

Abstract
Pannexin1 (Panx1) is a gap junction gene in vertebrates whose proteins mainly function as non-junctional channels on the cell surface. Panx1 channels can release ATP under physiological conditions and play critical roles in many physiological and pathological processes. Here, we report that Panx1 deficiency can reduce ATP release and endocochlear potential (EP) generation in the cochlea inducing hearing loss. Panx1 extensively expresses in the cochlea, including the cochlear lateral wall. We found that deletion of Panx1 in the cochlear lateral wall almost abolished ATP release under physiological conditions. Positive EP is a driving force for current through hair cells to produce auditory receptor potential. EP generation requires ATP. In the Panx1 deficient mice, EP and auditory receptor potential as measured by cochlear microphonics (CM) were significantly reduced. However, no apparent hair cell loss was detected. Moreover, defect of connexin hemichannels by deletion of connexin26 (Cx26) and Cx30, which are predominant connexin isoforms in the cochlea, did not reduce ATP release under physiological conditions. These data demonstrate that Panx1 channels dominate ATP release in the cochlea ensuring EP and auditory receptor potential generation and hearing. Panx1 deficiency can reduce ATP release and EP generation causing hearing loss.
Pannexins and connexins belong to gap junction gene families in vertebrates. However, they have completely different sequences1,2. Pannexins are homologous to innexins, which encode the gap junction proteins in invertebrates2. So far, three pannexin isoforms (Panx1, 2, and 3) have been cloned from the human and mouse genomes2. The profile of pannexin channel permeability is similar to that of connexin channels, which allow passage of ions and small molecules up to 1.5 kDa. However, unlike connexins, pannexins usually form non-junctional membrane channels on the cell surface to provide an intracellular-extracellular conduit3,4,5,6. Moreover, pannexin channels, particularly Panx1 channels, can open and function at physiological extracellular Ca++ levels, at which connexin channels are closed7. Panx1 channels can also be activated by mechanical stress, low oxygen, glutamate through NMDA receptors, elevation of extracellular K+ concentrations, and ATP binding to purinergic receptors3,8,9,10,11,12,13. These specific properties imply that pannexin channels can function in a wider range of physiological conditions.
Pannexins have ubiquitous cellular expression in many tissues and organs, including the inner ear. All three pannexin isoforms are expressed in the mammalian inner ear14. Panx1 expresses at the cochlear supporting cells, the spiral limbus, and the cochlear lateral wall. Panx2 only expresses at the basal cell layer in the stria vascularis, and Panx3 expression is restricted to the cochlear bone. These distinct distribution patterns suggest that pannexins play important roles in the inner ear. However, the function of pannexin in the cochlea and hearing has not been characterized yet and remains unclear.
It has been reported that Panx1 channels can release ATP under physiological and pathological conditions3,8,10,15,16 and play critical roles in many cellular and pathological processes, such as Ca++ homeostasis, immunological responses, cell apoptosis, migraine, ischemia, and some neurological disorders9,17,18,19,20,21,22,23,24. ATP also plays important functions in the cochlea and hearing. It has been reported that ATP can elevate intracellular Ca++ concentration in hair cells to modify neurotransmission and extend the dynamic range of hearing25,26,27,28. We also found that ATP can mediate outer hair cell (OHC) electromotility to regulate hearing sensitivity, gap junctional coupling between the cochlear supporting cells, and K+-recycling29,30,31,32.
ATP is also required for generation of positive endocochlear potential (EP, +100–110 mV)33,34,35,36. Positive EP is generated in the cochlear lateral wall33,36 and is a driving force that compels K+ ions in the endolymph through the transduction channels at stereocilia of hair cells to produce auditory receptor current and potential, thereby initiating hearing. In this study, we found that deletion of Panx1 in the cochlear lateral wall reduced ATP release and EP generation and thereby reduced auditory receptor potential leading to hearing loss. This indicates that Panx1 is required for EP generation and hearing.
Results
Panx1 deletion in the cochlea in Panx1 cKO mice
As we previously reported14, Panx1 had extensive expression in the cochlea, including the spiral limbus, the organ of Corti, and the cochlear lateral wall (Fig. 1a,b). In Panx1 conditional knockout (cKO) mice, Panx1 expression at the organ of Corti and the spiral limbus remained. Immunofluorescent staining showed that intense labeling for Panx1 was retained in these regions (Fig. 1c). However, Panx1 labeling at the cochlear lateral wall was absent (Fig. 1d). At the lateral wall of WT mouse cochlea, Panx1 was found to be mainly expressed at the type II fibrocytes in the spiral ligament (SPL) (Fig. 1b). In Panx1 cKO mice, Panx1 labeling at the type II fibrocytes was completely absent (indicated by an asterisk in Fig. 1d).
Figure 1. Panx1 deletion in the cochlea in Panx1 conditional knockout (cKO) mice.

