

Abstract
Background
The mechanical transmission of pathogenic bacteria by synanthropic filth flies is widely recognized. While many studies report the fate and the temporospatial distribution of ingested foodborne bacteria by filth flies, there is little evidence about the transmission dynamics of ingested foodborne bacteria by adult house flies (Musca domestica) to their progeny. In this study, we fed parental house fly adults with food contaminated with low, medium, and high concentrations of Salmonella enterica, Cronobacter sakazakii, Escherichia coli O157:H7, and Listeria monocytogenes and evaluated the probability of transmission of these pathogens to house fly eggs and the surface and the alimentary canal of their first filial (F1) generation adults.
Results
All foodborne pathogens were present in samples containing pooled house fly eggs. The probability of transmission was higher after parental house flies ingested food containing medium bacterial loads. Cronobacter sakazakii was 16, 6, and 3 times more likely to be transmitted to house fly eggs than S. enterica, E. coli O157:H7, and L. monocytogenes, respectively. Only S. enterica and C. sakazakii were transmitted to F1 generation adults and their presence was 2.4 times more likely on their body surfaces than in their alimentary canals. The highest probabilities of finding S. enterica (60 %) and C. sakazakii (28 %) on newly emerged F1 adults were observed after parental house flies ingested food containing medium and high levels of these pathogens, respectively.
Conclusion
Our study demonstrates that adult house flies that fed from food contaminated with various levels of foodborne bacteria were able to transmit those pathogens to their eggs and some were further transmitted to newly emerged F1 generation adults, enhancing the vector potential of these insects. Understanding the type of associations that synanthropic filth flies establish with foodborne pathogens will help to elucidate transmission mechanisms and possible ways to mitigate the spread of foodborne pathogens.
Electronic supplementary material
The online version of this article (doi:10.1186/s12866-015-0478-5) contains supplementary material, which is available to authorized users.
Background
The biology and ecology of synanthropic insects like flies make them efficient carriers of disease-causing microorganisms. Their breeding habits, mode of feeding and indiscriminate traveling between decomposed waste and human settings highly contribute to the dissemination of pathogens in the environment. Approximately 350 fly species in 29 families are potentially associated with the transmission of diseases of public health importance [1]. However, fewer numbers of fly species have been associated with the transmission of foodborne pathogens [1, 2]. Although there are scarce reports of filth flies being the causative agent of foodborne outbreaks, several studies have demonstrated a steady decrease in the incidence of foodborne diarrhea after suppressing fly populations [3–5], indirectly implicating filth flies as the source of the foodborne pathogen.
The presence of flies in food and food facilities has always been a concern of the U.S. Food and Drug Administration (FDA). The FDA’s regulatory action criteria for filth includes a five-attribute profile that needs to be fulfilled before including a particular fly species as reasonably likely to act as a contributing factor of the spread of foodborne pathogens. These five attributes are synanthropy, endophily, communicative behavior, attraction to filth and human food, and the isolation of pathogens from wild populations [1, 2]. Other fly species fulfilling at least four of those attributes are considered opportunistic pests and their presence in food and/or food-related environments is an indication of insanitation [2].
Foodborne pathogens transmitted by synanthropic filth flies are found not only externally on the fly surface (which includes body, head, legs, and wings), but also internally, mainly in the alimentary canal (which runs the length of the body, from pharynx to anus) [6]. In fact, we previously reported that foodborne pathogens were up to three times more likely to be found in the alimentary canal than on the body surface of wild flies caught in and around urban restaurant dumpsters [7]. Consequently, flies can contaminate food or food-contact surfaces mechanically or through regurgitation or defecation. The potential spread of foodborne pathogens increases when there is a focus of infection for a particular bacterium [8]. Our previous study showed a statistically significant association between the presence of Salmonella enterica, Listeria monocytogenes and Cronobacter spp. (former Enterobacter sakazakii) on the surface and in the guts of wild flies and the sites where those flies were collected [7]; thus, emphasizing that bacteria inhabiting the alimentary canal of flies are acquired from the surrounding environment. Filth flies also travel quickly and may move several miles [9]; therefore, they can rapidly intensify the risk of foodborne diseases by transporting pathogens from places where the pathogens pose no hazard to places where they do, such as food preparation areas [1].
The transmission process of a particular pathogen in populations of synanthropic filth flies determines the spread and persistence of that pathogen. Thus, information about the transmission dynamics of a particular pathogen within a fly population is essential to appropriately avoid the spread of foodborne diseases. It is important to note that understanding the epidemiology of an illness caused by a pathogen transmitted by flies, requires a deeper knowledge of the ecology, physiology, immunology, and genetics of the pathogen as well as the morphology, physiology, and behavior of the fly. Nevertheless, it is even more important to understand how pathogen and fly interact in a particular environment [10, 11].
Filth flies can be transient or definitive hosts of pathogens and, like vertebrates, they may be immune or susceptible to infection. Although flies can internally harbor foodborne bacteria, it is not well known if these pathogens are beneficial or harmful to them. However, flies have shown remarkable resilience to these pathogens. For instance, several species of Cronobacter have been isolated from the alimentary canal of several flies collected in the wild [7, 12–16] and have also shown to support the development of stable fly larvae in the absence of other microbes, by colonizing the alimentary canal of newly emerged flies [16]. Flies have also shown efficient and rapid responses to ingested Escherichia coli O157:H7 since excretion of this pathogen was observed 6 to 24 h after being ingested [17, 18].
There are plenty of studies reporting the mechanical transmission of foodborne pathogens by filth flies (some examples include [7, 19–22]) and there are other studies reporting the fate and the temporospatial distribution of ingested foodborne pathogens by flies [17, 18, 23–26]. However, there is little scientific information about the transmission dynamics of foodborne bacteria to the fly’s progeny after parental flies have ingested those pathogens. The objective of this study was to estimate the probability of transmission of four foodborne bacteria (S. enterica, C. sakazakii, E. coli O157:H7, and L. monocytogenes ) to the progeny of the common house fly, Musca domestica (Linneaus) (Diptera: Muscidae), after parental house flies were fed with food contaminated with low, medium, and high levels of each bacterium. The presence of each pathogen was evaluated on pooled house fly eggs laid by parental females and on the surface and in the alimentary canal of newly emerged first filial (F1) generation adults.
Results and discussion
All parental house flies used in our experiments were observed feeding from contaminated food and the presence of each pathogen was confirmed from all alimentary canals dissected from randomly selected parental females. Although the focus of this study was not to evaluate the dynamics of the parental population of adult house flies, anecdotal evidence suggests that the feeding and mating behaviors were not influenced by the ingestion of bacteria, and although not measured, we did not observe apparent increases in the mortality of parental flies or reductions in their oviposition rate, when compared to control groups. We observed clusters of house fly eggs on the oviposition substrate approximately 10-16 h after they were placed on the mesh of all jars.
The combined molecular and culture approach that we used to detect and isolate the foodborne pathogens from samples of pooled house fly eggs, and the surfaces and alimentary canals of single adult house flies was straightforward when evaluating for the presence of S. enterica, E. coli O157:H7, and L. monocytogenes. We easily obtained pure colonies of these three pathogens from the enrichment media of all PCR-positive samples. Likewise, we easily obtained pure C. sakazakii colonies from the enrichment media of all PCR-positives from pooled house fly eggs and alimentary canals of parental flies. However, the isolation of this pathogen from PCR-positive samples from F1 adults was more challenging and required several subculturing steps on selective media. Consequently, we could only obtain pure colonies of C. sakazakii from eight out of 15 PCR-positive samples from F1 adults.
