoded by coronaviruses, such as ADP-ribose-1″-phosphatase, are also present in higher eukaryotes, making their study relevant to understanding general aspects of molecular biology and biochemistry. Third, gaining a complete picture of the intricacies of the RTC will provide a framework for understanding the unique RNA replication process used by these viruses. Finally, defining the mechanism of how coronaviruses cause disease and understanding the host immunopathological response will significantly improve our ability to design vaccines and reduce disease burden.
Contributor Information
Helena Jane Maier, Email: helena.maier@pirbright.ac.uk.
Erica Bickerton, Email: erica.bickerton@pirbright.ac.uk.
Paul Britton, Email: paul.britton@pirbright.ac.uk.
Stanley Perlman, Email: stanley-perlman@uiowa.edu
References
- 1.Zhao L, Jha BK, Wu A, et al. Antagonism of the interferon-induced OAS-RNase L pathway by murine coronavirus ns2 protein is required for virus replication and liver pathology. Cell Host Microbe. 2012;11:607–616. doi: 10.1016/j.chom.2012.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Barcena M, Oostergetel GT, Bartelink W, et al. Cryo-electron tomography of mouse hepatitis virus: insights into the structure of the coronavirion. Proc Natl Acad Sci U S A. 2009;106:582–587. doi: 10.1073/pnas.0805270106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Neuman BW, Adair BD, Yoshioka C, et al. Supramolecular architecture of severe acute respiratory syndrome coronavirus revealed by electron cryomicroscopy. J Virol. 2006;80:7918–7928. doi: 10.1128/JVI.00645-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Beniac DR, Andonov A, Grudeski E, et al. Architecture of the SARS coronavirus prefusion spike. Nat Struct Mol Biol. 2006;13:751–752. doi: 10.1038/nsmb1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Delmas B, Laude H. Assembly of coronavirus spike protein into trimers and its role in epitope expression. J Virol. 1990;64:5367–5375. doi: 10.1128/jvi.64.11.5367-5375.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bosch BJ, van der Zee R, de Haan CA, et al. The coronavirus spike protein is a class I virus fusion protein: structural and functional characterization of the fusion core complex. J Virol. 2003;77:8801–8811. doi: 10.1128/JVI.77.16.8801-8811.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Collins AR, Knobler RL, Powell H, et al. Monoclonal antibodies to murine hepatitis virus-4 (strain JHM) define the viral glycoprotein responsible for attachment and cell–cell fusion. Virology. 1982;119:358–371. doi: 10.1016/0042-6822(82)90095-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Abraham S, Kienzle TE, Lapps W, et al. Deduced sequence of the bovine coronavirus spike protein and identification of the internal proteolytic cleavage site. Virology. 1990;176:296–301. doi: 10.1016/0042-6822(90)90257-R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Luytjes W, Sturman LS, Bredenbeek PJ, et al. Primary structure of the glycoprotein E2 of coronavirus MHV-A59 and identification of the trypsin cleavage site. Virology. 1987;161:479–487. doi: 10.1016/0042-6822(87)90142-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.de Groot RJ, Luytjes W, Horzinek MC, et al. Evidence for a coiled-coil structure in the spike proteins of coronaviruses. J Mol Biol. 1987;196:963–966. doi: 10.1016/0022-2836(87)90422-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Armstrong J, Niemann H, Smeekens S, et al. Sequence and topology of a model intracellular membrane protein, E1 glycoprotein, from a coronavirus. Nature. 1984;308:751–752. doi: 10.1038/308751a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Nal B, Chan C, Kien F, et al. Differential maturation and subcellular localization of severe acute respiratory syndrome coronavirus surface proteins S, M and E. J Gen Virol. 2005;86:1423–1434. doi: 10.1099/vir.0.80671-0. [DOI] [PubMed] [Google Scholar]
- 13.Neuman BW, Kiss G, Kunding AH, et al. A structural analysis of M protein in coronavirus assembly and morphology. J Struct Biol. 2011;174:11–22. doi: 10.1016/j.jsb.2010.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Godet M, L’Haridon R, Vautherot JF, et al. TGEV corona virus ORF4 encodes a membrane protein that is incorporated into virions. Virology. 1992;188:666–675. doi: 10.1016/0042-6822(92)90521-P. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.DeDiego ML, Alvarez E, Almazan F, et al. A severe acute respiratory syndrome coronavirus that lacks the E gene is attenuated in vitro and in vivo. J Virol. 2007;81:1701–1713. doi: 10.1128/JVI.01467-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nieto-Torres JL, Dediego ML, Verdia-Baguena C, et al. Severe acute respiratory syndrome coronavirus envelope protein ion channel activity promotes virus fitness and pathogenesis. PLoS Pathog. 