

Abstract
There are some who suggest that alcoholism and drug abuse are not diseases at all and that they are not consequences of a brain disorder as espoused recently by the American Society of Addiction Medicine (ASAM). Some would argue that addicts can quit on their own and moderate their alcohol and drug intake. When they present to a treatment program or enter the 12 Step Program & Fellowship, many addicts finally achieve complete abstinence. However, when controlled drinking fails, there may be successful alternatives that fit particular groups of individuals. In this expert opinion, we attempt to identify personal differences in recovery, by clarifying the molecular neurobiological basis of each step of the 12 Step Program. We explore the impact that the molecular neurobiological basis of the 12 steps can have on Reward Deficiency Syndrome (RDS) despite addiction risk gene polymorphisms. This exploration has already been accomplished in part by Blum and others in a 2013 Springer Neuroscience Brief. The purpose of this expert opinion is to briefly, outline the molecular neurobiological and genetic links, especially as they relate to the role of epigenetic changes that are possible in individuals who regularly attend AA meetings. It begs the question as to whether “12 steps programs and fellowship” does induce neuroplasticity and continued dopamine D2 receptor proliferation despite carrying hypodopaminergic type polymorphisms such as DRD2 A1 allele. “Like-minded” doctors of ASAM are cognizant that patients in treatment without the “psycho-social-spiritual trio,” may not be obtaining the important benefits afforded by adopting 12-step doctrines. Are we better off with coupling medical assisted treatment (MAT) that favors combining dopamine agonist modalities (DAM) as possible histone-deacetylase activators with the 12 steps followed by a program that embraces either one or the other? While there are many unanswered questions, at least we have reached a time when “science meets recovery,” and in doing so, can further redeem joy in recovery.
Keywords: 12 steps, Fellowship, Spirituality, Dopamine agonistic modalities (DAM), God, Genetic testing, Neuroepigenetics, Reward deficiency syndrome (RDS)
Introduction
The molecular neurobiological aspects of The Twelve Step Program adopted by self-help groups such as Alcoholics Anonymous (AA) and Narcotics Anonymous (NA) are the focus of this expert opinion. The purpose is to inform the addiction community that based on newly discovered functions of the reward circuitry of the brain, the neurobiological mechanisms at work within the 12-step doctrines may be understood.
We are cognizant that the one hundred alcoholics who developed these steps in the early to late 1930’s did so, empirically, without the tools of science we have today. The brain was a real mystery, very little was known about its workings especially the role of neurotransmitters and reward circuitry. Through the advent of 21st century science and medicine, especially neuroimaging technologies, science has finally caught up with “The 12 Step Program and Fellowship”. The mysteries that link the functioning of the brain and reward are being unraveled.
Understanding of the neuro-molecular biological underpinnings of The 12 steps and the work of various groups such as Al-Anon may indeed be a new and important step on the road to becoming and remaining clean and sober. Embracing principles of molecular neurobiology could ultimately lead to a better quality of life in recovery.
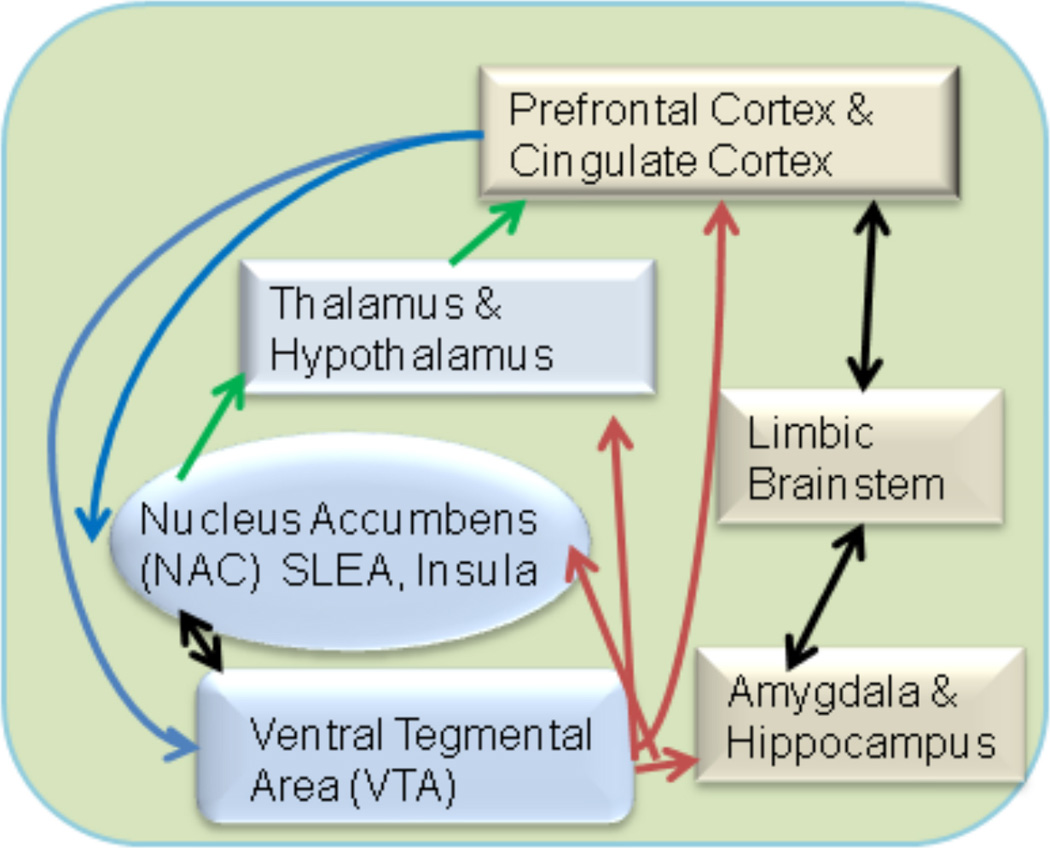
There is evidence that through the 12-step program and fellowship cross-talk between the Pre-Frontal Cortex-Cingulate (site of decision-making) and the Nucleus Accumbens (NAc) [site of craving behavior] is developed. Over half a century of dedicated and rigorous scientific research on the mesolimbic system provided insight into the neurogenetic mechanisms involved in the addictive brain and man’s quest for happiness. In brief, the site of the brain where one experiences feelings of well-being is called the Mesolimbic System and has been termed the reward center. The reward center is where chemical messages, including dopamine (DA), serotonin, enkephalins, and γ-aminobutyric acid (GABA), work together, to provide a net release of DA in the NAc. It is well known that genes control the synthesis, vesicular storage, metabolism, receptor formation, and catabolism of neurotransmitters [1–3]. Polymorphic variations in these genes can lead to an impairment of the neuronal events termed “The Brain Reward Cascade” that brings about the release of DA [4] (Figures 1a & 1b, 2, 3). A breakdown of this Cascade will lead to the dysregulation and dysfunction of DA homeostasis. Dopamine has been established as the pleasure and anti-stress molecule. Any reduction in DA function can result in a deficiency in reward that leads to substance seeking behavior [5].
Figure 1.
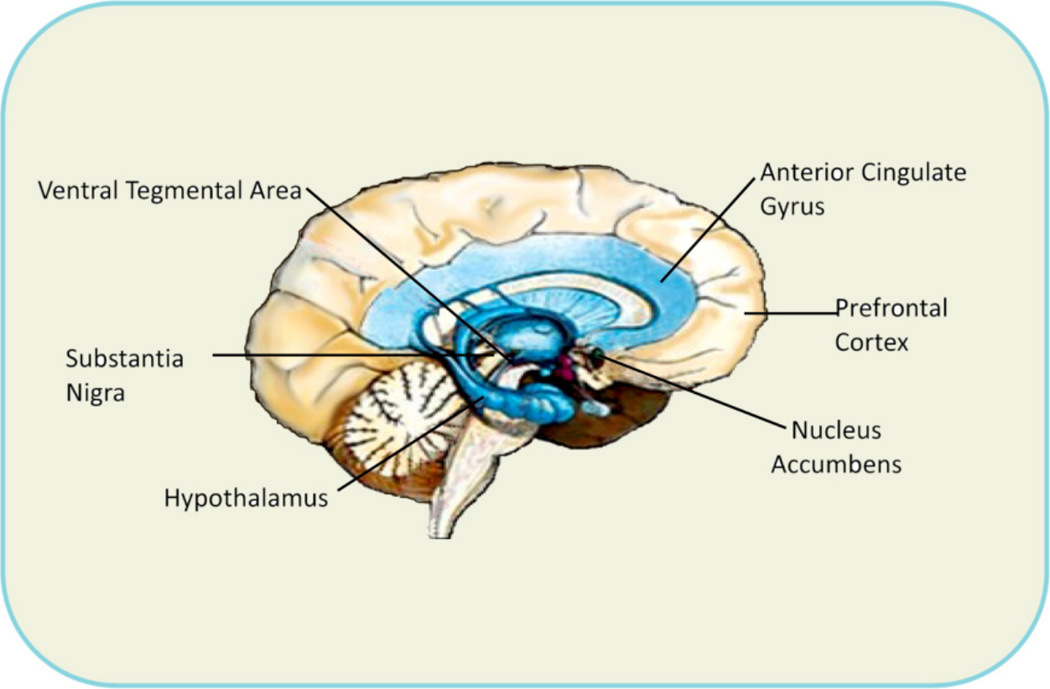
a: Brain Reward sites (Blum et al. [46] with permission).
b: Extended Brain Reward Circuitry (Blum et al. [46] with permission].
Figure 2.

Percent increase above rest of Dopamine release in the Claudate and Accumbens as Measured by Microdialysis.
Di Chiara G, Imperato A [15]-modified-Abbreviations: Me (Methadone); Mo (Morphine); A (Amphetamine); C (Cocaine); E (Ethanol); N (Nicotine) (with permission Blum et al. [46])
Figure 3.
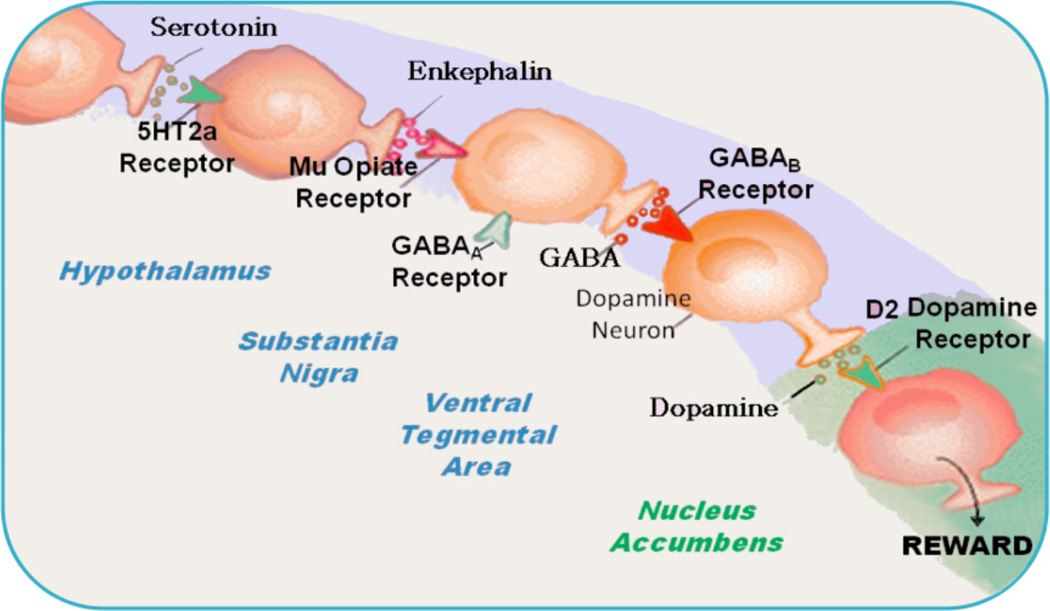
Brain Reward Cascade (with permission Blum et al. [46])
After 30,000 years, Homo sapiens are still evolving. We are biologically predisposed to drink, eat, reproduce, and desire pleasurable experiences. Humans have evolved rapidly; a few examples of recent traits are straight black hair, blue eyes, and lactose tolerance. The switch to agrarian from hunting and gathering based societies, allowed for new advantageous mutations due to enhanced reproduction. The human genome and future generations are likely to be mosaics of the past genome, due to epigenetics. The ability to build skyscrapers and cities suggest that we are different from our closest relative homo ergaster. While the brain of a chimp based on cognitive tests do as well as young children, the human brain size has quadrupled over 4 million years including structures around our reward system [6].
Impairment of the mechanisms involved in reward from these natural processes lead to multiple impulsive, compulsive, and addictive behaviors governed by genetic-polymorphic antecedents [7, 8]. There is a plethora of genetic variations at the level of mesolimbic activity. Polymorphisms of these genes are candidate genes, known to predispose individuals to excessive cravings and result in aberrant behaviors. They include; serotonergic 2A receptor (5-HTT2a); serotonergic transporter (5HTTLPR); DA D2 receptor (DRD2); DA D4 receptor (DRD4); DA transporter (DAT1); the catechol-O-methyltransferase (COMT), and monoamine-oxidase (MOA) genes [9].
In 1996, the term RDS was coined to define behaviors that associated with a common genetic variant involving DRD2 polymorphisms [10, 11] as a putative predictor of impulsive, compulsive, and addictive behaviors [12–14 [see Table 1].
Table 1.
Reward Deficiency Syndrome (from Blum et al. [45] with permission).
| Addictive Behaviors | Impulsive Behaviors | Obsessive Compulsive Behaviors |
Personality Disorders | ||
|---|---|---|---|---|---|
| Substance Related | Non Substance Related | Spectrum Disorders | Disruptive Impulsive | ||
| Alcohol | Thrill seeking (novelty) | Attention-deficit Hyperactivity |
Anti-social | Body Dysmorphic | Paranoid |
| Cannabis | Sexual Sadism | Tourette and Tic Syndrome |
Conduct | Hoarding | Schizoid |
| Opioids | Sexual Masochism | AuThism | Intermittent Explosive | Trichotillo-mania (hair pulling) | Borderline |
| Sedatives/Hypnotics | Hypersexual | Oppositional Defiant | Excoriation (skin picking) | Schizotypal | |
| Stimulants | Gambling | Exhibitionistic | Non-suicidal Self-Injury | Histrionic | |
| Tobacco | Internet Gaming | Narcissistic | |||
| Glucose | Avoidant | ||||
| Food | Dependent | ||||
Having genetic polymorphisms that, for example, result in reduced serotonergic and/or dopaminergic receptor densities or an increased rate of synaptic DA catabolism, because of high catabolic genotype of the COMT gene would have reduced DA availability. Drugs of abuse, are associated with the release of DA in the mesocorticolimbic system or reward pathway of the brain [15] [Figure 2].Without adequate DA function an individual would be predisposed to self medicate with any substance or behavior that will activate DA release including alcohol, nicotine, psychostimulants, opiates, glucose, sex, gambling, and even excessive internet gaming [16].
Dopaminergic system activation induces feelings of reward and pleasure [17, 18] impacted by epigenetic factors. However, hypodopaminergic functioning can, on the other hand, trigger drug-seeking and other RDS behaviors that seem to overlap [19–21]. Gene polymorphisms can induce hypodopaminergic functioning through, for example, reduced DA receptor density, blunted response to DA, or enhanced DA catabolism in the reward pathway [22]. Cessation of chronic drug use can also induce a hypodopaminergic state that prompts drug-seeking behaviors in an attempt to address the withdrawal-induced state [23].
Acute use of psychoactive substances can induce a feeling of well-being, sustained and prolonged abuse, unfortunately, leads to a toxic “high” and result in tolerance, disease, and discomfort. Thus, excessive cravings caused by carrying the DRD2 A1 allelic genotype that causes low DA receptors are compounded by consequential drug seeking behavior. Conversely, normal DA receptor densities, do not result in craving behaviors. The goal of preventing substance abuse or excessive glucose craving then may be accomplished in genetically prone individuals by the proliferation of DA D2 receptors [24]. In vitro, constant stimulation of the DA receptor system with a known D2 agonist in low doses results in significant D2 receptor proliferation despite genetic antecedents [25]. In essence, in the mesolimbic system D2 receptor stimulation signals negative feedback mechanisms to induce mRNA expression and causes the proliferation of D2 receptors. In humans, based on this molecular finding natural induction of DA release could be used to generate the same D2-directed mRNA to proliferate D2 receptors in order to, attenuate craving. In fact, this worked when a form of gene therapy; DNA-directed overexpression of the DRD2 receptors induced a significant reduction in both alcohol and cocaine craving behavior in animals [26–29].
The functional RDS hypothesis of drug-seeking and drug use is that regardless of its source, the presence of a hypodopaminergic state, is a primary cause of drug seeking behavior. Genetic polymorphisms that induce hypodopaminergic functioning are the primary causal mechanism of a predisposition to chronic drug use and relapse [30]. Long-term utilization of an approach that gently activates DA might become an effective and safe treatment for RDS behaviors including; substance use disorders (SUD), attention-deficit hyperactivity disorder (ADHD), and obesity.
Why Activate Dopamine?
Dopamine, as stated earlier, has been associated with pleasure and is the primary neurotransmitter modulating the activation of the reward system of the brain, It has been called the anti-stress molecule and the pleasure molecule [5, 31–33]. When DA is released into the synapse a number of receptors (D1-D5) are stimulated and feelings of well-being and stress reduction increase. The role of DRD2 gene in neuropsychiatric disorders and in alcoholism and other addictions [34], has been widely studied. Grasping the mechanism of motivated behavior and positive reinforcers requires an understanding of the neural circuitry of rewards [35].
