

Abstract
To support management decisions, molecular characterization of data and geo-reference of incidence records of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) were combined with data on the biology and ecology of the pest to estimate its climatic suitability and potential spread at regional and global scale. A CLIMEX model was developed and used for the global prediction of current and future climate-induced changes in the distributional shifts of T. absoluta. Results revealed that temperature and moisture characterized T. absoluta population growth while the pest ability to survive the cold, hot, wet and dry stress conditions are the primary characteristics defining its range frontiers. Simulated irrigation also played an important role in the model optimization. Model predictions suggest that T. absoluta represents an important threat to Africa, Asia, Australia, Northern Europe, New Zealand, Russian Federation and the United States of America (USA). Under climate change context, future predictions on distribution of T. absoluta indicated that the invasive nature of this pest will result in significant crop losses in certain locations whereas some parts of Africa may witness diminution in ranges. The following scenarios may occur: 1) T. absoluta damage potential may upsurge moderately in areas of Africa where the pest currently exists; 2) a range diminution in temperate to Sahel region with moderate upsurge in damage potential; 3) a range expansion in tropical Africa with reasonable upsurge of damage potential. These possible outcomes could be explained by the fact that the continent is already warm, with the average temperature in majority of localities near the threshold temperatures for optimal development and survival of T. absoluta. Outputs from this study should be useful in helping decision-makers in their assessment of site-specific risks of invasion and spread of T. absoluta with a view to developing appropriate surveillance, phytosanitary measures and management strategies.
Introduction
Tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae), is native to South America [1] and has been recorded from Argentina, Bolivia, Brazil, Chile, Colombia, Ecuador, Paraguay, Peru, Uruguay and Venezuela. The pest is predominantly found in areas below 1000 m above sea level. It is a devastating pest of tomato, Solanum lycopersicum L. (Solanaceae). Additionally the pest has also been found to attack other cultivated and wild solanaceae, such as pepper, Capsicum annuum L.; eggplant, Solanum melongena L.; tobacco, Nicotiana tabacum L.; potato, Solanum tuberosum L. and black nightshade, Solanum nigrum [2]. Recently, the pest was reported to expand its host range to garden bean, Phaseolus vulgaris L. (Fabaceae) [2]. Outside its native areas of distribution the pest has been reported for the first time in Spain in 2006, from where it is believed to have spread to other European countries including Italy (2008), France (2008), Albania (2009), Bulgaria (2009), Portugal (2009), Netherlands (2009), United Kingdom (2009) and Serbia (2011). In the Middle East region, it is known to occur in Israel (2010), Iran (2010), and Turkey (2010) [3–8]. It was later reported in the Canary Islands in 2012 [9]. In northern Africa, T. absoluta was found in Algeria (2008), Morroco (2008), Egypt (2009), Libya (2009), and Tunisia (2009) [1], [3], [10]. In 2012, Ethiopia, Niger, Senegal, and Sudan, were the countries in sub-Saharan Africa (SSA) region where T. absoluta was first detected [11–14]. Currently, the pest is found in Tanzania, Uganda and Kenya (S. A. Mohamed et al., unpublished data). Detections in Kenya were confirmed by using both morphological character states and molecular tool for species delineation (DNA barcoding). The fast spread of the pest across Africa may possibly be enhanced by trade, porous borders and fragile nature of the phytosanitary infrastructure coupled with inadequate implementation of quarantine measures.
Modeling of pest distribution is an important technique available to biologists and ecologists to understand and predict the past, current or future presence of species based on ecological observations and/or laboratory data. Theories so far developed for modeling species distribution can roughly be categorized in two main groups: the first class that considers climate (either directly or indirectly) as the major determining factor through its interactions with the biology of specific species [15–17], and the second class, takes into consideration multi-species interactions such as competition and predation as being the paramount component for the species presence in a region [16].
The present study is based on the first school of thought; which is characterized by two methodological approaches; namely mechanistic and correlative [18–19]. The mechanistic approach is constructed on the translation of environmental conditions into biologically relevant metrics (e.g. potential duration of activity) to capture environmental sensitivities of survivorship and fecundity for predicting species incidence in a region [18]. This modeling framework does not directly match climate; it rather models the species response to climatic factors and results into a geospatial metrics that indicates where the climate is suitable for the species development, survival and establishment [18]. The ‘correlative’ approach employs statistical or machine-learning algorithms to known occurrence records of the species with digital layers of environmental variables [19] for predictions. It uses datasets and simple functions to describe the species’ response to climatic factors [19]. Commonly, presence/absence data or occurrence data only, from different localities, are sufficient for generating risk maps. Such approaches project climate similarity (with no reference to biological response to climate) and the results indicate how well the climate in an area matches the climate where the species is located [20].
In assessing the potential risks of invasion and spread of T. absoluta in different countries of Africa, CLIMEX (Hearne Scientific Software Pty Ltd, Australia) was selected. This modeling platform combines both mechanistic and correlative approaches and provides distribution/information maps that use different spatial scales to envision the potential pest spread and dispersal. The objective of the modeling exercise in this context is to support strategic decision-making by mapping out where T. absoluta might potentially establish. Based on this hypothesis it is possible to provide global information where cultivated crops of the family Solanaceae may be subjected to damage by T. absoluta. Moreover, the study aims at identifying climatic factors, which determine the likelihood of success or failure of the development of T. absoluta. It is believed that the outcome of this study should be useful to better inform decisions on policy implementation and pest management practices such as pest surveillance and the implementation of appropriate phytosanitary measures to manage the spread of T. absoluta in Africa and beyond.
Materials and Methods
Ethics Statement
The study was not undertaken in national park or any protected areas. The crop tomato (S. lycopersicon) and the invasive insect pest (T. absoluta) involved in this study are not endangered or protected species.
Molecular Identification of Samples from Kenya
Trap catches from Isiolo Kenya (00 19.536 N, 037 33.233 E, 1160 masl) were preserved in 95% ethanol for DNA extraction and voucher specimen from the same samples have been kept at icipe Arthropod Pathology Unit. DNA was extracted from the whole moth using the Isolate II Genomic DNA Kit (Bioline GmbH) as per manufacturer’s instructions. The extracted DNA was stored at– 20°C until required for amplification. PCR was carried out using universal primers, forward primer (LCO1490) 5’-GGTCAACAAATCATAAAGATATTGG-3’ and reverse primer (HCO2198) 5’-TAAACTTCAGGGTGACCAAAAAATCA-3’ [21] to amplify a 658 bp fragment of the COI gene. The PCR amplification was carried out in a 20 μl volume containing 1x reaction Buffer, 200 μm of dNTP mix, 0.4 pmol/μl of each primer, 2.5 mM, MgCl2, 1 unit Taq DNA polymerase (MyTaq, Bioline) and 1 ng DNA template. Standard cycling conditions of 5 min at 94°C, then 35 cycles of 30 s at 94°C, 1 min at annealing temperature of 45°C and 1 min at 72°C, followed by a final elongation step of 5 min at 72°C were used. The products were purified using Isolate II PCR and Gel purification kit (Bioline GmbH) according to manufacturer’s instructions and subsequently sequenced in both directions using ABI 3700 genetic analyzers. Sequences obtained were assembled and edited using Chromas version 2.13 (Technelysium Pty ltd, Queensland, Australia). Consensus sequences from both the forward and reverse strands were generated and were then queried through BLASTN in the GenBank database provided by the National Centre of Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/) for identification purposes and to check for similarity with organisms already identified. Furthermore, additional sequences were obtained from GenBank and a multiple alignment was done using ClustalX version 2.1 [22]. A phylogenetic tree was constructed using MEGA version 6.0 [23] through the neighbor-joining method with a bootstrap test of 1000 replications [24] using Kimura 2-parameter (K2P) distance model [25].