2014;10:e1004077. doi: 10.1371/journal.ppat.1004077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chang CK, Sue SC, Yu TH, et al. Modular organization of SARS coronavirus nucleocapsid protein. J Biomed Sci. 2006;13:59–72. doi: 10.1007/s11373-005-9035-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hurst KR, Koetzner CA, Masters PS. Identification of in vivo-interacting domains of the murine coronavirus nucleocapsid protein. J Virol. 2009;83:7221–7234. doi: 10.1128/JVI.00440-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stohlman SA, Lai MM. Phosphoproteins of murine hepatitis viruses. J Virol. 1979;32:672–675. doi: 10.1128/jvi.32.2.672-675.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Stohlman SA, Baric RS, Nelson GN, et al. Specific interaction between coronavirus leader RNA and nucleocapsid protein. J Virol. 1988;62:4288–4295. doi: 10.1128/jvi.62.11.4288-4295.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Molenkamp R, Spaan WJ. Identification of a specific interaction between the coronavirus mouse hepatitis virus A59 nucleocapsid protein and packaging signal. Virology. 1997;239:78–86. doi: 10.1006/viro.1997.8867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kuo L, Masters PS. Functional analysis of the murine coronavirus genomic RNA packaging signal. J Virol. 2013;87:5182–5192. doi: 10.1128/JVI.00100-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hurst KR, Koetzner CA, Masters PS. Characterization of a critical interaction between the coronavirus nucleocapsid protein and nonstructural protein 3 of the viral replicase-transcriptase complex. J Virol. 2013;87:9159–9172. doi: 10.1128/JVI.01275-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sturman LS, Holmes KV, Behnke J. Isolation of coronavirus envelope glycoproteins and interaction with the viral nucleocapsid. J Virol. 1980;33:449–462. doi: 10.1128/jvi.33.1.449-462.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Klausegger A, Strobl B, Regl G, et al. Identification of a coronavirus hemagglutinin-esterase with a substrate specificity different from those of influenza C virus and bovine coronavirus. J Virol. 1999;73:3737–3743. doi: 10.1128/jvi.73.5.3737-3743.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cornelissen LA, Wierda CM, van der Meer FJ, et al. Hemagglutinin-esterase, a novel structural protein of torovirus. J Virol. 1997;71:5277–5286. doi: 10.1128/jvi.71.7.5277-5286.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kazi L, Lissenberg A, Watson R, et al. Expression of hemagglutinin esterase protein from recombinant mouse hepatitis virus enhances neurovirulence. J Virol. 2005;79:15064–15073. doi: 10.1128/JVI.79.24.15064-15073.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lissenberg A, Vrolijk MM, van Vliet AL, et al. Luxury at a cost? Recombinant mouse hepatitis viruses expressing the accessory hemagglutinin esterase protein display reduced fitness in vitro. J Virol. 2005;79:15054–15063. doi: 10.1128/JVI.79.24.15054-15063.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kubo H, Yamada YK, Taguchi F. Localization of neutralizing epitopes and the receptor-binding site within the amino-terminal 330 amino acids of the murine coronavirus spike protein. J Virol. 1994;68:5403–5410. doi: 10.1128/jvi.68.9.5403-5410.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cheng PK, Wong DA, Tong LK, et al. Viral shedding patterns of coronavirus in patients with probable severe acute respiratory syndrome. Lancet. 2004;363:1699–1700. doi: 10.1016/S0140-6736(04)16255-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Belouzard S, Chu VC, Whittaker GR. Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proc Natl Acad Sci U S A. 2009;106:5871–5876. doi: 10.1073/pnas.0809524106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Baranov PV, Henderson CM, Anderson CB, et al. Programmed ribosomal frameshifting in decoding the SARS-CoV genome. Virology. 2005;332:498–510. doi: 10.1016/j.virol.2004.11.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Brierley I, Digard P, Inglis SC. Characterization of an efficient coronavirus ribosomal frameshifting signal: requirement for an RNA pseudoknot. Cell. 1989;57:537–547. doi: 10.1016/0092-8674(89)90124-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Araki K, Gangappa S, Dillehay DL, et al. Pathogenic virus-specific T cells cause disease during treatment with the calcineurin inhibitor FK506: implications for transplantation. J Exp Med. 2010;207:2355–2367. doi: 10.1084/jem.20100124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ziebuhr J, Snijder EJ, Gorbalenya AE. Virus-encoded proteinases and proteolytic processing in the Nidovirales. J Gen Virol. 2000;81:853–879. doi: 10.1099/0022-1317-81-4-853. [DOI] [PubMed] [Google Scholar]
