

Abstract
Background
The Central Asian flora plays a significant role in Eurasia and the Northern Hemisphere. Calophaca, a member of this flora, includes eight currently recognized species, and is centered in Central Asia, with some taxa extending into adjacent areas. A phylogenetic analysis of the genus utilizing nuclear ribosomal ITS and plastid trnS-trnG and rbcL sequences was carried out in order to confirm its taxonomic status and reconstruct its evolutionary history.
Methodology/Principal Finding
We employed BEAST Bayesian inference for dating, and S-DIVA and BBM for ancestral area reconstruction, to study its spatiotemporal evolution. Our results show that Calophacais monophyletic and nested within Caragana. The divergence time of Calophaca is estimated at ca. 8.0 Ma, most likely driven by global cooling and aridification, influenced by rapid uplift of the Qinghai Tibet Plateau margins.
Conclusions/Significance
According to ancestral area reconstructions, the genus most likely originated in the Pamir Mountains, a global biodiversity hotspot and hypothesized Tertiary refugium of many Central Asian plant lineages. Dispersals from this location are inferred to the western Tianshan Mountains, then northward to the Tarbagatai Range, eastward to East Asia, and westward to the Caucasus, Russia, and Europe. The spatiotemporal evolution of Calophaca provides a case contributing to an understanding of the flora and biodiversity of the Central Asian mountains and adjacent regions.
Introduction
The Himalaya Range and Qinghai Tibet Plateau (QTP), as well as the associated Pamir and Tianshan mountains in Asia, have developed unparalleled alpine and montane floras, and therefore often have served as a source for plants adapted to higher elevations, even affecting the Alps of Europe[1–6]. Arid portions of these mountain ranges have developed drought adapted montane floras and enabled xeric plants to disperse more widely over geologic time [7,8]. However, this issue has received surprisingly little examination by modern methods of phylogenetic and biogeographical reconstruction [4].
From geological history perspectives, at the uplift of the QTP and withdrawal of the inland Paratethys Sea during the late Eocene [9], pronounced aridity appears to have begun in Central Asia [9], followed by a switch to monsoonal circulatory patterns, resulting in arid winter monsoon winds in the lee of the QTP in Central Asia by late Oligocene or early Miocene [10,11]. These circumstances resulted in the evolution of a well developed dryland Central Asian flora, which has yielded different evolutionary hypotheses, such as origins locally or from the Mediterranean, Africa, East Asia, or the QTP [1,4,6,7,12]. It appears that some of the elements of this flora proved to be preadapted as aridity increased across Eurasia in connection with global cooling during the Miocene and Pliocene [11,13]. Case studies of members of the Central Asian flora to test these evolutionary hypotheses, such as that of Artemisia (Asteraceae) [13], with linkage to paleogeography and paleoclimate events, and providing exact descriptions of spatiotemporal evolution, have to date been very few.
Calophaca Fisch. ex DC. is an ideal taxonomic group to investigate the Central Asian flora. It includes small trees or shrubs and eight currently described species distributed in semiarid montane and steppe environments, chiefly in Central Asia, with one species extending westward to southeastern Europe and Russia, and another in northern China [14–18]. The distribution of Calophaca occupies several global biodiversity conservation hotspots [19], i.e., the Pamir and western Tianshan mountains, the Caucasus, and East Asia, see Fig 1.
Fig 1. Distribution of Calophaca species as estimated from our field investigations in China, and floras and herbaria (PE, XIJI, LE, MW, MAH).
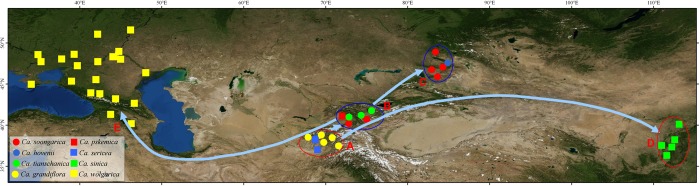
The place of origin of the genus, the Pamir—western Tianshan, and dispersal routes in three different directions are illustrated.
Regarding Calophaca systematics, Borissova [14] classified Calophaca in two series, Pubiflorae Boriss., with pubescence on the outside of the standard, and Glabriflorae Boriss., with the standard glabrous. Calophaca ser. Glabriflorae contains two species, C. grandiflora and C. sericea, and C. ser. Pubiflorae contains all of the remaining species of the genus except for C. sinica. Gorbunova [15] elevated these series as sections: sect. Glabriflora and sect. Calophaca (Table 1). According to traditional taxonomy [20], the genera Calophaca, Halimodendron, and Caragana belonged to tribe Galegeae. However, these genera were recently transferred to tribe Hedysareae on the basis of molecular evidence [21]. Chromosome data shows that Calophaca possesses a chromosome base number of x = 8, similar to Caragana and Astragalus. Of the species of the genus that have been sampled for chromosome number, C. wolgarica, C. kultiassovii (= C. tianschanica), and C. sinica are diploid [22–24], and C. soongorica is tetraploid [24]. Li et al. [24] and Chang et al. [25] studied the chromosomes, leaf epidermal features, pollen, carpodermis, and distribution of C. sinica and C. soongorica. They hypothesized that the narrow present distribution of C. sinica resulted from a gradual contraction of its distribution range accompanying the intensification of aridity since the Tertiary [25]. Considering the higher species density of Calophaca in Central Asia, some authors have proposed that this genus is a Tertiary relict from the region [25,26]. Molecular approaches coupled with phylogenetic and biogeographical reconstruction may shed more light on the still obscure systematic and evolutionary issues in Calophaca and Central Asian biodiversity, however, these have been lacking so far.
