

Abstract
Rationale
Obesity is a leading public health problem worldwide. Multiple lines of evidence associate deficits in the brain reward circuit with obesity.
Objective
Whether alterations in brain reward sensitivity precede or are a consequence of obesity is unknown. This study aimed to investigate both innate and obesity-induced differences in the sensitivity to the effects of an indirect dopaminergic agonist.
Methods
Rats genetically prone to diet-induced obesity (DIO) and their counterpart diet-resistant (DR) were fed a chow diet and their response to D-amphetamine on intracranial self-stimulation and food intake were assessed. The same variables were then evaluated after exposing the rats to a high-fat diet, after DIO rats selectively developed obesity. Finally, gene expression levels of dopamine receptor 1 and 2 as well as tyrosine hydroxylase were measured in reward-related brain regions.
Results
In a pre-obesity state, DIO rats showed innate decreased sensitivity to the reward-enhancing and anorectic effects of D-amphetamine, as compared to DR rats. In a diet-induced obese state, the insensitivity to the potentiating effects of D-amphetamine on ICSS threshold persisted and became more marked in DIO rats, while the anorectic effects were comparable between genotypes. Finally, innate and obesity-induced differences in the gene expression of dopamine receptors were observed.
Conclusions
Our results demonstrate that brain reward deficits antedate the development of obesity and worsen after obesity is fully developed, suggesting that these alterations represent vulnerability factors for its development. Moreover, our data suggests that the reward-enhancing and anorectic effects of D-amphetamine are dissociable in the context of obesity.
Keywords: Dopamine receptors, Dorsal Striatum, Food intake OR feeding, high-fat OR high fat, intracranial self-stimulation OR ICSS, obesity OR obese, Nucleus Accumbens, reward, obesity OR obese, Ventral Tegmental Area OR VTA
INTRODUCTION
Obesity is an epidemic medical condition defined by the World Health Organization as a body mass index (BMI) ≥30 kg/m. Obesity shows high comorbidity with a variety of diseases, raising both the economic impact of healthcare and the risk of mortality (Bray 2004; Lehnert et al. 2013). Obesity is a polygenic disorder and results from the complex interaction with an obesogenic environment (Speakman 2004).
In this context, a useful tool in preclinical research to study the interaction between genes and diet is represented by rats genetically prone to diet-induced obesity (DIO) and their diet-resistant (DR) counterpart (Levin et al. 1997). When fed a standard chow diet, DIO rats are lean; however, when exposed to high-fat diet, they disproportionally gain weight and become obese, while DR remain lean (Cottone et al. 2013; Levin et al. 1997). Therefore, the DIO/DR rodent model effectively mimics the polygenic, heritable differences in the vulnerability to diet-induced obesity observed in humans, allowing the study of pre-existing, predisposing factors leading to obesity (Cottone et al. 2007).
In addition to metabolic and homeostatic mechanisms, the brain reward system plays an equally important role in feeding. Indeed, the mesocorticolimbic dopaminergic pathway, which is activated in response to palatable food, has a major influence on feeding behavior.
The hypothesis that increased caloric intake and weight gain in obesity is driven by central mechanisms regulating reward is supported by evidence demonstrating that obese subjects show a deficit in the brain reward circuitry and a general hyposensitivity of dopaminergic systems (Johnson and Kenny 2010; South and Huang 2008; van de Giessen et al. 2014; Volkow et al. 2013b). These deficits may, in turn, lead obese subjects to compensate by eating more palatable food, as observed in drug addiction(Guo et al. 2014; Melis et al. 2005).
However, how exactly the brain reward system regulates food intake and body fatness is unclear. In particular, whether these alterations precede the development of obesity or are a consequence is unknown.
In this study we investigated the effect of D-amphetamine on brain reward function using intracranial self-stimulation (ICSS), as well as on food intake and body weight change, in order to assess the responsiveness of the dopaminergic system of DIO and DR rats in pre-obesity and obesity states. D-Amphetamine was chosen as a pharmacological tool to study the sensitivity of the dopaminergic system due its dopamine-releasing properties, and consequent elevations of extrasynaptic dopamine levels (Fleckenstein et al. 2007; Sulzer 2011). Acute D-Amphetamine administration lowers the threshold for electrical stimulation, indicating a potentiation of the rewarding properties of electrical brain stimulation (Esposito et al. 1980; Goodall and Carey 1975; Kornetsky and Esposito 1979).
Human brain imaging studies have demonstrated that obese patients have decreased dopamine 2 receptor (D2R) occupancy in the dorsal striatum, which negatively correlates with the body mass index (Wang et al. 2001). Alterations of the mesolimbic/mesocortical dopaminergic system have also been reported in environmental and genetic animal models of obesity (Geiger et al. 2009; Volkow et al. 2011; Vucetic and Reyes 2010). For the above reasons, here we also analyzed the gene expression of the dopaminergic system components in several bran regions, including the nucleus accumbens (NAcc), the dorsal striatum (DS), the ventral tegmental area (VTA), the ventromedial hypothalamus (VMH) and the lateral hypothalamus (LH) of both chow-fed and high-fat-fed DIO and DR rats. VTA, NAcc and DS were chosen because of their role in the rewarding and reinforcing properties of drugs and food (Kelley and Berridge 2002; Koob and Volkow 2010; Volkow et al. 2012); the LH and the VMH were chosen because of their role in the dopaminergic regulation of feeding behavior (Leibowitz 1975b; Mayer and Thomas 1967).
MATERIALS AND METHODS
Subjects
Diet-Induced Obesity (DIO) and Diet Resistant (DR) rats (n=44) were purchased from Taconic (Huston, NY) (Levin et al. 1997). Rats were housed in a 12h reverse light/dark cycle, AAALAC-approved vivarium. DIO and DR rats were fed either ad libitum fed chow (corn-based Harlan Teklad LM-485 Diet 7012 (Cottone et al. 2013; Cottone et al. 2007)) or ad libitum high-fat diet (D12266B Research Diets, New Brunswick, NJ) for 4 weeks, to allow DIO to develop obesity.(Ricci and Levin 2003) Procedures adhered to the NIH Guide for the Care and Use of Laboratory Animals and the Principles of Laboratory Animal Care and were approved by the Institutional Animal Care and Use Committee of Boston University. (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) (Pellegrino et al. 1979) Additional information is provided in the online Supplementary Information file.
Drugs
D-amphetamine doses (0, 0.1, 0.5 and 1 mg/kg) were administered intraperitoneally (i.p.) 10 min before experiments, using a Latin square within-subject design (Esposito et al. 1980; Grilly and Loveland 2001). Additional information is provided in the online Supplementary Information file.
Surgery for electrode implantation
Surgery for electrode implantation was performed as previously described (Dore et al. 2013; Iemolo et al. 2012; Kenny and Markou 2005; Schulteis et al. 1995). DIO and DR rats were unilaterally implanted with a 0.125mm diameter bipolar stainless steel electrode (Plastics One, Roanoke, VA) in the medial forebrain bundle at the level of the lateral hypothalamus, using the following coordinates from bregma: AP −0.5 mm, ML ±1.7 mm, DV −9.7 mm from skull, with the incisor bar 5.0 mm above the interaural line (Pellegrino et al. 1979). Additional information is provided in the online Supplementary Information file.
Intracranial self-stimulation procedure
ICSS took place in previously described operant test chambers (Med Associates Inc., St. Albans, VT) (Iemolo et al. 2012) and the ICSS procedure performed as previously reported (Dore et al. 2013; Iemolo et al. 2012) (see Supplementary Information file). Reward thresholds were assessed using a rate-independent discrete-trial current intensity procedure(Esposito and Kornetsky 1977). The brain reward threshold is the minimal current intensity able to produce a response that maintains self-stimulation (Markou and Koob 1991).
Food intake and body weight measurements
Pre-weighed food was provided at dark-cycle onset, 10min after drug administration, and food intake was recorded 2h, 6h, and 24h later. 24h rat body weight change was calculated.
Quantitative RT-PCR
RNA extraction, retrotranscription, and real-time PCR were performed as previously described (Baiamonte et al. 2014; Cottone et al. 2012). Additional information is available in the online Supplementary Information file.
