

Abstract
Interleukin-8 (IL-8) has been implicated in the pathogenesis of several human respiratory diseases, including tuberculosis (TB). Importantly and in direct relevance to the objectives of this report quite a few findings suggest that the presence of IL-8 may be beneficial for the host. IL-8 may aid with mounting an adequate response during infection with Mycobacterium tuberculosis (M. tb); however, the underlying mechanism remains largely unknown. The major goal of our study was to investigate the contribution of IL-8 to the inflammatory processes that are typically elicited in patients with TB. We have shown for the first time that IL-8 can directly bind to tubercle bacilli. We have also demonstrated that association of IL-8 with M. tb molecules leads to the augmentation of the ability of leukocytes (neutrophils and macrophages) to phagocyte and kill these bacilli. In addition, we have shown that significant amount of IL-8 present in the blood of TB patients associates with erythrocytes. Finally, we have noted that IL-8 is the major chemokine responsible for recruiting T lymphocytes (CD3+, CD4+, and CD8+ T cells). In summary, our data suggest that the association of IL-8 with M. tb molecules may modify and possibly enhance the innate immune response in patients with TB.
1. Introduction
Mycobacterium tuberculosis (M. tb) is an infectious agent that claims about three million lives each year [1]. Pathological manifestations of tuberculosis (TB) result from disregulated inflammatory responses. In contrast to adaptive (acquired) immunity, developed after contact with an antigen and mediated by T lymphocytes, the innate immune system is not dependent on memory of a previous exposure and therefore has no specificity. Thus, neutrophils, monocytes, macrophages, and natural killer cells constitute cellular effectors of innate immunity. Impairment of the functional integrity of this pivotal arm of the immune system leads to an increase in host susceptibility to infection with M. tb [2].
Interleukin-8 (IL-8) displays two major biological activities: chemoattraction and activation of several types of white blood cells. These properties of IL-8 can have important clinical consequences by affecting the pathogenesis of severe infectious diseases, including mycobacterial infections such as TB. IL-8 plays a central role in normal immune response to M. tb and has been shown to be absolutely required for granuloma formation [3]. Monocytes and macrophages infected with M. tb may be primary producers of IL-8 during the course of TB [4–6]; however, neutrophils as well as respiratory epithelial cells also have the ability to secrete this chemokine [3, 7, 8]. Moreover, IL-8 is most likely responsible for bringing neutrophils to sites of infection in patients with TB; for example, bronchoalveolar (BAL) fluids from these patients show a dramatic increase in neutrophil numbers which correlates with elevated concentrations of IL-8 [9, 10]. Similarly, the extent of expression of IL-8 mRNA in tuberculous lymph nodes is proportional to neutrophil infiltration [11]. The consequences of high levels of IL-8 secretion during pulmonary tuberculosis include the accumulation of neutrophils and the recruitment of T lymphocytes and monocytes [3, 12]. In addition, plasma IL-8 concentrations are higher in patients who died from TB than in survivors [9, 10]. On the other hand, IL-8 is required for effective host defense against M. tb. For instance, because of its angiogenic properties IL-8 could contribute to the development of new vessels that are found at the margins of tuberculous cavities during the healing process [13]. Moreover, Friedland et al. [14] have demonstrated that the inability to stimulate production of IL-8 ex vivo correlated with poor prognosis in patients with TB.
It is known that several cytokines and growth factors have the ability to directly bind to bacterial molecules [15–22]. Therefore, we hypothesized that IL-8 could interact with M. tb and modulate the proinflammatory properties of this pathogen, especially since high concentrations of IL-8 are typically detected in patients with active TB [9, 10].
2. Methods
2.1. Human Subjects
All studies involving human blood and bronchoalveolar lavage fluids were approved by Human Subjects Investigation Committees of University of Tyler Health Science Center and University of Cincinnati Medical Center. Informed consent was obtained from both healthy volunteers and patients. The diagnosis of TB was confirmed by positive culture or using nucleic acid amplification testing (PCR). The diagnosis of MAC lung disease was based on criteria published by the American Thoracic Society/Infectious Diseases Society of America which have symptomatic, radiographic, and microbiologic components, the latter being the most important [23].
2.2. Purification of Monocytes and T Lymphocytes
White blood cells containing monocytes and lymphocytes were prepared from blood of normal donors by centrifugation through Ficoll Paque Plus (Pharmacia, Piscataway, NJ). Monocytes were then separated from other cells, mainly lymphocytes, by adherence to plastic. The adherent cells (monocytes) were incubated overnight with heat-killed M. tb (ATCC, Rockville, MD) to induce cytokine production. Then, conditioned media were collected and stored for further analysis. T lymphocytes were obtained by incubation of nonadherent cells with magnetic beads conjugated to CD3 (Dynal, Lake Success, NY). A magnetic cell separator was used to positively select CD3+ cells. CD4+ and CD8+ T lymphocytes were purified in a similar manner. The purity of the cells was tested by cytofluorometric analysis.
2.3. Chemotaxis
Chemotaxis of T lymphocytes was performed using Boyden chambers. Each chamber consisted of two compartments separated by a membrane. The lower compartment was filled with the stimulant, including conditioned media from M. tb stimulated monocytes and purified IL-8 alone (positive control). Then the membrane (a five-micron pore size polycarbonate filter, Nucleopore, Pleasanton, CA) was placed on the surface and the chamber was assembled. A 200 μL aliquot of the T cell preparation was added to the top of the filter (the top compartment), and the chambers were incubated at 37°C for 4 h. After the incubation, the chambers were centrifuged for 20 min at 1,200 rpm to achieve settling of T lymphocytes, which migrated through the filters, at the bottom of the lower chambers. Then, the filter was removed, and the cells were counted using a hemocytometer (according to the manufacturer's instructions). In some experiments conditioned media from stimulated monocytes were incubated overnight with an antibody against IL-8 (R&D Systems, Minneapolis, MN), or monocyte chemotactic protein-1 (MCP-1) (R&D Systems, Minneapolis, MN), or macrophage inflammatory protein-1α (MIP-1α) (Serotec Inc., Raleigh, NC), or control antibody (mouse IgG1; Sigma, Chemical Co., St. Louis, MO) prior to performing the chemotactic assay.
