

Abstract
The role of Toll-like receptor-2 (TLR2) in radio-resistance remained largely unknown. TLR2 knockout (TLR2−/−) mice received radiation of 6.5 Gy, and then were studied. We found that radiation resulted in more severe mortality and morbidity rates in TLR2−/− mice. The cause of death in TLR2−/− mice may be severe and persistent bone marrow cell loss. Injection of the TLR2 agonist Pam3CSK4 into wild type (WT) mice induced radio-resistance. Myd88−/− mice were more susceptible to radiation. In conclusion, our data indicate that, similar to TLR4, TLR2 plays a critical role in radio-resistance.
Radiotherapy is one of the most commonly used anticancer therapies in the clinic1,2,3,4. However, the most serious adverse effect of ionizing radiation is the acute radiation syndromes especially the bone marrow cells (BMC) failure in the hematopoietic syndrome5,6. The extreme sensitivity of BMC to genotoxic stress is a key factor for the survival of the body. Although mechanisms of radiation injury have been elucidated and the adioprotectants for medical and biodefense applications have been, and continue to be developed5,6,7,8, knowledge in this area is still very limited.
Toll like receptors have a crucial role in the detection of microbial infection in mammals and insects9. To date, there are 13 types of TLRs with different functions in humans and mice10. In a previous study in 2008, Burdelya et al.5 indicated that a TLR5 agonist, CBLB502, showed radio-protective activity in mouse and monkey models by activation of the NF-κB pathway. Other studies also showed that the CBLB502 serves as a possible link between radio-resistance and the epithelial innate immune response11,12. In our previous study, we found that TLR4 played a critical role in radio-resistance13.
TLR2, TLR4 and TLR5 are in the same TLR family and they have many similarities in their signaling pathway14,15. TLR2 molecular expression is very broad and the function of TLR2 has been much studied14. Two studies demonstrated that TLR2 was involved in TBI sensitivity16,17, and Pam3CSK4 showed a significant in vivo radioprotective efficacy16. The signaling pathway involved in the reported TRL2- mediated radio-resistance remains unclear.
In this study, we confirmed that the radio-resistance effects of TLR2 by using TLR2−/− mice. As Myd88 plays a central role in the innate and adaptive immune response18, and is the important adaptor of TLR2 pathways14. In this study, we confirmed the radio-resistance effects of TLR2, and indicated that Myd88 was necessary for radio-resistance mediated by TLR2.
Material and Methods
Mice and treatments
C57BL/6 mice, 5–6 weeks of old, and TLR 2 knock out mice were purchased from The Model Animal Research Center, Nanjing University. All mice were housed in a Specific Pathogen-Free (SPF) facility for all experiments. All animal experiments were performed in accordance with the National Institute of Health “Guide for the Care and Use of Laboratory Animals”19, with the approval of the Laboratory Animal Center of the Second Military Medical University, Shanghai. All efforts were made to minimize the number of animals used as well as any suffering.
Reagents
Pam3CSK4 was obtained from EMC Microcollection (Tübingen, Germany)20. Flow cytometry staining buffer (eBioscience, San-Diego, CA, USA); Annexin V-FITC and propidium iodide (PI) double staining apoptosis assay kit were purchased from Bipec Biopharma (Cambridge, MA, USA). Anti-BrdU antibodies (Cell Signaling Technology, Cat. #5292) and rabbit anti-mouse IgG-horse radish peroxidase-conjugate antibodies were from Cell Signaling Technology; Ampicillin was from Changhai Hospital, (Shanghai, China). DMEM and RPMI 1640 medium and fetal calf serum were from PAA Laboratories, GmbH,(Pasching, Austria). BrdU and 2′7′-di-chlorofluorescein diacetate (DCFH-DA, Molecular Probes, Biyuntian, China).
Cell culture
Cells were cultured in 6-well plates at 37 °C in a humidified atmosphere of 5% CO2 with DMEM and 1640 Medium (PAA, Austria) containing 10% fetal calf serum (PAA, Austria). Exponentially growing cells were used for experiments.