Tuta absoluta Occurrence Data and Geo-Referencing
A search for information related to occurrence/incidence of T. absoluta was carried out from June 2013 to December 2014 using Web of Science, Google, PubMed and MEDLINE. The keywords used during the search comprised of the following: T. absoluta, tomato leafminer; South American tomato pinworm; pepper (C. annuum; Solanaceae); eggplant (S. melongena; Solanaceae); tobacco (N. tabacum; Solanaceae); potato (S. tuberosum; Solanaceae), solanaceous weeds and garden bean (P. vulgaris; Fabales: Fabaceae).
Geo-referencing a locality was centered on assigning geographical coordinates to the recorded description of that locality. The coordinates were assigned to records by searching the name of the site in the databases using Google Earth v.6.1 (GOOGLE) and on the maps with ArcGIS v.9.2 (ESRI). The coordinates were measured in decimal degrees and WGS 1984 datum. The combination of the analysis from database related to spatial interpretation and the output represented as map data were organized in 8 fields: continent, country, location, latitude, longitude, citations, references and the status of the species with categories in conformity with the European and Mediterranean Plant Protection Organization (EPPO) standards [26]. This information was used to generate MS Excel (Microsoft Corporation) datasets with detailed descriptions of the point locations where T. absoluta has been reported. Tuta absoluta incidence data obtained from collaborators in Eretria, Niger, Sudan, and Tunisia were also included in the datasets. The geo-referenced datasets of T. absoluta occurrence were used to produce a map depicting the current distribution of T. absoluta.
CLIMEX Modeling Platform
CLIMEX modeling theory is based on the species approximation responses to temperature, moisture and sometimes day-lengths and diapause [27]. The conceptual framework is entrenched on the assumption that knowledge on occurrence of species in a particular location can guide the inference of its tolerance to prevailing environmental conditions [16], which can further aid in predicting potential establishment range. CLIMEX attempts to mimic the mechanisms that limit species geographical distributions and determine their seasonal phenology and to a small magnitude their relative abundance [16–17], [27–28]. The model is parameterized with environmental observations and eco-physiological information of the species growth, development and survival [16–17]. CLIMEX platform uses a database of meteorological variables to derive weekly and annual indices [17]. The potential for population growth is described by a weekly growth index (GIW), which is combined to generate the annual growth index (GIA). Weekly stress indices (cold, hot, wet, dry and their interactions) are related to define the ability of a population to survive extreme circumstances [27]. Growth indices were inspired from the Sprengel–Liebig law of the minimum and Shelford’s law of tolerance [29–30] while stress indices are used to illustrate the observed patterns of species population response to extreme conditions [23]. The overall climatic suitability of a particular location for a given species is provided by an Eco-climatic Index (EI), which combines the annual potential for population growth (GI A), the stresses (SI) that limit species survival and the factors limiting interacting between these stresses (SX) [17], [31].
| (1) |
Meteorological Databases
CLIMEX modeling platform inputs weekly data from the averages of the following variables: maximum and minimum temperatures, relative humidity and rainfall. The climate data are sourced from the global change community through the Climatic Research Unit (CRU) in Norwich UK, available at http://www.cru.uea.ac.uk/. This data bank is made of point location with records from about 3,000 meteorological stations worldwide as well as interpolated climatic grids [31–32]. Due to low-density weather stations for Africa in CLIMEX database, additional weather data for East Africa were extracted from the FAOCLIM agro-climatic database [33]. With the supplemented datasets inputted into CLIMEX, a higher spatial resolution (5 arc-minutes) database was produced and used for developing the potential climatic suitability of T. absoluta in East Africa.
Scenarios of Irrigation and Climate Change
CLIMEX platform has features for analyzing the potential implications of irrigation and climate change on the seasonal phenology and survival of species as well as their future potential geographical distribution. When using the irrigation function, it treats all regions equally and only affects the amount of additional simulated rainfall [31]. In calibrating CLIMEX, a seasonal irrigation with a ‘top-up’ amount of water to increase the effective rainfall to 3.6 mm per day option was selected. For climate change analysis, a rise of 1.5°C Africa wide temperature and 10% increase in rainfall from March 2—September 30 and 10% decrease in rainfall for the rest of the year, were considered.
Potential Distribution of T. absoluta using CLIMEX
The developed model was fitted using inductive and deductive procedures. Inductive approach guided in fitting climatic trauma functions that describe T. absoluta range boundaries and deductive procedure was applied to express parameter values based on experiments of the species feedbacks to climatic factors and phenological observations [7], [34–35]. Practically, the model parameters were repetitively adjusted and the function "Compare location", which describes the potential geographical distribution of species, as controlled by weather variables was subsequently run until the estimated potential T. absoluta range best coincided with the known distribution of South America [16]. Calibration of the population growth and survival parameters was manually and iteratively conducted. The exercise was guided by biological information such as developmental threshold temperatures and the species’ feedback to different levels of natural and adequate wetness parameters [7], [34–35].
Population Growth Parameters
Eight (8) parameters were used to define conditions suitable for T. absoluta population growth. Lower temperature threshold (DV0) and upper temperature threshold (DV3) captured the temperature optima and bounds for growth. Lower soil moisture (SM0) and upper soil moisture threshold (SM3) represent ideal wetness and constraint for growth. Primary values for these parameters were sourced from published data on T. absoluta developmental characteristics [3], [34]. The final values of population growth were obtained when stress functions were able to explain a greater proportion of EI. Degree-days per generation (PPD) was used to approximate the potential number of generations T. absoluta can complete per year at a given location under defined conditions.
Survival Parameters
After the calibration of population growth parameters, mortality variables were inductively included to narrow the predicted distribution of T. absoluta to concur with known distribution patterns of South America. Three parameters (lower developmental temperature threshold (DVCS), weekly temperature threshold cold stress (TTCS) and weekly rate of accumulation of cold stress (THCS) were applied to characterize mortality due to extreme cold that prevents the distribution of T. absoluta in certain regions of South America. Additionally, cold stress accumulation rate (DHCS) and weekly degree-day threshold for cold stress (DTCS) were applied complementarily for taking into account mortality resulting from prolonged periods of cold, which may temporally occur and constraint T. absoluta incidence in unfavorable season. Due to lack of data, weekly heat stress temperature threshold (TTHS) and heat stress accumulation rate (THHS), which are additional CLIMEX parameters for inferring mortality resulting from extreme heat that constraints the distribution of species were not used. To eliminate areas of South America where there is no incidence of T. absoluta, the wet stress soil moisture threshold (SMWS), weekly wet stress accumulation rate (HWS), dry soil moisture dry stress (SMDS) and weekly rate of accumulation of dry stress (HDS) pressure factors were calibrated.
Model Evaluation
For testing the predictive ability of the developed model, a procedure using independent data; meaning records from different continents, not considered during model parameters estimation was adopted [16]. The parameters used for developing the EI potential distribution map for South America that correctly fit the geo-reference incidence datasets in this region were applied to predict the potential distribution of T. absoluta in Europe and Mediterranean basin. The obtained map in Europe and vicinity was then compared with the known incidence point locations in the region. Similar procedure was performed for Africa to generate the predictive EI map of T. absoluta, which was matched with geo-reference location points obtained from field surveys in particular regions of the continent. Additionally, a quantitative point-by-point geographical evaluation was conducted for different locations around the world (Brazil, Egypt, Turkey and Bulgaria) to estimate the potential number of generations per year that T. absoluta could complete at natural conditions. The selection of the above four locations was based on the available historical field and experimental data on the number of generations per year of the pest [36–39].