2.4. Neutrophil Infection and Viability Determination
Blood was drawn from healthy volunteers, and neutrophils were purified according to the protocol routinely used in our laboratory [24]. Neutrophils were maintained in RPMI-1640 media supplemented with 2 mM L-glutamic acid, 1 mM sodium pyruvate, and 10% fetal bovine serum (Sigma, St. Louis, MO) at 37°C and 5% CO2. Cells were infected with M. tbH37Rv (ATCC, Rockwille, MD) at MOI of 1 : 10 for 30 min. Unbound bacteria were washed off and the cells incubated for an additional 3 h. After 3 h, infected neutrophils were lysed with 1 mL of 0.1% sodium dodecyl sulfate (SDS) in PBS. Appropriate dilutions of cell lysates were plated onto Middlebrook 7H10 agar supplemented with 10% Oleic Albumin Dextrose Catalase (OADC) enrichment. After 21 days of culture, the number of colony forming units (CFU) was counted. Killing index was calculated as percent of CFU at 30 min according to the following formula: ([CFU at 30 min − CFU at 3 hr] × 100/CFU at 30 min). In some experiments, M. tbH37Rv were incubated with IL-8 (final concentration 100 ng/mL per 5 × 107 molecules of M. tb) overnight at 4°C, washed with PBS, and then used for infection.
2.5. Macrophage Infection and Viability Determination
Human monocytic leukemia cell line THP-1 (ATCC, Manassas, VA) was maintained in RPMI-1640 media supplemented with 2 mM L-glutamic acid, 1 mM sodium pyruvate, and 10% fetal bovine serum (Sigma, St. Louis, MO) at 37°C and 5% CO2. Prior to infection, THP-1 cells were exposed to 20 ng/mL phorbol-12-myristate-13-acetate (PMA) for 48 h to differentiate into macrophages (cells stop dividing and become adherent). THP-1 derived macrophages were infected with M. tbH37Rv (ATCC, Rockwille, MD) at MOI of 1 : 10 for 30 min. Unbound bacteria were washed off and the cells incubated for an additional 3 h. After 3 h, infected macrophages were lysed with 1 mL of 0.1% sodium dodecyl sulfate (SDS) in PBS. Appropriate dilutions of cell lysates were plated onto Middlebrook 7H10 agar supplemented with 10% Oleic Albumin Dextrose Catalase (OADC) enrichment. After 21 days of culture, the number of colony forming units (CFU) was counted. Killing index was calculated as percent of CFU at 30 min according to the following formula: ([CFU at 30 min − CFU at 3 hr] × 100/CFU at 30 min). In some experiments, M. tbH37Rv were incubated with IL-8 (final concentration 100 ng/mL per 5 × 107 molecules of M. tb) overnight at 4°C, washed with PBS, and then used for infection.
2.6. Respiratory Burst (Fluorescence Assay)
Neutrophils mounted on microscope slides and THP-1 derived macrophages growing on coverslips were incubated with anti-phospho-p40phox antibody (Sigma) followed by Alexa 568 conjugated secondary antibody. Stained cells were analyzed using a Nikon Eclipse TE2000-U inverted microscope with a UV filter set. Intensity scan was created using an Ultraview Program (Perkin Elmer, Waltham, MA).
2.7. Measurement of IL-8
IL-8 concentrations were measured in an ELISA assay using matched antibody pair (R&D Systems, Minneapolis, MN).
2.8. Western Blot
Western Blot was performed to detect binding of IL-8 to mycobacterial proteins. Bacterial extracts were loaded into a 4–15% gradient SDS-PAGE gel. After electrophoresis the gel was subjected to electrophoretic transfer to a nitrocellulose membrane. The membrane was then blocked and incubated with IL-8. Next, anti-IL-8 antibody was applied and followed by enhanced chemiluminescence (ECL) reagents (Biosource, Camarillo, CA). The membrane was then exposed to X-ray film (Fuji Super RX).
2.9. Interaction of IL-8 with M. tb Molecules
M. tb growing in Middlebrook 7H9 broth supplemented with OADC and 0.05% Tween 80 were incubated with IL-8 (100 ng/mL final concentration) overnight at 4°C. The presence of IL-8 attached to M. tb molecules was visualized using anti-IL-8 antibody followed by Alexa 568 conjugated secondary antibody. Heated killed M. tb molecules and bacteria not incubated with IL-8 served as controls. Fluorescence was analyzed using a Nikon Eclipse TE2000-U inverted microscope with a UV filter set.
2.10. Statistical Analysis
Differences between groups were analyzed by a simple one way analysis of variance (ANOVA), or if the data were not normally distributed by a Kruskal-Wallis ANOVA on ranks. The direct comparison between any two treatment groups was performed using Student's t-test, or the nonparametric Mann-Whitney test when the data sets were not normally distributed. A P value of 0.05 or less was considered significant. All statistics were performed using SIGMA STAT (SPSS Science Inc., Chicago, IL).