Total-body irradiation
A 60Co irradiator was used for total-body ionizing irradiation. Un-anaesthetized mice were placed in well-ventilated plastic boxes and exposed to the 60Co-γ radiation at a distance of 3 m 4 from the source. Four weekly sub-lethal doses of 5.5 6.5 7.5 8.5 9.5 Gy gamma-ray irradiation were delivered at a dose rate of 1 Gy/min as described previously13,21,22. The mice were then removed from the plastic boxes and allowed free access to food and drinking water.
Histological study
Tissues from bone marrow, testis, thymus, liver, lung, kidney, spleen and colon of euthanized TLR2−/− mice and TLR2−/− mice with different treatments were harvested and subjected to histological assays as described previously. Briefly, tissues were dehydrated in an ascending grade of ethanol, cleared and embedded in paraffin wax. Serial sections of 2–7 microns thick were obtained using a rotatory microtome. The deparaffinized sections were stained routinely using the hematoxylin and eosin (H and E) method. Photomicrographs of the desired sections were obtained using a digital research photographic microscope.
BrdU labeling
Crypt stem cell survival was determined 5, 14 and 20 days after irradiation by 5-bromo-2′-deoxyuridine (BrdU) incorporation into proliferating crypt cells, using a modification of the microcolony assay as described previously, with a minor change of the secondary antibody. S phase cells were labeled in vivo by administering BrdU (i.p., 120 mg/kg) to each mouse 2 hours before euthanasia. Mice were sacrificed and the bone marrow and small intestine were rapidly dissected, fixed in 10% neutral buffered formalin, and embedded in paraffin. Paraffin sections (2–5 μm) were cut perpendicular to the long axis of the intestines. Cells incorporating BrdU were visualized by immunohistochemistry using rat monoclonal anti-BrdU antibodies (Abcam, Cambridge, MA; Cat. # ab6326, dil.1:100) and secondary HRP-conjugated donkey anti-rat IgG antibody (Cell Signaling technology, Beverly, MA).
Apoptosis assay
Cells from each group were harvested at 48 h post-transfection. Cells were resuspended at a density of 1 × 106 cells/mL in PBS. After double staining with FITC-Annexin V and (PI) using the FITC Annexin V Apoptosis Detection Kit I (BestBio, Shanghai, China), cell were analyzed using FACScan flow cytometer (BD Biosciences) equipped with Cell Quest software (BD Biosciences). Annexin V + PI- cells were counted as apoptosis cells.
Quantitation of GM-CFU
Granulocyte-macrophage colony-forming units (GM-CFU was assayed in semisolid methylcellulose culture as described previously, with minor revisions. Mononuclear bone marrow cells (BMC) from femora and tibiae of non-irradiated TLR2−/− mice and TLR2−/− mice were pooled. Red blood cells were removed using RBC lysis buffer (eBioscience, San Diego, Ca). BMC were suspended in Iscove’s modified Dulbecco’s medium (IMDM) containing 30% fetal calf serum, 1% bovine serum albumin, 100 uM beta-mercaptoethanol, 10 ng/ml recombinant mouse granulocyte monocyte-colony stimulating factor (mGM-CSF; Biosource Cat.# PMC2016) and 1 ng/ml recombinant IL-3 (mIL-3; ProTech Rocky Hill NJ, USA). One-milliliter aliquots of the BMC suspension were plated in triplicate in 35-mm tissue culture dishes and incubated for 7 days in a humidified incubator at 37 °C with 5% CO2. Colonies were counted under a light microscope.
Statistical analysis
Comparisons between experimental groups and relevant controls (but not survival curves) were performed using a Student’s t-test. Differences in survival of the various groups of mice were assessed using Kaplan-Meier plus Cox Regression Analysis with the SPSS (Statistical Program for Social Sciences) software. The SPSS software generated a P value and Chi-Square value for each analysis; P < 0.05 was considered a statistically significant difference.