Results
Tuta absoluta Molecular Identification
Four (4) individuals were sequenced and the sequences were deposited at the GenBank with accession numbers KP324752 and KP324753. The Blast search through the NCBI BLASTN linked all the four individuals to T. absoluta sample from Serbia of accession number JN417242, with an E-value of 0 and with a probability of 99% match. This confirmed the identity of the moth as T. absoluta, an alien pest to Kenya. The optimal phylogenetic tree with the sum of branch length = 0.96778729 is shown in Fig 1. The analysis involved 8 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated. There were a total of 579 positions in the final dataset. The Kenyan sample linked closely to the Bosnian, USA and Serbian samples while the sample from Tunisia and India occupied a different branch.
Fig 1. Phylogenetic tree of T. absoluta samples from Kenya and others obtained from GenBank.

Tuta absoluta Occurrence Data and Geo-Referencing
A total of 498 occurrence records of T. absoluta belonging to 4 continents were obtained from the literature search as follow: Africa (79), Asia (52), Europe (132) and South America (236). Fig 2 shows the current known geo-reference occurrence location points of T. absoluta in the world. Point locations of South America, which were used for model parameters estimation (Table 1), are made of records from Argentina (33), Brazil (74), Chile (63), Ecuador (2), Peru (71) and Venezuela (2).
Fig 2. Global known geo-reference location points of T. absoluta.
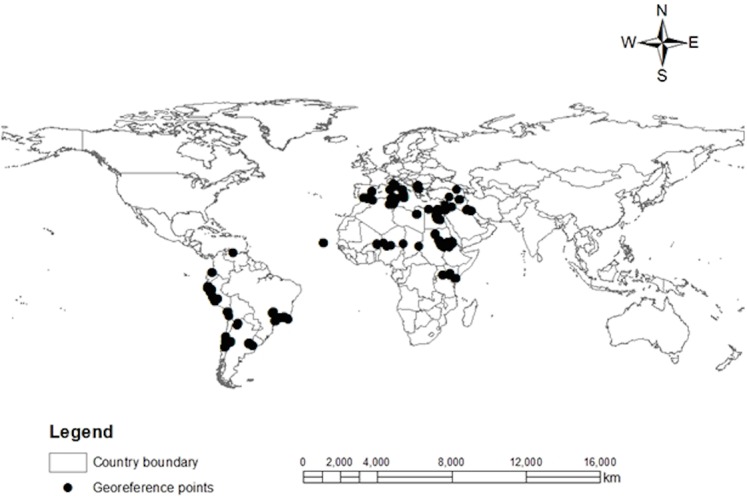
Point locations in South America correspond to its native distribution, and were obtained from the published literature. Point locations in Africa correspond to survey conducted by the authors of the article.
Table 1. Parameter values used to build a potential distribution for T. absoluta using CLIMEX modeling platform.
| Characteristic | Description—parameter | Value | Unit |
|---|---|---|---|
| Lower temperature threshold (DV0) | 8.0 | °C | |
| Lower optimum temperature (DV1) | 20 | °C | |
| Upper optimum temperature (DV2) | 30 | °C | |
| Upper temperature threshold (DV3) | 42 | °C | |
| Population growth | Lower soil moisture threshold (SM0) | 0.1 | SMC |
| Lower optimum soil moisture (SM1) | 0.4 | SMC | |
| Upper optimum soil moisture (SM2) | 1.5 | SMC | |
| Upper of soil moisture (SM3) | 4.0 | SMC | |
| Degree-days per generation (PDD) | 460 | °C days | |
| Lower developmental temperature threshold (DVCS) | 8.0 | °C | |
| Weekly temperature threshold cold stress (TTCS) | 3.0 | °C | |
| Weekly rate of accumulation of cold stress (THCS) | -0.001 | Week -1 | |
| Cold stress accumulation rate (DHCS)* | -0.0001 | Week -1 | |
| Survival | Weekly degree-day threshold for cold stress (DTCS)* | 15 | °C days |
| Weekly heat stress temperature threshold (TTHS) | - | °C | |
| Heat stress accumulation rate (THHS) | - | Week -1 | |
| Weekly wet stress soil moisture threshold (SMWS) | 2.0 | SMC | |
| Wet stress accumulation rate (HWS) | 0.002 | Week -1 | |
| Soil moisture dry stress (SMDS) | 0.1 | °C | |
| Weekly rate of accumulation of dry stress (HDS) | -0.01 | Week -1 |
* Alternative cold stress hypothesis
Population Growth Parameters
Table 1 contains the parameter values used to build the potential distribution of T. absoluta. DV0 was fixed to 8.0°C, as this was the minimum developmental threshold found in the literature [3], [40–41]. DV1 was fixed to 20°C and DV2 at 30°C. This range well encompasses the estimate of the optimum developmental threshold reported to be around the value of 25°C [37]. Many studies have reported the DV0 value for T. absoluta [41–42]; however, we have not encountered any study that showed the pest’s estimated value for DV3. Due to T. absoluta occurrence in cold temperate and warm tropical regions, we presumed that T. absoluta is highly adapted to wide range of temperatures, which offers the pest possibility to thrive under harsh conditions in Northern Sudan where daily temperatures constantly fluctuate and may occasionally reach up to 49°C in summer. Therefore, DV3 average weekly values was fixed to 42°C. Tuta absoluta may also be tolerant to dryness, requiring less water supply [3], [41], which may be the reason why it flourishes well in hot and dry areas. The lower moisture threshold was set to 0.1 and the optimum soil moisture at 4. PDD was allocated the value of 460-degree days as reported in literature [3], [41].
Survival Parameters
The values of survival parameters (Table 1) were adjusted such that the developmental cold stress (CS) temperature threshold was fixed to 8°C [3], [41]. TTCS was fixed to 3, denoting that T. absoluta can tolerate up to 3 cold days a week before it succumbs to CS [3] related damage. As the frequency and duration of coldness upsurge, the probability of damage in pest tissues increases. THCS was fine-tuned until the coldest known location where the pest has been reported was captured as being hardly suitable for this species persistence. CS was assumed to start accumulating at a rate of -0.0001 for DHCS per week with DTCS of 15°C days. Due to lack of data, mortality due to extreme heat; THHS and TTHS were not considered. WS was estimated to accumulate at a rate of 0.002 (HWS) per week with a value of 2 for SMWS, equivalent to the level of water stock aptitude. The average SMDS was set to 0.1, signifying that T. absoluta is sensitive to dry stress.
Potential Distribution Maps
CLIMEX model combines the growth and stress indices into an overall EI, ranging from 0 to 100 with the value of equal to 0 signifying unsuitable and EI of 100 representing optimal suitability for the year-round survival of a species [17]. The scaling for zones of suitability was made in accordance with the developed geo-reference map of T. absoluta incidence in South America. Four classes of EI were considered as shown in Fig 2: Class 1 (EI = (0–5)) indicates that the location is not suitable for T. absoluta survival. Class 2 (EI = (5–20)) represents zones with very moderate levels of suitability for the survival of T. absoluta. Class 3 (EI = (20–50)) indicates zones with high risk of establishment of T. absoluta. Class 4 (EI > 50) indicates the likelihood of long-term favorable zones that support the survival and establishment of T. absoluta.
Spatial predictions for South America locations within CLIMEX’s station database are shown in Fig 3. The predictions satisfactorily match the known distribution map for T. absoluta in the region and the geo-reference point locations belonging to class 3 and 4. Although the geo-reference records for Europe only include point location near the Mediterranean basin, T. absoluta potential distribution map in the region considers several countries with high risk for the pest invasion. By January 2015, T. absoluta has been reported in over 20 European countries [3], [39]; majority of which are in the southern, central and western parts of Europe. The present distribution map (Fig 4) showed that the northern countries (Belarus, Lithuania, Estonia, Denmark) might as well be at risk of T. absoluta invasion. A global projection (Fig 5) of the developed model indicated a high risk of invasion and establishment of T. absoluta in the world. From threshold developmental temperatures and degree-day accumulation, T. absoluta probably cannot survive winter conditions in the cold regions. However, transient populations may possibly survive in the field during the summer season and year round in greenhouses.