3. Results
3.1. Human IL-8 Interacts with Tubercle Bacilli
It has been previously reported that the concentration of IL-8 increases in patients with active tuberculosis [9, 10, 25]. Kurashima et al. [9], for example, showed that the level of IL-8 in BAL fluid from TB patients was as high as 559.7 pg/mg albumin. Moreover, some bacterial pathogens can associate with proinflammatory factors [15–22]. Based on these facts we hypothesized that IL-8 could interact with tubercle bacilli. We first incubated IL-8 with the bacilli as described in Section 2 and used fluorescent microscopy to detect IL-8 bound to M. tb molecules. IL-8 associated with bacteria was visualized with a specific anti-IL-8 antibody followed by a secondary antibody conjugated with Alexa 568. M. tb cells cultured without IL-8 served as a negative control (Figure 1, top histogram). Tubercle bacilli coated with IL-8 and incubated only with the secondary antibody were used to demonstrate the specificity of anti-IL-8 antibody (Figure 1, bottom histogram). Microscope analysis revealed the presence of IL-8 attached to M. tb molecules (Figure 1, red histogram). We also used thermally killed tubercle bacilli in which the protein components were denatured to determine whether IL-8 binds to a surface protein of M. tb. We found that heat killed bacteria bound no IL-8 (Figure 1, second histogram from the bottom). This observation suggests that the mycobacterial “IL-8 receptor” may be a protein.
Figure 1.

Binding of IL-8 to M. tb. Detection of IL-8 associated with M. tb using fluorescence microscopy.
To further confirm the ability of IL-8 to be associated with M. tb molecules, mycobacterial cell fractions were separated on a SDS PAGE gel. We noted that IL-8 bound to the membrane and whole cell lysate fractions but not to the cytosolic fraction (Figure 2). Detection of free IL-8 (Figure 2, left panel) served as a control of specificity of the anti-IL-8 antibody.
Figure 2.

Binding of IL-8 to M. tb. Detection of IL-8 associated with cellular fractions of M. tb using Western Blot. The vertical bar chart depicts densitometric analysis of protein bands with Quantity One 1D Analysis Software (Bio-Rad).
3.2. The Direct Association of IL-8 with M. tb Cells Leads to the Enhancement of the Ability of Inflammatory Cells to Phagocytose and Kill M. tb
Neutrophils are among the first cells attracted to a site of inflammation and play a critical role in the restricting of bacterial spread and in controlling the initial replication of bacteria [26]. Further, neutrophils which accumulate in the area of infection possess antimycobacterial qualities. These include the ability to phagocytize bacteria and elicit the oxidative burst which results in the production of reactive oxygen intermediates and degranulation, leading to the release of potent antimicrobial enzymes [12, 27, 28].
We tested proinflammatory functions of M. tb molecules coated with IL-8 using purified neutrophils. Neutrophils purified from blood of healthy volunteers [24] were infected with tubercle bacilli coated with IL-8 (100 ng of IL-8 per 5 × 107 bacteria) or not coated. An index of phagocytosis was determined after 30 min of infection using fluorescent microscopy (M. tb conjugated with fluorescein—FITC). At least 100 neutrophils were evaluated. We also assessed phagocytosis by counting colony forming units (CFU). We found that phagocytic uptake of IL-8/M. tb complexes by neutrophils was substantially enhanced (P < 0.01). Accordingly, the phagocytic index (number of ingested bacteria per cell) was 5.0 for IL-8-coated bacteria (M. tb/IL-8) and 3.0 for control bacteria (M. tb). Therefore, the association of IL-8 with M. tb triggered a substantial increase (approximately 1.5 times) in the ability of neutrophils to phagocyte bacteria (Figure 3(a)). Furthermore, analysis of 3 h infection indicated that the presence of IL-8 downregulated the infectious potential of M. tb. As shown in Figure 3(b), neutrophils killed bacilli opsonized with IL-8 more effectively than IL-8-free bacteria. Analysis of the CFU showed that the total number of M. tb was lower at both time points (30 min and 3 h) for bacteria preincubated with IL-8 in comparison to the number of IL-8-free M. tb (P < 0.05 and < 0.01, resp.) This observation was further confirmed by calculating killing index as shown in Figure 3(c) (P < 0.05).
Figure 3.

Effect of direct association of IL-8 with M. tb molecules on the ability of neutrophils to phagocytose and kill M. tb. (a) Phagocytosis of M. tb only (M. tb), or M. tb bound to IL-8 (M. tb/IL-8) presented as a phagocytosis index/increase fold over M. tb only. Image of neutrophil phagocyting M. tb conjugated with FITC photographed under fluorescent microscope. (b) Phagocytosis and killing of M. tb only (M. tb), or M. tb associated with IL-8 (M. tb/IL-8) calculated after 30 min and 3 h of infection using colony forming units (CFU) and presented as a CFU/fold over M. tb only. (c) Killing index (%) calculated for M. tb cytokine-free (M. tb) and M. tb associated with IL-8 (M. tb/IL-8) according to the formula ([CFU at 30 min − CFU at 3 h] × 100/CFU at 30 min).
Macrophages are the second major group of inflammatory cells involved in the innate immunity response against microorganisms. They are implicated in phagocytosing and killing mycobacteria and are considered the primary host cells for mycobacteria [29]. THP-1 cells have become one of the most widely used cell lines to investigate biological functions of monocytes and macrophages as they relate to various diseases, including TB [30]. In our studies, THP-1 cells were differentiated into macrophages with PMA and then infected with M. tb coated with IL-8 (100 ng of IL-8 per 5 × 107 bacteria) or uncoated bacteria. Analysis of phagocytic index showed that the association of IL-8 with M. tb enhanced the ability of macrophages to ingest bacteria 1.22 times but this difference did not reach statistical significance (P = 0.07; Figure 4(a)). The analysis of the CFU at 3 h indicated that the total number of M. tb/IL-8 was substantially (P < 0.001) decreased in comparison to the number of IL-8-free M. tb bacilli (Figure 4(b)). However, the killing index did not differ for these 2 groups as shown in Figure 4(c) (P = 0.054).