Fat pad and body composition analysis
DIO and DR rats, after prolonged ad libitum feeding with chow diet or high-fat diet, were euthanized and carcasses were transferred to the University of Alabama–Birmingham where the body composition analysis was carried out as previously described (Cottone et al. 2013; Cottone et al. 2007). Additional information is available in the online Supplementary Information file.
Statistical analysis
ICSS, food intake and body weight change data were analyzed using mixed design two-way ANOVAs. Following a statistically significant overall effect and/or interaction, separate one-way ANOVAs were performed. Pairwise post-hoc were either Student’s t test (to compare two groups) or Fisher’s least significant difference (LSD) analysis (for all others), after confirming significant omnibus effect (p≤0.05). Additional information is available in the online Supplementary Information file.
RESULTS
Effects of D-amphetamine on ICSS threshold in chow-fed DIO and DR rats
Basal, spontaneous ICSS threshold did not differ between genotypes during a 4-day baseline period (Genotype: F[1,14]=1.38, n.s.; 4-day average, mean ± SEM: DIO 90.3±10.2 μA, DR: 107.7±13.7 μA, not shown). Fig.1 shows that D-amphetamine treatment significantly lowered the ICSS threshold in both chow-fed DIO and DR rats (Treatment: F[3,42]=10.14, p≤0.001; Genotype*Treatment: F[3,42]=1.46, n.s.). Pairwise comparisons revealed that while in DIO rats only the highest dose of D-amphetamine significantly lowered the ICSS threshold, both the middle and the highest dose did in DR rats. The percent reduction following the 0.5 and 1 mg/kg doses, compared to vehicle, was in DIO 11% and 28%, and in DR rats 27% and 38%. As shown in Figure 1B, D-amphetamine had no effect on the latency to respond (Friedman test: DIO χ2=4.47, n.s.; DR χ2=2.49, n.s), a measure of locomotor activity.
Figure 1.
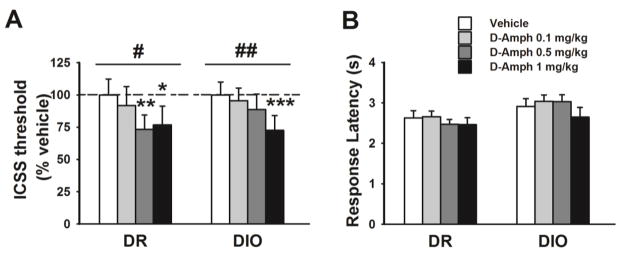
Effects of D-amphetamine administration on ICSS threshold (A) and response latency (B) in chow-fed DIO and DR rats (n=7–9/genotype). Data represent M±SEM. # p≤ 0.05, ## p≤0.01 represents a main effect of Treatment. * p≤0.05, ** p≤0.01, *** p≤0.001 vs. vehicle.
Effects of D-amphetamine on food intake and body weight gain in chow-fed DIO and DR rats
Similarly to what previously reported by Ricci and Levin (2003), the chow intake of DIO rats was slightly but significantly higher as compared to DR rats (DIO: 99.3 ±3.1 kcal, DR: 86.8 ±3.8 kcal; t(16)=2.56 p<0.05, mean ± SEM).
As shown in Fig.2A, D-amphetamine exerted an anorectic effect in both chow-fed DIO and DR rats. The two-way ANOVA revealed that D-amphetamine reduced 2h food intake differentially in DIO and DR rats (Genotype*Treatment: F[3,48]=2.72, p≤0.05; Treatment: F[3,48]=15.45, p≤0.001; Genotype: F[1,16]=4.00, n.s.). Separate one-way ANOVAs suggested that D-amphetamine reduced food intake less potently in DIO rats compared to DR (Treatment: DIO, F[3,27]=3.58, p≤0.03; DR, F[3,21]=14.06, p≤0.0001). Indeed, while in DIO rats only the highest dose of D-amphetamine was effective, both the middle and the highest were in DR rats. The percent reduction following the 0.5 and 1 mg/kg doses, compared to vehicle, was in DIO 4.3% and 36.3%, and in DR 37.6% and 75.8%.
Figure 2.

Effects of D-amphetamine administration on caloric intake and body weight change in chow-fed DIO and DR rats (n=8–10/genotype). Data represent M±SEM of food intake measurements at the 2h (A), 6h (B), and 24h (C) time points. (D) 24h body weight change. # p≤0.05, ### p≤0.001 represent main effects of Treatment. ** p≤0.01, *** p≤0.001 vs. vehicle.
Although main effects of the genotype and drug treatment were detected by the two-way ANOVA on the 6h measures (Treatment: F[3,48]=3.28, p≤0.03; Genotype*Treatment: F[3,48]=0.12, n.s.; Genotype: F[1,16]=12.39, p≤0.003), the magnitude of the effect was such that separate one way ANOVAs did not confirm them (DIO Treatment: F[3,27]=1.45, n.s.; DR Treatment: F[3,21]=2.16, n.s.; Fig.2B).
At the 24h time point, the drug effect on food intake was no longer observed (Treatment: F[3,48]=2.36, n.s.; Genotype*Treatment: F[3,48]=0.17, n.s.; Genotype: F[1,16]=12.70, p≤0.003; Fig.2C). In addition, body weight change was not affected by D-amphetamine treatment in either genotype fed with chow (Treatment: F[3,48]=0.71, n.s.; Genotype*Treatment: F[1,16]=0.08, n.s.; Genotype: F[1,16]=1.22, n.s.; Fig.2D).
Effects of D-amphetamine on ICSS threshold in high-fat-fed DIO and DR rats
4-day spontaneous baseline ICSS threshold did not differ between obese DIO and DR fed with high-fat diet (Genotype: F[1,12]=1.90, n.s.; 4-day average, Mean±SEM: DIO 80.9±9.8, DR: 102.1±10.9, not shown). D-amphetamine treatment decreased ICSS threshold of high-fat-fed rats (Treatment: F[3,36]=8.46, p≤0.001; Genotype*Treatment: F[3,36]=1.95, n.s.; Genotype: F[1,12]=0.26, n.s.). As shown in Fig.3, one-way ANOVAs on individual genotypes revealed that D-amphetamine administration lowered the ICSS threshold dose-dependently in DR rats, while it had no effect in obese DIO rats (DR Treatment: F[3,18]=15.65, p≤0.001; DIO Treatment: F[3,18]=1.53, n.s.). Pairwise comparisons showed that in DR rats all three doses tested significantly reduced the ICSS threshold. The percent reduction following the 0.5 and 1 mg/kg doses, compared to vehicle, was in DIO 12% and 19%, and in DR 31% and 32%. While a linear contrast analyses did not show a significant Genotype x Dose interaction under chow conditions (F(1,14)= 0.18; n.s.), this was instead statistically significant under high-fat diet (F(1,12)= 5.29; p<0.05), confirming the concept that differences in the response to the psychostimulants between genotypes became much more evident after exposure to high-fat food. As shown in Figure 3B, the ICSS response latency was unaffected by drug administration (Friedman test: DIO χ2=3.86, n.s.; DR χ2=3.86, n.s).
Figure 3.
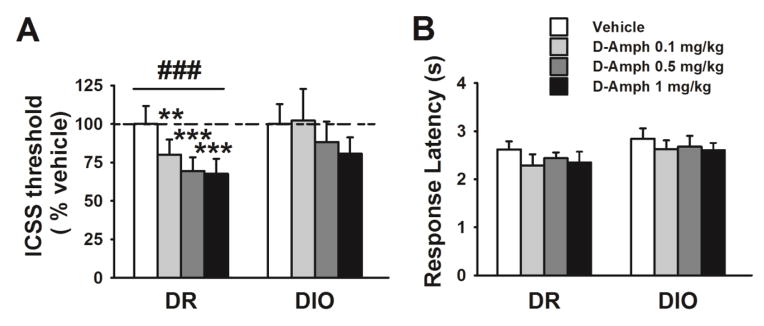
Effects of D-amphetamine administration on ICSS threshold (A) and response latency (B) in high-fat-fed DIO and DR rats (n=7/genotype). Data represents M±SEM. ### p≤0.001 represents main effect of Treatment. ** p≤0.01, *** p≤0.001 vs. vehicle.