Fig 3. CLIMEX climatic suitability indices for T. absoluta in South America considered as the native region of the pest.
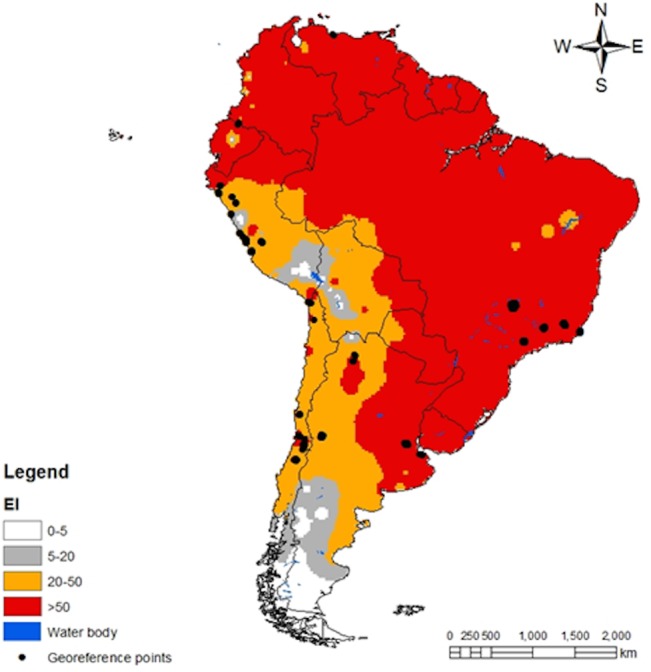
Predictions are based on the eco-climatic index (EI), a measure of climatic suitability scaled from 1–100, for locations within CLIMEX’s platform station database. EI = (0–5) location is not suitable; EI = (5–20) moderate level of suitability; EI = (20–50) high risk of establishment and EI > 50 very high likelihood of long-term survival. The black dotes are the geo-reference points obtained from literatures.
Fig 4. CLIMEX climatic suitability indices for T. absoluta in Europe.
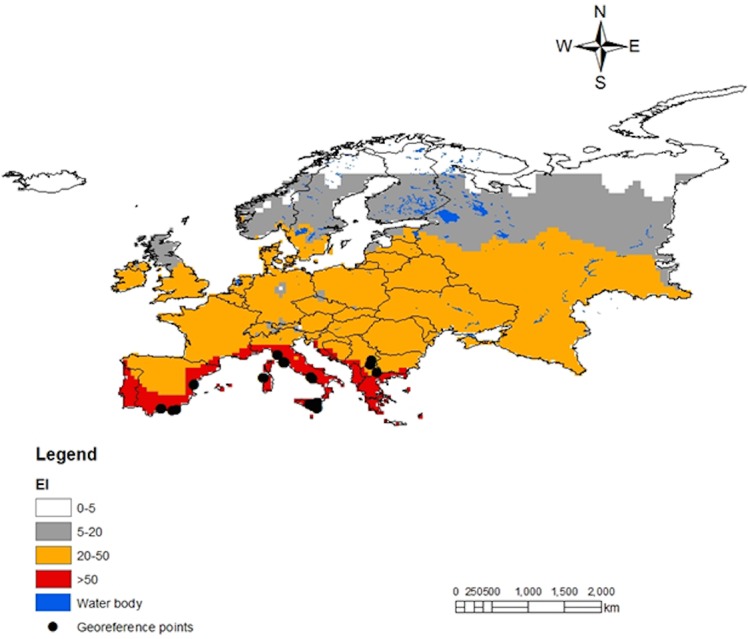
Predictionsare based on the eco-climatic index (EI), a measure of climatic suitability scaled from 1–100, for locations within CLIMEX’s platform station database. EI = (0–5) location is not suitable; EI = (5–20) moderate level of suitability; EI = (20–50) high risk of establishment and EI > 50 very high likelihood of long-term survival. The black dotes are the geo-reference points obtained from literatures.
Fig 5. CLIMEX climatic suitability indices for T. absoluta in the world.
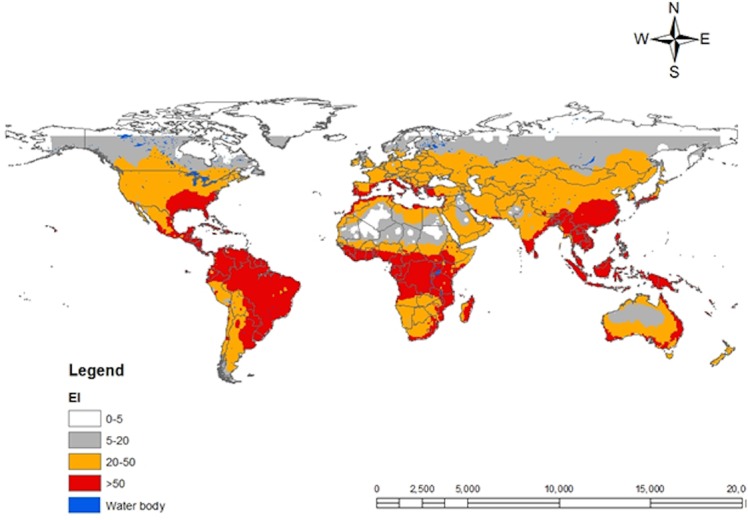
Predictions are based on the eco-climatic index (EI), a measure of climatic suitability scaled from 1–100, for locations within CLIMEX’s station database. EI = (0–5) location is not suitable; EI = (5–20) moderate level of suitability; EI = (20–50) high risk of establishment and EI > 50 very high likelihood of long-term survival. The black dotes are the geo-reference points obtained from literatures.
Tuta absoluta has been reported in many countries in the Middle East (Fig 4) and in Africa (Fig 6). The model predictions of the pest incidence in Africa reasonably match the current known distribution of T. absoluta. Warmer climates (similar to South America) that prevail in these regions may lead to soaring damage of the crops by this pest. The supplement datasets at higher spatial resolution of 5 arc-minutes from East Africa provided more accuracy in predictions for East Africa (Fig 7) as compared to the predictions for Africa (Fig 6). Referring to the classification of EI above, East Africa region has high probability for the pest to invade and naturalize.
Fig 6. CLIMEX climatic suitability indices for T. absoluta at in Africa.
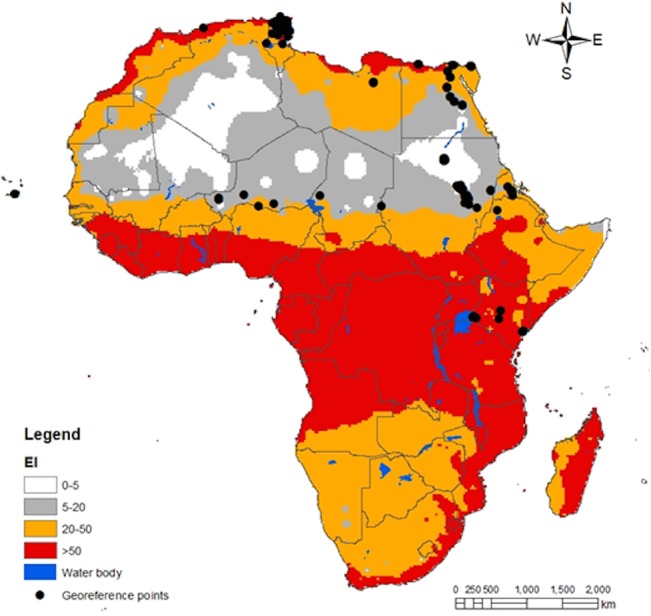
Predictions are based on the eco-climatic index (EI), a measure of climatic suitability scaled from 1–100, for locations within CLIMEX’s station database. EI = (0–5) location is not suitable; EI = (5–20) moderate level of suitability; EI = (20–50) high risk of establishment and EI > 50 very high likelihood of long-term survival. The black dotes are the geo-reference location points obtained from surveys.