- 36.Mielech AM, Chen Y, Mesecar AD, et al. Nidovirus papain-like proteases: multifunctional enzymes with protease, deubiquitinating and deISGylating activities. Virus Res. 2014 doi: 10.1016/j.virusres.2014.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Snijder EJ, Bredenbeek PJ, Dobbe JC, et al. Unique and conserved features of genome and proteome of SARS-coronavirus, an early split-off from the coronavirus group 2 lineage. J Mol Biol. 2003;331:991–1004. doi: 10.1016/S0022-2836(03)00865-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sethna PB, Hofmann MA, Brian DA. Minus-strand copies of replicating coronavirus mRNAs contain antileaders. J Virol. 1991;65:320–325. doi: 10.1128/jvi.65.1.320-325.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Brown CG, Nixon KS, Senanayake SD, et al. An RNA stem-loop within the bovine coronavirus nsp1 coding region is a cis-acting element in defective interfering RNA replication. J Virol. 2007;81:7716–7724. doi: 10.1128/JVI.00549-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Guan BJ, Wu HY, Brian DA. An optimal cis-replication stem-loop IV in the 5′ untranslated region of the mouse coronavirus genome extends 16 nucleotides into open reading frame 1. J Virol. 2011;85:5593–5605. doi: 10.1128/JVI.00263-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Liu P, Li L, Keane SC, et al. Mouse hepatitis virus stem-loop 2 adopts a uYNMG(U)a-like tetraloop structure that is highly functionally tolerant of base substitutions. J Virol. 2009;83:12084–12093. doi: 10.1128/JVI.00915-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Raman S, Bouma P, Williams GD, et al. Stem-loop III in the 5′ untranslated region is a cis-acting element in bovine coronavirus defective interfering RNA replication. J Virol. 2003;77:6720–6730. doi: 10.1128/JVI.77.12.6720-6730.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Liu Q, Johnson RF, Leibowitz JL. Secondary structural elements within the 3′ untranslated region of mouse hepatitis virus strain JHM genomic RNA. J Virol. 2001;75:12105–12113. doi: 10.1128/JVI.75.24.12105-12113.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Goebel SJ, Miller TB, Bennett CJ, et al. A hypervariable region within the 3′ cis-acting element of the murine coronavirus genome is nonessential for RNA synthesis but affects pathogenesis. J Virol. 2007;81:1274–1287. doi: 10.1128/JVI.00803-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Williams GD, Chang RY, Brian DA. A phylogenetically conserved hairpin-type 3′ untranslated region pseudoknot functions in coronavirus RNA replication. J Virol. 1999;73:8349–8355. doi: 10.1128/jvi.73.10.8349-8355.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hsue B, Masters PS. A bulged stem-loop structure in the 3′ untranslated region of the genome of the coronavirus mouse hepatitis virus is essential for replication. J Virol. 1997;71:7567–7578. doi: 10.1128/jvi.71.10.7567-7578.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hsue B, Hartshorne T, Masters PS. Characterization of an essential RNA secondary structure in the 3′ untranslated region of the murine coronavirus genome. J Virol. 2000;74:6911–6921. doi: 10.1128/jvi.74.15.6911-6921.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sawicki SG, Sawicki DL, Siddell SG. A contemporary view of coronavirus transcription. J Virol. 2007;81:20–29. doi: 10.1128/JVI.01358-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bentley K, Keep SM, Armesto M, et al. Identification of a noncanonically transcribed subgenomic mRNA of infectious bronchitis virus and other gammacoronaviruses. J Virol. 2013;87:2128–2136. doi: 10.1128/JVI.02967-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Keck JG, Makino S, Soe LH, et al. RNA recombination of coronavirus. Adv Exp Med Biol. 1987;218:99–107. doi: 10.1007/978-1-4684-1280-2_11. [DOI] [PubMed] [Google Scholar]
- 51.Lai MM, Baric RS, Makino S, et al. Recombination between nonsegmented RNA genomes of murine coronaviruses. J Virol. 1985;56:449–456. doi: 10.1128/jvi.56.2.449-456.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Krijnse-Locker J, Ericsson M, Rottier PJM, et al. Characterization of the budding compartment of mouse hepatitis virus: evidence that transport from the RER to the Golgi complex requires only one vesicular transport step. J Cell Biol. 1994;124:55–70. doi: 10.1083/jcb.124.1.55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Tooze J, Tooze S, Warren G. Replication of coronavirus MHV-A59 in sac-cells: determination of the first site of budding of progeny virions. Eur J Cell Biol. 1984;33:281–293. [PubMed] [Google Scholar]
- 54.de Haan CA, Rottier PJ. Molecular interactions in the assembly of coronaviruses. Adv Virus Res. 2005;64:165–230. doi: 10.1016/S0065-3527(05)64006-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Bos EC, Luytjes W, van der Meulen HV, et al. The production of recombinant infectious DI-particles of a murine coronavirus in the absence of helper virus. Virology. 1996;218:52–60. doi: 10.1006/viro.1996.0165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Siu YL, Teoh KT, Lo J, et al. The M, E, and N structural proteins of the severe acute respiratory syndrome coronavirus are required for efficient assembly, trafficking, and release of virus-like particles. J Virol. 