a-b: Immunofluorescent staining for Panx1 in the WT mouse cochlea. c-d: Immunofluorescent staining for Panx1 in the Panx1 cKO mouse cochlea. An asterisk indicates that the area of type II fibrocytes in the cochlear lateral wall has no Panx1 labeling. OC: the organ of Corti, OS: outer sulcus cell, SLM: spiral limbus, SPL: spiral ligament. Scale bars: 25 μm.
Deletion of Panx1 did not affect connexin expression. Cx26 and Cx30 are predominant connexin isoforms in the cochlea37,38,39. In Panx1 cKO mice, Cx26 and Cx30 expression at the cochlear lateral wall appeared normal (supplementary Fig. S2). There were no significant differences in Cx26 and Cx30 expression at the cochlear lateral wall between Panx1 cKO mice and WT mice (Cx26: P = 0.38, Cx30: P = 0.13, one-way ANOVA).
Hearing loss in Panx1 cKO mice
Panx1 cKO mice had hearing loss (Fig. 2). The thresholds of auditory brainstem response (ABR) in Panx1 cKO mice at 8, 16, 24, 32, and 40 kHz were 64.7 ± 2.11, 60.6 ± 2.77, 68.3 ± 3.45, 84.2 ± 2.19, and 92.2 ± 1.41 dB SPL, respectively (Fig. 2b). In comparison with wild-type (WT) mice, the ABR thresholds in Panx1 cKO mice at 8, 16, 24, 32, and 40 kHz were elevated by 19.7 ± 2.11, 26.1 ± 2.77, 28.3 ± 3.45, 37.7 ± 2.19, and 43.2 ± 1.41 dB SPL, respectively (P < 0.001, one-way ANOVA with a Bonferroni correction). The hearing loss appeared severe at higher frequencies.
Figure 2. Hearing loss in Panx1 cKO mice.

a: ABR waveforms recorded from Panx1 cKO and wild-type (WT) mice. b: ABR thresholds were measured at age P40-60. **P < 0.001, one-way ANOVA with a Bonferroni correction.
EP reduction in Panx1 cKO mice
The cochlear lateral wall is responsible for generation of positive EP. Fig. 3 shows that deletion of Panx1 at the cochlear lateral wall reduced EP. EP in Panx1 cKO mice and WT mice was 57.6 ± 6.03 and 98.2 ± 1.41 mV, respectively (Fig. 3b). In comparison with WT mice, the EP in Panx1 cKO mice was significantly reduced by 40% (P < 0.001, t-test). However, deletion of Panx1 did not affect the lateral wall development. The thickness of the cochlear lateral wall in Panx1 cKO mice was similar to that in WT mice (P = 0.49, one-way ANOVA, supplementary Fig. S3).
Figure 3. Reduction of EP in Panx1 cKO mice.
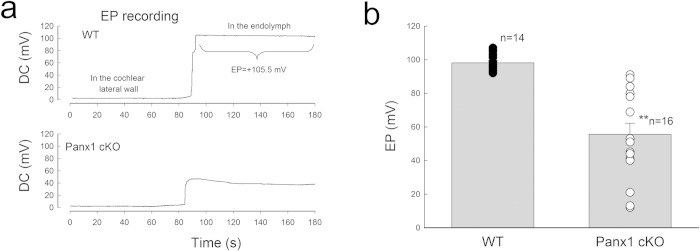
a: Time-sequence of DC potential changes in electrode penetrating through the cochlear lateral wall. b: EP reduction in Panx1 cKO mice. EP was recorded at age P40-60. ** P < 0.001, t-test.
Reduction of auditory receptor potential in Panx1 cKO mice
EP is the driving force for generation of auditory receptor current and potential. Fig. 4 shows that the auditory receptor potential as measured by cochlear microphonic (CM) in Panx1 cKO mice was reduced. In WT mice, the magnitudes of the recorded CM were 600–700 μV and had a slight increase at 26 kHz near the recording place. In Panx1 cKO mice, the magnitudes of the recorded CM were ~250 μV and significantly reduced by 60–70% (P < 0.001, one-way ANOVA with a Bonferroni correction) as compared with WT mice (Fig. 4b).
Figure 4. CM reduction in Panx1 cKO mice.
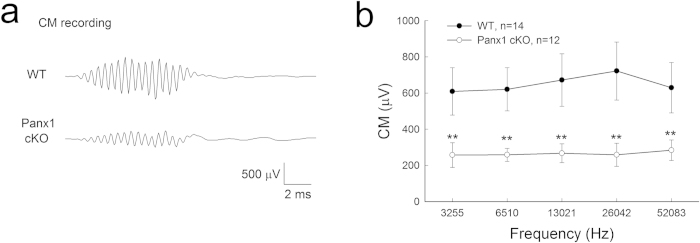
Mice were P40-60 old. a: The recorded CM waveforms from WT and Panx1 cKO mice. b: CM reduction in Panx1 cKO mice. ** P < 0.001, one-way ANOVA with a Bonferroni correction.