Fig 7. CLIMEX climatic suitability indices for T. absoluta in East Africa.
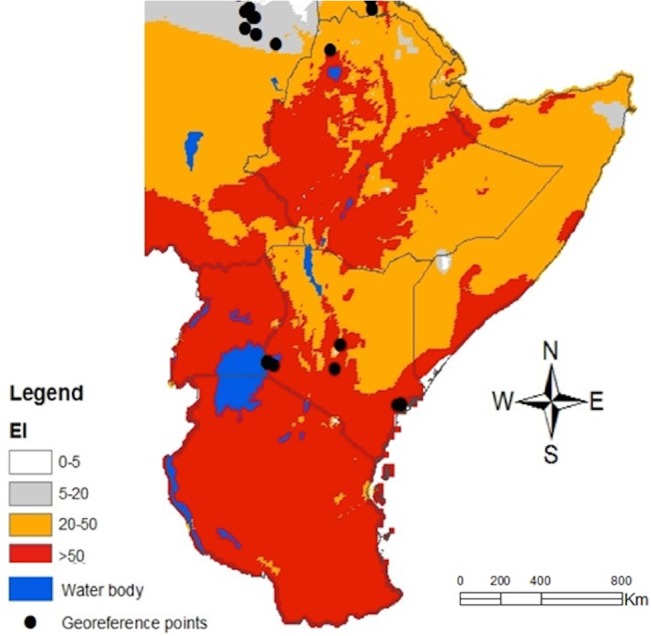
Predictions are based on the eco-climatic index (EI), a measure of climatic suitability scaled from 1–100, for locations within CLIMEX’s station database. EI = (0–5) location is not suitable; EI = (5–20) moderate level of suitability; EI = (20–50) high risk of establishment and EI > 50 very high likelihood of long-term survival. The black dotes are the geo-reference points obtained from surveys.
T. absoluta Potential Number of Generations per Year
Published and predicted data on the number of generations’ that T. absoluta can produce per year is shown in Table 1. The model prediction in different locations yielded the following number of generations per year of T. absoluta in different countries and localities: Brazil, Permambuco 10.1; Egypt, El-Behaira 11.6, Giza 12.3, Fayoum 11.4 and Qena 13.4 generations per year; Turky, Sanliurfa 4.9; Bulgaria, Pleven 4.8, Plovdiv 5.1, Sliven 4.9, Sofiya 3.9 and Varna 4.9. Empirical results [36–39] at these locations revealed similar number of generations per year as shown in Table 2.
Table 2. Number of T. absoluta generations produced per year in different locations around the world.
Comparison of the developed model outputs using CLIMEX and experimental results published in the literatures.
| Locality/Country | Number of generation per year | References | |
|---|---|---|---|
| Model | Experimental | ||
| Pernambuco (Brazil) | 10.1 | 9 | Vivan et al. 2002 |
| El-Behaira (Egypt) | 11.6 | 11 | Abolmaaty et al (2010) |
| Giza (Egypt) | 12.3 | 12 | Abolmaaty et al (2010) |
| Fayoum (Egypt) | 11.4 | 12 | Abolmaaty et al (2010) |
| Qena (Egypt) | 13.4 | 13 | Abolmaaty et al (2010) |
| Sanliurfa (Turky) | 4.9 | 4 | Mamay and Yanik (2012) |
| Pleven (Bulgaria) | 4.8 | 2–5 | Karadjova et al. (2013) |
| Plovdiv (Bulgaria) | 5.1 | 2–5 | Karadjova et al. (2013) |
| Sliven (Bulgaria) | 4.9 | 2–5 | Karadjova et al. (2013) |
| Sofiya (Bulgaria) | 3.9 | 2–5 | Karadjova et al. (2013) |
| Varna (Bulgaria) | 4.9 | 2–5 | Karadjova et al. (2013) |
Potential Irrigation and Climate Change Induced Shift for T. absoluta Distribution in Africa
Topping up the precipitation via irrigation to 3.6 mm per day in the growing season amplifies the climatic suitability for T. absoluta in several regions such as the African Sahel and therefore improves the model predictions. For example, EI values of dry regions like Niger (Maradi, Ouallam, Tohoua and Zinder) and Sudan (Blue Nile, Elabas, Gazira, Kassala, Khartoum) with no or low level of suitability become probable habitat for T. absoluta. Furthermore, these localities have recorded severe damage by the pest, proving their suitability as habitats of T. absoluta, despite their low EI values. Tuta absoluta would possibly show a wide annual population density fluctuation in these localities [17].
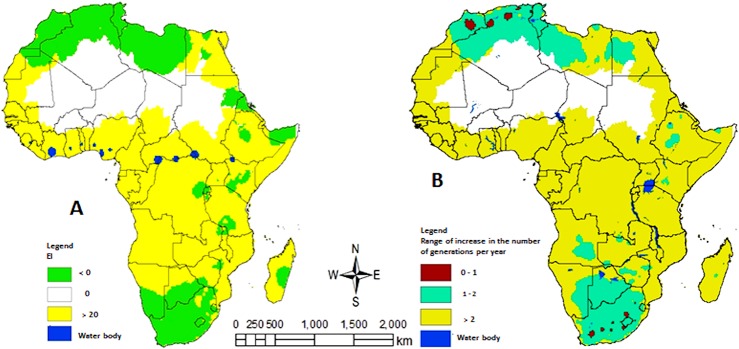
The map in Fig 8(A) shows the difference between the values of EI from predicted future distribution of T. absoluta (obtained when applying climate change criteria) and the distribution of the pest originated from the model inputs with climate datasets of the year 2000 in Africa. A value of EI = 0 demonstrates no range shift. A negative value of EI signifies a reduction of climatic suitability for T. absoluta to persist in a region, whereas a positive EI represents an upsurge in the probability of survival and permanent establishment of the pest. Based on the total number of degree-days above the lower threshold temperature for T. absoluta growth, 1 to 2.5 generations were estimated as the potential value of increase in number of generations under the selected climate change scenario (Fig 8(B)). The temperate zones (southern and northern) of Africa may, with time, reduce in level of climatic suitability for T. absoluta establishment while significant area of the continent (western, central and eastern) will be more favorable for its establishment consequently contributing to long-term survival of T. absoluta in the continent.
Fig 8. (A) Potential range shifts in the distribution of T. absoluta in Africa using the eco-climatic indices EI under climate change scenario (a rise of 1.5°C Africa wide temperature and 10% increase of rainfall from March 2—September 30 and 10% decrease in the rest of the year). The map was produced from the difference between the values of EI of the predicted future T. absoluta distribution (obtained when applying climate change criteria) and the distribution of the pest originated from current climate (year 2000) in Africa. EI = 0 demonstrates no range shift; EI < 0 signifies a reduction of climatic suitability EI > 0 represents an increase in the likelihood of survival and permanent establishment of the species. (B) Potential range of increase in number of generations per year of T. absoluta under the selected climate change scenario.
Discussion
Molecular Identification
A pre-requisite to any management strategies is proper identification of the pest species. Molecular tools have been utilized for species identification due to limitations of morphological features [44]. The current molecular tool of choice is DNA Barcoding, which is a system that employs sequence diversity in short, standardized gene regions aiding in identification of species [45]. This standardized method for identifications of species focuses sequencing efforts on one target gene, cytochrome c oxidase subunit I (COI) [44–46]. DNA barcoding successfully identified the moth’s sample as T. absoluta. The pest is most likely on a downward incursion from North Africa towards SSA.