Results
TLR2−/− Mice were more susceptible to radiation-induced mortality
We analyzed the difference in radio-sensitivity between wild type (WT) and TLR2−/− mice under 0, 5.5, 6.5, 7.5, 8.5 Gy of γ-radiation (Fig. 1A). Initially, without radiation, there was no difference between TLR2−/− and WT mice. TLR2−/− mice were able to survive over a year under specific pathogen-free (SPF) conditions. Then, to investigate survivability after exposure to ionizing radiation (IR), age- and sex- matched TLR2−/−and WT mice were exposed to 0, 5.5, 6.5, 7.5, 8.5 Gy of γ-radiation. After radiation, the survival rates of TLR2−/− and WT mice were compared. At the dose of 6.5 Gy WT mice had a survival rate of 100%, whereas TLR2−/− mice had a survival rate of only 60% (p < 0.05). Consistently, TLR2−/− mice also showed higher mortality than WT mice following exposure to 7.5 or 8.5 Gy radiation. Regression analysis of the survival after radiation exposure confirmed that under the same dose of radiation, there were more deaths in TLR2−/− mice group (Fig. 1B). Taken together, these data, indicated that TLR2−/− mice were more susceptible to radiation-induced mortality.
Figure 1. Increased mortality in TLR2−/− mice exposed to IR.

Survival of TLR2−/− and WT mice to month 12 without IR in SPF conditions (n = 8). TLR2−/− (n = 40) and WT (n = 40) mice were randomly divided into four groups and exposed to 5.5, 6.5, 7.5 or 8.5 Gy 60Co-γ radiation (dose rate: 1 Gy/min). Survival was monitored until day 30 after IR (A) Liner regression analysis of the survival rate for TLR2−/− and WT control mice after IR. The equation and the correlation coefficient for the line are given (B) All data are expressed as the mean ± s.d. of three separate experiments. *P < 0.05.
Mortality of TLR2−/− mice after radiation was associated with a severe and persistent loss of BMC
To investigate the reason for the increased mortality of TLR2−/− mice, TLR2−/− and WT mice were processed to perform mass biopsy for a histological study of the radiation-induced tissue damage. A 6.5 Gy TBI was found to induce tissue damage in multiple organs of both TLR2−/− and WT mice, but the TLR2−/− mice showed greater injury in bone marrow (Fig. 2A), stomach, spleen, and kidney (Supplementary Figure S1). The histology inspection of these organs suggested that cause of death in TLR2−/− mice may be the severe loss of BMC (Fig. 2A). Hence, to further confirm this result, the number of BMC and peripheral blood mononuclear cell (PBMC) in TLR2−/− were counted after exposure to 6.5 Gy of radiation. We found that 6.5 Gy exposure led more BMC and PBMC loss in TLR2−/− mice. The BM showed the greatest damage and cell loss at day 5 post-radiation, while, by day 28, the bone marrow from WT mice was well repaired and the numbers of cells were partly recovered, this was not the case in TLR2−/− mice (Fig. 2B,C). These data consistently indicate that the main cause of death in TLR2−/− may be a severe and persistent BMC loss after radiation.
Figure 2. Increased mortality of TLR2−/− mice after IR associated with a severe and persistent loss of BMC.

TLR2−/− and WT mice were irradiated with 6.5 Gy and the femur was collected and stained with H and E at day 0, 1, 5, 14 and 28 post-IR. All images are × 100. The figure shows a typical H and E image of three independent experiments (n = 8) (A) The number of BMC was counted on day 0, 1, 5, 14 and 28 post-IR (B) The number of PBMC was counted at day 0, 1, 5, 14 and 28 post-IR (C) *P < 0.05.
