

Abstract
Background
Haemaphysalis longicornis is a major vector of Theileria spp., Anaplasma phagocytophilum, Babesia spp. and Coxiella burnetti in East Asian countries. All life stages of ixodid ticks have a destructive pool-feeding style in which they create a pool-feeding site by lacerating host tissue and secreting a variety of biologically active compounds that allows the tick to evade host responses, enabling the uptake of a blood meal. The identification and functional characterization of tick saliva proteins can be useful to elucidate the molecular mechanisms involved in tick development and to conceive new anti-tick control methods.
Methods
H. longicornis tick saliva was collected from fully engorged nymphs and fully engorged adults induced by dopamine or pilocarpine, respectively. Saliva was digested with trypsin for LC-MS/MS sequencing and peptides were searched against tick and rabbit sequences.
Results
A total of 275 proteins were identified, of which 135 were tick and 100 were rabbit proteins. Of the tick proteins, 30 proteins were identified exclusively in fully engorged nymph saliva, 74 in fully engorged adult females, and 31 were detected in both stages. The identified tick proteins include heme/iron metabolism-related proteins, oxidation/detoxification proteins, enzymes, proteinase inhibitors, tick-specific protein families, and cytoskeletal proteins. Proteins involved in signal transduction, transport and metabolism of carbohydrate, energy, nucleotide, amino acids and lipids were also detected. Of the rabbit proteins, 13 were present in nymph saliva, 48 in adult saliva, and 30 were present in both. The host proteins include immunoglobulins, complement system proteins, antimicrobial proteins, serum albumin, peroxiredoxin, serotransferrin, apolipoprotein, hemopexin, proteinase inhibitors, and hemoglobin/red blood cells-related products.
Conclusions
This study allows the identification of H. longicornis saliva proteins. In spontaneously detached tick saliva various proteins were identified, although results obtained with saliva of fully engorged ticks need to be carefully interpreted. However, it is interesting to note that proteins identified in this study were also described in other tick saliva proteomes using partially engorged tick saliva, including hemelipoprotein, proteases, protease inhibitors, proteins related to structural functions, transporter activity, metabolic processes, and others. In conclusion, these data can provide a deeper understanding to the biology of H. longicornis.
Electronic supplementary material
The online version of this article (doi:10.1186/s13071-015-0918-y) contains supplementary material, which is available to authorized users.
Keywords: Tick, Proteomic, Saliva, Tick-host relationship
Background
The hard tick Haemaphysalis longicornis is a medically and veterinary important vector of many tick-borne disease (TBD), transmitting pathogens such as Ehrlichia chaffeensis [1], Anaplasma bovis [2], A. phagocytophilum [3], Coxiella burnetii [4], and Spotted fever group rickettsiae [5]. Of significant veterinary importance, this tick species is considered the primary vector of theileriosis caused by Theileria spp. and of babesiosis caused by Babesia spp. in both sheep and cattle in East Asia [6, 7]. As a three-host tick, it has a wide range of hosts, from birds and lagomorphs (as immature ticks), and large domestic and wild mammals (as adult parasites). This tick is vastly distributed throughout Eastern Asian countries such as China, Korea, Japan, New Zealand, and Australia [8].
The tick feeding process is initiated when the tick engages and attaches onto its host. As a pool feeder, upon attachment the tick determines a suitable feeding site, and prepares it by lacerating small blood vessels. Feeding occurs by sucking up the blood that flows to the pool. This feeding style triggers host defense mechanisms such as pain or itching, hemostasis, inflammatory reactions, tissue repair, and immune rejection [9–12]. To control the feeding site and counteract the host defenses, ticks secrete and inject saliva into its host, of which contains hundreds of different proteins [7, 13–16] and other pharmacologically active molecules that confer anti-hemostatic, anti-inflammatory and immunomodulatory properties, supporting blood feeding [17–20].
During the feeding process, infected ticks may transmit TBD-causing pathogens. Besides being a critical component of the feeding process, saliva has also been shown to play a role in pathogen transmission [21]. Therefore, the identification and characterization of novel H. longicornis saliva proteins could point to candidates for the development of anti-tick and transmission-blocking vaccines [22–26] and of new pharmacological active molecules for medical application [18, 20, 27–29].
Currently the saliva proteome of Amblyomma americanum, Ixodes scapularis, Ornithodoros moubata, Rhipicephalus sanguineus sensu lato, R. microplus, and Dermacentor andersoni [13–16, 30–33] tick species have been analyzed. However, H. longicornis tick saliva proteome has not been the object of any analysis. The objectives of this study were to identify secreted proteins in the saliva of fully engorged nymphs (nymphs) and fully engorged adult females (adults) of H. longicornis ticks, comparing the protein profile of these developmental tick stages to evaluate the variation in tick saliva during feeding of different life stages. This affords to identify tick saliva proteins shared by the two developmental stages. Such proteins may play an important role in the success of both developmental stages in their feeding cycle. The novel catalog of tick saliva proteins identified in this study provides a deeper understanding to the biology of H. longicornis.
Methods
Ethics statement ethical approval
All animals used in these experiments were housed in Laboratory of Veterinary Parasitology, College of Veterinary Medicine and Bio-Safety Research Institute in Chonbuk National University, Jeonju 561–756, Republic of Korea. All animal studies and protocols are in agreement with the ethical principles for animal research and approved by the Chonbuk Animal Care and Use Committee (CBNU 2015–003).
Ticks and saliva collection
The Jeju strain of the hard tick H. longicornis has been maintained on rabbits in our laboratory since 2003. To feed, H. longicornis ticks were placed onto the ears of specific pathogen-free (SPF) New Zealand White rabbits. Ticks were restricted to the ear using cloth pocket-like socks attached on ear ending with tape. Approximately 150 nymphs and 150 adults were placed in their respective feeding apparatuses and allowed to feed upon full engorgement and spontaneous detachment. Saliva was collected from 90 nymphs and 50 adults that were fully engorged and that detached from the rabbits spontaneously. Ticks were rinsed with sterile distilled water and induced to salivate by dorsal injection (posterior to fourth coxae in the region of epimeral and anal plates) of 5.0 to 7.0 μL 0.2 % dopamine or 1.5 to 3.0 μl 2 % pilocarpine (in 10 mM Tris-EDTA buffer) for nymphs and adults, respectively. Injections were applied using a micro-fine especially homemade glass needle. Then, ticks were maintained at 37 °C in an 85 % relative humidity chamber, and saliva was periodically collected for approximately 4 h using a pipette tip. Tick saliva was lyophilized and stored at −80 °C for LC-MS/MS analysis. Saliva protein concentrations were determined by Qubitfluorometer (Life Technologies, Carlsbad, CA, USA).
Protein digestion and sample preparation
Saliva of H. longicornis nymphs and adult females was digested in solution with trypsin. Lyophilized salivary proteins were dissolved in 8 M urea/0.1 M Tris, pH 8.5, reduced with 5 mMTris(2-carboxyethyl)phosphine hydrochloride (TCEP, Sigma-Aldrich, St Louis, MO, USA) and alkylated with 25 mM iodoaceamide (Sigma-Aldrich). Proteins were digested overnight at 37 °C in 2 M urea/0.1 M Tris pH 8.5, 1 mM CaCl2 with trypsin (Promega, Madison, WI, USA) at a final 1:20 ratio (enzyme:substrate). Digestions were stopped with formic acid (5 % final concentration) and centrifuged for debris removal.
Precolumns and analytical columns
Reversed phase pre-columns were prepared by first creating a Kasil frit at one end of a deactivated 250-μm ID/360-μm OD capillary (Agilent Technologies, Santa Clara, CA, USA). Kasil frits were prepared by dipping 20 cm capillary in 300 μL Kasil 1624 (PQ Corporation, Malvern, PA, USA) and 100 μL formamide solution, curing at 100 °C for 3 h, and cutting the frit to a length of 2 mm. Pre-columns were packed in-house with 5 μm ODS-AQ C18 (YMC America, INC., Allentown, PA, USA) particles from particle slurries in methanol upon reaching the height of 2 cm. Analytical reversed phase columns were assembled by pulling a 100-μm ID/360-μm OD (Molex Polymicro Technologies™, Austin, TX, USA) silica capillary to a 5 μm ID tip. The same packing material was packed directly into the pulled column until the length of 20 cm was reached. Reversed phase precolumns and analytical columns were connected using a zero-dead volume union (IDEX Corp., Upchurch Scientific, Oak Harbor, WA, USA).
LC-MS/MS
Peptide mixtures were analyzed by nanoflow liquid chromatography mass spectrometry using an Easy NanoLC II and a Q Exactive mass spectrometer (Thermo Scientific, Waltham, MA, USA). Peptides eluted from the analytical column were electrosprayed directly into the mass spectrometer. Buffer A and B consisted of 5 % acetonitrile/0.1 % formic acid and 80 % acetonitrile/0.1 % formic acid, respectively. The flow rate was set to 400 nL/min. H. longicornis digested saliva samples (1.5 μg per injection) were separated in 155 min chromatographic runs, with linear gradient from 1 to 10 % of buffer B for 10 min followed by an increase to 40 % of buffer B in 100 min, an increase to 50 % of buffer B in 10 min and finally an increase to 90 % of buffer B for additional 10 min. Column was held at 90 % of buffer B for 10 min, reduced to 1 % of buffer B and re-equilibrated prior to the next injection.
The mass spectrometer was operated in a data dependent mode, collecting a full MS scan from 400 to 1200 m/z at 70,000 resolution and an AGC target of 1 × 106. The 10 most abundant ions per scan were selected for MS/MS at 17,500 resolution and AGC target of 2 × 105 and an underfill ratio of 0.1 %. Maximum fill times were 20 and 120 ms for MS and MS/MS scans, respectively, with dynamic exclusion of 15 s. Normalized collision energy was set to 25.
Data analysis
Tandem mass spectra were extracted from raw files using RawExtract 1.9.9.2 [34] and searched with ProLuCID [35] against a combined non-redundant database containing (i) Ixodidae database from NCBI; (ii) R. microplus transcriptome database (Rm-INCT-EM, containing 22,009 sequences produced by our research group using Illumina Sequencing technology – BioProject ID PRJNA232001 at Transcriptome Shotgun Assembly (TSA) database – GenBank); (iii) Oryctolagus cuniculus database from SwissProt and (iv) reverse sequences of all database entries. Searches were done using an Integrated Proteomics Pipeline – IP2 (Integrated Proteomics Applications) for The Scripps Institute (La Jolla, CA, USA). The search space included all fully-tryptic and half-tryptic peptide candidates. Carbamidomethylation on cysteine was used as static modification. Data was searched with 50-ppm precursor ion tolerance and 20-ppm fragment ion tolerance.
The validity of the peptide spectrum matches (PSMs) generated by ProLuCID was assessed using Search Engine Processor (SEPro) [36]. Identifications were grouped by charge state and tryptic status, resulting in four distinct subgroups. For each group, ProLuCIDXCorr, DeltaCN, DeltaMass, ZScore, number of peaks matched and Spec Count Score values were used to generate a Bayesian discriminator. The identifications were sorted in a nondecreasing order according to the discriminator score. A cutoff score was established to accept a false discovery rate (FDR) of 1 % based on the number of decoys. This procedure was independently performed on each data subset, resulting in a false-positive rate that was independent of tryptic status or charge state. Additionally, a minimum sequence length of six residues per peptide was required. Results were post-processed to only accept PSMs with < 10 ppm precursor mass error.
A Volcano plot was generated by a pairwise comparison between nymphs and fully engorged female H. longicornis tick saliva using the TFold module from PatternLab for Proteomics platform [37]. The following parameters were used to select differentially expressed proteins: proteins were grouped by maximum parsimony, spectral count data was normalized using normalized spectral abundance factor (NSAF) [38], and two nonzero replicate values were required for each condition (at least two out of four replicates). A BH q-value was set at 0.02 (2 % FDR). A variable fold-change cutoff for each individual protein was calculated according to the t-test p-value using an F-Stringency value automatically optimized by the TFold software. Low abundant proteins were removed using an L-stringency value of 0.4.
Venn diagrams were manually generated from the output of PatternLab’s Birds Eye view report. Proteins were grouped by maximum parsimony and the presence of proteins in at least two out of four replicates was required for each condition.
Functional annotation and classification
To gain insight on the nature of the identified protein sequences, BLASTp searches against several databases were performed. To check the identity of tick saliva proteins detected, several databases were used for screening: non-redundant (NR), Acari and refseq-invertebrate from NCBI; Acari from Swissprot; the GeneOntology (GO) FASTA subset [39]; MEROPS database; and the conserved domains database of NCBI [40] containing the KOG [41], PFAM [42], and SMART motifs [43]. To check rabbit proteins identity, we used Oryctolagus cuniculus and refseq-vertebrates databases from NCBI; the conserved domains database of NCBI [40] containing the KOG [41], PFAM [42], and SMART motifs [43]; and the GeneOntology (GO) FASTA subset [39]. To functionally classify the protein sequences, a program written and provided by Dr. José M. C. Ribeiro in Visual Basic 6.0 (Microsoft, Redmond, Washington, USA) was used [44]. The functionally annotated catalog for each dataset was manually curated and plotted in a hyperlinked Excel spreadsheet (Additional file 1: Table S1 and Additional file 2: Table S2).
Results and discussion
H. longicornis is the primary tick known to transmit disease pathogens to humans and animals in East Asian countries [45]. Like several other blood-sucking parasites, its saliva secretion is a mixture of proteins produced in different salivary gland acinar cells [46]. Tick salivation can be studied in vivo by injection of dopamine, a neurotransmitter that stimulates fluid secretion by salivary gland, or by using pilocarpine, a cholinomimetic agent that induces the release of dopamine from the salivary nerves, resulting in salivation [47–49]. In Ornithodoros moubata, comparative analysis by SDS-PAGE of several pilocarpine- and dopamine-induced saliva batches demonstrated reproducibility between both protocols used to induce tick salivation [13]. We have tested the use of pilocarpine and/or dopamine in both adult and nymph saliva collection. However, using pilocarpine for nymph saliva collection did not result in a substantial amount of saliva. When dopamine was used in adult ticks, the saliva acquired a darker color (data not shown). This observation using dopamine for adult saliva stimulation is in accordance with results described for R. sanguineus s.l. [15]. However, differently from data obtained for R. sanguineus s.l. adult tick saliva, we successfully collected and identified proteins from H. longicornis nymph saliva induced by dopamine. These differences could be associated to stage and/or species of ticks used in the salivation induction. Few tick saliva proteomes have been published, most of which using adult ticks and, therefore, knowledge about nymph saliva collection is not available.