Effects of D-amphetamine on food intake and body weight gain in high-fat-fed DIO and DR rats
The high-fat food intake did not differ between genotypes (DIO: 101.9 ± 4.4 kcal, DR: 89.7 ± 8.0 kcal; t(16)=1.42 n.s.), consistently with previous reports (Cottone et al. 2013; Madsen et al. 2010).
As shown in Fig.4, the effects of D-amphetamine administration in high-fat diet fed DIO and DR rats on food intake were more profound compared to chow (Fig.2). The two-way ANOVAs revealed that D-amphetamine administration exerted a significant effect at all the time points (Fig.4A 2h: Treatment: F[3,48]=56.57, p≤0.001; Genotype*Treatment: F[3,48]=0.35, n.s.; Genotype: F[1,16]=2.80, n.s. Fig.4B 6h: Treatment: F[3,48]=22.22, p≤0.001; Genotype*Treatment: F[3,48]=2.37, n.s.; Genotype: F[1,16]=6.36, p≤0.03. Fig.4C 24h: Treatment: F[3,48]=13.26, p≤0.001; Genotype*Treatment: F[3,48]=0.20, n.s.; Genotype: F[1,16]=4.62, p≤0.05). Separate one-way ANOVAs confirmed the anorectic effect of D-amphetamine in both genotypes (2h: DIO Treatment: F[3,27]=52.72, p≤0.001; DR Treatment: F[3,21]=15.76, p≤0.001. 6h: DIO Treatment: F[3,27]=20.28, p≤0.001; DR Treatment: F[3,21]=5.85, p≤0.005. 24h: DIO Treatment: F[3,27]=10.75, p≤0.001; DR Treatment: F[3,21]=4.16, p≤0.02). Post-hoc analysis revealed that the treatment significantly and dose-dependently reduced the high-fat intake during the entire 24h period at the middle and highest doses tested. The lowest dose significantly reduced food intake in both genotypes only at the 2h time point.
Figure 4.

Effects of D-amphetamine administration on caloric intake and body weight change in high-fat-fed DIO and DR rats (n=8–10/genotype). Data represent M±SEM of food intake measurements at the 2h (A), 6h (B), and 24h (C) time points. (D) 24h body weight change. # p≤0.05, ## p≤0.01, ### p≤0.001 represent main effects of Treatment. * p≤0.05, ** p≤0.01 and *** p≤ 0.001 vs. vehicle.
As shown in Fig.4D, D-amphetamine treatment decreased the body weight gain of both high-fat-fed DIO and DR rats (Treatment: F[3,48]=9.57, p≤0.001; Genotype*Treatment: F[3,48]=2.17, n.s.; Genotype: F[1,16]=1.14, n.s.). Separate one-way ANOVAs confirmed the efficacy of D-amphetamine in both genotypes (DIO Treatment: F[3,27]=8.15, p≤0.001; DR Treatment: F[3,27]=4.30, p≤0.02). Post-hoc analyses revealed that in DR rats the highest dose, while in DIO both the middle and the highest were.
Gene expression in chow or high-fat-fed rats
As shown in Fig.5, lean chow-fed DIO rats showed higher D1R and D2R mRNA levels, compared to chow-fed DR rats, in the VTA and in the DS, but not in the NAcc (VTA D1R: t(15)=2.22, p≤0.04; DS D1R: t(17)=2.93, p≤0.01; NAcc D1R: t(16)=0.71, n.s.; VTA D2R: t(15)=2.33, p≤0.03; DS D2R: t(16)=3.45, p≤0.003; NAcc D2R: t(15)=1.36, n.s.). Moreover, in the VTA chow-fed DIO rats showed higher mRNA levels of the biosynthetic enzyme TH compared to the DR rats (t(16)=2.48, p≤0.03).
Figure 5.

Dopamine 1 receptor (D1R), dopamine 2 receptor (D2R) and tyrosine hydroxylase (TH) mRNA expression in the ventral tegmental area (VTA), in the nucleus accumbens (NAcc), and in the dorsal striatum (DS) in chow-fed (A, B, and C) and high-fat diet fed (D, E, and F) DIO and DR rats (n=7–9/genotype). Data represent M±SEM expressed as percent of DR rats. * p≤0.05, ** p≤0.01, ** p≤0.001 vs. DR rats.
Interestingly, obese DIO had lower D1R mRNA level than the lean high-fat-fed DR rats in the VTA (t(9)=2.44, p≤0.03) and in the NAcc (t(10)=5.10, p≤0.0005), but not in the DS (t(12)=1.26 n.s.). High-fat-fed obese DIO rats did not differ from their lean counterpart in either D2R or TH mRNA levels in any brain area (VTA D2R: t(11)=0.01, n.s.; DS D2R: t(11)=0.65 n.s.; NAcc D2R: t(10)=1.65, n.s; VTA TH: t(10)=0.38 n.s.).
Body composition analysis
DIO rats were significantly smaller upon arrival compared to the DR rats, despite being of the exact same age (mean ± SEM, DIO: 266 ±5.0 g; DR: 289 ± 8.6 g; t(18)=2.28, p≤0.05; not shown). However, DIO rats gained weight faster compared to DR rats over a 2–month period on chow (Genotype*Time: F[1,16]=14.66, p≤0.001; Genotype: F[1,16]=1.57, n.s.). The body weights of chow-fed DIO and DR rats did not significantly differ at sacrifice (mean ± SEM, DIO: 357 ± 4.6 g; DR: 370 ± 10.2 g; t(18)=1.16, n.s.).
As previously shown (Cottone et al. 2007), chow-fed DIO rats had smaller whole carcass adiposity, measured as either grams or percentages of body weight, compared to DR. Fat pad analysis revealed that DIO subcutaneous fat pads were significantly smaller compared to DR rats (t(18)=4.05, p≤0.001, Table 1). Inguinal, mesenteric, and the brown fat pads from chow-fed DIO rats tended to be smaller compared to DR (t(18)>1.89, p≤0.07). Thus, the adiposity values in both genotypes were consistent with a lean state (Cottone et al. 2007).
Table 1.
Body weight and adiposity in chow-fed (n=12/genotype) and high-fat-fed (n=9–10/genotype) DIO and DR rats.
| Parameters | Chow | High-fat | ||
|---|---|---|---|---|
| DR | DIO | DR | DIO | |
| Carcass weight (g) | 338.4±9.2 | 325±4.9 | 572.7±31.8 | 750.7±31.5 *** |
| White fat pad (g) | ||||
| Inguinal | 8.2±0.6 | 6.8±0.4 | 27.8±4.2 | 55.5±4.1 *** |
| Subcutaneous | 5.4±0.2 | 4.0±0.3 *** | 18.2±3.7 | 44.9±5.1 *** |
| Mesenteric | 4.4±0.4 | 3.5±0.2 | 13.6±3.1 | 26.4±2.9 ** |
| Gonadal | 3.6±0.2 | 3.8±0.2 | 16.1±2.1 | 20.9±1.6 |
| Retroperitoneal | 4.1±0.4 | 3.5±0.3 | 18.8±2.6 | 36.2±3.2 *** |
| Total | 25.8±0.8 | 21.6±0.6 | 94.5±2.4 | 184.0±6.2 * |
| Brown fat pad (g) | 0.3±0.07 | 0.3±0.02 | 0.7±0.1 | 0.9±0.1 * |
| White fat pad (% body weight) | ||||
| Inguinal | 2.4±0.1 | 2.1±0.1 | 4.7±0.5 | 7.5±0.3 *** |
| Subcutaneous | 1.6±0.05 | 1.2±0.08 *** | 3.0±0.5 | 5.6±0.6 *** |
| Mesenteric | 1.3±0.1 | 1.1±0.04 | 2.3±0.4 | 3.4±0.3 * |
| Gonadal | 1.1±0.05 | 1.2±0.1 | 2.7±0.3 | 2.7±0.1 |
| Retroperitoneal | 1.2±0.1 | 1.1±0.07 | 3.2±0.3 | 4.7±0.3 *** |
| Total | 7.6±0.2 | 6.7±0.2 | 0.1±0.003 | 0.1±0.01 |
| Brown fat pad (% body weight) | 0.1±0.01 | 0.09±0.004 | 16.0±0.4 | 24.3±0.07 *** |
| Whole carcass adiposity | ||||
| Fat (g) | 27.64±2.1 | 22.37±1.1 * | 99.6±15.8 | 215.8±18.8 *** |
| Fat (% body weight) | 9.1±0.5 | 7.6±0.4 * | 17.8±2.2 | 29.3±1.6 *** |
Values represent M±SEM.
p≤0.05,
p≤0.01,
p≤0.001 vs. DR rats.
