

Abstract
The retrovirus integrase (IN) is responsible for integration of the reverse transcribed linear cDNA into the host DNA genome. First, IN cleaves a dinucleotide from the 3’ OH blunt ends of the viral DNA exposing the highly conserved CA sequence in the recessed ends. IN utilizes the 3’ OH ends to catalyze the concerted integration of the two ends into opposite strands of the cellular DNA producing 4 to 6 bp staggered insertions, depending on the retrovirus species. The staggered ends are repaired by host cell machinery that results in a permanent copy of the viral DNA in the cellular genome. Besides integration, IN performs other functions in the replication cycle of several studied retroviruses. The proper organization of IN within the viral internal core is essential for the correct maturation of the virus. IN plays a major role in reverse transcription by interacting directly with the reverse transcriptase and by binding to the viral capsid protein and a cellular protein. Recruitment of several other host proteins into the viral particle are also promoted by IN. IN assists with the nuclear transport of the preintegration complex across the nuclear membrane. With several retroviruses, IN specifically interacts with different host protein factors that guide the preintegration complex to preferentially integrate the viral genome into specific regions of the host chromosomal target. Human gene therapy using retrovirus vectors is directly affected by the interactions of IN with these host factors. Inhibitors directed against the human immunodeficiency virus (HIV) IN bind within the active site of IN containing viral DNA ends thus preventing integration and subsequent HIV/AIDS.
Keywords: Retrovirus integrase, Integration, Host factors, Atomic structure, Human immunodeficiency virus integrase inhibitors
Core tip: This review examines the multifunctional properties of retrovirus integrase (IN) besides its key function of integrating the viral DNA into host chromosomes. IN has a major role in the maturation of the virus, reverse transcription and nuclear transport of the preintegration complex. IN binds to cellular cofactors for uncoating of the core and to other cellular proteins that guide the preintegration complex to preferred regions on the host genome for integration. Understanding these IN functions has resulted in the production of clinical IN strand transfer inhibitors to prevent human immunodeficiency virus (HIV/AIDS) and development of retrovirus vectors for human gene therapy.
INTRODUCTION
Retroviruses are ubiquitous in nature and are found in yeast to insects and to the animal and plant kingdoms. In retroviruses, the single stranded viral RNA genome is reverse transcribed by the viral reverse transcriptase into a double strand DNA copy that is subsequently integrated into the host genome. The integration process that inserts the viral DNA into the host DNA is mediated by the viral encoded integrase (IN). Retroviruses are mobile elements that distinguish themselves from other viruses because their DNA genomes become an integral part of the host genome, an essential requirement in their replication cycle. There are slight variations in this integration process in the above mentioned biological systems, some of which are called Ty1/copia and Ty3/gypsy elements or LTR retrotransposons. This review will focus on IN found in alpharetroviruses, gammaretroviruses, lentiviruses which includes human immunodeficiency virus (HIV), and spumaviruses. The purpose of the review will be to briefly highlight the most current salient features of the many functional facets of the retrovirus IN and identify critical papers and reviews that provide in-depth information for a specific subject.
HIV is the most studied retrovirus with regards to its biology, biochemistry and pathogenesis. A general retrovirus replication cycle is depicted in Figure 1 along with location of steps for FDA approved HIV drug therapies. Each step from the binding of the virus particle to a cellular receptor to the release of a new virus particle is described in the legend. In Figure 1, IN plays a role in step 3: uncoating of the internalized viral core, step 4: reverse transcription of the viral RNA, step 5: nuclear transport of the preintegration complex (PIC), step 6: integration of viral DNA into the host genome, and step 13: maturation.
Figure 1.
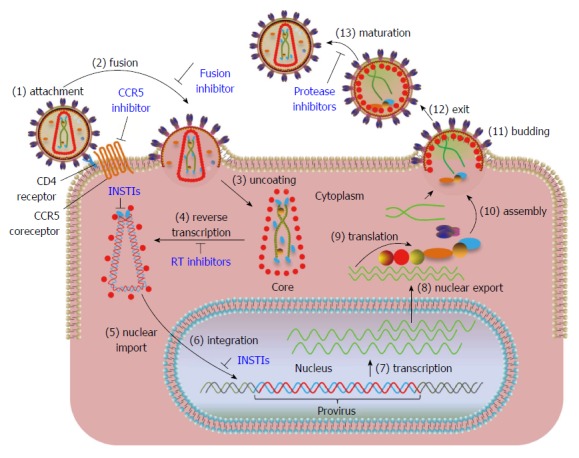
Overview of human immunodeficiency virus type 1 replication cycle. Steps in the viral replication cycle are numbered. The steps where FDA approved drugs exist are marked in blue. (1) The trimeric envelope glycoprotein (gp120/gp41) of human immunodeficiency virus (HIV) binds to the CD4 receptor and CCR5 co-receptors present on host cell surface. Maraviroc, a CCR5 receptor antagonist blocks the binding of viral gp120 to CCR5 and prevents the entry of virus into cells; (2) Viral membrane fuses with cellular membrane allowing the entry of the virus into the cytoplasm. Enfuvirtide, a fusion inhibitor binds to gp41 and prevents the formation of an entry pore for the capsid of the virus, thus blocking the entry of virus into the cytoplasm; (3) The virus undergoes uncoating of the core; (4) HIV RNA is reverse-transcribed by reverse transcriptase into double stranded DNA within the core. Reverse-transcriptase inhibitors block the synthesis of viral DNA; (5) Integrase (IN) cleaves two nucleotides adjacent to conserved CA nucleotides from both 3’-ends of viral DNA (referred as 3’-processing in text) and forms the preintegration complex (PIC). The PIC is transported into the nucleus through a cellular nuclear import pathway. IN strand transfer inhibitors (INSTIs) bind to the IN-DNA complex after 3’-processing rendering the cytoplasmic PIC defective for integration but not nuclear transport; (6) IN facilitates the joining of viral DNA to the host DNA (termed strand transfer in text). Integrated viral DNA is referred to as “provirus”. A cellular protein, lens-epithelium derived growth factor (LEDGF/p75), preferentially targets the integration of viral DNA within transcriptionally active regions. INSTIs block the joining of processed viral DNA ends into cellular DNA; (7) The viral DNA is transcribed by cellular RNA pol II into viral RNAs; (8) Full-length and alternatively spliced RNAs are transported out of nucleus; (9) Viral proteins are synthesized from the RNA template; (10) Full-length viral RNA and newly synthesized proteins begin the assembly of new virus particles at the inner surface of cell membrane; (11) Immature viral particles bud out of the membrane and released (12); and (13) Proteolytic processing of viral proteins results in the maturation of viral particle. Protease inhibitors block the formation of mature proteins by inhibiting the proteolytic processing of the viral polyproteins.