2008;82:11318–11330. doi: 10.1128/JVI.01052-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Raamsman MJ, Locker JK, de Hooge A, et al. Characterization of the coronavirus mouse hepatitis virus strain A59 small membrane protein E. J Virol. 2000;74:2333–2342. doi: 10.1128/jvi.74.5.2333-2342.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Corse E, Machamer CE. Infectious bronchitis virus E protein is targeted to the Golgi complex and directs release of virus-like particles. J Virol. 2000;74:4319–4326. doi: 10.1128/jvi.74.9.4319-4326.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Fischer F, Stegen CF, Masters PS, et al. Analysis of constructed E gene mutants of mouse hepatitis virus confirms a pivotal role for E protein in coronavirus assembly. J Virol. 1998;72:7885–7894. doi: 10.1128/jvi.72.10.7885-7894.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Boscarino JA, Logan HL, Lacny JJ, et al. Envelope protein palmitoylations are crucial for murine coronavirus assembly. J Virol. 2008;82:2989–2999. doi: 10.1128/JVI.01906-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ye Y, Hogue BG. Role of the coronavirus E viroporin protein transmembrane domain in virus assembly. J Virol. 2007;81:3597–3607. doi: 10.1128/JVI.01472-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hurst KR, Kuo L, Koetzner CA, et al. A major determinant for membrane protein interaction localizes to the carboxy-terminal domain of the mouse coronavirus nucleocapsid protein. J Virol. 2005;79:13285–13297. doi: 10.1128/JVI.79.21.13285-13297.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Perlman S, Netland J. Coronaviruses post-SARS: update on replication and pathogenesis. Nat Rev Microbiol. 2009;7:439–450. doi: 10.1038/nrmicro2147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mihindukulasuriya KA, Wu G, St LJ, et al. Identification of a novel coronavirus from a beluga whale by using a panviral microarray. J Virol. 2008;82:5084–5088. doi: 10.1128/JVI.02722-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.He B, Zhang Y, Xu L, et al. Identification of diverse alphacoronaviruses and genomic characterization of a novel severe acute respiratory syndrome-like coronavirus from bats in china. J Virol. 2014;88:7070–7082. doi: 10.1128/JVI.00631-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Nga PT, Parquet Mdel C, Lauber C, et al. Discovery of the first insect nidovirus, a missing evolutionary link in the emergence of the largest RNA virus genomes. PLoS Pathog. 2011;7:e1002215. doi: 10.1371/journal.ppat.1002215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lauber C, Ziebuhr J, Junglen S, et al. Mesoniviridae: a proposed new family in the order Nidovirales formed by a single species of mosquito-borne viruses. Arch Virol. 2012;157:1623–1628. doi: 10.1007/s00705-012-1295-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Levy GA, Liu M, Ding J, et al. Molecular and functional analysis of the human prothrombinase gene (HFGL2) and its role in viral hepatitis. Am J Pathol. 2000;156:1217–1225. doi: 10.1016/S0002-9440(10)64992-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lampert PW, Sims JK, Kniazeff AJ. Mechanism of demyelination in JHM virus encephalomyelitis. Acta Neuropathol. 1973;24:76–85. doi: 10.1007/BF00691421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Weiner LP. Pathogenesis of demyelination induced by a mouse hepatitis virus (JHM virus) Arch Neurol. 1973;28:298–303. doi: 10.1001/archneur.1973.00490230034003. [DOI] [PubMed] [Google Scholar]
- 71.Wu GF, Dandekar AA, Pewe L, et al. CD4 and CD8 T cells have redundant but not identical roles in virus-induced demyelination. J Immunol. 2000;165:2278–2286. doi: 10.4049/jimmunol.165.4.2278. [DOI] [PubMed] [Google Scholar]
- 72.Wang F, Stohlman SA, Fleming JO. Demyelination induced by murine hepatitis virus JHM strain (MHV-4) is immunologically mediated. J Neuroimmunol. 1990;30:31–41. doi: 10.1016/0165-5728(90)90050-W. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Wu GF, Perlman S. Macrophage infiltration, but not apoptosis, is correlated with immune-mediated demyelination following murine infection with a neurotropic coronavirus. J Virol. 1999;73:8771–8780. doi: 10.1128/jvi.73.10.8771-8780.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.McIntosh K, Becker WB, Chanock RM. Growth in suckling-mouse brain of “IBV-like” viruses from patients with upper respiratory tract disease. Proc Natl Acad Sci U S A. 1967;58:2268–2273. doi: 10.1073/pnas.58.6.2268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Bradburne AF, Bynoe ML, Tyrell DAJ. Effects of a “new” human respiratory virus in volunteers. Br Med J. 1967;3:767–769. doi: 10.1136/bmj.3.5568.767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hamre D, Procknow JJ. A new virus isolated from the human respiratory tract. Proc Soc Exp Biol Med. 1966;121:190–193. doi: 10.3181/00379727-121-30734. [DOI] [PubMed] [Google Scholar]