During the 4-weeks of exposure on high-fat diet, DIO rats disproportionally gained more weight compared to DR (Genotype*Time: F[1,16]=22.49, p≤0.001; Genotype: F[1,16]=14.57, p≤0.001) (4-week body weight gain, mean ± SEM, DIO: 101.8 ± 6.3 g; DR: 54.19 ± 8.0 g; t(16)=4.05, p≤0.001; not shown). At the time of sacrifice the high-fat-fed DIO rats were significantly heavier than high-fat-fed DR (mean ± SEM, DIO: 804 ± 32.5 g; DR: 626 ± 32.5 g; t(17)=3.86, p≤0.001; not shown).
As shown in Table 1, the fat pad analysis revealed that all the DIO adiposity parameters analyzed were significantly higher when compared to DR (t(16)=2.47, p≤0.03) consistent with the development of the adipose state (Levin 1999; Levin et al. 1997).
DISCUSSION
The results of the present study were as follows: (I) before the onset of obesity DIO rats show an innate decreased sensitivity to the effects of D-amphetamine on ICSS threshold and feeding, as compared to DR rats; (II) In a diet-induced obesity state, DIO rats show insensitivity to the D-amphetamine effect on ICSS, but show equal sensitivity to its anorectic effects, compared to lean DR; (III) DIO rats show differences in gene expression of components of the dopaminergic system compared to DR rats, both before and after the development of obesity.
By releasing dopamine and inhibiting uptake, the indirect agonist D-amphetamine potentiates the rewarding properties of electrical brain stimulation, observed as a decrease in the ICSS threshold (Esposito et al. 1980; Goodall and Carey 1975). In this study we showed that chow-fed DIO rats, in a pre-obesity state, exhibit an innate reduced sensitivity to the effects of D-amphetamine on ICSS threshold, as compared to DR rats. Specifically, D-amphetamine was less potent in lean DIO rats than in DR. Therefore these results reveal that a deficit in the brain reward function happens before the onset of obesity in DIO rats, being pre-existing and independent from the exposure to energy-dense food. Thus, an innate hypofunctional dopaminergic brain reward system could be a contributing factor in the development of obesity. In support of this hypothesis, brain imaging studies in obese patients reported decreased D2R density in striatal brain regions that was negatively correlated with the BMI (Guo et al. 2014; Wang et al. 2001), and a polymorphism of Taq I A1 allele of D2R has been associated with obesity in humans (Blum et al. 1996). However, while human studies present limitations in their ability to detect vulnerability factors which can predict the onset of diseases (O’Brien and Gardner 2005), preclinical animal studies allow a greater control of the experimental variables (Levin et al. 1997; Sabino et al. 2009; Velazquez-Sanchez et al. 2014).
Previous reports have suggested that hyposensitivity of the dopaminergic system is associated with obesity also in animal models. Multiple studies have shown in obese animals a lower brain reward function, decreased levels of components of the dopaminergic machinery in limbic areas of the brain, and/or a decreased responsiveness to dopamine receptors ligands (Alsio et al. 2010; Davis et al. 2008; Geiger et al. 2009; Johnson and Kenny 2010; Koyama et al. 2014; South and Huang 2008). Therefore, the difference between lean DIO and DR rats in ICSS threshold observed following D-amphetamine treatment provides further valuable information about which phenotypes may represent risk factors for the development of obesity, suggesting a causal relationship between the brain reward deficit and the predisposition to obesity.
In this study we observed that the resistance to the reward-enhancing effects of D-amphetamine in DIO rats was exacerbated following exposure to the high-fat diet, when rats were in an obvious obesity state, as demonstrated by the carcass analysis. While all doses lowered ICSS threshold in lean high-fat-fed DR rats, obese DIO rats did not respond to any. Based on these data, we can conclude that the decreased reward sensitivity, hypothesized as being a contributing factor for the development of obesity, results both from predisposing heritable factors and the interaction with high-energy foods (Johnson and Kenny 2010; Wang et al. 2001). These findings further corroborate the hypothesis that obesity is a polygenic disorder resulting from the interaction between an innate vulnerable phenotype and the increased availability of energy-dense food (Speliotes et al. 2010; Volkow et al. 2013a). The reduced responsivity of DIO to the potentiating effects of D-amphetamine on the brain reward system function but the absence of spontaneous decreases in brain reward function in DIO rats suggests that the reward deficiency in DIO rats becomes evident only after the system is challenged by a dopaminergic agent, and this result is in conflict with previous observations by Johnson and Kenny (2010) who demonstrated that obese rats overeating a cafeteria diet show elevated ICSS threshold compared to rats fed chow. Yet, a direct comparison between the two studies is difficult as that study was performed in outbred Wistar rats, while our study was performed in inbred Sprague-Dawley. Our study is in agreement with reports showing a lower response of both obesity-prone rats as well as Sprague–Dawley rats on a cafeteria-style diet to D-amphetamine-induced dopamine release in the NAcc (Geiger et al. 2008; Geiger et al. 2009).
The possible mechanism responsible for the reduced sensitivity to the reward potentiating effects of the dopaminergic agent D-Amphetamine may involve leptin. Dopaminergic neurons of the VTA were shown to be responsive to leptin, and the diminished locomotor response to amphetamine of ob/ob mice to be reversed by leptin (Fulton et al. 2006a). Therefore it can be hypothesized that the reduced dopaminergic sensitivity in DIO rats may be due to a defective central leptin signaling, which is present already before obesity onset.
It is noteworthy to mention that specific areas of the LH, region where the electrode was placed, may be differentially sensitive to diets and feeding signals compared to others (Fulton et al. 2006a); indeed, the reward produced by stimulating the LH is affected by food restriction and body weight, and it is attenuated by leptin (Fulton et al. 2006b; Shizgal et al. 2001). Therefore, slightly different histological location on the tip of the electrode could account for the variability of neuronal excitation along the medial forebrain bundle.
Similarly to what we observed in the ICSS, D-amphetamine decreased feeding less potently in lean, chow-fed DIO rats when compared to DR. The lower potency in the anorectic effect in DIO rats is not surprising. Indeed, resistance to multiple anorexigens was observed before obesity onset in obesity-prone rats (Irani et al. 2007; Levin et al. 2004; Levin et al. 2003). DIO rats show central insulin resistance and lower anorectic response to leptin even before obesity (Clegg et al. 2005; Levin et al. 2004). In addition, lean chow-fed DIO rats show decreased anorexia by CRF2 agonist stimulation compared to DR (Cottone et al. 2007). Another phenotype existing prior to the development of obesity is represented by profound differences in meal pattern in lean chow-fed DIO, compared to DR; DIO rats show signs of a preexisting deficit in the maintenance of post-meal satiety which resembles human ‘snacking’ behavior (Cottone et al. 2007). Interestingly, ‘snacking’ behavior, a phenotype typically observed in children, represents a risk factor for adult obesity (Francis et al. 2003). Therefore, dopamine hyposensitivity, together with altered metabolic factors may play a role in the development of obesity in DIO rats.
DIO and DR rats have previously been compared for spontaneous motor activity in an open field as well as for spontaneous physical activity in presence of a running wheel and in both cases no differences between genotypes were found while on chow diet (Levin 1991; Levin and Dunn-Meynell 2006)\\bumc.bu.edu\bumc\BUSM\Pharmacology\Dept\LAD\cottone-sabino\Valentina\My Office\My Papers\Submitted\DIO-DR Valenza paper\DIO DR_Psychopharmacol\Resubmission\Levin and - _ENREF_36. Furthermore, exposure to high-fat diet reduced running activity in equal degree in DIO and DR rats. These results suggest that the DIO’s vulnerability to obesity and their hyposensitive dopaminergic system is likely not associated with altered physical activity.