Figure 4.

Effect of direct association of IL-8 with M. tb molecules on the ability of THP-1 cells to phagocytose and kill M. tb. (a) Phagocytosis of M. tb only (M. tb), or M. tb bound to IL-8 (M. tb/IL-8) presented as a phagocytosis index/increase fold over M. tb only. Image of THP-1 cell phagocyting M. tb conjugated with FITC photographed under fluorescent microscope. (b) Phagocytosis and killing of M. tb only (M. tb), or M. tb associated with IL-8 (M. tb/IL-8) calculated after 30 min and 3 h of infection using colony forming units (CFU) method and presented as a CFU/fold over M. tb only. (c) Killing index (%) calculated for M. tb cytokine-free (M. tb) and M. tb associated with IL-8 (M. tb/IL-8) according to the formula ([CFU at 30 min − CFU at 3 h] × 100/CFU at 30 min).
The protein p40phox is a major constituent of NADPH-oxidase, which is a multicomponent enzyme system responsible for the oxidative burst [31, 32]. As a consequence of stimulation of inflammatory cells p40phox is phosphorylated and translocates from the cytosol to the cell membrane to form an enzymatic complex which produces oxygen radicals. Analysis of the level of phospho-p40phox in neutrophils and THP-1 derived macrophages infected with M. tb was performed using fluorescent microscopy. As shown in Figure 5(a), the presence of IL-8 associated with M. tb molecules caused the increase (approximately 1.38 times) in the level of pp40phox in neutrophils compared to chemokine-free bacteria but the difference was not statistically significant (P = 0.28). Similar observations were made using THP-1 derived macrophages stimulated with IL-8 coated or uncoated M. tb cells (Figure 5(b)). Analysis of fluorescence intensity showed that cells infected with M. tb opsonized with IL-8 appeared to be more potent (1.57 times) producers of phospho-p40phox (P < 0.001; Figure 5(b)).
Figure 5.

Respiratory burst presented as a level of pp40phox component detected using fluorescent microscopy. (a) Level of pp40phox molecule in neutrophils infected with M. tb cytokine-free (M. tb) or M. tb associated with IL-8 (M. tb/IL-8) presented as an intensity scan/increase fold over normal cells. (b) Level of pp40phox molecule in THP-1 cells infected with M. tb cytokine-free (M. tb) or M. tb associated with IL-8 (M. tb/IL-8) presented as an intensity scan/increase fold over normal cells.
3.3. IL-8 Concentration in Blood and Lung Fluids from TB Patients: Clinical Aspect of IL-8 in Tuberculosis
IL-8 concentrations have been measured by several investigators in both plasma and BAL fluids from TB patients. However, the results are quite variable [9, 10, 33–35]. Therefore we evaluated concentrations of this chemokine in blood and BAL fluids from patients with TB and for comparison in samples from patients with mycobacteriosis caused by Mycobacterium avium-complex (MAC). It has to be noted that IL-8 produced by white blood cells is present in blood in two forms: soluble (plasma IL-8) and cell-associated (IL-8 associated with red blood cells). It has been shown that a significant percentage of IL-8 in blood is associated with red blood cells. Erythrocytes express a receptor that binds multiple chemokines, including IL-8 [36, 37]. We measured both types of IL-8 in blood from 7 healthy volunteers, 14 TB patients, and 13 patients infected with MAC. We showed for the first time that levels of IL-8 associated with red blood cells from TB patients, MAC patients, and healthy donors were significantly different (P < 0.05) (Table 1). In agreement with previous reports [24] we found that IL-8 concentrations were significantly higher in plasma from TB patients than in plasma from normal subjects (P < 0.02) (Table 2). Our data also demonstrated (Table 2) that amounts of IL-8 were substantially elevated in plasma from patients with MAC compared to the amounts of this chemokine in plasma from normal subjects. Moreover, we evaluated concentrations of IL-8 in BAL fluids from TB patients, patients with MAC, and healthy subjects (Table 3). The levels of IL-8 were significantly higher (P < 0.001) in the two patient groups. Previous reports also demonstrated that lung fluids from TB patients contained significant amounts of IL-8 [9, 10]. We also noted that concentrations of IL-8 were lower in plasma than in bronchoalveolar lung lavage fluids.
Table 1.
Concentration of IL-8 (pg/mL) associated with red blood cells in human blood from: normal subjects (7 subjects), patients infected with MAC (13 patients), and patients infected with TB (14 patients).
| IL-8 associated with red blood cells (pg/mL) | ||
|---|---|---|
| Normal | MAC | TB |
| 53.8 ± 17.6 | 103.8 ± 82.5* | 125.3 ± 91.3** |
Values are means ± SD.
* P < 0.05 compared with normal.
** P < 0.05 compared with normal.
Table 2.
Concentration of IL-8 (pg/mL) in human samples; A/IL-8 in human plasma from: normal subjects (7 subjects), patients infected with Mycobacterium Avium Complex (MAC, 13 patients), and patients infected with TB (14 patients).
| IL-8 in plasma (pg/mL) | ||
|---|---|---|
| Normal | MAC | TB |
| 20.7 ± 26.2 | 64.0 ± 85.4* | 64.1 ± 77.6** |
Values are means ± SD.
* P < 0.02 compared with normal.
** P < 0.02 compared with normal.
Table 3.