IN DOMAIN ORGRANIZATION
The retrovirus IN is located at the C-terminal end of the Pol protein that is part of a larger precursor polyprotein (Gag-Pol) encapsulated within the virus particle. The retrovirus IN was first identified and purified from an alpharetrovirus[1] and genetically shown to be necessary for integration[2-5]. The avian retrovirus or Rous sarcoma virus (RSV) and HIV IN proteins are 286 and 288 residues in length, respectively, while the prototype foamy virus (PFV) IN is 392 residues[6] (Figure 2). RSV IN possesses three structural domains comprising an N-terminal domain (NTD) (residues 1-44) that contains a zinc-binding HHCC motif, a catalytic core domain (CCD) (residues 58-213) and a C-terminal domain (CTD) (residues 222-268). HIV IN has similar size domains and linkers as RSV IN. Both PFV and murine leukemia virus (MLV) IN (408 residues) possess an additional N-terminal extension domain (NED) (approximately 50 residues) attached adjacent to the NTD of IN.
Figure 2.
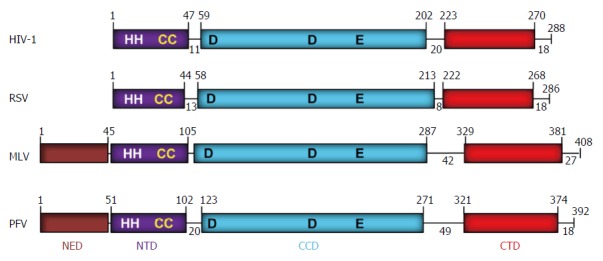
Domain organization of retroviral integrases. The domains (NED: N-terminal extension domain; NTD: N-terminal domain; CCD: Catalytic core domain; and CTD: C-terminal domain) are depicted with different colors. The number of residues in each domain are indicated. The domains are separated by protein linkers of various sizes. NTD contains the zinc binding HH-CC motif and the D-D-35-E motif in CCD binds Mg2+ which constitutes the active site. The CTD shows a topology of SH3-like domain, a characteristic feature associated with protein-protein and protein-DNA interactions. The exact MLV domain sizes have not been determined experimentally except for the CTD[99]. A short tail of disordered amino acids is located on the C-terminal end of each IN. IN: Integrase; HIV: Human immunodeficiency virus; RSV: Rous sarcoma virus; MLV: Murine leukemia; PFV: Prototype foamy virus.
BIOLOGICAL FUNCTIONS OF IN
Role of IN in viral maturation
The retrovirus structural Gag proteins are approximately 80% of the total mass of the viral proteins. The reverse transcriptase (RT) and IN are derived from a separate Gag-Pol precursor polyprotein produced by translational frameshifts. This precursor polyprotein accounts for approximately 2% to 3% of the total viral protein mass (Figure 3). Despite this lower level of IN in virus particles, studies using site-directed mutagenesis of HIV IN highly suggest that deletion and missense mutations affect HIV assembly and release[7]. The mechanisms associated with these defects are not well understood but may include activation of the viral protease located within Gag-Pol and dimerization of this precursor polyprotein.
Figure 3.
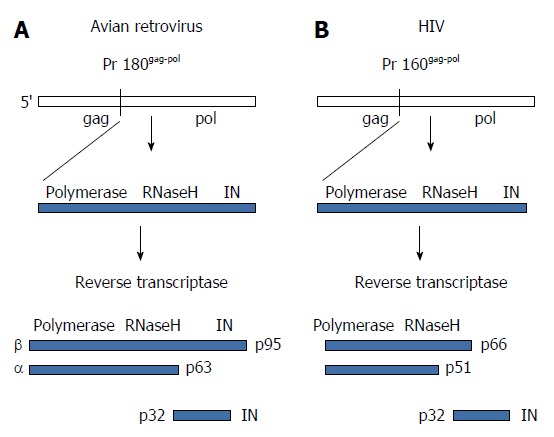
Structural organization and proteolytic processing of reverse transcriptase and integrase. A: The precursor Gag-Pol protein is processed by the viral protease into an intermediate polyprotein. The avian polyprotein exists as a dimer of the β subunit that is processed into the active αβ subunits (RT) and integrase (IN). The α subunit contains the active polymerase and RNaseH sites while β retains IN residues. RT does not possess integration capabilities; B: Cleavage of the human immunodeficiency virus (HIV) polyprotein produces the active dimeric RT and IN. The HIV p66 subunit possesses the polymerase and RNaseH active sites. RT: Reverse transcriptase.