D-amphetamine treatment decreased high-fat diet intake of DIO and DR rats similarly. Interestingly, drug treatment was more potent in reducing body weight in the obese DIO rats as compared to lean DR. These findings are in line with previous observations obtained in a different model of obesity, which show that genetically obese Zucker rats are more sensitive to D-amphetamine anorexic effect than lean rats (Grinker et al. 1980). An interesting observation is the anorectic effect of D-amphetamine treatment was more potent, efficient, and long-lasting in both DIO and DR rats under a high-fat feeding regimen compared to when on standard chow diet. Since previous studies have shown a lack of interaction between the anorectic properties of amphetamine and diet palatability / energy density (Wellman et al. 1982), we hypothesize that the long-term exposure to the high-fat diet may have induced neuroadaptations in the dopaminergic system, which were responsible for the sensitized anorectic effect to D-amphetamine observed here. In support of this hypothesis are previous reports that repeated access to candies in baboons increases their sensitivity to the effects of D-amphetamine on the intake of both candies and regular food (Foltin 2011).
D-amphetamine is believed to exert its anorectic action by potentiating dopaminergic transmission in the hypothalamus, where it plays an important role in the regulation of food intake (Leibowitz 1975a; Vucetic and Reyes 2010). In addition, a role for central noradrenergic systems in the anorectic effects of amphetamine has been proposed (Ahlskog 1974; Leibowitz 1975b; Sahakian et al. 1983). Therefore, our results reveal that the effects of D-amphetamine on ICSS and feeding are dissociable in the context of obesity. While the development of obesity exacerbated the hyposensitivity to the rewarding effects of D-amphetamine, therefore enhancing differences between genotypes, on the other hand the exposure to the high-fat diet increased the drug’s anorectic properties.
In this study qPCR showed that lean, chow-fed DIO rats showed high levels of D1R and D2R mRNA in both the DS and the VTA as compared to chow-fed DR rats. In addition, gene expression of TH was higher in lean DIO rats vs. DR. These molecular data are not necessarily in contrast with the pharmacological data as they can be interpreted as compensatory responses to counterbalance the decreased availability of dopamine. qPCR results showed that under chow conditions the VTA DA synthetic enzyme, TH, was increased in DIO rats, compared to DR, suggesting an up-regulated, rather than down-regulated system as instead suggested by the behavioral experiments. While we cannot completely rule out the possibility that the mesolimbic DAergic system is up-regulated in DIO rats, it can also be hypothesized that the increase in TH is a compensatory response to the functional downstream hyposensitivity of the dopaminergic system(Volkow et al. 2008). Geiger and colleagues reported lower TH mRNA and protein expression in VTA primary neuronal cultures obtained by chow-fed DIO rats compared to cells derived from chow-fed DR rats (Geiger et al. 2008). However, differences in the sample source and processing could obviously account for this discrepancy. Notably, differences in D1R, D2R and TH gene expression observed in pre-obesity were not observed in obese DIO rats. The selectivity of these effects is in line with evidence showing that certain altered features are observed in lean, obesity-prone rats but normalize following development of obesity (e.g. NPY and alpha-2 adrenoreceptors expressions, norepinephrine turnover, etc.) (Levin 1999; Levin and Dunn-Meynell 2000). This phenomenon is believed to represent a neuroadaptative mechanism relevant for the defense of adiposity and body weight (Levin 1999). Therefore, neuroadaptations occurring after prolonged exposure to high-fat diet might also explain why D-amphetamine retained its anorectic effect during high-fat food intake experiment in both DIO and DR rats.
In high-fat-fed DIO rats, after obesity was fully developed, we found significantly lower D1R gene expression levels in both the NAcc and the VTA, compared to high-fat-fed DR rats. Our results confirm previous observations by Sharma and Fulton (2012) who reported decreased D1R protein expression in the NAcc of high-fat-fed DIO mice as compared to low-fat fed controls. Our results are also in agreement with those of Vucetic, showing reduced D1R in the VTA and the NAcc of obese mice (Vucetic et al. 2012; Vucetic and Reyes 2010). Based on the role that the mesolimbic D1R plays in mediating the effects of ICSS (Ikemoto and Panksepp 1999; Markou and Koob 1992; Steinberg et al. 2014), it can be hypothesized that the hyposensitivity of obese DIO rats to the reward-ehnancing effect of D-amphetamine be due to lower D1R expression.
Obese DIO rats did not show reduced D2R mRNA expression in the DS compared to high-fat fed DR rats. This observation contrasts some evidence collected in obese patients and rodents (Wang et al. 2001). A noticeable discrepancy can be found in literature in relation to the expression of D2R in animal models of obesity; for instance, Sharma and Fulton (2012) did not find differences in the expression of D2R in the DS, and two other studies have reported elevated D2R expression level in the DS of obesity-prone animals (Alsio et al. 2010; South and Huang 2008). Another important factor may be represented by the relative contribution of the two isoforms of D2R, short and long (Geiger et al. 2008; Johnson and Kenny 2010). Indeed, Johnson and Kenny (2010) reported that rats fed a cafeteria diet show a lower striatal protein expression of the mature and fully glycosilated (70 KDa) membrane-bound D2R isoform compared to chow-fed rats, while no differences in the expression of either the unglycosylated immature (D2S, ~39 kDa) or the intermediate glycosylated cytoplasmic (D2L, ~51 kDa) forms of D2R were found. Another difference with previous reports is represented by the fact that differences in the expression of the receptor protein were assessed, while the present study investigated mRNA levels. Therefore, further studies are needed in order to better elucidate the role of D2R in the mesolimbic system in obesity.
In contrast, no differences in the gene expression levels of dopaminergic receptors were found in either LH or VMH between the two genotypes under chow or high-fat diet. Therefore, the differential anorectic effect of D-amphetamine on chow intake in DIO and DR rats may be not related to hypothalamic levels of these receptors.
One limitation of the present study is that the samples from either chow-fed or high-fat fed DIO/DR rats were analyzed in separate qPCR experiments, therefore making a direct comparison between expression levels under chow and high-fat condition not possible. Even though qPCR is considered a techniques which provides absolute measures of gene expression, measured are deeply affected by inter-assay differences. Hence while the present mRNA data provide important information regarding putative difference between genotypes under either feeding condition, they do not allow drawing conclusions about the effect that the high-fat diet exposure had on gene expression in each genotype.
In conclusion, our data suggests that obesity-prone rats show a heritable, pre-existing deficit of the brain reward system, which is exacerbated by the exposure to a high-fat diet and the development of obesity. A better understanding of pre-existing dysfunctions in the dopaminergic brain reward system and its interaction with energy-dense food gives new insights into the disease pathogenesis and may yield new therapeutic opportunities.
Supplementary Material
Acknowledgments
We thank Aditi R. Narayan, Jina Kwak and Aditya Khedkar for technical assistance, and Andrew Kim, Jeffrey Santos and Angela Tung for editorial assistance. This publication was made possible by grant numbers DA030425, MH091945, and MH093650A1 from the National Institute on Drug Abuse (NIDA), and the National Institute of Mental Health (NIMH), and by the Peter Paul Career Development Professorship (P.C.). Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the National Institutes of Health. We gratefully acknowledge Dr. Timothy R. Nagy, Maria S. Johnson and the UAB Small Animal Phenotyping Core for performing the rat carcass analysis, which was founded by grant numbers P30DK056336 (NORC) and P60DK079626 (DRTC).
Footnotes
The authors declare no conflict of interest.
Supplementary information is available online.