A positive reinforcer is operationally defined as an event known to increase the probability of a subsequent positive response involving DA networks and drugs of abuse are considered to be stronger positive reinforcers than natural reinforcers (like food and sex) [36–38]. There is an important distinction between natural and unnatural rewards. Natural rewards include satisfaction of physiological drives (like hunger and reproduction and exploratory locomotion), and unnatural rewards are learned and involve satisfaction of acquired drives [39]. Acquired drives involve hedonic sensations and pleasure derived from alcohol, other drugs, as well as, from gambling and other risk-taking behaviors [2, 3, 36].
The reinforcing effects of drugs of abuse such as cocaine, alcohol, nicotine, food, and music are mediated in the NAc, a site within the ventral striatum. Indeed, it is believed that this structure directs motivated behaviors, elicited by natural rewards or incentive stimuli. The main tenets of positive reinforcement is that motor responses will increase in magnitude and vigor when followed by a rewarding event. Our hypothesis is that a mechanism of action for the powerful effects that drugs, music, food, and sex have on human motivation may be in part due to low DA function in the ventral striatum [40].
The human drive for the three necessary motivated behaviors, hunger, thirst, and sex, may all have common molecular-genetic antecedents that, if impaired, lead to aberrant behaviors. We hypothesize based on a plethora of scientific support that sexual activity like drugs, food, and music activates brain mesolimbic reward circuitry. Moreover, dopaminergic genes and possibly other candidate neurotransmitter-related genes and their polymorphisms affect both hedonic and anhedonic behavioral outcomes. As such we anticipate that future genetic studies of sex addiction will provide evidence for polymorphic associations with specific clustering of sexual typologies based on assessments using clinical instruments. We encourage both academic and clinical scientists to embark on neuroimaging studies of natural dopaminergic agonistic agents (like KB220Z™) to systematically target specific gene polymorphisms and normalize hyper-or hypo-sexual response [41–43].
Drug-microinjection studies have shown that opioids in brain reward regions especially in the Ventral Medial Striatum amplify the liking of sweet-taste rewards. Hedonic hot spots have been identified within the accumbens and pallidum using Fos plume mapping. These hotspots are where opioids are especially tuned to magnify the liking of food rewards. Hedonic hot spots in different brain structures may interact with each other within the larger functional circuitry that connects them [44]. Excessive hedonic liking for particular rewards might contribute to excessive consumption, and disorders such as RDS.
With this brief introduction to mesolimbic reward circuitry illustrated in Figure 3 it provides a framework for understanding the potential role of neurogenetics and neurotransmission, involving DA and the subsequent development of well-being. With this in mind we have explored the molecular neurobiology that may impact the 12 step doctrine as a model for recovery in earlier publications [45].
Alcoholics Anonymous/Narcotics Anonymous
Alcoholics Anonymous (AA) founded in 1935 by Bill Wilson and Dr. Bob Smith (Bill W. & Dr. Bob) in Akron Ohio is an international mutual aid movement. The primary stated purpose of AA is to encourage alcoholics “to stay sober and help other alcoholics achieve sobriety”. Wilson and Smith with help from other early members developed AA’s Twelve-Step program of spiritual and character development. In 1946, The Twelve Traditions were introduced to help AA strengthen and grow. The Traditions recommend that in public media groups and members remain anonymous, include all who wish to stop drinking and altruistically help other alcoholics. The Traditions also recommend that AA members, acting on behalf of the fellowship steer clear of dogma, involvement in public issues and governing hierarchies. Subsequent fellowships such as Narcotics Anonymous have adopted and adapted the Twelve Steps and the Twelve Traditions to their respective primary purposes [47].
Although, AA generally avoids discussing the medical nature of alcoholism AA is regarded as a proponent and popularizer of the older disease theory of alcoholism [48]. The American Psychiatric Association recommended AA’s program or similar community resources, in conjunction with sustained treatment for chronic alcoholics unresponsive to brief treatment. AA’s data states that 64% drop out of AA in their first year [49, 50].
AA membership since 1935 has spread “across diverse cultures holding different beliefs and values”, including geopolitical areas resistant to grass-root movements. AA claims more than 2 million members. While there is, a difference between the 12-step program and AA/NA fellowship both can play an important part in successful recovery. In this article we have interchanged the words “fellowship” and “program” because there are those that believe they are synonymous in a real true sense. AA’s name derived from its first book, informally called “The Big Book,” originally titled “Alcoholics Anonymous: The story of how more than one-hundred men have recovered from Alcoholism”. While there may be a real allergy to ethanol potentially due to one’s genetic makeup, people may not be doomed. We now know the importance of environmental impact on our polymorphic genes especially those involved in the brain reward circuitry. We are also aware that many people may be able to embrace the 12-step programs but although there is no magic bullet are we getting closer to “hatching the addiction egg”? [51, 52].
Addiction Epigenetics
An earlier simple understanding of genetics and environment held that P = G + E where P = any phenotype; G = Genes and E = environmental elements is the basis for understanding why we are not doomed because of our DNA polymorphisms. While it is believed that our genes contribute approximately 50–70% of the variance to RDS the environment seems to play a significant role in terms of gene expression and as such behaviors “normal” or aberrant”. Through extensive research during the last ten years we are beginning to understand the impact of the environment onto our genome [53].
Importantly, evidence indicates that epigenetic mechanisms are involved in drug addiction. Enzymes involved in chromatin remodeling have been recently studied. Simon-O’Brien et al. [54] found that histone deacetylase (HDAC) inhibitors (HDACi) had significant effects on ethanol intake and relapse. Specifically, they found that excessive alcohol intake of dependent (but not non-dependant) rats in the operant ethanol self-administration paradigm was significantly decreased by Sodium Butyrate (NaB) and MS-275. NaB reduced excessive drinking and prevented the escalation of ethanol intake in the intermittent access to 20% ethanol paradigm and completely blocked the increase of ethanol consumption induced by an alcohol deprivation. These results demonstrated a preventive effect of NaB on relapse.
In addition, Febo et al. [55] found that acute exposure to cocaine resulted in widespread BOLD activation in fore-and midbrain, however, chronic exposure did not. Pretreatment with the histone deacetylase inhibitor NaB restored BOLD signals in the forebrain after repeated cocaine exposure. Areas of activation included, the hippocampus/amygdala, various portions of limbic and sensory cortex and a pronounced activation in the anterior thalamus. These findings suggest that HDACi modulation after repeated stimulant exposure involves corticolimbic circuitry regulating emotion, motivation, and memory.
Since it is well-known that memory of the drug experience is an important cue for reinstatement of drug seeking and negative consequences also are cues to block reinstatement. In this regard Sen [56], reported that down-regulation of genes due to alterations in epigenetics leads to cognitive deficiencies that may play a role in the addictive process. Kenny’s group [57] suggest that there is evidence that DNA methylation plays a central role in these processes, likely by directly influencing the expression of genes involved in synaptic plasticity.
It is well-established that abuse of opiates, induce synaptic adaptation in a number of brain regions including ventral tegmental area (VTA). These adaptations may underlay the initiation and maintenance of opioid dependence and addiction in humans and animal models. Wang et al. [58], has shown that certain genes involved in glutaminergic function are altered by morphine. Through epigenetic mechanisms morphine alters a protein involved in postsynaptic density called “protein 95” (PSD-95). This protein is critically involved in the glutamatergic synaptic maturation and plasticity in the central neurons.
Scientists worldwide all agree that acute and chronic ethanol exposure may involve chromatin remodeling resulting from covalent histone modifications and DNA methylation in the neuronal circuits involving the amygdala brain region [59]. In this regard, Pandey et al. [60] revealed a novel role for amygdaloidal chromatin remodeling in the process of alcohol addiction. They further suggest that HDAC inhibitors may be potential therapeutic agents in treating alcohol withdrawal symptoms.
Importantly, microRNAs are small non-coding RNA molecules that regulate decrease or increase polypeptide formation as a function of mRNAs expression. They exert this function through base-pairing with partially complementary sequences in the 3’-UTR of target mRNAs. Since the first discovery of miR, lin-4 in Caenorhabditis Elegans, hundreds of miRs have been identified from humans in viruses, which have provided a pervasive layer of post-transcriptional gene regulation. The human nervous system is a rich source of miR expression, with a diversity of miR functions in fundamental neurobiological processes including neuronal development, plasticity, metabolism, and apoptosis and addiction [61]. It is also known that for example, expression of the NMDA receptor 2B (NR2B) gene is upregulated following chronic intermittent ethanol (CIE) treatment and withdrawal. This upregulation underlies behavioral alterations in addiction [62]. In some histone methyltransferases (HMTs) Qiang et al. [62] found a significant down-regulation at both the global level and the local chromatin of the NR2B gene following CIE treatment. In addition, it was also found that in the chromatin of the NR2B gene promoter, a decrease in G9a, Suv39 h1 and HDAC1-3 is responsible for the altered H3K9 modifications caused by CIE. Modifications in H3K9 show an increase in methylation with acute ethanol and a subsequent reduction of methylation during withdrawal with an increase in histone acetylation. Importantly, this is another example of how changes in H3K9 modifications in the local chromatin of the NR2B gene underlie alcohol-induced neuroadaptation. Moreover, Taqi et al. [63] revealed that following alcoholism in post-mortem tissue there was methylation in the alcoholics compared to controls. However, the methylation was found in the non-risk allele of prodynorphine-gene (PDYN). Thus, alcohol per se could affect even gene expression by altering the activation of PDYN transcription and vulnerability of individuals with the C, non-risk allele(s) to develop alcohol dependence.
There have been a number of animal studies showing that chronic cannabis smoking can result in molecular neurobiological modifications in the reward circuitry leading to prolonged behavioral problems. Further, this has now been confirmed by the recent work of Szutorisz et al. [64]. They found that parental exposure to the main psychoactive component of cannabis (not the same as grown cannabis) Δ(9)-tetrahydrocannabinol (THC), leads to compulsive heroin seeking behavior and changed striatal synaptic plasticity in subsequent generations. Germline THC exposure was found to decrease mRNA, with a concomitant reduction in NMDA receptor binding observed in the dorsal striatum of adult offspring. These results further suggest that THC exposure influences the molecular characteristics of the striatum and can impact offspring phenotype, leading to the augmented risk for psychiatric disorders in the subsequent generation through neuro epigenetic effects.
The effect of environmental elements on mRNA transcription is an important area of investigation. Specifically, MicroRNAs (miRNAs) are a type of non-protein-coding single-stranded RNA, typically 20–25 nt in length. Undoubtable, miRNAs play significant roles in many biological processes, including development, cell proliferation, differentiation, and apoptosis. Xu et al. [65] found that miR-212 expression level was constantly elevated during cocaine administration. Along similar lines Bahi & Dreyer [66] showed that striatal miR124a and BDNF signaling have crucial roles in alcohol consumption and ethanol conditioned reward. Zhang et al. [67] reported on the important role of miR-190 in the regulation of morphine function through its impact on OPRM1 expression.
The take-home message is that genetic variability may increase the risk of addictive behaviors in an individual and exposure to a drug results in neuroadaptations in interconnected brain circuits. In genetically susceptible individuals, these neuroadaptations underlie the transition to, and maintenance of, an addicted state. Moreover, these adaptations occur at the cellular, molecular, or epigenetic level and are associated with synaptic plasticity and modified gene expression. These effects on gene expression can occur via factors influencing translation (epigenetics) and transcription (non-coding micro RNAs) of the DNA or even RNA itself [68–70].
Are There Epigenetic Effects in the 12 Step Programs & Fellowship?
While it may be difficult to prove because of the fact that “anonymous” prevents potential real-time exploration, there are many aspects of this important addition to the recovery process that suggest that epigenetics may have profound neurobiological influences on the reward circuitry. In the book “Molecular Neurobiology of Recovery: 12 Steps Programs and Fellowship” Blum et al. [46] adequately addresses this issue.
As we pointed out, a growing body of evidence supports the claim that the AA and the 12-step programs do work for many but not for all. One interesting notation is that those who attend regular meetings seem to adapt to recovery with “a brand new psyche”. This possible outcome will translate to a new and improved life of sobriety and or clean time and acceptance of others without judgment. We believe that through fellowship there must be powerful epigenetic effects. The “love” of another possibly even through preferential release of the bonding chemical oxytocin may induce a “synaptic change” leading to a degree of new found happiness. In addition, Michael Meaney and associates of McGill University, showed that effects of maternal behavior are mediated to some degree, through epigenetics. Specifically, rat mothers that display high levels of nurturing behavior, licking and grooming their pups, result in offspring that are less anxious and produce less stress hormone than pup’s raised by less caring mothers. The basic reason for this involves differential levels of methylation linked to the glucocorticoid receptor in the hippocampus. Less caring caused more methylation and reduced receptor numbers. This results in enhanced production of cortisol, with concomitant exacerbated stress [71].
However, we must ask the following unanswered questions
Are there neuroplastic and lasting brain changes in regular meeting goers?
Is there preferential release of DA/oxytocin during AA attendance?
Is there proliferation of DRD2 receptors even in carriers of the DRD2 A1 (30–40% reduced D2 receptors) when these individuals regularly attend AA meetings;
Can we manipulate the stress level of recovering addicts with holistic approaches such as: KB220Z.to decrease methylation on the glucocorticord receptor. [KB220Z is a complex that activates functional connectivity of the brain even at rest, hyperbaric oxygenation, yoga, meditation, diet, exercise , music therapy, sound therapy, drum therapy, trauma relief therapy, cognitive behavioral therapy among other know modalities];
Through attendance at AA meetings can we actually proliferate D2 receptors?
D2 receptor proliferation is one example of inducing a neuroplasticity that could result in reduction of norepinephrine induced stress, reduce cravings, enhance decision-making, enhance social bonding, reduce immature defense styling (lying and or manipulation). It could also regulate Prefrontal cortices cingulate. to prevent relapse, increase focus, expand memory, augment self-esteem and confidence, reduce crime, reduce unprotected sex, enhance brain reward white/ grey matter density, and finally induce spirituality and new awakenings [45, 52].
So Does It Make Sense to Incorporate the 12-step Program & Fellowship along with Medical Assisted Treatment (MAT)?
We should also ask the important question-If the current FDA approved drugs favor blocking DA function; Why would we want to block DA in the long-term, especially while actively seeking help through the 12 steps? The answer to this conundrum is to at least embrace the concept that blocking DA is not the best approach, until we find an appropriate form of MAT with enhanced pleasure and anti-stress to assist those in recovery. One goal would be to reduce “white knuckle sobriety” in the recovering addict.
Importantly, there is now a molecular basis for the gateway hypothesis especially for nicotine [72]. It has always been thought that young people use drugs of abuse in stages and that certain drugs like marijuana and nicotine may be gateway substance leading to heavier psychoactive drugs such as heroin and cocaine [73]. We now know that nicotine induces “hyperacetylation” in the brain and alters the expression of FOSB, which is a trigger for addiction. It is also feasible that activation of histone deacetylase (HDAC) may have some benefit in treating addiction since by doing so can decrease FOSB expression in response to cocaine. While modifying HDAC activators to target specifically the striatum would be most desirable, we must be cognizant that systemic treatment with HDAC activators or histone acetyltransferase inhibitors could be dangerous especially for cognition. In addition, e-cigarettes may be equally harmful as smoking, except for cancer potential, because it contains pure nicotine and as such could act as a gateway. These facts, have real relevance to the recovering community as it relates to continued use of tobacco and now e-cigarettes and may indeed interfere with abstinence due to setting the brain up to reinstate alcohol or other abusable licit or illicit substances through a known gateway neuro epigenetic mechanism.
Controversy
For clarity Harvard Professor George Vaillant, surprisingly did not find evidence for effectiveness of the AA program relative to a control having no AA treatment.
Vaillant concluded that
“AA may be a good and comfortable fit for a few people who have a problem with alcohol, the majority of people with alcohol problems appear to do better with a different approach. We would love to see a study of why so many people dropped out of AA. We hypothesize that this may be due to the fact that AA’s theological notions of the powerlessness of humanity and of the need for a rescuing God are unpalatable not only to many atheists and agnostics but to almost all theists who are not Calvinists as well”.
Furthermore Vaillant suggests based on his research that: “It may also be the case that the AA philosophy of “powerlessness” over alcohol and slogans such as “one drink, one drunk”, “one is too many and a thousand is never enough” and “alcohol is cunning, baffling, and powerful” actually set people up to binge drink rather than to practice damage control when they slip up and fail to abstain as intended. More data on this topic is definitely needed”.
A summary of Valliant’s
AA is a good fit for a small number of people with alcohol problems and helps them to abstain.
AA is a poor fit for the majority of people with alcohol problems and can make some people worse.
AA is better at creating “true believers” than it is at eliminating problem drinking.
Whether or not AA is a good fit for a person has little if anything to do with how much a person drinks or the number of alcohol-related problems that a person has-the essential factor is personality type.
AA is a good fit for black-and-white thinkers who accept proof by authority.
AA is a poor fit for people who think in shades of gray and demand experimental evidence and scientific proof.
We assert it may be possible in the future to test for specific genes that would better match individuals to accepting the doctrines of AA [74]. There are examples of why AA does not work for everybody [75]. Interestingly, a PUBMED search (9-5-14) using terminology “why alcoholics anonymous does NOT work” did not retrieve any results. However, Kelly [75] pointed out that:
“Regarding subpopulations, current evidence suggests non-or less-religious individuals benefit as much from self-help groups as more religious individuals and women become as involved and benefit as much as men. However, participation in, and effects from, traditional self-help groups for dually diagnosed patients may be moderated by type of psychiatric comorbidity. Some youth appear to benefit, but remain largely unstudied. Dropout and nonattendance rates are high, despite clinical recommendations to attend.”