The recent discovery of inhibitors that do not interact within the catalytic active site of HIV IN has provided unforeseen information on a structural role for IN in virus particle maturation. These inhibitors called LEDGINS[8] or ALLINIS (allosteric IN inhibitors)[9] prevent interactions of IN with a cellular chromatin binding protein lens epithelium-derived growth factor (LEDGF/p75), a transcriptional cofactor that guides the HIV PIC (Figure 1) to active transcription sites for integration (see host factors). In addition to inhibiting integration, significant structural defects are caused by these inhibitors resulting in abnormal formation of electron dense HIV virus cores thus producing non-infectious particles upon egress[10-12] (Figure 1). Other studies have shown that these inhibitors significantly induce HIV IN monomers to produce inactive oligomers in vitro[10,13,14]. These results highly suggest that IN plays a critical role in virus maturation (Figure 1).
Reverse transcription requires IN functions
RT and IN are linked together covalently at the C-terminal end of Gag-Pol precursor polyprotein (Figure 3). The processing of the precursor protein to RT and IN by the viral protease in the virus particle is different between RSV and HIV. For RSV, the Pol product from the polyprotein is a 95 kDa polypeptide termed β that forms dimers (Figure 3). The viral protease cleaves the dimers into active RT, termed αβ, which causes the release and activation of the 32 kDa IN. The αβ polypeptide still contains a single copy of IN residues that facilitates a processive mechanism for reverse transcription and its associated RNaseH activity. In contrast, HIV IN is released entirely from the polyprotein producing RT (p66/p51) (Figure 3), similarly to MLV IN (not shown).
Site-specific mutations introduced into the cleaved HIV IN result in reverse transcription defects but not integration[15]. The exact mechanisms associated with these RT defects are mostly unknown. HIV IN (Figure 3) appears to have a physical interaction with RT (p66/p51) that is necessary for normal viral DNA synthesis[16-19]. In normal HIV infected cells, IN interacts with the major structural viral capsid protein (CA) whose interactions affect the normal uncoating process of the core and thus reverse transcription[20]. Following the above theme that viral core uncoating affects reverse transcription, recent results suggest that the cellular protein DYNLL1 is required for early viral DNA synthesis and uncoating via a physical interaction of DYNLL1 with HIV IN[21]. As these authors suggested, IN-DYNLL1 interactions may be a potential target for future inhibitors.
Role of IN in the preintegration complex, its nuclear transport and in integration
Although IN has many roles in the retrovirus replication cycle, IN is the catalytic component necessary for concerted integration of the viral DNA ends into the host genome (Figures 4 and 5). A pivotal study clearly demonstrated that linear MLV DNA acts as the substrate for integration in vivo[22]. The high molecular weight PIC containing a copy of the linear viral DNA was isolated from MLV infected cells (Figures 1 and 4). This complex is capable of integrating the viral ends in a concerted fashion into a target substrate in vitro. DNA sequence analysis of the integrated viral DNA demonstrated that IN inserted the 3’ OH ends into opposite strands of the target DNA producing the 4 bp host site duplication found associated with the integrated MLV provirus in vivo or a 6 bp duplication with RSV (Figure 5). The host site duplication for HIV is 5 bp.
Figure 4.
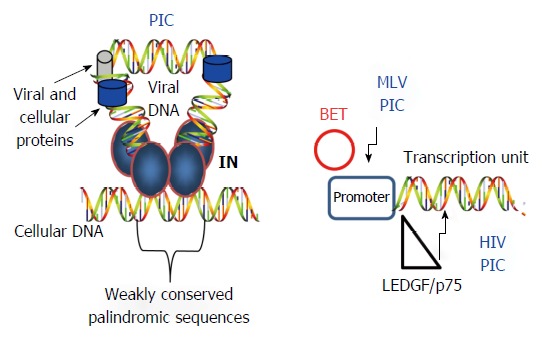
Selection of cellular DNA target sites for integration. Left: IN within the PIC is guided to host sites by several mechanisms including recognition of weak palindromic sequences by IN, nuclear topography of the cellular host DNA site, chromatin structure, and several kinds of associated histone tails. Only IN (labeled), RT (blue circle) and LEDGF/p75 (gray cylinder) are identified in the figure; Right: Cell proteins (BET and LEDGF/p75) influence the PIC to preferentially insert the viral DNA into specific regions of the host genome. IN: Integrase; PIC: Preintegration complex; HIV: Human immunodeficiency virus; MLV: Murine leukemia virus.
Figure 5.
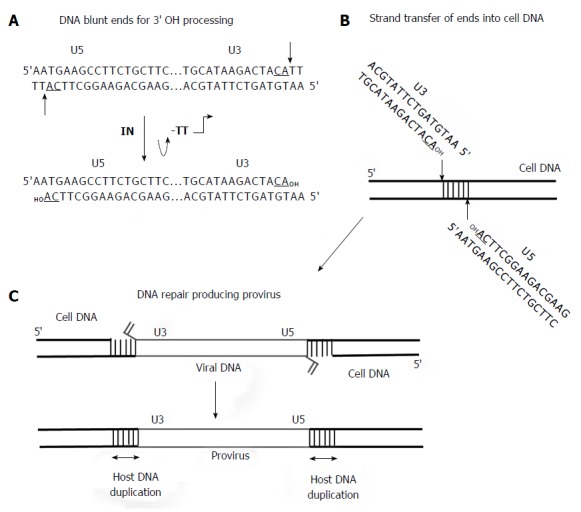
Requirement for 3’ OH processing and strand transfer activities to facilitate concerted integration of viral DNA by integrase into target DNA. A: The linear approximately 10 kb DNA with only the ends is shown. The U3 and U5 RSV DNA blunt ends with the underlined conserved CA dinucleotide on the catalytic strand are indicated. The thin arrows indicate 3’ OH processing of two nucleotides by IN producing the recessed substrate for strand transfer activity; B: Concerted integration of the viral linear DNA ends by integrase (IN) into the target DNA is by a transesterification mechanism; C: The insertion of viral DNA into the target produces a 6 bp stagger cut by RSV IN that is repaired by cellular proteins producing the integrated provirus. RSV: Rous sarcoma virus.