Table 1. Voucher information for Calophaca and four genera as the outgroups.
| Taxon | Voucher | Source | GenBank accession number (ITS, rbcL, trnS-G) |
|---|---|---|---|
| Calophaca | |||
| Sect. Glabriflora (Boriss.) Gorb. | |||
| Calophaca grandiflora Regel | M.G. Vasileva 5.8.1983 (MW) | Between rivers Kyzylsu and Yakhsu, Kushtek, S. Tajikistan | KP862569, KP862557, KP862545 |
| C. sericea B. Fedtsch. ex. Boriss. | E.A. Ra 28.7.1949 (LE) | Pamir, Tajikistan | KP862571, KP862559, KP862547 |
| Sect. Calophaca | |||
| C. tianshanica (B. Fedtsch.) Boriss. | I.I. Rusanovich 6.06.1989 (MHA) | Algabas, Shymkent, Kazakhstan | KP862578, KP862566, KP862554 |
| C. pskemica Gorbunova | L. Popova 7.6.1977 (MW) | Sandalash, Pskem, Kyrgistan | KP862581,————,———— |
| C. soongorica Kar. & Kir. | E.E. Pyoahobeq & L.A. Kpamapehko 5-14-1984 (PE) | Semiipalatinskaya, Kazakhstan | FJ537288, FJ537236, FJ537189 |
| C. soongorica Kar. & Kir.1 | H.X. Zhang et al.20110502 (XIJI) | Tacheng, Xinjiang, China | KP862574, KP862562, KP862550 |
| C. soongorica Kar. & Kir. 2 | H.X. Zhang et al. 20110602 (XIJI) | Tacheng, Xinjiang, China | KP862575, KP862563, KP862551 |
| C. soongorica Kar. & Kir. 3 | M. Pimenova, L. Kamenskih, L. Sdobnina 21.7.1975 (MW) | W. Tarbagatay, Saysu, Kazakhstan | KP862576, KP862564, KP862552 |
| C. soongorica Kar. & Kir. 4 | V.I.Grubov 25.06.1986 (LE) | Ili-Balkhash, Ayagoz, Kazakhstan | KP862577, KP862565, KP862553 |
| C. hovenii Schrenk. | E. Kluykov 22.8.1979 (MW) | Tarbagatay, Urzharsky, Samipalatinsk, Kazakhstan | KP862570, KP862558, KP862546 |
| C. wolgarica (L.f.) DC. 1 | M.G. Pimenov 10.06.2005 (MW) | Ergeney, Kalmykia, Russia | KP862579, KP862567, KP862555 |
| C. wolgarica (L.f.) DC. 2 | N. Bintapu 25.05.1980 (LE) | Caucasus, Russia | KP862580, KP862568, KP862556 |
| Sect. Trichomeae M.L. Zhang b | |||
| C. sinica Rehd. 1 | J.F. Huang 2010020 (XIJI) | Jiaocheng, Shanxi, China | KP862572, KP862560, KP862548 |
| C. sinica Rehd. 2 | J.F. Huang 2010022 (XIJI) | Jiaocheng, Shanxi, China | KP862573, KP862561, KP862549 |
| Outgroups | |||
| Caragana | |||
| Sect. Caragana | |||
| Ser. Caragana | |||
| Car. arborescens Lam. | M.L. Zhang 00–201 (PE) | Altai, Xinjiang, China | FJ537262, FJ537211, FJ537164 |
| Car. boisii C. K. Schneid. | M.L. Zhang & Y. Kang 00–121 (PE) | Lixian, Sichuan, China | FJ537259, FJ537208, FJ537161 |
| Car. turkestanica Kom. | M.L. Zhang 00-101(PE) | Cultivated, Bergius Botanical Garden, Stockholm, Sweden | FJ537256, FJ537206, FJ537158 |
| Ser. Microphyllae (Kom.) Pojark. | |||
| Car. bungei Ledeb. | M.L. Zhang et al. 99–225 (PE) | Bajanchongor, Mongolia | FJ537267, FJ537216, FJ537169 |
| Car. microphylla Lam. | M.L. Zhang et al. 99–214 (PE) | Lhongcheng, Mongolia | FJ537264, FJ537213, FJ537166 |
| Sect. Bracteolatae (Kom.) M. L. Zhang | |||
| Ser. Bracteolatae Kom. | |||
| Car. bicolor Kom. | M.L. Zhang & Y. Kang Y 99–178 (PE) | Markang, Sichuan, China | FJ537246, FJ537197, FJ537147 |
| Car. brevispina Benth. | M.L. Zhang 281-05-8414/101 (PE) | Cultivated, Berlin Botanical Garden, Germany (originally from Kashmir) | FJ537248, FJ537200, FJ537150 |
| Car. sukiensis C. K. Schneid. | S.G. Miehe & K. Kock s.n. (NHM) | Donkardzong, Nepal | FJ537247, FJ537199, FJ537149 |
| Sect. Jubatae (Kom.) Y. Z. Zhao | |||
| Ser. Jubatae Kom. | |||
| Car. jubata (Pall.) Poir. | M.L. Zhang 00279 (PE) | Zhaosu (Tianshan), Xinjiang, China | FJ537242, FJ537194, FJ537143 |
| Car. pleiophylla (Regel) Pojark. | M.L. Zhang 10–146 (PE) | Tekes, Xinjiang, China | FJ537253, FJ537203, FJ537155 |
| Ser. Leucospinae Y. Z. Zhao | |||
| Car. tibetica (Maxim. ex C. K. Schneid.) Kom. | M.L. Zhang 00–89 (PE) | Uhai, Nei Mongol, China | FJ537244, FJ537195, FJ537145 |
| Sect. Frutescentes (Kom.) Sanchir | |||
| Ser. Frutescentes Kom. | |||
| Car. kirghisorum Pojark. | C.Y. Chang et al. 2004219 (WUG) | Khorgos, Xinjiang, China | FJ537820, FJ537229, FJ537181 |
| Car. opulens Kom. | M.L. Zhang & Y. Kang 99–123 (PE) | Daofu, Sichuan, China | FJ537282, FJ537231, FJ537183 |
| Ser. Chamlagu Pojark. | |||
| Car. rosea Turcz. ex Maxim. | M.L. Zhang 99–45 (PE) | Beihuashan, Beijing, China | FJ537272, FJ537221, FJ537174 |
| Car. sinica (Buc’hoz) Rehder | M.L. Zhang 99–49 (PE) | Xiangshan, Beijing, China | FJ537284, FJ537233, FJ537185 |
| Ser. Pygmaeae Kom. | |||
| Car. brevifolia Kom. | Q.L. Ho et al. 2498 (NHM) | Yushu, Qinghai, China | FJ537268, FJ537217, FJ537170 |
| Car. stenophylla Pojark. | M.L. Zhang 00–78 (PE) | Hangjinqi, Nei Mongol, China | FJ537274, FJ537223, FJ537176 |
| Car. versicolor Benth. | S. Miehe 99-62-06 (NHM) | Upper Dolpo, Nepal | FJ537271, FJ537220, FJ537173 |
| Sect. Spinosae (Kom.) Y. Z. Zhao | |||
| Ser. Spinosae Kom. | |||
| Car. hololeuca Bunge ex Kom. | M.L. Zhang 00–153 (PE) | Cultivated, Turfan Botanical Garden, Xinjiang, China | FJ537240, FJ537192, FJ537141 |
| Car. spinosa (L.) Hornem. | C.Y. Chang et al. 2004503 (WUG) | Qinghe, Xinjiang, China | FJ537241, FJ537193, FJ537142 |
| Ser. Acanthophyllae Pojark. | |||
| Car. acanthophylla Kom. | M.L. Zhang 00–154 (PE) | Cultivated, Turfan Botanical Garden, Xinjiang, China | FJ537238, FJ537191, FJ537139 |
| Halimodendron halodendron (Pall.) Voss | M.L. Zhang 00–279 (PE) | Cultivated, Urumqi Botanical Garden, Xinjiang, China | FJ537289, FJ537237, FJ537190 |
| Hedysarum alpinum L. | M. Riewe 182 (CAS) | Northwest Territories, Canada | FJ537287, FJ537235, FJ537188 |
| Astragalus coluteocarpus Boiss. | Qinghai-Xizang Expedition Team 76–8083 (PE) | Zada, Ali, Xizang, China | FJ537286,————, FJ537187 |
Classification system of Caragana follows Zhang (1997) (See also Zhang et al. 2009, Table 1). Classification system of Calophaca follows Gorbanova (1987) concerning two sections, and a new section, Sect. Trichomeae M.L. Zhang is yielded from this paper.