- 77.Woo PC, Lau SK, Chu CM, et al. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J Virol. 2005;79:884–895. doi: 10.1128/JVI.79.2.884-895.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.van der Hoek L, Pyrc K, Jebbink MF, et al. Identification of a new human coronavirus. Nat Med. 2004;10:368–373. doi: 10.1038/nm1024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.van der Hoek L, Sure K, Ihorst G, et al. Croup is associated with the novel coronavirus NL63. PLoS Med. 2005;2:e240. doi: 10.1371/journal.pmed.0020240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Chibo D, Birch C. Analysis of human coronavirus 229E spike and nucleoprotein genes demonstrates genetic drift between chronologically distinct strains. J Gen Virol. 2006;87:1203–1208. doi: 10.1099/vir.0.81662-0. [DOI] [PubMed] [Google Scholar]
- 81.Vijgen L, Keyaerts E, Lemey P, et al. Circulation of genetically distinct contemporary human coronavirus OC43 strains. Virology. 2005;337:85–92. doi: 10.1016/j.virol.2005.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Guan Y, Zheng BJ, He YQ, et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science. 2003;302:276–278. doi: 10.1126/science.1087139. [DOI] [PubMed] [Google Scholar]
- 83.Lau SK, Woo PC, Li KS, et al. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc Natl Acad Sci U S A. 2005;102:14040–14045. doi: 10.1073/pnas.0506735102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Li W, Shi Z, Yu M, et al. Bats are natural reservoirs of SARS-like coronaviruses. Science. 2005;310:676–679. doi: 10.1126/science.1118391. [DOI] [PubMed] [Google Scholar]
- 85.Ge XY, Li JL, Yang XL, et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature. 2013;503:535–538. doi: 10.1038/nature12711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Peiris JS, Yuen KY, Osterhaus AD, et al. The severe acute respiratory syndrome. N Engl J Med. 2003;349:2431–2441. doi: 10.1056/NEJMra032498. [DOI] [PubMed] [Google Scholar]
- 87.Peiris JS, Chu CM, Cheng VC, et al. Clinical progression and viral load in a community outbreak of coronavirus-associated SARS pneumonia: a prospective study. Lancet. 2003;361:1767–1772. doi: 10.1016/S0140-6736(03)13412-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Spiegel M, Schneider K, Weber F, et al. Interaction of severe acute respiratory syndrome-associated coronavirus with dendritic cells. J Gen Virol. 2006;87:1953–1960. doi: 10.1099/vir.0.81624-0. [DOI] [PubMed] [Google Scholar]
- 89.Law HK, Cheung CY, Ng HY, et al. Chemokine upregulation in SARS coronavirus infected human monocyte derived dendritic cells. Blood. 2005;106:2366–2376. doi: 10.1182/blood-2004-10-4166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Lau YL, Peiris JSM. Pathogenesis of severe acute respiratory syndrome. Curr Opin Immunol. 2005;17:404–410. doi: 10.1016/j.coi.2005.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Roberts A, Paddock C, Vogel L, et al. Aged BALB/c mice as a model for increased severity of severe acute respiratory syndrome in elderly humans. J Virol. 2005;79:5833–5838. doi: 10.1128/JVI.79.9.5833-5838.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Zhao J, Zhao J, Perlman S. T cell responses are required for protection from clinical disease and for virus clearance in severe acute respiratory syndrome coronavirus-infected mice. J Virol. 2010;84:9318–9325. doi: 10.1128/JVI.01049-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Zhao J, Zhao J, Legge K, et al. Age-related increases in PGD(2) expression impair respiratory DC migration, resulting in diminished T cell responses upon respiratory virus infection in mice. J Clin Invest. 2011;121:4921–4930. doi: 10.1172/JCI59777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Zaki AM, van Boheemen S, Bestebroer TM, et al. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med. 2012;367:1814–1820. doi: 10.1056/NEJMoa1211721. [DOI] [PubMed] [Google Scholar]
- 95.van Boheemen S, de Graaf M, Lauber C et al (2012) Genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans. MBio 3. doi:10.1128/mBio.00473-12 [DOI] [PMC free article] [PubMed]
- 96.Meyer B, Muller MA, Corman VM, et al. Antibodies against MERS coronavirus in dromedary camels, United Arab Emirates, 2003 and 2013. Emerg Infect Dis. 2014;20:552–559. doi: 10.3201/eid2004.131746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Eckerle I, Corman VM, Muller MA, et al. Replicative capacity of MERS coronavirus in livestock cell lines. Emerg Infect Dis. 2014;20:276–279. doi: 10.3201/eid2002.131182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Memish ZA, Cotten M, Meyer B, et al. Human infection with MERS coronavirus after exposure to infected camels, Saudi Arabia, 2013. Emerg Infect Dis. 2014;20:1012–1015. doi: 10.3201/eid2006.140402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Azhar EI, El-Kafrawy SA, Farraj SA, et al. Evidence for camel-to-human transmission of MERS coronavirus. N Engl J Med. 2014 doi: 10.1056/NEJMoa1401505. [DOI] [PubMed] [Google Scholar]