No apparent hair cell degeneration in Panx1 cKO mice
Fig. 5 shows that there was no apparent hair cell loss in Panx1 cKO mice. At the end of the basal turn (very high frequency region), Panx1 cKO mice had a significant increase in hair cell loss as compared with WT mice (P = 0.022, one-way ANOVA with a Bonferroni correction). However, the loss of hair cells was less than 10% (Fig. 5e), and could not produce broad, severe hearing loss as shown in Fig. 2.
Figure 5. No substantial hair cell degeneration in Panx1 cKO mice.
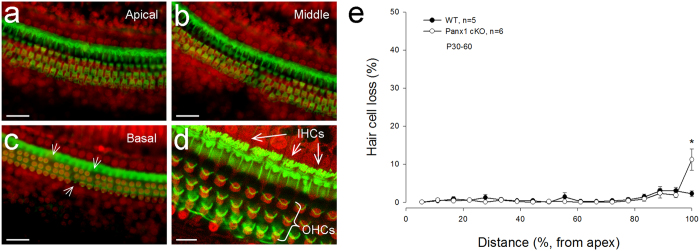
a-c: The cochlear sensory epithelia of Panx1 cKO mice staining with phalloidin-Alexa Fluor-488 (green) and propidium iodide (PI, red). White arrows in panel c indicate scattered loss of outer hair cells (OHCs) in the basal turn. Scale bars: 50 μm. d: A high-magnification image. Inner hair cell’s (IHC) and OHC’s hair bundles are clearly visible. Scale bars: 20 μm. e: Hair cell loss accounting in Panx1 cKO and WT mice. Mice were P60-90 old. * P < 0.05, one-way ANOVA with a Bonferroni correction.
Reduction of ATP release in Panx1 cKO mice
EP generation requires ATP34,35. Fig. 6 shows that deletion of Panx1 at the cochlear lateral wall almost abolished ATP release under physiological conditions. At 2 mM extracellular Ca++ concentration, the amount of ATP release in Panx1 cKO and WT mice was 1.28 ± 0.23 and 10.58 ± 0.42 fmoles, respectively (Fig. 6a). Compared to WT mice, ATP release in Panx1 cKO mice was significantly reduced by ~8-fold (P < 0.001, one-way ANOVA with a Bonferroni correction). Moreover, application of carbenoxolone (CBX), which can block pannexin channels7, could also reduce ATP release in WT mice (Fig. 6a). The amount of ATP release under 0.1 mM CBX treatment in the WT mice was significantly reduced to 1.71 ± 0.41 fmoles (P < 0.001, one-way ANOVA with a Bonferroni correction), similar to values obtained in Panx1 cKO mice. However, deletion of Cx26 or Cx30 did not reduce ATP release under the physiological level of Ca++ concentration (Fig. 6b). The measured ATP release at 2 mM extracellular Ca++ concentration in Cx26 KO mice and Cx30 KO mice was 10.87 ± 0.32 and 10.21 ± 0.41 fmoles, respectively. There was no significant difference in the amount of ATP released from WT and Cx26 KO mice or Cx30 KO mice (P = 0.98, one-way ANOVA).
Figure 6. Reduction of ATP release in the cochlea in Panx1 cKO mice.
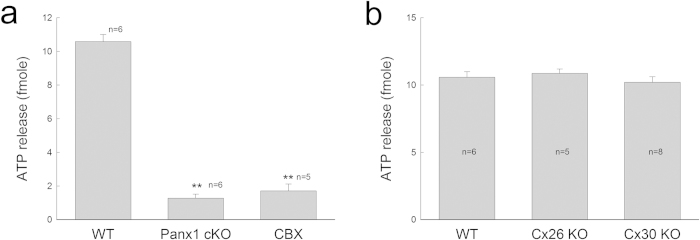
Mice were P40-60 old. a: ATP release is reduced in Panx1 cKO mice and inhibited by application of 0.1 mM CBX. **P < 0.001, one-way ANOVA with a Bonferroni correction. b: Connexin deletion does not reduce ATP release in the cochlea.
Discussion
In this study, we found that Panx1 deficiency can cause hearing loss (Figs. 1 and 2). Deletion of Panx1 at the cochlear lateral wall abolished ATP release, reduced EP and auditory receptor potential generation, and eventually led to hearing loss (Figs. 3,4,6). These data reveal that Panx1 channel-mediated ATP release is required for EP generation and auditory receptor potential producing. This study also provides the first evidence that Panx1 is required for hearing.