A previous phylogenetic study of the nuclear ITS region and two chloroplast regions (trnS-trnG and rbcL) indicated Calophaca to be nested within Caragana [27], but only one species was sampled from Calophaca. Here, sampling all of the species of Calophaca, we conduct a phylogenetic analysis to assess its systematic position, monophyly, and inter-species relationships, and its putative geographical origin and dispersion across Eurasia.
Materials and Methods
Taxon sampling
Fourteen individuals representing all eight species currently recognized in Calophaca were sampled for this study [14,17,25], shown in Table 1 and deposited in the herbaria. According to the previous phylogenetic studies [27–30], Calophaca and Halimodendron nest within Caragana. Thus the genera Halimodendron, Caragana, Hedysarum, and Astragalus were included as outgroups in this study (Table 1), with Astragalus serving to root the trees. To assess the monophyly of Calophaca, 21 species of the genus Caragana were also sampled, giving a total of 38 samples that were included in the study. All species materials vouchers in Table 1 are deposited in the public herbaria PE (Institute of Botany, Chinese Academy of Sciences, Beijing, China), XIJI (Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi, China), WUG (Northwest Institute of Botany, Yangling, Shaanxi, China), LE (Komarov Botanical Institute, Russian Academy of Sciences, St. Petersburg, Russia), MW (Moscow University, Moscow, Russia) and MAH (Main Botanical Garden, Russian Academy of Sciences, Moscow, Russia).
We state that all use of species materials were permitted by the authority of these herbaria. Our study did not concern Human Subject Research or Animal Research. We can declare that the leaf materials did not come from conservation parks, and none of the samples involved endangered or protected species.
DNA sequencing
Total genomic DNA was extracted using the CTAB method [31]. For primers of the ITS region, trnS-trnG spacer and rbcL, see Zhang et al. [27]. The polymerase chain reaction (PCR) was used for amplification of double stranded DNA. The 25 μl reaction system contained 0.25 μl of Ex Taq, 2.5 μl of 10× Ex Taq buffer (Mg2+ concentration of 25 mM), 2.0 μl of dNTP mix (2.5 mM concentration for each dNTP), 1 μl of the forward and reverse primers at 5 umol/μl, and 0.5 μl of template DNA. The protocol for amplification consisted of an initial hot start at 95°C for 2 min, followed by 30 cycles of denaturing at 94°C for 30 s, annealing at 52°C for 30 s, extension at 72°C for 90 s, and a final extension at 72°C for 10 min. PCR products were purified using the PEG precipitation procedure [32] and sequenced using an ABI Prism 3770 Genetic Analyzer (Shanghai Shenggong Biological Engineering Technology & Service, Shanghai, China).
Sequences were aligned with CLUSTAL X [33] and then adjusted manually. All gaps were treated as missing data cells. Finally, the combined 3-genic region data set comprised 3417 aligned nucleotide characters.
Phylogenetic analysis and divergence time estimate
The incongruence length difference (ILD) test [34] was carried out in PAUP* [35] to assess potential conflicts between data set partitions. This was implemented with 100 partition-homogeneity test replicates, a heuristic search option with simple addition of taxa, TBR branch swapping, and MaxTrees set to 1000. Phylogenetic analyses were performed using maximum likelihood (ML) and Bayesian inference. ML analysis was implemented with PAUP; clade support was estimated with 1,000 heuristic bootstrap replicates (100 random addition cycles per replicate, with tree bisection-reconnection and branch-swapping) [36,37]. For ML analysis, Modeltest 3.06 [38] was used to estimate the appropriate model of DNA substitution for sequence data. The models selected by the Akaike information criterion (AIC) were TIM+I+G. The related parameters of Modeltest were used for the ML analysis.
Bayesian phylogenetic analysis and divergence time estimates were implemented in BEAST 1.5.4 [39,40]. We used the uncorrelated lognormal relaxed clock model with a Yule process for the speciation model, and GTR+I+G for the substitution model (estimated for the data set). The age of tree prior (normal distribution Mean = 28, Stdev = 1) for Hedysareae and Astragaleae, i.e., the ancestral node of Astragalus and Caragana was defined as the root at 28 Ma following Lavin et al. [41], Wojciechowski [30] and Zhang and Fritsch [42]. A Markov chain Monte Carlo was run for 50 million generations and sampled every 1,000 generations. Two independent runs were performed to confirm convergence of the analysis. The stationarity of each run was examined using the effective sampling size of each parameter (>200). The last 40 million generations were used to construct the maximum clade credibility tree and the associated 95% highest posterior density distributions around the estimated node ages using program TreeAnnotator 1.5.4, and visualized using FigTree 1.3.1.