- 100.Raj VS, Mou H, Smits SL, et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature. 2013;495:251–254. doi: 10.1038/nature12005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Zhao J, Li K, Wohlford-Lenane C, et al. Rapid generation of a mouse model for Middle East respiratory syndrome. Proc Natl Acad Sci U S A. 2014;111:4970–4975. doi: 10.1073/pnas.1323279111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Emery SL, Erdman DD, Bowen MD, et al. Real-time reverse transcription-polymerase chain reaction assay for SARS-associated coronavirus. Emerg Infect Dis. 2004;10:311–316. doi: 10.3201/eid1002.030759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Gaunt ER, Hardie A, Claas EC, et al. Epidemiology and clinical presentations of the four human coronaviruses 229E, HKU1, NL63, and OC43 detected over 3 years using a novel multiplex real-time PCR method. J Clin Microbiol. 2010;48:2940–2947. doi: 10.1128/JCM.00636-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Cinatl J, Morgenstern B, Bauer G, et al. Treatment of SARS with human interferons. Lancet. 2003;362:293–294. doi: 10.1016/S0140-6736(03)13973-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Stockman LJ, Bellamy R, Garner P. SARS: systematic review of treatment effects. PLoS Med. 2006;3:e343. doi: 10.1371/journal.pmed.0030343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Laude H, Van Reeth K, Pensaert M. Porcine respiratory coronavirus: molecular features and virus-host interactions. Vet Res. 1993;24:125–150. [PubMed] [Google Scholar]
- 107.Saif LJ. Animal coronavirus vaccines: lessons for SARS. Dev Biol (Basel) 2004;119:129–140. [PubMed] [Google Scholar]
- 108.Wang L, Junker D, Collisson EW. Evidence of natural recombination within the S1 gene of infectious bronchitis virus. Virology. 1993;192:710–716. doi: 10.1006/viro.1993.1093. [DOI] [PubMed] [Google Scholar]
- 109.Vennema H, de Groot RJ, Harbour DA, et al. Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization. J Virol. 1990;64:1407–1409. doi: 10.1128/jvi.64.3.1407-1409.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Zust R, Cervantes-Barragan L, Kuri T, et al. Coronavirus non-structural protein 1 is a major pathogenicity factor: implications for the rational design of coronavirus vaccines. PLoS Pathog. 2007;3:e109. doi: 10.1371/journal.ppat.0030109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Netland J, DeDiego ML, Zhao J, et al. Immunization with an attenuated severe acute respiratory syndrome coronavirus deleted in E protein protects against lethal respiratory disease. Virology. 2010;399:120–128. doi: 10.1016/j.virol.2010.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.de Haan CA, Volders H, Koetzner CA, et al. Coronaviruses maintain viability despite dramatic rearrangements of the strictly conserved genome organization. J Virol. 2002;76:12491–12502. doi: 10.1128/JVI.76.24.12491-12502.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Yount B, Roberts RS, Lindesmith L, et al. Rewiring the severe acute respiratory syndrome coronavirus (SARS-CoV) transcription circuit: engineering a recombination-resistant genome. Proc Natl Acad Sci U S A. 2006;103:12546–12551. doi: 10.1073/pnas.0605438103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Graham RL, Becker MM, Eckerle LD, et al. A live, impaired-fidelity coronavirus vaccine protects in an aged, immunocompromised mouse model of lethal disease. Nat Med. 2012;18:1820–1826. doi: 10.1038/nm.2972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Yeager CL, Ashmun RA, Williams RK, et al. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature. 1992;357:420–422. doi: 10.1038/357420a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Hofmann H, Pyrc K, van der Hoek L, et al. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci U S A. 2005;102:7988–7993. doi: 10.1073/pnas.0409465102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Delmas B, Gelfi J, L’Haridon R, et al. Aminopeptidase N is a major receptor for the entero-pathogenic coronavirus TGEV. Nature. 1992;357:417–420. doi: 10.1038/357417a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Li BX, Ge JW, Li YJ. Porcine aminopeptidase N is a functional receptor for the PEDV coronavirus. Virology. 2007;365:166–172. doi: 10.1016/j.virol.2007.03.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Tresnan DB, Levis R, Holmes KV. Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I. J Virol. 1996;70:8669–8674. doi: 10.1128/jvi.70.12.8669-8674.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Benbacer L, Kut E, Besnardeau L, et al. Interspecies aminopeptidase-N chimeras reveal species-specific receptor recognition by canine coronavirus, feline infectious peritonitis virus, and transmissible gastroenteritis virus. J Virol. 1997;71:734–737. doi: 10.1128/jvi.71.1.734-737.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Nedellec P, Dveksler GS, Daniels E, et al. Bgp2, a new member of the carcinoembryonic antigen-related gene family, encodes an alternative receptor for mouse hepatitis viruses. J Virol. 1994;68:4525–4537. doi: 10.1128/jvi.68.7.4525-4537.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Williams RK, Jiang GS, Holmes KV. Receptor for mouse hepatitis virus is a member of the carcinoembryonic antigen family of glycoproteins. Proc Natl Acad Sci U S A. 1991;88:5533–5536. doi: 10.1073/pnas.88.13.5533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Schultze B, Herrler G. Bovine coronavirus uses N-acetyl-9-O-acetylneuraminic acid as a receptor determinant to initiate the infection of cultured cells. J Gen Virol. 1992;73(Pt 4):901–906. doi: 10.1099/0022-1317-73-4-901. [DOI] [PubMed] [Google Scholar]