At the time study was carried out, H. longicornis genome and transcriptome sequences were not available, therefore we screened against available Ixodidae databases from NCBI and an in-house R. microplus transcriptome database to identify tick salivary proteins by shotgun proteomics. Additionally, saliva was screened for host proteins using available rabbit protein sequences from Uniprot. The comparison in proteomic content between samples from different life cycle stages may show unique or increased levels of particular proteins that are important for tick biology. Moreover, it is possible to provide information about semi-quantitative variations on the levels of the specific proteins. Comparative proteomics of spontaneously detached fully engorged nymph and fully engorged adult females H. longicornis have shown alterations in protein salivary content through different life cycle stages, generating new insights into tick physiology. The main objective in this study was to evaluate the H. longicornis saliva proteome, showing saliva protein content in nymphs, as an immature stage, and in fully engorged adult females, as an experimental end-point, evaluating not only differences, but also the similarities in saliva contents at different stages of tick development. It is important to observe that analyses of saliva of fully engorged ticks have to be carefully interpreted, since they are at the end of the feeding process. However, this analysis can provide useful information, since various categories of proteins identified in this study were also described in other tick saliva proteomes using partially engorged tick saliva [13–16], including: proteins related to heme/iron metabolism (hemelipoprotein, ferritin), proteases (cathepsin, trypsin-like, metalloprotease), protease inhibitors (serpin, cystatin, alpha-2-macroglobulin, TIL), proteins related to structural functions, transporter activity, metabolic processes, protein modification machinery (heat shock proteins), and others. In addition, other proteins from fully engorged ticks have been characterized as anticoagulant molecules, like microphilin [27], BmAP [28], and haemalin [29]. Therefore, despite the limitation in using fully engorged adult saliva, the biological interpretation of these data can provide a deeper understanding to the biology of H. longicornis.
An overview of identified saliva proteins
Tick saliva was obtained by dopamine and pilocarpine stimulations from H. longicornis nymphs and adult females, respectively. The saliva accumulated in the mouthparts was periodically collected from the ticks using a pipette tip (Fig. 1a and b). Crystal particles formed around the nymph mouthpart were also collected (Fig. 1c). Collected saliva was subjected to tryptic digestion and analyzed by shotgun proteomics in quadruplicate. A total of 135 proteins were identified matching tick databases, and 100 proteins matching rabbit database (Fig. 2 and Tables 1, 2 and 3). The identified tick proteins were classified and divided into groups according to their putative functions (Fig. 3 and Tables 1, 2 and 3), consistent with previously published tick sialomes [9]. In the set of tick specific proteins, 30 proteins were identified exclusively in nymph saliva, 74 proteins were identified exclusively in adult saliva, and 31 proteins were detected in both stages (Fig. 2). Of those 31 identified tick proteins detected in both stages, 11 had statistical differential expression confirmed (Fig. 4 and Table 4). This finding is discussed below. These proteins identified in nymph saliva as well as in adult saliva can be secreted through developmental stages throughout the tick feeding process, since proteomic studies using saliva from other tick species collected during feeding process have described the presence of similar classes of proteins as identified in this study [14–16]. Moreover, some of these proteins have been used in anti-tick vaccination experiments, e.g. glutathione S-transferase [50, 51], cystatin [52], ferritin [53, 54], serpins [23, 55–59], and hemelipoproteins [60, 61].
Fig. 1.

Haemaphysalis longicornis saliva collection. a Saliva collection from a fully engorged adult female. b Saliva collection from a fully engorged nymph. c After application of dopamine, saliva appears as crystals in nymph mouthparts (arrows). Insert shows a magnified image of crystals in mouthparts. Bars = 100 μm
Fig. 2.
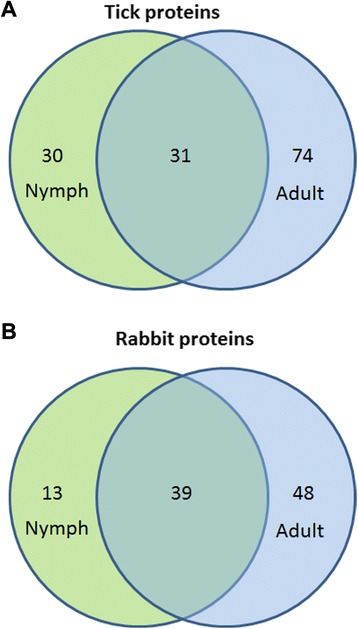
Venn diagram of Haemaphysalis longicornis saliva proteins identified in fully engorged nymphs (nymph) and fully engorged adult females. a Tick and b rabbit proteins identified in tick saliva. The overlap between circles shows the proteins present in both stages
Table 1.
Tick and host proteins identified exclusively in nymph saliva
| Contig numbera | Annotation | Class | Spec count |
|---|---|---|---|
| TICK | |||
| BAF36722 | glycine-rich cell wall structural protein | Glycine-rich protein | 1.00 |
| Rm-6837 | glycine proline-rich secreted protein | Glycine-rich protein | 5.25 |
| Rm-21891 | actin-depolymerizing factor | Cytoskeletal | 3.00 |
| JAA63693 | substrate adhesion-dependent cell spreading | Extracellular matrix/cell adhesion | 2.50 |
| JAA55046 | toll-like receptor 5 | Immunity | 5.00 |
| ACX33152 | catalytically inactive chitinase-like lectins | Metabolism, carbohydrate | 5.00 |
| JAA63210 | lysosomal alpha-mannosidase-like isoform X2 | Metabolism, carbohydrate | 2.00 |
| JAA54599 | phospholipase A2 | Metabolism, lipid | 6.00 |
| Rm-32843 | lysosomal & prostatic acid phosphatase | Protein modification machinery | 4.66 |
| BAF31119 | secreted protein | Secreted conserved protein | 7.75 |
| AEO33293 | antigen 5/SCP | Secreted conserved protein | 5.75 |
| AEO36255 | secreted protein | Secreted conserved protein | 1.66 |
| JAA60619 | secreted protein | Secreted conserved protein | 5.50 |
| Rm-9069 | secreted protein | Secreted conserved protein | 1.50 |
| BAE02551 | lipocalin | Lipocalin | 5.50 |
| BAF43801 | longipain | Proteinase | 5.75 |
| DAA34687 | cathepsin L-like | Proteinase | 1.50 |
| JAA62284 | tick serine protease | Proteinase | 1.50 |
| Rm-44814 | serine carboxypeptidase | Proteinase | 2.00 |
| ACJ26770 | alpha-2-macroglobulin | Proteinase inhibitor | 27.75 |
| EEC05896 | serpin | Proteinase inhibitor | 1.00 |
| AEO34218 | serpin | Proteinase inhibitor | 2.33 |
| AEO34349 | serpin | Proteinase inhibitor | 4.75 |
| JAB70612 | alpha-2-macroglobulin | Proteinase inhibitor | 1.25 |
| Rm-12491 | serpin | Proteinase inhibitor | 4.00 |
| AEO35364 | cystatin | Proteinase inhibitor | 1.25 |
| EEC02492 | TGF-beta-induced protein ig-h3 | Signal transduction | 5.25 |
| JAA58175 | chorion peroxidase | Heme/iron metabolism | 5.50 |
| BAN13552 | ferritin | Heme/iron metabolism | 5.00 |
| JAA54618 | metabotropic glutamate receptor | Transporters/receptors | 2.25 |
| RABBIT | |||
| G1T4V7 | myosin tail | Cytoskeletal | 6.25 |
| G1T4A5 | collagen alpha-1 (I) chain-like | Extracellular matrix/cell adhesion | 4.00 |
| P25227 | alpha-1-acid glycoprotein 1 | Immunity | 1.66 |
| G1TJG6 | glyceraldehyde-3-phosphate dehydrogenase | Metabolism, energy | 1.50 |
| G1TFX2 | alpha-1-antiproteinase | Proteinase inhibitor | 10.00 |
| Q28665 | alpha-1-antiproteinase | Proteinase inhibitor | 10.00 |
| G1SDN2 | keratin 17 | Keratin | 35.50 |
| G1SHY2 | keratin 75 - extracellular vesicular exosome | Keratin | 71.00 |
| G1SHZ4 | keratin type II cytoskeletal 7 | Keratin | 48.50 |
| G1SPP3 | keratin type II cytoskeletal 4 | Keratin | 59.50 |
| G1SUH1 | keratin type II cytoskeletal 72 isoform X1 | Keratin | 7.66 |
| G1T8T1 | keratin type I cytoskeletal 28 | Keratin | 13.25 |
| G1U758 | keratin type I cytoskeletal 18 | Keratin | 16.25 |
aProtein and spectral count of host and tick proteins identified in nymph Haemaphysalis longicornis saliva. Annotation and accession numbers of best match identities obtained using BLASTP against several protein databases can be checked with more details in Additional file 1: Table S1 and Additional file 2: Table S2
Table 2.
Tick and host proteins identified exclusively in adult saliva
| Contig numbera | Annotation | Class | Spec count |
|---|---|---|---|
| TICK | |||
| JAA54320 | glycine-rich protein | Glycine-rich protein | 1.50 |
| EEC14126 | radixin/ezrin/moesin | Cytoskeletal | 2.00 |
| ACX53929 | putative beta thymosin | Cytoskeletal | 1.33 |
| DAA34555 | microtubule-binding protein | Cytoskeletal | 1.00 |
| AEO32824 | actin depolymerizing factor | Cytoskeletal | 1.50 |
| AEO33976 | beta tubulin | Cytoskeletal | 6.50 |
| AFR32950 | paramyosin | Cytoskeletal | 5.33 |
| JAB76162 | dynein light chain | Cytoskeletal | 1.00 |
| JAB80373 | myosin class i heavy chain | Cytoskeletal | 12.25 |
| AAY42205 | troponin T | Cytoskeletal | 1.00 |
| Rm-1533 | beta tubulin partial | Cytoskeletal | 6.33 |
| Rm-80704 | myosin class ii heavy chain | Cytoskeletal | 4.67 |
| ACF35539 | glutathione S-transferase | Oxidant metabolism/detoxification | 6.50 |
| ACG76272 | aldehyde dehydrogenase | Oxidant metabolism/detoxification | 1.00 |
| AAQ74441 | glutathione S-transferase | Oxidant metabolism/detoxification | 1.50 |
| AEO33057 | sulfotransferase | Oxidant metabolism/detoxification | 2.00 |
| AEO35358 | catalase | Oxidant metabolism/detoxification | 8.00 |
| JAA63098 | putative aldehyde dehydrogenase | Oxidant metabolism/detoxification | 3.00 |
| Rm-14504 | aldehyde dehydrogenase | Oxidant metabolism/detoxification | 14.75 |
| Rm-46289 | aldehyde dehydrogenase | Oxidant metabolism/detoxification | 2.00 |
| ADN34303 | cuticular protein | Extracellular matrix/cell adhesion | 4.75 |
| AEO34631 | phosphoserine aminotransferase | Metabolism, amino acid | 1.50 |
| Rm-73466 | homocysteine S-methyltransferase | Metabolism, amino acid | 2.50 |
| AEO32930 | transketolase | Metabolism, carbohydrate | 2.33 |
| ACH88101 | glyceraldehyde-3-phosphate dehydrogenase | Metabolism, energy | 2.67 |
| AEO32901 | ATP synthase subunit beta | Metabolism, energy | 7.75 |
| AEO34579 | 3-phosphoglycerate dehydrogenase | Metabolism, energy | 3.75 |
| AEO35473 | Isocitrate dehydrogenase NADP | Metabolism, energy | 6.00 |
| JAA60091 | 3-phosphoglycerate dehydrogenase | Metabolism, energy | 3.33 |
| JAA60302 | Isocitrate dehydrogenase NADP | Metabolism, energy | 7.00 |
| JAA62224 | imp cyclohydrolase/methylglyoxal synthase | Metabolism, energy | 8.75 |
| JAA66712 | C-1-tetrahydrofolate synthase | Metabolism, energy | 2.00 |
| JAA68969 | glyceraldehyde-3-phosphate dehydrogenase | Metabolism, energy | 2.50 |
| JAB74970 | cytochrome B5 | Metabolism, energy | 1.33 |
| Rm-26196 | Malate dehydrogenase | Metabolism, energy | 2.67 |
| Rm-18758 | 15-hydroxyprostaglandin dehydrogenase | Metabolism, lipid | 1.50 |
| Rm-73635 | farnesoic acid o-methyltransferase | Metabolism, lipid | 1.67 |
| JAA59537 | 3’(2’)5’-bisphosphate nucleotidase | Metabolism, nucleotide | 1.00 |
| JAA60240 | GDP dissociation inhibitor | Metabolism, nucleotide | 1.50 |
| Inosine-5’-monophosphate dehydrogenase | Metabolism, nucleotide | 4.75 | |
| Rm-32846 | purine nucleoside phosphorylase transferase | Metabolism, nucleotide | 2.00 |
| Rm-6587 | SAICAR synthase | Metabolism, nucleotide | 2.00 |
| AEO35504 | annexin | Protein export machinery | 1.00 |
| BAF63673 | protein disulfide-isomerase | Protein modification machinery | 2.50 |
| EEC14106 | multifunctional chaperone | Protein modification machinery | 2.50 |
| JAA60036 | heat shock protein | Protein modification machinery | 1.50 |
| JAA67522 | heat shock protein hsp 90-alpha isoform | Protein modification machinery | 2.50 |
| JAB73342 | methyltransferase | Protein modification machinery | 1.50 |
| JAB75807 | cysteine s-methyltransferase | Protein modification machinery | 2.00 |
| Rm-24571 | heat shock protein 70 cognate | Protein modification machinery | 6.50 |
| Rm-62470 | heat shock-related protein | Protein modification machinery | 3.75 |
| JAA62377 | gmp synthase | Protein modification machinery | 2.50 |
| AEO32591 | elongation factor 1-alpha | Protein synthesis machinery | 8.33 |
| AEO32633 | AAA+ ATPase | Protein synthesis machinery | 1.00 |
| Rm-50382 | elongation factor 2-like | Protein synthesis machinery | 4.00 |
| AEO34773 | secreted protein | Secreted conserved protein | 3.00 |
| AEO32884 | lipocalin | Lipocalin | 7.25 |
| Rm-26539 | lipocalin | Lipocalin | 5.00 |
| BAF51711 | tick legumain | Proteinase | 8.50 |
| BAH86062 | cathepsin L-like | Proteinase | 2.33 |
| JAA60171 | metalloexopeptidase | Proteinase | 3.00 |
| JAA65860 | CNDP dipeptidase | Proteinase | 2.33 |
| EEC20000 | alpha-2-macroglobulin | Proteinase inhibitor | 6.00 |
| BAH02683 | haemalin | Proteinase inhibitor | 4.50 |
| AEO35673 | TIL | Proteinase inhibitor | 5.00 |
| BAG41813 | cyclophilin 1 | Signal transduction | 3.50 |
| AEO32842 | regulatory protein mlp | Signal transduction | 2.00 |
| JAB75372 | calmodulin | Signal transduction | 1.67 |
| Rm-69447 | neuropeptide-like protein 31 | Signal transduction | 1.00 |
| Rm-79606 | Regucalcin | Signal transduction | 2.50 |
| Rm-154439 | hemelipoprotein | Heme/iron metabolism | 1.67 |
| JAA62029 | zinc finger protein | Transcription machinery | 1.00 |
| EEC05877 | apolipophorin | Transporters/receptors | 2.75 |
| JAB78095 | glutamate receptor 1-like | Transporters/receptors | 1.67 |
| RABBIT | |||
| G1U9R8 | gelsolin | Cytoskeletal | 2.50 |
| P27170 | serum paraoxonase/arylesterase 1 | Oxidant metabolism/detoxification | 1.00 |
| G1SR65 | myeloperoxidase | Oxidant metabolism/detoxification | 1.50 |
| G1T049 | collagen alpha-6(IV) chain | Extracellular matrix/cell adhesion | 1.00 |
| G1TZA1 | type X collagen alpha 1-like | Extracellular matrix/cell adhesion | 2.25 |
| O77791 | protein S100-A12 | Immunity | 4.50 |
| P01840 | Ig kappa-b4 chain C region | Immunity | 10.25 |
| P01870 | Ig gamma-1 chain C region | Immunity | 48.50 |
| P04221 | Ig mu chain C region | Immunity | 4.00 |
| G1SS91 | complement C4-A | Immunity | 3.75 |
| G1T3X1 | complement component C9 | Immunity | 2.00 |
| G1TEC1 | Ig kappa chain V-I | Immunity | 4.75 |
| G1TFU1 | Ig kappa chain V-I | Immunity | 5.25 |
| G1TKP3 | immunoglobulin lambda-like | Immunity | 4.00 |
| G1TM51 | Ig kappa chain V-I | Immunity | 4.00 |
| G1TPF2 | T-cell surface glycoprotein CD8 -like | Immunity | 5.00 |
| G1TPZ1 | galectin-1 | Immunity | 2.33 |
| G1TRW8 | neutrophil gelatinase-associated lipocalin | Immunity | 8.50 |
| G1TUX5 | C1q and TNF related protein 7-like | Immunity | 2.50 |
| G1TVN7 | Ig kappa chain V-I | Immunity | 5.75 |
| G1TVZ5 | T-cell surface glycoprotein CD8 -like | Immunity | 1.50 |
| Q9GK63 | mammaglobin-B | Immunity | 2.00 |
| Q9GK67 | secretoglobin | Immunity | 3.67 |
| U3KM01 | Ig kappa chain V-I | Immunity | 4.00 |
| G1SQA8 | ATP synthase mitochondrial precursor | Metabolism, energy | 3.50 |
| G1TS29 | triosephosphate isomerase | Metabolism, energy | 1.33 |
| G1TYA7 | L-lactate dehydrogenase B chain | Metabolism, energy | 2.67 |
| G1TB24 | peroxysomal Fatty Acyl CoA Transporter | Metabolism, lipid | 1.50 |
| G1SN21 | purine nucleoside phosphorylase | Metabolism, nucleotide | 2.00 |
| G1TXI1 | nucleophosmin | Metabolism, nucleotide | 3.33 |
| G1U9T4 | nucleoside diphosphate kinase B | Metabolism, nucleotide | 3.33 |
| G1U155 | histone H2B type 1-like | Nuclear regulation | 3.00 |
| P51662 | annexin A1 | Protein export machinery | 5.50 |
| G1TDI4 | transthyretin | Protein export machinery | 3.00 |
| G1SJZ9 | thioredoxin | Protein modification machinery | 1.33 |
| G1SEK8 | metalloendopeptidase inhibitor | Proteinase inhibitor | 6.75 |
| G1SIK0 | antithrombin III | Proteinase inhibitor | 8.67 |
| G1SZA4 | inter-alpha-trypsin inhibitor heavy chain H1 | Proteinase inhibitor | 2.00 |
| G1TM88 | alpha-1-antitrypsin | Proteinase inhibitor | 2.33 |
| G1U6R8 | negative regulation of endopeptidase | Proteinase inhibitor | 8.25 |
| Q45GR2 | alpha-2-antiplasmin | Proteinase inhibitor | 3.50 |
| G1SNP6 | serine/threonine-protein kinase | Signal transduction | 1.50 |
| P09809 | apolipoprotein A-I | Heme/iron metabolism | 4.25 |
| G1TFW8 | lactotransferrin | Heme/iron metabolism | 5.75 |
| G1SU82 | vitamin D-binding protein | Transporters/receptors | 4.75 |
| G1T4K1 | sodium channel protein type 3 isoform X1 | Transporters/receptors | 2.00 |
| G1TP66 | keratin type II cuticular Hb3 | Keratin | 1.50 |
| P00919 | carbonic anhydrase 2 | Hemoglobin/RBC products | 3.50 |
aProtein and spectral count of host and tick proteins identified in adult Haemaphysalis longicornis saliva. Annotation and accession numbers of best match identities obtained using BLASTP against the several protein databases can be checked with more details in Additional file 1: Table S1 and Additional file 2: Table S2
Table 3.