Biogeographical analysis
We designated five biogeographical areas based on the distributions of the Calophaca species, especially its disjunctions, see Fig 1. Most of the species occurring in these areas are endemic, for example, C. grandiflora and C. sericea are endemic to the Pamir mountains, C. wolgarica to the Caucasus, and C. sinica to East Asia. The areas chosen are natural regions, and often independent biodiversity hotspots [19], for example, the Pamir and Tianshan mountains and the Caucasus. These areas also have distinct vegetation and floras; especially different are those of the Pamir, western Tianshan, and Tarbagatai mountains, see Fig 1. The five designated areas are thus: A: the Pamirs, Tadjikistan; B: the western Tianshan Mountains, including portions of Kyrgistan and Kazakhstan; C: the Tarbagatai, including portions of northwestern China and Kazakhstan; D: East Asia, and montane portions of Shanxi Province, China; E: the Caucasus, and plains along the Volga and Don Rivers.
To infer vicariance and dispersal events, a Bayesian parsimony-based method (S-DIVA) and BBM (Bayesian Binary Method) were employed [43,44]. DIVA is an event-based method that optimizes ancestral distributions by assuming a vicariance explanation, while incorporating the potential contributions of dispersal and extinction [45]. Nylander et al. [46] proposed a modified approach to DIVA named Bayes-DIVA that integrates biogeographical reconstructions of DIVA over the posterior distribution of a Bayesian MCMC sample of tree topologies. Bayes-DIVA is also referred to as S-DIVA [43]. BBM infers ancestral areas using a full hierarchical Bayesian approach; it hypothesizes a special “null distribution” which means that an ancestral range contains none of the unit areas [44]. S-DIVA and BBM can be performed in RASP (Reconstruct Ancestral State in Phylogenies) 2.0 beta. http://mnh.scu.edu.cn/soft/blog/RASP.
The BEAST molecular dating tree (Fig 2) was treated as a fully resolved phylogram for use as the basis for S-DIVA and BBM, with 1000 post-burnin trees derived from the BEAST analysis used for ancestral area reconstruction in the program RASP. Multiple samples of the same taxon in a clade are combined into one branch, or many identical terminal areas in a clade are regarded as one; consequently, the tree is reduced to maximum simplicity. RASP was performed with various constraints of maximum areas 2 at each node, to infer possible ancestral areas and potential vicariance and dispersal events [47–49].
Fig 2. Chronogram of Calophaca and outgroups Caragana, Halimodendron, Hedysarum, and Astragalus, with maximum clade credibility performed by BEAST.

Dates of divergence are shown to the right of nodes, and posterior probability values are shown to the left of nodes.
The outgroups used in the phylogeny were also used in the biogeographical analyses throughout data processing, but are not shown in the resulting Fig 3.
Fig 3. Ancestral area optimizations performed with S-DIVA and BBM.

Pie chartsat nodes are conducted from S-DIVA (on the branch above) and BBM (on the branch below). The three thicked arrowheaded lines at branches show four dispersals.
Area labels, as stated in the text: A: Pamir Mountains, Tadjikistan; B: western Tianshan Mountains, plus parts of Kyrgistan and Kazakhstan; C: Tarbagatai Mountains, including adjacent regions of China and Kazakhstan; D: East Asia, mountains of Shanxi Province, China; E: Caucasus, and plains along the Volga River and Don River.
Results
Phylogenetic analysis and divergence time estimate
The data partitions of the three genes were not significantly incongruent on the basis of the ILD tests (P = 0.133). ML analysis yielded three optimal trees. Topologies of the three were on the whole equivalent to the Bayesian BEAST tree. Since the BEAST tree also has a dating result, it is used to illustrate phylogenetic relationships and the dating results (see Fig 2).
Our phylogenetic analyses show that Calophaca is monophyletic, with high bootstrap (bt) support and posterior probability (pp) (bt = 96%, pp = 1.00) (node I), and is nested within Caragana (Fig 2). The phylogenetic analysis yielded a topology largely in accordance with the current infrageneric classification of Calophaca. The two species of C. sect. Glabriflorae formed a clade (bt = 100%, pp = 1.00), which is sister to the other taxa, and the two samples of C. sinica also formed a clade (bt = 98%, pp = 1.00). Samples of C. soongorica, C. hovenii, C. pskemica, C. wolgarica, and C. tianshanica comprised a clade (bt = 97%, pp = 1.00) (node III), equivalent to section Calophaca. These three clades within Calophaca had high bootstrap and posterior probability values (Fig 2).
The estimated divergence (stem) age of Calophaca was ca. 8.0 (95% HPD: 3.44–11.09) Ma, the crown age between the Pamirian sect. Glabriflorae and the other two sections was ca. 7.0 (95% HPD: 1.66–7.29, crown age) Ma (node I), and the age between the East Asian sect. Trichomeae and the Tianshan Mountains sect. Calophaca was ca. 6.0 Ma (Fig 2, node II, crown age). The diversification time range within sect. Calophaca was from Pliocene to Pleistocene, i.e., 2.6–0.2 Ma (Fig 2, node III). The diversification time of the two species of sect. Glabriflorae was ca. 0.4 Ma. Calophaca wolgarica, distributed from the Caucasus to Russian Europe, was formed more recently at ca. 0.2 Ma.
Ancestral area reconstructions
Within Calophaca sect. Calophaca, the ancestral area (node III, Fig 3) estimated from S-DIVA (B and BC at equal probability) and BBM (B), thus we chose it as B (western Tianshan Mountains). The remaining area C (Tarbagatai Mountains) within this section (Fig 3), is shown as a dispersal many times. The most incongruent and uncertain node of the ancestral distribution estimates is at the root of Calophaca (node I). Considering S-DIVA, showing the likely ancestral area as AD (union of Pamir Mountains and East Asia) and AB (union of the Pamir and western Tianshan mountains), A is the intersection, occurring in both ancestral areas. Concerning the value of AD at node I, and the reason for the selection of A (ignoring D), was that we mainly thought A, with C. sericea and C. grandiflora at the phylogenetic tree base (stem age ca. 6.98 Ma, see Fig 2), to be more “old” than D of C. sinica (stem age ca. 5.98 Ma). This indicates that A (Pamir) should be the ancestral area instead of D (East Asia). In addition, ignoring AD as an ancestral area would mean ignoring an impossibly large ancestral area union, covering two separate portions, the Pamir and East Asia (North China, Shanxi Province), and consequently the first speciation of Calophaca would have had to be speculated as a vicariance between Pamir and East Asia, in contrast with the unique place of origin of the Pamirs. Therefore, we felt we should choose the single area A as ancestral. In addition, BBM, with a most likely estimated area of A, confirmed and justified the S-DIVA result. Therefore, it appeares preferable to choose A as the ancestral area of Calophaca.