Geo-Reference Information of T. absoluta
The information used to generate the worldwide distribution of T. absoluta contains numerous challenges; for instance, details of some records were given only at national and regional level making them unusable for geo-referencing exercise. Nevertheless, the produced T. absoluta geo-reference incidence and establishment map showed that the pest may be restricted to the 45° north and south latitudes of the temperate zones, which are above the borders of subtropical regions. However, the pest has been found without detailed knowledge of permanent establishment in certain countries marginally above this range (Ukraine, Russia [43] and United Kingdom [47]).
CLIMEX Model Match with Tuta absoluta Native and Invaded Distribution
Among the various procedures and tools, accessible for pest risk assessment [48–50], CLIMEX was favored because of its track record of being able to model the potential distribution of invasive species with some level of reliability and accuracy [51–54]. CLIMEX platform offers an easy option for its outputs to be directly exported to ArcGIS v.9.2 (ESRI) for standardized spatial analysis. The representation of the outputs became practical, showing T. absoluta distribution ranges, which facilitate the analysis of results for explicit conclusions [16]. Another advantage of using CLIMEX is the ability of the tool to provide useful predictions with minimum input information. This was demonstrated by using only 230 geo-reference records for the whole of South America to develop and fine-tune the model of T. absoluta that offers acceptable results globally. In addition, it is appropriate that a model of species prediction at continental level is verified using independent incidence data sets of the same species from different continents [16], [31]. CLIMEX offers features to easily conduct such evaluation; thus the value of parameters produced to predict T. absoluta incidences in South America were reused to project its potential invasion and establishment in Europe and other parts of the world.
Tuta absoluta is reported to be a multi-voltine species with a high reproductive potential [3], [42]. Larvae do not undergo diapause when food and suitable ecological conditions are available; the species can produce up to 12 generations per year [42]. The high reproductive capacity contributes to the invasive nature of T. absoluta, which offers plausible justification of the assessment of its potential for establishment and spread. This article complements previous efforts, which were only limited to Europe and the Mediterranean basin, of calibrating CLIMEX mechanistic platform [3], [39]. A major advantage in the present model is the formulation of new hypotheses, including irrigation as a potential vehicle for the spread and permanent establishment of T. absoluta in other areas. This factor that was not considered in previous studies [3], [39], was very useful in this study as it affects both the growth and survival of T. absoluta by enabling populations to persist and florish in season and region with suboptimal climatic conditions. In warm regions such as North Sudan, crops are predominantly under irrigated cultivation, which may have contributed to the successful propagation and potential establishment of the pest in this country. In addition, T. absoluta may be able to establish permanent populations inside greenhouses with controlled temperatures, if irrigated host plants are available throughout the year; consequently, providing adequate justification for the presence of the pest in some cold countries of Europe [3–8]. Simulation of irrigation considerably increased the extent to which T. absoluta could invade areas with little rainfall and where tomato fruits and Solanaceous plants are cultivated in Africa, Asia and Australia. By including irrigation module into our model, it enhanced the level of suitability of the Sahel, an important region of tomato production that is witnessing a wider range of incidence for T. absoluta. With this module, even the marginally suitable habitats in parts of the Middle East and Australia also became potential habitats for invasion and potential establishment of T. absoluta.
The present model further considers that T. absoluta might tolerate higher moisture, threshold temperatures and temperature summation needed for development. These assumptions make the potential distribution patterns of the species more accurate in capturing incidence data from South America. Projections in Europe and other areas suggested that countries located at approximately 60° north (Denmark, Estonia, Latvia, Norway, Sweden, sections of United Kingdom and the Russian Federation) are also at risk of invasion by T. absoluta. Such findings are in agreement with the literature that mentioned erratic outbreaks of T. absoluta in some of these countries [3], [8], [43–47]. Regional trade of agricultural products may be playing an important role in the dispersal of T. absoluta. It has also been reported that the adult pest is capable of flying between farms [3], [47] during summer and potentially finding other suitable plants such as garden Solanaceae as alternative hosts [3], [47]. This further explains the spontaneous occurrence of T. absoluta in some of the cultivated plants in greenhouses and open fields in northern Europe.
Globally, the potential of invasion by T. absoluta is very high: Asia, Middle East, New Zealand, United Stated of America and a large section of Australia present high areas of suitability, persistence and spread of the pest. It is suggested that cold winters and lack of soil moisture may be critical factors determining the survival of T. absoluta in a region. Dry stress limits the climatic suitability in low precipitation zones and, likely, cold stress prevented the permanent establishment of the pest in the following regions: northern parts of Europe; the Russian Federation and Canada. Records of T. absoluta occurrence in hot and dry areas like Zinder (Niger) and Khartoum (Sudan) in Africa suggest that this pest is extremely heat-tolerant. It could survive in areas with little annual rainfall as long as host plants are available.
Tuta absoluta as a Threat to Africa: Zooming to the Horn of the Continent
Surveys conducted in different parts of Africa showed that T. absoluta is considerably spreading very fast across the continent. The pest is currently reported in Algeria, Canary Island, Eritrea, Ethiopia, Egypt, Libya, Morocco, Niger, Senegal, Sudan [1], [3], [9–14] and in South Sudan, Kenya, Tanzania and Uganda (S.A. Mohamed et al., unpublished data). There is a high probability that T. absoluta is already in Somalia where it transited from Ethiopia and invaded Kenya. The potential distribution of this pest in Africa as observed from the developed model showed that it can invade and establish in most areas of the continent. This rapid spread may be attributed to the intensive cultivation and cross border trade of tomato fruits, which is the primary host of T. absoluta. In addition, the ecological and climatic conditions of Africa are similar to those of South America countries, the native region of the pest. According to published data, adult T. absoluta were found more than 6 miles (10 kilometers) away from tomato fields, suggesting that they can move long distances to colonize new areas by flying or drifting with the wind currents [3].
CLIMEX climate database only contains 720 stations for the whole Africa. Around the Sahara desert region, weather data is very scanty, probably justifying the failure of predictions to capture some localities where T. absoluta has been recorded. Due to such misrepresentation, supplementary datasets were added to generate a high-resolution predictive map for East African region. Zooming in, at countries of the horn of Africa showed that this region has a very high risk of invasion and permanent establishment of the pest. Tuta absoluta therefore, represents a significant threat to many horticultural crops considered as cash products in the region. The abundance of smallholder farmers may as well facilitate its rapid spread. Exercising vigilant phytosanitary inspections of imported plant germplasms and putting in place strict quarantine measures on exchange of planting materials and fresh harvest from endemic to non-endemic countries are recommended to curb the spread of this devastating pest.
Tuta absoluta Potential Number of Generation per Year
The model predictions for the potential establishment of T. absoluta were validated with known geo-referenced distribution data. However, it was found convenient to further evaluate the predictive ability of the model by estimating the potential number of generations that T. absoluta may develop in different localities of the world. The outcome of this second validation technique was based on the findings from the literatures [36–37]. Hence increasing the expectation of the predictive ability of the model developed for T. absoluta.
Change in Climate-Induced Distribution of Tuta absoluta in Africa
The International Panel for Climate Change (IPCC) has made a number of predictions for future climate change [55]. A warming of about 0.2°C per decade projected for the next two decades may possibly drive the expected range shifts of insects alongside the emergence of new pests in natural ecosystems which may change in response to altered temperature and precipitation profiles [56]. Africa is assumed to be the most vulnerable continent to climate change [55]; thus there is an urgent need to analyze possible impacts of climate change-induced changes on T. absoluta in this region. The present analysis and predictions on the future distribution of T. absoluta indicated that the invasiveness of this pest would be intensified and the number of the pest generation per year will increase, resulting in high yield losses in some locations. Depending on climatic zones (temperate, tropical, arid and semi-arid), the following possible scenarios may concurrently or individually occur: i) Tuta absoluta damage potential may progressively upsurge in regions of Africa where the pest already occurs; ii) a range diminution in temperate and semi-arid regions with moderate increase in damage potential; and iii) a range extension in tropical Africa with rationally high increase of the pest damage potential. It was reported [57] that the effects caused by global warming, precisely, temperature change on the metabolism of insects, are non-linear and may accelerate in warmer regions with little increase in temperature [57]. Such findings may be used to substantiate why future damage potential of T. absoluta is expected to rise in tropical than temperate Africa as predicted by the model. Another justification may be due to the fact that the continent is considerably warm, with the average temperature in many locations already near the threshold temperatures for optimum development and survival of the pest.