Tick and host proteins identified in nymph and adult saliva
| Contig number | Annotation | Class | Spec count | |
|---|---|---|---|---|
| Nymph | Adult | |||
| TICK | ||||
| ABQ96858 | tropomyosin | Cytoskeletal | 3.00 | 5.25 |
| BAF98180 | actin | Cytoskeletal | 21.75 | 18.75 |
| AEO32669 | alpha tubulin | Cytoskeletal | 3.25 | 11.00 |
| AGC13075 | glutathione peroxidase | Oxidant metabolism/detoxification | 10.50 | 4.33 |
| AEO34612 | enolase | Metabolism, carbohydrate | 2.00 | 4.25 |
| Rm-10851 | ATP synthase subunit alpha | Metabolism, energy | 1.50 | 4.67 |
| AEO34838 | histone H4 | Nuclear regulation | 5.75 | 7.33 |
| AEO34879 | histone H2A | Nuclear regulation | 4.00 | 10.00 |
| AEO32095 | heat shock 70 kDa protein | Protein modification machinery | 5.50 | 5.67 |
| AEO32791 | heat shock protein | Protein modification machinery | 9.75 | 11.00 |
| AEO34048 | protein disulfide-isomerase A6-like | Protein modification machinery | 1.50 | 1.00 |
| JAA62581 | heat shock protein | Protein modification machinery | 3.00 | 3.50 |
| ADG86641 | lysosomal acid phosphatase | Protein modification machinery | 9.50 | 26.75 |
| JAA73257 | ubiquitin/40s ribosomal protein s27a | Proteasome machinery | 17.25 | 4.75 |
| ADK47399 | secreted protein | Secreted conserved protein | 2.67 | 3.00 |
| AGH08176 | AV422 | Secreted conserved protein | 4.75 | 4.25 |
| BAE53722 | aspartic protease | Proteinase | 4.00 | 3.25 |
| BAD11156| | serpin | Proteinase inhibitor | 15.50 | 3.50 |
| JAA60430 | alpha-2-macroglobulin | Proteinase inhibitor | 41.00 | 12.33 |
| JAA64973 | alpha-2-acroglobulin | Proteinase inhibitor | 17.50 | 3.50 |
| Rm-7619 | alpha-2-macroglobulin | Proteinase inhibitor | 26.25 | 9.00 |
| ABZ89554. | cystatin | Proteinase inhibitor | 6.33 | 2.67 |
| Rm-69112 | 14–3–3 zeta | Signal transduction | 4.33 | 2.33 |
| BAG12081 | hemelipoprotein | Heme/iron metabolism | 796.25 | 323.25 |
| BAJ21514 | hemelipoprotein | Heme/iron metabolism | 405.00 | 224.00 |
| BAJ21515 | hemelipoprotein | Heme/iron metabolism | 98.50 | 6.25 |
| BAL42280 | hemelipoprotein | Heme/iron metabolism | 141.75 | 60.75 |
| JAA59652 | hemelipoprotein | Heme/iron metabolism | 19.67 | 14.25 |
| ABD83654 | hemelipoprotein | Heme/iron metabolism | 12.50 | 12.25 |
| Rm-72548 | hemelipoprotein | Heme/iron metabolism | 32.25 | 14.75 |
| JAA61676 | plexins functional semaphorin receptor | Transporters/receptors | 12.50 | 2.50 |
| RABBIT | ||||
| G1T229 | filaggrin-2 | Cytoskeletal | 17.00 | 1.50 |
| G1T6W7 | catalase | Oxidant metabolism/detoxification | 5.25 | 7.50 |
| P16973 | lysozyme C | Immunity | 3.00 | 7.50 |
| P25230 | antimicrobial protein CAP18 | Immunity | 1.50 | 5.33 |
| P50117 | protein S100-A9 | Immunity | 3.75 | 3.00 |
| G1SUZ1 | complement C3 | Immunity | 5.25 | 15.00 |
| G1SYM4 | alpha-1B-glycoprotein | Immunity | 3.75 | 3.00 |
| G1THZ6 | Ig gamma-1 chain C region | Immunity | 15.00 | 62.00 |
| G1T0Z2 | histone H2A type 1-A | Nuclear regulation | 4.00 | 5.67 |
| G1T9M9 | heat shock cognate 71 kDa protein | Protein modification machinery | 9.00 | 10.00 |
| G1SQ70 | alpha-2-macroglobulin | Proteinase inhibitor | 10.75 | 32.50 |
| G1TFV7 | alpha-1-antiproteinase | Proteinase inhibitor | 10.25 | 37.25 |
| Q07298 | alpha-1-antiproteinase | Proteinase inhibitor | 10.00 | 32.67 |
| P19134 | serotransferrin | Heme/iron metabolism | 33.75 | 95.25 |
| P20058 | hemopexin | Heme/iron metabolism | 12.00 | 36.50 |
| G1SQ02 | peroxiredoxin-1 | Heme/iron metabolism | 3.00 | 3.25 |
| G1SWF6 | haptoglobin - hemoglobin binding | Heme/iron metabolism | 7.50 | 20.00 |
| G1TVS4 | hemopexin | Heme/iron metabolism | 12.00 | 38.25 |
| G1U9S2 | serum albumin | Heme/iron metabolism | 156.00 | 665.50 |
| G1SGQ5 | alpha-2-HS-glycoprotein | Transporters/receptors | 3.00 | 1.50 |
| G1SLY0 | HCO3- transporter family | Transporters/receptors | 1.50 | 3.00 |
| U3KMC6 | ceruloplasmin | Transporters/receptors | 3.67 | 8.50 |
| G1SKE3 | keratin type II cytoskeletal 6A | Keratin | 84.50 | 20.00 |
| G1SUH8 | keratin 2 | Keratin | 228.75 | 52.00 |
| G1SWB8 | keratin type I cytoskeletal 27 | Keratin | 23.67 | 14.00 |
| G1SY72 | keratin type II cytoskeletal | Keratin | 68.50 | 17.33 |
| G1T1V0 | keratin type I cytoskeletal 10 | Keratin | 108.50 | 58.50 |
| G1T1Y7 | keratin type I cytoskeletal 14 isoform X1 | Keratin | 56.00 | 12.50 |
| G1T4R6 | keratin type I cytoskeletal 16 | Keratin | 57.50 | 13.25 |
| G1T4S1 | keratin 15 | Keratin | 51.75 | 17.00 |
| G1TDN6 | keratin type II cytoskeletal 5 | Keratin | 99.75 | 14.75 |
| G1U754 | histidine-rich glycoprotein | Heme/iron metabolism | 11.75 | 25.25 |
| G1U9I8 | keratin type II cytoskeletal 1 | Keratin | 104.25 | 39.75 |
| G1T0W8 | fibrinogen beta chain | Fibrinogen | 5.25 | 26.50 |
| G1T0X2 | fibrinogen alpha chain | Fibrinogen | 9.00 | 13.50 |
| G1TKX3 | fibrinogen gamma chain | Fibrinogen | 6.25 | 34.75 |
| P01948 | hemoglobin subunit alpha-1/2 | Hemoglobin/RBC products | 87.00 | 544.00 |
| P02057 | hemoglobin subunit beta | Hemoglobin/RBC products | 100.75 | 517.50 |
| P07452 | carbonic anhydrase 1 | Hemoglobin/RBC products | 5.00 | 11.00 |
aProtein and spectral count of host and tick proteins identified both in nymph and adult Haemaphysalis longicornis saliva. Annotation and accession numbers of best match identities obtained using BLASTP against several protein databases can be checked with more details in Additional file 1: Table S1 and Additional file 2: Table S2
Fig. 3.
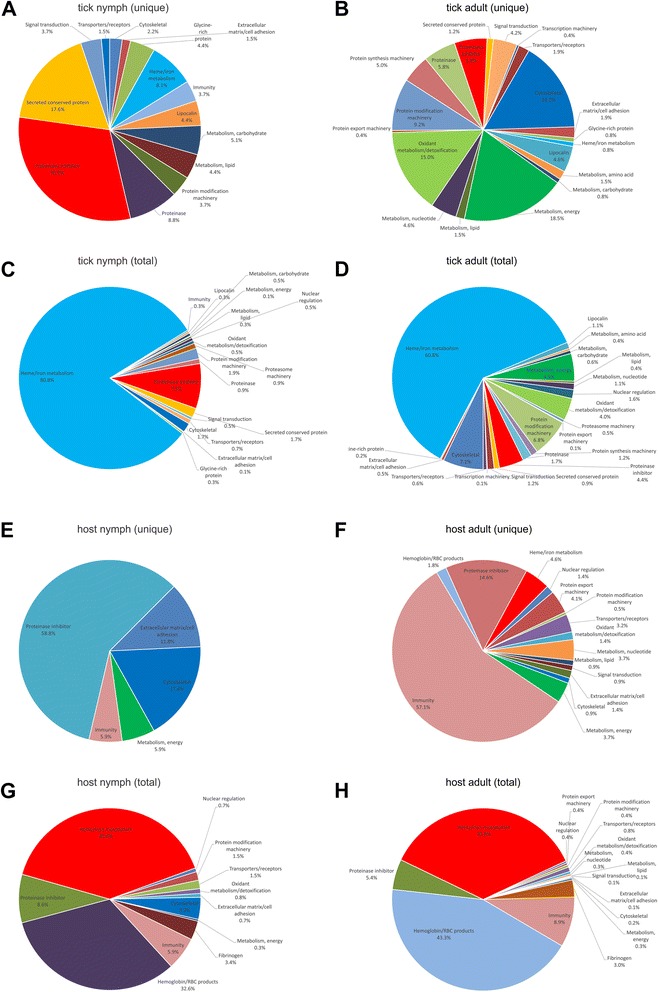
Functional classification of Haemaphysalis longicornis salivary proteins identified in fully engorged nymph (nymph) and fully engorged adult females (adult). a Tick proteins detected in nymph (a and c) and adult (b and d). Proteins were divided as detected both in nymph (c) and adult (d) and proteins detected exclusively in nymph (a) or adult (b) and classified in groups according to their function and/or protein family. Pie charts represent the percentage of proteins found in each group with respect to normalized spectral counting for each sample. b Host proteins detected in nymph (e and g) and adult (f and h). Proteins were divided as detected both in nymph (g) and adult (h) and proteins detected exclusively in nymph (e) or adult (f) and classified in groups according to their function and/or protein family. Pie charts represent the percentage of proteins found in each group with respect to normalized spectral counting for each sample. Keratin was removed before data interpretation. (Additional file 3: Figure S1 shows data with keratin)
Fig. 4.
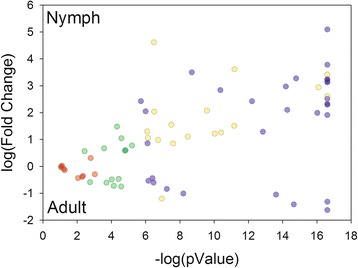
Volcano plot abundance changes analysis of Haemaphysalis longicornis saliva proteins identified both in fully engorged nymph (nymph) and fully engorged adult females (adult). Each point represents the difference in expression (log2 fold difference) between nymph and adult plotted against the level of statistical significance. Proteins represented by (blue dot) had an identification that satisfied both fold and statistical criteria; (yellow dot) had an identifications that was filtered out by the L-stringency; (green dot) had an identification satisfied the fold criteria but, most likely, this happened by chance; and (red dot) had identification did not meet the fold and p-value criteria
Table 4.