Concentration of IL-8 (pg/mL) in lung fluids from: normal subjects (10 subjects), patients infected with MAC (10 patients), and patients infected with TB (15 patients).
| IL-8 in lung fluids (pg/mL) | ||
|---|---|---|
| Normal | MAC | TB |
| 4.0 ± 4.4 | 141.3 ± 334.6* | 365.6 ± 794.9** |
Values are means ± SD.
* P < 0.001 compared with normal.
** P < 0.001 compared with normal.
3.4. IL-8 Triggers Chemotaxis of T Lymphocytes
Since the role of IL-8 as a chemoattractant for T lymphocytes remains controversial [38–41], we evaluated the ability of IL-8 to recruit these cells. We observed that IL-8 induced a significant chemotaxis of T lymphocytes (Figure 6). This response was highly reproducible and dose dependent (Figure 6). Moreover, both CD4+ and CD8+ T cell subsets play a central role in developing resistance to M. tb [42]; thus, CD4+ and CD8+ T lymphocytes were also tested in the chemotactic assay (Figure 6). These cells were isolated using magnetic beads conjugated to CD4 or CD8 (purity of >95%). We found that both CD4+ and CD3+ T cells reached maximum response at an IL-8 concentration of 10−10 M (Figure 6). The response of CD8+ T lymphocytes was weaker than of other cell types and reached a maximum at an IL-8 concentration of 10−11 M (Figure 6).
Figure 6.
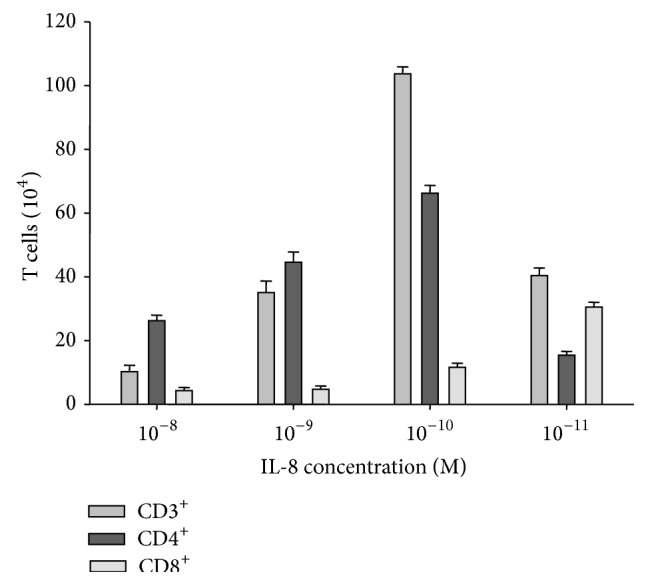
IL-8-induced chemotaxis of T lymphocytes.
TB is considered a granulomatous disease due to the accumulation of significant numbers of neutrophils, monocytes, and T lymphocytes in the infected area [2]. However, specific chemokines responsible for recruiting these cells have not yet been identified. We used conditioned media from human monocytes stimulated with heat-killed M. tb to evaluate the contribution of IL-8 to chemotactic responses of T lymphocytes. We noted that monocytes stimulated with M. tb produced approximately 100 ng/mL of IL-8. Conditioned media induced a significant chemotaxis of CD3+, CD4+, and CD8+ T cells. Furthermore, we found that the chemotactic response of T lymphocytes was substantially inhibited by a monoclonal anti-IL-8 antibody (P < 0.05) and not by the control antibody. To examine the possibility that other chemokines, which are produced by the stimulated monocytes, contribute to the migratory activity of the conditioned media, we tested specific neutralizing antibodies against chemokines with known activity towards T cells. These include macrophage inflammatory protein-1α (MIP-1α) and macrophage chemotactic protein-1 (MCP-1). Our results indicated that anti-MIP-1 α antibody or anti-MCP-1 antibody is less effective (P < 0.05) than anti-IL-8 antibody in suppressing the chemotactic response of CD3+ (Figure 7(a)) as well as CD4+ and CD8+ T lymphocytes (Figures 7(b) and 7(c)).
Figure 7.

Chemotaxis of CD3+ (a), CD4+ (b), and CD8+ (c) cells from a healthy subject triggered by conditioned media from M. tb stimulated monocytes. Effect of anti-IL-8, anti-MIP-1α, and anti-MCP-1 antibodies.
4. Discussion
The function of IL-8 in the pathogenesis of TB is controversial and a major goal in the following studies was to investigate the involvement of IL-8 in modulating the immune and inflammatory responses during infection with M. tb. There are several studies showing that some growth factors, cytokines, or other mediators have the ability to interact with bacterial pathogens affecting the course of inflammation [15–22]. C3-binding molecules, for example, were detected on the surface of several intracellular pathogens, including Legionella pneumophila, Chlamydia trachomatis, Leishmania, and Mycobacterium leprae [18–20]. Bermudez et al. [21, 22] studied epidermal growth factor-binding protein in Mycobacterium avium and M. tb. In addition, Luo et al. [16] investigated TNF-α binding to Shigella flexneri. We analyzed the interaction between IL-8 and tubercule bacilli and showed for the first time that IL-8 could directly associate with M. tb. We also found that as a consequence of this interaction antimicrobial activities of neutrophils and macrophages were enhanced. Moreover, the amount of IL-8 that was used to coat the bacteria appears to be physiologically relevant (discussed in the next paragraph).