Beyound Vaillant
Since Valliant has reviewed the potential of the 12 steps in reducing relapse, there have been many reports to the contrary revealing the importance of meditation, personality, transcendence, mindfulness and spirituality. In fact our laboratory has recently reported that as belief in spirituality rises in an individual so does remission to substance abuse [76]. There have been studies directed at understanding from a neurotransmitter level with mixed results. Whereby, Finnish scientists found no association between 5-HT-1A receptors and spiritual experiences in both patients with major depression and healthy controls [77], others did find an association. Specifically, Borg et al. [78] did find that spiritual acceptance correlated significantly with the several-fold availability of 5-HT-1A receptor density and “may explain why people vary greatly in spiritual zeal”. Along similar lines, it was found that boys and girls with the combination of presence of the short 5-HTTLPR, and homozygosity for the long AP-2beta genotype scored significantly lower on Self-Transcendence and Spiritual Acceptance [79].
Other work by Davidson’s group on mindfulness reveals the importance of mediation in terms of brain activation of the reward circuitry. They found that Expert Meditators activated to a greater degree fMRI adapted Stroop Word-Colour Task (SWCT), which requires attention and impulse control compared to novices. Understanding this could suggest that meditation coupled with enhance spiritual belief may indeed induce DA release at the VTA and cingulate gyrus that could translate to better clinical outcomes and reduced relapse [80, 81].
Certainly the genes have a role in substance use disorder as well as behavioral addictions and these subsets of RDS seem to be inheritable. However, RDS is not a monogenetic disorder with one gene causing this complex mental condition. It is polygenic with multiple inducible epigenetic effects on DNA chromatin structure and function. Belcher et al. [82] have identified three-high order personality traits that seem tied to specific brain regions and gene polymorphisms. Obviously these polymorphisms influence and effect the workings of these brain regions and ultimately reflect an individual’s personality [83] and even belief system. This could translate to either vulnerability or resilience to developing RDS.
Undoubtedly most treatment facilities would embrace the 12-step program including the helper principle [84] but its role as the only treatment option has been questioned. In spite of the fact that the use of MAT is not usually endorsed by AA/ NA or similar organizations, Chappel and Dupont [47] not only embraced the 12 step program for recovering addicts but also suggested the importance of legally prescribed treatment medications as well, especially for co-morbid psychiatric conditions. They also point out the importance of Galanters’ involvement of family members and friends in the network therapy to prevent relapse. As suggested by Scott A. Teitelbaum in his book “Addiction: A family Affair (2011) since there is no known “cure” for addiction, it is imperative that prevention must start with the family. Finally Galanter et al. [85] reported that patients more oriented toward a spiritual than a formerly religious affiliation toward other 12-step members (a spiritual awakening) associated with lower rates of substance seeking behavior.
Evidence continues to emerge regarding our understanding of brain function especially the reward circuitry and all addictive behaviors and clinicians are encouraged to review some of the select molecular neurogenetic literature [86–105].
Summary of Molecular Neurobiology of the 12 Steps
The summary of how the molecular neurobiology effects each step has been covered in detail in Blum et al. [46]. Here we provide a brief synopsis of the main message linked to each step (see Table 2 with references).
Table 2.
The Molecular Neurobiological Summary of the 12 Steps (taken with permission from Blum et al. [46]).
| STEP | Summary with References |
|---|---|
| First-Powerless | While the concept of POWERLESSNESS may be controversial in the field, the first step admitting personal powerlessness over addiction is supported by the actual mechanisms involved in the neurobiological circuits of our brain. It begins with genetic vulnerability to addiction and compounded by epigenetically induced environmental elements. Stress and the toxic effects of the drugs and compulsive behaviors themselves induce changes in the neuroanatomy, neurophysiology, and neurochemistry of the brain that effect hedonic tone, physical dependence, craving and relapse. Although genetic factors play a very significant role in the process of addiction and especially in risk for developing reward dependence behaviors, there are strong epigenetic effects of powerful substances. Those substances like alcohol etc. profoundly affect brain reward homeostasis and an unmanageable desire to self-administer drugs of abuse. This leads to powerlessness, an inability to control behaviors in face of harm that ultimately influences every aspect of one’s life [4, 104, 107–135]. |
| Second-Restore Us to Sanity |
Sanity (sound judgment) or insanity (repetitive behavior in spite of harm) may be impaired even at birth and could be due to deficient brain reward circuitry function especially resulting in a hypodopaminergic trait. This poor judgment could be a root cause for aberrant substance-seeking behavior in the face of harm’s way. This becomes further complicated when other environmental factors are present including drug availability, non-nurturing parents, social economic burdens and stress. Importantly the ability to behave sanely also may be impacted by an individual’s relationship with a power greater than themselves. In terms of relapse, it is well known that the prefrontal cortex and cingulate gyrus are very important brain regions that could regulate relapse. Poor judgment stemming from impairments in the neurochemical functioning of these regions due to genes and/or toxic substances and/or behaviors, impedes recovery and induces relapse. Understanding the molecular biology of the brain reward system (genes and environment) highlights the importance of positive input from fellowship (self-help) programs and other treatment modalities that can offset unwanted gene expression, lift spirits, and assist in enabling the individual to achieve a state of sanity and make good choices [5, 9, 16, 74, 78, 79, 104, 136–179]. |
| Third-Turn our lives over to GOD |
Will-power is not simple to control, especially if you are born with a compromised reward system, especially low levels of endorphins. Genetically predisposed individuals seek out drugs such as alcohol, heroin, cocaine, nicotine, and even sugar, because these substances all activate reward substrates (i.e., enkephalins, DA pathways) and provide a pseudo temporary feeling of well-being (so called “normalization”). Will power is based on both the interplay of genes and environmental elements in society. This includes stress as an adult and surprising during the prenatal phase. This early stress could lead to aberrant substance use disorders in adult life. Since it is not easy to fight the hard wiring of our brain reward circuitry, for the recovering addict it seems obvious to look for reward outside of our genome [i.e. alcohol, drugs, sex, and food] [180–186]. |
| Fourth-Fearless & Moral Inventory of Ourselves |
Fearless moral inventory must include not only the drug of choice but other Reward Deficiency Syndrome related behaviors. This is so because the phenotype is not any particular drug or behavior of choice; it is indeed Reward Deficiency Syndrome. However, the inventory the individual is completing cannot be “right” or “wrong,” because it his/her own list of resentments, and evaluation of self. Moreover, the Big Book states, “No one among us has been able to maintain perfect adherence to any of these principles. The point is that we are willing to grow along spiritual lines. The principles we have set down are guides to progress. We claim spiritual progress rather than spiritual perfection.” Several fourth steps may be taken by an individual over the course of his/her sobriety. Moreover, it is literally almost impossible for early recovering addicts to embrace Step 4 due to protracted abstinent impairments of brain reward circuitry, for example, in alcoholics, heroin addicts, and cocaine addicts. Unfortunately this could be due to the chronic abuse of these powerful substances as an epigenetic phenomena, as well as possible inherited reward gene polymorphisms that occur at birth. It has been reasoned that one therapeutic target involves continued natural DA D2 activation as reflected in the preliminary fMRI research being conducted in China using KB220Z [51, 15, 24, 29, 31, 41, 67, 102, 106, 115, 187–195]. |
| Fifth-Admitted Exact Nature of our Wrong Doings |
Understanding our natural desire to obtain pleasure states and to admit “wrong doing” to God, ourselves, and those around us is no simple task and involves the consideration of not just our issue with “getting high” but rather with the toxic effects produced in the brain by continual exposure to these powerful substances. Their impact on brain reward networks is indeed physiological (e.g., increase in brain DBI). This can result in mental effects (anxiety and aggression) that also result in harmful behaviors with harmful and sometimes fatal consequences not only to one-self but to others [187, 196–208]. |
| Sixth-Removing Defects in Character |
Although it is possible to define character in a moralistic sense, it is very difficult to assign responsibility for defects of character and the bad decisions and consequence since character is shaped by genetic (evolutionary) forces far beyond a person’s control. With this stated it is argued that environmental elements especially in childhood may also require rethinking in terms of blame and or even praise of an individual act. This idea supports the idea in the sixth step that the removal of character defects is the province of a higher power. Clinicians should be cognizant that for the individual, achievement of this step requires deep character analysis, painful realization, and ability to dissociate oneself (present) from the past self. It should also be noted that carriers of the DRD2 gene polymorphism (risk for addiction) will have great difficulty in achieving honesty [209–225]. |
| Seventh-Removing Shortcomings |
Being humble must be accompanied with both gratitude and grace. The concept of ‘turning it over’ and let GOD remove our shortcomings is not easily accomplished. To be humble is akin to having gratitude for the things we have the idea of moving forward. Statements of spiritual faith and being humble challenge the recovering person to face the fact that good intentions and honest effort alone will not always succeed in getting him or her what is truly wanted from life. I n turn and supported by genetic predisposition, this could lead to chronic depression and relapse. However, the 12-step program and the traditions together ask the person to believe that evil and brutishness, injustice and cruelty will not necessarily win out in the end. Being humble and having faith advocates neither passivity nor hopelessness; on the contrary, they express the belief that our shortcomings can be removed by our willingness to believe that things can work out for the best in the long term. Having positive feelings about GOD translates to positive epigenetics which enhance the chances that we could remove our shortcomings by expressing “good” genes rather than “bad genes” [10, 11, 99, 226–244]. |
| Eighth-Making Amends of Harms |
It is not easy to make amends especially to people who are not only our friends but people whom we love. Step 8 does not come early in one’s sobriety but only after periods of being clean and sober. However, once an individual accomplishes this arduous task he or she will be able to move forward in the path of recovery. In terms of connecting the dots, it is important for clinicians to realize that the old adage of “Birds of a feather flock together” may be inheritable by virtue of friends seeking friends who not only have similar characteristic (maybe even drinking, drugging, and eating), but similar genotypes, such as the DRD2 A1 allele. So that when the alcoholic, for example, is asked to make amends and also eliminate certain friends that would not be conducive to their recovery, we need to be cognizant about going against the genetic grain. Thus, on a molecular neurobiological level, it is easily said but not easily done. A form of happiness is that people live in social networks that are comfortable. Making amends for the hurt may not reestablish trust but may help assuage guilt and shame. Here it may be helpful to consider the genetic predisposition of families to Reward Deficiency Syndrome behaviors [46, 104, 165, 216, 245–267]. |
| Ninth-Direct Amends to Such People without Injury to Them |
It is not easy to achieve happiness and peace especially when the alcoholic or addict is faced with taking responsibility for hurting others with whom he or she has relationships while drinking and drugging. An obvious source of injury to relationships caused by addiction is the “abandonment” of a spouse or significant other for alcohol and/or drugs. Victims of Reward Deficiency Syndrome must take responsibility for this abandonment of loved ones. Furthermore, addicts may have been very abusive (both physically and emotionally) during their active addiction. Before any amends can be made, the addict is asked in Step 8 to take an inventory of all persons harmed, which can easily evoke intense feelings of guilt and shame. It also requires overcoming denial and being willing to make amends. In Step 9, the achievement of making amends (except where doing so would cause no further injury) is subject to correlations among genes, friendships, and relationships. As noted in the research summarized above, relationships and happiness are based on neuronal hard wiring, and this presents both a formidable challenge and clarity as to how to achieve effective healing in recovery. The degree to which the person can make amends to others (without harm or hurt) is tantamount to a healthy recovery, and importantly, the attainment of happiness. This can be facilitated through the positive natural release of DA in reward centers of the brain [34, 137, 268–271]. |
| Tenth-Take Personal Inventory and Admit to being wrong |
The tenth step can be a pressure-relief valve. Addicts work this step while the day’s ups and downs are still fresh in mind. They list what they have done and try not to rationalize their actions. The first thing they must do is stop! Then they must take the time to allow themselves the privilege of thinking. They work this step continuously. It presents a way of avoiding grief. The individual monitors feelings, emotions, fantasies, and actions. By constantly looking at these things they may be able to avoid repeating the actions that make them feel bad (Narcotics Anonymous Basic Text, Chapter 4/Step 10). Step 10 is the maintenance step for Steps 4 and 5 and encourages the taking of a personal inventory, which, for recovering persons, should be a daily process. It is important that addicts realize that if they do carry a genetic risk, for example the DRD2 A1 allele among other gene deficits with 30–40 % less D2 receptor density, taking inventory and feeling good about it is a temporarily “dopamine fix.” As such, addicts must continue to “work the steps” on a day-to-day basis to replenish DA. [25, 272–274]. |
| Eleventh-Prayer & Meditation to Contact God |
Doing the work required in Step 11 continuously through both the meditative and prayer process increases the release of DA at the synaptic level. In addition, working Step 11 on a daily basis will offset the genetically induced “hypodopaminergic brain function” by continued DA release in the synapse. Increased DA will result in a subsequent proliferation of DA D2 receptors even in carriers of the DRD2 A1 allele and other reward gene polymorphisms. The increase in D2 receptors translates to enhanced DA function, which will ultimately promote greater confidence in the recovering addict, enabling a better understanding of the written word of the twelve-step fellowship. This will lead to an anti-stress effect and as such reduce the chance for relapse especially in dysfunctional and co-dependent families [25, 32, 42, 156, 169, 258, 272, 274, 275–278]. |
| Twelfth-Spiritual Awakening & Carrying the Message to Others |
Step 12-occurs when the recovering person had done the work, and truly understands all the preceding steps in the program. It has been said that working all the steps will allow an individual to have spiritual awakening. We point out that for people who have addiction is dependent on both genes and environmental conditions, attaining this awakening may be more or less difficult. One of the most fulfilling experiences one could get is sharing emotions with others especially as it relates to carrying the message of the fellowship to other addicts. It is important to realize this experience maybe impacted by the synthesis and release of the brain chemical oxytocin. Unfortunately, independent of one’s genetic makeup, alcohol and opiates significantly impair the synthesis and release of this important human bonding neuropeptide. Finally, clinicians should be cognizant that any lifestyle change is significantly impacted by both polymorphic genes and traumatic events [79, 279–286]. |
Step 1-We admitted that we were powerless over alcohol-that our lives had become unmanageable
While the concept of POWERLESSNESS may be controversial in the field, the first step admitting personal powerlessness over addiction is supported by the actual mechanisms involved in the neurobiological circuits of our brain. Genetic vulnerability to addiction and compulsive behaviors, compounded by epigenetically induced environmental elements. Stress and the toxic-effects of the drugs themselves induce changes in the neuroanatomy, neurophysiology, and neurochemistry of the brain that change hedonic tone, physical dependence, craving, and relapse. In essence, it is very true that indeed a person is powerless. The addicted person has no control over drug seeking and other damaging behaviors despite their denial of loss of control over drug abuse and erroneous thoughts concerning their “pseudo-power” over their unwanted behavior.
Although genetic factors play a very significant role in the process of addiction and especially in risk for developing reward dependence behaviors, as we see from experiments published above these powerful substances have strong epigenetic effects of. Those effects profoundly disrupt brain-reward homeostasis and cause an UNMANAGEABLE desire to self-administer drugs of abuse. The unmanageable desire manifests as powerlessness, an inability to control behaviors that influence every aspect of one’s life.
Step 2-Came to believe that a Power greater than ourselves could restore us to sanity
Sanity (sound judgment) or insanity (repetitive behavior despite the harm) may be impaired even at birth and could be due to deficient brain reward circuitry function especially resulting in a hypodopaminergic trait. This poor judgment could be a cause of aberrant substance seeking behavior in the face of harm’s way. Poor decisions compounded by environmental factors including drug availability, non-nurturing parents, social-economic burdens, and stress. Importantly the ability to behave sanely also may be impacted by an individual’s relationship with a power greater than themselves. In terms of relapse, it is well known that the prefrontal cortex and cingulate gyrus are critical areas of the brain involved in relapse regulation. Poor judgment caused by impairments in the neurochemical functioning of these regions due to genes and/ or toxic substances and/or behaviors impede recovery and induce relapse. Understanding the molecular biology of the brain reward system (genes and environment) highlights the importance of positive input from fellowship (self-help) programs and other treatment modalities. Positive input from fellowship can offset unwanted gene expression, lift spirits, and assist in enabling the individual to achieve a state of sanity and make right choices.
Step 3-Made a decision to turn our will and our lives over to the care of God as we understood him
Will-power is difficult to control, especially in individuals born with a compromised reward system, and low levels of endorphins. Genetically predisposed individuals seek out drugs such as alcohol, heroin, cocaine, nicotine, and even sugar. These substances all activate the reward substrates (like serotonin, enkephalins, GABA and DA pathways) and provide a pseudo temporary feeling of well-being (so called “normalization”). Willpower is based on both the interplay of genes and environmental elements in society. Stress as an adult and surprising during the prenatal phase are environmental elements. This early stress can lead to aberrant substance use disorders in adult life as seen with epigenetic effects on Glucocorticoid receptor express. Since it is difficult to fight the hard wiring of our brain reward circuitry, for the recovering addict it seems obvious to look for reward outside of our genome (i.e. alcohol, drugs, sex, and food).
Step 4-Made a searching and fearless moral inventory of ourselves
Fearless moral inventory must include the drug of choice and other RDS related behaviors because the phenotype is not any particular drug or behavior; it is indeed RDS. However, the inventory the individual is completing cannot be “right” or “wrong,” because it his/her own evaluation of self and list of resentments. Moreover, the Big Book states (How It Works, Page 60), “No one among us has been able to maintain perfect adherence to any of these principles. The point is that we are willing to grow along spiritual lines. The principles we have set down are guides to progress. We claim spiritual progress rather than spiritual perfection.” Several “fourth steps” may be taken by an individual over the course of his/her sobriety. Moreover, it is almost impossible for addicts in early recovery, to embrace Step 4. Impairments of brain reward circuitry are protracted and amplified during withdrawal and early recovery, for example, in alcoholics, heroin addicts, and cocaine addicts. Unfortunately, this could be due to the chronic abuse of these powerful substances as epigenetic phenomena, as well as, possible inherited reward gene polymorphisms that occur at birth. It has been reasoned that one therapeutic target involves continued natural DA D2 activation as reflected in the preliminary fMRI research being conducted in China using KB220Z [106].