There are numerous publications (many not indicated here) describing the integration capabilities of the PIC for MLV[22,23] and later for HIV[24-27] at the biochemical, genetic and inhibition of strand transfer by IN inhibitors (Figure 1). The PIC is composed of linear DNA, viral proteins IN, RT, CA, matrix (MA), viral protein R (Vpr) and several cellular associated proteins (Figure 4). In the cytoplasm, IN cleaves a dinucleotide from the 3’ OH ends of blunt DNA before transport of the PIC into the nucleus (Figure 5). HIV IN also appears to play a role in the nuclear transport of the PIC along with other viral and cellular proteins[28,29]. After transport, IN inserts the 3’ OH recessed DNA ends into the cellular DNA genome. As described later, the HIV IN strand transfer inhibitors (STIs) interact within the active site of IN and the 3’ processed LTR ends in the cytoplasmic PIC, thus inactivating the complex preventing integration into cellular DNA[27]. The retrovirus PIC has provided major insights into retrovirus integration but has a severe limitation for providing sufficient quantities necessary for biochemical studies.
IN interactions with host factors and cellular target site selection
IN is the dominant factor for selection of weakly conserved palindromic sequences at the target site in vivo[30-32] (Figure 4). Whole genomic sequencing of DNA target sites demonstrates that HIV IN prefers to integrate its viral DNA into active transcriptional units[30] while MLV IN favors integration sites near the promoter region of genes and not into transcriptional units[33]. Further studies demonstrated that RSV IN essentially integrates its DNA into the host genome at random sites[34,35]. These results suggested that HIV and MLV IN interact with specific host factors that direct IN to preferred sites in the host genome.
Major studies have centered on the ability of LEDGF/p75, a transcriptional co-activator, to guide the HIV PIC to integrate its DNA into active transcriptional sites[36-41] (Figure 4). Besides LEDGF/p75, cellular Nup153 is indispensable for the integration of HIV DNA into the peripheral edge of the nuclear DNA suggesting that nuclear topography is also an important for HIV integration site selection[42]. For MLV IN, recent studies demonstrated that IN interacts with the chromatin BET (bromo- and extra-terminal domain) proteins (BRD2, BRD3 and BRD4) directing it to the promoter region of genes[39,43-45] (Figure 4). To date, there are no reports that have identified cellular proteins involved in host site selection by avian retroviruses. In summary, IN is capable of integrating its viral DNA into numerous segments of the host genome but is preferentially directed to specific regions by host factors.
BIOCHEMICAL AND STRUCTURAL PROPERTIES OF IN
Solution properties of IN
Biochemical and biophysical studies of retrovirus IN has been extensive over the last 37 years. Most studies have used recombinant RSV, HIV, MLV and PFV IN. Only the avian retrovirus IN has been purified from virus particles[1,46]. As stated above, IN cleaves a dinucleotide from both 3’ OH blunt ends of the viral DNA and subsequently inserts the recessed ends into a target DNA by a transesterification reaction (Figure 5). The biochemistry of these reactions and other structural properties of IN were recently reviewed[47].
The subunit structure of IN is varied and highly dependent on the purification process and different salt requirements to maintain solubility for each expressed recombinant IN. PFV IN is soluble (10-15 mg/mL) in 0.2 mol/L NaCl[48,49] while RSV IN is highly soluble (30 mg/mL) in 0.2 mol/L (NH4)2SO4[50]. PFV IN is monomeric[49] while RSV IN is a dimer in solution[50-52]. Recombinant HIV IN has been purified as a monomer, dimer and tetramer[47,53,54]. Recent studies suggest that the active form of HIV IN is a monomer that is responsible for assembly of the synaptic complex (SC) or intasome capable of efficient concerted integration of two oligonucleotides into a target substrate in vitro[53]. The HIV IN monomer purified in the presence of EDTA is converted into a tetramer in the presence of Zn2+ without the formation of a dimer intermediate[53,54]. Studies have demonstrated that a tetramer of IN is associated with HIV, PFV and RSV SC capable of concerted integration[50,55-57].
The oligomeric state of retrovirus IN play key roles in virus maturation upon release of the virus particle, enzymatic activities for 3’ OH processing and concerted integration of viral DNA and possibly for reverse transcription in the cytoplasmic PIC.
Retroviral IN superfamily and PFV intasome structure
The retrovirus IN superfamily comprises proteins that are involved in a variety of activities like replication and repair of DNA, V(D)J recombination, and DNA transposition. The DDE domain of retrovirus IN (Figure 2) is highly conserved in proteins capable of similar catalytic reactions including MuA transposase, human RAG1 recombinase, the RNaseH domain on the retrovirus RT (Figure 3) and other bacterial and human endonucleases[58,59]. Structures of many of the above protein have been resolved at the atomic level in complex with DNA. However, among the retrovirus INs only the four-domain PFV intasome (PFV IN-DNA complex) has been resolved (Figure 6).
Figure 6.