We have previously reported that C. sakazakii colonies could not be recovered from some PCR-positive samples while using this combined approach, likely due to the PCR being positive when other closely related bacterial genera, such as Citrobacter freundii, are present in the samples [7, 27]. Additionally, besides C. sakazakii a number of other Enterobacteriaceae are α-glucosidase positive, therefore the co-isolation of those organisms from samples with highly complex microbiota (such as the fly’s alimentary canal) could lower the efficiency of recovery of C. sakazakii from the chromogenic media used [28]. As a result, only those samples from which pure C. sakazakii colonies were isolated, were considered positive for the presence of the pathogen and included for statistical analysis. No pathogens were observed on chromogenic media from any of the PCR-negative samples that were randomly selected.
Pure colonies of S. enterica, L. monocytogenes, and E. coli O157:H7 isolated from PCR-positive samples were confirmed to be identical to the strains ingested by parental house flies by showing indistinguishable PFGE profiles (see Additional file 1). Likewise, matching nucleotide sequences were obtained from pure colonies of C. sakazakii when performing nucleotide comparison of the amplified fragment (463 bp) of the cgcA C. sakazakii gene.
Probability of bacterial transmission to house fly eggs
Our study reports the probability of the presence of the target pathogen in a sample containing pooled house fly eggs laid by several females fed from contaminated food. This study does not attempt to report the transmission rate of individual eggs laid by one or several female flies. The stepwise selection model of the logistic regression analysis indicated that the predicted probability of the presence of bacteria in samples of pooled house fly eggs was associated with the type of foodborne pathogen and the level of bacterial contamination of the food given to parental house flies. However, there was not a significant interaction between these two variables; thus, the interaction was removed from the full model described in Eq. 1. The model fit statistics and the AUC value of 0.89 (excellent discrimination) shows that our data fit the model relatively well. Results from the analysis of the maximum likelihood estimates of the parameters included in the logistic regression model and the model fit statistics for house fly eggs are included in Additional file 2(A).
For all bacteria evaluated, there was a higher chance of the presence of the pathogen in samples with house fly eggs after parental flies received food containing medium levels of bacteria (Table 1A). In fact, when parental house flies received food containing medium bacterial loads, the pathogens were two and six times more likely to be present in the samples than when parental flies fed from food contaminated with high and low bacterial levels, respectively. Therefore, there was not a positive correlation between the levels of contaminated food given to parental flies and the presence of the pathogens in samples with pooled house fly eggs.
Table 1.
Odds ratios estimates of the presence of foodborne pathogens
| Foodborne pathogen | Bacterial levels in food | Fly’s body part | Odds ratio (95 % CL) | |
|---|---|---|---|---|
| A) House fly eggs | ||||
| Medium vs. high | 1.9 (0.5, 6.8) | |||
| Medium vs. low | 6.0 (1.7, 20.4) | |||
| High vs. low | 3.2 (1.1, 9.6) | |||
| C. sakazakii vs. S. enterica | 15.5 (2.9, 82.6) | |||
| C. sakazakii vs. E. coli O157:H7 | 5.7 (1.0, 31.3) | |||
| C. sakazakii vs. L. monocytogenes | 3.0 (0.5, 16.4) | |||
| L. monocytogenes vs. S. enterica | 5.2 (1.5, 18.7) | |||
| L. monocytogenes vs. E. coli O157:H7 | 1.9 (0.5, 7.2) | |||
| E. coli O157:H7 vs. S. enterica | 2.7 (0.8, 8.6) | |||
| B) F1 female adults | ||||
| S. enterica | Medium vs. high | 2.4 (1.7, 3.4) | ||
| Surface vs. alimentary canal | 2.4 (1.7, 3.4) | |||
| C. sakazakii | High vs. medium | 2.2 (1.3, 3.5) | ||
| Surface vs. alimentary canal | 2.4 (1.5, 3.8) | |||
(A) house fly eggs and (B) first filial (F1) generation adults
The transmission potential of ingested bacteria to the house fly progeny is a very complex process. Flies harbor many microorganisms (including human pathogens) in their alimentary canals and they require the ingestion of live bacteria for their development. However, feeding from contaminated food does not imply that flies will become infected themselves or that ingested pathogens will survive, proliferate, and/or invade the reproductive system to be transovarially transmitted to house fly eggs and to subsequent life stages or generations. House flies can fight ingested opportunistic invaders by using physical barriers (i.e. the type II peritrophic matrix of the midgut epithelium), physiological defenses (i.e. digestive processes: pH, and digestive enzymes such as lysozyme), and innate immune response (i.e. the secretion of antimicrobial peptides, AMPs, by the fat body) [23, 29]. House flies also carry symbiotic bacteria from one source to another and from one generation to the next [30]. Other studies have suggested that the presence of inherited symbiotic bacteria in insects increases the insect’s resistance to pathogens; thus, inherited symbionts may have important effects on the ecology and evolutionary dynamics of host-pathogen interactions [31–33]. For instance, symbiotic Klebsiella oxytoca has been associated with house fly eggs. This bacterium is deposited on the surface of the eggs, inducing female oviposition. However, when K. oxytoca is above the threshold abundance levels, it causes oviposition inhibition [34]. Nonetheless, the threshold levels of many other ingested bacteria that will trigger a particular defense mechanism (s) or particular behaviors in the house fly are not yet well known. Nayduch and Joyner [35] detected lysozyme protein in adult house flies that ingested 1.2×105 CFU/μl of Staphylococcus aureus, and in their life history stages (eggs, larval instars, and F1 adults), providing evidence that the digestive and defensive dual role of lysozymes was activated by the ingestion of high levels of these bacteria. These facts could help to explain the lower rates of contamination found in samples containing pooled house fly eggs laid by females that ingested high levels of contaminated food. However, more research is needed to determine the role of specific foodborne bacteria in house flies and the threshold levels that will trigger defense mechanisms or behaviors in these insects.
The highest rates of contamination of house fly eggs were observed when parental flies fed from food contaminated with C. sakazakii (Fig. 1). Percentages of contamination of 87, 98, and 96 % were observed after parental house flies fed from food containing low, medium, and high levels of C. sakazakii, respectively. This was followed by the ingestion of food contaminated with L. monocytogenes and E. coli O157:H7. The contamination rate of house fly eggs with S. enterica was lower than other pathogens: 30, 72, and 58 %, after parental house flies received food containing low, medium, and high levels of this pathogen, respectively (Fig. 1). Regardless of the level of contamination of the food given to parental house flies, C. sakazakii was 16, 6, and 3 times more likely to contaminate house fly eggs than S. enterica, E. coli O157:H7, and L. monocytogenes, respectively. Similarly, L. monocytogenes was 5 and 2 times more probable to contaminate house fly eggs than S. enterica and E. coli O157:H7, respectively (Table 1A).
Fig. 1.
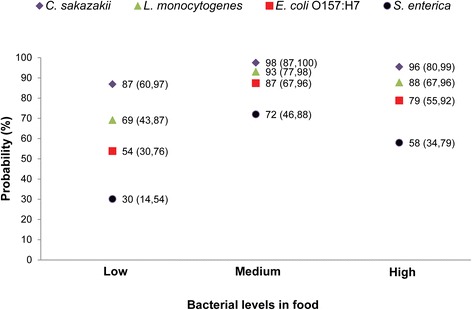
Probability of bacterial transmission to house fly eggs. Numbers in parenthesis represent lower and upper 95 % confidence limits (CL)
Although the groups of collected house fly eggs were surface-disinfected and we obtained no bacterial growth from aliquots of water from the last rinse of the surface-disinfection process, this only demonstrates that no more bacteria could be dislodged from the surface of the eggs (also known as chorion). To verify that bacterial cells were not adsorbed onto the surface of house fly eggs, we randomly selected several surface-disinfected eggs and used them either to take scanning electron microscopy (SEM) images or to plate them on the surface of chromogenic media specific for each pathogen. Even though SEM images of individual eggs did not reveal the attachment of bacterial cells to the chorion (see Additional file 3), we observed the presence of typical bacterial colonies surrounding some of the surface-disinfected house fly eggs that were individually plated. Thus, indicating that some bacterial cells remained attached to the chorion of surface-disinfected eggs.