BMC loss in TLR2−/− mice was associated with cell apoptosis and increased oxidative stress
To investigate whether the loss of BMC is due to the cell apoptosis, we assayed the cell apoptosis rates of BMC from TLR2−/− and WT mice 24 h after exposure to different doses of radiation. We found that that radiation induced BMC apoptosis in a dose-dependent manner in both TLR2−/− and WT mice, and radiation induced more cell apoptosis in TLR2−/−mice (Fig. 3A,). TUNEL assays also indicated there were more cell apoptosis in colon, testis and lung in TLR2−/− mice than in WT mice (Fig. 3B). Besides, DCFH-DA assay showed that there were more oxidative stress in TLR2−/− mice than in WT mice (Fig. 3C) .These data indicated that TLR2 was required for radio-resistance by suppression of apoptosis in many tissues, and especially the BMC.
Figure 3. Increased oxidative stress and apoptosis rate of TLR2−/− mice after IR BMC were prepared 24 h after IR exposure, stained with AnnV-FITV, and PI apoptosis was detected by FACS analysis.
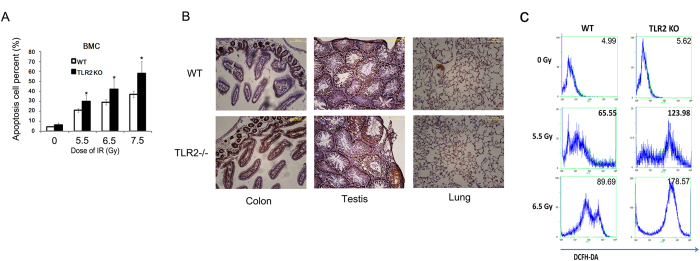
(A) The TLR2−/− and WT mice were irradiated with 6.5 Gy then 1 day later, colon, testis and lung were collected and subjected to TUNEL assay (B) The TLR2−/− mice and WT mice were irradiated with 0, 5,5, 6.5 Gy; next, 24 h later, BMC were prepared and analyzed for oxidative stress using the DCFH-DA assay (C) All data are expressed as the mean ± s.d. of three separate experiments. *P < 0.05.
BMC from TLR2−/− mice showed impaired proliferation capacity
We also performed GM-CFU units and 5-bromo-2′-deoxyuridines (BrdU) to explore why the BM loss was more persistent in TLR2−/− mice. We found that BMC from TLR2−/− mice showed significantly impaired proliferation capacity without IR (Fig. 4A). In addition, the results of the BrdU assay on days 1, 5, 14 and 28 after 5 Gy, revealed that not only the total number of BMC decreased in TLR2−/− mice after TBI, but also irradiated BMC from TLR2−/− mice showed significantly impaired tissues repair capacity when compared to WT mice(Fig. 4B).
Figure 4. BMC from TLR2−/− mice showed impaired proliferation capacity Colony-forming units Granulocyte/macrophage was quantified in BMC obtained from TLR2−/− and WT mice with or without exposure to IR (6.5 Gy).