- 124.Li W, Moore MJ, Vasilieva N, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426:450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Huang C, Lokugamage KG, Rozovics JM, et al. Alphacoronavirus transmissible gastroenteritis virus nsp1 protein suppresses protein translation in mammalian cells and in cell-free HeLa cell extracts but not in rabbit reticulocyte lysate. J Virol. 2011;85:638–643. doi: 10.1128/JVI.01806-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Kamitani W, Huang C, Narayanan K, et al. A two-pronged strategy to suppress host protein synthesis by SARS coronavirus Nsp1 protein. Nat Struct Mol Biol. 2009;16:1134–1140. doi: 10.1038/nsmb.1680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Kamitani W, Narayanan K, Huang C, et al. Severe acute respiratory syndrome coronavirus nsp1 protein suppresses host gene expression by promoting host mRNA degradation. Proc Natl Acad Sci U S A. 2006;103:12885–12890. doi: 10.1073/pnas.0603144103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Tanaka T, Kamitani W, DeDiego ML, et al. Severe acute respiratory syndrome coronavirus nsp1 facilitates efficient propagation in cells through a specific translational shutoff of host mRNA. J Virol. 2012;86:11128–11137. doi: 10.1128/JVI.01700-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Graham RL, Sims AC, Brockway SM, et al. The nsp2 replicase proteins of murine hepatitis virus and severe acute respiratory syndrome coronavirus are dispensable for viral replication. J Virol. 2005;79:13399–13411. doi: 10.1128/JVI.79.21.13399-13411.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Cornillez-Ty CT, Liao L, Yates JR, 3rd, et al. Severe acute respiratory syndrome coronavirus nonstructural protein 2 interacts with a host protein complex involved in mitochondrial biogenesis and intracellular signaling. J Virol. 2009;83:10314–10318. doi: 10.1128/JVI.00842-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Chatterjee A, Johnson MA, Serrano P, et al. Nuclear magnetic resonance structure shows that the severe acute respiratory syndrome coronavirus-unique domain contains a macrodomain fold. J Virol. 2009;83:1823–1836. doi: 10.1128/JVI.01781-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Egloff MP, Malet H, Putics A, et al. Structural and functional basis for ADP-ribose and poly(ADP-ribose) binding by viral macro domains. J Virol. 2006;80:8493–8502. doi: 10.1128/JVI.00713-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Eriksson KK, Cervantes-Barragan L, Ludewig B, et al. Mouse hepatitis virus liver pathology is dependent on ADP-ribose-1″-phosphatase, a viral function conserved in the alpha-like supergroup. J Virol. 2008;82:12325–12334. doi: 10.1128/JVI.02082-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Frieman M, Ratia K, Johnston RE, et al. Severe acute respiratory syndrome coronavirus papain-like protease ubiquitin-like domain and catalytic domain regulate antagonism of IRF3 and NF-kappaB signaling. J Virol. 2009;83:6689–6705. doi: 10.1128/JVI.02220-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Neuman BW, Joseph JS, Saikatendu KS, et al. Proteomics analysis unravels the functional repertoire of coronavirus nonstructural protein 3. J Virol. 2008;82:5279–5294. doi: 10.1128/JVI.02631-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Serrano P, Johnson MA, Almeida MS, et al. Nuclear magnetic resonance structure of the N-terminal domain of nonstructural protein 3 from the severe acute respiratory syndrome coronavirus. J Virol. 2007;81:12049–12060. doi: 10.1128/JVI.00969-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Serrano P, Johnson MA, Chatterjee A, et al. Nuclear magnetic resonance structure of the nucleic acid-binding domain of severe acute respiratory syndrome coronavirus nonstructural protein 3. J Virol. 2009;83:12998–13008. doi: 10.1128/JVI.01253-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Ziebuhr J, Thiel V, Gorbalenya AE. The autocatalytic release of a putative RNA virus transcription factor from its polyprotein precursor involves two paralogous papain-like proteases that cleave the same peptide bond. J Biol Chem. 2001;276:33220–33232. doi: 10.1074/jbc.M104097200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Clementz MA, Kanjanahaluethai A, O’Brien TE, et al. Mutation in murine coronavirus replication protein nsp4 alters assembly of double membrane vesicles. Virology. 