Positive EP (+100-110 mV) in the cochlear endolymph is a driving force that propels K+ ions through transduction channels in hair cells to produce auditory receptor current and potential. Positive EP is generated in the cochlear lateral wall by a complex process33,36. Based on a widely accepted “two-cell” model (Fig. 7), EP generation is initiated at fibrocyte cells in the spiral ligament. Na+/K+-ATPases and Na+, K+, 2Cl- cotransporters at the type II fibrocyte cells depolarize cells to ~ –5 mV. Then, the intermediate cells in the stria vascularis (SV) are consequently depolarized to ~ –5 mV through gap junctional coupling, which is formed by Cx26 and Cx3039. Subsequently, ATP-dependent Kir4.1 K+ channels at the apical membrane of the intermediate cells 39,40,41 generate a 105-110 mV transmembrane potential (Nernst’s K+ equilibrium potential) between the intracellular space and the intrastrial space, i.e., +110-115 mV in the intrastrial space with respect to normal extracellular space, since the K+ concentration in the intrastrial space is lower (1-2 mM). Finally, this positive intrastrial potential eventually leads to positive EP (+100-110 mV) in the endolymph in the scala media (Fig. 7).
Figure 7. Schematic drawing of EP generation with Panx1 ATP release in the cochlear lateral wall.

a: Based on the “two-cell” model, EP is generated by Kir4.1 in the apical membrane of intermediate cells in conjunction with Kir5.1 channels, Na+/K+ ATPases, and Na+, K+, 2Cl--cotransporters in the fibrocytes through gap junctional coupling. b: A simplified “two-cell” model of EP generation with Panx1 ATP release. ATP may activate purinergic (P2) receptors and then subsequently activate Kir K channels to ensure Na+/K+-ATPase activity and Na+, K+, 2Cl- cotransporter activity to produce EP. MC: marginal cell; BC: basal cell; FC: fibrocyte; IC: intermediate cell; IS: intrastrial space; GJ: gap junction.
Evidently, ATP is necessary and required for EP generation. It has been reported that intracochlear perfusion of ATP can significantly increase EP42. As mentioned above, depolarization of fibrocyte cells by co-activation of Na+/K+-ATPases and Na+, K+, 2Cl– -cotransporters is the first step for EP generation. Although Na+/K+-ATPase is driven by intracellular ATP, it has been reported that extracellular ATP can also stimulate Na+/K+-ATPase activity through activation of purinergic receptors and Src family kinase (SFK)43. Moreover, the function of the primary active Na+/K+-ATPase requires K channel coupling to recycle K+ (Fig. 7b). This “pump coupling” was postulated in 1958 and later corroborated by experimental data44,45,46. Recently, it has been found that extracellular ATP can also activate ATP-sensitive (Kir) K channels in hippocampal CA3 pyramidal neurons and lung epithelial cells to co-operate with Na+/K+-ATPases activity47,48. ATP may primarily activate P2X receptors and then subsequently activate Kir K channels and Na+/K+-ATPases (Fig. 7b). In this study, we found that Panx1 deletion reduced ATP release and EP (Figs. 3and 6). These findings indicate that Panx1 channel-mediated ATP release plays an important role in the EP generation. We previously also found that ATP is required for K+-recycling in the cochlea31,32. Deletion of Panx1 reduced ATP release (Fig. 6) and could also consequently compromise K+-recycling to impair EP generation (Fig. 7a).
In this study, we found that deletion of Panx1 or application CBX in WT mice abolished ATP release in the cochlear lateral wall at physiological extracellular Ca++ (2 mM) levels (Fig. 6). CBX can block both connexin and pannexin channels7,13. However, the connexin hemichannels are already closed at this high Ca++ level7. Thus, CBX in this experiment mainly blocked Panx1 channel activity. These data indicate that Panx1 channels dominate ATP release in the cochlear lateral wall under physiological conditions, even thought other ATP release mechanisms may exist. This concept is further supported by the fact that Cx26 or Cx30 deletion did not reduce ATP release at 2 mM physiological extracellular Ca++ level (Fig. 6b), at which concentration connexin hemichannels are closed7. Thus, deletion of Cx26 or Cx30 had little effect on ATP release under physiological conditions (Fig. 6b). Cx26 or Cx30 deletion could also reduce or abolish EP. However, the EP reduction results from the disruption of gap junctional coupling49,50.
Gap junctions play an important role in hearing. Connexin mutations can induce a high incidence of hearing loss, responsible for >50% of nonsyndromic deafness51. However, pannexin mutation-induced hearing loss in humans has not been identified yet. In this study, we found that Panx1 deficiency could cause hearing loss in mice (Fig. 2). This strongly suggests that Panx1 deficiency may be able to induce hearing loss in humans as well, which requires further study in the future.