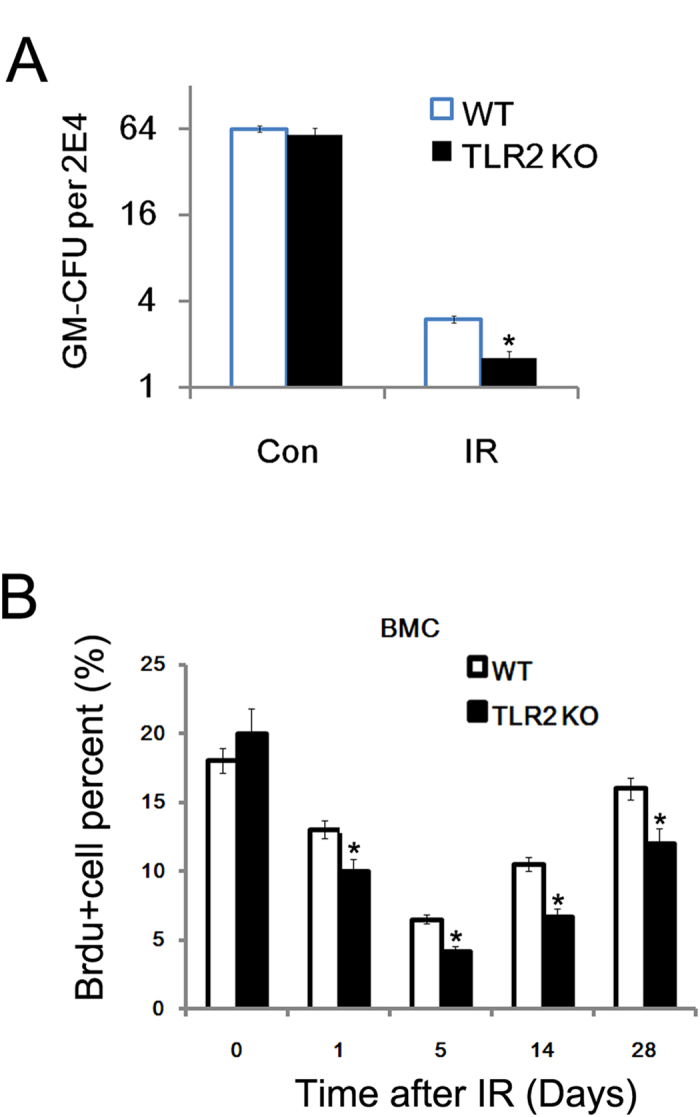
(A) The TLR2−/− and WT mice were exposed to 6.5 Gy, femurs were harvested 5, 14, and 20 days later, and the proliferation of BMC was measured by the BrdU assay. All data are expressed as the mean ± s.d. of three separate experiments. *P < 0.05.
The role of TLR2 agonist Pam3CSK4 in radio-protection
Pam3CSK4 is an agonist of TLR2, and here it was applied to investigate the role of TLR2 in radio-resistance. When given 24 h before radiation exposure, Pam3CSK4 displayed significant radio-protective effects on C57BL/10 mice exposed to doses of 7.5, 8.5 or 9.5 Gy as shown in Fig. 5A
Figure 5. TLR2 induced radio-protection via Myd88 C57BL/10 mice (20 g in weight, n = 20 per group) were treated with Pam3CSK4 (50 ng/ml) and then 24 h later exposed to 7.5, 8.5 and 9.5 Gy TBI as indicated, while control mice received PBS.

Survival was recorded (A). Linear regression analysis of the survival rate for mice treated with Pam3CSK4. The equation and the correlation coefficient for the line are given (B). Pam3CSK4 protects C57BL/10 mice from radiation injury. Survival was recorded (C). Myd88−/− and WT mice were exposed to 0 and 6.5 IR, and the survival was recorded. All data are expressed as the mean ± s.d. of three separate experiments. *P < 0.05.
In addition, the calculated dose-reduction factor (DRF) of Pam3CSK4 was 1.2 (Pam3CSK4 versus PBS)(Fig. 5B), and when Pam3CSK4 was given to TLR2−/− mice 24 h before exposure to doses of 7.5 Gy, the results revealed that Pam3CSK4 did not show a radio-protective effects on TLR2−/− mice (Fig. 5C). Additionally, as the Myd88 dependent pathway is the downstream pathways of TLR214, in vivo experiments in Myd88−/− mice were conducted to explored whether Myd88 participate in TLR2-induced radio-protection. The Myd88−/− mice was studies. Myd88−/− mice and WT mice were exposed to 6.5 IR, and the survival was recorded (Fig. 5D). Our data indicated that TLR2 induced radio-protection via Myd88.
Discussion
In this study, we found that mice deficient in TLR2−/− were more susceptible to IR-induced mortality and morbidity. Mortality in TLR2−/− mice was associated with a severe and persistent loss of BMC. Treatment with the TLR2 agonist Pam3CSK4 induced radio-resistance. Myd88 may be the critical adaptor for TLR2 mediated radio-protection. Taken together, our data suggested that TLR2 and its agonist Pam3CSK4 were required for radio-resistance.