After deciding the ancestral areas at the nodes (Fig 3), several dispersals can be recognized, see Figs 3 and 1. First, two dispersals are inferred from area A (Pamir) (node I) to B (western Tianshan) and D (East Asia). A dispersal is shown from node III to node V, and finally to the Caucasus. Another dispersal is from B (node IV) to C (Tarbagatai).
Discussion
Classification of Calophaca
The monophyly of Calophaca was verified with high bootstrap (bt) support and posterior probability (pp) (bt = 96%, pp = 1.00, node I, Fig 2). The present phylogenetic tree (Fig 2) indicates three clades with high support within Calophaca, one consisting of C. grandiflora and C. sericea, a second containing C. sinica, and a third containing C. soongorica, C. tianschanica, C. hovenii, C. wolgorica, and C. pskemica. This is well consistent with previous morphological classification except for C. sinica. According to Gorbunova’s [15] classification system for the genus, C. grandiflora and C. sericea are included in sect. Glabriflora, while C. soongorica and C. wolgorica, etc. belong to sect. Calophaca. Calophaca sinica has a combination of distinctive characters, with dense glandular trichomes on the standard, peduncle, calyx, ovary, style, and a legume different from other species within the genus, and has an endemic distribution in East Asia. Therefore, we can use it to establish a new section Trichomeae, to contrast with the two sections of Gorbanova [15], see S1 File. This effectively completes the classification for the genus, with a total of three sections.
The five accessions of Calophaca soongorica formed a clade with the inclusion of C. hovenii. Calophaca hovenii Schrenk, C. tianshanica (B. Fedtsch.) Borissova, and C. soongorica (C. soongorica 1 and 2 = C. chinensis Borissova, see Table 1) were included within C. soongorica as varieties [14,15,18]. Clearly, C. hovenii and the others should be placed within C. soongorica.
In the traditional taxonomy, Calophaca, Halimodendron, and Caragana are regarded as related and distinctive genera [20,21]. Previously the three were included in subtribe Astragaleae of tribe Galegeae [20], and later changed to tribe Hedysareae [21]. Diagnostic characters of Calophaca and Caragana are that while both are shrubs with pinnate leaflets, Calophaca is imparipinnate but Caragana is paripinnate. Contrasting with the flat pods of Calophaca and Caragana, Halimodendron has an inflated pod. Calophaca, particularly, has a distribution of montane grassland, shrubland, and dry forest.
Since Calophaca and Halimodendron are nested within Caragana (Fig 2), it results in Caragana being paraphyletic. In terms of the taxonomic identities of Calophaca, Halimodendron, and Caragana, Caragana should be subdivided into segregate taxa. Currently it has been recognized as six phylogenetic clades (Fig 2), which include the three sections labeled, as well as clades representative by Car. jubata, Car. acanthophylla, and Car. spinosa. However, in view of the limitation of sampled taxa in our study, further research should be conducted examining sufficient taxa and more genes. Essentially, the present paper focuses on Calophaca.
Origin and diversification
Our results yielded a divergence time (stem age, or origin age) of ca. 8.0 Ma for Calophaca (Fig 2), and a place of origin in the Pamir Mountains (Fig 3), which implies that a global cooling and drying process at 8–7 Ma in the late Miocene [10,13], most likely drove generic origin. It is also possible more local events played a part in the origin, especially major tectonism in the Tianshan range at about 6.5 Ma [50,51]; the Pamir mass had experienced severe deformation and uplift from the India-Asia impact at a previous time [52]. Progressive aridification in connection with global cooling appears to have played an important role in the later Miocene [13]. The Pamir-western Tianshan mountains experienced progressively more arid climates. This climate transition possibly led to range restriction and isolation of organisms, and Calophaca may be one of the many lineages that then evolved and diversified in the Pamirs.
Compared with the diversification time of Caragana [42], the crown age (diversification age) of Calophaca at ca. 6.98 Ma coincided with the diversification times of the three sections within Caragana, whereas diversification times of the three Calophaca sections are younger at 2.6, 0.4 and 0.24 Ma (Fig 2). Most Central Asian Calophaca species, such as C. grandiflora, C. sericea, C. soongorica, and C. wolgarica, have young crown ages i.e., Pliocene to Pleistocene 0.84–0.2 Ma (Fig 2). These show that the genus is a relatively young taxon among lineages of temperate legume shrubs and the Central Asian flora.
In general, the Central Asian flora is speculated to have formed from Tertiary Tethys relicts [12]. Hippophae rhamnoides (Elaeagnaceae) distributed in Eurasia, was similarly shown to have originated from the QTP and migrated to Central Asia and Asia Minor ⁄ Europe [6]. Artemisia has been shown to have originated from the arid-semiarid middle latitudes of Asia in the late Eocene, and then spread westward and eastward in the Oligocene as the result of early QTP uplift and accompanying aridification [9]. Calophaca presents another pattern of Asian origin and dispersal that is somewhat different from that of Hippophae rhamnoides and Artemisia.
Parts of the QTP may have experienced rapid uplift contributing to global cooling during past 8–7 Ma [13,53–60]. Responding to this QTP uplift event and global cooling and drying, many biological cases mainly occurring in situ in the QTP have been contributed, for example, the Ligularia Cremanthodium—Parasenecio complex (Asteraceae) [61], Rheum (Polygonaceae) [62], Androsace (Primulaceae) [3] and Saussurea (Asteraceae) [63], and the glyptosternoid catfishes [64]. Even though absent in the QTP, Calophaca has a diversification crown age of ca. 6.98 Ma and a distribution located to the north of the QTP, and it can be supposed that the origin and evolution of Calophaca were driven by the same paleogeographic and paleoclimatic conditions.
Dispersal and disjunction
According to the biogeographical analysis (Fig 3), we can hypothesize that dispersal may be a dominant speciation mode in Calophaca. There are four remarkable dispersals from the late Miocene to Pleistocene in this genus, i.e., from the Pamir to the western Tianshan mountains, and to East Asia at ca. 7.0 Ma, northward to the Tarbagatai Mountains at ca. 2.2 Ma, and from the western Tianshan Mountains westward to the Caucasus—Russian Europe at 2.6 Ma (Fig 1). The great arid and semiarid belt, comprising the Tarim Basin, Junggar Basin, Hexi Corridor, and Loess Plateau in arid northwestern China forms a barrier between the Pamir-western Tianshan mountains and East Asia. Similarly, the Turan lowland and desert separates the Pamir—western Tianshan mountains and the Caucasus. The presence of these arid belts most likely resulted from the intensified Asian Interior aridification which is associated with global cooling and drying since the Middle Miocene optimum 8–7 Ma [13], and consequently to have fragmented the ranges of the Calophaca species and also eliminated the traces of their dispersals.