Ingested bacteria could be adsorbed onto the surface of house fly eggs during or after oviposition because in female house flies the vaginal opening is in close proximity to the anal opening [36], which may facilitate contamination of the egg’s surface with waste products of the fly’s digestive tract. Bacterial cells could remain attached to the chorion due to the adhesive fluid that covers the eggs when they are laid. This fluid is secreted by the accessory glands of the female reproductive system and causes the eggs to adhere to each other and to the material where they were laid [37]. Additionally, the chorionic sculpture of house fly eggs has minute hexagonal markings, distinct curved rib-like thickenings (the hatching line), and some elevations and depressions [36, 37] that could hinder the dislodgement of bacterial cells during the surface-disinfection process.
We did not perform histological studies or transmission electron microscopy (TEM) to demonstrate the presence and/or possible development of the target bacteria in the internal tissues of the eggs. Thus, in this study we cannot confirm that the presence of pathogens in samples containing pooled house fly eggs was due to the transfer of bacteria at early stages of oogenesis and embryogenesis, as required during true transovarial transmission. Instead, the presence of pathogens in samples with pooled house fly eggs was probably due to the adsorption of bacterial cells onto the surface of the eggs during or after oviposition. Bacteria adsorbed on the surface of the eggs can proliferate in the larval rearing substrate and re-contaminate the hatching larvae, creating new focus of infection from where the newly hatched larvae can re-acquire the pathogen. In fact, random samples from larval rearing substrates taken the same day that pupae were removed from the rearing chambers evidenced the presence of the target pathogens (data not shown). Bacteria associated with house fly eggs have been found to supplement the rearing substrate of the developing larvae [38]. However, in this study we did not evaluate the presence of pathogens in any of the F1 larval stages. Future studies in our lab will assess the temporospatial fate of green fluorescent protein (GFP)-expressing S. enterica and/or C. sakazakii from individual eggs laid by female house flies fed with contaminated food. We will also evaluate the presence of the pathogen on the surface and internal tissues of the developing stages of the house fly (three larval instars, puparia, and newly-emerged adults) to have a better understanding of the trans-stadial transmission of those pathogens during metamorphosis.
Probability of bacterial transmission to house fly F1 adults
House fly F1 adults were observed in all treatments, indicating the successful completion of the house fly’s life cycle. No pathogens were detected from the surface or the alimentary canal of any of the adult specimens that were sampled from the control groups. Even though L. monocytogenes and E. coli O157:H7 were present in samples of pooled house fly eggs (Fig. 1), they were not detected from either the surface or the alimentary canal of any of the house fly F1 adults that were sampled. Therefore, no statistics were computed for these two pathogens when included in the model, because all observations had the same response.
We previously reported that L. monocytogenes was found in 3 % of wild filth flies [7] and later confirmed that isolated strains belonged to serotype 4b (unpublished data), responsible for most major outbreaks of human listeriosis [39]. However, studies providing evidence of L. monocytogenes being vectored by synanthropic filth flies are scarce. The innate immune response elicited by L. monocytogenes infections has shown that this bacterium is rapidly detected by the insect, inducing autophagy and inhibiting its intracellular growth to enhance insect survival [40, 41]. Additionally, L. monocytogenes are not restricted to localized tissues or specialized cells within the insect [42] and their release from the alimentary canal during metamorphosis may induce both localized and humoral insect immune responses [43], decreasing the overall bacterial population [44]. Thus, the absence of L. monocytogenes from house fly F1 adults was probably due to the flies’ innate immune response towards this foodborne pathogen. However, the ubiquitous abundance of L. monocytogenes in the environment, their ability to survive for long periods of time in acidic soils containing high endogenous microbiota [45], and their capability to attach to environmental surfaces and form biofilms [46] gives them the ability to create new focus of infection that can be used by filth flies to widely spread this pathogen.
Escherichia coli O157:H7 was also absent from house fly F1 generation adults. While some studies have demonstrated that house flies that ingested high E. coli O157:H7 concentrations (109 CFU/ml), retained this pathogen inside the alimentary canal for up to three days [18, 47], some others have reported that immune molecular effectors such as AMPs and lysozymes prevent the proliferation of this pathogen in the fly’s alimentary canal [17]. Thus, the question that E. coli O157:H7 is pathogenic to house flies needs to be further investigated. Although E. coli O157:H7 was present in samples containing pooled house fly eggs, this pathogen did not persist throughout metamorphosis. This finding was opposite to other studies that have reported the ingestion of non-pathogenic E. coli by house fly larva and their persistence throughout pupae and newly emerged adults [48, 49]. However, in this study we did not quantify the amount of E. coli O157:H7 present in the larval rearing substrate; hence, the levels of this pathogen that were likely to be ingested by house fly larvae were unknown and probably low enough to avoid their persistence through the house fly life cycle. Nevertheless, the association of synanthropic filth flies with E. coli O157:H7 is broadly supported [21, 50–54], strongly suggesting that house flies can indiscriminately disseminate this foodborne pathogen.
Salmonella enterica and C. sakazakii were the only pathogens present on F1 generation adults and only when parental house flies were given food contaminated with medium and high bacterial loads. The analysis of the maximum likelihood estimates of the parameters of this logistic regression model and the model fit statistics are included in Additional file 2(B). As shown by the model fit statistics and AUC values of 0.87 and 0.82 (excellent discrimination) for S. enterica and C. sakazakii, respectively, our data fit the model in Eq. 2 relatively well. The estimated probability of transmitting these pathogens to any single female adult fly from the F1 generation was associated with the bacterial concentration given to parental flies and the body part of the fly.
The presence of S. enterica and C. sakazakii was 2.4 times more likely on the body surface than in the alimentary canal of newly emerged F1 adults (Table 1B). This is in agreement with early studies performed by Radvan [55] who determined that some bacteria including Bacillus anthracis, B. subtilis, Shigella sonnei, and non-pathogenic E. coli were mainly located on the surface of recently emerged flies. This is probably due to the release of the intestinal content of the larvae into the pupal cavity, one of the changes that take place while the larvae re-organizes into an adult house fly [43, 56].
The probability of finding S. enterica on a single F1 adult house fly was greater than the probability of finding C. sakazakii (Fig. 2a, b). When parental flies received food with medium levels of S. enterica the probability of finding this pathogen on the surface and in the alimentary canal of a single F1 adult fly was 60 and 38 %, respectively. However, this probability decreased 38 and 20 % for the body surface and the alimentary canal, respectively, when parental flies fed from food with high S. enterica levels (Fig. 2a). Overall, it was 2.4 times more likely to find S. enterica on F1 adults after parental flies fed from food containing medium bacterial loads (Table 1B). Even though the presence of S. enterica in samples containing pooled house fly eggs was lower than other bacteria evaluated, this pathogen has developed strategies to deal with environmental changes brought on by the whole microbial community of a specific niche [57]. This could allow Salmonella to colonize the larval rearing substrate, be re-acquired by the developing larvae and persist through the adult stage.
Fig. 2.