References
- Ahlskog JE. Food intake and amphetamine anorexia after selective forebrain norepinephrine loss. Brain research. 1974;82:211–40. doi: 10.1016/0006-8993(74)90600-3. [DOI] [PubMed] [Google Scholar]
- Alsio J, Olszewski PK, Norback AH, Gunnarsson ZE, Levine AS, Pickering C, Schioth HB. Dopamine D1 receptor gene expression decreases in the nucleus accumbens upon long-term exposure to palatable food and differs depending on diet-induced obesity phenotype in rats. Neuroscience. 2010;171:779–87. doi: 10.1016/j.neuroscience.2010.09.046. [DOI] [PubMed] [Google Scholar]
- Baiamonte BA, Valenza M, Roltsch EA, Whitaker AM, Baynes BB, Sabino V, Gilpin NW. Nicotine dependence produces hyperalgesia: Role of corticotropin-releasing factor-1 receptors (CRF1Rs) in the central amygdala (CeA) Neuropharmacology. 2014;77:217–23. doi: 10.1016/j.neuropharm.2013.09.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blum K, Braverman ER, Wood RC, Gill J, Li C, Chen TJ, Taub M, Montgomery AR, Sheridan PJ, Cull JG. Increased prevalence of the Taq I A1 allele of the dopamine receptor gene (DRD2) in obesity with comorbid substance use disorder: a preliminary report. Pharmacogenetics. 1996;6:297–305. doi: 10.1097/00008571-199608000-00003. [DOI] [PubMed] [Google Scholar]
- Bray GA. Medical consequences of obesity. The Journal of clinical endocrinology and metabolism. 2004;89:2583–9. doi: 10.1210/jc.2004-0535. [DOI] [PubMed] [Google Scholar]
- Clegg DJ, Benoit SC, Reed JA, Woods SC, Dunn-Meynell A, Levin BE. Reduced anorexic effects of insulin in obesity-prone rats fed a moderate-fat diet. American journal of physiology Regulatory, integrative and comparative physiology. 2005;288:R981–6. doi: 10.1152/ajpregu.00675.2004. [DOI] [PubMed] [Google Scholar]
- Cottone P, Sabino V, Nagy TR, Coscina DV, Levin BE, Zorrilla EP. Centrally administered urocortin 2 decreases gorging on high-fat diet in both diet-induced obesity-prone and -resistant rats. Int J Obes (Lond) 2013;37:1515–23. doi: 10.1038/ijo.2013.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cottone P, Sabino V, Nagy TR, Coscina DV, Zorrilla EP. Feeding microstructure in diet-induced obesity susceptible versus resistant rats: central effects of urocortin 2. The Journal of physiology. 2007;583:487–504. doi: 10.1113/jphysiol.2007.138867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cottone P, Wang X, Park JW, Valenza M, Blasio A, Kwak J, Iyer MR, Steardo L, Rice KC, Hayashi T, Sabino V. Antagonism of Sigma-1 Receptors Blocks Compulsive-Like Eating. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology. 2012 doi: 10.1038/npp.2012.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis JF, Tracy AL, Schurdak JD, Tschop MH, Lipton JW, Clegg DJ, Benoit SC. Exposure to elevated levels of dietary fat attenuates psychostimulant reward and mesolimbic dopamine turnover in the rat. Behavioral neuroscience. 2008;122:1257–63. doi: 10.1037/a0013111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dore R, Iemolo A, Smith KL, Wang X, Cottone P, Sabino V. CRF mediates the anxiogenic and anti-rewarding, but not the anorectic effects of PACAP. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology. 2013;38:2160–9. doi: 10.1038/npp.2013.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esposito R, Kornetsky C. Morphine lowering of self-stimulation thresholds: lack of tolerance with long-term administration. Science. 1977;195:189–91. doi: 10.1126/science.831268. [DOI] [PubMed] [Google Scholar]
- Esposito RU, Perry W, Kornetsky C. Effects of d-amphetamine and naloxone on brain stimulation reward. Psychopharmacology. 1980;69:187–91. doi: 10.1007/BF00427648. [DOI] [PubMed] [Google Scholar]
- Fleckenstein AE, Volz TJ, Riddle EL, Gibb JW, Hanson GR. New insights into the mechanism of action of amphetamines. Annual review of pharmacology and toxicology. 2007;47:681–98. doi: 10.1146/annurev.pharmtox.47.120505.105140. [DOI] [PubMed] [Google Scholar]
- Foltin RW. Consumption of palatable food decreases the anorectic effects of serotonergic, but not dopaminergic drugs in baboons. Physiology & behavior. 2011;103:493–500. doi: 10.1016/j.physbeh.2011.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis LA, Lee Y, Birch LL. Parental weight status and girls’ television viewing, snacking, and body mass indexes. Obesity research. 2003;11:143–51. doi: 10.1038/oby.2003.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fulton S, Pissios P, Manchon RP, Stiles L, Frank L, Pothos EN, Maratos-Flier E, Flier JS. Leptin regulation of the mesoaccumbens dopamine pathway. Neuron. 2006a;51:811–22. doi: 10.1016/j.neuron.2006.09.006. [DOI] [PubMed] [Google Scholar]
- Fulton S, Woodside B, Shizgal P. Potentiation of brain stimulation reward by weight loss: evidence for functional heterogeneity in brain reward circuitry. Behavioural brain research. 2006b;174:56–63. doi: 10.1016/j.bbr.2006.07.009. [DOI] [PubMed] [Google Scholar]
- Geiger BM, Behr GG, Frank LE, Caldera-Siu AD, Beinfeld MC, Kokkotou EG, Pothos EN. Evidence for defective mesolimbic dopamine exocytosis in obesity-prone rats. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2008;22:2740–6. doi: 10.1096/fj.08-110759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geiger BM, Haburcak M, Avena NM, Moyer MC, Hoebel BG, Pothos EN. Deficits of mesolimbic dopamine neurotransmission in rat dietary obesity. Neuroscience. 2009;159:1193–9. doi: 10.1016/j.neuroscience.2009.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodall EB, Carey RJ. Effects of d- versus l-amphetamine, food deprivation, and current intensity on self-stimulation of the lateral hypothalamus, substantia nigra, and medial frontal cortex of the rat. Journal of comparative and physiological psychology. 1975;89:1029–45. doi: 10.1037/h0077187. [DOI] [PubMed] [Google Scholar]
- Grilly DM, Loveland A. What is a “low dose” of d-amphetamine for inducing behavioral effects in laboratory rats? Psychopharmacology. 2001;153:155–69. doi: 10.1007/s002130000580. [DOI] [PubMed] [Google Scholar]
- Grinker JA, Drewnowski A, Enns M, Kissileff H. Effects of d-amphetamine and fenfluramine on feeding pattens and activity of obese and lean Zucker rats. Pharmacology, biochemistry, and behavior. 1980;12:265–75. doi: 10.1016/0091-3057(80)90367-6. [DOI] [PubMed] [Google Scholar]
- Guo J, Simmons WK, Herscovitch P, Martin A, Hall KD. Striatal dopamine D2-like receptor correlation patterns with human obesity and opportunistic eating behavior. Molecular psychiatry. 2014;19:1078–84. doi: 10.1038/mp.2014.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iemolo A, Valenza M, Tozier L, Knapp CM, Kornetsky C, Steardo L, Sabino V, Cottone P. Withdrawal from chronic, intermittent access to a highly palatable food induces depressive-like behavior in compulsive eating rats. Behavioural pharmacology. 2012;23:593–602. doi: 10.1097/FBP.0b013e328357697f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikemoto S, Panksepp J. The role of nucleus accumbens dopamine in motivated behavior: a unifying interpretation with special reference to reward-seeking. Brain research Brain research reviews. 1999;31:6–41. doi: 10.1016/s0165-0173(99)00023-5. [DOI] [PubMed] [Google Scholar]
- Irani BG, Dunn-Meynell AA, Levin BE. Altered hypothalamic leptin, insulin, and melanocortin binding associated with moderate-fat diet and predisposition to obesity. Endocrinology. 2007;148:310–6. doi: 10.1210/en.2006-1126. [DOI] [PubMed] [Google Scholar]