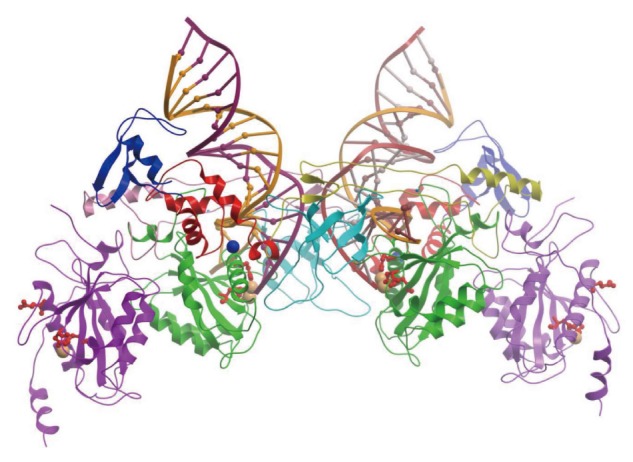
Prototype foamy virus intasome structure. The intasome consist of an integrase (IN) tetramer with two viral DNA ends. The 17/19 bp DNA interacts extensively with the two inner monomers of IN. Among the inner monomers, the N-terminal extension domain, N-terminal domain, catalytic core domain (CCD) and C-terminal domains are shown in blue, red, green and cyan, respectively. The DDE residues are identified as red amino acids. Among the outer protomers, only the CCD (shown in purple) was resolved. The cartoon was redrawn from the PDB ID 3OY9 in Molsoft ICM-Browser. The Zn2+ and Mg2+ metal ions are shown with blue and orange spheres, respectively.
The crystallographic study of the full-length PFV IN with bound viral DNA revealed, for the first time, the architecture of retrovirus intasome assembly and atomic details of the IN-DNA interactions[57,60] (Figure 6). The PFV intasome contains a tetramer of IN, which consists of a dimer of the “inner” subunits that make extensive protein-protein and protein-DNA interactions, and a pair of the “outer” subunits that appear to play structural roles. All four domains of the IN subunits and the inter-domain linkers interact extensively with DNA to bridge between the two viral DNA ends and capture the target DNA. The engagement of the viral DNA sequence and catalysis in trans is a feature shared with related transposase systems[59,61].
The determination of the atomic structure of the PFV intasome greatly enhanced our understanding of IN interactions with 3’ OH recessed ends in the active site and showing how a tetramer of IN catalyzes the concerted integration reaction[57]. Other structural studies have shown how PFV IN is bound to the viral DNA in its ground state prior to 3’ OH processing or strand transfer[62]. Lastly, the PFV intasome in complex with target DNA after insertion of the 3’ OH ends (strand transfer complex) demonstrated how severely the target DNA is bent upon insertion of the viral DNA ends[60,63].
HIV IN INHIBITORS
Strand transfer inhibitors and their interactions within the active site of HIV IN
The requirement for integration of the viral DNA into the host genome for replication renders IN as a natural target for development of inhibitors. Numerous studies in the 1990s to identify relevant HIV IN inhibitors culminated with a study demonstrating that IN STI prevent the integration of the viral DNA into the cell genome resulting in inhibition of HIV replication in cells[27]. Further studies established that the STI competed with target DNA binding to IN suggesting a unique conformation of IN for the strand transfer reaction[64]. The diketo acid pharmacophore of the STI also bound to the divalent metal ion in the active site of IN[65].
Crystallization studies of the four-domain PFV IN intasome without (Figure 6) and with STIs demonstrated the interactions of these inhibitors within the active site of IN[57,62,66,67] (see Figures 7 and 8 for brief description of STIs and their mechanism). There are currently three clinical STIs approved by the FDA for treatment of HIV/AIDS (Figure 7). They are Raltegravir (RAL), Elvitegravir (EVG) and Dolutegravir (DTG). The HIV IN STIs are interfacial inhibitors whose mechanisms are similar to other compounds that target bacterial DNA gyrase and topoisomerase IV as well as anticancer drugs targeting human topoisomerases[68,69]. Interfacial inhibitors, like the HIV STIs, bind to a site produced at the interface of two (or more) molecules bound together resulting in a functional complex. In the case of STIs, the interfaces are formed by the viral terminal CA nucleotides and specific IN residues in the PFV intasome[57,67] (Figure 8). A recent book[70] and several reviews of HIV IN inhibitors provide an extensive overview of the discovery of STIs and their mechanisms[47,71-73].
Figure 7.
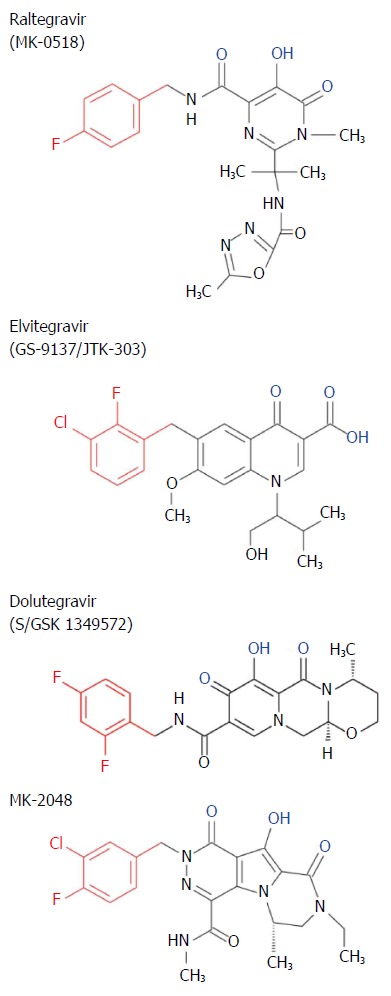
Chemical structure of strand transfer inhibitors. Conserved metal chelating moiety similar to diketo acid is shown in blue and a conserved halogenated benzyl group that interacts with the penultimate nucleotide of DNA is shown in red.
Figure 8.