Montane distribution and refugia
Extant Calophaca species of the Pamir and western Tianshan mountains, such as C. grandiflora, C. sericea, C. tianshanica, C. pskemica, and C. soongorica, often occur in middle or low montane belts at altitudes of 800–2800 m, sometimes in meadows or steppe, with most individuals occurring in juniper forests or shrublands [14,17]. Calophaca sinica, in the northern China mountains, likewise grows in forest or shrubland [65]. Calophaca wolgarica, a shrub endemic to the Caucasus and eastern Europe, occurs on steppe, plains, and limestone hills [14,17]. In conclusion, the predominant habitat of the genus is the lower montane, in the Pamir—western Tianshan mountains, the Caucasus, and the northern China mountains.
These montane distribution centers of Calophaca, occupy several global biodiversity hotspots [19], especially the Pamir—western Tianshan mountains and the Caucasus. These areas are also regarded as plant refugia of the Quaternary, since the mountains have maintained a somewhat moist habitat, and thus have an advantage in protection of species from destruction caused by enhanced cooling and drying in the Quaternary [66–73]. The high level of genetic and taxonomic diversity of this area is indicative of its ecological stability [67,69]. Therefore, Calophaca is in fact a representative of the global biodiversity hotspots and refugia.
Supporting Information
(PDF)
Acknowledgments
We are grateful to the herbaria staffs of Komarov Botanical Institute, Russian Academy of Sciences (St. Petersburg), Moscow University (Moscow), Main Botanical Garden, Russian Academy of Sciences (Moscow), Institute of Botany (Beijing), Chinese Academy of Sciences, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences (Urumqi, China), Northwest Institute of Botany (Yangling, Shaanxi, China) for our convenient checking of specimens. We are deeply appreciative of Dr. Wei Wang, an anonymous reviewer, and Dr. Qi Wang for their helpful and constructive comments and suggestions.
Data Availability
All data are available in the paper.
Funding Statement
This study was funded by China National Basic Research Special Program (2012FY111500), Innovation Research Group Program of Chinese Academy of Sciences and State Administration of Foreign Experts Affairs of China (KZCX2-YW-T09), State International Science and Technology Cooperation Program of China (2010DFA92720), and Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Takhtajan A. Flowering Plants, Origin and Dispersal. Edinburgh: Oliver and Boyd; 1969. [Google Scholar]
- 2. Agakhanjanz O, Breckle SW. Origin and evolution of the mountain flora in Middle Asia and neighbouring mountain regions. Ecol Studies. 1995; 113: 63–80. [Google Scholar]
- 3. Wang YJ, Li XJ, Hao G, Liu JQ. Molecular phylogeny and biogeography of Androsace (Primulaceae) and the convergent evolution of cushion morphology. Acta Phytotaxon Sin. 2004; 42: 481–499. [Google Scholar]
- 4. Kadereit JW, Licht W, Uhink CH. Asian relationships of the flora of the European Alps. Plant Ecol Diver. 2008; 1: 171–U160. [Google Scholar]
- 5. Malyshev LI. Phenetics of the subgenera and sections in the genus Oxytropis DC. (Fabaceae) bearing on ecology and phylogeny. Contemp Prob Ecol. 2008; 1: 440–444. [Google Scholar]
- 6. Jia D-R, Abbott RJ, Liu T-L, Mao K-S, Bartish IV, Liu J-Q. Out of the Qinghai-Tibet Plateau: evidence for the origin and dispersal of Eurasian temperate plants from a phylogeographic study of Hippophae rhamnoides (Elaeagnaceae). New Phytologist. 2012; 194: 1123–1133. 10.1111/j.1469-8137.2012.04115.x [DOI] [PubMed] [Google Scholar]
- 7. Grubov VI. Plants of Central Asia Vol. 1. New Hampshire: Science Publishers; 1999. [Google Scholar]
- 8. Pyankov VI, Gunin PD, Tsoog S, Black CC. C4 plants in the vegetation of Mongolia: their natural occurrence and geographical distribution in relation to climate. Oecologia. 2000; 123: 15–31. [DOI] [PubMed] [Google Scholar]
- 9. Miao Y, Meng Q, Fang X, Yan X, Wu F, Song C. Origin and development of Artemisia (Asteraceae) in Asia and its implications for the uplift history of the Tibetan Plateau: A review. Quatern Internat. 2011; 236: 3–12. [Google Scholar]
- 10. Guo ZT, Sun B, Zhang ZS, Peng SZ, Xiao GQ, Ge JY, et al. A major reorganization of Asian climate by the early Miocene. Climate Past. 2008; 4: 153–174. [Google Scholar]
- 11. Sun J, Ye J, Wu W, Ni X, Bi S, Zhang Z, et al. Late Oligocene-Miocene mid-latitude aridification and wind patterns in the Asian interior. Geology. 2010; 38: 515–518. [Google Scholar]
- 12. Wulff EV. Historical plant geography: history of the world flora. Waltham, MA: Chronica Botanica; 1944. [Google Scholar]
- 13. Miao Y-F, Herrmann M, Wu F-L, Yan X-L, Yang S-L. What controlled Mid-Late Miocene long-term aridification in Central Asia?—Global cooling or Tibetan Plateau uplift: A review. Earth Sci Revi. 2012; 112: 155–172. [Google Scholar]
- 14. Borissova AG. Flora of the USSR, vol. 11. In Komarov VL, Schishkin BK eds. Moscow, URSS: Academiae Scientiarum, 327–368; 1945. [Google Scholar]
- 15. Gorbunova NN. Systematics: Caragana and Calophaca in Kirgistan . Frunze: Academy of Kirgistan, Institute of Biology; 1987. [Google Scholar]
- 16. Zhang Z-W. Calophaca In: Fu K-Ted. Flora Reipublicae Popularis Sinicae 42(1). Beijing: Science Press; 67–71; 1993. [Google Scholar]