2008;375:118–129. doi: 10.1016/j.virol.2008.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Gadlage MJ, Sparks JS, Beachboard DC, et al. Murine hepatitis virus nonstructural protein 4 regulates virus-induced membrane modifications and replication complex function. J Virol. 2010;84:280–290. doi: 10.1128/JVI.01772-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Lu Y, Lu X, Denison MR. Identification and characterization of a serine-like proteinase of the murine coronavirus MHV-A59. J Virol. 1995;69:3554–3559. doi: 10.1128/jvi.69.6.3554-3559.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Oostra M, Hagemeijer MC, van Gent M, et al. Topology and membrane anchoring of the coronavirus replication complex: not all hydrophobic domains of nsp3 and nsp6 are membrane spanning. J Virol. 2008;82:12392–12405. doi: 10.1128/JVI.01219-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Zhai Y, Sun F, Li X, et al. Insights into SARS-CoV transcription and replication from the structure of the nsp7-nsp8 hexadecamer. Nat Struct Mol Biol. 2005;12:980–986. doi: 10.1038/nsmb999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Imbert I, Guillemot JC, Bourhis JM, et al. A second, non-canonical RNA-dependent RNA polymerase in SARS coronavirus. EMBO J. 2006;25:4933–4942. doi: 10.1038/sj.emboj.7601368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Egloff MP, Ferron F, Campanacci V, et al. The severe acute respiratory syndrome-coronavirus replicative protein nsp9 is a single-stranded RNA-binding subunit unique in the RNA virus world. Proc Natl Acad Sci U S A. 2004;101:3792–3796. doi: 10.1073/pnas.0307877101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Bouvet M, Debarnot C, Imbert I, et al. In vitro reconstitution of SARS-coronavirus mRNA cap methylation. PLoS Pathog. 2010;6:e1000863. doi: 10.1371/journal.ppat.1000863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Decroly E, Debarnot C, Ferron F, et al. Crystal structure and functional analysis of the SARS-coronavirus RNA cap 2′-O-methyltransferase nsp10/nsp16 complex. PLoS Pathog. 2011;7:e1002059. doi: 10.1371/journal.ppat.1002059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Xu X, Liu Y, Weiss S, et al. Molecular model of SARS coronavirus polymerase: implications for biochemical functions and drug design. Nucleic Acids Res. 2003;31:7117–7130. doi: 10.1093/nar/gkg916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Ivanov KA, Thiel V, Dobbe JC, et al. Multiple enzymatic activities associated with severe acute respiratory syndrome coronavirus helicase. J Virol. 2004;78:5619–5632. doi: 10.1128/JVI.78.11.5619-5632.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Ivanov KA, Ziebuhr J. Human coronavirus 229E nonstructural protein 13: characterization of duplex-unwinding, nucleoside triphosphatase, and RNA 5′-triphosphatase activities. J Virol. 2004;78:7833–7838. doi: 10.1128/JVI.78.14.7833-7838.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Eckerle LD, Becker MM, Halpin RA, et al. Infidelity of SARS-CoV Nsp14-exonuclease mutant virus replication is revealed by complete genome sequencing. PLoS Pathog. 2010;6:e1000896. doi: 10.1371/journal.ppat.1000896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Eckerle LD, Lu X, Sperry SM, et al. High fidelity of murine hepatitis virus replication is decreased in nsp14 exoribonuclease mutants. J Virol. 2007;81:12135–12144. doi: 10.1128/JVI.01296-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Minskaia E, Hertzig T, Gorbalenya AE, et al. Discovery of an RNA virus 3′->5′ exoribonuclease that is critically involved in coronavirus RNA synthesis. Proc Natl Acad Sci U S A. 2006;103:5108–5113. doi: 10.1073/pnas.0508200103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Chen Y, Cai H, Pan J, et al. Functional screen reveals SARS coronavirus nonstructural protein nsp14 as a novel cap N7 methyltransferase. Proc Natl Acad Sci U S A. 2009;106:3484–3489. doi: 10.1073/pnas.0808790106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Bhardwaj K, Sun J, Holzenburg A, et al. RNA recognition and cleavage by the SARS coronavirus endoribonuclease. J Mol Biol. 2006;361:243–256. doi: 10.1016/j.jmb.2006.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Ivanov KA, Hertzig T, Rozanov M, et al. Major genetic marker of nidoviruses encodes a replicative endoribonuclease. Proc Natl Acad Sci U S A. 2004;101:12694–12699. doi: 10.1073/pnas.0403127101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Decroly E, Imbert I, Coutard B, et al. Coronavirus nonstructural protein 16 is a cap-0 binding enzyme possessing (nucleoside-2′O)-methyltransferase activity. J Virol. 2008;82:8071–8084. doi: 10.1128/JVI.00407-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Zust R, Cervantes-Barragan L, Habjan M, et al. Ribose 2′-O-methylation provides a molecular signature for the distinction of self and non-self mRNA dependent on the RNA sensor Mda5. Nat Immunol. 2011;12:137–143. doi: 10.1038/ni.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]