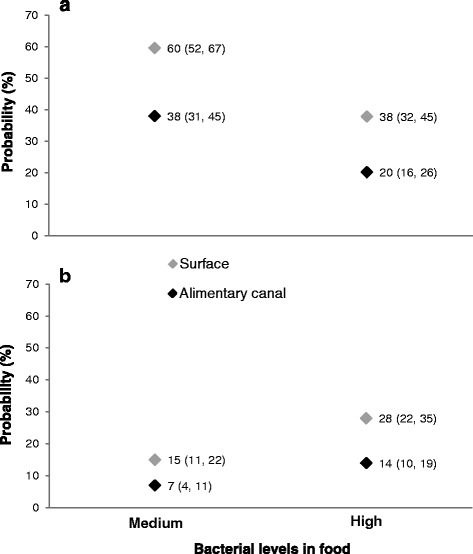
Probability of bacterial transmission to house fly first filial (F1) generation adults. a Salmonella enterica and (b) Cronobacter sakazakii. Numbers in parenthesis represent lower and upper 95 % confidence limits (CL)
Contrary to our findings with S. enterica, it was 2.2 times more likely to find C. sakazakii in a single F1 adult house fly after parental flies fed from food contaminated with high levels of this pathogen (Table 1B). The probabilities of finding C. sakazakii on the fly’s body surface and in the alimentary canal were 28 and 14 %, respectively, after parental flies fed from highly contaminated food (Fig. 2b). This probability decreased 15 and 7 % for the body surface and the alimentary canal, respectively, when parental flies fed from food contaminated with medium C. sakazakii levels. Thus, our results emphasize that pathogen concentration is an important parameter to determine the transmission of bacteria to the house fly progeny. Other authors have also stressed the significance of bacterial concentrations in the transmission of microorganisms since low bacterial inocula are insufficient to colonize the insect and the ingestion of excessive bacteria may be either pathogenic [48, 58, 59] or alter population dynamics or behavior [60].
Cronobacter sakazakii and S. enterica have probably evolved several mechanisms to evade the fly’s immune system. Bacteria that are associated with food can access the fly’s digestive tract and if they tolerate digestive processes and evade the immune system, they are able to access an environment that allows them to disseminate via regurgitation or defecation [32, 61]. Some ingested pathogenic bacteria can also produce a chronic infection in the host that makes it difficult to distinguish between a pathogenic or beneficial insect-microbe association [32, 62]. If C. sakazakii and S. enterica provide some benefit to synanthropic filth flies needs to be studied further. Examples of beneficial facultative symbionts by several arthropods include Serratia symbiotica in the pea aphid, Acyrthosiphon pisum (Hemiptera: Aphididae), which confers resistance against natural enemies such as parasitic wasps [63–65], and Hamiltonella defensa in whiteflies Bemisia tabaci (Hemiptera: Aleyrodidae) that increases the development and fitness of the host [66]. Consequently, the transmission mechanisms of both C. sakazakii and S. enterica need to be studied through more than one generation of flies to elucidate the type of associations these bacteria can potentially establish with these insects. Additionally, the interactions of these foodborne pathogens with other microorganisms present in flies need to be further explored. Understanding the type of associations that synanthropic flies establish with foodborne pathogens will help to elucidate transmission mechanisms as well as possible ways to mitigate the spread of foodborne pathogens.
It is important to mention that there is zero tolerance for the presence of S. enterica, C. sakazakii, E. coli O157:H7, or L. monocytogenes in foods. The mere presence of any of these four foodborne pathogens deems the food to be adulterated. Because the concentration of these pathogens in foods is usually not quantified, it is difficult to associate the three levels of contaminated food given to flies, to contamination concentrations of these pathogens in foods. Interestingly, this study demonstrated that adult house flies feeding from food contaminated with levels of bacteria as low as 100 cells/ml are able to transfer ingested pathogens to their progeny. Even though food can become contaminated at any point during production, the presence of pests, such as flies, increases the potential risk of pathogen transmission. Synanthropic filth flies that feed from any level of contaminated food are able to disseminate pathogens indiscriminately, not only mechanically or through regurgitation and defecation but also to their progeny, greatly increasing their vector potential.
To better protect public health, it is important to highlight the need for effective preventative measures that minimize the hazard posed by pests that may come in contact with food or food-contact surfaces and utensils. The implementation of pest control programs is one of the frequently and highly recommended measures to avoid the indirect transmission of foodborne pathogens by synanthropic insects like flies. The effectiveness of the program should be constantly monitored and filthy breading sites should be eliminated. By targeting control measures towards synanthropic filth flies, the potential transmission of foodborne pathogens can be interrupted, contributing to the prevention of future foodborne illness outbreaks.
Conclusion
In this study, we demonstrated that adult house flies that fed from food contaminated with low, medium, and high levels of S. enterica, C. sakazakii, E. coli O157:H7 or L. monocytogenes transmit these pathogens to their eggs. Salmonella enterica and C. sakazakii were further transmitted to F1 generation house fly adults, and they were more commonly found on the surface than in the alimentary canal of newly emerged house flies. Results from this research emphasize the public health significance and the regulatory importance of the presence of flies in food and food facilities.
Methods
House fly source
House fly (M. domestica) puparia were obtained from Spider Pharm, Inc. (Yarnell, AZ) and placed in plastic cages inside a Percival growth chamber at 30 °C and 16:8 h light:dark (L:D) photoperiod until eclosion. Emerged house flies were fed with a dry mixture of 1:1 granulated sugar and powdered milk. Cotton balls soaked in autoclaved water were also provided as a water source. Adult house flies (2-4 days old) were immobilized by placing the plastic cages at -30 °C for 5-7 min. Groups of approximately 40 adults (mixed sex) were transferred to autoclaved wide-mouth quart Mason glass jars. A disinfected 6-inch square piece of fiberglass window screen (New York Wire, Hanover, PA) was placed on top of each jar and secured with a rubber band. All glass jars were kept in the Percival growth chamber under the same conditions described above.
Preparation of contaminated food
Four bacterial foodborne pathogens (S. enterica, C. sakazakii, E. coli O157:H7, and L. monocytogenes) were used in our study. Information about bacterial strains, serotypes, and their origin is specified in Additional file 4. Bacterial strains were reconstituted from 30 % glycerol stock cultures, plated on Trypticase Soy Agar (TSA; Oxoid, Cambridge, UK), and incubated at 37 °C overnight. Stock suspensions of each bacterium were prepared by scraping bacterial cells from overnight cultures and adding them to buffered peptone water (BPW; Difco, Becton, Dickinson and Company, Sparks, MD). The optical density of the stock suspension was measured at 600 nm (OD600) using a GENESYS™ 20 Spectrophotometer (Thermo Fisher Scientific, Rochester, NY), and the bacterial concentration was calculated assuming that 0.1 OD600 = 108 bacterial cells/ml [67, 68]. A known volume of the stock bacterial suspension was added to a known volume of liquid fly food (18 g of dried powdered milk, 4 g of sugar, 2 g of protein powder, and 200 ml sterile distilled water) to obtain final bacterial concentrations of 108, 104, and 102 CFU/ml of each foodborne pathogen.
Adult house fly feeding bioassay
For each pathogen, approximately ten ml of fly food with the corresponding level of bacteria was added to three autoclaved cotton balls that were previously placed in the base of a sterile 60 mm diameter Petri dish. Fly food with no bacteria was used to feed the control groups. Fly food was given to parental house flies by inverting the Petri dish onto the mesh screen on top of each glass jar (see Additional file 5(A)), replacing with the corresponding fresh food after 18-20 h. Jars were kept in the Percival growth chamber under the same conditions described before and adult house flies were allowed to mate and feed ad libitum for a total of 30-32 h. Although the level of bacterial contamination of the fly food provided to parental house flies was known, the amount of bacteria actually ingested by adult house flies was not quantified. Thus, fly food containing final bacterial concentrations of 108, 104, and 102 CFU/ml will be referred hereinafter as high, medium, and low, respectively. After completing the feeding time, the Petri dish and cotton balls were removed and the mesh screen was thoroughly cleaned and disinfected with 70 % ethanol before adding the oviposition substrate.