- Johnson PM, Kenny PJ. Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nature neuroscience. 2010;13:635–41. doi: 10.1038/nn.2519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelley AE, Berridge KC. The neuroscience of natural rewards: relevance to addictive drugs. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2002;22:3306–11. doi: 10.1523/JNEUROSCI.22-09-03306.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenny PJ, Markou A. Conditioned nicotine withdrawal profoundly decreases the activity of brain reward systems. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2005;25:6208–12. doi: 10.1523/JNEUROSCI.4785-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koob GF, Volkow ND. Neurocircuitry of addiction. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology. 2010;35:217–38. doi: 10.1038/npp.2009.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kornetsky C, Esposito RU. Euphorigenic drugs: effects on the reward pathways of the brain. Federation proceedings. 1979;38:2473–6. [PubMed] [Google Scholar]
- Koyama S, Mori M, Kanamaru S, Sazawa T, Miyazaki A, Terai H, Hirose S. Obesity attenuates D2 autoreceptor-mediated inhibition of putative ventral tegmental area dopaminergic neurons. Physiological reports. 2014;2:e12004. doi: 10.14814/phy2.12004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehnert T, Sonntag D, Konnopka A, Riedel-Heller S, Konig HH. Economic costs of overweight and obesity. Best practice & research Clinical endocrinology & metabolism. 2013;27:105–15. doi: 10.1016/j.beem.2013.01.002. [DOI] [PubMed] [Google Scholar]
- Leibowitz SF. Amphetamine: possible site and mode of action for producing anorexia in the rat. Brain research. 1975a;84:160–7. doi: 10.1016/0006-8993(75)90811-2. [DOI] [PubMed] [Google Scholar]
- Leibowitz SF. Catecholaminergic mechanisms of the lateral hypothalamus: their role in the mediation of amphetamine anorexia. Brain research. 1975b;98:529–45. doi: 10.1016/0006-8993(75)90371-6. [DOI] [PubMed] [Google Scholar]
- Levin BE. Spontaneous motor activity during the development and maintenance of diet-induced obesity in the rat. Physiology & behavior. 1991;50:573–81. doi: 10.1016/0031-9384(91)90548-3. [DOI] [PubMed] [Google Scholar]
- Levin BE. Arcuate NPY neurons and energy homeostasis in diet-induced obese and resistant rats. The American journal of physiology. 1999;276:R382–7. doi: 10.1152/ajpregu.1999.276.2.R382. [DOI] [PubMed] [Google Scholar]
- Levin BE, Dunn-Meynell AA. Defense of body weight against chronic caloric restriction in obesity-prone and -resistant rats. American journal of physiology Regulatory, integrative and comparative physiology. 2000;278:R231–7. doi: 10.1152/ajpregu.2000.278.1.R231. [DOI] [PubMed] [Google Scholar]
- Levin BE, Dunn-Meynell AA. Differential effects of exercise on body weight gain and adiposity in obesity-prone and -resistant rats. Int J Obes (Lond) 2006;30:722–7. doi: 10.1038/sj.ijo.0803192. [DOI] [PubMed] [Google Scholar]
- Levin BE, Dunn-Meynell AA, Balkan B, Keesey RE. Selective breeding for diet-induced obesity and resistance in Sprague-Dawley rats. The American journal of physiology. 1997;273:R725–30. doi: 10.1152/ajpregu.1997.273.2.R725. [DOI] [PubMed] [Google Scholar]
- Levin BE, Dunn-Meynell AA, Banks WA. Obesity-prone rats have normal blood-brain barrier transport but defective central leptin signaling before obesity onset. American journal of physiology Regulatory, integrative and comparative physiology. 2004;286:R143–50. doi: 10.1152/ajpregu.00393.2003. [DOI] [PubMed] [Google Scholar]
- Levin BE, Dunn-Meynell AA, Ricci MR, Cummings DE. Abnormalities of leptin and ghrelin regulation in obesity-prone juvenile rats. American journal of physiology Endocrinology and metabolism. 2003;285:E949–57. doi: 10.1152/ajpendo.00186.2003. [DOI] [PubMed] [Google Scholar]
- Madsen AN, Hansen G, Paulsen SJ, Lykkegaard K, Tang-Christensen M, Hansen HS, Levin BE, Larsen PJ, Knudsen LB, Fosgerau K, Vrang N. Long-term characterization of the diet-induced obese and diet-resistant rat model: a polygenetic rat model mimicking the human obesity syndrome. The Journal of endocrinology. 2010;206:287–96. doi: 10.1677/JOE-10-0004. [DOI] [PubMed] [Google Scholar]
- Markou A, Koob GF. Postcocaine anhedonia. An animal model of cocaine withdrawal. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology. 1991;4:17–26. [PubMed] [Google Scholar]
- Markou A, Koob GF. Bromocriptine reverses the elevation in intracranial self-stimulation thresholds observed in a rat model of cocaine withdrawal. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology. 1992;7:213–24. [PubMed] [Google Scholar]
- Mayer J, Thomas DW. Regulation of food intake and obesity. Science. 1967;156:328–37. doi: 10.1126/science.156.3773.328. [DOI] [PubMed] [Google Scholar]
- Melis M, Spiga S, Diana M. The dopamine hypothesis of drug addiction: hypodopaminergic state. International review of neurobiology. 2005;63:101–54. doi: 10.1016/S0074-7742(05)63005-X. [DOI] [PubMed] [Google Scholar]
- O’Brien CP, Gardner EL. Critical assessment of how to study addiction and its treatment: human and non-human animal models. Pharmacology & therapeutics. 2005;108:18–58. doi: 10.1016/j.pharmthera.2005.06.018. [DOI] [PubMed] [Google Scholar]
- Pellegrino LJ, Pellegrino AS, Cushman AJ. A stereotaxic atlas of the rat brain. 2. Plenum Press; New York: 1979. [Google Scholar]
- Ricci MR, Levin BE. Ontogeny of diet-induced obesity in selectively bred Sprague-Dawley rats. American journal of physiology Regulatory, integrative and comparative physiology. 2003;285:R610–8. doi: 10.1152/ajpregu.00235.2003. [DOI] [PubMed] [Google Scholar]
- Sabino V, Cottone P, Zhao Y, Steardo L, Koob GF, Zorrilla EP. Selective reduction of alcohol drinking in Sardinian alcohol-preferring rats by a sigma-1 receptor antagonist. Psychopharmacology. 2009;205:327–35. doi: 10.1007/s00213-009-1548-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sahakian BJ, Winn P, Robbins TW, Deeley RJ, Everitt BJ, Dunn LT, Wallace M, James WP. Changes in body weight and food-related behaviour induced by destruction of the ventral or dorsal noradrenergic bundle in the rat. Neuroscience. 1983;10:1405–20. doi: 10.1016/0306-4522(83)90122-7. [DOI] [PubMed] [Google Scholar]
- Schulteis G, Markou A, Cole M, Koob GF. Decreased brain reward produced by ethanol withdrawal. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:5880–4. doi: 10.1073/pnas.92.13.5880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma S, Fulton S. Diet-induced obesity promotes depressive-like behaviour that is associated with neural adaptations in brain reward circuitry. Int J Obes (Lond) 2012 doi: 10.1038/ijo.2012.48. [DOI] [PubMed] [Google Scholar]
- Shizgal P, Fulton S, Woodside B. Brain reward circuitry and the regulation of energy balance. International journal of obesity and related metabolic disorders : journal of the International Association for the Study of Obesity 25 Suppl. 2001;5:S17–21. doi: 10.1038/sj.ijo.0801906. [DOI] [PubMed] [Google Scholar]
- South T, Huang XF. High-fat diet exposure increases dopamine D2 receptor and decreases dopamine transporter receptor binding density in the nucleus accumbens and caudate putamen of mice. Neurochemical research. 2008;33:598–605. doi: 10.1007/s11064-007-9483-x. [DOI] [PubMed] [Google Scholar]
- Speakman JR. Obesity: the integrated roles of environment and genetics. The Journal of nutrition. 2004;134:2090S–2105S. doi: 10.1093/jn/134.8.2090S. [DOI] [PubMed] [Google Scholar]