Materials and Methods
Creation of Panx1 cKO mice and genotyping
Panx1tm1a(KOMP)Wtsi mice, in which Exon2 is floxed with loxPs (supplementary Fig. S1a), were purchased from KOMP (Knock Out Mouse Project, David, CA). They have a hypomorphic phenotype52 and retain Panx1 expression in the cochlea (supplementary Fig. S4). We further crossed Panx1tm1a(KOMP)Wtsi mice with Foxg1-Cre transgenic mice (Stock No. 004337, Jackson Lab, http://jaxmice.jax.org/strain/004337.html) to create Panx1 cKO mice. The genotyping of Panx1tm1a(KOMP)Wtsi mice was identified by PCR amplification with the following primers: Panx1-Mut1 (common mutant primer): 5’-GAG ATG GCG CAA CGC AAT TAA T-3’, Panx1-Mut2 (gene specific primer): 5’-CTG GCT CTC ATA ATT CTT GCC CTG-3’, Panx1-WF (wildtype-F): 5’-CTG TAT CAC ACA ACC ACT TCA GAG AAG G-3’, and Panx1-WR (wildtype-R): 5’-GAG CTG ACC CCT TTC CAT TCA ATA G-3’, and generated a 579 bp WT band and a 381 bp mutation band (supplementary Fig. S1b). The Cre transgene in the generated Panx1-Foxg1 mice was identified using following primers: CreF: 5’-GCA TTA CCG GTC GAT GCA-3’ and CreR: 5’-GAA CCT GGTCGA AAT CAG-3’, which produced ~400 bp positive band for the Cre transgene (Fig. S1c). The post-Cre Exon2-deleted Panx1 allele in Panx1-Foxg1 mice was identified by Panx1-Mut1a (post-Cre mutant primer): 5’-CAC TGC ATT CTA GTT GTG GTT TGT CC-3’ and Panx1-Mut2 (gene specific primer): 5’-CTG GCT CTC ATA ATT CTT GCC CTG-3’, which generated a 421 bp band after deletion of Exon2. The reaction would be negative for WT or no Exon2 deletion (supplementary Fig. S1d). We used homozygous Foxg1Cre:Panx1f/f mice as Panx1 cKO mice in this study. The WT littermate mice served as WT controls in the experiment. The experimental procedures were approved by the University of Kentucky’s Animal Care & Use Committee and conducted according to the standards of the NIH Guidelines for the Care and Use of Laboratory Animals.
Cx26 KO and Cx30 KO mice
As we previously reported53, Cx26 KO mice were generated by crossing Cx26loxP/loxP mice (EM00245, European Mouse Mutant Archive) with the Pax2-Cre mouse line (the Mutation Mouse Regional Center, Chapel Hill, NC). The Cx26 floxed allele and Pax2-Cre transgene were detected on tail genomic DNA by PCR amplification using the following primers: Cx26F: 5’-CTT TCC AAT GCT GGT GGA GTG-3’ and Cx26R: 5’-ACA GAA ATG TGT TGG TGA TGG-3’ for the Cx26 floxed allele; Pax2-CreF: 5’-GCC TGC ATT ACC GGT CGA TGC AAC GA- 3’ and Pax2-CreR: 5’-GTG GCA GAT GGC GCG GCA ACA CCA TT- 3’ for Pax2-Cre transgene. Cx26loxP/loxP and WT mice generated 400 and 300 bps bands, respectively. The band of Pax2-Cre was 700 bps.
Cx30 KO mice were also purchased from European Mouse Mutant Archive (EM0032349,50, ). Primer pairs for detecting Cx30 KO were Cx30 KO-1 (LACZ e Neo): 5’-GGT ACC TTC TAC TAA TTA GCT TGG -3’; Cx30 KO2 (LACZ e Neo): 5’-AGG TGG TAC CCA TTG TAG AGG AAG -3’; Cx30 KO-3 (LACZ e Neo) 5’-AGC GAG TAA CAA CCC GTC GGA TTC -3’. The bands of Cx30 KO and WT mice are located at 460 and 544 bps, respectively.
ABR measurement
ABR was measured by a Tucker-Davis’ ABR workstation (Tucker-Davis Tech. Alachua, FL)50,53,54,55. Mice were anaesthetized by intraperitoneal injection with a mixture of ketamine and xylazine (8.5 ml saline + 1 ml Ketamine + 0.55 ml Xylazine, 0.1 ml per 10 g). Body temperature was maintained at 37–38 oC. Two subdermal needle electrodes were inserted at the vertex (an active electrode) and ventrolaterally to the right or left ear (a reference electrode). The ground needle electrode was inserted in the right leg. ABR was measured by clicks in alternative polarity and series tone bursts (8 – 40 kHz) from 80 to 10 dB SPL in a 5-dB step. The ABR threshold was determined by the lowest level at which an ABR can be recognized. If the ABR threshold was greater than 75 dB SPL, the acoustic stimuli from 110 to 70 dB SPL were added.
EP and CM recording
Mice were anaesthetized as described above and the body temperature was maintained at 37–38 oC. The trachea was exposed and cut along the middle line. The tracheal tube was put into the trachea. Then, the cochlea was exposed by a ventral approach and the bone over the spiral ligament was gently picked to form a small hole50,54,55. A glass pipette filled with a K+-based intracellular solution was inserted into the hole. The DC potential was continually recorded as the electrode pipette penetrated through the lateral wall (Fig. 3a).
For CM recording, acoustic tone bursts (3.25 – 52 kHz) were delivered through an ES-1 high frequency speaker (Tucker Davis Tech. FL). The CM response was recorded by MultiClamp 700A amplifier (Molecular Devices, CA) and digitized utilizing a Digidata 1322A (Molecular Devices, CA).