Collection of house fly eggs
To create an oviposition substrate, several pieces of dehydrated beef liver (approximately 1 cubic inch and hydrated overnight) were placed on top of the mesh screen of each glass jar and covered with the lid of a sterile Petri dish to prevent dehydration (see Additional file 5(B)). Once fly eggs were visible on the surface of the liver, the glass jars were removed from the Percival growth chamber and clusters of approximately 100 eggs (laid by several females) were carefully removed using autoclaved forceps. To remove microbiota from the outer surface of the eggs, each cluster of eggs was transferred to a two ml tube with 70 % ethanol for 1 min, then submersed in 0.05 % bleach for 1 min, and finally rinsed three times with autoclaved distilled water (see Additional file 5(C)). One-hundred μl aliquots of water from the last rinse were plated on chromogenic media specific for the target foodborne pathogen (see Additional file 4). Surface-disinfected house fly eggs were divided in two groups approximately equal in number (~40-50). To assess the presence of pathogens, the first group of pooled eggs was added to one ml of enrichment media specific for each bacterial pathogen and incubated accordingly (see Additional file 4). The second group of eggs was added to a larval rearing substrate and allowed to hatch and complete their life cycle to evaluate the presence of foodborne pathogens in adult house flies of the F1 generation.
Validation that parental house fly adults ingested bacteria
After eggs were collected, glass jars containing parental house flies were placed at -20 °C for 5-7 min until flies were immobilized. Immobilized flies were then transferred to a disposable Petri dish containing 70 % alcohol for 2 min. Using a dissecting scope, three adult female house flies were randomly sub-sampled per each glass jar (n = 48 per each foodborne bacterium) and individually transferred to an autoclaved two ml tube to be surface-disinfected and their alimentary canals dissected as described by Pava-Ripoll, et al. [27]. The alimentary canals of maternal house flies were individually evaluated for the presence of the target bacteria as described in sections below.
House fly F1 offspring rearing procedure
The larval rearing substrate was prepared by pre-mixing dry ingredients (1 cup of autoclaved alfalfa pellets, 1 cup of autoclaved wheat bran, 1 cup of autoclaved bone meal, 1 cup of autoclaved poultry litter, 1/3 cup of dried milk powder, and 1 teaspoon of Brewer’s yeast) and adding 4 ½ cups of autoclaved tap water. Half cup of the prepared larval rearing substrate was added to individual plastic containers and then the group of surface-disinfected house fly eggs was added to the substrate using a disposable plastic pipette. The container was then nested in a larger plastic container that was approximately 1/8th filled with autoclaved sand to give fly larvae a dry place to pupate (see Additional file 5(D)). The rearing chambers were covered with an autoclaved paper towel, secured with a rubber band and placed in a Percival growth chamber at 32-35 °C and 16:8 h L:D photoperiod until pupation (approximately 4-5 days; see Additional file 5(E)). Using a disposable 1000 μl sterile pipette tip the larval substrate was gently mixed every day to inhibit mold growth. House fly pupae from each rearing chamber were carefully separated from the sand using sterile forceps and transferred to an extra-deep sterile, disposable Petri dish (Fisherbrand, Thermo Fisher Scientific, Rochester, NY) to allow F1 adults to emerge avoiding cross-contamination with the larval rearing substrate (see Additional file 5(F)). Petri dishes containing pupae were kept in the Percival growth chamber under same conditions until emergence of F1 generation adults (approximately 2-3 additional days; see Additional file 5(G)).
Collection of female F1 generation house fly adults
Recently emerged (0-1 days old) F1 adults were immobilized by placing extra-deep Petri dishes at -20 °C for 5-7 min. Under a dissecting scope, three females were randomly sub-sampled per each Petri dish (n = 48 per each foodborne bacterium) and individually transferred to autoclaved two ml tubes containing one ml of enrichment media specific for the target pathogen (see Additional file 4) to collect microbiota from the surface of the newly emerged house fly. Each house fly was then removed from the enrichment media, surface-disinfected and their alimentary canals aseptically dissected as described by Pava-Ripoll, et al. [27]. Tubes with enrichment media containing microbiota from the surface (s) and the alimentary canal (ac) of each F1 adult house fly were incubated at times and temperatures recommended for each bacterial pathogen (see Additional file 4).
Detection and isolation of the target bacteria
Enriched samples were assessed for the presence/absence of the target bacteria using a combined molecular and culture approach. The molecular approach was performed using a commercial PCR cycler/detector system (BAX® System Q7, DuPont Qualicon, Wilmington, DE) and assay kits specific for each bacteria (see Additional file 4) following manufacturer’s instructions and as described by Pava-Ripoll, et al. [27]. Each assay kit contains PCR-ready tablets with an intercalating dye that emits a fluorescence signal when binding to the target double-stranded DNA. The signal is detected by the PCR system and interpreted by the software as positive or negative. The culture approach was performed by plating ten μl of the enrichment media of PCR-positive samples on chromogenic media specific for each bacterium (see Additional file 4) until pure colonies were obtained. The culture approach was performed to confirm that isolated pathogens were the same strains given to parental house flies. Isolated S. enterica, L. monocytogenes, and E. coli O157:H7 were confirmed through pulsed-field gel electrophoresis (PFGE), following the protocols described by PulseNet and only using primary enzyme restriction [69, 70]. Isolated C. sakazakii was confirmed by polymerase chain reaction (PCR) amplification of the diguanylate cyclase (cgcA) gene using primers Cmstu-825 F and Csak-1317R as described by Carter, et al. [71]. Amplicons of expected size (463 bp) of the singleton PCR reaction were purified and sequenced by Retrogen, Inc. (San Diego, CA) and sequence files were imported into Sequencher 5.0 (GeneCodes, Ann Arbor, MI) to be processed and assembled. Contigs were exported and aligned using the CLUSTALX software (Lasergene, Madison, WI) and aligned sequences were used to generate a variance table report (Sequencher 5.0) where nucleotide bases of each sequence were compared to the reference C. sakazakii sequence. Four to five randomly selected PCR-negative samples were also plated on specific chromogenic media to confirm the absence of the target pathogen.
Experimental design
This experiment was set up as a completely randomized design and was performed at four different times with a one-month interlude. One foodborne pathogen (S. enterica, C. sakazakii, E. coli O157:H7, or L. monocytogenes) and fly food with three levels of bacterial contamination (high, medium, and low) plus a control, consisting of fly food with no bacteria, were evaluated each time. Each treatment combination (foodborne pathogen by levels of contaminated food) was replicated four times. Thus, 16 glass jars containing parental generation of adult flies were prepared each time the experiment was run. The presence/absence of the target bacterium was assessed as follows: a) from the alimentary canals of three parental females that were randomly sub-sampled per replicate (n = 48 per each foodborne pathogen); b) from pooled house fly eggs laid by several parental females (n = 16 per each foodborne pathogen); and c) from body surfaces and alimentary canals of three F1 female house flies that were randomly sub-sampled per replicate (n surface = 48 and n alimentary canal = 48 per each foodborne pathogen).
Statistical analysis
We used the SAS logistic regression procedure (PROC LOGIT; SAS Institute Inc., 2005) to predict the probability of bacterial contamination to house fly eggs and to the surface and the alimentary canal of F1 female adults. The presence/absence of foodborne pathogens was the categorical dichotomous response variable and its relationship with the predictor variables was analyzed using the two full logistic probability models described in Eq. 1 (for house fly eggs) and Eq. 2 (for F1 generation house fly adults).
| 1 |
| 2 |
Where logit (P) = ln [P/1-P], ln is the natural log, P is the probability of the presence of bacteria, β0 is the P intercept, βi are regression coefficients. The predictor variables for the probabilistic model of house fly eggs (Eq. 1) were the type foodborne pathogen, the level of bacterial contamination of the food given to parental house flies and their interaction. The predictor variables for the probabilistic model of F1 generation of house fly adults were the level of bacterial contamination of the food given to parental house flies and the fly’s body part (surface and alimentary canal) and the model was analyzed by each foodborne pathogen. The stepwise selection method with analysis of maximum likelihood estimates based on a Wald Chi-square p value <0.05 was used to determine the best probability model. The receiver operating characteristics (ROC) curve was used as a measurement of the goodness-of-fit of the model. The ROC curve quantifies the power of the predicted values using the area under the ROC curve (AUC). AUC values >0.7 are considered acceptable discrimination, >0.8 are considered excellent discrimination and >0.9 are considered outstanding discrimination [72].