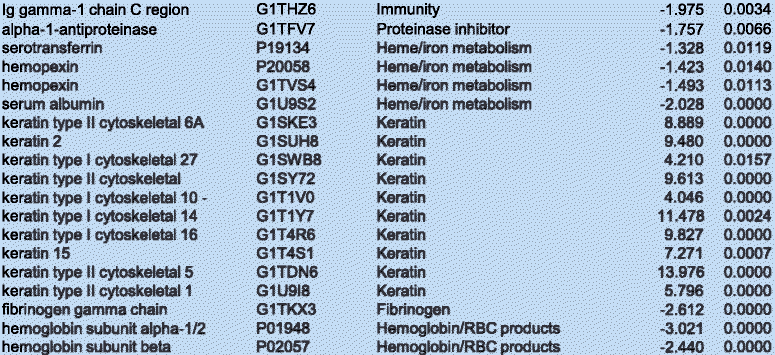
Differential abundance of nymph and adult saliva proteins (determined using the TFold)
| Annotation | Contig numbera | Class | Fold changeb | p value |
|---|---|---|---|---|
| TICK | ||||

| ||||
| RABBIT | ||||
| ||||
| TICK | ||||
| ||||
| RABBIT | ||||

| ||||
| TICK | ||||
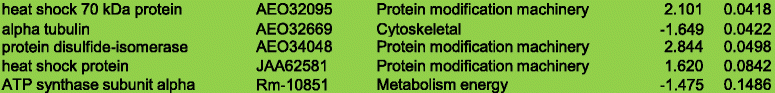
| ||||
| RABBIT | ||||

| ||||
| TICK | ||||

| ||||
| RABBIT | ||||

| ||||
Blue: identifications that satisfied both fold and statistical criteria
Yellow: identifications were filtered out by the L‐stringency
Green: identifications satisfied the fold criteria but, most likely, this happened by chance
Red: identifications did not meet the fold and p‐value criteria
aAccession numbers of best matches identities obtained using BLASTP against the non‐redundant protein database in GenBank
bPositive number means the fold increased expression in nymph (relation between nymph and adult). Negative number means the fold increased expression in adults (relation between adult and nymph)
The identified tick proteins include (i) heme/iron metabolism-related proteins, including hemelipoproteins and ferritin; (ii) proteins related to oxidation/detoxification functions, including glutathione S-transferase, aldehyde dehydrogenase, glutathione peroxidase, and catalase; (iii) enzymes such as serine proteinases, cysteine proteinases, metalloexopeptidases, carboxypeptidases, and aspartic proteases; (iv) proteinase inhibitors of the serpin, cystatin, alpha-2-macroglobulin, trypsin inhibitor-like (TIL), and Kunitz families; (v) tick-specific protein families including lipocalin (histamine-binding proteins), glycine-rich proteins, and the group of secreted conserved proteins [9]. In addition, proteins related to cytoskeletal functions, protein modification machinery, signal transduction, transporters and receptors, metabolism of carbohydrate, energy, nucleotide, amino acids, and lipid were identified (Fig. 3 and Tables 1, 2 and 3).
Heme/iron metabolism-related proteins
The most abundant proteins identified in H. longicornis saliva are hemelipoproteins, which are associated with heme/iron metabolic processes. This finding is in agreement with previous studies describing hemelipoproteins in proteomic saliva of Ornithodoros moubata, Rhipicephalus microplus, Ixodes scapularis [13, 16, 30, 31], as well as with a study about A. americanum immunoproteome [32], which showed that hemelipoproteins are the major proteins in tick saliva.
Hemelipoproteins were first described as heme-binding proteins from tick hemolymph, being able to transport cholesterol, phospholipids, and free fatty acids, in addition to heme [62, 63]. These proteins are the most abundant proteins in spontaneously detached fully engorged H. longicornis saliva, which were relatively highly expressed in nymph saliva (Fig. 3 and Table 4). This data is in accordance with the findings observed for R. microplus, where hemelipoproteins were highly expressed in partially engorged adult ticks, showing a higher expression in the early developmental stages of tick feeding [16]. The physiological role of hemelipoproteins in blood meal acquisition is not completely understood. Tissue and vessel dilaceration produced by style of tick feeding and the presence of hemoglobin digestive enzymes (such as cathepsin and legumain) in tick saliva cause hemolysis and heme release in the feeding site. It is known that heme activates innate immune cells such as macrophages and neutrophils through activation of innate immune receptors [64–66], thus a role of hemelipoproteins being injected into the feeding site to prevent heme-induced inflammation is suggested. In addition, these proteins could be involved in a heme-excretory system, removing heme excess from tick and re-injecting it into the host. However, the presence of heme in tick saliva needs further investigation.
Proteinases
Several proteinases classes were identified in H. longicornis saliva: (i) serine proteinases; (ii) cysteine proteinases (including longipain, legumain and cathepsin L); (iii) aspartic proteinases; (iv) serine carboxypeptidases; and (v) proteins that belong to peptidase family M20 from MEROPS (including dipeptidases, and metalloexopeptidases). This set of different proteins present in tick saliva could have multiple modes of action during blood feeding. Serine proteinases may interfere with host inflammation and blood clotting. As shown by the presence of I. scapularis saliva protein C activator, a protein acting in the production of activated protein C, a potent anticoagulant that also regulates a myriad of inflammatory responses through protease activated receptors activation [67].
Cysteine proteinases, aspartic proteinases, serine carboxypeptidases and legumains have been described mainly as digestive enzymes with a role in hemoglobin digestion [68, 69] and pathogen transmission in ticks [70]. Thus, the presence of these enzymes in the feeding site may indicate that they act as digestive enzymes secreted into the host, digesting blood components at the tick attachment site and facilitating pathogen transmission during tick feeding. The presence of putative digestive enzymes in tick saliva was also observed in R. microplus [16].
In general, dipeptidases hydrolyze the late products of protein degradation to complete the conversion of proteins to free amino acids. In ticks, dipeptidases were reported to be responsible for the destruction of bradykinin, a potent pain inductor [71]. These enzymes are related to the kininase activity found in I. scapularis saliva, which may be responsible for the lack of host pain response subsequent to attachment and feeding [72].
Proteinase inhibitors
Host defense responses triggered by tick feeding are mainly dependent on the action of several proteinases, such as procoagulant (thrombin, factor Xa and other coagulation factors), pro-inflammatory (neutrophil elastase, proteinase-3, chymase, tryptase, kallikrein, cathepsin L, cathepsin B, cathespin S, cathepsin C, and cathepsin G) and complement enzymes (factors B, C, D, and component 2) [9–12]. Proteinases from these pathways are regulated by specific endogenous inhibitors, maintaining homeostasis. From this perspective, proteinase inhibitors secreted into the feeding site act by disrupting host defenses, facilitating blood meal acquisition. Several families of protease inhibitor domains were found in saliva of H. longicornis (Tables 1, 2 and 3).
Serpins are a superfamily of serine proteinase inhibitors involved in blood coagulation, fibrinolysis, inflammation, and complement activation in mammals [73, 74]. These proteins were found mostly in nymph saliva (Table 1). Tick serpins are secreted into the feeding site to disrupt host defenses against tick feeding, including anticoagulant [75–77] and immunomodulatory responses [78–80]. The potential effect of these proteins on host systems is supported by several studies that demonstrate the mortality and reduced feeding efficiency when several tick species were fed on host immunized with tick recombinant serpins [55–59]. Additionally, serpins in Aedes aegypti and Anopheles stephensi have been shown to play a role in pathogen transmission [81, 82].
Cystatins form a large superfamily of reversible and tight-binding inhibitors that interact with papain-like cysteine proteases and legumains [83]. Tick salivary cystatins have been described as immunosuppressive and anti-inflammatory proteins [84–87]. Moreover, the importance of cystatins in tick physiology was observed in studies that showed that neutralization of cystatin reduces tick feeding ability [52, 85, 88, 89].
Haemalin is a member of Kunitz-type inhibitors identified in H. longicornis saliva. This protein has been described as a thrombin inhibitor, delaying bovine plasma clotting time and inhibiting both thrombin-induced fibrinogen clotting and thrombin-induced platelet aggregation. This protein was described as a midgut protein [29], and this is the first time it is described in H. longicornis saliva. Taking into account haemalin function, we suggested that this protein acts as an anticoagulant salivary protein during tick feeding.
TIL (trypsin inhibitor-like) proteins have been reported in several tick sialomes [9, 10] and are described as elastase inhibitors, which also have antimicrobial activity [90, 91]. Alpha-2-macroglobulin are a group of proteins that have been found to inhibit several serum proteinases in vertebrates, including thrombin, factor Xa and kallikreins [77, 92–94], mediating T-cell proliferation and activating macrophages [95, 96]. Thus, as these proteins are secreted both in nymph and in adult tick saliva, they could act as anticoagulant and/or immunomodulatory proteins during blood feeding.
Tick-specific protein families
Advances in transcriptomic and proteomic studies of tick salivary gland have created new opportunities to identify the variety of tick salivary transcripts and proteins. Many proteins are described to have no similarities to non-tick proteins from the NCBI database [9]. The secreted conserved protein group is composed mainly of tick proteins containing a signal peptide predicted and with similarities to proteins identified in other ticks [9]. Most of proteins included in this group have unknown functions and were described only in gene or protein sequencing projects, having their expression up-regulated after blood acquisition [97, 98]. A functionally described member of tick secreted conserved protein group is A. americanum AV422 protein. This protein was first described as an up-regulated protein in response to tick host exposure and/or to feeding stimuli in rabbits [99]. AV422 is secreted into the host during tick feeding, acting as an anticoagulant and anti-complement protein [100, 101]. In H. longicornis, this protein is identified in both the spontaneously detached fully engorged nymph and adult saliva (Table 3). Additionally, an AV422-like protein was identified in partially and fully engorged R. microplus female saliva proteome [16]. Based on the high amino acid conservation and on its presence in other tick proteomes, it will be interesting to determine whether or not H. longicornis AV422-like protein is functionally similar to that of A. americanum.
The antigen 5 protein family is a group of cysteine-rich secreted proteins [102]. This group is described in the salivary glands of blood-sucking insects and ticks, with functions that remain mostly unknown [10, 103, 104], being identified exclusively in nymph saliva (Table 1). Glycine-rich proteins are extracellular matrix proteins and/or structural proteins with an important role in attachment to the host, since it they are present in cement material secreted by salivary glands during feeding process [105, 106]. The expression levels of these proteins are associated with size of mouthparts and the number of hosts used by tick during its life cycle. Ticks with short mouthparts and one-host ticks express more glycine-rich protein. It could be implied that one-host ticks are more consistently pressed to sustain attachment on the host’s skin [107]. In this sense, glycine-rich proteins were identified mostly in nymph saliva (Table 1). These proteins are also involved in the defense against pathogens, since they are inducible antibacterial proteins predominantly active by Gram-negative bacteria presents in insect and tick hemolymph and salivary gland [108].
Tick proteins of the lipocalin family are classified based on their homology with tick proteins containing the characteristic tick histamine-binding domain (PF02098) [109, 110]. They are a group of multifunctional secreted proteins that bind several types of small hydrophobic ligands. The role as transport proteins is well studied, however it is clear that the arthropod lipocalin family is involved in various other physiological functions such as cell growth and metabolism, regulation of the immune response, and tissue repair [111]. Lipocalins were detected in several tick saliva proteomes [13, 16], and H. longicornis saliva contains at least three proteins described as lipocalin. The presence of lipocalins in tick saliva is related to control of inflammatory processes and interference with host hemostatic functions [112–114]. Furthermore, the importance of lipocalins in nymph and adult saliva has been demonstrated, since the expression of lipocalins in insects and ticks is up-regulated in response to injury [115], as well as to viral [116] or bacterial infections [117], enhancing tick immune responses and resilience to infection.
Detoxification/oxidation
In nymph and adult saliva, various proteins involved in processes of detoxification and redox buffering were detected, including glutathione S-transferase, aldehyde dehydrogenase, sulfotransferase, and catalase. These proteins could detoxify oxidants generated during blood meal acquisition and/or host oxidants associated with inflammation. The antioxidant expression levels differ throughout arthropod development [118]. In H. longicornis saliva, the abundance of antioxidant proteins is higher in adults, when compared to nymphs (Tables 1, 2 and 3). A glutathione peroxidase was the only protein of this class identified in nymph saliva. This protein has been well characterized for its antioxidant and anti-inflammatory activity in mammals [119], and could be related as an immunomodulatory protein from H. longicornis saliva. Sulfotransferase may inactivate dopamine, the secretagogue found in the salivary gland of ticks [120]. Glutathione S-transferase expression has been associated with resistance to acaricides and insecticides in many species [121, 122]. In addition, it has been proposed that GST secreted by parasites has immunomodulatory activity due to the alteration of the cytokine gene expression profile, modulation of immune cell proliferation, and decrease in oxidative ability of phagocytes [123, 124]. The roles of GST and other proteins in the detoxification of endogenous toxin preventing and repairing the damage of ROS generated by hemoglobin degradation has been described [125], and the induction of the expression of these proteins in response to oxidative stress have been observed, supporting the antioxidant physiological role [126, 127]. Furthermore, immunization experiments showed the potential use of tick GST to protect hosts against tick infestation [50, 51].
Cytoskeletal
Proteins in H. longicornis saliva associated with cytoskeletal and structural cellular function were identified, including actin, tubulin, paramyosin, among others, which are fundamental to intracellular transport and cellular division. Notably, these proteins were abundantly expressed in adult saliva, when compared to nymph saliva, suggesting that the presence of such proteins has a physiological explanation, as opposed to tick/host tissue contamination during saliva collection (Table 4). Since these are intracellular proteins, and considering that most lack signal peptides, it may be hypothesized that they are released as consequence of damage, degeneration, or apoptosis of salivary gland acines [128, 129]. Furthermore, the presence of apocrine secretion in tick salivary gland is described [130]. Another structural protein has been identified in R. microplus saliva is paramyosin. This protein is secreted in saliva and recognized during the tick infestation, further suggesting that it may possess additional, non-muscle functions in the tick-host relationship [131]. Troponin I-like molecule was detected in H. longicornis saliva, with angiogenesis inhibitor activity impairing host tissue repair and helping the tick feeding process [132]. These observations suggested that these proteins could have roles other than structural functions.
Metabolism
A wide variety of enzymes and proteins related to carbohydrate, lipid and amino acid metabolism and to the energetic pathways was observed in H. longicornis saliva. This finding is in accordance with other tick saliva proteomes. Similarly to cytoskeletal proteins, proteins that belong to this class are predominant in adult tick saliva (Table 1, 2 and 3). Although functional activity of these proteins remains unknown concerning tick feeding, the activity of some proteins related to metabolism has been characterized.
A salivary enolase from O. moubata was described acting as a plasminogen receptor, and may play a role stimulating host fibrinolysis and maintaining blood fluidity during tick feeding [133]. Similar activity was described for enolases from other parasites [134, 135].