Step 5-Admitted to God, to ourselves, and to another human being the exact nature of our wrongs
This step involves the consideration of our issues with “getting high”, as well as, the toxic effects of continual exposure to these powerful substances. Their impact on brain reward networks is indeed physiological (e.g., increase in brain DBI). Physiological changes can result in psychological effects (anxiety and aggression) that are behind behaviors with harmful and sometimes fatal consequences not only to ones-self but others.
Step 6-Were entirely ready to have God remove all these defects of character
Although it is possible to define character in a moralistic sense, it is very difficult to assign responsibility for defects of character and the wrong-decisions and consequence since character is shaped by genetic (evolutionary) forces far beyond a person’s control. With this stated it is argued that environmental elements especially in childhood may also require rethinking in terms of blame and or even praise of an individual act. This idea supports the idea in the sixth step that the removal of character defects is the province of a higher power. Clinicians should be cognizant that for the individual, achievement of this step requires deep character analysis, painful realization, and ability to dissociate oneself (present) from the past self. It should also be noted that carriers of the DRD2 gene polymorphism (risk for addiction) will have great difficulty being honest.
Step 7-Humbly asked Him to remove our shortcomings
Being humble must be accompanied with both gratitude and grace. The concept of ‘turning it over’ and let GOD remove our shortcomings is not easily accomplished. To be humble is akin to having gratitude for the things we have the idea of moving forward. Statements of spiritual faith and being humble challenge the recovering person to face the fact that good intentions and honest effort alone will not always succeed in getting him or her what is truly wanted from life. In turn, and supported by genetic predisposition, this could lead to chronic depression and relapse. However, the 12-step program and the traditions together ask the person to believe that evil and brutishness, injustice and cruelty will not necessarily win out in the end. Being humble and having faith, advocates neither passivity nor hopelessness; on the contrary, they express the belief that our shortcomings can be removed by our willingness to believe that things can work out for the best in the long term. Having positive feelings about GOD translates to positive epigenetics that enhance the chances that we could remove our shortcomings by expressing “good” genes rather than “bad” genes.
Step 8-Made a list of all persons we had harmed, and became willing to make amends to them all
It is not easy to make amends especially to people who are not only our friends but people whom we love. Step 8 does not come early in one’s sobriety but only after periods of being clean and sober. However, once an individual accomplishes this arduous task he or she will be able to move forward on the path to recovery. In terms of connecting the dots, it is important for clinicians to realize the old adage of “Birds of a feather flock together” may be an effect of a genetic association. By virtue of friends seeking friends who, not only have similar characteristic (maybe even drinking, drugging, and eating), but similar genotypes, such as the DRD2 A1 allele So that when the alcoholic, for example, is asked to make amends and also eliminate certain friends that would not be conducive to their recovery, we need to be cognizant of going against the genetic grain. Thus, on a molecular neurobiological level, it is easily said but not easily done. A form of happiness is that people live in social networks that are comfortable. Making amends for the hurt may not reestablish trust but may help assuage guilt and shame. Here it may be helpful to consider the genetic predisposition of families to RDS behaviors.
Step 9-Made direct amends to such people wherever possible, except when to do so would injure them or others
It is not easy to achieve happiness and peace especially when the alcoholic or addict is faced with taking responsibility for hurting others with whom he or she had a relationships while drinking and drugging. An obvious source of injury to relationships caused by addiction is the “abandonment” of a spouse or significant other for alcohol and/or drugs. Victims of RDS must take responsibility for this abandonment of loved ones. Furthermore, addicts may have been very abusive (both physically and emotionally) during their active addiction. In Step 8 before any amends can be made, the addict is asked to take an inventory of all persons harmed, which can easily evoke intense feelings of guilt and shame. It also requires overcoming denial and being willing to make amends. In Step 9, the achievement of making amends (except where doing so would cause no further injury) is subject to correlations among genes, friendships, and relationships. As noted in the research summarized above, relationships and happiness are based on neuronal hard wiring, and this presents both a formidable challenge and clarity as to how to achieve healing during recovery. The degree to which the person can make amends to others (without harm or hurt) is tantamount to a healthy recovery, and importantly, the attainment of happiness. Making amends can be facilitated by the active natural release of DA in reward centers of the brain.
Step 10-Take personal inventory and admit to being wrong
The tenth step can be a pressure-relief valve. Addicts work this step while the day’s ups and downs are still fresh in mind. They list what they have done and try not to rationalize their actions. The first thing they must do is stop! Then they must take the time to allow themselves the privilege of thinking. They work this step continuously. It presents a way of avoiding grief. The individual monitors feelings, emotions, fantasies, and actions. By constantly looking at these things they may be able to avoid repeating the actions that make them feel bad (Narcotics Anonymous Basic Text, Chapter 4/Step 10). Step 10 is the maintenance for Steps 4 and 5 and “encourages the taking of a personal inventory, which, for recovering persons, should be a daily process”. It is important that addicts realize that if they do carry a genetic risk, for example, the DRD2 A1 allele with 30–40% less D2 receptor density, taking inventory and feeling good about it is a temporarily “dopamine fix”. As such, addicts must continue to “work the steps” on a daily basis to replenish DA.
Step 11-Sought through prayer and meditation to improve our conscious contact with God, as we understood Him, praying only for knowledge of His will for us and the power to carry that out
Doing the work required in Step 11 continuously through both the meditative and prayer process increases the release of DA at the synaptic level. In addition, working Step 11 on a daily basis will offset the genetically induced “hypodopaminergic brain function” by continued DA release in the synapse. Increased DA will result in a subsequent proliferation of DA D2 receptors even in carriers of the DRD2 A1 allele and other reward gene polymorphisms. The increase in D2 receptors translates to enhanced DA function, which will ultimately promote greater confidence in the recovering addict, enabling a better understanding of the written word of the twelve-step fellowship. This will lead to an anti-stress effect and as such reduce the chance for relapse especially in dysfunctional and co-dependent families.
Step 12-Having had a spiritual awakening as the result of these steps, we tried to carry this message to alcoholics and to practice these principles in all our affairs
Step 12 occurs when the recovering person had done the work, and truly understands all the preceding steps in the program. It has been said that working all the steps will allow an individual to have spiritual awakening. We point out that for people who have addiction is dependent on both genes and environmental conditions, attaining this awakening may be more or less difficult. One of the most fulfilling experiences one could get is sharing emotions with others especially as it relates to carrying the message of the fellowship to other addicts. It is important to realize this experience may be impacted by the synthesis and release of the brain chemical oxytocin. Unfortunately, independent of one’s genetic makeup, alcohol and opiates significantly impair the synthesis and release of this important human bonding neuropeptide. Finally, clinicians should be cognizant that any lifestyle change is significantly impacted by both polymorphic genes and traumatic events.
Although still controversial one way to sum up the benefits of the 12 step program & fellowship is embedded and so reflected in these statements by addiction professionals working in the field of addiction medicine (see Figure 5).
Figure 5.

Professional’s views of the 12 Step Program & Fellowship.
Conclusion
While it would be easy to say that all addicted individuals would benefit from the 12 step doctrines, this may not be the case. In fact when it comes to spirituality there are a number of genes and associated polymorphisms that load onto one’s beliefs related to GOD [46, 74, 79, 137, 287–289]. One member stated that “The program is perfect and it does not fail-people will.” (Anonymous) .
As we stated before “Finding happiness may not only reside in our genome but may indeed be impacted by positive meditative practices, positive psychology, spiritual acceptance, love of others and self, and taking inventory of ourselves-one day at a time” [46].
Acknowledgements
The authors are appreciative of the support of Mary Hauser, Lisa Marzili and Chris Campanella of Dominion Diagnostics, LLC.
Funding Sources
The writing of this paper was supported in part by funds from the National Institutes of Health, NIAAA (RO1-AA07112 and K05-AA00219) and the Medical Research Service of the US Department of Veterans Affairs (MOB). The authors appreciate the editorial assistance of Margaret A. Madigan and Paula J. Edge. The University of Florida Foundation supported the present research. Rajendra D. Badgaiyan is supported by the National Institutes of Health grants 1R01NS073884 and 1R21MH073624; and VA Merit Review Awards CX000479 and CX000780. Kenneth Blum is the recipient of a grant to PATH FOUNDATION NY, by Life Extension Foundation, Ft/ Lauderdale, Florida.
Footnotes
Conflict of Interest
Kenneth Blum, PhD is the holder of a number of US and Foreign patents issued and pending related to Nutrigenomics and Nutraceuticals. Through IGENE LLC., Dr. Blum exclusively licensed the Genetic Addiction Risk Score (GARS)™ to Dominion Diagnostics, LLC. Dr. Blum is also an officer and stock holder of IGENE, LLC and is a paid consultant of Dominion Diagnostics, LLC, IGENE, Malibu Recovery Center. Dr. Blum is a paid consultant of Malibu Recovery Center. Dr. Blum is a member of the scientific advisory board of Dominion Diagnostics, LLC and is Chief Scientific Advisor of Dominion Diagnostics, and Chief Scientific Officer of RDSolutions and Victory Nutrition International, LLC.
Authors Contribution
The initial manuscript was written by KB, JG, MSG, ST, MOB, JF. Clinical review by DES, AKR, ZD, GA and JF. All authors approved the manuscript prior to submission. MAM (Margaret A. Madigan) provided final writing and edits.
Dedication
To the “like minded Docs” of ASAM.
References
- 1.Hodge CW, Chappelle AM, Samson HH. Dopamine receptors in the medial prefrontal cortex influence ethanol and sucrose-reinforced responding. Alcohol Clin Exp Res. 1996;20(9):1631–1638. doi: 10.1111/j.1530-0277.1996.tb01709.x. [DOI] [PubMed] [Google Scholar]
- 2.Hodge CW, Slawecki CJ, Aiken AS. Norepinephrine and serotonin receptors in the paraventricular nucleus interactively modulate ethanol consumption. Alcohol Clin Exp Res. 1996;20(9):1669–1674. doi: 10.1111/j.1530-0277.1996.tb01714.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hodge CW, Cox AA. The discriminative stimulus effects of ethanol are mediated by NMDA and GABA(A) receptors in specific limbic brain regions. Psychopharmacology (Berl) 1998;139(1–2):95–107. doi: 10.1007/s002130050694. [DOI] [PubMed] [Google Scholar]
- 4.Blum K, Kozlowski GP. Ethanol and neuromodulator interactions: A cascade model of reward. In: Ollat H, Parvez H, editors. Alchol and Behavior. Utrecht and Netherlands: Prog Alcohol Res VSP; 1990. pp. 131–149. [Google Scholar]
- 5.Blum K, Braverman ER, Holder JM, Lubar JF, Monastra VJ, et al. Reward deficiency syndrome: a biogenetic model for the diagnosis and treatment of impulsive, addictive, and compulsive behaviors. J Psychoactive Drugs. 2000;32(Suppl: i-iv):1–112. doi: 10.1080/02791072.2000.10736099. [DOI] [PubMed] [Google Scholar]
- 6.Hawks J. Still evolving (after all these years) Sci Am. 2014;311(3):86–91. doi: 10.1038/scientificamerican0914-86. [DOI] [PubMed] [Google Scholar]
- 7.Blum K, Sheridan PJ, Wood RC, Braverman ER, Chen TJ, et al. Dopamine D2 receptor gene variants: association and linkage studies in impulsive-addictive-compulsive behaviour. Pharmacogenetics. 1995;5(3):121–141. doi: 10.1097/00008571-199506000-00001. [DOI] [PubMed] [Google Scholar]
- 8.Blum K, Wood RC, Braverman ER, Chen TJ, Sheridan PJ. The D2 dopamine receptor gene as a predictor of compulsive disease: Bayes’ theorem. Funct Neurol. 1995;10(1):37–44. [PubMed] [Google Scholar]
- 9.Blum K, Sheridan PJ, Wood RC, Braverman ER, Chen TJ, et al. The D2 dopamine receptor gene as a determinant of reward deficiency syndrome. J R Soc Med. 1996;89(7):396–400. doi: 10.1177/014107689608900711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Grandy DK, Litt M, Allen L, Bunzow JR, Marchionni M, et al. The human dopamine D2 receptor gene is located on chromosome 11 at q22-q23 and identifies a TaqI RFLP. Am J Hum Genet. 1989;45(5):778–785. [PMC free article] [PubMed] [Google Scholar]
- 11.Smith GP, Schneider LH. Relationships between mesolimbic dopamine function and eating behavior. Ann N Y Acad Sci. 1988;537:254–261. doi: 10.1111/j.1749-6632.1988.tb42111.x. [DOI] [PubMed] [Google Scholar]
- 12.Blum K, Braverman ER, Wood RC, Gill J, Li C, et al. Increased prevalence of the Taq I A1 allele of the dopamine receptor gene (DRD2) in obesity with comorbid substance use disorder: a preliminary report. Pharmacogenetics. 1996;6(4):297–305. doi: 10.1097/00008571-199608000-00003. [DOI] [PubMed] [Google Scholar]
- 13.Braverman ER, Blum K. Substance use disorder exacerbates brain electrophysiological abnormalities in a psychiatrically-ill population. Clin Electroencephalogr. 1996;27(47):5–27. doi: 10.1177/1550059496027s0402. [DOI] [PubMed] [Google Scholar]
- 14.Gyollai A, Griffiths MD, Barta C, Vereczkei A, Urbán R, et al. The genetics of problem and pathological gambling: a systematic review. Curr Pharm Des. 2014;20(25):3993–3999. doi: 10.2174/13816128113199990626. [DOI] [PubMed] [Google Scholar]
- 15.Di Chiara G, Imperato A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci U S A. 1988;85(14):5274–5278. doi: 10.1073/pnas.85.14.5274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Comings DE, Gonzales N, Saucier G, Johnson JP, MacMurray JP. The DRD4 gene and the spiritual transcendence scale of the character temperament index. Psychiatr Genet. 2000;10(4):185–189. doi: 10.1097/00041444-200010040-00006. [DOI] [PubMed] [Google Scholar]
- 17.Rutter JL, Volkow ND. Re-defininG AddiC(CH3)Tion: genomics and epigenomics on substance use disorders. Mol Genet Genomic Med. 2014;2(4):273–279. doi: 10.1002/mgg3.93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Eisenberg DT, Campbell B, Mackillop J, Lum JK, Wilson DS. Season of birth and dopamine receptor gene associations with impulsivity, sensation seeking and reproductive behaviors. PLoS One. 2007;2(11):e1216. doi: 10.1371/journal.pone.0001216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tomasi D, Wang GJ, Wang R, Caparelli EC, Logan J, et al. Overlapping patterns of brain activation to food and cocaine cues in cocaine abusers: association to striatal D2/D3 receptors. Hum Brain Mapp. 2015;36(1):120–136. doi: 10.1002/hbm.22617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dackis CA, Gold MS. New concepts in cocaine addiction: the dopamine depletion hypothesis. Neurosci Biobehav Rev. 1985;9(3):469–477. doi: 10.1016/0149-7634(85)90022-3. [DOI] [PubMed] [Google Scholar]