ATP release measurement
As we previously reported29, the mouse temporal bone was micro-dissected in a sterile normal extracellular solution (ECS) (142 NaCl, 5.37 KCl, 1.47 MgCl2, 2 CaCl2, 10 HEPES in mM, osmolarity 310 mOsm and pH 7.2). The inner ear was opened from its apex to base. After removal of the bone, the exposed cochlear lateral wall was dissected and put into an incubation chamber. For testing Panx1 channel ATP release, the isolated lateral wall was incubated in 200 μL ECS, which contains 2 mM Ca++ and 1.47 mM Mg++ to block connexin hemichannels, for 10 minutes. Every 5 minutes, the whole incubation solution (200 μL) was collected and replaced with fresh solution. The osmolarity of all solutions was measured by a micro-computer controlled osmometer (Model 3300, Advanced Instruments Inc. Norwood, MA) and adjusted by Dextrose. All experiments were performed at room temperature (23 oC).
The collected incubation solutions were kept on ice. As we previously reported29, the amount of ATP was measured by a bioluminescence method with a luciferin-luciferase assay kit (FL-ASC, Sigma) using a black 96-well plate to avoid optical cross-talk. The bioluminescence was read by a Biotek Synergy 4 Hybrid Microplate Reader (Biotek Instruments Inc, Winooski, VT, USA). The amount of ATP was calculated from the ATP standard curve, which was established from measurements of serial-diluted ATP standards (supplementary Fig. S5). The standard curve also served as an internal control for bioluminescence measurement.
Immunofluorescent staining and quantitative measurement
The detailed methods and procedures of immunofluorescent staining can be found in our previous reports14,38,39. The cochlea was fixed with 4% paraformaldehyde. The cochlear cryostat sections were washed with PBS for 5 min twice and incubated in a blocking solution (10% goat serum and 1% bovine serum albumin) with 0.1% Triton X-100 for 30 min. Then, the tissue sections were incubated with chicken anti-human Panx1 antibody (1:500; #4515, a gift from Dr. Gerhard Dahl at the University of Miami Medical School) in the blocking solution overnight. For double immunofluorescent staining, monoclonal mouse anti-Cx26 antibody (1: 400, Cat# 33-5800, Invitrogen, CA) and polyclonal rabbit anti-Cx30 antibody (1:400, #71-2200, Invitrogen, CA) were used. After being washed with PBS three times, the sections were reacted with corresponding Alexa Fluor 488- or 568 secondary antibodies (1:500, Molecular Probes) for 2 hr at room temperature (23 oC). The staining was observed under a fluorescent microscope or laser confocal microscope. The images were saved in TIFF format for analysis and presentation.
For quantitative measurement of Cx26 and C30 labeling at the cochlear lateral wall, ImageJ software (NIH, Bethesda, MD) was used38,55. The density of labeling at the lateral wall was measured and compared between WT and Panx1 cKO mice.
Statistical analysis
Data were expressed as mean ± s.e.m. unless otherwise indicated in text and plotted by SigmaPlot (SPSS Inc. Chicago, IL). The statistical analyses were performed by SPSS v18.0 (SPSS Inc. Chicago, IL) using one-way ANOVA with a Bonferroni correction or t-test.
Additional Information
How to cite this article: Chen, J. et al. Pannexin1 channels dominate ATP release in the cochlea ensuring endocochlear potential and auditory receptor potential generation and hearing. Sci. Rep. 5, 10762; doi: 10.1038/srep10762 (2015).
Supplementary Material
Acknowledgments
We are grateful to Dr. Willis at University of California Davis for technical support on Panx1 knockout and genotyping, Dr. Dahl at Miami University for kindly providing anti-Panx1 antibody, and Dr. Liang Zong for technical help on mouse breeding and genotyping at the early stage of this study. This work was supported by NIH (R01) DC 05989.
Footnotes
Author Contributions H.-B.Z. conceived the general framework of this study. J.C., Y.Z., C.L., J.C. and H.-B.Z. performed the experiments and analyzed data. HBZ wrote the paper. All authors have read the manuscript and provided input.