Another group of metabolism-related proteins with a possible role in tick feeding is formed by chitinases. Chitinases are either active or inactive, based on the functional domain. Active chitinases are mostly described as responsible for the hydrolytic cleavage of the β-glycosidic linkages between GlcNAc residues of chitin, involved in molting and growth of arthropods [136]. On the other hand, inactive chitinases were suggested to be involved in the maintenance of a stable feeding site and in the activity of a potential immunoglobulin G binding protein in A. americanum [137]. It is interesting to note that we show the presence of a putative inactive chitinase only in nymphs (Table 1). However, previous studies showed the presence of an inactive chitinase in adults, as a secreted saliva protein [16, 138].
Enzymes of the o phospholipase A2 family play important roles in phospholipid digestion, rearrangement of cellular membrane phospholipid structures, inflammatory responses, defense and predation mechanisms, and signal transduction [139]. These enzymatic activities have been identified in tick saliva, and are speculated to stimulate tick prostaglandin E2 production [140]. Tick saliva proteomic studies have identified these proteins [16], pointing to their presence in tick saliva. A phospholipase A2 was identified exclusively in nymph saliva (Table 1). These proteins could provide the anti-inflammatory, anti-hemostatic, and vasodilator activity required for long-term blood feeding.
It should be noted that these metabolism-related proteins have been identified in tick saliva, and may be classified as moonlighting proteins, so they can have other distinct functions [141, 142]. Several studies on moonlighting proteins are being carried out, and the discovery of new functions could afford deeper insights into metabolism-related proteins in tick feeding physiology.
Differential expression between nymph and adult female saliva proteins
Ixodid ticks begin attachment by cutting into the host skin, followed by secretion of cement, a process that may take from one to two days. When completely affixed to the wound site, these ticks feed slowly from the pooled blood formed, for several days [143–145]. During feeding, the salivary gland of adult ixodid ticks undergoes remarkable growth and differentiation, which is accompanied by significant increases in the rate of proteins synthesis [46, 146]. It has been proposed that different tick feeding conditions might affect salivary gland transcription of hard ticks. This feature is related to different vertebrate host exposure and distinct developmental stages, leading to changes in salivary transcription dynamics, as shown previously [14, 16, 32, 99, 147–150].
As shown in other tick saliva proteomes [14, 16], the protein content of tick saliva in different developmental stages varies, such as in this case from nymph to adult female ticks in H. longicornis. Of the 135 proteins detected in H. longicornis saliva, 30 proteins were identified in nymph saliva, 74 proteins were identified in fully engorged adult females, and 31 were detected in both stages (Fig. 2, Tables 1, 2 and 3). Nymph-specific proteins are represented mostly by proteinase inhibitors and a secreted conserved proteins group (Fig. 3 and Table 1), while adult specific proteins are represented primarily by proteins related to energy metabolism, oxidant/detoxification metabolism, and cytoskeleton (Fig. 3 and Table 2). This finding in adult saliva may be related to salivary gland degeneration starting after adult ticks detach from the host [128, 151].
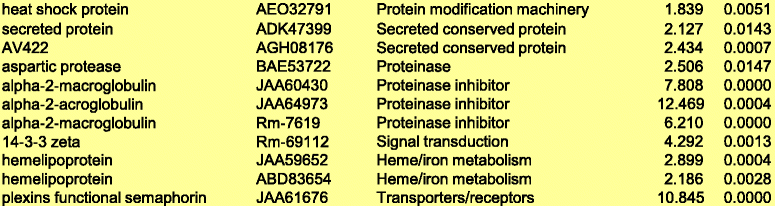
Changes in expression levels were observed for 11 of the 31 identified tick proteins detected in both stages. The changes in the protein secretion were determined by pairwise comparison between nymphs and fully engorged H. longicornis female tick saliva using the TFold module from PatternLab for Proteomics platform [37], and were shown to be statistically significant (Fig. 4 and Table 4). The range in fold change was shown to be greater in the specific-tick proteins than in host protein secreted in nymph saliva. The most significantly affected nymph up-regulated proteins were a hemelipoprotein (up-regulated 34.69-fold), a serpin (up-regulated 8.99-fold), a ribosomal protein s27a (up-regulated 7.98-fold), a cystatin (up-regulated 5.47-fold), and a glutathione peroxidase (up-regulated 5.46-fold). The physiological meaning of these differences is not clear. However, as discussed previously, hemelipoproteins are known by their heme-binding function [63], and expression in early developmental stages of tick feeding observed here is in accordance with that found in R. microplus [16]. Proteinase inhibitors were described as early stage secreted saliva proteins in tick, with anticoagulant and immunomodulatory properties, modulating host defense systems trigged against tick feeding [14, 16, 75, 76, 78, 87]. This observation can be related to the nature of the blood feeding process, throughout the different stages of tick as they have to use unique proteins to counteract host defenses, especially for fully engorged nymphs, which need to molt into adults and prepare to feed on another host.
Host proteins
As shown previously for other tick species [13, 15, 16, 30, 31], a large number of host proteins were identified both in nymph and adult H. longicornis saliva (Table 1, 2 and 3). It was demonstrated that ticks transport intact proteins across the digestive system to the hemolymph. After blood ingestion, host blood proteins such as albumin and immunoglobulin cross the midgut epithelium of ticks, and are detected in tick tissues, including secretion into saliva [15, 152, 153]. This finding suggests that the presence of host proteins in tick saliva may be a real and common recycling system present in ticks, not a result of contamination during saliva collection.
In H. longicornis saliva, 100 proteins matched the rabbit database. Host identified proteins in tick saliva included proteins related to (i) immunity, such as immunoglobulins, complement system proteins, and antimicrobial proteins; (ii) heme/iron metabolism-related proteins, like serum albumin, peroxiredoxin, serotransferrin, apolipoprotein, and hemopexin; (iii) proteinase inhibitors of the serpin and alpha-2-macroglobulin superfamilies; and (iv) hemoglobin/red blood cells-related products. A set of 13 rabbit proteins was found only in nymph saliva samples, mostly rabbit keratin (Table 4, Fig. 3, and Additional file 3: Figure S1), suggesting that host keratin from tick mouthparts reached saliva during collection, since cleaning of host tissue in nymphs’ mouthparts is more critical than in adults, due to its smaller size. Forty-eight rabbit proteins were only found in this adult saliva, and 30 rabbit proteins were present in both samples (Fig. 2).
The presence of different classes of host proteins in the saliva of the two tick developmental stages suggests the existence of a selective uptake process for host proteins (Fig. 3 and Additional file 3: Figure S1, Tables 1, 2, and 3) as observed in other studies [15, 16]. Furthermore, the relationship between concentrations of these proteins in saliva is different from that observed in host blood. This data is similar to findings observed in other tick species [30, 31, 153, 154]. An explanation for the presence of host proteins in tick saliva is that ticks recycle pivotal host proteins in order to subvert their role in the host and/or using host proteins in specific tick physiologic systems. Utilization of host hemoglobin as substrate to generate antimicrobial peptides against microorganisms was observed in R. microplus [155] and O. moubata [156]. As observed in R. microplus, here we found the same profile for proteins related to heme/iron metabolism. While the major tick heme-binding protein is secreted into saliva from nymphs, its expression decreases in adults. Reduction of heme-binding proteins in adults was accompanied by an increase in the host heme-binding proteins serum albumin, hemopexin, apolipoprotein, and peroxiredoxin (Fig. 3, Table 1, 2 and 3). These observations could suggest that the tick replaces hemelipoproteins by host derived heme-binding proteins, since hemelipoproteins are used for vitellogenesis at the end of the feeding process [62]. The host-derived transferrin was described in the hemolymph of D. variabilis and in whole nymphal ticks of A. americanum [157, 158]. Recently, a study showed the movement of host-derived transferrin H. longicornis, particularly from the midgut to the ovary, via hemolymph [159]. In the same way, R. microplus re-use heme from blood meal to synthetize heme proteins during protein synthesis [160].
Mammalian serpins are described endogenous regulators of host defenses against tick feeding [161, 162]. Host proteins of the serpin superfamily were identified in saliva, including alpha-1-antiprotease, antithrombin III, and alpha-2-antiplasmin. These proteins regulate enzymes such as neutrophil elastase, thrombin, and plasmin. It is important to find out whether these host proteins have the potential to inhibit their own serine proteinases. The presence of immunoglobulin chains could be explained as a tick self-defense system, since antibodies remain in an active form in tick hemolymph [152].
These observations suggest that the use of host proteins in tick physiology is not an unusual occurrence, and that these proteins may have an important physiology role in tick feeding process.
Conclusions
H. longicornis tick saliva has not been previously studied due to the considerable difficulty to collect saliva. Previously, researchers used salivary gland extract instead of saliva for protein analysis. This study describes the first proteome analysis of saliva of nymph and adult H. longicornis. Despite the use of saliva of fully engorged ticks, we could identify several tick proteins that can provide useful information for basic and applied aspects of the host-parasite interaction. The role of saliva proteins in the contact between a tick and the host is crucial during feeding process, and the knowledge about salivary components may improve the understanding of tick physiology, aiding the identification of a new target for tick control.
Acknowledgments
This project was supported by FAPERJ, INCT-Entomologia Molecular, CNPq and CAPES from Brazil; National Center for Research Resources (5P41RR011823), National Institute of General Medical Sciences (8P41GM103533) from USA, and research fund of Chonbuk National University in 2015 from Korea. Authors would like to thank Dr. José M. C. Ribeiro for providing the VB programs used in protein annotation.
Additional files
Tick proteins in H. longicornis saliva identified by mass spectrometry. Name, classification based on function and/or protein family and annotation and accession numbers of best match identities obtained using BLASTP against the several protein databases are provided in the accompanying spreadsheet (Hyperlinked Excel spreadsheet, zipped).
Host proteins in H. longicornis saliva identified by mass spectrometry. Name, classification based in function and/or protein family and annotation and accession numbers of best match identities obtained using BLASTP against the several protein databases are provided in the accompanying spreadsheet (Hyperlinked Excel spreadsheet, zipped).
Functional classification of Haemaphysalis longicornis salivary proteins identified in fully engorged nymph (nymph) and fully engorged adult females (adult). Host proteins detected in nymph (A and C) and adult (B and D). Proteins divided as detected both in nymph (C) and adult (D) or detected exclusively in nymph (A) or adult (B) and classified in groups according to their function and/or protein family. Pie charts represent the percentage of proteins found in each group with respect to normalized spectral counting for each sample.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
LT, MSI, MY, and ISV designed the study. LT, MSI, JKD, and AFMP carried out the experiments. LT, MSI, TK, JKD, JRY, AFMP, AM, MY, and ISV analyzed the data. LT, MSI, TK, AFMP, AM, MY, and ISV drafted the manuscript. All authors read and approved the final manuscript.
Authors’ information
Lucas Tirloni and Mohammad Saiful Islam are co-first authors.
Myung-Jo You and Itabajara da Silva Vaz Jr are co-senior authors.
Contributor Information
Lucas Tirloni, Email: ltirloni@gmail.com.
Mohammad Saiful Islam, Email: saifulhstu@gmail.com.
Tae Kwon Kim, Email: taekim009@gmail.com.
Jolene K. Diedrich, Email: jdiedric@scripps.edu
John R. Yates, III, Email: jyates@scripps.edu.
Antônio F. M. Pinto, Email: pinto.afm@gmail.com
Albert Mulenga, Email: AMulenga@cvm.tamu.edu.
Myung-Jo You, Email: tick@jbnu.ac.kr.