- 21.Blum K, Eubanks JD, Wallace JE, Schwertner HA. Suppression of ethanol withdrawal by dopamine. Experientia. 1976;32(4):493–495. doi: 10.1007/BF01920816. [DOI] [PubMed] [Google Scholar]
- 22.Hietala J, West C, Syvälahti E, Någren K, Lehikoinen P, et al. Striatal D2 dopamine receptor binding characteristics in vivo in patients with alcohol dependence. Psychopharmacology (Berl) 1994;116(3):285–290. doi: 10.1007/BF02245330. [DOI] [PubMed] [Google Scholar]
- 23.Melis M, Spiga S, Diana M. The dopamine hypothesis of drug addiction: hypodopaminergic state. Int Rev Neurobiol. 2005;63:101–154. doi: 10.1016/S0074-7742(05)63005-X. [DOI] [PubMed] [Google Scholar]
- 24.Rothman RB, Blough BE, Baumann MH. Dual dopamine/ serotonin releasers as potential medications for stimulant and alcohol addictions. AAPS J. 2007;9(1):E1–E10. doi: 10.1208/aapsj0901001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Boundy VA, Pacheco MA, Guan W, Molinoff PB. Agonists and antagonists differentially regulate the high affinity state of the D2L receptor in human embryonic kidney 293 cells. Mol Pharmacol. 1995;48(5):956–964. [PubMed] [Google Scholar]
- 26.Thanos PK, Volkow ND, Freimuth P, Umegaki H, Ikari H, et al. Overexpression of dopamine D2 receptors reduces alcohol self-administration. J Neurochem. 2001;78(5):1094–1103. doi: 10.1046/j.1471-4159.2001.00492.x. [DOI] [PubMed] [Google Scholar]
- 27.Thanos PK, Taintor NB, Rivera SN, Umegaki H, Ikari H, et al. DRD2 gene transfer into the nucleus accumbens core of the alcohol preferring and nonpreferring rats attenuates alcohol drinking. Alcohol Clin Exp Res. 2004;28(5):720–728. doi: 10.1097/01.alc.0000125270.30501.08. [DOI] [PubMed] [Google Scholar]
- 28.Thanos PK, Rivera SN, Weaver K, Grandy DK, Rubinstein M, et al. Dopamine D2R DNA transfer in dopamine D2 receptor-deficient mice: effects on ethanol drinking. Life Sci. 2005;77(2):130–139. doi: 10.1016/j.lfs.2004.10.061. [DOI] [PubMed] [Google Scholar]
- 29.Thanos PK, Michaelides M, Umegaki H, Volkow ND. D2R DNA transfer into the nucleus accumbens attenuates cocaine self-administration in rats. Synapse. 2008;62(7):481–486. doi: 10.1002/syn.20523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Merlo LJ, Gold MS. Prescription opioid abuse and dependence among physicians: hypotheses and treatment. Harv Rev Psychiatry. 2008;16(3):181–194. doi: 10.1080/10673220802160316. [DOI] [PubMed] [Google Scholar]
- 31.Blum K, Noble EP, Sheridan PJ, Montgomery A, Ritchie T, et al. Allelic association of human dopamine D2 receptor gene in alcoholism. JAMA. 1990;263(15):2055–2060. [PubMed] [Google Scholar]
- 32.Madrid GA, MacMurray J, Lee JW, Anderson BA, Comings DE. Stress as a mediating factor in the association between the DRD2 TaqI polymorphism and alcoholism. Alcohol. 2001;23(2):117–122. doi: 10.1016/s0741-8329(00)00138-5. [DOI] [PubMed] [Google Scholar]
- 33.Bau CH, Almeida S, Hutz MH. The TaqI A1 allele of the dopamine D2 receptor gene and alcoholism in Brazil: association and interaction with stress and harm avoidance on severity prediction. Am J Med Genet. 2000;96(3):302–306. doi: 10.1002/1096-8628(20000612)96:3<302::aid-ajmg13>3.0.co;2-i. [DOI] [PubMed] [Google Scholar]
- 34.Blum K, Braverman ER, Wu S, Cull JG, Chen TJ, et al. Association of polymorphisms of dopamine D2 receptor (DRD2), and dopamine transporter (DAT1) genes with schizoid/avoidant behaviors (SAB) Mol Psychiatry. 1997;2(3):239–246. doi: 10.1038/sj.mp.4000261. [DOI] [PubMed] [Google Scholar]
- 35.Robbins TW, Everitt BJ. Neurobehavioural mechanisms of reward and motivation. Curr Opin Neurobiol. 1996;6(2):228–236. doi: 10.1016/s0959-4388(96)80077-8. [DOI] [PubMed] [Google Scholar]
- 36.Wightman RM, Robinson DL. Transient changes in mesolimbic dopamine and their association with ‘reward’. J Neurochem. 2002;82(4):721–735. doi: 10.1046/j.1471-4159.2002.01005.x. [DOI] [PubMed] [Google Scholar]
- 37.Epping-Jordan MP, Watkins SS, Koob GF, Markou A. Dramatic decreases in brain reward function during nicotine withdrawal. Nature. 1998;393(6680):76–79. doi: 10.1038/30001. [DOI] [PubMed] [Google Scholar]
- 38.Cooper ML, Frone MR, Russell M, Mudar P. Drinking to regulate positive and negative emotions: a motivational model of alcohol use. J Pers Soc Psychol. 1995;69(5):990–1005. doi: 10.1037//0022-3514.69.5.990. [DOI] [PubMed] [Google Scholar]
- 39.Yamada M, Uddin LQ, Takahashi H, Kimura Y, Takahata K, et al. Superiority illusion arises from resting-state brain networks modulated by dopamine. Proc Natl Acad Sci U S A. 2013;110(11):4363–4367. doi: 10.1073/pnas.1221681110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Heinz A, Schäfer M, Higley JD, Krystal JH, Goldman D. Neurobiological correlates of the disposition and maintenance of alcoholism. Pharmacopsychiatry. 2003;36(Suppl 3):S255–S258. doi: 10.1055/s-2003-45139. [DOI] [PubMed] [Google Scholar]
- 41.Blum K, Chen TJ, Morse S, Giordano J, Chen AL, et al. Overcoming qEEG abnormalities and reward gene deficits during protracted abstinence in male psychostimulant and polydrug abusers utilizing putative dopamine D2 agonist therapy: Part 2. Postgrad Med. 2010;122(6):214–226. doi: 10.3810/pgm.2010.11.2237. [DOI] [PubMed] [Google Scholar]
- 42.Blum K, Chen ALC, Chen TJH, Braverman ER, Reinking J, et al. Activation instead of blocking mesolimbic dopaminergic reward circuitry is a preferred modality in the long term treatment of reward deficiency syndrome (RDS): a commentary. Theor Biol Med Model. 2008;5:24. doi: 10.1186/1742-4682-5-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Koob GF. Neurobiological substrates for the dark side of compulsivity in addiction. Neuropharmacology. 2009;56(Suppl 1):18–31. doi: 10.1016/j.neuropharm.2008.07.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Peciña S, Smith KS, Berridge KC. Hedonic hot spots in the brain. Neuroscientist. 2006;12(6):500–511. doi: 10.1177/1073858406293154. [DOI] [PubMed] [Google Scholar]
- 45.Blum K, Oscar-Berman M, Jacobs W, McLaughlin T, Gold M. Buprenorphine Response as a Function of Neurogenetic Polymorphic Antecedents: Can Dopamine Genes Affect Clinical Outcomes in Reward Deficiency Syndrome (RDS)? J Addict Res Ther. 2014;5:185. doi: 10.4172/2155-6105.1000185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Blum K, Femino J, Teitlebaum S, Giordano J, Oscar-Berman M, et al. Molecular Neurobiology of Addiction Recovery: The 12 Steps Program & Fellowship. New York, USA: Springer; 2013. [Google Scholar]
- 47.Chappel JN, DuPont RL. Twelve-step and mutual-help programs for addictive disorders. Psychiatr Clin North Am. 1999;22(2):425–446. doi: 10.1016/s0193-953x(05)70085-x. [DOI] [PubMed] [Google Scholar]
- 48.Blum K. Alcohol and the Addictive Brain. New York, USA: The Free Press; 1991. [Google Scholar]
- 49.Timko C, Laudet A, Moos RH. Newcomers to Al-Anon family groups: Who stays and who drops out? Addict Behav. 2014;39(6):1042–1049. doi: 10.1016/j.addbeh.2014.02.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Timko C, DeBenedetti A. A randomized controlled trial of intensive referral to 12-step self-help groups: one-year outcomes. Drug Alcohol Depend. 2007;90(2–3):270–279. doi: 10.1016/j.drugalcdep.2007.04.007. [DOI] [PubMed] [Google Scholar]
- 51.Kalivas PW, Brady K. Getting to the core of addiction: hatching the addiction egg. Nat Med. 2012;18(4):502–503. doi: 10.1038/nm.2726. [DOI] [PubMed] [Google Scholar]
- 52.Blum K, Thompson B, Oscar-Berman M, Giordano J, Braverman E, et al. Genospirituality: Our Beliefs, Our Genomes, and Addictions. J Addict Res Ther. 2013;5(4) doi: 10.4172/2155-6105.1000162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Demers CH, Bogdan R, Agrawal A. The Genetics, Neurogenetics and Pharmacogenetics of Addiction. Curr Behav Neurosci Rep. 2014;1(1):33–44. doi: 10.1007/s40473-013-0004-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Simon-O’Brien E, Alaux-Cantin S, Warnault V, Buttolo R, Naassila M, et al. The histone deacetylase inhibitor sodium butyrate decreases excessive ethanol intake in dependent animals. Addict Biol. 2014 doi: 10.1111/adb.12161. [DOI] [PubMed] [Google Scholar]
- 55.Febo M, Akbarian S, Schroeder FA, Ferris CF. Cocaine-induced metabolic activation in cortico-limbic circuitry is increased after exposure to the histone deacetylase inhibitor, sodium butyrate. Neurosci Lett. 2009;465(3):267–271. doi: 10.1016/j.neulet.2009.07.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sen N. Epigenetic Regulation of Memory by Acetylation and Methylation of Chromatin: Implications in Neurological Disorders, Aging, and Addiction. Neuromolecular Med. 2014 doi: 10.1007/s12017-014-8306-x. [DOI] [PubMed] [Google Scholar]
- 57.Bali P, Im HI, Kenny PJ. Methylation, memory and addiction. Epigenetics. 2011;6(6):671–674. doi: 10.4161/epi.6.6.15905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Wang Z, Yan P, Hui T, Zhang J. Epigenetic upregulation of PSD-95 contributes to the rewarding behavior by morphine conditioning. Eur J Pharmacol. 2014;732:123–129. doi: 10.1016/j.ejphar.2014.03.040. [DOI] [PubMed] [Google Scholar]
- 59.Starkman BG, Sakharkar AJ, Pandey SC. Epigenetics-beyond the genome in alcoholism. Alcohol Res. 2012;34(3):293–305. doi: 10.35946/arcr.v34.3.04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pandey SC, Ugale R, Zhang H, Tang L, Prakash A. Brain chromatin remodeling: a novel mechanism of alcoholism. J Neurosci. 2008;28(14):3729–3737. doi: 10.1523/JNEUROSCI.5731-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.He Y, Wang ZJ. Let-7 microRNAs and Opioid Tolerance. Front Genet. 2012;3:110. doi: 10.3389/fgene.2012.00110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Qiang M, Denny A, Lieu M, Carreon S, Li J. Histone H3K9 modifications are a local chromatin event involved in ethanol-induced neuroadaptation of the NR2B gene. Epigenetics. 2011;6(9):1095–1104. doi: 10.4161/epi.6.9.16924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Taqi MM, Bazov I, Watanabe H, Sheedy D, Harper C, et al. Prodynorphin CpG-SNPs associated with alcohol dependence: elevated methylation in the brain of human alcoholics. Addict Biol. 2011;16(3):499–509. doi: 10.1111/j.1369-1600.2011.00323.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Szutorisz H, DiNieri JA, Sweet E, Egervari G, Michaelides M, et al. Parental THC exposure leads to compulsive heroin-seeking and altered striatal synaptic plasticity in the subsequent generation. Neuropsychopharmacology. 2014;39(6):1315–13123. doi: 10.1038/npp.2013.352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Xu LF, Wang J, Lv FB, Song Q. Functions of microRNA in response to cocaine stimulation. Genet Mol Res. 2013;12(4):6160–6167. doi: 10.4238/2013.December.4.2. [DOI] [PubMed] [Google Scholar]
- 66.Bahi A, Dreyer JL. Striatal modulation of BDNF expression using microRNA124a–expressing lentiviral vectors impairs ethanol-induced conditioned-place preference and voluntary alcohol consumption. Eur J Neurosci. 2013;38(2):2328–2337. doi: 10.1111/ejn.12228. [DOI] [PubMed] [Google Scholar]
- 67.Zhang Y, Tian J, Yuan K, Liu P, Zhuo L, et al. Distinct resting-state brain activities in heroin-dependent individuals. Brain Res. 2011;1402:46–53. doi: 10.1016/j.brainres.2011.05.054. [DOI] [PubMed] [Google Scholar]
- 68.Archer T, Oscar-Berman M, Blum K. Epigenetics in Developmental Disorder: ADHD and Endophenotypes. J Genet Syndr Gene Ther. 2011;2(1):1000104. doi: 10.4172/2157-7412.1000104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Archer T, Oscar-Berman M, Blum K, Gold M. Neurogenetics and Epigenetics in Impulsive Behaviour: Impact on Reward Circuitry. J Genet Syndr Gene Ther. 2012;3(3):1000115. doi: 10.4172/2157-7412.1000115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Archer T, Oscar-Berman M, Blum K, Gold M. Epigenetic Modulation of Mood Disorders. J Genet Syndr Gene Ther. 2013;4(1):1000120. doi: 10.4172/2157-7412.1000120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Zhang TY, Labonté B, Wen XL, Turecki G, Meaney MJ. Epigenetic mechanisms for the early environmental regulation of hippocampal glucocorticoid receptor gene expression in rodents and humans. Neuropsychopharmacology. 2013;38(1):111–123. doi: 10.1038/npp.2012.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Kandel ER, Kandel DB. Shattuck Lecture. A molecular basis for nicotine as a gateway drug. N Engl J Med. 2014;371:932–943. doi: 10.1056/NEJMsa1405092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kandel D. Stages in adolescent involvement in drug use. Science. 1975;190:912–914. doi: 10.1126/science.1188374. [DOI] [PubMed] [Google Scholar]
- 74.Comings DE, Gade-Andavolu R, Gonzalez N, Wu S, Muhleman D, et al. A multivariate analysis of 59 candidate genes in personality traits: the temperament and character inventory. Clin Genet. 2000;58(5):375–385. doi: 10.1034/j.1399-0004.2000.580508.x. [DOI] [PubMed] [Google Scholar]
- 75.Kelly JF. Self-help for substance-use disorders: history, effectiveness, knowledge gaps, and research opportunities. Clin Psychol Rev. 2003;23(5):639–663. doi: 10.1016/s0272-7358(03)00053-9. [DOI] [PubMed] [Google Scholar]
- 76.Schoenthaler SJ, Blum K, Braverman ER, Giordano J, Thompson B, et al. NIDA-Drug Addiction Treatment Outcome Study (DATOS) Relapse as a Function of Spirituality. J Reward Defic Syndr. 2015;1(1) doi: 10.17756/jrds.2015-007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Karlsson H, Hirvonen J, Salminen JK, Hietala J. No association between serotonin 5-HT 1A receptors and spirituality among patients with major depressive disorders or healthy volunteers. Mol Psychiatry. 2011;16(3):282–285. doi: 10.1038/mp.2009.126. [DOI] [PubMed] [Google Scholar]
- 78.Borg J, Andrée B, Soderstrom H, Farde L. The serotonin system and spiritual experiences. Am J Psychiatry. 2003;160(11):1965–1969. doi: 10.1176/appi.ajp.160.11.1965. [DOI] [PubMed] [Google Scholar]
- 79.Nilsson KW, Damberg M, Ohrvik J, Leppert J, Lindström L, et al. Genes encoding for AP-2beta and the Serotonin Transporter are associated with the Personality Character Spiritual Acceptance. Neurosci Lett. 2007;411(3):233–237. doi: 10.1016/j.neulet.2006.10.051. [DOI] [PubMed] [Google Scholar]
- 80.Brefczynski-Lewis JA, Lutz A, Schaefer HS, Levinson DB, Davidson RJ. Neural correlates of attentional expertise in long-term meditation practitioners. Proc Natl Acad Sci U S A. 2007;104(27):11483–11488. doi: 10.1073/pnas.0606552104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Kozasa EH, Sato JR, Lacerda SS, Barreiros MA, Radvany J, et al. Meditation training increases brain efficiency in an attention task. Neuroimage. 2012;59(1):745–749. doi: 10.1016/j.neuroimage.2011.06.088. [DOI] [PubMed] [Google Scholar]
- 82.Belcher AM, Volkow ND, Moeller FG, Ferré S. Personality traits and vulnerability or resilience to substance use disorders. Trends Cogn Sci. 2014;18(4):211–217. doi: 10.1016/j.tics.2014.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Hides L, Lubman DI, Dawe S. Models of co-occurring substance misuse and psychosis: are personality traits the missing link? Drug Alcohol Rev. 2004;23(4):425–432. doi: 10.1080/09595230412331324545. [DOI] [PubMed] [Google Scholar]
- 84.Zemore SE, Kaskutas LA, Ammon LN. In 12-step groups, helping helps the helper. Addiction. 2004;99(8):1015–1023. doi: 10.1111/j.1360-0443.2004.00782.x. [DOI] [PubMed] [Google Scholar]
- 85.Galanter M, Dermatis H, Post S, Sampson C. Spirituality-based recovery from drug addiction in the twelve-step fellowship of narcotics anonymous. J Addict Med. 2013;7(3):189–195. doi: 10.1097/ADM.0b013e31828a0265. [DOI] [PubMed] [Google Scholar]