References
- Bruzzone R., Hormuzdi S. G., Barbe M. T., Herb A. & Monyer H. Pannexins, a family of gap junction proteins expressed in brain. Proc Natl Acad Sci USA 100, 13644–13649 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baranova A. et al. The mammalian pannexin family is homologous to the invertebrate innexin gap junction proteins. Genomics 83, 706–716 (2004). [DOI] [PubMed] [Google Scholar]
- Locovei S., Bao L. & Dahl G. Pannexin 1 in erythrocytes: function without a gap. Proc Natl Acad Sci USA 103, 7655–7659 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boassa D. et al. Pannexin1 channels contain a glycosylation site that targets the hexamer to the plasma membrane. J Biol Chem. 282, 31733–31743 (2007). [DOI] [PubMed] [Google Scholar]
- Penuela S. et al. Pannexin 1 and pannexin 3 are glycoproteins that exhibit many distinct characteristics from the connexin family of gap junction proteins. J Cell Sci. 120, 3772–3783 (2007). [DOI] [PubMed] [Google Scholar]
- Sosinsky G. E. et al. Pannexin channels are not gap junction hemichannels. Channels (Austin) 5, 193–197 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruzzone R., Barbe M. T., Jakob N. J. & Monyer H. Pharmacological properties of homomeric and heteromeric pannexin hemichannels expressed in Xenopus oocytes. J Neurochem 92, 1033–1043 (2005). [DOI] [PubMed] [Google Scholar]
- Bao L., Locovei S. & Dahl G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett 572, 65–68 (2004). [DOI] [PubMed] [Google Scholar]
- Thompson R. J., Zhou N. & MacVicar B. A. Ischemia opens neuronal gap junction hemichannels. Science 312, 924–927 (2006). [DOI] [PubMed] [Google Scholar]
- Sridharan M. et al. Pannexin 1 is the conduit for low oxygen tension-induced ATP release from human erythrocytes. Am J Physiol Heart Circ Physiol. 299, H1146–1152, (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bargiotas P. et al. Pannexins in ischemia-induced neurodegeneration. Proc Natl Acad Sci USA. 108, 20772–20777 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahl G. & Keane R. W. Pannexin: from discovery to bedside in 11±4 years? Brain Res 1487, 150–159 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penuela S., Gehi R. & Laird D. W. The biochemistry and function of pannexin channels. Biochim Biophys Acta 1828, 15–22 (2013). [DOI] [PubMed] [Google Scholar]
- Wang X. H., Streeter M., Liu Y. P. & Zhao H. B. Identification and characterization of pannexin expression in the mammalian cochlea. J Comp Neurol 512, 336–346 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Y. J. et al. The role of pannexin 1 hemichannels in ATP release and cell-cell communication in mouse taste buds. Proc Natl Acad Sci USA 104, 6436–6441 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ransford G. A. et al. Pannexin 1 contributes to ATP release in airway epithelia. Am J Respir Cell Mol Biol 41, 525–534 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanden-Abeele F. et al. Functional implications of calcium permeability of the channel formed by pannexin1. J Cell Biol 174, 535–546 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silverman W. R. et al. The pannexin 1 channel activates the inflammasome in neurons and astrocytes. J Biol Chem. 284, 18143–18151 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chekeni F. B. et al. Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during apoptosis. Nature 467, 863–867 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishikawa M. et al. Pannexin 3 functions as an ER Ca2+ channel, hemichannel, and gap junction to promote osteoblast differentiation. J Cell Biol 193, 1257–1274 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qu Y. et al. Pannexin-1 is required for ATP release during apoptosis but not for inflammasome activation. J Immunol 186, 6553–6561 (2011). [DOI] [PubMed] [Google Scholar]
- Gulbransen B. D. et al. Activation of neuronal P2×7 receptor-pannexin-1 mediates death of enteric neurons during colitis. Nat Med 18, 600–604 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karatas H. et al. Spreading depression triggers headache by activating neuronal Panx1 channels. Science 339, 1092–1095 (2013). [DOI] [PubMed] [Google Scholar]
- Jackson D. G., Wang J., Keane R. W., Scemes E. & Dahl G. ATP and potassium ions: a deadly combination for astrocytes. Sci Rep 4, 4576. 10.1038/srep04576 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashmore J. F. & Ohmori H. Control of intracellular calcium by ATP in isolated outer hair cells of the guinea-pig cochlea. J Physiol 428, 109–131 (1990). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dulon D., Mollard P. & Aran J. M. Extracellular ATP elevates cytosolic Ca2+ in cochlear inner hair cells. Neuroreport 2, 69–72 (1991). [DOI] [PubMed] [Google Scholar]
- Housley G. D., Bringmann A. & Reichenbach A. Purinergic signaling in special senses. Trends Neurosci. 32, 128–141 (2009). [DOI] [PubMed] [Google Scholar]