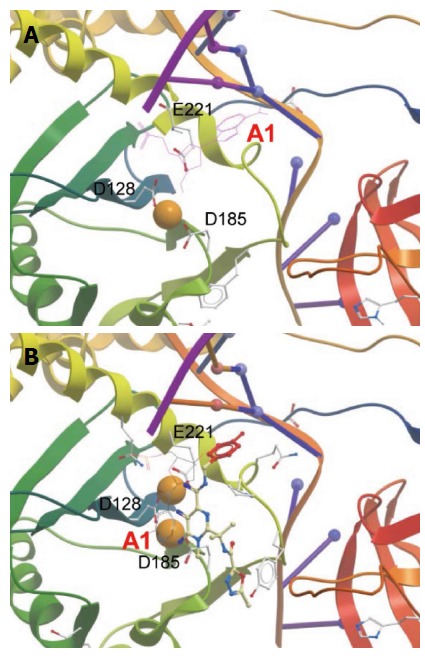
Active site within prototype foamy virus intasome in apo (A) and Raltegravir-bound state (B). Orange spheres indicate Mg2+. Active sites residues are labeled. Only the terminal 3’ adenosine (marked A1) is displayed in its chemical form to show the displacement upon RAL binding. The diketo acid group of Raltegravir (RAL) interacts with the metal ions. Adenine is π-stacked against the RAL metal-chelating scaffold. The halobenzoyl group of RAL is marked red. The figures were constructed from PDB entries 3L2R and 3OYA, respectively, in Molsoft ICM-Browser.
HIV and RSV synaptic complexes are kinetically stabilized by STIs
The atomic structures of the three-domain HIV and RSV IN (Figure 2) in complex with viral DNA are unknown. Crystallization of these INs in complex with their viral DNA substrates at high IN concentrations has not been successful for numerous biochemical reasons including aggregation of HIV IN and limited stability of the complexes.
The HIV SC at low nM concentrations can be physically trapped in the presence of HIV IN STIs as analyzed by native agarose gel electrophoresis[74,75]. There are strong structural similarities in the CCD between HIV, PFV and RSV IN (Figure 2)[47,51,52,57,67,76,77] suggesting that under appropriate conditions the highly soluble RSV IN could also be trapped in the presence of the appropriate oligonucleotide (ODN) substrate and STIs. Assembly of a trapped RSV SC at high IN concentrations (1.5 mg/mL; 45 μmol/L) in solution required the presence of 3’ OH recessed viral ODN ranging in sizes (16/18R to 18/20R) and DTG, RAL or MK-2048[50] (Figure 9). DTG and MK-2048 (an investigational inhibitor) possess a significantly longer dissociative half-life than RAL within an HIV IN-DNA complex allowing these STIs to capture and stabilize the IN-DNA complex[78,79]. Without the STI presence, the transient RSV SC produced in the concerted integration pathway could not be isolated highly suggesting the STI-trapped SC is kinetically stabilized. Efforts are ongoing to resolve the atomic structure of this kinetically stabilized STI-trapped SC and the RSV strand transfer complex.
Figure 9.
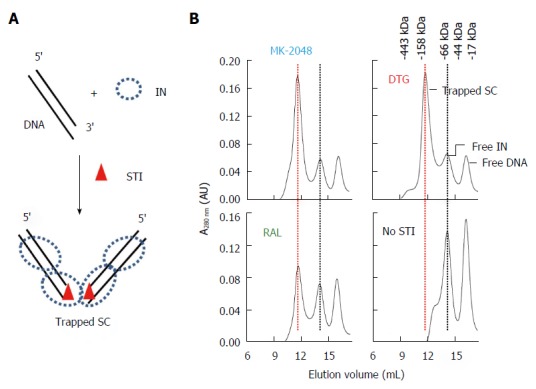
Identification of strand transfer inhibitor-trapped Rous sarcoma virus synaptic complex by size-exclusion chromatography. A: Schematic of Rous sarcoma virus (RSV) IN, 3’ OH recessed ODN, and strand transfer inhibitor (STI) used to assemble STI-trapped synaptic complex (SC); B: MK-2048 and Dolutegravir (DTG) efficiently produce trapped SC while Raltegravir had moderate efficiency. No trapped SC is produced in the absence of STI. Elution positions of STI-trapped SC (red line), free IN and DNA are marked. Molecular weight markers are indicated.
Clinical results using HIV IN STIs and drug resistance
The previously described three clinical STIs (Figure 7) are the newest class of inhibitors to treat HIV/AIDS that must be used in combinational drug therapy with reverse transcriptase, protease and entry inhibitors. RAL and EVG were FDA approved for clinical use in 2007 and 2012, respectively, while DTG was approved in 2013. IN STIs are superior to other regimens in first-line therapies[80,81]. All three IN STIs have a high safety record and display strong efficacy[81].
HIV has a high mutational rate, approximately 3 × 10-5 mutations/base pair per cycle[82]. Thus, like with RT and protease inhibitors, the use of IN STIs has resulted in the emergence of drug-resistant viral strains that causes a loss of drug effectiveness. Both RAL and EVG possess a low genetic barrier giving rise to drug-resistant IN mutants (G140S/Q148H, N155S, Q148K, F121Y, T66I/S153Y) in tissue culture as well as in treated patients[72,83]. In contrast, DTG has a high genetic barrier and to date significant resistant IN mutants have been nearly absent in treatment-naïve patients[81,84,85]. The higher genetic barrier of DTG in patients may be due to its longer dissociative half-life from the HIV IN-DNA complex suggesting that DTG extended binding may be a significant factor for prevention of drug-resistance[86]; also shown to be a factor in producing kinetically stabilized RSV SC (Figure 9). HIV carrying the rare signature IN R263K mutation (others are H51Y and E138K) observed with DTG apparently cannot co-exist in combination with many of the above classical IN resistance mutations[87]. HIV is apparently unable to compensate for these mutations which induce a fitness cost that prevents the virus from evading inhibitor pressure.