- 17. Yakovlev GP, Sytin AK, Roskov JR. Legumes of Northern Eurasia, a check-list. Kew: Royal Botanic Gardens; 1996. [Google Scholar]
- 18. Zhu X-Y, Larsen K. Calophaca Pages 527–528 in: Wu Z-Y, Raven PH eds. Flora of China, vol 10 St. Louis & Beijing: Science Press, and Missouri Botanical Garden Press; 2010. [Google Scholar]
- 19. Zachos FE, Habel JC. Biodiversity hotspots. Distribution and protection of conservation priority areas. Berlin: Springer-Verlag; 2011. [Google Scholar]
- 20. Polhill RM. Galegeae Pages 357–363 in: Polhill RM, Raven PH eds. Advances in Legume Systematics, part 1 Kew: Royal Botanical Gardens; 1981. [Google Scholar]
- 21. Lock JM. Tribe Hedysareae Pages 489–495 in: Lewis G, Schrire B, MacKinder B, Lock M eds Legumes of the world Kew: Royal Botanic Gardens; 2005. [Google Scholar]
- 22. Kreuter E. Chromosome atlas of flowering plants. In: Darlingto CD, Wylie AP eds. London: George Allen & Unwin Ltd, 163; 1955. [Google Scholar]
- 23. Magulaev YA. Chromosome numbers of some legumes of northern Caucasus. Bot Zhur. 1980; 65: 836–843. [Google Scholar]
- 24. Li S-F, Chang Z-Y, Li B, Xu L-R. The cytogeography study of one pairing species of Calophaca Fisch. in west and east sides of Mongolia-Xinjiang Desert. Acta Bot Bor-Occid Sin. 1998; 18: 58–60. [Google Scholar]
- 25. Chang Z-Y, Xu L-R, Wu Z-H. Comparative morphology, ecology and geographical distribution of Calophaca sinica and C. soongorica . Acta Bot Bor-Occid Sin. 2004; 24: 2312–2320. [Google Scholar]
- 26. Xu L-R, Fu K-T, He S-B, Chang C-W. Study on the flora of Leguminosae in Loess Plateau. Acta Bot Bor-Occid Sin. 1992; 12: 149–153. [Google Scholar]
- 27. Zhang M, Fritsch PW, Cruz BC. Phylogeny of Caragana (Fabaceae) based on DNA sequence data from rbcL, trnS-trnG, and ITS. Mol Phylogen Evol. 2009; 50: 547–559. 10.1016/j.ympev.2008.12.001 [DOI] [PubMed] [Google Scholar]
- 28. Wojciechowski MF, Sanderson MJ, Steele KP, Liston A. Molecular phylogeny of the "Temperate Herbaceous Tribes" of Papilionoid legumes: a supertree approach In: Herendeen PS, Bruneau A (eds), Advances in Legume Systematics 9. Kew, UK: Royal Botanic Gardens, 277–298; 2000. [Google Scholar]
- 29. Wojciechowski MF, Lavin M, Sanderson MJ. A phylogeny of legumes (Leguminosae) based on analyses of the plastid matK gene resolves many well-supported subclades within the family. Amer J Bot 91: 1846–1862; 2004. [DOI] [PubMed] [Google Scholar]
- 30. Wojciechowski MF. Astragalus (Fabaceae): A molecular phylogenetic perspective. Brittonia. 2005; 57: 382–396. [Google Scholar]
- 31. Doyle JJ, Doyle JL. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 1987; 19: 11–15. [Google Scholar]
- 32. Johnson LA, Soltis DE. Phylogenetic inference in Saxifragaceae sensu stricto and Gilia (Polemoniaceae) using matK sequences. Ann Missouri Bot Gard. 1995; 82: 149–175. [Google Scholar]
- 33. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl Acid Res. 1997; 25: 4876–4882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Farris JS, Källersjö M, Kluge AG, Bult C. Testing significance of incongruence. Cladistics. 1994; 10: 315–319. [Google Scholar]
- 35. Swofford DL. PAUP*. phylogenetic analysis using parsimony (*and other methods). Version 4. Sunderland: Sinauer Associates; 2003. [Google Scholar]
- 36. Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985; 39: 783–791. [DOI] [PubMed] [Google Scholar]
- 37. Hillis DM, Bull JJ. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst Biol. 1993; 42: 182–192. [Google Scholar]
- 38. Posada D, Crandall KA. Modeltest: testing the model of DNA substitution. Bioinformatics. 1998; 14: 817–818. [DOI] [PubMed] [Google Scholar]
- 39. Drummond AJ, Ho SY, Phillips MJ, Rambaut A. Relaxed phylogenetics and dating with confidence. PLoS Biol. 2006; 4: e88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Drummond AJ, Rambaut A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol Biol. 2007; 7: 214 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Lavin M, Herendeen PS, Wojciechowski MF. Evolutionary rates analysis of Leguminosae implicates a rapid diversification of lineages during the tertiary. Syst Biol. 2005; 54: 575–594. [DOI] [PubMed] [Google Scholar]
- 42. Zhang M-L, Fritsch PW. Evolutionary response of Caragana (Fabaceae) to Qinghai-Tibetan Plateau uplift and Asian interior aridification. Plant Syst Evol. 2010; 288: 191–199. [Google Scholar]
- 43.Yu Y, Harris AJ, He XJ. RASP (Reconstruct Ancestral State in Phylogenies). Version 2.0; 2011. [DOI] [PubMed]
- 44. Ali SS, Yu Y, Pfosser M, et al. Inferences of biogeographical histories within subfamily Hyacinthoideae using S-DIVA and Bayesian binary MCMC analysis implemented in RASP (Reconstruct Ancestral State in Phylogenies). Ann Bot. 2012; 109(1): 95–107. 10.1093/aob/mcr274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Ronquist F. Dispersal-vicariance analysis: A new approach to the quantification of historical biogeography. Syst Biol. 1997; 46: 195–203. [Google Scholar]
- 46. Nylander JAA, Olsson U, Alstrom P, Sanmartin I. Accounting for phylogenetic uncertainty in biogeography: A Bayesian approach to dispersal-vicariance analysis of the thrushes (Aves: Turdus). Syst Biol. 2008; 57: 257–268. 10.1080/10635150802044003 [DOI] [PubMed] [Google Scholar]
- 47. Clayton JW, Soltis PS, Soltis DE. Recent long-distance dispersal overshadows ancient biogeographical patterns in a pantropical angiosperm family (Simaroubaceae, Sapindales). Syst Biol. 2009; 58: 395–410. 10.1093/sysbio/syp041 [DOI] [PubMed] [Google Scholar]