Acknowledgments
Financial disclosure
The authors received no specific funding for this work.
Additional files
Pulsed-field gel electrophoresis (PFGE) profiles. The PFGE fingerprinting shows an indistinguishable pattern between the bacterial strains used to feed parental flies and the bacterial colonies isolated from the alimentary canal of parental flies (Pac), house fly eggs (e), and the surface (s) and alimentary canal (ac) of adult flies from the first filial (F1) generation. Profiles were obtained from (A) Salmonella enterica serotype Schwarzengrund (strain SAL3542; PFGE PulseNet pattern JM6X01.0289); (B) enterohemorrhagic Escherichia coli O157:H7 (strain ESC0786; PFGE PulseNet pattern EXHX01.0125); and (C) Listeria monocytogenes serotype 4b (strain LIS0150; PFGE PulseNet combined pattern GX6A16.0059_GX6A12.1652).
Analysis of Maximum Likelihood Estimates (MLE) of the logistic regression model. (A) house fly eggs and (B) house fly first filial (F1) generation of adults.
Scanning Electron Microscopy (SEM) of surface-disinfected house fly ( Musca domestica ) eggs (A) house fly egg; (B) the hatching line, with distinct curved rib-like thickenings; and (C) adhesive fluid on the egg surface.
Information about foodborne bacteria, culture media, incubation conditions, and PCR-based kits used in this study.
Experimental setup. (A) feeding of the parental population of house flies, (B) oviposition substrate, (C) collected house fly eggs, (D) surface-disinfected eggs placed in the larval rearing substrate, (E) house fly larval rearing container, (F) transfer of house fly pupae to plates, (G) emergence of first filial (F1) generation of house fly adults.
Footnotes
Competing interests
The authors certify that there is no competing interest with any financial organization regarding the materials discussed in this manuscript. The use of specified instrumentation is not an endorsement by the U.S. Food and Drug Administration.
Authors’ contributions
MPR and REGP conceived and designed the experiments. MPR, REGP, and AKM carried out laboratory work including feeding bioassays, rearing procedures, and detected and isolated bacteria from individual flies. MPR performed PCR analysis of C. sakazakii. BDT performed Scanning Electron Microscopy (SEM). CEK performed pulsed-field gel electrophoresis (PFGE) profiles of S. enterica, L. monocytogenes, and E. coli O157:H7. BDT and GCZ contributed by giving their point of view to the discussion of the results. MPR performed statistical analysis and wrote the manuscript. All authors read and approved the final manuscript.
Contributor Information
Monica Pava-Ripoll, Email: Monica.Pava-Ripoll@fda.hhs.gov.
Rachel E. Goeriz Pearson, Email: Rachel.Pearson@fda.hhs.gov.
Amy K. Miller, Email: Amy.Miller@fda.hhs.gov
Ben D. Tall, Email: Ben.Tall@fda.hhs.gov
Christine E. Keys, Email: Christine.Keys@fda.hhs.gov
George C. Ziobro, Email: George.Ziobro@fda.hhs.gov
References
- 1.Olsen AR. Regulatory action criteria for filth and other extraneous materials III. Review of flies and foodborne enteric disease. Regul Toxicol Pharmacol. 1998;28(3):199–211. doi: 10.1006/rtph.1998.1271. [DOI] [PubMed] [Google Scholar]
- 2.Olsen AR, Gecan JS, Ziobro GC, Bryce JR. Regulatory action criteria for filth and other extraneous materials V. Strategy for evaluating hazardous and nonhazardous filth. Regul Toxicol Pharmacol. 2001;33(3):363–92. doi: 10.1006/rtph.2001.1472. [DOI] [PubMed] [Google Scholar]
- 3.Emerson PM, Lindsay SW, Walraven GEL, Faal H, Bogh C, Lowe K, et al. Effect of fly control on trachoma and diarrhoea. Lancet. 1999;353(9162):1401–3. doi: 10.1016/S0140-6736(98)09158-2. [DOI] [PubMed] [Google Scholar]
- 4.Esrey SA, Potash JB, Roberts L, Shiff C. Effects of improved water-supply and sanitation on ascariasis, diarrhea, dracunculiasis, hookworm infection, schistosomiasis, and trachoma. Bull WHO. 1991;69(5):609–21. [PMC free article] [PubMed] [Google Scholar]
- 5.Cohen D, Green M, Block C, Slepon R, Ambar R, Wasserman SS, et al. Reduction of transmission of shigellosis by control of house flies (Musca domestica) Lancet. 1991;337(8748):993–7. doi: 10.1016/0140-6736(91)92657-N. [DOI] [PubMed] [Google Scholar]
- 6.Steinhaus EA. Insect microbiology. vol. November. New York: Hafner Publishing Co Ltd.; 1967.
- 7.Pava-Ripoll M, Pearson REG, Miller AK, Ziobro GC. Prevalence and relative risk of Cronobacter spp., Salmonella spp., and Listeria monocytogenes associated with the body surfaces and guts of individual filth flies. Appl Environ Microbiol. 2012;78(22):7891–902. doi: 10.1128/AEM.02195-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Greenberg B, Bornstein AA. Fly dispersion from rural Mexican slaughterhouse. Am J Trop Med Hyg. 1964;13(6):881. doi: 10.4269/ajtmh.1964.13.881. [DOI] [PubMed] [Google Scholar]
- 9.Savage EP. Disease Vector. In: Purdom PW, editor. Environ Health. London, U.K: Academic Pres; 1971. p. 48. [Google Scholar]
- 10.Gorham JR. Food, filth, and disease: A review. In: Foodborne disease handbook: Diseases caused by hazardous substances. Edited by Hui YH, Gorham JR, Murrell KD, Cliver DO, vol. 3: New York, USA: Marcel Dekker, Inc.; 1994: 627–634.