There is some information in the literature concerning the concentration of IL-8 in plasma and BAL fluids of patients with active TB [9, 10, 25, 33–35]. Sadek et al. [10] showed that levels of IL-8 were increased in BAL fluid by 8.9-fold, whereas we found that IL-8 concentrations were increased 91 times in BAL fluids of patient with TB in comparison to healthy subjects (Table 3). It is important to mention that the increase of IL-8 concentration would be even more significant considering the fact that the BAL method dilutes alveolar fluids by 50- to 100-fold [43].
In agreement with previous findings [44] we have shown that the concentration of IL-8 in plasma and BAL fluids from patients with MAC disease was significantly increased in comparison to normal subjects. It should be noted that we are the first group to show that high concentration of IL-8 associates with red blood cells in TB patients and subjects with MAC disease.
There are a limited number of studies reporting the role of IL-8 as a chemoattractant for T cells [38–41]. Xu et al. [38] showed that IL-8 was as potent as RANTES, MIP-1α, and MIP-1β in inducing chemotaxis of CD3+ T lymphocytes. They tested several concentrations of IL-8 and found that the optimal concentration of IL-8 ranged from 10 to 50 ng/mL (1–6 × 10−9 M) [38]. We demonstrated that IL-8 present in culture media of monocytes stimulated with heat-killed M. tb was significantly more effective in inducing CD3+ T cell chemotaxis than MIP-1α and MCP-1 (Figure 7). This is in contrast to Sadek et al. [10], who reported similar activity for IL-8, RANTES, MCP-1, and MIP-1α. Furthermore, we observed that the chemotactic migration of CD3+ T cells was concentration dependent and reached a maximum response at an IL-8 concentration of 10−10 M (Figure 6).
Zachariae et al. [39] questioned the ability of IL-8 to attract CD4+ and CD8+ T lymphocytes equally well. In addition, the authors stated that IL-8 at concentration of 100 ng/mL (1.2 × 10−8 M) attracted CD8+ T lymphocytes more efficiently. Taub et al. [40] demonstrated that IL-8 was a potent T lymphocytes migratory agent in vivo in SCID mice. However the effect which IL-8 had on T lymphocytes chemotaxis in vitro was indirect. According to the authors [40] IL-8 induced neutrophil stimulation and degranulation which further caused T cells migration. The neutralization analysis showed that the majority of neutrophil granule-induced T cell migration was not due to chemokines [40]. We found that IL-8 was a potent attractant for CD4+ and CD8+ T lymphocytes and that migration of these cells towards IL-8 reached a maximum at concentrations of 10−10 M and 10−11 M for CD4+ and CD8+ T cells, respectively (Figure 6).
In summary, chemokines, including IL-8, are produced in lungs in response to M. tb infection. They induce immune cell recruitment to the lungs and play an important role in granuloma formation. They also act as key regulators of host defense against M. tb infection. Our novel observations indicate that IL-8 has the ability to directly interact with M. tb and in this way enhance antimicrobial functions of proinflammatory cells, that is, macrophages and neutrophils.
Acknowledgment
This paper was spported in part by a grant from National Science Center, Poland (UMO-2011/01/M/NZ6/01764).
Conflict of Interests
The authors declare no commercial or financial conflict of interests.
Authors' Contribution
Anna K. Kurdowska, Agnieszka Krupa, David Griffith, Robert P. Baughman, and Jaroslaw Dziadek were responsible for conception and design; Agnieszka Krupa, Marek Fol, Bozena R. Dziadek, Ewa Kepka, Dominika Wojciechowska, Anna Brzostek, and Agnieszka Torzewska were responsible for acquisition of data; Anna K. Kurdowska, Agnieszka Krupa, Marek Fol, and Bozena R. Dziadek were responsible for analysis and interpretation of data; David Griffith and Robert P. Baughman were responsible for providing patient samples; Anna K. Kurdowska and Agnieszka Krupa were responsible for the drafting of the paper; Anna K. Kurdowska, Agnieszka Krupa, and Jaroslaw Dziadek were responsible for the critical revision of the paper.
References
- 1.World Health Organization. WHO Report 2000. WHO/CDS/TB/2000.275. Geneva, Switzerland: World Health Organization; 2000. Global tuberculosis control. [Google Scholar]
- 2.Dannenberg A. M., Rook G. A. W. Pathogenesis of pulmonary tuberculosis: an interplay of tissue-damaging and macrophage-activating immune responses. In: Bloom B. R., editor. Tuberculosis Pathogenesis, Protection. Washington, DC, USA: ASM Press; 1994. pp. 459–483. [Google Scholar]
- 3.Miranda M. S., Breiman A., Allain S., Deknuydt F., Altare F. The tuberculous granuloma: an unsuccessful host defence mechanism providing a safety shelter for the bacteria? Clinical and Developmental Immunology. 2012;2012:14. doi: 10.1155/2012/139127.139127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zhang Y., Broser M., Cohen H., et al. Enhanced interleukin-8 release and gene expression in macrophages after exposure to Mycobacterium tuberculosis and its components. The Journal of Clinical Investigation. 1995;95(2):586–592. doi: 10.1172/jci117702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ameixa C., Friedland J. S. Interleukin-8 secretion from Mycobacterium tuberculosis-infected monocytes is regulated by protein tyrosine kinases but not by ERK1/2 or p38 mitogen-activated protein kinases. Infection and Immunity. 