Taken these demonstrated properties of DTG, investigators have slightly modified its structure to formulate a long-acting injectable nanoparticle (200 nm) suitable for clinical administration on a quarterly basis[88]. In these preclinical studies, GSK744 provided high-level protection against repeated simian/human immunodeficiency virus challenge in rhesus macaques. This study suggests that GSK744 in combination with a similar injectable RT inhibitor could decrease adherence problems associated with pre-exposure prophylaxis.
RETROVIRAL VECTORS
Human gene therapy
Retroviruses are natural vectors for transfer of genetic information between cells. For example, the insertion of an oncogene derived from one cell by a retrovirus into another cell occurs naturally[82]. Numerous commercial and investigative retroviral vectors have been developed to accomplish a wide variety of experimental protocols as well as the study of human diseases[89]. This permanent transfer of genetic information by retroviruses has led to an explosion of ideas for human gene therapies using gammaretroviral (MLV) and lentiviral (HIV) vectors.
The selection of DNA target sites by IN in cells is highly influenced by the interactions of IN with host cofactors and nuclear import properties of the PIC (Figure 4, see above). The selective property of MLV IN to preferentially integrate its viral DNA in the promoter regions of genes had placed a hold on human gene therapy using MLV vectors to treat SCID-X1 (x-linked severe combined immunodeficiency) in boys, which was successful[90-92]. Insertional mutagenesis lead to T-cell acute lymphoblastic leukemia in 5 of the 20 patients caused by transactivation of LMO2 or CCND2 proto-oncogenes[93,94]. Recently, a highly modified self-inactivating (SIN) MLV vector with deleted viral U3 enhancer sequences provided the same efficacy as the original MLV vectors and with no leukemogenesis produced after 33 mo[95]. Future observations of these patients using this modified MLV vector will determine further use of this kind of SIN vector. SIN HIV vectors have been used in human trials and have the advantage that they can effectively transfect resting cells due to their nuclear import properties[89,96]. Finally, SIN avian retrovirus vectors have been developed but have not been used in human gene therapy[97]. Consistent with the observation that avian retroviruses integrate randomly into cellular DNA[34,35], genome-wide analysis of integration sites by a SIN avian retrovirus vector in human hematopoietic stem/progenitor cells demonstrated that integration is random and not near hotspots containing proto-oncogenes like LMO2[98].
CONCLUSION
This review has outlined the multiple functions that the retrovirus IN has in the life cycle of retroviruses besides the key function of DNA integration. The development of the three clinical STIs (Figure 7) directed against HIV IN has provided a strong premise for supporting basic scientific research[73]. The future development of advanced STIs for pre-exposure prophylaxis, the search for new IN inhibitors and the discovery other IN functions are currently underway. The potential utilization of retrovirus vectors for human gene therapy only adds to this future spectrum for scientific investigation of the retrovirus IN.
Footnotes
Supported by Partially National Institutes of Health grants from NIAID (AI100682 to Grandgenett DP) and NIGMS (GM109770 to Aihara H).
Conflict-of-interest statement: Authors declare no conflict of interests for this article.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: May 13, 2015
First decision: June 24, 2015
Article in press: July 27, 2015
P- Reviewer: Martinez-Costa OH, O’Connor TR S- Editor: Ji FF L- Editor: A E- Editor: Wang CH
References
- 1.Grandgenett DP, Vora AC, Schiff RD. A 32,000-dalton nucleic acid-binding protein from avian retrovirus cores possesses DNA endonuclease activity. Virology. 1978;89:119–132. doi: 10.1016/0042-6822(78)90046-6. [DOI] [PubMed] [Google Scholar]
- 2.Donehower LA, Varmus HE. A mutant murine leukemia virus with a single missense codon in pol is defective in a function affecting integration. Proc Natl Acad Sci USA. 1984;81:6461–6465. doi: 10.1073/pnas.81.20.6461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Panganiban AT, Temin HM. The retrovirus pol gene encodes a product required for DNA integration: identification of a retrovirus int locus. Proc Natl Acad Sci USA. 1984;81:7885–7889. doi: 10.1073/pnas.81.24.7885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Schwartzberg P, Colicelli J, Goff SP. Construction and analysis of deletion mutations in the pol gene of Moloney murine leukemia virus: a new viral function required for productive infection. Cell. 1984;37:1043–1052. doi: 10.1016/0092-8674(84)90439-2. [DOI] [PubMed] [Google Scholar]
- 5.Quinn TP, Grandgenett DP. Genetic evidence that the avian retrovirus DNA endonuclease domain of pol is necessary for viral integration. J Virol. 1988;62:2307–2312. doi: 10.1128/jvi.62.7.2307-2312.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li X, Krishnan L, Cherepanov P, Engelman A. Structural biology of retroviral DNA integration. Virology. 2011;411:194–205. doi: 10.1016/j.virol.2010.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Engelman A. Neamati N, editor. Pleiotrophic nature of HIV-1 integrase mutations, Chapter 6, in HIV-1 Integrase, Mechanism and Inhibitor Design. New Jersey: John Wiley and Sons; 2011. pp. 67–81. [Google Scholar]
- 8.Christ F, Voet A, Marchand A, Nicolet S, Desimmie BA, Marchand D, Bardiot D, Van der Veken NJ, Van Remoortel B, Strelkov SV, et al. Rational design of small-molecule inhibitors of the LEDGF/p75-integrase interaction and HIV replication. Nat Chem Biol. 2010;6:442–448. doi: 10.1038/nchembio.370. [DOI] [PubMed] [Google Scholar]