Itabajara Da Silva Vaz, Jr., Phone: +55 51 3308 6078, Email: itabajara.vaz@ufrgs.br
References
- 1.Sun J, Liu Q, Lu L, Ding G, Guo J, Fu G, et al. Coinfection with four genera of bacteria (Borrelia, Bartonella, Anaplasma, and Ehrlichia) in Haemaphysalis longicornis and Ixodes sinensis ticks from China. Vector Borne Zoonotic Dis. 2008;8:791–5. doi: 10.1089/vbz.2008.0005. [DOI] [PubMed] [Google Scholar]
- 2.Lee MJ, Chae JS. Molecular detection of Ehrlichia chaffeensis and Anaplasma bovis in the salivary glands from Haemaphysalis longicornis ticks. Vector Borne Zoonotic Dis. 2010;10:411–3. doi: 10.1089/vbz.2008.0215. [DOI] [PubMed] [Google Scholar]
- 3.Zou Y, Wang Q, Fu Z, Liu P, Jin H, Yang H, et al. Detection of spotted fever group Rickettsia in Haemaphysalis longicornis from Hebei Province, China. J Parasitol. 2011;97:960–2. doi: 10.1645/GE-2751.1. [DOI] [PubMed] [Google Scholar]
- 4.Lee JH, Park HS, Jang WJ, Koh SE, Park TK, Kang SS, et al. Identification of the Coxiella sp. detected from Haemaphysalis longicornis ticks in Korea. Microbiol Immunol. 2004;48:125–30. doi: 10.1111/j.1348-0421.2004.tb03498.x. [DOI] [PubMed] [Google Scholar]
- 5.Lee JH, Park HS, Jung KD, Jang WJ, Koh SE, Kang SS, et al. Identification of the spotted fever group rickettsiae detected from Haemaphysalis longicornis in Korea. Microbiol Immunol. 2003;47:301–4. doi: 10.1111/j.1348-0421.2003.tb03399.x. [DOI] [PubMed] [Google Scholar]
- 6.Guan G, Moreau E, Liu J, Hao X, Ma M, Luo J, et al. Babesia sp. BQ1 (Lintan): molecular evidence of experimental transmission to sheep by Haemaphysalis qinghaiensis and Haemaphysalis longicornis. Parasitol Int. 2010;59:265–7. doi: 10.1016/j.parint.2009.12.002. [DOI] [PubMed] [Google Scholar]
- 7.Li Y, Luo J, Guan G, Ma M, Liu A, Liu J, et al. Experimental transmission of Theileria uilenbergi infective for small ruminants by Haemaphysalis longicornis and Haemaphysalis qinghaiensis. Parasitol Res. 2009;104:1227–31. doi: 10.1007/s00436-009-1347-7. [DOI] [PubMed] [Google Scholar]
- 8.Hoogstraal H, Roberts FH, Kohls GM, Tipton VJ. Review of Haemaphysalis (kaiseriana) longicornis Neumann (resurrected) of Australia, New Zealand, New Caledonia, Fiji, Japan, Korea, and Northeastern China and USSR, and its parthenogenetic and bisexual populations (Ixodoidea, Ixodidae) J Parasitol. 1968;54:1197–213. [PubMed] [Google Scholar]
- 9.Francischetti IM, Sa-Nunes A, Mans BJ, Santos IM, Ribeiro JM. The role of saliva in tick feeding. Front Biosci (Landmark Ed) 2009;14:2051–88. doi: 10.2741/3363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mans BJ. Evolution of vertebrate hemostatic and inflammatory control mechanisms in blood-feeding arthropods. J Innate Immun. 2011;3:41–51. doi: 10.1159/000321599. [DOI] [PubMed] [Google Scholar]
- 11.Ribeiro JM. Blood-feeding arthropods: live syringes or invertebrate pharmacologists? Infect Agents Dis. 1995;4:143–52. [PubMed] [Google Scholar]
- 12.Ribeiro JM, Francischetti IM. Role of arthropod saliva in blood feeding: sialome and post-sialome perspectives. Annu Rev Entomol. 2003;48:73–88. doi: 10.1146/annurev.ento.48.060402.102812. [DOI] [PubMed] [Google Scholar]
- 13.Diaz-Martin V, Manzano-Roman R, Valero L, Oleaga A, Encinas-Grandes A, Perez-Sanchez R. An insight into the proteome of the saliva of the argasid tick Ornithodoros moubata reveals important differences in saliva protein composition between the sexes. J Proteomics. 2013;80C:216–35. doi: 10.1016/j.jprot.2013.01.015. [DOI] [PubMed] [Google Scholar]
- 14.Mudenda L, Pierle SA, Turse JE, Scoles GA, Purvine SO, Nicora CD, et al. Proteomics informed by transcriptomics identifies novel secreted proteins in Dermacentor andersoni saliva. Int J Parasitol. 2014;44:1029–37. doi: 10.1016/j.ijpara.2014.07.003. [DOI] [PubMed] [Google Scholar]
- 15.Oliveira CJ, Anatriello E, de Miranda-Santos IK, Francischetti IM, Sa-Nunes A, Ferreira BR, et al. Proteome of Rhipicephalus sanguineus tick saliva induced by the secretagogues pilocarpine and dopamine. Ticks Tick Borne Dis. 2013;4(6):469–77. doi: 10.1016/j.ttbdis.2013.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tirloni L, Reck J, Terra RM, Martins JR, Mulenga A, Sherman NE, et al. Proteomic analysis of cattle tick Rhipicephalus (Boophilus) microplus saliva: a comparison between partially and fully engorged females. PLoS One. 2014;9:e94831. doi: 10.1371/journal.pone.0094831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Carvalho-Costa T, Mendes M, Da SM, Da CT, Tiburcio M, Anhe A, et al. Immunosuppressive effects of Amblyomma cajennense tick saliva on murine bone marrow-derived dendritic cells. Parasit Vectors. 2015;8:22. doi: 10.1186/s13071-015-0634-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Maritz-Olivier C, Stutzer C, Jongejan F, Neitz AW, Gaspar AR. Tick anti-hemostatics: targets for future vaccines and therapeutics. Trends Parasitol. 2007;23:397–407. doi: 10.1016/j.pt.2007.07.005. [DOI] [PubMed] [Google Scholar]
- 19.Oliveira CJ, Sa-Nunes A, Francischetti IM, Carregaro V, Anatriello E, Silva JS, et al. Deconstructing tick saliva: non-protein molecules with potent immunomodulatory properties. J Biol Chem. 2011;286:10960–9. doi: 10.1074/jbc.M110.205047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Steen NA, Barker SC, Alewood PF. Proteins in the saliva of the Ixodida (ticks): pharmacological features and biological significance. Toxicon. 2006;47:1–20. doi: 10.1016/j.toxicon.2005.09.010. [DOI] [PubMed] [Google Scholar]
- 21.Nuttall PA, Labuda M. Tick-host interactions: saliva-activated transmission. Parasitology. 2004;129(Suppl):S177–89. doi: 10.1017/s0031182004005633. [DOI] [PubMed] [Google Scholar]
- 22.Parizi LF, Githaka NW, Logullo C, Konnai S, Masuda A, Ohashi K, et al. The quest for a universal vaccine against ticks: cross-immunity insights. Vet J. 2012;194:158–65. doi: 10.1016/j.tvjl.2012.05.023. [DOI] [PubMed] [Google Scholar]
- 23.Imamura S, Konnai S, Vaz IS, Yamada S, Nakajima C, Ito Y, et al. Effects of anti-tick cocktail vaccine against Rhipicephalus appendiculatus. Jpn J Vet Res. 2008;56:85–98. [PubMed] [Google Scholar]
- 24.Havlikova S, Roller L, Koci J, Trimnell AR, Kazimirova M, Klempa B, et al. Functional role of 64P, the candidate transmission-blocking vaccine antigen from the tick, Rhipicephalus appendiculatus. Int J Parasitol. 2009;39:1485–94. doi: 10.1016/j.ijpara.2009.05.005. [DOI] [PubMed] [Google Scholar]
- 25.Labuda M, Trimnell AR, Lickova M, Kazimirova M, Davies GM, Lissina O, et al. An antivector vaccine protects against a lethal vector-borne pathogen. PLoS Pathog. 2006;2:e27. doi: 10.1371/journal.ppat.0020027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Neelakanta G, Sultana H. Transmission-blocking vaccines: focus on anti-vector vaccines against tick-borne diseases. Arch Immunol Ther Exp (Warsz) 2014;63(3):169–79. doi: 10.1007/s00005-014-0324-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ciprandi A, de Oliveira SK, Masuda A, Horn F, Termignoni C. Boophilus microplus: its saliva contains microphilin, a small thrombin inhibitor. Exp Parasitol. 2006;114:40–6. doi: 10.1016/j.exppara.2006.02.010. [DOI] [PubMed] [Google Scholar]
- 28.Horn F, Dos Santos PC, Termignoni C. Boophilus microplus anticoagulant protein: an antithrombin inhibitor isolated from the cattle tick saliva. Arch Biochem Biophys. 2000;384:68–73. doi: 10.1006/abbi.2000.2076. [DOI] [PubMed] [Google Scholar]
- 29.Liao M, Zhou J, Gong H, Boldbaatar D, Shirafuji R, Battur B, et al. Hemalin, a thrombin inhibitor isolated from a midgut cDNA library from the hard tick Haemaphysalis longicornis. J Insect Physiol. 2009;55:164–73. doi: 10.1016/j.jinsphys.2008.11.004. [DOI] [PubMed] [Google Scholar]
- 30.Madden RD, Sauer JR, Dillwith JW. A proteomics approach to characterizing tick salivary secretions. Exp Appl Acarol. 2002;28:77–87. [PubMed] [Google Scholar]
- 31.Valenzuela JG, Francischetti IM, Pham VM, Garfield MK, Mather TN, Ribeiro JM. Exploring the sialome of the tick Ixodes scapularis. J Exp Biol. 2002;205:2843–64. doi: 10.1242/jeb.205.18.2843. [DOI] [PubMed] [Google Scholar]
- 32.Radulovic ZM, Kim TK, Porter LM, Sze SH, Lewis L, Mulenga A. A 24–48 h fed Amblyomma americanum tick saliva immuno-proteome. BMC Genomics. 2014;15:518. doi: 10.1186/1471-2164-15-518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lewis LA, Radulovic Z, Kim TK, Porter L, Mulenga A. Identification of 24h Ixodes scapularis immunogenic tick saliva proteins. Ticks Tick Borne Dis. 2015;6(3):424–34. doi: 10.1016/j.ttbdis.2015.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.McDonald WH, Tabb DL, Sadygov RG, MacCoss MJ, Venable J, Graumann J, et al. MS1, MS2, and SQT-three unified, compact, and easily parsed file formats for the storage of shotgun proteomic spectra and identifications. Rapid Commun Mass Spectrom. 2004;18:2162–8. doi: 10.1002/rcm.1603. [DOI] [PubMed] [Google Scholar]
- 35.Xu T, Venable JD, Park SK, Conciorva D, Lu B, Liao L, Wohlschlegel J, Hewel J, Yates JR., III ProLuCID, a fast and sensitive tandem mass spectra-based protein identification program. Mol Cell Proteomics. 2006;5:S174. [Google Scholar]
- 36.Carvalho PC, Fischer JS, Xu T, Cociorva D, Balbuena TS, Valente RH, et al. Search engine processor: filtering and organizing peptide spectrum matches. Proteomics. 2012;12:944–9. doi: 10.1002/pmic.201100529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Carvalho PC, Fischer JS, Xu T, Yates JR, III, Barbosa VC. PatternLab: from mass spectra to label-free differential shotgun proteomics. Curr Protoc Bioinformatics. 2012;40:13.19:13.19.1–13.19.18. [DOI] [PubMed]
- 38.Zybailov B, Mosley AL, Sardiu ME, Coleman MK, Florens L, Washburn MP. Statistical analysis of membrane proteome expression changes in Saccharomyces cerevisiae. J Proteome Res. 2006;5:2339–47. doi: 10.1021/pr060161n. [DOI] [PubMed] [Google Scholar]
- 39.Lewis S, Ashburner M, Reese MG. Annotating eukaryote genomes. Curr Opin Struct Biol. 2000;10:349–54. doi: 10.1016/s0959-440x(00)00095-6. [DOI] [PubMed] [Google Scholar]
- 40.Marchler-Bauer A, Lu S, Anderson JB, Chitsaz F, Derbyshire MK, DeWeese-Scott C, et al. CDD: a Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011;39:D225–9. doi: 10.1093/nar/gkq1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tatusov RL, Fedorova ND, Jackson JD, Jacobs AR, Kiryutin B, Koonin EV, et al. The COG database: an updated version includes eukaryotes. BMC Bioinformatics. 2003;4:41. doi: 10.1186/1471-2105-4-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bateman A, Birney E, Cerruti L, Durbin R, Etwiller L, Eddy SR, et al. The Pfam protein families database. Nucleic Acids Res. 2002;30:276–80. doi: 10.1093/nar/30.1.276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Schultz J, Copley RR, Doerks T, Ponting CP, Bork P. SMART: a web-based tool for the study of genetically mobile domains. Nucleic Acids Res. 2000;28:231–4. doi: 10.1093/nar/28.1.231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Karim S, Singh P, Ribeiro JM. A deep insight into the sialotranscriptome of the gulf coast tick, Amblyomma maculatum. PLoS One. 2011;6:e28525. doi: 10.1371/journal.pone.0028525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yu Z, Wang H, Wang T, Sun W, Yang X, Liu J. Tick-borne pathogens and the vector potential of ticks in China. Parasit Vectors. 2015;8:24. doi: 10.1186/s13071-014-0628-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Binnington KC. Sequential changes in salivary gland structure during attachment and feeding of the cattle tick, Boophilus microplus. Int J Parasitol. 1978;8:97–115. doi: 10.1016/0020-7519(78)90004-8. [DOI] [PubMed] [Google Scholar]
- 47.Kaufman W. The influence of various factors on fluid secretion by in vitro salivary glands of ixodid Ticks. J Exp Biol. 1976;64:727–42. doi: 10.1242/jeb.64.3.727. [DOI] [PubMed] [Google Scholar]
- 48.McSwain JL, Essenberg RC, Sauer JR. Oral secretion elicited by effectors of signal transduction pathways in the salivary glands of Amblyomma americanum (Acari: Ixodidae) J Med Entomol. 1992;29:41–8. doi: 10.1093/jmedent/29.1.41. [DOI] [PubMed] [Google Scholar]
- 49.Sauer JR, McSwain JL, Bowman AS, Essenberg RC. Tick salivary gland physiology. Annu Rev Entomol. 1995;40:245–67. doi: 10.1146/annurev.en.40.010195.001333. [DOI] [PubMed] [Google Scholar]
- 50.Parizi LF, Utiumi KU, Imamura S, Onuma M, Ohashi K, Masuda A, et al. Cross immunity with Haemaphysalis longicornis glutathione S-transferase reduces an experimental Rhipicephalus (Boophilus) microplus infestation. Exp Parasitol. 2011;127:113–8. doi: 10.1016/j.exppara.2010.07.001. [DOI] [PubMed] [Google Scholar]
- 51.Parizi LF, Reck J, Jr, Oldiges DP, Guizzo MG, Seixas A, Logullo C, et al. Multi-antigenic vaccine against the cattle tick Rhipicephalus (Boophilus) microplus: a field evaluation. Vaccine. 2012;30:6912–7. doi: 10.1016/j.vaccine.2012.08.078. [DOI] [PubMed] [Google Scholar]
- 52.Kotsyfakis M, Anderson JM, Andersen JF, Calvo E, Francischetti IM, Mather TN, et al. Cutting edge: Immunity against a “silent” salivary antigen of the Lyme vector Ixodes scapularis impairs its ability to feed. J Immunol. 2008;181:5209–12. doi: 10.4049/jimmunol.181.8.5209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Galay RL, Miyata T, Umemiya-Shirafuji R, Maeda H, Kusakisako K, Tsuji N, et al. Evaluation and comparison of the potential of two ferritins as anti-tick vaccines against Haemaphysalis longicornis. Parasit Vectors. 2014;7:482. doi: 10.1186/s13071-014-0482-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hajdusek O, Almazan C, Loosova G, Villar M, Canales M, Grubhoffer L, et al. Characterization of ferritin 2 for the control of tick infestations. Vaccine. 2010;28:2993–8. doi: 10.1016/j.vaccine.2010.02.008. [DOI] [PubMed] [Google Scholar]
- 55.Imamura S, da Silva VJ I, Sugino M, Ohashi K, Onuma M. A serine protease inhibitor (serpin) from Haemaphysalis longicornis as an anti-tick vaccine. Vaccine. 2005;23:1301–11. doi: 10.1016/j.vaccine.2004.08.041. [DOI] [PubMed] [Google Scholar]
- 56.Imamura S, Namangala B, Tajima T, Tembo ME, Yasuda J, Ohashi K, et al. Two serine protease inhibitors (serpins) that induce a bovine protective immune response against Rhipicephalus appendiculatus ticks. Vaccine. 2006;24:2230–7. doi: 10.1016/j.vaccine.2005.10.055. [DOI] [PubMed] [Google Scholar]
- 57.Jittapalapong S, Kaewhom P, Pumhom P, Canales M, de la Fuente J, Stich RW. Immunization of rabbits with recombinant serine protease inhibitor reduces the performance of adult female Rhipicephalus microplus. Transbound Emerg Dis. 2010;57:103–6. doi: 10.1111/j.1865-1682.2010.01108.x. [DOI] [PubMed] [Google Scholar]