2002;70(8):4743–4746. doi: 10.1128/IAI.70.8.4743-4746.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Riedel D. D., Kaufmann S. H. E. Chemokine secretion by human polymorphonuclear granulocytes after stimulation with Mycobacterium tuberculosis and lipoarabinomannan. Infection and Immunity. 1997;65(11):4620–4623. doi: 10.1128/iai.65.11.4620-4623.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lin Y., Zhang M., Barnes P. F. Chemokine production by a human alveolar epithelial cell line in response to Mycobacterium tuberculosis . Infection and Immunity. 1998;66(3):1121–1126. doi: 10.1128/iai.66.3.1121-1126.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wickremasinghe M. I., Thomas L. H., Friedland J. S. Pulmonary epithelial cells are a source of IL-8 in the response to Mycobacterium tuberculosis: essential role of IL-1 from infected monocytes in a NF-κB-dependent network. The Journal of Immunology. 1999;163(7):3936–3947. [PubMed] [Google Scholar]
- 9.Kurashima K., Mukaida N., Fujimura M., et al. Elevated chemokine levels in bronchoalveolar lavage fluid of tuberculosis patients. American Journal of Respiratory and Critical Care Medicine. 1997;155(4):1474–1477. doi: 10.1164/ajrccm.155.4.9105097. [DOI] [PubMed] [Google Scholar]
- 10.Sadek M. I., Sada E., Toossi Z., Schwander S. K., Rich E. A. Chemokines induced by infection of mononuclear phagocytes with mycobacteria and present in lung alveoli during active pulmonary tuberculosis. American Journal of Respiratory Cell and Molecular Biology. 1998;19(3):513–521. doi: 10.1165/ajrcmb.19.3.2815. [DOI] [PubMed] [Google Scholar]
- 11.Bergeron A., Bonay M., Kambouchner M., et al. Cytokine patterns in tuberculous and sarcoid granulomas: correlations with histopathologic features of the granulomatous response. The Journal of Immunology. 1997;159(6):3034–3043. [PubMed] [Google Scholar]
- 12.Baggiolini M. Chemokines and leukocyte traffic. Nature. 1998;392(6676):565–568. doi: 10.1038/33340. [DOI] [PubMed] [Google Scholar]
- 13.Koch A. E., Polverini P. J., Kunkel S. L., et al. Interleukin-8 as a macrophage-derived mediator of angiogenesis. Science. 1992;258(5089):1798–1801. doi: 10.1126/science.1281554. [DOI] [PubMed] [Google Scholar]
- 14.Friedland J. S., Hartley J. C., Hartley C. G. C., Shattock R. J., Griffin G. E. Inhibition of ex vivo proinflammatory cytokine secretion in fatal Mycobacterium tuberculosis infection. Clinical & Experimental Immunology. 1995;100(2):233–238. doi: 10.1111/j.1365-2249.1995.tb03659.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wilson M., Seymour R., Henderson B. Bacterial perturbation of cytokine networks. Infection and Immunity. 1998;66(6):2401–2409. doi: 10.1128/iai.66.6.2401-2409.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Luo G., Niesel D. W., Shaban R. A., Grimm E. A., Klimpel G. R. Tumor necrosis factor alpha binding to bacteria: evidence for a high-affinity receptor and alteration of bacterial virulence properties. Infection and Immunity. 1993;61(3):830–835. doi: 10.1128/iai.61.3.830-835.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mueller-Ortiz S. L., Wanger A. R., Norris S. J. Mycobacterial protein HbhA binds human complement component C3. Infection and Immunity. 2001;69(12):7501–7511. doi: 10.1128/iai.69.12.7501-7511.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schlesinger L. S., Horwitz M. A. Phenolic glycolipid-1 of Mycobacterium leprae binds complement component C3 in serum and mediates phagocytosis by human monocytes. The Journal of Experimental Medicine. 1991;174(5):1031–1038. doi: 10.1084/jem.174.5.1031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bellinger-Kawahara C., Horwitz M. A. Complement component C3 fixes selectively to the major outer membrane protein (MOMP) of Legionella pneumophila and mediates phagocytosis of liposome-MOMP complexes by human monocytes. The Journal of Experimental Medicine. 1990;172(4):1201–1210. doi: 10.1084/jem.172.4.1201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hall R. T., Strugnell T., Wu X., Devine D. V., Stiver H. G. Characterization of kinetics and target proteins for binding of human complement component C3 to the surface-exposed outer membrane of Chlamydia trachomatis serovar L2. Infection and Immunity. 1993;61(5):1829–1834. doi: 10.1128/iai.61.5.1829-1834.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bermudez L. E., Petrofsky M., Shelton K. Epidermal growth factor-binding protein in Mycobacterium avium and Mycobacterium tuberculosis: a possible role in the mechanism of infection. Infection and Immunity. 1996;64(8):2917–2922. doi: 10.1128/iai.64.8.2917-2922.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Parker A. E., Bermudez L. E. Sequence and characterization of the glyceraldehyde-3-phosphate dehydrogenase of Mycobacterium avium: correlation with an epidermal growth factor binding protein. Microbial Pathogenesis. 2000;28(3):135–144. doi: 10.1006/mpat.1999.0335. [DOI] [PubMed] [Google Scholar]
- 23.Griffith D. E., Aksamit T., Brown-Elliott B. A., et al. An official ATS/IDSA statement: diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. American Journal of Respiratory and Critical Care Medicine. 2007;175(4):367–416. doi: 10.1164/rccm.200604-571st. [DOI] [PubMed] [Google Scholar]
- 24.Miller E. J., Kurdowska A., Nagao S., et al. A synthetic peptide which specifically inhibits heat-treated interleukin-8 binding and chemotaxis for neutrophils. Agents and Actions. 1993;40(3-4):200–208. doi: 10.1007/bf01984062. [DOI] [PubMed] [Google Scholar]