- 86.Barr CL, Kidd KK. Population frequencies of the A1 allele at the dopamine D2 receptor locus. Biol Psychiatry. 1993;34(4):204–209. doi: 10.1016/0006-3223(93)90073-m. [DOI] [PubMed] [Google Scholar]
- 87.Blum K, Gold MS. Neuro-chemical activation of brain reward meso-limbic circuitry is associated with relapse prevention and drug hunger: a hypothesis. Med Hypotheses. 2011;76(4):576–584. doi: 10.1016/j.mehy.2011.01.005. [DOI] [PubMed] [Google Scholar]
- 88.Blum K, Chen TJ, Downs BW, Bowirrat A, Waite RL, et al. Neurogenetics of dopaminergic receptor supersensitivity in activation of brain reward circuitry and relapse: proposing “deprivation-amplification relapse therapy” (DART) Postgrad Med. 2009;121(6):176–196. doi: 10.3810/pgm.2009.11.2087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Bossert JM, Gray SM, Lu L, Shaham Y. Activation of group II metabotropic glutamate receptors in the nucleus accumbens shell attenuates context-induced relapse to heroin seeking. Neuropsychopharmacology. 2006;31(10):2197–2209. doi: 10.1038/sj.npp.1300977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Budygin EA, Park J, Bass CE, Grinevich VP, Bonin KD, et al. Aversive stimulus differentially triggers subsecond dopamine release in reward regions. Neuroscience. 2012;201:331–337. doi: 10.1016/j.neuroscience.2011.10.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.David V, Matifas A, Gavello-Baudy S, Decorte L, Kieffer BL, et al. Brain regional FOS expression elicited by the activation of mu-but not delta-opioid receptors of the ventral tegmental area: evidence for an implication of the ventral thalamus in opiate reward. Neuropsychopharmacology. 2008;33(7):1746–1759. doi: 10.1038/sj.npp.1301529. [DOI] [PubMed] [Google Scholar]
- 92.Filip M, Zaniewska M, Frankowska M, Wydra K, Fuxe K. The importance of the adenosine A(2A) receptor-dopamine D(2) receptor interaction in drug addiction. Curr Med Chem. 2012;19(3):317–355. doi: 10.2174/092986712803414231. [DOI] [PubMed] [Google Scholar]
- 93.Ikemoto S. Brain reward circuitry beyond the mesolimbic dopamine system: a neurobiological theory. Neurosci Biobehav Rev. 2010;35(2):129–150. doi: 10.1016/j.neubiorev.2010.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.LaForge KS, Shick V, Spangler R, Proudnikov D, Yuferov V, et al. Detection of single nucleotide polymorphisms of the human mu opioidreceptor gene by hybridization or single nucleotide extension on custom oligonucleotide gelpad microchips: potential in studies of addiction. Am J Med Genet. 2000;96(5):604–615. doi: 10.1002/1096-8628(20001009)96:5<604::aid-ajmg5>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- 95.Lobo MK, Covington HE3rd, Chaudhury D, Friedman AK, Sun H, et al. Cell type-specific loss of BDNF signaling mimics optogenetic control of cocaine reward. Science. 2010;330(6002):385–390. doi: 10.1126/science.1188472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Myers RD, Robinson DE. Mµ and D2 receptor antisense oligonucleotides injected in nucleus accumbens suppress high alcohol intake in genetic drinking HEP rats. Alcohol. 1999;18(2–3):225–233. doi: 10.1016/s0741-8329(99)00015-4. [DOI] [PubMed] [Google Scholar]
- 97.Johnson PM, Kenny PJ. Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat Neurosci. 2010;13(5):635–641. doi: 10.1038/nn.2519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Nestler EJ. Molecular neurobiology of addiction. Am J Addict. 2001;10(3):201–217. doi: 10.1080/105504901750532094. [DOI] [PubMed] [Google Scholar]
- 99.Noble EP, Blum K, Ritchie T, Montgomery A, Sheridan PJ. Allelic association of the D2 dopamine receptor gene with receptor-binding characteristics in alcoholism. Arch Gen Psychiatry. 1991;48(7):648–654. doi: 10.1001/archpsyc.1991.01810310066012. [DOI] [PubMed] [Google Scholar]
- 100.Palomo T, Archer T, Kostrzewa RM, Beninger RJ. Comorbidity of substance abuse with other psychiatric disorders. Neurotox Res. 2007;12(1):17–27. doi: 10.1007/BF03033898. [DOI] [PubMed] [Google Scholar]
- 101.Pierce RC, Kumaresan V. The mesolimbic dopamine system: the final common pathway for the reinforcing effect of drugs of abuse? Neurosci Biobehav Rev. 2006;30(2):215–238. doi: 10.1016/j.neubiorev.2005.04.016. [DOI] [PubMed] [Google Scholar]
- 102.Wang GJ, Smith L, Volkow ND, Telang F, Logan J, et al. Decreased dopamine activity predicts relapse in methamphetamine abusers. Mol Psychiatry. 2012;17(9):918–925. doi: 10.1038/mp.2011.86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Wahlstrom D, White T, Luciana M. Neurobehavioral evidence for changes in dopamine system activity during adolescence. Neurosci Biobehav Rev. 2010;34(5):631–648. doi: 10.1016/j.neubiorev.2009.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Blum K, Chen AL, Oscar-Berman M, Chen TJ, Lubar J, et al. Generational association studies of dopaminergic genes in reward deficiency syndrome (RDS) subjects: selecting appropriate phenotypes for reward dependence behaviors. Int J Environ Res Public Health. 2011;8(12):4425–4459. doi: 10.3390/ijerph8124425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Yacubian J, Sommer T, Schroeder K, Gläscher J, Kalisch R, et al. Gene-gene interaction associated with neural reward sensitivity. Proc Natl Acad Sci U S A. 2007;104(19):8125–8130. doi: 10.1073/pnas.0702029104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Blum K, Liu Y, Wang W, Wang Y, Zhang Y, et al. rsfMRI effects of KB220Z™ on neural pathways in reward circuitry of abstinent genotyped heroin addicts. Postgrad Med. 2015;127(2):232–241. doi: 10.1080/00325481.2015.994879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Gowan T, Whetstone S, Andic T. Addiction, agency, and the politics of self-control: doing harm reduction in a heroin users’ group. Soc Sci Med. 2012;74(8):1251–1260. doi: 10.1016/j.socscimed.2011.11.045. [DOI] [PubMed] [Google Scholar]
- 108.Ford GG. An existential model for promoting life change. Confronting the disease concept. J Subst Abuse Treat. 1996;13(2):151–158. doi: 10.1016/0740-5472(96)00041-4. [DOI] [PubMed] [Google Scholar]
- 109.Greene W, Gold MS. Drug Abuse. In: Bope ET, Kellerman RD, editors. Conn’s Current Therapy. Philadelphia, PA, USA: Saunders, Inc; 2012. pp. 946–951. [Google Scholar]
- 110.Koob GF, Volkow ND. Neurocircuitry of addiction. Neuropsychopharmacology. 2010;35(1):217–238. doi: 10.1038/npp.2009.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Goldstein RZ, Volkow ND. Dysfunction of the prefrontal cortex in addiction: neuroimaging findings and clinical implications. Nat Rev Neurosci. 2011;12:652–669. doi: 10.1038/nrn3119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Herkov MJ, Nias MF, Gold MS. Addiction and Dependence. In: Bruce Jennings., editor. Encyclopedia of Bioethics-4th edition. Farmington Hills, MI, USA: Macmillan Reference; 2013. [Google Scholar]
- 113.Mandyam CD, Koob GF. The addicted brain craves new neurons: putative role for adult-born progenitors in promoting recovery. Trends Neurosci. 2012;35(4):250–260. doi: 10.1016/j.tins.2011.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Hyman SE. The neurobiology of addiction: implications for voluntary control of behavior. Am J Bioeth. 2007;7(1):8–11. doi: 10.1080/15265160601063969. [DOI] [PubMed] [Google Scholar]
- 115.Harris GJ, Jaffin SK, Hodge SM, Kennedy D, Caviness VS, et al. Frontal white matter and cingulum diffusion tensor imaging deficits in alcoholism. Alcohol Clin Exp Res. 2008;32(6):1001–1013. doi: 10.1111/j.1530-0277.2008.00661.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Makris N, Oscar-Berman M, Jaffin SK, Hodge SM, Kennedy DN, et al. Decreased volume of the brain reward system in alcoholism. Biol Psychiatry. 2008;64(3):192–202. doi: 10.1016/j.biopsych.2008.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kienast T, Wrase J, Heinz A. [Neurobiology of substance-related addiction: findings of neuroimaging] Fortschr Neurol Psychiatr. 2008;76(Suppl 1):S68–S76. doi: 10.1055/s-2008-1038141. [DOI] [PubMed] [Google Scholar]
- 118.Yaryura-Tobias JA, Neziroglu FA, Kaplan S. Self-mutilation, anorexia, and dysmenorrhea in obsessive compulsive disorder. Int J Eat Disord. 1995;17:33–38. doi: 10.1002/1098-108x(199501)17:1<33::aid-eat2260170104>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- 119.Koob GF, Le Moal M. Review. Neurobiological mechanisms for opponent motivational processes in addiction. Philos Trans R Soc Lond B Biol Sci. 2008;363(1507):3113–3123. doi: 10.1098/rstb.2008.0094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Broderick PA, Barr GA, Sharpless NS, Bridger WH. Biogenic amine alterations in limbic brain regions of muricidal rats. Res Commun Chem Pathol Pharmacol. 1985;48(1):3–15. [PubMed] [Google Scholar]
- 121.Blum K, Liu Y, Shriner R, Gold MS. Reward circuitry dopaminergic activation regulates food and drug craving behavior. Curr Pharm Des. 2011;17(12):1158–1167. doi: 10.2174/138161211795656819. [DOI] [PubMed] [Google Scholar]
- 122.Gardner EL. Addiction and brain reward and antireward pathways. Adv Psychosom Med. 2011;30:22–60. doi: 10.1159/000324065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Brickman P, Campbell DT. Hedonic relativism and planning the good society. In: Apley MH, editor. Adaptation Level Theory: A Symposium. New York: Academic Press; 1971. pp. 287–302. [Google Scholar]
- 124.Miller NS, Gold MS. Dissociation of “conscious desire” (craving) from and relapse in alcohol and cocaine dependence. Ann Clin Psychiatry. 1994;6(2):99–106. doi: 10.3109/10401239409148988. [DOI] [PubMed] [Google Scholar]
- 125.Diana M. The dopamine hypothesis of drug addiction and its potential therapeutic value. Front Psychiatry. 2011;2:64. doi: 10.3389/fpsyt.2011.00064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Panksepp J, Knutson B, Burgdorf J. The role of brain emotional systems in addictions: a neuro-evolutionary perspective and new ‘self-report’ animal model. Addiction. 2002;97(4):459–469. doi: 10.1046/j.1360-0443.2002.00025.x. [DOI] [PubMed] [Google Scholar]
- 127.Smith DE. The process addictions and the new ASAM definition of addiction. J Psychoactive Drugs. 2012;44(1):1–4. doi: 10.1080/02791072.2012.662105. [DOI] [PubMed] [Google Scholar]
- 128.Vetulani J. Drug addiction. Part II. Neurobiology of addiction. Pol J Pharmacol. 2001;53(4):303–317. [PubMed] [Google Scholar]
- 129.Blum K, Briggs AH, DeLallo L, Elston SF, Ochoa R. Whole brain methionine-enkephalin of ethanol-avoiding and ethanol-preferring c57BL mice. Experientia. 1982;38(12):1469–1470. doi: 10.1007/BF01955775. [DOI] [PubMed] [Google Scholar]
- 130.Blum K, DeLallo L, Briggs AH, Hamilton MG. Opioid responses of isoquinoline alkaloids (TIQs) Prog Clin Biol Res. 1982;90:387–398. [PubMed] [Google Scholar]
- 131.Schulz R, Wüster M, Duka T, Herz A. Acute and chronic ethanol treatment changes endorphin levels in brain and pituitary. Psychopharmacology (Berl) 1980;68(3):221–227. doi: 10.1007/BF00428107. [DOI] [PubMed] [Google Scholar]
- 132.Turchan J, LasoÅ W, Budziszewska B, PrzewÅocka B. Effects of single and repeated morphine administration on the prodynorphin, proenkephalin and dopamine D2 receptor gene expression in the mouse brain. Neuropeptides. 1997;31(1):24–28. doi: 10.1016/s0143-4179(97)90015-9. [DOI] [PubMed] [Google Scholar]
- 133.Duka T, Wüster M, Herz A. Rapid changes in enkephalin levels in rat striatum and hypothalamus induced by diazepam. Naunyn Schmiedebergs Arch Pharmacol. 1979;309(1):1–5. doi: 10.1007/BF00498749. [DOI] [PubMed] [Google Scholar]
- 134.Wüster M, Schulz R, Herz A. Inquiry into endorphinergic feedback mechanisms during the development of opiate tolerance/dependence. Brain Res. 1980;189(2):403–411. doi: 10.1016/0006-8993(80)90100-6. [DOI] [PubMed] [Google Scholar]
- 135.Sharpe LG, Pilotte NS, Shippenberg TS, Goodman CB, London ED. Autoradiographic evidence that prolonged withdrawal from intermittent cocaine reduces mu-opioid receptor expression in limbic regions of the rat brain. Synapse. 2000;37(4):292–297. doi: 10.1002/1098-2396(20000915)37:4<292::AID-SYN6>3.0.CO;2-V. [DOI] [PubMed] [Google Scholar]
- 136.Kurtz E. The collected Ernie Kurtz. New York: Hindsfoot Foundation Series on Treatment and Recovery, Authors Choice; 2008. Research on alcoholics anonymous: the historical context; pp. 1–22. [Google Scholar]
- 137.Charlton BG. Genospirituality: genetic engineering for spiritual and religious enhancement. Med Hypotheses. 2008;71(6):825–828. doi: 10.1016/j.mehy.2008.06.006. [DOI] [PubMed] [Google Scholar]
- 138.McNamara P. The Neuroscience of Religious Experience. Cambridge, UK: Cambridge University Press; 2009. [Google Scholar]
- 139.Wildman WJ. Religious and Spiritual Experiences. Cambridge, UK: Cambridge University Press; 2011. [Google Scholar]
- 140.Braverman ER. The religious medical model: holy medicine and the Spiritual Behavior Inventory. South Med J. 1987;80(4):415–420. doi: 10.1097/00007611-198704000-00002. 425. [DOI] [PubMed] [Google Scholar]
- 141.Sulmasy DP. Spiritual issues in the care of dying patients: “… it’s okay between me and god”. JAMA. 2006;296(11):1385–1392. doi: 10.1001/jama.296.11.1385. [DOI] [PubMed] [Google Scholar]
- 142.Kirk KM, Eaves LJ, Martin NG. Self-transcendence as a measure of spirituality in a sample of older Australian twins. Twin Res. 1999;2(2):81–87. doi: 10.1375/136905299320565942. [DOI] [PubMed] [Google Scholar]
- 143.Comings DE. Is Your Spiritual Brain in Peace with Your Thinking Brain? Durate, California, USA: Hope Press; 2008. Did Man Create God? [Google Scholar]
- 144.Newberg A. The brain and the biology of belief: An interview with Andrew Newberg, MD. Interview by Nancy Nachman-Hunt. Adv Mind Body Med. 2009;24(1):32–36. [PubMed] [Google Scholar]
- 145.Comings DE, Blum K. Reward deficiency syndrome: genetic aspects of behavioral disorders. Prog Brain Res. 2000;126:325–341. doi: 10.1016/S0079-6123(00)26022-6. [DOI] [PubMed] [Google Scholar]
- 146.Chopra D. Life after Death. New York, USA: Three Rivers Press; 2006. [Google Scholar]
- 147.Cloninger CR, Svrakic DM, Przybeck TR. A psychobiological model of temperament and character. Arch Gen Psychiatry. 1993;50(12):975–990. doi: 10.1001/archpsyc.1993.01820240059008. [DOI] [PubMed] [Google Scholar]
- 148.Asghari V, Sanyal S, Buchwaldt S, Paterson A, Jovanovic V, et al. Modulation of intracellular cyclic AMP levels by different human dopamine D4 receptor variants. J Neurochem. 1995;65(3):1157–1165. doi: 10.1046/j.1471-4159.1995.65031157.x. [DOI] [PubMed] [Google Scholar]
- 149.Gelernter J, Kranzler H, Coccaro E, Siever L, New A, et al. D4 dopamine-receptor (DRD4) alleles and novelty seeking in substance-dependent, personality-disorder, and control subjects. Am J Hum Genet. 1997;61(5):1144–1152. doi: 10.1086/301595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Gold MS, Dackis CA. New insights and treatments: opiate withdrawal and cocaine addiction. Clin Ther. 1984;7(1):6–21. [PubMed] [Google Scholar]
- 151.Shultes RE, Hoffman A, Ratsch C. Plants of the Gods. Their Sacred Healing and Hallucinogenic Powers. Rochester, NY, USA: Healing Arts Press; 1998. [Google Scholar]
- 152.Fromm E. The Sane Society. New York, USA: Rinehart; 1955. [Google Scholar]
- 153.Bowirrat A, Chen TJ, Oscar-Berman M, Madigan M, Chen AL, et al. Neuropsychopharmacology and neurogenetic aspects of executive functioning: should reward gene polymorphisms constitute a diagnostic tool to identify individuals at risk for impaired judgment? Mol Neurobiol. 2012;45(2):298–313. doi: 10.1007/s12035-012-8247-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Bertolino A, Taurisano P, Pisciotta NM, Blasi G, Fazio L, et al. Genetically determined measures of striatal D2 signaling predict prefrontal activity during working memory performance. PLoS One. 2010;5(2):e9348. doi: 10.1371/journal.pone.0009348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Markett SA, Montag C, Reuter M. The association between dopamine DRD2 polymorphisms and working memory capacity is modulated by a functional polymorphism on the nicotinic receptor gene CHRNA4. J Cogn Neurosci. 2010;22(9):1944–1954. doi: 10.1162/jocn.2009.21354. [DOI] [PubMed] [Google Scholar]