- 48. Almeida EAB, Pie MR, Brady SG, Danforth BN. Biogeography and diversification of colletid bees (Hymenoptera: Colletidae): emerging patterns from the southern end of the world. J Biogeogr. 2012; 39: 526–544. 10.1016/j.ijantimicag.2012.02.014 [DOI] [PubMed] [Google Scholar]
- 49. Mao K, Milne RI, Zhang L, Peng Y, Liu J, Thomas P. et al. Distribution of living Cupressaceae reflects the breakup of Pangea. PNAS. 2012; 109: 7793–7798. 10.1073/pnas.1114319109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Sun J, Zhang Z. Syntectonic growth strata and implications for late Cenozoic tectonic uplift in the northern Tian Shan, China. Tectonophysics. 2009; 463: 60–68. [Google Scholar]
- 51. Sun J, Li Y, Zhang Z, Fu B. Magnetostratigraphic data on Neogene growth folding in the foreland basin of the southern Tianshan Mountains. Geology. 2009; 37: 1051–1054. [Google Scholar]
- 52. Ducea MN, Lutkov V, Minaev VT, Hacker B, Ratschbacher L, Luffi P, et al. Building the Pamirs: The view from the underside. Geology. 2003; 31: 849–852. [Google Scholar]
- 53. Harrison TM, Copeland P, Kidd WSF, Yin A. Raising Tibet. Science. 1992; 255: 1663–1670. [DOI] [PubMed] [Google Scholar]
- 54. Cerling TE, Wang Y, Quade J. Expansion of C4 ecosystems as an indicator of global ecological change in the late Miocene. Nature. 1993; 361: 344–345. [Google Scholar]
- 55. Molnar P, England P, Martiod J. Mantle dynamics, uplift of the Tibetan Plateau and the Indian monsoon development. Review Geophysics. 1993; 34: 357–396. [Google Scholar]
- 56. Quade J, Cerling TE. Expansion of C4 grasses in the Late Miocene of Northern Pakistan: evidence from stable isotopes in paleosols. Palaeogeol Palaeoclim Palaeoecol. 1995; 115: 91–116. [Google Scholar]
- 57. Cerling TE, Harris JM, MacFadden BJ, Leakey MG, Quade J, Eisenmann V, et al. Global vegetation change through the Miocene/Pliocene boundary. Nature 389: 153–158. [Google Scholar]
- 58. Zheng H-B, Powell CM, An Z-S, Zhou J, Dong G-R (2000) Pliocene uplift of the northern Tibetan Plateau. J Geol. 1997; 28: 715–718. [Google Scholar]
- 59. An ZS, Kutzbach JE, Prell WL, Porter SC. Evolution of Asian monsoons and phased uplift of the Himalayan Tibetan plateau since Late Miocene times. Nature. 2001; 411: 62–66. [DOI] [PubMed] [Google Scholar]
- 60. Molnar P. Mio-Pliocene growth of the Tibetan Plateau and evolution of East Asian climate. Palaeontol Electron 8; 2005. [Google Scholar]
- 61. Liu JQ, Wang YJ, Wang AL, Hideaki O, Abbott RJ. Radiation and diversification within the Ligularia-Cremanthodium-Parasenecio complex (Asteraceae) triggered by uplift of the Qinghai-Tibetan Plateau. Mol Phylogen Evol. 2006; 38: 31–49. [DOI] [PubMed] [Google Scholar]
- 62. Wang AL, Yang MH, Liu JQ. Molecular phylogeny, recent radiation and evolution of gross morphology of the rhubarb genus Rheum (Polygonaceae) inferred from chloroplast DNA trnL-F sequences. Ann Bot. 2005; 96: 489–498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Wang Y-J, Susanna A, Von Raab-Straube E, Milne R, Liu J-Q. Island-like radiation of Saussurea (Asteraceae: Cardueae) triggered by uplifts of the Qinghai-Tibetan Plateau. Biol J Linn Soc. 2009; 97: 893–903. [Google Scholar]
- 64. Peng ZG, Ho SYW, Zhang YG, He SP. Uplift of the Tibetan plateau: Evidence from divergence times of glyptosternoid catfishes. Mol Phylogen Evol. 2006; 39: 568–572. [DOI] [PubMed] [Google Scholar]
- 65. Ma Q, Duan Y, Shangguan T, Li H, Guo H, Zhang Q. Relationship Among Species Diversities and Soil Properties in Endangered Calophaca sinica habitats in Tianlong Mountain, Shanxi, China. Chin J Appl Environ Biol. 2012; 18: 391–398. [Google Scholar]
- 66. Comes HP, Kadereit JW. The effect of Quaternary climatic changes on plant distribution and evolution. Trends Pl Sci. 1998; 3: 432–438. [Google Scholar]
- 67. Medail F, Quezel P. Biodiversity hotspots in the Mediterranean basin: Setting global conservation priorities. Conser Biol. 1999; 13: 1510–1513. [Google Scholar]
- 68. Hewitt GM. Genetic consequences of climatic oscillations in the Quaternary. Philosophical Transactions of the Royal Society: Biol Sci. 2004; 359: 183–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Schonswetter P, Stehlik I, Holderegger R, Tribsch A. Molecular evidence for glacial refugia of mountain plants in the European Alps. Mol Ecol. 2005; 14: 3547–3555. [DOI] [PubMed] [Google Scholar]
- 70. Medail F, Diadema K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J Biogeogr. 2009; 36: 1333–1345. [Google Scholar]
- 71. Stewart JR, Lister AM, Barnes I, Dalen L. Refugia revisited: individualistic responses of species in space and time. Proc Roy Soc B-Biol Sci. 2010; 277: 661–671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Qiu Y-X, Fu C-X, Comes HP. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world's most diverse temperate flora. Mol Phylogen Evol. 2011; 59: 225–244. [DOI] [PubMed] [Google Scholar]
- 73. Liu X-J, Lai Z-P, Yu L-P, Sun Y-J, Madsen D. Luminescence chronology of aeolian deposits from the Qinghai Lake area in the northeastern Qinghai-Tibetan Plateau and its paleoenvironmental implications. Quatern Geochronol. 2012; 10: 37–43. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(PDF)
Data Availability Statement
All data are available in the paper.