Acknowledgments
The authors thank the German Ministry of Economic Coperation and Development (BMZ), for financial support to the Tuta absoluta IPM project. Our thanks also go to Mohammedelnazeir Mahmoud, Ensaf S.I. Mohamed, Khaled Abbes, Adugna Haile (Eritrea), Mbaye Ndiaye, Jurgen Kroschel, Thomas Gachie and Jaris Veneros for providing geo-reference points for model development and validation.
Data Availability
Four individuals were sequenced and the sequences were deposited at the GenBank with accession numbers KP324752 and KP324753. The Blast search through the NCBI BLASTN linked all the four individuals to a T. absoluta sample from Serbia of accession number JN417242, with an E-value of 0 and with a probability of 99% match.
Funding Statement
The authors thank the German Ministry of Economic Cooperation and Development (BMZ) for financial support to the Tuta absoluta IPM project.
References
- 1. Cabello T, Gallego JR, Fernandez FJ, Gamez M, Vila E, Del Pino M, et al. Biological control strategies for the South American tomato moth (Lepidoptera: Gelechiidae) in greenhouse tomatoes. J Econ Entomol. 2012; 105: 2085–2096. [DOI] [PubMed] [Google Scholar]
- 2. Ferracini C, Ingegno BL, Navone P, Ferrari E, Mosti M, Tavella L, et al. Adaptation of indigenous larval parasitoids to Tuta absoluta (Lepidoptera: Gelechiidae) in Italy. J Econ Entomol. 2012; 105: 1311–1319. [DOI] [PubMed] [Google Scholar]
- 3. Desneux N, Wajnberg E, Wyckhuys KAG, Burgio G, Arpaia S, Narváez-Vasquez CA, et al. Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. J Pest Sci. 2010; 83: 197–215. 10.1007/s10340-010-0321-6 [DOI] [Google Scholar]
- 4. Desneux N, Luna MG, Guillemaud T, Urbaneja A. The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: the new threat to tomato world production. J Pest Sci. 2011; 84: 403–408. 10.1007/s10340-011-0398-6 [DOI] [Google Scholar]
- 5. DJurić Z, Delić D, Hrnčić S, Radonjić S. Distribution and molecular identification of Tuta absoluta (Meyrick, 1917)(Lepidoptera, Gelechiidae) populations in Bosnia and Herzegovina and Montenegro. Pol J Entomol. 2014; 83: 121–129. [Google Scholar]
- 6. Hrnčić S, Radonjić S. Tomato leafminer–Tuta absoluta Meyrick (Lepidoptera, Gelechiidae)–current status in Montenegro. EPPO Bull. 2012; 42: 341–343. [Google Scholar]
- 7. Lobos E, Occhionero M, Werenitzky D, Fernandez J, Gonzalez LM, Rodriguez C, et al. Optimization of a Trap for Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) and Trials to Determine the Effectiveness of Mass Trapping. Neotrop Entomol. 2013; 42: 448–457. 10.1007/s13744-013-0141-5 [DOI] [PubMed] [Google Scholar]
- 8.EPPO. EPPO Reporting Service. New data on quarantine pests and pests of the EPPO Alert List. 2014/015.
- 9. Polaszek A, Rugman-Jones PF, Stouthamer R, Hernandez-Suarez E, Cabello T, del Pino Pérez M. Molecular and morphological diagnoses of five species of Trichogramma: biological control agents of Chrysodeixis chalcites (Lepidoptera: Noctuidae) and Tuta absoluta (Lepidoptera: Gelechiidae) in the Canary Islands. BioControl. 2012; 57: 21–35. [Google Scholar]
- 10. Abbes K, Harbi A, Chermiti B. The tomato leafminer Tuta absoluta, (Meyrick) in Tunisia: current status and management strategies. EPPO Bull. 2012; 42: 226–233. [Google Scholar]
- 11. Pfeiffer DG, Muniappan R, Sall D, Diatta P, Diongue A, Dieng EO. First Record of Tuta absoluta (Lepidoptera: Gelechiidae) in Senegal. Fla Entomol. 2013; 96: 661–662. [Google Scholar]
- 12.Tomato leaf miner, Tuta absoluta,Tomato pest, potato pest,Tuta absoluta pheromone trap, potato,Aubergine, pest,Green house tomato pest,Tuta in Spain, Italy, France,Morocco,Algeria,Tunisia, Tuta absoluta on delta trap,water trap, pheromone trap,open field [Internet]. [cited 2 Jul 2014]. Available: http://www.tutaabsoluta.com/.
- 13. Mohamed ESI, Mohamed ME, Gamiel SA. First record of the tomato leafminer, Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae) in Sudan. EPPO Bull. 2012; 42: 325–327. [Google Scholar]
- 14. Brévault T, Sylla S, Diatte M, Bernadas G, Diarra K. Tuta absoluta Meyrick (Lepidoptera: Gelechiidae): a new threat to tomato production in sub-Saharan Africa: short communications. Afr Entomol. 2014; 22: 441–444. [Google Scholar]
- 15. Venette RC, Kriticos DJ, Magarey RD, Koch FH, Baker RH, Worner SP, et al. Pest risk maps for invasive alien species: a roadmap for improvement. BioScience. 2010; 60: 349–362. [Google Scholar]
- 16. Tonnang HEZ, Kangalawe RYM, Yanda PZ. Predicting and mapping malaria under climate change scenarios: the potential redistribution of malaria vectors in Africa. Malar J. 2010; 9: 111 10.1186/1475-2875-9-111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Sutherst RW. Prediction of species geographical ranges. J Biogeogr. 2003; 30: 805–816. 10.1046/j.1365-2699.2003.00861.x [DOI] [Google Scholar]
- 18. Miller J. Species Distribution Modeling. Geogr Compass. 2010; 4: 490–509. 10.1111/j.1749-8198.2010.00351.x [DOI] [Google Scholar]
- 19. Kearney MR, Wintle BA, Porter WP. Correlative and mechanistic models of species distribution provide congruent forecasts under climate change. Conserv Lett. 2010; 3: 203–213. 10.1111/j.1755-263X.2010.00097.x [DOI] [Google Scholar]
- 20. Rodda GH, Jarnevich CS, Reed RN. Challenges in Identifying Sites Climatically Matched to the Native Ranges of Animal Invaders. PLoS ONE. 2011; 6: e14670 10.1371/journal.pone.0014670 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994; 3: 294–299. [PubMed] [Google Scholar]
- 22. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research. 1997; 24: 4876–4882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S, MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution 2013, 30: 2725–2729. 10.1093/molbev/mst197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution. 1987; 4: 406–425. [DOI] [PubMed] [Google Scholar]
- 25. Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of molecular evolution. 1980; 16: 111–120. [DOI] [PubMed] [Google Scholar]
- 26. Normes EPPO Standards. EPPO Bull. 2014; 44: 3–4. 10.1111/epp.12088 [DOI] [Google Scholar]
- 27. Sutherst RW, Maywald GF. A computerised system for matching climates in ecology. Agric Ecosyst Environ. 1985; 13: 281–299. 10.1016/0167-8809(85)90016-7 [DOI] [Google Scholar]
- 28. Kriticos DJ, Morin L, Leriche A, Anderson RC, Caley P. Combining a Climatic Niche Model of an Invasive Fungus with Its Host Species Distributions to Identify Risks to Natural Assets: Puccinia psidii Sensu Lato in Australia. PLoS ONE. 2013; 8: e64479 10.1371/journal.pone.0064479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. The Ecology of North America Victor E. Shelford. University of Illinois Press, Urbana, 1963. 610 pp. Illus. [Internet]. [cited 2 Jul 2014]. Available: http://www.sciencemag.org/content/144/3617/401.2.citation. [Google Scholar]
- 30. Van der Ploeg RR, Bo¨hm W, Kirkham MB. On the Origin of the Theory of Mineral Nutrition of Plants and the Law of the Minimum. Soil Sci Soc Am J. /9; 63: 1055–1062. [Google Scholar]
- 31. Sutherst R W, Maywald G F, Kriticos D J. CLIMEX Version 3: User’s Guide. Melbourne, Australia: Hearne Scientific Software; Melbourne, Australia: Hearne Scientific Software; 2007: 131 p. [Google Scholar]
- 32. Kriticos DJ, Webber BL, Leriche A, Ota N, Macadam I, Bathols J, et al. CliMond: global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol Evol. 2012; 3: 53–64. [Google Scholar]
- 33.CLIMPAG | DATA and MAPS | FAOCLIM 2: world-wide agroclimatic data [Internet]. [cited 18 Jul 2014]. Available: http://www.fao.org/nr/climpag/pub/en1102_en.asp.