Previous study indicated that monocytes incubated with Pam3CSK4 produced IL-6, IL-8, IL-1β and IL-1020, and that IL-6 was a radioprotective cytokine5. Indeed, Pam3CSK4 is the potent inducer of IL-12p35 and IL-10 gene expression in murine bone marrow-derived dendritic cells (DCs), as well as in purified oral myeloid DCs. Moreover, sublingual administration of Pam3CSK4, together with the antigen in BALB/c mice sensitized to OVA, dramatically decreases airway hyperresponsiveness as well as OVA-specific T-helper type 2 (Th2) responses in cervical lymph nodes23. As our data indicated that it is Myd88 which is involved in the mechanism of the TLR2-induced radio-protection, thus we hypothesized that TLR2-mediated radioprotection is likely to involve multiple mechanisms.
In this and previous study, TLR2−/− or TLR4−/− mice received TBI, and mortality in both mice strain was associated with severe and persistent loss of BMC. Interestingly, exposure of TLR2−/− or TLR4−/− mice to 18-Gy whole thorax radiation has shown that the combined deficiency of these receptors decreases survival time and enhances the development of fibrosis24.
Our data indicated the radioprotective role of Myd88. Previous study showed that MyD88 is important for host survival from radiation-induced injury and that in the absence of MyD88, cells accumulated in the lung, which ultimately displayed a fibrotic phenotype by 24 weeks following radiation exposure25. These results demonstrate that MyD88 is important for regulating non-infectious inflammatory processes so as to promote healthy tissue regeneration. Besides, there is evidence suggesting MyD88 in regulating hematopoiesis and cell replenishment programs26,27,28.
In conclusion, our data suggest that TLR2 signaling may play a critical role in radio-resistance, at least partly via MyD88.
Additional Information
How to cite this article: Gao, F. et al. A critical role of toll-like receptor 2 (TLR2) and its' in vivo ligands in radio-resistance. Sci. Rep. 5, 13004; doi: 10.1038/srep13004 (2015).
Supplementary Material
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (Grants No. 31100605, No.31170827 and No. 81402630).
Footnotes
Author Contributions F.G., C.Z. and C.L. involved in study concept and design, carried out experiments, W.S., X.L., P.Z., J.H., L.X., D.B., H.L., Y.C., B.L. and J.C. carried out experiments; C.L. and C.Z. drafted the manuscript. C.Z., C.L. and J.C. involved in study design, obtained funding.
References
- Moding E. J., Kastan M. B. & Kirsch D. G. Strategies for optimizing the response of cancer and normal tissues to radiation. Nat Rev Drug Discov 12, 526–542 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barcellos-Hoff M. H., Park C. & Wright E. G. Radiation and the microenvironment—tumorigenesis and therapy. Nat Rev Cancer 5, 867–875 (2005). [DOI] [PubMed] [Google Scholar]
- Thoms J. et al. Neoadjuvant radiotherapy for locally advanced and high-risk prostate cancer. Nat Rev Clin Oncol 8, 107–113 (2011). [DOI] [PubMed] [Google Scholar]
- Eifel P. J. Radiotherapy: Intermediate-risk endometrial cancer--adjuvant treatment. Nat Rev Clin Oncol 7, 553–554 (2010). [DOI] [PubMed] [Google Scholar]
- Burdelya L. G. et al. An agonist of toll-like receptor 5 has radioprotective activity in mouse and primate models. Science 320, 226–230 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirsch D. G. et al. p5 controls radiation-induced gastrointestinal syndrome in mice independent of apoptosis. Science 327, 593–596 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J. H. & Park J. W. A manganese porphyrin complex is a novel radiation protector. Free Radic Biol Med 37, 272–283 (2004). [DOI] [PubMed] [Google Scholar]