- 156.Frank MJ, Hutchison K. Genetic contributions to avoidance-based decisions: striatal D2 receptor polymorphisms. Neuroscience. 2009;164(1):131–140. doi: 10.1016/j.neuroscience.2009.04.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Drew MR, Simpson EH, Kellendonk C, Herzberg WG, Lipatova O, et al. Transient overexpression of striatal D2 receptors impairs operant motivation and interval timing. J Neurosci. 2007;27(29):7731–7739. doi: 10.1523/JNEUROSCI.1736-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Xu TX, Sotnikova TD, Liang C, Zhang J, Jung JU, et al. Hyperdopaminergic tone erodes prefrontal long-term potential via a D2 receptor-operated protein phosphatase gate. J Neurosci. 2009;29(45):14086–14099. doi: 10.1523/JNEUROSCI.0974-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Stelzel C, Basten U, Montag C, Reuter M, Fiebach CJ. Effects of dopamine-related gene-gene interactions on working memory component processes. Eur J Neurosci. 2009;29(5):1056–1063. doi: 10.1111/j.1460-9568.2009.06647.x. [DOI] [PubMed] [Google Scholar]
- 160.Blanchard MM, Chamberlain SR, Roiser J, Robbins TW, Müller U. Effects of two dopamine-modulating genes (DAT1 9/10 and COMT Val/Met) on n-back working memory performance in healthy volunteers. Psychol Med. 2011;41(3):611–618. doi: 10.1017/S003329171000098X. [DOI] [PubMed] [Google Scholar]
- 161.Stroth S, Reinhardt RK, Thöne J, Hille K, Schneider M, et al. Impact of aerobic exercise training on cognitive functions and affect associated to the COMT polymorphism in young adults. Neurobiol Learn Mem. 2010;94(3):364–372. doi: 10.1016/j.nlm.2010.08.003. [DOI] [PubMed] [Google Scholar]
- 162.Bousman CA, Cherner M, Atkinson JH, Heaton RK, Grant I, et al. COMT Val158Met Polymorphism, Executive Dysfunction, and Sexual Risk Behavior in the Context of HIV Infection and Methamphetamine Dependence. Interdiscip Perspect Infect Dis. 2010 doi: 10.1155/2010/678648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Blum K, Briggs AH, Wallace JE, Hall CW, Trachtenberg MA. Regional brain [Met]-enkephalin in alcohol-preferring and non-alcohol preferring inbred strains of mice. Experientia. 1987;43(4):408–410. doi: 10.1007/BF01940430. [DOI] [PubMed] [Google Scholar]
- 164.Bowirrat A, Oscar-Berman M. Relationship between dopaminergic neurotransmission, alcoholism, and Reward Deficiency syndrome. Am J Med Genet B Neuropsychiatr Genet. 2005;132B(1):29–37. doi: 10.1002/ajmg.b.30080. [DOI] [PubMed] [Google Scholar]
- 165.Blum K, Chen ALC, Chen TJH, et al. Genes and Happiness. Gen Ther Mol Biol. 2009;13:91–129. [Google Scholar]
- 166.Oscar-Berman M, Marinkovic K. Alcohol: effects on neurobehavioral functions and the brain. Neuropsychol Rev. 2007;17(3):239–257. doi: 10.1007/s11065-007-9038-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Lyvers M, Tobias-Webb J. Effects of acute alcohol consumption on executive cognitive functioning in naturalistic settings. Addict Behav. 2010;35(11):1021–1028. doi: 10.1016/j.addbeh.2010.06.022. [DOI] [PubMed] [Google Scholar]
- 168.Barnes JJ, Dean AJ, Nandam LS, O’Connell RG, Bellgrove MA. The molecular genetics of executive function: role of monoamine system genes. Biol Psychiatry. 2011;69(12):e127–e143. doi: 10.1016/j.biopsych.2010.12.040. [DOI] [PubMed] [Google Scholar]
- 169.Zhang Y, Bertolino A, Fazio L, Blasi G, Rampino A, et al. Polymorphisms in human dopamine d2 receptor gene affect gene expression, splicing, and neuronal activity during working memory. Proc Natl Acad Sci U S A. 2007;104(51):20552–20557. doi: 10.1073/pnas.0707106104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Arnsten AF. Toward a new understanding of attention-deficit hyperactivity disorder pathophysiology: an important role for prefrontal cortex dysfunction. CNS Drugs. 2009;23(Suppl 1):33–41. doi: 10.2165/00023210-200923000-00005. [DOI] [PubMed] [Google Scholar]
- 171.Dickinson D, Elvevåg B. Genes, cognition and brain through a COMT lens. Neuroscience. 2009;164(1):72–87. doi: 10.1016/j.neuroscience.2009.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Haluk DM, Floresco SB. Ventral striatal dopamine modulation of different forms of behavioral flexibility. Neuropsychopharmacology. 2009;34(8):2041–2052. doi: 10.1038/npp.2009.21. [DOI] [PubMed] [Google Scholar]
- 173.MacDonald SW, Li SC, Bäckman L. Neural underpinnings of within-person variability in cognitive functioning. Psychol Aging. 2009;24(4):792–808. doi: 10.1037/a0017798. [DOI] [PubMed] [Google Scholar]
- 174.Pinto E, Ansseau M. [Genetic factors of alcohol-dependence] Encephale. 2009;35(5):461–469. doi: 10.1016/j.encep.2008.03.008. [DOI] [PubMed] [Google Scholar]
- 175.Pinto E, Reggers J, Gorwood P, Boni C, Scantamburlo G, et al. The TaqI A DRD2 polymorphism in type II alcohol dependence: a marker of age at onset or of a familial disease? Alcohol. 2009;43(4):271–275. doi: 10.1016/j.alcohol.2009.02.006. [DOI] [PubMed] [Google Scholar]
- 176.Heck A, Pfister H, Czamara D, Müller-Myhsok B, Pütz B, et al. Evidence for Associations between MDGA2 Polymorphisms and Harm Avoidance-Replication and Extension of a Genome-wide Association Finding. Psychiatric Genetics. 2011;21(5):257–260. doi: 10.1097/YPG.0b013e3283457bfb. [DOI] [PubMed] [Google Scholar]
- 177.Dreyer JK, Herrik KF, Berg RW, Hounsgaard JD. Influence of phasic and tonic dopamine release on receptor activation. J Neurosci. 2010;30(42):14273–14283. doi: 10.1523/JNEUROSCI.1894-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Lawson-Yuen A, Saldivar JS, Sommer S, Picker J. Familial deletion within NLGN4 associated with autism and Tourette syndrome. Eur J Hum Genet. 2008;16(5):614–618. doi: 10.1038/sj.ejhg.5202006. [DOI] [PubMed] [Google Scholar]
- 179.Sundaram SK, Huq AM, Wilson BJ, Chugani HT. Tourette syndrome is associated with recurrent exonic copy number variants. Neurology. 2010;74(20):1583–1590. doi: 10.1212/WNL.0b013e3181e0f147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.McClearn GE. Genetics as a tool in alcohol research. Ann N Y Acad Sci. 1972;197:26–31. doi: 10.1111/j.1749-6632.1972.tb28114.x. [DOI] [PubMed] [Google Scholar]
- 181.McBride WJ, Li TK. Animal models of alcoholism: neurobiology of high alcohol-drinking behavior in rodents. Crit Rev Neurobiol. 1998;12(4):339–369. doi: 10.1615/critrevneurobiol.v12.i4.40. [DOI] [PubMed] [Google Scholar]
- 182.Blum K, Topel H. Opioid peptides and alcoholism: genetic deficiency and chemical management. Funct Neurol. 1986;1(1):71–83. [PubMed] [Google Scholar]
- 183.Blum K, Wallace JE, Briggs AH, Trachtenberg MC. Evidence for the importance of the “genotype” theory in alcohol seeking behavior: a commentary. Alcohol Drug Res. 1985;6(6):455–461. [PubMed] [Google Scholar]
- 184.Blum K, Elston SF, DeLallo L, Briggs AH, Wallace JE. Ethanol acceptance as a function of genotype amounts of brain [Met]enkephalin. Proc Natl Acad Sci U S A. 1983;80(21):6510–6512. doi: 10.1073/pnas.80.21.6510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Blum K, Briggs AH, Trachtenberg MC, Delallo L, Wallace JE. Enkephalinase inhibition: regulation of ethanol intake in genetically predisposed mice. Alcohol. 1987;4(6):449–456. doi: 10.1016/0741-8329(87)90084-x. [DOI] [PubMed] [Google Scholar]
- 186.Campbell JC, Szumlinski KK, Kippin TE. Contribution of early environmental stress to alcoholism vulnerability. Alcohol. 2009;43(7):547–554. doi: 10.1016/j.alcohol.2009.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Blum K, Noble EP, Sheridan PJ, Finley O, Montgomery A, et al. Association of the A1 allele of the D2 dopamine receptor gene with severe alcoholism. Alcohol. 1991;8(5):409–416. doi: 10.1016/0741-8329(91)90693-q. [DOI] [PubMed] [Google Scholar]
- 188.Volkow ND, Wang GJ, Begleiter H, Porjesz B, Fowler JS, et al. High levels of dopamine D2 receptors in unaffected members of alcoholic families: possible protective factors. Arch Gen Psychiatry. 2006;63:999–1008. doi: 10.1001/archpsyc.63.9.999. [DOI] [PubMed] [Google Scholar]
- 189.Volkow ND, Fowler JS, Wang GJ. The addicted human brain: insights from imaging studies. J Clin Invest. 2003;111:1444–1451. doi: 10.1172/JCI18533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Yuan Y, Zhu Z, Shi J, Zou Z, Yuan F, et al. Gray matter density negatively correlates with duration of heroin use in young lifetime heroin-dependent individuals. Brain Cogn. 2009;71(3):223–228. doi: 10.1016/j.bandc.2009.08.014. [DOI] [PubMed] [Google Scholar]
- 191.Bell RP, Foxe JJ, Nierenberg J, Hoptman MJ, Garavan H. Assessing white matter integrity as a function of abstinence duration in former cocaine-dependent individuals. Drug Alcohol Depend. 2011;114(2–3):159–168. doi: 10.1016/j.drugalcdep.2010.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Wang GJ, Geliebter A, Volkow ND, Telang FW, Logan J, et al. Enhanced striatal dopamine release during food stimulation in binge eating disorder. Obesity (Silver Spring) 2011;19(8):1601–1608. doi: 10.1038/oby.2011.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Goldstein RZ, Leskovjan AC, Hoff AL, Hitzemann R, Bashan F, et al. Severity of neuropsychological impairment in cocaine and alcohol addiction: association with metabolism in the prefrontal cortex. Neuropsychologia. 2004;42(11):1447–1458. doi: 10.1016/j.neuropsychologia.2004.04.002. [DOI] [PubMed] [Google Scholar]
- 194.Zijlstra F, Booij J, van den Brink W, Franken IH. Striatal dopamine D2 receptor binding and dopamine release during cue-elicited craving in recently abstinent opiate-dependent males. Eur Neuropsychopharmacol. 2008;18(4):262–270. doi: 10.1016/j.euroneuro.2007.11.002. [DOI] [PubMed] [Google Scholar]
- 195.Ruiz SM, Oscar-Berman M, Sawyer KS, Valmas MM, Urban T, et al. Drinking history associations with regional white matter volumes in alcoholic men and women. Alcohol Clin Exp Res. 2013;37(1):110–122. doi: 10.1111/j.1530-0277.2012.01862.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Pritchard JK, Di Rienzo A. Adaptation-not by sweeps alone. Nat Rev Genet. 2010;11(10):665–667. doi: 10.1038/nrg2880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Moore LG, Zamudio S, Zhuang J, Droma T, Shohet RV. Analysis of the myoglobin gene in Tibetans living at high altitude. High Alt Med Biol. 2002;3(1):39–47. doi: 10.1089/152702902753639531. [DOI] [PubMed] [Google Scholar]
- 198.Chen TJ, Blum K, Mathews D, Fisher L, Schnautz N, et al. Are dopaminergic genes involved in a predisposition to pathological aggression? Hypothesizing the importance of “super normal controls” in psychiatricgenetic research of complex behavioral disorders. Med Hypotheses. 2005;65:703–707. doi: 10.1016/j.mehy.2005.04.037. [DOI] [PubMed] [Google Scholar]
- 199.Costa E, Guidotti A. Molecular mechanisms in the receptor action of benzodiazepines. Annu Rev Pharmacol Toxicol. 1979;19:531–545. doi: 10.1146/annurev.pa.19.040179.002531. [DOI] [PubMed] [Google Scholar]
- 200.Costa E, Corda MG, Guidotti A. On a brain polypeptide functioning as a putative effector for the recognition sites of benzodiazepine and beta-carboline derivatives. Neuropharmacology. 1983;22(12B):1481–1492. doi: 10.1016/0028-3908(83)90116-8. [DOI] [PubMed] [Google Scholar]
- 201.Guidotti A, Forchetti CM, Corda MG, Konkel D, Bennett CD, et al. Isolation, characterization, and purification to homogeneity of an endogenous polypeptide with agonistic action on benzodiazepine receptors. Proc Natl Acad Sci U S A. 1983;80(11):3531–3535. doi: 10.1073/pnas.80.11.3531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.van Kammen DP, Guidotti A, Kelley ME, Gurklis J, Guarneri P, et al. CSF diazepam binding inhibitor and schizophrenia: clinical and biochemical relationships. Biol Psychiatry. 1993;34(8):515–522. doi: 10.1016/0006-3223(93)90193-h. [DOI] [PubMed] [Google Scholar]
- 203.Ferrero P, Costa E, Conti-Tronconi B, Guidotti A. A diazepam binding inhibitor (DBI)-like neuropeptide is detected in human brain. Brain Res. 1986;399(1):136–142. doi: 10.1016/0006-8993(86)90607-4. [DOI] [PubMed] [Google Scholar]
- 204.Alho H, Miyata M, Korpi E, Kiianmaa K, Guidotti A. Studies of a brain polypeptide functioning as a putative endogenous ligand to benzodiazepine recognition sites in rats selectively bred for alcohol related behavior. Alcohol Alcohol. 1987;(Suppl 1):637–641. [PubMed] [Google Scholar]
- 205.Blum K, Chen TJ, Meshkin B, Waite RL, Downs BW, et al. Manipulation of catechol-O-methyl-transferase (COMT) activity to influence the attenuation of substance seeking behavior, a subtype of Reward Deficiency Syndrome (RDS), is dependent upon gene polymorphisms: a hypothesis. Med Hypotheses. 2007;69(5):1054–1060. doi: 10.1016/j.mehy.2006.12.062. [DOI] [PubMed] [Google Scholar]
- 206.Linnoila M, Virkkunen M, Scheinin M, Nuutila A, Rimon R, et al. Low cerebrospinal fluid 5-hydroxyindoleacetic acid concentration differentiates impulsive from nonimpulsive violent behavior. Life Sci. 1983;33(26):2609–2614. doi: 10.1016/0024-3205(83)90344-2. [DOI] [PubMed] [Google Scholar]
- 207.Muehlenkamp F, Lucion A, Vogel WH. Effects of selective serotonergic agonists on aggressive behavior in rats. Pharmacol Biochem Behav. 1995;50:671–674. doi: 10.1016/0091-3057(95)00351-7. [DOI] [PubMed] [Google Scholar]
- 208.Barbaccia ML, Costa E, Ferrero P, Guidotti A, Roy A, et al. Diazepam-binding inhibitor. A brain neuropeptide present in human spinal fluid: studies in depression, schizophrenia, and Alzheimer’s disease. Arch Gen Psychiatry. 1986;43(12):1143–1147. doi: 10.1001/archpsyc.1986.01800120029007. [DOI] [PubMed] [Google Scholar]
- 209.Campbell V, Bond R. Evaluation of a character education curriculum. In: McClelland D, editor. Education for Values. New York, USA: Irvington Publishers; 1982. [Google Scholar]
- 210.Anscombe GEM. Modern moral philosophy 1. Philosophy. 1958;33(124):1–19. [Google Scholar]
- 211.Perry JC, Körner AC. Impulsive phenomena, the impulsive character (der Triebhafte Charakter) and DSM personality disorders. J Pers Disord. 2011;25(5):586–606. doi: 10.1521/pedi.2011.25.5.586. [DOI] [PubMed] [Google Scholar]
- 212.Doris JM. Lack of Character: Personality and Moral Behavior. Cambridge University Press; 2000. p. 284. [Google Scholar]
- 213.Homiak M. Moral character. In: Zalta EN, editor. The Stanford Encyclopedia of Philosophy (Fall Edition) 2008. [Google Scholar]
- 214.Huitt W. Educational Psychology Interactive. Valdosta, GA: Valdosta State University; 2004. Moral and Character Development. [Google Scholar]
- 215.Lawrence LE. Reality is reality. J Natl Med Assoc. 1994;86(6):417–419. [PMC free article] [PubMed] [Google Scholar]
- 216.Comings DE, MacMurray J, Johnson P, Dietz G, Muhleman D. Dopamine D2 receptor gene (DRD2) haplotypes and the defense style questionnaire in substance abuse, Tourette syndrome, and controls. Biol Psychiatry. 1995;37(11):798–805. doi: 10.1016/0006-3223(94)00222-O. [DOI] [PubMed] [Google Scholar]
- 217.Bond M, Gardner ST, Christian J, Sigal JJ. Empirical study of self-rated defense styles. Arch Gen Psychiatry. 1983;40(3):333–338. doi: 10.1001/archpsyc.1983.01790030103013. [DOI] [PubMed] [Google Scholar]
- 218.Andrews G, Pollock C, Stewart G. The determination of defense style by questionnaire. Arch Gen Psychiatry. 1989;46(5):455–460. doi: 10.1001/archpsyc.1989.01810050069011. [DOI] [PubMed] [Google Scholar]
- 219.Frank E, Salchner P, Aldag JM, Salomé N, Singewald N, et al. Genetic predisposition to anxiety-related behavior determines coping style, neuroendocrine responses, and neuronal activation during social defeat. Behavioral Neuroscience. 2006;120(1):60–71. doi: 10.1037/0735-7044.120.1.60. [DOI] [PubMed] [Google Scholar]
- 220.Bond MP, Vaillant JS. An empirical study of the relationship between diagnosis and defense style. Arch Gen Psychiatry. 1986;43(3):285–288. doi: 10.1001/archpsyc.1986.01800030103012. [DOI] [PubMed] [Google Scholar]
- 221.Vaillant GE, Bond M, Vaillant CO. An empirically validated hierarchy of defense mechanisms. Arch Gen Psychiatry. 1986;43(8):786–794. doi: 10.1001/archpsyc.1986.01800080072010. [DOI] [PubMed] [Google Scholar]