- 25.Algood H. M. S., Chan J., Flynn J. L. Chemokines and tuberculosis. Cytokine & Growth Factor Reviews. 2003;14(6):467–477. doi: 10.1016/s1359-6101(03)00054-6. [DOI] [PubMed] [Google Scholar]
- 26.Appelberg R. Mycobacterial infection primes T cells and macrophages for enhanced recruitment of neutrophils. Journal of Leukocyte Biology. 1992;51(5):472–477. doi: 10.1002/jlb.51.5.472. [DOI] [PubMed] [Google Scholar]
- 27.Berry M. P. R., Graham C. M., McNab F. W., et al. An interferon-inducible neutrophil-driven blood transcriptional signature in human tuberculosis. Nature. 2010;466(7309):973–977. doi: 10.1038/nature09247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Eum S.-Y., Kong J.-H., Hong M.-S., et al. Neutrophils are the predominant infected phagocytic cells in the airways of patients with active pulmonary TB. Chest. 2010;137(1):122–128. doi: 10.1378/chest.09-0903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Guirado E., Schlesinger L. S., Kaplan G. Macrophages in tuberculosis: friend or foe. Seminars in Immunopathology. 2013;35(5):563–583. doi: 10.1007/s00281-013-0388-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Theus S. A., Cave M. D., Eisenach K. D. Activated THP-1 cells: an attractive model for the assessment of intracellular growth rates of Mycobacterium tuberculosis isolates. Infection and Immunity. 2004;72(2):1169–1173. doi: 10.1128/iai.72.2.1169-1173.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Forman H. J., Torres M. Signaling by the respiratory burst in macrophages. IUBMB Life. 2001;51(6):365–371. doi: 10.1080/152165401753366122. [DOI] [PubMed] [Google Scholar]
- 32.Quinn M. T., Gauss K. A. Structure and regulation of the neutrophil respiratory burst oxidase: comparison with nonphagocyte oxidases. Journal of Leukocyte Biology. 2004;76(4):760–781. doi: 10.1189/jlb.0404216. [DOI] [PubMed] [Google Scholar]
- 33.Casarini M., Ameglio F., Alemanno L., et al. Cytokine levels correlate with a radiologic score in active pulmonary tuberculosis. American Journal of Respiratory and Critical Care Medicine. 1999;159(1):143–148. doi: 10.1164/ajrccm.159.1.9803066. [DOI] [PubMed] [Google Scholar]
- 34.Thillai M., Eberhardt C., Lewin A. M., et al. Sarcoidosis and tuberculosis cytokine profiles: indistinguishable in bronchoalveolar lavage but different in blood. PLoS ONE. 2012;7(7) doi: 10.1371/journal.pone.0038083.e38083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nakaya M., Yoneda T., Yoshikawa M., et al. The evaluation of interleukin-8 (IL-8) and tumor necrosis factor-α (TNF-α) level in peripheral blood of patients with active pulmonary tuberculosis. Kekkaku. 1995;70(8):461–466. [PubMed] [Google Scholar]
- 36.Darbonne W. C., Rice G. C., Mohler M. A., et al. Red blood cells are a sink for interleukin 8, a leukocyte chemotaxin. The Journal of Clinical Investigation. 1991;88(4):1362–1369. doi: 10.1172/jci115442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Neote K., Darbonne W., Ogez J., Horuk R., Schall T. J. Identification of a promiscuous inflammatory peptide receptor on the surface of red blood cells. The Journal of Biological Chemistry. 1993;268(17):12247–12249. [PubMed] [Google Scholar]
- 38.Xu L., Kelvin D. J., Ye G. Q., et al. Modulation of IL-8 receptor expression on purified human T lymphocytes is associated with changed chemotactic responses to IL-8. Journal of Leukocyte Biology. 1995;57(2):335–342. doi: 10.1002/jlb.57.2.335. [DOI] [PubMed] [Google Scholar]
- 39.Zachariae C. O. C., Jinquan T., Nielsen V., Kaltoft K., Thestrup-Pedersen K. Phenotypic determination of T-lymphocytes responding to chemotactic stimulation from fMLP, IL-8, human IL-10, and epidermal lymphocyte chemotactic factor. Archives of Dermatological Research. 1992;284(6):333–338. doi: 10.1007/bf00372035. [DOI] [PubMed] [Google Scholar]
- 40.Taub D. D., Anver M., Oppenheim J. J., Longo D. L., Murphy W. J. T lymphocyte recruitment by interleukin-8 (IL-8). IL-8-induced degranulation of neutrophils releases potent chemoattractants for human T lymphocytes both in vitro and in vivo. The Journal of Clinical Investigation. 1996;97(8):1931–1941. doi: 10.1172/jci118625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wilkinson P. C., Newman I. Identification of IL-8 as a locomotor attractant for activated human lymphocytes in mononuclear cell cultures with anti-CD3 or purified protein derivative of Mycobacterium tuberculosis . The Journal of Immunology. 1992;149(8):2689–2694. [PubMed] [Google Scholar]
- 42.Lalvani A., Brookes R., Wilkinson R. J., et al. Human cytolytic and interferon γ-secreting CD8+ T lymphocytes specific for Mycobacterium tuberculosis . Proceedings of the National Academy of Sciences of the United States of America. 1998;95(1):270–275. doi: 10.1073/pnas.95.1.270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Martin T. R., Goodman R. B. Chemokines in the Lung. Vol. 172. New York, NY, USA: CRC Press; 2003. Chemokines in acute lung injury; pp. 189–220. (Lung Biology in Health and Disease). [Google Scholar]
- 44.Yamazaki Y., Kubo K., Sekiguchi M., Honda T. Analysis of BAL fluid in M. avium-intracellulare infection in individuals without predisposing lung disease. European Respiratory Journal. 1998;11(6):1227–1231. doi: 10.1183/09031936.98.11061227. [DOI] [PubMed] [Google Scholar]