- 11.Reisen WK. Landscape epidemiology of vector-borne diseases. Annu Rev Entomol. 2010;55:461–83. doi: 10.1146/annurev-ento-112408-085419. [DOI] [PubMed] [Google Scholar]
- 12.Butler JF, Garcia-Maruniak A, Meek F, Maruniak JE. Wild Florida house flies (Musca domestica) as carriers of pathogenic bacteria. Fla Entomol. 2010;93(2):218–23. doi: 10.1653/024.093.0211. [DOI] [Google Scholar]
- 13.Gupta AK, Nayduch D, Verma P, Shah B, Ghate HV, Patole MS, et al. Phylogenetic characterization of bacteria in the gut of house flies (Musca domestica L.) FEMS Microbiol Ecol. 2012;79(3):581–93. doi: 10.1111/j.1574-6941.2011.01248.x. [DOI] [PubMed] [Google Scholar]
- 14.Hamilton JV, Lehane MJ, Braig HR. Isolation of Enterobacter sakazakii from midgut of Stomoxys calcitrans. Emerging Infect Dis. 2003;9(10):1355–6. doi: 10.3201/eid0910.030218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mramba F, Broce A, Zurek L. Isolation of Enterobacter sakazakii from stable flies, Stomoxys calcitrans L. (Diptera:Muscidae) J Food Prot. 2006;69(3):671–3. doi: 10.4315/0362-028x-69.3.671. [DOI] [PubMed] [Google Scholar]
- 16.Mramba F, Broce AB, Zurek L. Vector competence of stable flies, Stomoxys calcitrans L. (Diptera:Muscidae), for Enterobacter sakazakii. J Vector Ecol. 2007;32(1):134–9. doi: 10.3376/1081-1710(2007)32[134:VCOSFS]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 17.Fleming A. Spatial and temporal immune response in house flies in response to ingestion of Bacillus cereus and Eschericha coli O57:H7. Electronic Theses & Dissertations: Georgia Southern University; 2012. [Google Scholar]
- 18.Kobayashi M, Sasaki T, Saito N, Tamura K, Suzuki K, Watanabe H, et al. Houseflies: Not simple mechanical vectors of enterohemorrhagic Escherichia coli O157:H7. Am J Trop Med Hyg. 1999;61(4):625–9. doi: 10.4269/ajtmh.1999.61.625. [DOI] [PubMed] [Google Scholar]
- 19.Bidawid SP, Edeson JFB, Ibrahim J, Matossian RM. Role of non-biting flies in transmission of enteric pathogens (Salmonella species and Shigella species) in Beirut, Lebanon. Ann Trop Med Parasitol. 1978;72(2):117–21. doi: 10.1080/00034983.1978.11719291. [DOI] [PubMed] [Google Scholar]
- 20.Mian LS, Maag H, Tacal JV. Isolation of Salmonella from muscoid flies at commercial animal establishments in San Bernardino county, California. J Vector Ecol. 2002;27(1):82–5. [PubMed] [Google Scholar]
- 21.Moriya K, Fujibayashi T, Yoshihara T, Matsuda A, Sumi N, Umezaki N, et al. Verotoxin-producing Escherichia coli O157:H7 carried by the housefly in Japan. Med Vet Entomol. 1999;13(2):214–6. doi: 10.1046/j.1365-2915.1999.00161.x. [DOI] [PubMed] [Google Scholar]
- 22.Olsen AR, Hammack TS. Isolation of Salmonella spp. from the housefly, Musca domestica L., and the dump fly, Hydrotaea aenescens (Wiedemann) (Diptera:Muscidae), at caged-layer houses. J Food Prot. 2000;63(7):958–60. doi: 10.4315/0362-028x-63.7.958. [DOI] [PubMed] [Google Scholar]
- 23.Fleming A, Kumar HV, Joyner C, Reynolds A, Nayduch D. Temporospatial fate of bacteria and immune effector expression in house flies fed GFP-Escherichia coli O157:H7. Med Vet Entomol. 2014;28(4):364–71. doi: 10.1111/mve.12056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Olafson PU, Lohmeyer KH, Edrington TS, Loneragan GH. Survival and Fate of Salmonella enterica serovar Montevideo in Adult Horn Flies (Diptera: Muscidae) J Med Entomol. 2014;51(5):993–1001. doi: 10.1603/ME13217. [DOI] [PubMed] [Google Scholar]
- 25.Joyner C, Mills MK, Nayduch D. Pseudomonas aeruginosa in Musca domestica L.: Temporospatial examination of bacteria population dynamics and house fly antimicrobial responses. Plos One. 2013, 8(11):e79224. [DOI] [PMC free article] [PubMed]
- 26.Nayduch D, Cho H, Joyner C. Staphylococcus aureus in the House Fly: temporospatial fate of bacteria and expression of the antimicrobial peptide defensin. J Med Entomol. 2013;50(1):171–8. doi: 10.1603/ME12189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pava-Ripoll M, Pearson REG, Miller AK, Ziobro GC. Detection of foodborne bacterial pathogens from individual filth flies. J Vis Exp. 2015;96:e52372. doi: 10.3791/52372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Iversen C, Forsythe SJ. Comparison of media for the isolation of Enterobacter sakazakii. Appl Environ Microbiol. 2007;73(1):48–52. doi: 10.1128/AEM.01562-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Brandt SM, Dionne MS, Khush RS, Pham LN, Vigdal TJ, Schneider DS. Secreted bacterial effectors and host-produced eiger/TNF drive death in a Salmonella-infected fruit fly. PLoS Biol. 2004;2(12):2067–75. doi: 10.1371/journal.pbio.0020418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kellner R. The role of microorganisms for eggs and progeny. In: Chemoecology of Insect Eggs and Egg Deposition. Edited by Hilker M, Meiners T: Oxford, UK: Blackwell Publishing; 2002: 149–164.
- 31.Brownlie JC, Johnson KN. Symbiont-mediated protection in insect hosts. Trends Microbiol. 2009;17(8):348–54. doi: 10.1016/j.tim.2009.05.005. [DOI] [PubMed] [Google Scholar]
- 32.Eleftherianos I, Atri J, Accetta J, Castillo JC. Endosymbiotic bacteria in insects: Guardians of the immune system? Front Physiol. 2013, 4(46):1–10. [DOI] [PMC free article] [PubMed]
- 33.Hurst GDD, Darby AC. The inherited microbiota of arthropods, and their importance in understanding resistance and immunity. In: Insect Infection and Immunity. Evolution, Ecology, and Mechanisms. Edited by Rolff J, Reynolds SE: Oxford, UK: Oxford University Press 2009: 119-135.
- 34.Lam K, Babor D, Duthie B, Babor EM, Moore M, Gries G. Proliferating bacterial symbionts on house fly eggs affect oviposition behaviour of adult flies. Anim Behav. 2007;74:81–92. doi: 10.1016/j.anbehav.2006.11.013. [DOI] [Google Scholar]
- 35.Nayduch D, Joyner C. Expression of lysozyme in the life history of the house Fly (Musca domestica L.) J Med Entomol. 2013;50(4):847–52. doi: 10.1603/ME12167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Leopold RA, Meola S, Degrugillier ME. The egg fertilization site within the house fly, Musca domestica (L.) (Diptera: Muscidae) Int J Insect Morphol Embryol. 1978;7(2):111–20. doi: 10.1016/0020-7322(78)90011-9. [DOI] [Google Scholar]
- 37.Hewitt CG. The housefly, its structure, habits, development, relation to disease and control: Cambridge, UK: Cambridge University Press; 1914.
- 38.Lam K, Geisreiter C, Gries G. Ovipositing female house flies provision offspring larvae with bacterial food. Entomol Exp Appl. 2009;133(3):292–5. doi: 10.1111/j.1570-7458.2009.00928.x. [DOI] [Google Scholar]
- 39.Pan YW, Breidt F, Kathariou S. Competition of Listeria monocytogenes Serotype 1/2a and 4b strains in mixed-culture biofilms. Appl Environ Microbiol. 2009;75(18):5846–52. doi: 10.1128/AEM.00816-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tindwa H, Patnaik BB, Kim DH, Mun S, Jo YH, Lee BL, et al. Cloning, characterization and effect of TmPGRP-LE gene silencing on survival of Tenebrio molitor against Listeria monocytogenes infection. Int J Mol Sci. 2013;14(11):22462–82. doi: 10.3390/ijms141122462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Yano T, Mita S, Ohmori H, Oshima Y, Fujimoto Y, Ueda R, et al. Autophagic control of Listeria through intracellular innate immune recognition in Drosophila. Nat Immunol. 2008;9(8):908–16. doi: 10.1038/ni.1634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kikuchi Y. Endosymbiotic bacteria in insects: their diversity and culturability. Microbes Environ. 2009;24(3):195–204. doi: 10.1264/jsme2.ME09140S. [DOI] [PubMed] [Google Scholar]