- 58.Prevot PP, Couvreur B, Denis V, Brossard M, Vanhamme L, Godfroid E. Protective immunity against Ixodes ricinus induced by a salivary serpin. Vaccine. 2007;25:3284–92. doi: 10.1016/j.vaccine.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 59.Sugino M, Imamura S, Mulenga A, Nakajima M, Tsuda A, Ohashi K, et al. A serine proteinase inhibitor (serpin) from ixodid tick Haemaphysalis longicornis; cloning and preliminary assessment of its suitability as a candidate for a tick vaccine. Vaccine. 2003;21:2844–51. doi: 10.1016/s0264-410x(03)00167-1. [DOI] [PubMed] [Google Scholar]
- 60.Taheri M, Nabian S, Ranjbar M, Mazaheri NR, Gerami SA, Sazmand A. Study of vitellogenin in Boophilus annulatus tick larvae and its immunological aspects. Trop Biomed. 2014;31:398–405. [PubMed] [Google Scholar]
- 61.Tellam RL, Kemp D, Riding G, Briscoe S, Smith D, Sharp P, et al. Reduced oviposition of Boophilus microplus feeding on sheep vaccinated with vitellin. Vet Parasitol. 2002;103:141–56. doi: 10.1016/s0304-4017(01)00573-8. [DOI] [PubMed] [Google Scholar]
- 62.Logullo C, Moraes J, Dansa-Petretski M, Vaz IS, Masuda A, Sorgine MH, et al. Binding and storage of heme by vitellin from the cattle tick, Boophilus microplus. Insect Biochem Mol Biol. 2002;32:1805–11. doi: 10.1016/s0965-1748(02)00162-5. [DOI] [PubMed] [Google Scholar]
- 63.Maya-Monteiro CM, Daffre S, Logullo C, Lara FA, Alves EW, Capurro ML, et al. HeLp, a heme lipoprotein from the hemolymph of the cattle tick, Boophilus microplus. J Biol Chem. 2000;275:36584–9. doi: 10.1074/jbc.M007344200. [DOI] [PubMed] [Google Scholar]
- 64.Dutra FF, Alves LS, Rodrigues D, Fernandez PL, de Oliveira RB, Golenbock DT, et al. Hemolysis-induced lethality involves inflammasome activation by heme. Proc Natl Acad Sci U S A. 2014;111:E4110–8. doi: 10.1073/pnas.1405023111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Dutra FF, Bozza MT. Heme on innate immunity and inflammation. Front Pharmacol. 2014;5:115. doi: 10.3389/fphar.2014.00115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Graca-Souza AV, Arruda MA, de Freitas MS, Barja-Fidalgo C, Oliveira PL. Neutrophil activation by heme: implications for inflammatory processes. Blood. 2002;99:4160–5. doi: 10.1182/blood.v99.11.4160. [DOI] [PubMed] [Google Scholar]
- 67.Pichu S, Ribeiro JM, Mather TN, Francischetti IM. Purification of a serine protease and evidence for a protein C activator from the saliva of the tick, Ixodes scapularis. Toxicon. 2014;77:32–9. doi: 10.1016/j.toxicon.2013.10.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Franta Z, Frantova H, Konvickova J, Horn M, Sojka D, Mares M, et al. Dynamics of digestive proteolytic system during blood feeding of the hard tick Ixodes ricinus. Parasit Vectors. 2010;3:119. doi: 10.1186/1756-3305-3-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sojka D, Franta Z, Horn M, Caffrey CR, Mares M, Kopacek P. New insights into the machinery of blood digestion by ticks. Trends Parasitol. 2013;29:276–85. doi: 10.1016/j.pt.2013.04.002. [DOI] [PubMed] [Google Scholar]
- 70.Tsuji N, Miyoshi T, Battsetseg B, Matsuo T, Xuan X, Fujisaki K. A cysteine protease is critical for Babesia spp. transmission in Haemaphysalis ticks. PLoS Pathog. 2008;4:e1000062. doi: 10.1371/journal.ppat.1000062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature. 2001;413:203–10. doi: 10.1038/35093019. [DOI] [PubMed] [Google Scholar]
- 72.Ribeiro JM, Mather TN. Ixodes scapularis: salivary kininase activity is a metallo dipeptidyl carboxypeptidase. Exp Parasitol. 1998;89:213–21. doi: 10.1006/expr.1998.4296. [DOI] [PubMed] [Google Scholar]
- 73.Gettins PG. Serpin structure, mechanism, and function. Chem Rev. 2002;102:4751–804. doi: 10.1021/cr010170+. [DOI] [PubMed] [Google Scholar]
- 74.Mulenga A, Khumthong R, Blandon RA. Molecular and expression analysis of a family of the Amblyomma americanum tick Lospins. J Exp Biol. 2007;210:3188–98. doi: 10.1242/jeb.006494. [DOI] [PubMed] [Google Scholar]
- 75.Ibelli AM, Kim TK, Hill CC, Lewis LA, Bakshi M, Miller S, et al. A blood meal-induced Ixodes scapularis tick saliva serpin inhibits trypsin and thrombin, and interferes with platelet aggregation and blood clotting. Int J Parasitol. 2014;44:369–79. doi: 10.1016/j.ijpara.2014.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Mulenga A, Kim T, Ibelli AM. Amblyomma americanum tick saliva serine protease inhibitor 6 is a cross-class inhibitor of serine proteases and papain-like cysteine proteases that delays plasma clotting and inhibits platelet aggregation. Insect Mol Biol. 2013;22:306–19. doi: 10.1111/imb.12024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Prevot PP, Adam B, Boudjeltia KZ, Brossard M, Lins L, Cauchie P, et al. Anti-hemostatic effects of a serpin from the saliva of the tick Ixodes ricinus. J Biol Chem. 2006;281:26361–9. doi: 10.1074/jbc.M604197200. [DOI] [PubMed] [Google Scholar]
- 78.Chmelar J, Oliveira CJ, Rezacova P, Francischetti IM, Kovarova Z, Pejler G, et al. A tick salivary protein targets cathepsin G and chymase and inhibits host inflammation and platelet aggregation. Blood. 2011;117:736–44. doi: 10.1182/blood-2010-06-293241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Leboulle G, Crippa M, Decrem Y, Mejri N, Brossard M, Bollen A, et al. Characterization of a novel salivary immunosuppressive protein from Ixodes ricinus ticks. J Biol Chem. 2002;277:10083–9. doi: 10.1074/jbc.M111391200. [DOI] [PubMed] [Google Scholar]
- 80.Prevot PP, Beschin A, Lins L, Beaufays J, Grosjean A, Bruys L, et al. Exosites mediate the anti-inflammatory effects of a multifunctional serpin from the saliva of the tick Ixodes ricinus. FEBS J. 2009;276:3235–46. doi: 10.1111/j.1742-4658.2009.07038.x. [DOI] [PubMed] [Google Scholar]
- 81.Chisenhall DM, Christofferson RC, McCracken MK, Johnson AM, Londono-Renteria B, Mores CN. Infection with dengue-2 virus alters proteins in naturally expectorated saliva of Aedes aegypti mosquitoes. Parasit Vectors. 2014;7:252. doi: 10.1186/1756-3305-7-252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Williams AR, Zakutansky SE, Miura K, Dicks MD, Churcher TS, Jewell KE, et al. Immunisation against a serine protease inhibitor reduces intensity of Plasmodium berghei infection in mosquitoes. Int J Parasitol. 2013;43:869–74. doi: 10.1016/j.ijpara.2013.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Abrahamson M, Alvarez-Fernandez M, Nathanson CM. Cystatins. Biochem Soc Symp. 2003;70:179–99. doi: 10.1042/bss0700179. [DOI] [PubMed] [Google Scholar]
- 84.Kotsyfakis M, Sa-Nunes A, Francischetti IM, Mather TN, Andersen JF, Ribeiro JM. Antiinflammatory and immunosuppressive activity of sialostatin L, a salivary cystatin from the tick Ixodes scapularis. J Biol Chem. 2006;281:26298–307. doi: 10.1074/jbc.M513010200. [DOI] [PubMed] [Google Scholar]
- 85.Kotsyfakis M, Karim S, Andersen JF, Mather TN, Ribeiro JM. Selective cysteine protease inhibition contributes to blood-feeding success of the tick Ixodes scapularis. J Biol Chem. 2007;282:29256–63. doi: 10.1074/jbc.M703143200. [DOI] [PubMed] [Google Scholar]
- 86.Parizi LF, Sabadin GA, Alzugaray MF, Seixas A, Logullo C, Konnai S, et al. Rhipicephalusmicroplus and Ixodesovatus cystatins in tick blood digestion and evasion of host immune response. Parasit Vectors. 2015;8:122. doi: 10.1186/s13071-015-0743-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Sa-Nunes A, Bafica A, Antonelli LR, Choi EY, Francischetti IM, Andersen JF, et al. The immunomodulatory action of sialostatin L on dendritic cells reveals its potential to interfere with autoimmunity. J Immunol. 2009;182:7422–9. doi: 10.4049/jimmunol.0900075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Karim S, Miller NJ, Valenzuela J, Sauer JR, Mather TN. RNAi-mediated gene silencing to assess the role of synaptobrevin and cystatin in tick blood feeding. Biochem Biophys Res Commun. 2005;334:1336–42. doi: 10.1016/j.bbrc.2005.07.036. [DOI] [PubMed] [Google Scholar]
- 89.Kotsyfakis M, Horka H, Salat J, Andersen JF. The crystal structures of two salivary cystatins from the tick Ixodes scapularis and the effect of these inhibitors on the establishment of Borrelia burgdorferi infection in a murine model. Mol Microbiol. 2010;77:456–70. doi: 10.1111/j.1365-2958.2010.07220.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Fogaca AC, Almeida IC, Eberlin MN, Tanaka AS, Bulet P, Daffre S. Ixodidin, a novel antimicrobial peptide from the hemocytes of the cattle tick Boophilus microplus with inhibitory activity against serine proteinases. Peptides. 2006;27:667–74. doi: 10.1016/j.peptides.2005.07.013. [DOI] [PubMed] [Google Scholar]
- 91.Sasaki SD, Cotrin SS, Carmona AK, Tanaka AS. An unexpected inhibitory activity of Kunitz-type serine proteinase inhibitor derived from Boophilus microplus trypsin inhibitor on cathepsin L. Biochem Biophys Res Commun. 2006;341:266–72. doi: 10.1016/j.bbrc.2005.12.178. [DOI] [PubMed] [Google Scholar]
- 92.Cvirn G, Gallistl S, Koestenberger M, Kutschera J, Leschnik B, Muntean W. Alpha 2-macroglobulin enhances prothrombin activation and thrombin potential by inhibiting the anticoagulant protein C/protein S system in cord and adult plasma. Thromb Res. 2002;105:433–9. doi: 10.1016/s0049-3848(02)00042-7. [DOI] [PubMed] [Google Scholar]
- 93.Harpel PC. Human plasma alpha 2-macroglobulin: an inhibitor of plasma kallikrein. J Exp Med. 1970;132:329–52. doi: 10.1084/jem.132.2.329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Meijers JC, Tijburg PN, Bouma BN. Inhibition of human blood coagulation factor Xa by alpha 2-macroglobulin. Biochemistry. 1987;26:5932–7. doi: 10.1021/bi00392a053. [DOI] [PubMed] [Google Scholar]
- 95.Banks RE, Evans SW, Van LF, Alexander D, McMahon MJ, Whicher JT. Measurement of the ‘fast’ or complexed form of alpha 2 macroglobulin in biological fluids using a sandwich enzyme immunoassay. J Immunol Methods. 1990;126:13–20. doi: 10.1016/0022-1759(90)90006-h. [DOI] [PubMed] [Google Scholar]
- 96.Bonacci GR, Caceres LC, Sanchez MC, Chiabrando GA. Activated alpha(2)-macroglobulin induces cell proliferation and mitogen-activated protein kinase activation by LRP-1 in the J774 macrophage-derived cell line. Arch Biochem Biophys. 2007;460:100–6. doi: 10.1016/j.abb.2007.01.004. [DOI] [PubMed] [Google Scholar]
- 97.Anatriello E, Ribeiro JM, de Miranda-Santos IK, Brandao LG, Anderson JM, Valenzuela JG, et al. An insight into the sialotranscriptome of the brown dog tick, Rhipicephalus sanguineus. BMC Genomics. 2010;11:450. doi: 10.1186/1471-2164-11-450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Gibson AK, Smith Z, Fuqua C, Clay K, Colbourne JK. Why so many unknown genes? Partitioning orphans from a representative transcriptome of the lone star tick Amblyomma americanum. BMC Genomics. 2013;14:135. doi: 10.1186/1471-2164-14-135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Mulenga A, Blandon M, Khumthong R. The molecular basis of the Amblyomma americanum tick attachment phase. Exp Appl Acarol. 2007;41:267–87. doi: 10.1007/s10493-007-9064-3. [DOI] [PubMed] [Google Scholar]
- 100.Mulenga A, Kim TK, Ibelli AM. Deorphanization and target validation of cross-tick species conserved novel Amblyomma americanum tick saliva protein. Int J Parasitol. 2013;43:439–51. doi: 10.1016/j.ijpara.2012.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Xu XL, Cheng TY, Yang H, Yan F, Yang Y. De Novo Sequencing, Assembly and Analysis of Salivary Gland Transcriptome of Haemaphysalis flava and Identification of Sialoprotein Genes. Infect Genet Evol. 2015;32:135–142. doi: 10.1016/j.meegid.2015.03.010. [DOI] [PubMed] [Google Scholar]
- 102.Megraw T, Kaufman TC, Kovalick GE. Sequence and expression of Drosophila Antigen 5-related 2, a new member of the CAP gene family. Gene. 1998;222:297–304. doi: 10.1016/s0378-1119(98)00489-2. [DOI] [PubMed] [Google Scholar]
- 103.Garcia GR, Gardinassi LG, Ribeiro JM, Anatriello E, Ferreira BR, Moreira HN, et al. The sialotranscriptome of Amblyomma triste, Amblyomma parvum and Amblyomma cajennense ticks, uncovered by 454-based RNA-seq. Parasit Vectors. 2014;7:430. doi: 10.1186/1756-3305-7-430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Anderson JM, Oliveira F, Kamhawi S, Mans BJ, Reynoso D, Seitz AE, Lawyer P, Garfield M, Pham M, Valenzuela JG. Comparative salivary gland transcriptomics of sandfly vectors of visceral leishmaniasis. BMC Genomics. 2006;7:52. doi: 10.1186/1471-2164-7-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Bishop R, Lambson B, Wells C, Pandit P, Osaso J, Nkonge C, et al. A cement protein of the tick Rhipicephalus appendiculatus, located in the secretory e cell granules of the type III salivary gland acini, induces strong antibody responses in cattle. Int J Parasitol. 2002;32:833–42. doi: 10.1016/s0020-7519(02)00027-9. [DOI] [PubMed] [Google Scholar]
- 106.Zhou J, Gong H, Zhou Y, Xuan X, Fujisaki K. Identification of a glycine-rich protein from the tick Rhipicephalus haemaphysaloides and evaluation of its vaccine potential against tick feeding. Parasitol Res. 2006;100:77–84. doi: 10.1007/s00436-006-0243-7. [DOI] [PubMed] [Google Scholar]
- 107.Maruyama SR, Anatriello E, Anderson JM, Ribeiro JM, Brandao LG, Valenzuela JG, et al. The expression of genes coding for distinct types of glycine-rich proteins varies according to the biology of three metastriate ticks, Rhipicephalus (Boophilus) microplus, Rhipicephalus sanguineus and Amblyomma cajennense. BMC Genomics. 2010;11:363. doi: 10.1186/1471-2164-11-363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Liu XY, Bonnet SI. Hard tick factors implicated in pathogen transmission. PLoS Negl Trop Dis. 2014;8:e2566. doi: 10.1371/journal.pntd.0002566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Mans BJ, Neitz AW. Exon-intron structure of outlier tick lipocalins indicate a monophyletic origin within the larger lipocalin family. Insect Biochem Mol Biol. 2004;34:585–94. doi: 10.1016/j.ibmb.2004.03.006. [DOI] [PubMed] [Google Scholar]
- 110.Paesen GC, Adams PL, Harlos K, Nuttall PA, Stuart DI. Tick histamine-binding proteins: isolation, cloning, and three-dimensional structure. Mol Cell. 1999;3:661–71. doi: 10.1016/s1097-2765(00)80359-7. [DOI] [PubMed] [Google Scholar]
- 111.Ganfornina MD, Kayser H, Sanchez D. Lipocalins in Arthropoda: diversification and functional explorations. In: Madame Curie Bioscinece Database. [Internet]. Austin (TX). Edited by Lands Bioscience: Lands Bioscience; 2000. Available from: http://www.ncbi.nlm.nih.gov/books/NBK6576/.