- 222.Pollock C, Andrews G. Defense styles associated with specific anxiety disorders. Am J Psychiatry. 1989;146(11):1500–1502. doi: 10.1176/ajp.146.11.1500. [DOI] [PubMed] [Google Scholar]
- 223.Brennan J, Andrews G, Morris-Yates A, Pollock C. An examination of defense style in parents who abuse children. J Nerv Ment Dis. 1990;178(9):592–595. doi: 10.1097/00005053-199009000-00007. [DOI] [PubMed] [Google Scholar]
- 224.Kesby G, Parker G, Barrett E. Personality and coping style as influences on alcohol intake and cigarette smoking during pregnancy. Med J Aust. 1991;155(4):229–233. doi: 10.5694/j.1326-5377.1991.tb142227.x. [DOI] [PubMed] [Google Scholar]
- 225.Andrews G. The changing nature of psychiatry. Aust N Z J Psychiatry. 1991;25(4):453–459. doi: 10.3109/00048679109064438. [DOI] [PubMed] [Google Scholar]
- 226.Dean JC, Poremba GA. The alcoholic stigma and the disease concept. Int J Addict. 1983;18(5):739–751. doi: 10.3109/10826088309027366. [DOI] [PubMed] [Google Scholar]
- 227.Iversen SD, Alpert JE. Functional organization of the dopamine system in normal and abnormal behavior. Adv Neurol. 1982;35:69–76. [PubMed] [Google Scholar]
- 228.Louilot A, Le Moal M, Simon H. Opposite influences of dopaminergic pathways to the prefrontal cortex or the septum on the dopaminergic transmission in the nucleus accumbens. An in vivo voltammetric study. Neuroscience. 1989;29(1):45–56. doi: 10.1016/0306-4522(89)90331-x. [DOI] [PubMed] [Google Scholar]
- 229.Blackburn JR, Pfaus JG, Phillips AG. Dopamine functions in appetitive and defensive behaviours. Prog Neurobiol. 1992;39(3):247–279. doi: 10.1016/0301-0082(92)90018-a. [DOI] [PubMed] [Google Scholar]
- 230.Lorenzi M, Karam JH, McIlroy MB, Forsham PH. Increased growth hormone response to dopamine infusion in insulin-dependent diabetic subjects: indication of possible blood-brain barrier abnormality. J Clin Invest. 1980;65(1):146–153. doi: 10.1172/JCI109644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Routtenberg A. The reward system of the brain. Sci Am. 1978;239(5):154–164. doi: 10.1038/scientificamerican1178-154. [DOI] [PubMed] [Google Scholar]
- 232.Wise RA, Rompre PP. Brain dopamine and reward. Annu Rev Psychol. 1989;40:191–225. doi: 10.1146/annurev.ps.40.020189.001203. [DOI] [PubMed] [Google Scholar]
- 233.Bunzow JR, Van Tol HH, Grandy DK, Albert P, Salon J, et al. Cloning and expression of a rat D2 dopamine receptor cDNA. Nature. 1988;336(6201):783–787. doi: 10.1038/336783a0. [DOI] [PubMed] [Google Scholar]
- 234.Hauge XY, Grandy DK, Eubanks JH, Evans GA, Civelli O, et al. Detection and characterization of additional DNA polymorphisms in the dopamine D2 receptor gene. Genomics. 1991;10(3):527–530. doi: 10.1016/0888-7543(91)90431-d. [DOI] [PubMed] [Google Scholar]
- 235.Uhl G, Blum K, Noble E, Smith S. Substance abuse vulnerability and D2 receptor genes. Trends Neurosci. 1993;16(3):83–88. doi: 10.1016/0166-2236(93)90128-9. [DOI] [PubMed] [Google Scholar]
- 236.Noble EP. The D2 dopamine receptor gene: a review of association studies in alcoholism. Behav Genet. 1993;23(2):119–129. doi: 10.1007/BF01067416. [DOI] [PubMed] [Google Scholar]
- 237.Smith SS, O’Hara BF, Persico AM, Gorelick DA, Newlin DB, et al. Genetic vulnerability to drug abuse. The D2 dopamine receptor Taq I B1 restriction fragment length polymorphism appears more frequently in polysubstance abusers. Arch Gen Psychiatry. 1992;49(9):723–727. doi: 10.1001/archpsyc.1992.01820090051009. [DOI] [PubMed] [Google Scholar]
- 238.Comings DE, Flanagan SD, Dietz G, Muhleman D, Knell E, et al. The dopamine D2 receptor (DRD2) as a major gene in obesity and height. Biochem Med Metab Biol. 1993;50(2):176–185. doi: 10.1006/bmmb.1993.1059. [DOI] [PubMed] [Google Scholar]
- 239.O’Hara BF, Smith SS, Bird G, Persico AM, Suarez BK, et al. Dopamine D2 receptor RFLPs, haplotypes and their association with substance use in black and Caucasian research volunteers. Hum Hered. 1993;43(4):209–218. doi: 10.1159/000154133. [DOI] [PubMed] [Google Scholar]
- 240.Nyman ES, Loukola A, Varilo T, Taanila A, Hurtig T, et al. Sex-specific influence of DRD2 on ADHD-type temperament in a large population-based birth cohort. Psychiatr Genet. 2012;22(4):197–201. doi: 10.1097/YPG.0b013e32834c0cc8. [DOI] [PubMed] [Google Scholar]
- 241.Comings DE, Rosenthal RJ, Lesieur HR, Rugle LJ, Muhleman D, et al. A study of the dopamine D2 receptor gene in pathological gambling. Pharmacogenetics. 1996;6(3):223–234. doi: 10.1097/00008571-199606000-00004. [DOI] [PubMed] [Google Scholar]
- 242.Myung W, Lim SW, Kim J, Lee Y, Song J, et al. Serotonin transporter gene polymorphisms and chronic illness of depression. J Korean Med Sci. 2010;25(12):1824–1827. doi: 10.3346/jkms.2010.25.12.1824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.McLellan AT, Lewis DC, O’Brien CP, Kleber HD. Drug dependence, a chronic medical illness: implications for treatment, insurance, and outcomes evaluation. JAMA. 2000;284(13):1689–1695. doi: 10.1001/jama.284.13.1689. [DOI] [PubMed] [Google Scholar]
- 244.Wojnar M, Brower KJ, Strobbe S, Ilgen M, Matsumoto H, et al. Association between Val66Met brain-derived neurotrophic factor (BDNF) gene polymorphism and post-treatment relapse in alcohol dependence. Alcohol Clin Exp Res. 2009;33(4):693–702. doi: 10.1111/j.1530-0277.2008.00886.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Fowler JH, Dawes CT, Christakis NA. Model of genetic variation in human social networks. Proc Natl Acad Sci U S A. 2009;106(6):1720–1724. doi: 10.1073/pnas.0806746106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Jackson KJ, McIntosh JM, Brunzell DH, Sanjakdar SS, Damaj MI. The role of alpha6-containing nicotinic acetylcholine receptors in nicotine reward and withdrawal. J Pharmacol Exp Ther. 2009;331(2):547–554. doi: 10.1124/jpet.109.155457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Kendler KS, Baker JH. Genetic influences on measures of the environment: a systematic review. Psychol Med. 2007;37(5):615–626. doi: 10.1017/S0033291706009524. [DOI] [PubMed] [Google Scholar]
- 248.Iervolino AC, Pike A, Manke B, Reiss D, Hetherington EM, et al. Genetic and environmental influences in adolescent peer socialization: evidence from two genetically sensitive designs. Child Dev. 2002;73(1):162–174. doi: 10.1111/1467-8624.00398. [DOI] [PubMed] [Google Scholar]
- 249.Guo G. Genetic similarity shared by best friends among adolescents. Twin Res Hum Genet. 2006;9(1):113–121. doi: 10.1375/183242706776402920. [DOI] [PubMed] [Google Scholar]
- 250.Dudley SA, File AL. Kin recognition in an annual plant. Biol Lett. 2007;3(4):435–438. doi: 10.1098/rsbl.2007.0232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Burt A. A mechanistic explanation of popularity: genes, rule breaking, and evocative gene-environment correlations. J Pers Soc Psychol. 2009;96(4):783–794. doi: 10.1037/a0013702. [DOI] [PubMed] [Google Scholar]
- 252.Dick DM, Pagan JL, Holliday C, Viken R, Pulkkinen L, et al. Gender differences in friends’ influences on adolescent drinking: a genetic epidemiological study. Alcohol Clin Exp Res. 2007;31(12):2012–2019. doi: 10.1111/j.1530-0277.2007.00523.x. [DOI] [PubMed] [Google Scholar]
- 253.Fowler JH, Settle JE, Christakis NA. Correlated genotypes in friendship networks. Proc Natl Acad Sci U S A. 2011;108(5):1993–1997. doi: 10.1073/pnas.1011687108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Diener E, Lucas RE, Scollon CN. Beyond the hedonic treadmill: revising the adaptation theory of well-being. Am Psychol. 2006;61(5):305–314. doi: 10.1037/0003-066X.61.4.305. [DOI] [PubMed] [Google Scholar]
- 255.Haller J, Makara GB, Kovács JL. The effect of alpha 2 adrenoceptor blockers on aggressive behavior in mice: implications for the actions of adrenoceptor agents. Psychopharmacology (Berl) 1996;126(4):345–350. doi: 10.1007/BF02247386. [DOI] [PubMed] [Google Scholar]
- 256.Bruijnzeel AW, Repetto M, Gold MS. Neurobiological mechanisms in addictive and psychiatric disorders. Psychiatr Clin North Am. 2004;27(4):661–674. doi: 10.1016/j.psc.2004.06.005. [DOI] [PubMed] [Google Scholar]
- 257.Kahneman D, Krueger AB, Schkade D, Schwarz N, Stone AA. Would you be happier if you were richer? A focusing illusion. Science. 2006;312(5782):1908–1910. doi: 10.1126/science.1129688. [DOI] [PubMed] [Google Scholar]
- 258.Guo G, Roettger ME, Shih JC. Contributions of the DAT1 and DRD2 genes to serious and violent delinquency among adolescents and young adults. Hum Genet. 2007;121(1):125–136. doi: 10.1007/s00439-006-0244-8. [DOI] [PubMed] [Google Scholar]
- 259.Haeffel GJ, Getchell M, Koposov RA, Yrigollen CM, Deyoung CG, et al. Association between polymorphisms in the dopamine transporter gene and depression: evidence for a gene-environment interaction in a sample of juvenile detainees. Psychol Sci. 2008;19(1):62–69. doi: 10.1111/j.1467-9280.2008.02047.x. [DOI] [PubMed] [Google Scholar]
- 260.Mehrabian A, Blum JS. Temperament and personality as functions of age. Int J Aging Hum Dev. 1996;42(4):251–269. doi: 10.2190/1QCY-VETT-JJAY-3EY1. [DOI] [PubMed] [Google Scholar]
- 261.Martinez D, Orlowska D, Narendran R, Slifstein M, Liu F, et al. Dopamine type 2/3 receptor availability in the striatum and social status in human volunteers. Biol Psychiatry. 2010;67(3):275–278. doi: 10.1016/j.biopsych.2009.07.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Sapolsky RM. The influence of social hierarchy on primate health. Science. 2005;308(5722):648–652. doi: 10.1126/science.1106477. [DOI] [PubMed] [Google Scholar]
- 263.Moors A, De Houwer J. Automatic processing of dominance and submissiveness. Exp Psychol. 2005;52(4):296–302. doi: 10.1027/1618-3169.52.4.296. [DOI] [PubMed] [Google Scholar]
- 264.Zink CF, Tong Y, Chen Q, Bassett DS, Stein JL, et al. Know your place: neural processing of social hierarchy in humans. Neuron. 2008;58(2):273–283. doi: 10.1016/j.neuron.2008.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Fowler JH, Christakis NA. Dynamic spread of happiness in a large social network: longitudinal analysis over 20 years in the Framingham Heart Study. BMJ. 2008;337:a2338. doi: 10.1136/bmj.a2338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Chen TJ, Blum K, Chen AL, Bowirrat A, Downs WB, et al. Neurogenetics and clinical evidence for the putative activation of the brain reward circuitry by a neuroadaptagen: proposing an addiction candidate gene panel map. J Psychoactive Drugs. 2011;43(2):108–127. doi: 10.1080/02791072.2011.587393. [DOI] [PubMed] [Google Scholar]
- 267.Wang GJ, Volkow ND, Logan J, Pappas NR, Wong CT, et al. Brain dopamine and obesity. Lancet. 2001;357(9253):354–357. doi: 10.1016/s0140-6736(00)03643-6. [DOI] [PubMed] [Google Scholar]
- 268.van der Zwaluw CS, Engels RC, Vermulst AA, Franke B, Buitelaar J, et al. Interaction between dopamine D2 receptor genotype and parental rule-setting in adolescent alcohol use: evidence for a gene-parenting interaction. Mol Psychiatry. 2010;15(7):727–735. doi: 10.1038/mp.2009.4. [DOI] [PubMed] [Google Scholar]
- 269.Carioppo JT, Patrick W. Loneliness: Human Nature and the Need for Social Connection. New York, London: W. W. Norton; 2008. [Google Scholar]
- 270.Weiss RD, O’malley SS, Hosking JD, Locastro JS, Swift R COMBINE Study Research Group. Do patients with alcohol dependence respond to placebo? Results from the COMBINE Study. J Stud Alcohol Drugs. 2008;69(6):878–884. doi: 10.15288/jsad.2008.69.878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Røysamb E, Tambs K, Reichborn-Kjennerud T, Neale MC, Harris JR. Happiness and health: environmental and genetic contributions to the relationship between subjective well-being, perceived health, and somatic illness. J Pers Soc Psychol. 2003;85(6):1136–1146. doi: 10.1037/0022-3514.85.6.1136. [DOI] [PubMed] [Google Scholar]
- 272.Lou HC, Nowak M, Kjaer TW. The mental self. Prog Brain Res. 2005;150:197–204. doi: 10.1016/S0079-6123(05)50014-1. [DOI] [PubMed] [Google Scholar]
- 273.Miller JK. A Hunger for Healing: The Twelve Steps as a Classic Model for Christian Spiritual Growth. New York: HarperCollins; 1992. [Google Scholar]
- 274.Boundy VA, Lu L, Molinoff PB. Differential coupling of rat D2 dopamine receptor isoforms expressed in Spodoptera frugiperda insect cells. J Pharmacol Exp Ther. 1996;276(2):784–794. [PubMed] [Google Scholar]
- 275.Comings DE, Ferry L, Bradshaw-Robinson S, Burchette R, Chiu C, et al. The dopamine D2 receptor (DRD2) gene: a genetic risk factor in smoking. Pharmacogenetics. 1996;6(1):73–79. doi: 10.1097/00008571-199602000-00006. [DOI] [PubMed] [Google Scholar]
- 276.Esposito-Smythers C, Spirito A, Rizzo C, McGeary JE, Knopik VS. Associations of the DRD2 TaqIA polymorphism with impulsivity and substance use: preliminary results from a clinical sample of adolescents. Pharmacol Biochem Behav. 2009;93(3):306–312. doi: 10.1016/j.pbb.2009.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Bowirrat A, Chen TJ, Oscar-Berman M, Madigan M, Chen AL, et al. Neuropsychopharmacology and neurogenetic aspects of executive functioning: should reward gene polymorphisms constitute a diagnostic tool to identify individuals at risk for impaired judgment? Mol Neurobiol. 2012;45(2):298–313. doi: 10.1007/s12035-012-8247-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278.Kjaer TW, Bertelsen C, Piccini P, Brooks D, Alving J, et al. Increased dopamine tone during meditation-induced change of consciousness. Brain Res Cogn Brain Res. 2002;13(2):255–259. doi: 10.1016/s0926-6410(01)00106-9. [DOI] [PubMed] [Google Scholar]
- 279.de Boer A, van Buel EM, Ter Horst GJ. Love is more than just a kiss: a neurobiological perspective on love and affection. Neuroscience. 2012;201:114–124. doi: 10.1016/j.neuroscience.2011.11.017. [DOI] [PubMed] [Google Scholar]
- 280.Lane A, Luminet O, Rimé B, Gross JJ, de Timary P, et al. Oxytocin increases willingness to socially share one’s emotions. Int J Psychol. 2013;48(4):676–681. doi: 10.1080/00207594.2012.677540. [DOI] [PubMed] [Google Scholar]
- 281.Gillath O, Shaver PR, Baek JM, Chun DS. Genetic correlates of adult attachment style. Pers Soc Psychol Bull. 2008;34(10):1396–1405. doi: 10.1177/0146167208321484. [DOI] [PubMed] [Google Scholar]
- 282.Insel TR, Winslow JT, Wang Z, Young LJ. Oxytocin, vasopressin, and the neuroendocrine basis of pair bond formation. Adv Exp Med Biol. 1998;449:215–224. doi: 10.1007/978-1-4615-4871-3_28. [DOI] [PubMed] [Google Scholar]
- 283.Silva SM, Madeira MD, Ruela C, Paula-Barbosa MM. Prolonged alcohol intake leads to irreversible loss of vasopressin and oxytocin neurons in the paraventricular nucleus of the hypothalamus. Brain Res. 2002;925(1):76–88. doi: 10.1016/s0006-8993(01)03261-9. [DOI] [PubMed] [Google Scholar]
- 284.Ortiz-Miranda S, Dayanithi G, Custer E, Treistman SN, Lemos JR. Micro-opioid receptor preferentially inhibits oxytocin release from neurohypophysial terminals by blocking R-type Ca2+ channels. J Neuroendocrinol. 2005;17(9):583–590. doi: 10.1111/j.1365-2826.2005.01346.x. [DOI] [PubMed] [Google Scholar]
- 285.Shapira NA, Ferguson MA, Frost-Pineda K, Gold MS. Gambling and Problem Gambling Prevalence Among Adolescents in Florida. A 100 page Report to the Florida Council on Compulsive Gambling, Inc; 2002. [Google Scholar]
- 286.Scherrer JF, Xian H, Shah KR, Volberg R, Slutske W, et al. Effect of genes, environment, and lifetime co-occurring disorders on health-related quality of life in problem and pathological gamblers. Arch Gen Psychiatry. 2005;62(6):677–683. doi: 10.1001/archpsyc.62.6.677. [DOI] [PubMed] [Google Scholar]
- 287.Silveira LA. Experimenting with spirituality: analyzing The God Gene in a nonmajors laboratory course. CBE Life Sci Educ. 2008;7(1):132–145. doi: 10.1187/cbe.07-05-0029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Beck M. Illness, disease and sin: the connection between genetics and spirituality. Christ Bioeth. 2007;13(1):67–89. doi: 10.1080/13803600701283052. [DOI] [PubMed] [Google Scholar]
- 289.Hamer D. The God Gene: How Faith is Hardwired into Our Genes. New York: Doubleday, USA; 2004. [Google Scholar]