- 43.Regan JC, Brandão AS, Leitão AB, Mantas Dias AR, Sucena E, Jacinto A, et al. Steroid hormone signaling is essential to regulate innate immune cells and fight bacterial infection in Drosophila. PLoS Path. 2013;9(10):e1003720. doi: 10.1371/journal.ppat.1003720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Greenberg B. Persistence of bacteria in the developmental stages of the housefly. 3. Quantitative distribution in prepupae and pupae. Am J Trop Med Hyg. 1959;8(6):613–7. doi: 10.4269/ajtmh.1959.8.613. [DOI] [PubMed] [Google Scholar]
- 45.Locatelli A, Spor A, Jolivet C, Piveteau P, Hartmann A. Biotic and abiotic soil properties influence survival of Listeria monocytogenes in soil. Plos One. 2013, 8(10):e75969. [DOI] [PMC free article] [PubMed]
- 46.Lemon KP, Higgins DE, Kolter R. Flagellar motility is critical for Listeria monocytogenes biofilm formation. J Bacteriol. 2007;189(12):4418–24. doi: 10.1128/JB.01967-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sasaki T, Kobayashi M, Agui N. Epidemiological potential of excretion and regurgitation by Musca domestica (Diptera:Muscidae) in the dissemination of Escherichia coli O157:H7 to food. J Med Entomol. 2000;37(6):945–9. doi: 10.1603/0022-2585-37.6.945. [DOI] [PubMed] [Google Scholar]
- 48.Rochon K, Lysyk TJ, Selinger LB. Retention of Escherichia coli by house fly and stable fly (Diptera:Muscidae) during pupal metamorphosis and eclosion. J Med Entomol. 2005;42(3):397–403. doi: 10.1093/jmedent/42.3.397. [DOI] [PubMed] [Google Scholar]
- 49.Schuster GL, Donaldson JR, Buntyn JO, Duoss HA, Callaway TR, Carroll JA, et al. Use of bioluminescent Escherichia coli to determine retention during the life cycle of the Hhusefly, Musca domestica (Diptera: Muscidae, L) Foodborne Pathog Dis. 2013;10(5):442–7. doi: 10.1089/fpd.2012.1326. [DOI] [PubMed] [Google Scholar]
- 50.Ahmad A, Nagaraja TG, Zurek L. Transmission of Escherichia coli O157:H7 to cattle by house flies. Prev Vet Med. 2007;80(1):74–81. doi: 10.1016/j.prevetmed.2007.01.006. [DOI] [PubMed] [Google Scholar]
- 51.Alam MJ, Zurek L. Association of Escherichia coli O157:H7 with houseflies on a cattle farm. Appl Environ Microbiol. 2004;70(12):7578–80. doi: 10.1128/AEM.70.12.7578-7580.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Iwasa M, Makino S, Asakura H, Kobori H, Morimoto Y. Detection of Escherichia coli O157:H7 from Musca domestica (Diptera: Muscidae) at a cattle farm in Japan. J Med Entomol. 1999;36(1):108–12. doi: 10.1093/jmedent/36.1.108. [DOI] [PubMed] [Google Scholar]
- 53.Rahn K, Renwick SA, Johnson RP, Wilson JB, Clarke RC, Alves D, et al. Persistence of Escherichia coli O157:H7 in dairy cattle and the dairy farm environment. Epidemiol Infect. 1997;119(2):251–9. doi: 10.1017/S0950268897007929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Talley JL, Wayadande AC, Wasala LP, Gerry AC, Fletcher J, DeSilva U, et al. Association of Escherichia coli O157:H7 with filth flies (Muscidae and Calliphoridae) captured in leafy greens fields and experimental transmission of E. coli O157:H7 to spinach leaves by house flies (Diptera: Muscidae) J Food Prot. 2009;72(7):1547–52. doi: 10.4315/0362-028x-72.7.1547. [DOI] [PubMed] [Google Scholar]
- 55.Radvan R. Persistence of bacteria during development in flies. III. Localization of the bacteria and transmission after emergence of the fly. Folia Microbiol. 1960;5:149–56. doi: 10.1007/BF02931776. [DOI] [Google Scholar]
- 56.Radvan R. Persistence of bacteria during development in flies. I. Basic possibilities of survival. Folia Microbiol. 1960;5:50–6. doi: 10.1007/BF02930896. [DOI] [Google Scholar]
- 57.Anjam Khan CM. The dynamic interactions between Salmonella and the microbiota, within the challenging niche of the gastrointestinal tract. Int Sch Res Notices. 2014;2014(846049):1–23. [DOI] [PMC free article] [PubMed]
- 58.Hosokawa T, Kikuchi Y, Fukatsu T. How many symbionts are provided by mothers, acquired by offspring, and needed for successful vertical transmission in an obligate insect-bacterium mutualism? Mol Ecol. 2007;16(24):5316–25. doi: 10.1111/j.1365-294X.2007.03592.x. [DOI] [PubMed] [Google Scholar]
- 59.Min KT, Benzer S. Wolbachia, normally a symbiont of Drosophila, can be virulent, causing degeneration and early death. P Natl Acad Sci USA. 1997;94(20):10792–6. doi: 10.1073/pnas.94.20.10792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Goodacre SL, Martin OY. Modification of insect and Arachnid behaviours by vertically transmitted endosymbionts: Infections as drivers of behavioural change and evolutionary novelty. Insects. 2012;3(1):246–61. doi: 10.3390/insects3010246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Douglas AE, Francois CLMJ, Minto LB. Facultative ‘secondary’ bacterial symbionts and the nutrition of the pea aphid, Acyrthosiphon pisum. Physiol Entomol. 2006;31(3):262–9. doi: 10.1111/j.1365-3032.2006.00516.x. [DOI] [Google Scholar]
- 62.Mateos M, Castrezana SJ, Nankivell BJ, Estes AM, Markow TA, Moran NA. Heritable endosymbionts of Drosophila. Genetics. 2006;174(1):363–76. doi: 10.1534/genetics.106.058818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Oliver KM, Degnan PH, Burke GR, Moran NA. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu Rev Entomol. 2010;55:247–66. doi: 10.1146/annurev-ento-112408-085305. [DOI] [PubMed] [Google Scholar]
- 64.Oliver KM, Moran NA, Hunter MS. Costs and benefits of a superinfection of facultative symbionts in aphids. P Roy Soc B-Biol Sci. 2006;273(1591):1273–80. doi: 10.1098/rspb.2005.3436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Oliver KM, Russell JA, Moran NA, Hunter MS. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. P Natl Acad Sci USA. 2003;100(4):1803–7. doi: 10.1073/pnas.0335320100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Su Q, Oliver KM, Pan H, Jiao X, Liu B, Xie W, et al. Facultative symbiont Hamiltonella confers benefits to Bemisia tabaci (Hemiptera: Aleyrodidae), an invasive agricultural pest worldwide. Environ Entomol. 2013;42(6):1265–71. doi: 10.1603/EN13182. [DOI] [PubMed] [Google Scholar]
- 67.Li WR, Xie XB, Shi QS, Zeng HY, Ou-Yang YS, Chen YB. Antibacterial activity and mechanism of silver nanoparticles on Escherichia coli. Appl Microbiol Biotechnol. 2010;85(4):1115–22. doi: 10.1007/s00253-009-2159-5. [DOI] [PubMed] [Google Scholar]
- 68.Sondi I, Salopek-Sondi B. Silver nanoparticles as antimicrobial agent: a case study on E. coli as a model for Gram-negative bacteria. J Colloid Interface Sci. 2004;275(1):177–82. doi: 10.1016/j.jcis.2004.02.012. [DOI] [PubMed] [Google Scholar]
- 69.Halpin JL, Garrett NM, Ribot EM, Graves LM, Cooper KL. Re-evaluation, optimization, and multilaboratory validation of the PulseNet-standardized pulsed-field gel electrophoresis protocol for Listeria monocytogenes. Foodborne Pathog Dis. 2010;7(3):293–8. doi: 10.1089/fpd.2009.0394. [DOI] [PubMed] [Google Scholar]
- 70.Ribot EM, Fair MA, Gautom R, Cameron DN, Hunter SB, Swaminathan B, et al. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157: H7 Salmonella, and Shigella for PulseNet. Foodborne Pathog Dis. 2006;3(1):59–67. doi: 10.1089/fpd.2006.3.59. [DOI] [PubMed] [Google Scholar]
- 71.Carter L, Lindsey LA, Grim CJ, Sathyamoorthy V, Jarvis KG, Gopinath G, et al. Multiplex PCR assay targeting a diguanylate cyclase-encoding gene, cgcA, to differentiate species within the genus Cronobacter. Appl Environ Microbiol. 2013;79(2):734–7. doi: 10.1128/AEM.02898-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hosmer DW, Lemeshow S. Applied logistic regression. 2. New York: John Wiley and Sons; 2000. [Google Scholar]