- 112.Beaufays J, Adam B, Menten-Dedoyart C, Fievez L, Grosjean A, Decrem Y, et al. Ir-LBP, an Ixodes ricinus tick salivary LTB4-binding lipocalin, interferes with host neutrophil function. PLoS One. 2008;3:e3987. doi: 10.1371/journal.pone.0003987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Mans BJ, Ribeiro JM. Function, mechanism and evolution of the moubatin-clade of soft tick lipocalins. Insect Biochem Mol Biol. 2008;38:841–52. doi: 10.1016/j.ibmb.2008.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Mans BJ, Ribeiro JM, Andersen JF. Structure, function, and evolution of biogenic amine-binding proteins in soft ticks. J Biol Chem. 2008;283:18721–33. doi: 10.1074/jbc.M800188200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Kim HJ, Je HJ, Cheon HM, Kong SY, Han J, Yun CY, et al. Accumulation of 23 kDa lipocalin during brain development and injury in Hyphantria cunea. Insect Biochem Mol Biol. 2005;35:1133–41. doi: 10.1016/j.ibmb.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 116.McNally KL, Mitzel DN, Anderson JM, Ribeiro JM, Valenzuela JG, Myers TG, et al. Differential salivary gland transcript expression profile in Ixodes scapularis nymphs upon feeding or flavivirus infection. Ticks Tick Borne Dis. 2012;3:18–26. doi: 10.1016/j.ttbdis.2011.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Cheng PH, Mans BJ, Neitz AW, Gaspar AR. Savicalin, a lipocalin from hemocytes of the soft tick, Ornithodoros savignyi. Exp Appl Acarol. 2010;52:313–26. doi: 10.1007/s10493-010-9368-6. [DOI] [PubMed] [Google Scholar]
- 118.Heekin AM, Guerrero FD, Bendele KG, Saldivar L, Scoles GA, Dowd SE, et al. The ovarian transcriptome of the cattle tick, Rhipicephalus (Boophilus) microplus, feeding upon a bovine host infected with Babesia bovis. Parasit Vectors. 2013;6:276. doi: 10.1186/1756-3305-6-276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Chu FF, Esworthy RS, Doroshow JH. Role of Se-dependent glutathione peroxidases in gastrointestinal inflammation and cancer. Free Radic Biol Med. 2004;36:1481–95. doi: 10.1016/j.freeradbiomed.2004.04.010. [DOI] [PubMed] [Google Scholar]
- 120.Pichu S, Yalcin EB, Ribeiro JM, King RS, Mather TN. Molecular characterization of novel sulfotransferases from the tick, Ixodes scapularis. BMC Biochem. 2011;12:32. doi: 10.1186/1471-2091-12-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Coles TB, Dryden MW. Insecticide/acaricide resistance in fleas and ticks infesting dogs and cats. Parasit Vectors. 2014;7:8. doi: 10.1186/1756-3305-7-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.da Silva VI, Jr, Torino LT, Michelon A, Sanchez Ferreira CA, de Freitas DR J, Termignoni C, et al. Effect of acaricides on the activity of a Boophilus microplus glutathione S-transferase. Vet Parasitol. 2004;119:237–45. doi: 10.1016/j.vetpar.2003.11.004. [DOI] [PubMed] [Google Scholar]
- 123.Ouaissi A, Ouaissi M, Sereno D. Glutathione S-transferases and related proteins from pathogenic human parasites behave as immunomodulatory factors. Immunol Lett. 2002;81:159–64. doi: 10.1016/s0165-2478(02)00035-4. [DOI] [PubMed] [Google Scholar]
- 124.Plumas-Marty B, Verwaerde C, Loyens M, Velge P, Taibi A, Cesbron MF, et al. Trypanosoma cruzi glutathione-binding proteins: immunogenicity during human and experimental Chagas’ disease. Parasitology. 1992;104(Pt 1):87–98. doi: 10.1017/s0031182000060832. [DOI] [PubMed] [Google Scholar]
- 125.Toh SQ, Glanfield A, Gobert GN, Jones MK. Heme and blood-feeding parasites: friends or foes? Parasit Vectors. 2010;3:108. doi: 10.1186/1756-3305-3-108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Freitas DR, Rosa RM, Moraes J, Campos E, Logullo C, da Silva VI, Jr, et al. Relationship between glutathione S-transferase, catalase, oxygen consumption, lipid peroxidation and oxidative stress in eggs and larvae of Boophilus microplus (Acarina: Ixodidae) Comp Biochem Physiol A Mol Integr Physiol. 2007;146:688–94. doi: 10.1016/j.cbpa.2006.04.032. [DOI] [PubMed] [Google Scholar]
- 127.Wei SH, Clark AG, Syvanen M. Identification and cloning of a key insecticide-metabolizing glutathione S-transferase (MdGST-6A) from a hyper insecticide-resistant strain of the housefly Musca domestica. Insect Biochem Mol Biol. 2001;31:1145–53. doi: 10.1016/s0965-1748(01)00059-5. [DOI] [PubMed] [Google Scholar]
- 128.Freitas DR, Rosa RM, Moura DJ, Seitz AL, Colodel EM, Driemeier D, et al. Cell death during preoviposition period in Boophilus microplus tick. Vet Parasitol. 2007;144:321–7. doi: 10.1016/j.vetpar.2006.10.017. [DOI] [PubMed] [Google Scholar]
- 129.Furquim KC, Bechara GH, Camargo Mathias MI. Degeneration of salivary glands of males of the tick Rhipicephalus sanguineus (Latreille, 1806) (Acari, Ixodidae) Vet Parasitol. 2008;154:325–35. doi: 10.1016/j.vetpar.2008.03.013. [DOI] [PubMed] [Google Scholar]
- 130.Coons LB, Roshdy MA. Fine structure of the salivary glands of unfed male Dermacentor variabilis (Say) (Ixodoidea: Ixodidae) J Parasitol. 1973;59:900–12. [PubMed] [Google Scholar]
- 131.Leal BF, Seixas A, Mattos RT, Coutinho ML, Masuda A, da Silva VI, Jr, et al. Tissue expression and the host’s immunological recognition of a Rhipicephalus microplus paramyosin. Vet Parasitol. 2013;197:304–11. doi: 10.1016/j.vetpar.2013.06.020. [DOI] [PubMed] [Google Scholar]
- 132.Fukumoto S, Sakaguchi T, You M, Xuan X, Fujisaki K. Tick troponin I-like molecule is a potent inhibitor for angiogenesis. Microvasc Res. 2006;71:218–21. doi: 10.1016/j.mvr.2006.02.003. [DOI] [PubMed] [Google Scholar]
- 133.Diaz-Martin V, Manzano-Roman R, Oleaga A, Encinas-Grandes A, Perez-Sanchez R. Cloning and characterization of a plasminogen-binding enolase from the saliva of the argasid tick Ornithodoros moubata. Vet Parasitol. 2013;191:301–14. doi: 10.1016/j.vetpar.2012.09.019. [DOI] [PubMed] [Google Scholar]
- 134.Jolodar A, Fischer P, Bergmann S, Buttner DW, Hammerschmidt S, Brattig NW. Molecular cloning of an alpha-enolase from the human filarial parasite Onchocerca volvulus that binds human plasminogen. Biochim Biophys Acta. 2003;1627:111–20. doi: 10.1016/s0167-4781(03)00083-6. [DOI] [PubMed] [Google Scholar]
- 135.Vanegas G, Quinones W, Carrasco-Lopez C, Concepcion JL, Albericio F, Avilan L. Enolase as a plasminogen binding protein in Leishmania mexicana. Parasitol Res. 2007;101:1511–6. doi: 10.1007/s00436-007-0668-7. [DOI] [PubMed] [Google Scholar]
- 136.Hamid R, Khan MA, Ahmad M, Ahmad MM, Abdin MZ, Musarrat J, et al. Chitinases: an update. J Pharm Bioallied Sci. 2013;5:21–9. doi: 10.4103/0975-7406.106559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Kim TK, Curran J, Mulenga A. Dual silencing of long and short Amblyomma americanum acidic chitinase forms weakens the tick cement cone stability. J Exp Biol. 2014;217:3493–503. doi: 10.1242/jeb.107979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Kotsyfakis M, Schwarz A, Erhart J, Ribeiro JM. Tissue- and time-dependent transcription in Ixodes ricinus salivary glands and midguts when blood feeding on the vertebrate host. Sci. Rep. 2015;5:9103. doi: 10.1038/srep09103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Burke JE, Dennis EA. Phospholipase A2 structure/function, mechanism, and signaling. J Lipid Res. 2009;50(Suppl):S237–42. doi: 10.1194/jlr.R800033-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Bowman AS, Gengler CL, Surdick MR, Zhu K, Essenberg RC, Sauer JR, et al. A novel phospholipase A2 activity in saliva of the lone star tick, Amblyomma americanum (L.) Exp Parasitol. 1997;87:121–32. doi: 10.1006/expr.1997.4201. [DOI] [PubMed] [Google Scholar]
- 141.Huberts DH, van der Klei IJ. Moonlighting proteins: an intriguing mode of multitasking. Biochim Biophys Acta. 2010;1803:520–5. doi: 10.1016/j.bbamcr.2010.01.022. [DOI] [PubMed] [Google Scholar]
- 142.Mani M, Chen C, Amblee V, Liu H, Mathur T, Zwicke G, et al. MoonProt: a database for proteins that are known to moonlight. Nucleic Acids Res. 2015;43:D277–82. doi: 10.1093/nar/gku954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Anderson JF, Magnarelli LA. Biology of ticks. Infect Dis Clin North Am. 2008;22:195–215. doi: 10.1016/j.idc.2007.12.006. [DOI] [PubMed] [Google Scholar]
- 144.Ribeiro JM. Role of saliva in blood-feeding by arthropods. Annu Rev Entomol. 1987;32:463–78. doi: 10.1146/annurev.en.32.010187.002335. [DOI] [PubMed] [Google Scholar]
- 145.Ribeiro JM. How ticks make a living. Parasitol Today. 1995;11:91–3. doi: 10.1016/0169-4758(95)80162-6. [DOI] [PubMed] [Google Scholar]
- 146.McSwain JL, Essenberg RC, Sauer JR. Protein changes in the salivary glands of the female lone star tick, Amblyomma americanum, during feeding. J Parasitol. 1982;68:100–6. [PubMed] [Google Scholar]
- 147.Schwarz A, von Reumont BM, Erhart J, Chagas AC, Ribeiro JM, Kotsyfakis M. De novo Ixodes ricinus salivary gland transcriptome analysis using two next-generation sequencing methodologies. FASEB J. 2013;27:4745–56. doi: 10.1096/fj.13-232140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Schwarz A, Tenzer S, Hackenberg M, Erhart J, Gerhold-Ay A, Mazur J, et al. A systems level analysis reveals transcriptomic and proteomic complexity in Ixodes ricinus midgut and salivary glands during early attachment and feeding. Mol Cell Proteomics. 2014;13:2725–35. doi: 10.1074/mcp.M114.039289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Chmelar J, Anderson JM, Mu J, Jochim RC, Valenzuela JG, Kopecky J. Insight into the sialome of the castor bean tickm, Ixodes ricinus. BMC Genomics. 2008;9:233. doi: 10.1186/1471-2164-9-233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Leboulle G, Rochez C, Louahed J, Ruti B, Brossard M, Bollen A, et al. Isolation of Ixodes ricinus salivary gland mRNA encoding factors induced during blood feeding. Am J Trop Med Hyg. 2002;66:225–33. doi: 10.4269/ajtmh.2002.66.225. [DOI] [PubMed] [Google Scholar]
- 151.Bowman AS, Sauer JR. Tick salivary glands: function, physiology and future. Parasitology. 2004;129(Suppl):S67–81. doi: 10.1017/s0031182004006468. [DOI] [PubMed] [Google Scholar]
- 152.Vaz JI, Martinez RH, Oliveira A, Heck A, Logullo C, Gonzales JC, et al. Functional bovine immunoglobulins in Boophilus microplus hemolymph. Vet Parasitol. 1996;62:155–60. doi: 10.1016/0304-4017(95)00851-9. [DOI] [PubMed] [Google Scholar]
- 153.Wang H, Nuttall PA. Excretion of host immunoglobulin in tick saliva and detection of IgG-binding proteins in tick haemolymph and salivary glands. Parasitology. 1994;109(Pt 4):525–30. doi: 10.1017/s0031182000080781. [DOI] [PubMed] [Google Scholar]
- 154.Francischetti IM, Anderson JM, Manoukis N, Pham VM, Ribeiro JM. An insight into the sialotranscriptome and proteome of the coarse bontlegged tick, Hyalomma marginatum rufipes. J Proteomics. 2011;74:2892–908. doi: 10.1016/j.jprot.2011.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Fogaca AC, da Silva PIJ, Miranda MT, Bianchi AG, Miranda A, Ribolla PE, et al. Antimicrobial activity of a bovine hemoglobin fragment in the tick Boophilus microplus. J Biol Chem. 1999;274:25330–4. doi: 10.1074/jbc.274.36.25330. [DOI] [PubMed] [Google Scholar]
- 156.Nakajima Y, Ogihara K, Taylor D, Yamakawa M. Antibacterial hemoglobin fragments from the midgut of the soft tick, Ornithodoros moubata (Acari: Argasidae) J Med Entomol. 2003;40:78–81. doi: 10.1603/0022-2585-40.1.78. [DOI] [PubMed] [Google Scholar]
- 157.Ackerman S, Clare FB, McGill TW, Sonenshine DE. Passage of host serum components, including antibody, across the digestive tract of Dermacentor variabilis (Say) J Parasitol. 1981;67:737–40. [PubMed] [Google Scholar]
- 158.Wickramasekara S, Bunikis J, Wysocki V, Barbour AG. Identification of residual blood proteins in ticks by mass spectrometry proteomics. Emerg Infect Dis. 2008;14:1273–5. doi: 10.3201/eid1408.080227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Mori H, Galay RL, Maeda H, Matsuo T, Umemiya-Shirafuji R, Mochizuki M, et al. Host-derived transferrin is maintained and transferred from midgut to ovary in Haemaphysalis longicornis ticks. Ticks Tick Borne Dis. 2014;5:121–6. doi: 10.1016/j.ttbdis.2013.09.004. [DOI] [PubMed] [Google Scholar]
- 160.Braz GR, Coelho HS, Masuda H, Oliveira PL. A missing metabolic pathway in the cattle tick Boophilus microplus. Curr Biol. 1999;9:703–6. doi: 10.1016/s0960-9822(99)80312-1. [DOI] [PubMed] [Google Scholar]
- 161.Mulenga A, Sugino M, Nakajim M, Sugimoto C, Onuma M. Tick-Encoded serine proteinase inhibitors (serpins); potential target antigens for tick vaccine development. J Vet Med Sci. 2001;63:1063–9. doi: 10.1292/jvms.63.1063. [DOI] [PubMed] [Google Scholar]
- 162.Rau JC, Beaulieu LM, Huntington JA, Church FC. Serpins in thrombosis, hemostasis and fibrinolysis. J Thromb Haemost. 2007;5(Suppl 1):102–15. doi: 10.1111/j.1538-7836.2007.02516.x. [DOI] [PMC free article] [PubMed] [Google Scholar]