- 34. Miranda MMM, Picanço M, Zanuncio JC, Guedes RNC. Ecological Life Table of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Biocontrol Sci Technol. 1998; 8: 597–606. 10.1080/09583159830117 [DOI] [Google Scholar]
- 35. Spasov D, Spasova D, Atanasova B, Mitev V. Control of population number of tomato leaf miner tuta absoluta (Meyrick, 1917) in the Strumica region. Godisen Zb-2011. 2011; 79–87. [Google Scholar]
- 36. Vivan LM, Torres JB, Barros R, Veiga A. Tasa de crecimiento poblacional del chinche depredador Podisus nigrispinus (Heteroptera: Pentatomidae) y de la presa Tuta absoluta (Lepidoptera: Gelechiidae) en invernadero. Rev Biol Trop. 2002; 50: 145–154. [PubMed] [Google Scholar]
- 37. Abolmaaty SM, Khalil AA, Amna MH. Using degree–day unit accumulation to predict potato tuber worm incidence under climate change conditions in Egypt. Nat Sci. 2011; 9: 156–160. [Google Scholar]
- 38. MAMAY M, YANIK E. Şanlıurfa’da domates alanlarında Domates güvesi [Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae)]’nin ergin popülasyon gelişimi. Türk Entomol Bült. 2012; 2: 189–198. [Google Scholar]
- 39. Karadjova O, Ilieva Z, Krumov V, Petrova E, Ventsislavov V, others. Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae): potential for entry, establishment and spread in Bulgaria. Bulg J Agric Sci. 2013; 19: 563–571. [Google Scholar]
- 40.Erdoğan P, Babaroğlu NE. Life Table of the Tomato Leaf Miner, Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae). 2014; Available: http://earsiv.gop.edu.tr/xmlui/handle/123456789/175.
- 41. Barrientos Z R, Apablaza H J, Norero S A, Estay P P. Threshold temperature and thermal constant for the development of the South american tomato moth, Tuta absoluta (Lepidoptera: Gelechiidae) Cienc E Investig Agrar Chile; 1998; Available: http://agris.fao.org/agris-search/search.do?RecordID=CL2000000279. [Google Scholar]
- 42.Cherif A, Mansour R, Grissa-Lebdi K. Biological aspects of tomato leafminer Tuta absoluta (Lepidoptera: Gelechiidae) in conditions of Northeastern Tunisia: possible implications for pest management. Available: http://eeb.lu.lv/EEB/201312/EEB_11_Cherif.pdf.
- 43.Tuta absoluta. [Distribution map]. [Internet]. [cited 17 Jul 2014]. Available: http://www.cabi.org/isc/abstract/20143031650.
- 44. Hebert PD, Cywinska A, Ball SL, others. Biological identifications through DNA barcodes. Proc R Soc Lond B Biol Sci. 2003; 270: 313–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Hebert PD, Ratnasingham S, de Waard JR. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc R Soc Lond B Biol Sci. 2003; 270: S96–S99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Ratnasingham S, Hebert PD. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol Ecol Notes. 2007; 7: 355–364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Cuthbertson AG, Mathers JJ, Blackburn LF, Korycinska A, Luo W, Jacobson RJ, et al. Population Development of Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae) under Simulated UK Glasshouse Conditions. 2013; Available: http://citeseerx.ist.psu.edu/viewdoc/summary? doi: = 10.1.1.361.5061 [DOI] [PMC free article] [PubMed]
- 48.Phillips SJ, Dudík M, Schapire RE. A maximum entropy approach to species distribution modeling. Proceedings of the twenty-first international conference on Machine learning. ACM; 2004. p. 83. Available: http://dl.acm.org/citation.cfm?id=1015412.
- 49. Sporleder M, Tonnang HEZ, Carhuapoma P, Gonzales JC, Juarez H, Kroschel J, et al. Insect Life Cycle Modelling (ILCYM) software-a new tool for regional and global insect pest risk assessments under current and future climate change scenarios. Potential Invasive Pests Agric Crops. 2013; 412–427. [Google Scholar]
- 50. Elith J, Leathwick JR. Species distribution models: ecological explanation and prediction across space and time. Annu Rev Ecol Evol Syst. 2009; 40: 677. [Google Scholar]
- 51. Jaramillo J, Muchugu E, Vega FE, Davis A, Borgemeister C, Chabi-Olaye A. Some like it hot: the influence and implications of climate change on coffee berry borer (Hypothenemus hampei) and coffee production in East Africa. PLoS One. 2011; 6: e24528 10.1371/journal.pone.0024528 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Macfadyen S, Kriticos DJ. Modelling the Geographical Range of a Species with Variable Life-History. PLoS ONE. 2012; 7: e40313 10.1371/journal.pone.0040313 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Parsa S, Kondo T, Winotai A. The Cassava Mealybug (Phenacoccus manihoti) in Asia: First Records, Potential Distribution, and an Identification Key. PLoS ONE. 2012; 7: e47675 10.1371/journal.pone.0047675 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. De Villiers M, Hattingh V, Kriticos DJ. Combining field phenological observations with distribution data to model the potential distribution of the fruit fly Ceratitis rosa Karsch (Diptera: Tephritidae). Bull Entomol Res. 2013; 103: 60–73. 10.1017/S0007485312000454 [DOI] [PubMed] [Google Scholar]
- 55. Parry ML. Climate Change 2007: impacts, adaptation and vulnerability: contribution of Working Group II to the fourth assessment report of the Intergovernmental Panel on Climate Change [Internet]. Cambridge University Press; 2007. Available: http://books.google.com/books. [Google Scholar]
- 56. Porter JH, Parry ML, Carter TR. The potential effects of climatic change on agricultural insect pests. Agric For Meteorol. 1991; 57: 221–240. [Google Scholar]
- 57. Dillon ME, Wang G, Huey RB. Global metabolic impacts of recent climate warming. Nature. 2010; 467: 704–706. 10.1038/nature09407 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Four individuals were sequenced and the sequences were deposited at the GenBank with accession numbers KP324752 and KP324753. The Blast search through the NCBI BLASTN linked all the four individuals to a T. absoluta sample from Serbia of accession number JN417242, with an E-value of 0 and with a probability of 99% match.