- Housley G. D. et al. ATP-gated ion channels mediate adaptation to elevated sound levels. Proc Natl Acad Sci USA. 110, 7494–7499 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao H. B., Yu N. & Fleming C. R. Gap junctional hemichannel-mediated ATP release and hearing controls in the inner ear. Proc Natl Acad Sci USA. 102, 18724–18729 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu N. & Zhao H. B. ATP activates P2x receptors and requires extracellular Ca++ participation to modify outer hair cell nonlinear capacitance. Pflugers Arch. 457, 453–461 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Y. & Zhao H. B. ATP-mediated potassium recycling in the cochlear supporting cells. Purinergic Signal. 6, 221–229 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Y. & Zhao H. B. ATP activates P2X receptors to mediate gap junctional coupling in the cochlea. Biochem Biophys Res Commun. 426, 528–532 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wangemann P. & Schacht J. Homeostatic mechanisms in the cochlea., in The Cochlea. (eds Dallos P., Popper A. N. & Fay R. R. ) 130–185 (New York, Springer-Verlag; 1996). [Google Scholar]
- Muñoz D. J., Thorne P. R. & Housley G. D. P2X receptor-mediated changes in cochlear potentials arising from exogenous adenosine 5’-triphosphate in endolymph. Hear Res. 138, 56–64 (1999). [DOI] [PubMed] [Google Scholar]
- Telang R. S. et al. Reduced P2x(2) receptor-mediated regulation of endocochlear potential in the ageing mouse cochlea. Purinergic Signal. 6, 263–272 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J. & Zhao H. B. The role of an inwardly rectifying K+ channel (Kir4.1) in the inner ear and hearing loss. Neuroscience 265, 137–146 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forge A. et al. Gap junctions in the inner ear: comparison of distribution patterns in different vertebrates and assessement of connexin composition in mammals. J Comp Neurol. 467, 207–231 (2003). [DOI] [PubMed] [Google Scholar]
- Zhao H. B. & Yu N. Distinct and gradient distributions of connexin26 and connexin30 in the cochlear sensory epithelium of guinea pigs. J Comp Neurol 499, 506–518 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y. P. & Zhao H. B. Cellular characterization of Connexin26 and Connexin30 expression in the cochlear lateral wall. Cell Tissue Res 333, 395–403 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ando M. & Takeuchi S. Immunological identification of an inward rectifier K+ channel (Kir4.1) in the intermediate cell (melanocyte) of the cochlear stria vascularis of gerbils and rats. Cell Tissue Res 298, 179–183 (1999). [DOI] [PubMed] [Google Scholar]
- Nin F. et al. The endocochlear potential depends on two K+ diffusion potentials and an electrical barrier in the stria vascularis of the inner ear. Proc Natl Acad Sci USA. 105, 1751–1756 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sueta T., Paki B., Everett A. W. & Robertson D. Purinergic receptors in auditory neurotransmission. Hear Res. 183, 97–108 (2003). [DOI] [PubMed] [Google Scholar]
- Shahidullah M., Mandal A., Beimgraben C. & Delamere N. A. Hyposmotic stress causes ATP release and stimulates Na,K-ATPase activity in porcine lens. J Cell Physiol. 227, 1428–1437 (2012). [DOI] [PubMed] [Google Scholar]
- Koefoed-Johnsen V. & Ussing H. H. The nature of the frog skin potential. Acta Physiol Scand. 42, 298–308 (1958). [DOI] [PubMed] [Google Scholar]
- Dawson D. C. & Richards N. W. Basolateral K conductance: role in regulation of NaCl absorption and secretion. Am J Physiol. 259, C181–C195 (1990). [DOI] [PubMed] [Google Scholar]
- Tsuchiya K., Wang W., Giebisch G. & Welling P. A. ATP is a coupling modulator of parallel Na,K-ATPase-K-channel activity in the renal proximal tubule. Proc Natl Acad Sci USA. 89, 6418–6422 (1992). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawamura M. Jr., Ruskin D. N. & Masino S. A. Metabolic autocrine regulation of neurons involves cooperation among pannexin hemichannels, adenosine receptors, and KATP channels. J Neurosci. 30, 3886–3895 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richter K., Kiefer K. P., Grzesik B. A., Clauss W. G. & Fronius M. Hydrostatic pressure activates ATP-sensitive K+ channels in lung epithelium by ATP release through pannexin and connexin hemichannels. FASEB J. 28, 45–55 (2014). [DOI] [PubMed] [Google Scholar]
- Teubner B. et al. Connexin30 (Gjb6)-deficiency causes severe hearing impairment and lack of endocochlear potential. Hum Mol Genet 12, 13–21 (2003). [DOI] [PubMed] [Google Scholar]
- Chen J., Chen J., Zhu Y., Liang C. & Zhao H. B. Deafness induced by Connexin26 (GJB2) deficiency is not determined by endocochlear potential (EP) reduction but is associated with cochlear developmental disorders. Biochem Biophys Res Commun. 448, 28–32 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castillo F. J. & Castillo I. The DFNB1 subtype of autosomal recessive non-syndromic hearing impairment. Front Biosci 17, 3252–3274 (2011). [DOI] [PubMed] [Google Scholar]
- Hanstein R. et al. Promises and pitfalls of a Pannexin1 transgenic mouse line. Front Pharmacol. 4, 61. 10.3389/fphar.2013.00061 (2013). [DOI] [PMC free article] [PubMed]
- Liang C., Zhu Y., Zong L., Lu G. J. & Zhao H. B. Cell degeneration is not a primary causer for Connexin26 (GJB2) deficiency associated hearing loss. Neurosci Lett. 528, 36–41 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Y. et al. Active cochlear amplification is dependent on supporting cell gap junctions. Nat Commun 4, 1786. 10.1038/ncomms2806 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Y. et al. Connexin26 (GJB2) deficiency reduces active cochlear amplification leading to late-onset hearing loss. Neuroscience. 284, 719–729 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.