- Lee J. H. & Park J. W. Oxalomalate regulates ionizing radiation-induced apoptosis in mice. Free Radic Biol Med 42, 44–51 (2007). [DOI] [PubMed] [Google Scholar]
- Medzhitov R. Toll-like receptors and innate immunity. Nat Rev Immunol 1, 135–145 (2001). [DOI] [PubMed] [Google Scholar]
- Barton G. M. & Medzhitov R. Toll-like receptor signaling pathways. Science 300, 1524–1525 (2003). [DOI] [PubMed] [Google Scholar]
- Janot L. et al. Radioresistant cells expressing TLR5 control the respiratory epithelium’s innate immune responses to flagellin. Eur J Immunol 39, 1587–1596 (2009). [DOI] [PubMed] [Google Scholar]
- Vijay-Kumar M. et al. Flagellin treatment protects against chemicals, bacteria, viruses, and radiation. J Immunol 180, 8280–8285 (2008). [DOI] [PubMed] [Google Scholar]
- Liu C. et al. A critical role of toll-like receptor 4 (TLR4) and its’ in vivo ligands in basal radio-resistance. Cell Death Dis 4, e649 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akira S. & Takeda K. Toll-like receptor signalling. Nat Rev Immunol 4, 499–511 (2004). [DOI] [PubMed] [Google Scholar]
- O’Neill L. A. & Bowie A. G. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat Rev Immunol 7, 353–364 (2007). [DOI] [PubMed] [Google Scholar]
- Shakhov A. N. et al. Prevention and mitigation of acute radiation syndrome in mice by synthetic lipopeptide agonists of Toll-like receptor 2 (TLR2). PLoS One 7, e33044 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh V. K. et al. CBLB613: a TLR 2/6 agonist, natural lipopeptide of Mycoplasma arginini, as a novel radiation countermeasure. Radiation research 177, 628–642 (2012). [DOI] [PubMed] [Google Scholar]
- Yamamoto M. et al. Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science 301, 640–643 (2003). [DOI] [PubMed] [Google Scholar]
- Barthold S. W., Bayne K. & Davis M. Guide for the care and use of laboratory animals. (ed^(eds). Washington, National Academy Press (2011). [Google Scholar]
- Funderburg N. T. et al. The Toll-like receptor 1/2 agonists Pam(3) CSK(4) and human beta-defensin-3 differentially induce interleukin-10 and nuclear factor-kappaB signalling patterns in human monocytes. Immunology 134, 151–160 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu Z. et al. Expression changes of ERK1/2, STAT3 and SHP-2 in bone marrow cells from gamma-ray induced leukemia mice. J Radiat Res (Tokyo) 47, 121–130 (2006). [DOI] [PubMed] [Google Scholar]
- Liu C. et al. Gamma-ray irradiation impairs dendritic cell migration to CCL19 by down-regulation of CCR7 and induction of cell apoptosis. Int J Biol Sci 7, 168–179 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lombardi V. et al. Toll-like receptor 2 agonist Pam3CSK4 enhances the induction of antigen-specific tolerance via the sublingual route. Clin Exp Allergy 38, 1819–1829 (2008). [DOI] [PubMed] [Google Scholar]
- Paun A. et al. Combined Tlr2 and Tlr4 deficiency increases radiation-induced pulmonary fibrosis in mice. Int J Radiat Oncol Biol Phys 77, 1198–1205 (2010). [DOI] [PubMed] [Google Scholar]
- Brickey W. J. et al. MyD88 provides a protective role in long-term radiation-induced lung injury. International journal of radiation biology 88, 335–347 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagai Y. et al. Toll-like receptors on hematopoietic progenitor cells stimulate innate immune system replenishment. Immunity 24, 801–812 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGettrick A. F. & O’Neill L. A. Toll-like receptors: key activators of leucocytes and regulator of haematopoiesis. British journal of haematology 139, 185–193 (2007). [DOI] [PubMed] [Google Scholar]
- Kenny E. F. & O’Neill L. A. Signalling adaptors used by Toll-like receptors: an update. Cytokine 43, 342–349 (2008). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.