- Speliotes EK, Willer CJ, Berndt SI, Monda KL, Thorleifsson G, Jackson AU, Lango Allen H, Lindgren CM, Luan J, Magi R, Randall JC, Vedantam S, Winkler TW, Qi L, Workalemahu T, Heid IM, Steinthorsdottir V, Stringham HM, Weedon MN, Wheeler E, Wood AR, Ferreira T, Weyant RJ, Segre AV, Estrada K, Liang L, Nemesh J, Park JH, Gustafsson S, Kilpelainen TO, Yang J, Bouatia-Naji N, Esko T, Feitosa MF, Kutalik Z, Mangino M, Raychaudhuri S, Scherag A, Smith AV, Welch R, Zhao JH, Aben KK, Absher DM, Amin N, Dixon AL, Fisher E, Glazer NL, Goddard ME, Heard-Costa NL, Hoesel V, Hottenga JJ, Johansson A, Johnson T, Ketkar S, Lamina C, Li S, Moffatt MF, Myers RH, Narisu N, Perry JR, Peters MJ, Preuss M, Ripatti S, Rivadeneira F, Sandholt C, Scott LJ, Timpson NJ, Tyrer JP, van Wingerden S, Watanabe RM, White CC, Wiklund F, Barlassina C, Chasman DI, Cooper MN, Jansson JO, Lawrence RW, Pellikka N, Prokopenko I, Shi J, Thiering E, Alavere H, Alibrandi MT, Almgren P, Arnold AM, Aspelund T, Atwood LD, Balkau B, Balmforth AJ, Bennett AJ, Ben-Shlomo Y, Bergman RN, Bergmann S, Biebermann H, Blakemore AI, Boes T, Bonnycastle LL, Bornstein SR, Brown MJ, Buchanan TA, Busonero F, Campbell H, Cappuccio FP, Cavalcanti-Proenca C, Chen YD, Chen CM, Chines PS, Clarke R, Coin L, Connell J, Day IN, den Heijer M, Duan J, Ebrahim S, Elliott P, Elosua R, Eiriksdottir G, Erdos MR, Eriksson JG, Facheris MF, Felix SB, Fischer-Posovszky P, Folsom AR, Friedrich N, Freimer NB, Fu M, Gaget S, Gejman PV, Geus EJ, Gieger C, Gjesing AP, Goel A, Goyette P, Grallert H, Grassler J, Greenawalt DM, Groves CJ, Gudnason V, Guiducci C, Hartikainen AL, Hassanali N, Hall AS, Havulinna AS, Hayward C, Heath AC, Hengstenberg C, Hicks AA, Hinney A, Hofman A, Homuth G, Hui J, Igl W, Iribarren C, Isomaa B, Jacobs KB, Jarick I, Jewell E, John U, Jorgensen T, Jousilahti P, Jula A, Kaakinen M, Kajantie E, Kaplan LM, Kathiresan S, Kettunen J, Kinnunen L, Knowles JW, Kolcic I, Konig IR, Koskinen S, Kovacs P, Kuusisto J, Kraft P, Kvaloy K, Laitinen J, Lantieri O, Lanzani C, Launer LJ, Lecoeur C, Lehtimaki T, Lettre G, Liu J, Lokki ML, Lorentzon M, Luben RN, Ludwig B, Manunta P, Marek D, Marre M, Martin NG, McArdle WL, McCarthy A, McKnight B, Meitinger T, Melander O, Meyre D, Midthjell K, Montgomery GW, Morken MA, Morris AP, Mulic R, Ngwa JS, Nelis M, Neville MJ, Nyholt DR, O’Donnell CJ, O’Rahilly S, Ong KK, Oostra B, Pare G, Parker AN, Perola M, Pichler I, Pietilainen KH, Platou CG, Polasek O, Pouta A, Rafelt S, Raitakari O, Rayner NW, Ridderstrale M, Rief W, Ruokonen A, Robertson NR, Rzehak P, Salomaa V, Sanders AR, Sandhu MS, Sanna S, Saramies J, Savolainen MJ, Scherag S, Schipf S, Schreiber S, Schunkert H, Silander K, Sinisalo J, Siscovick DS, Smit JH, Soranzo N, Sovio U, Stephens J, Surakka I, Swift AJ, Tammesoo ML, Tardif JC, Teder-Laving M, Teslovich TM, Thompson JR, Thomson B, Tonjes A, Tuomi T, van Meurs JB, van Ommen GJ, Vatin V, Viikari J, Visvikis-Siest S, Vitart V, Vogel CI, Voight BF, Waite LL, Wallaschofski H, Walters GB, Widen E, Wiegand S, Wild SH, Willemsen G, Witte DR, Witteman JC, Xu J, Zhang Q, Zgaga L, Ziegler A, Zitting P, Beilby JP, Farooqi IS, Hebebrand J, Huikuri HV, James AL, Kahonen M, Levinson DF, Macciardi F, Nieminen MS, Ohlsson C, Palmer LJ, Ridker PM, Stumvoll M, Beckmann JS, Boeing H, Boerwinkle E, Boomsma DI, Caulfield MJ, Chanock SJ, Collins FS, Cupples LA, Smith GD, Erdmann J, Froguel P, Gronberg H, Gyllensten U, Hall P, Hansen T, Harris TB, Hattersley AT, Hayes RB, Heinrich J, Hu FB, Hveem K, Illig T, Jarvelin MR, Kaprio J, Karpe F, Khaw KT, Kiemeney LA, Krude H, Laakso M, Lawlor DA, Metspalu A, Munroe PB, Ouwehand WH, Pedersen O, Penninx BW, Peters A, Pramstaller PP, Quertermous T, Reinehr T, Rissanen A, Rudan I, Samani NJ, Schwarz PE, Shuldiner AR, Spector TD, Tuomilehto J, Uda M, Uitterlinden A, Valle TT, Wabitsch M, Waeber G, Wareham NJ, Watkins H, Wilson JF, Wright AF, Zillikens MC, Chatterjee N, McCarroll SA, Purcell S, Schadt EE, Visscher PM, Assimes TL, Borecki IB, Deloukas P, Fox CS, Groop LC, Haritunians T, Hunter DJ, Kaplan RC, Mohlke KL, O’Connell JR, Peltonen L, Schlessinger D, Strachan DP, van Duijn CM, Wichmann HE, Frayling TM, Thorsteinsdottir U, Abecasis GR, Barroso I, Boehnke M, Stefansson K, North KE, McCarthy MI, Hirschhorn JN, Ingelsson E, Loos RJ. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nature genetics. 2010;42:937–48. doi: 10.1038/ng.686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinberg EE, Boivin JR, Saunders BT, Witten IB, Deisseroth K, Janak PH. Positive reinforcement mediated by midbrain dopamine neurons requires D1 and D2 receptor activation in the nucleus accumbens. PloS one. 2014;9:e94771. doi: 10.1371/journal.pone.0094771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sulzer D. How addictive drugs disrupt presynaptic dopamine neurotransmission. Neuron. 2011;69:628–49. doi: 10.1016/j.neuron.2011.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van de Giessen E, Celik F, Schweitzer DH, van den Brink W, Booij J. Dopamine D2/3 receptor availability and amphetamine-induced dopamine release in obesity. J Psychopharmacol. 2014;28:866–73. doi: 10.1177/0269881114531664. [DOI] [PubMed] [Google Scholar]
- Velazquez-Sanchez C, Ferragud A, Moore CF, Everitt BJ, Sabino V, Cottone P. High trait impulsivity predicts food addiction-like behavior in the rat. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology. 2014;39:2463–72. doi: 10.1038/npp.2014.98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Baler RD. Reward, dopamine and the control of food intake: implications for obesity. Trends in cognitive sciences. 2011;15:37–46. doi: 10.1016/j.tics.2010.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Fowler JS, Telang F. Overlapping neuronal circuits in addiction and obesity: evidence of systems pathology. Philosophical transactions of the Royal Society of London Series B, Biological sciences. 2008;363:3191–200. doi: 10.1098/rstb.2008.0107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Fowler JS, Tomasi D, Baler R. Food and drug reward: overlapping circuits in human obesity and addiction. Current topics in behavioral neurosciences. 2012;11:1–24. doi: 10.1007/7854_2011_169. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Tomasi D, Baler RD. The addictive dimensionality of obesity. Biological psychiatry. 2013a;73:811–8. doi: 10.1016/j.biopsych.2012.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Tomasi D, Baler RD. Obesity and addiction: neurobiological overlaps. Obesity reviews : an official journal of the International Association for the Study of Obesity. 2013b;14:2–18. doi: 10.1111/j.1467-789X.2012.01031.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vucetic Z, Carlin JL, Totoki K, Reyes TM. Epigenetic dysregulation of the dopamine system in diet-induced obesity. Journal of neurochemistry. 2012;120:891–8. doi: 10.1111/j.1471-4159.2012.07649.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vucetic Z, Reyes TM. Central dopaminergic circuitry controlling food intake and reward: implications for the regulation of obesity. Wiley interdisciplinary reviews Systems biology and medicine. 2010;2:577–93. doi: 10.1002/wsbm.77. [DOI] [PubMed] [Google Scholar]
- Wang GJ, Volkow ND, Logan J, Pappas NR, Wong CT, Zhu W, Netusil N, Fowler JS. Brain dopamine and obesity. Lancet. 2001;357:354–7. doi: 10.1016/s0140-6736(00)03643-6. [DOI] [PubMed] [Google Scholar]
- Wellman PJ, Pittenger DJ, Wikler KC. Diet palatability and amphetamine-induced anorexia. Physiol Psychol. 1982;10:117–121. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.