- 9.Jurado KA, Engelman A. Multimodal mechanism of action of allosteric HIV-1 integrase inhibitors. Expert Rev Mol Med. 2013;15:e14. doi: 10.1017/erm.2013.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jurado KA, Wang H, Slaughter A, Feng L, Kessl JJ, Koh Y, Wang W, Ballandras-Colas A, Patel PA, Fuchs JR, et al. Allosteric integrase inhibitor potency is determined through the inhibition of HIV-1 particle maturation. Proc Natl Acad Sci USA. 2013;110:8690–8695. doi: 10.1073/pnas.1300703110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Balakrishnan M, Yant SR, Tsai L, O’Sullivan C, Bam RA, Tsai A, Niedziela-Majka A, Stray KM, Sakowicz R, Cihlar T. Non-catalytic site HIV-1 integrase inhibitors disrupt core maturation and induce a reverse transcription block in target cells. PLoS One. 2013;8:e74163. doi: 10.1371/journal.pone.0074163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Feng L, Larue RC, Slaughter A, Kessl JJ, Kvaratskhelia M. HIV-1 integrase multimerization as a therapeutic target. Curr Top Microbiol Immunol. 2015;389:93–119. doi: 10.1007/82_2015_439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Desimmie BA, Schrijvers R, Demeulemeester J, Borrenberghs D, Weydert C, Thys W, Vets S, Van Remoortel B, Hofkens J, De Rijck J, et al. LEDGINs inhibit late stage HIV-1 replication by modulating integrase multimerization in the virions. Retrovirology. 2013;10:57. doi: 10.1186/1742-4690-10-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Feng L, Sharma A, Slaughter A, Jena N, Koh Y, Shkriabai N, Larue RC, Patel PA, Mitsuya H, Kessl JJ, et al. The A128T resistance mutation reveals aberrant protein multimerization as the primary mechanism of action of allosteric HIV-1 integrase inhibitors. J Biol Chem. 2013;288:15813–15820. doi: 10.1074/jbc.M112.443390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Engelman A. In vivo analysis of retroviral integrase structure and function. Adv Virus Res. 1999;52:411–426. doi: 10.1016/s0065-3527(08)60309-7. [DOI] [PubMed] [Google Scholar]
- 16.Zhu K, Dobard C, Chow SA. Requirement for integrase during reverse transcription of human immunodeficiency virus type 1 and the effect of cysteine mutations of integrase on its interactions with reverse transcriptase. J Virol. 2004;78:5045–5055. doi: 10.1128/JVI.78.10.5045-5055.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wu X, Liu H, Xiao H, Conway JA, Hehl E, Kalpana GV, Prasad V, Kappes JC. Human immunodeficiency virus type 1 integrase protein promotes reverse transcription through specific interactions with the nucleoprotein reverse transcription complex. J Virol. 1999;73:2126–2135. doi: 10.1128/jvi.73.3.2126-2135.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dobard CW, Briones MS, Chow SA. Molecular mechanisms by which human immunodeficiency virus type 1 integrase stimulates the early steps of reverse transcription. J Virol. 2007;81:10037–10046. doi: 10.1128/JVI.00519-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wilkinson TA, Chow S. Neamati N, editor.Functional interaction between human immunodeficiency virus type 1 reverse transcriptase and integrase, in HIV-1 Integrase, Mechanism and Inhibitor Design. New Jersey: John Wiley and Sons, Inc; 2011. pp. 95–103. [Google Scholar]
- 20.Briones MS, Dobard CW, Chow SA. Role of human immunodeficiency virus type 1 integrase in uncoating of the viral core. J Virol. 2010;84:5181–5190. doi: 10.1128/JVI.02382-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jayappa KD, Ao Z, Wang X, Mouland AJ, Shekhar S, Yang X, Yao X. Human immunodeficiency virus type 1 employs the cellular dynein light chain 1 protein for reverse transcription through interaction with its integrase protein. J Virol. 2015;89:3497–3511. doi: 10.1128/JVI.03347-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Brown PO, Bowerman B, Varmus HE, Bishop JM. Correct integration of retroviral DNA in vitro. Cell. 1987;49:347–356. doi: 10.1016/0092-8674(87)90287-x. [DOI] [PubMed] [Google Scholar]
- 23.Wei SQ, Mizuuchi K, Craigie R. Footprints on the viral DNA ends in moloney murine leukemia virus preintegration complexes reflect a specific association with integrase. Proc Natl Acad Sci USA. 1998;95:10535–10540. doi: 10.1073/pnas.95.18.10535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Farnet CM, Haseltine WA. Integration of human immunodeficiency virus type 1 DNA in vitro. Proc Natl Acad Sci USA. 1990;87:4164–4168. doi: 10.1073/pnas.87.11.4164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chen H, Wei SQ, Engelman A. Multiple integrase functions are required to form the native structure of the human immunodeficiency virus type I intasome. J Biol Chem. 1999;274:17358–17364. doi: 10.1074/jbc.274.24.17358. [DOI] [PubMed] [Google Scholar]
- 26.Chen H, Engelman A. Asymmetric processing of human immunodeficiency virus type 1 cDNA in vivo: implications for functional end coupling during the chemical steps of DNA transposition. Mol Cell Biol. 2001;21:6758–6767. doi: 10.1128/MCB.21.20.6758-6767.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hazuda DJ, Felock P, Witmer M, Wolfe A, Stillmock K, Grobler JA, Espeseth A, Gabryelski L, Schleif W, Blau C, et al. Inhibitors of strand transfer that prevent integration and inhibit HIV-1 replication in cells. Science. 2000;287:646–650. doi: 10.1126/science.287.5453.646. [DOI] [PubMed] [Google Scholar]
- 28.Ao Z, Huang G, Yao H, Xu Z, Labine M, Cochrane AW, Yao X. Interaction of human immunodeficiency virus type 1 integrase with cellular nuclear import receptor importin 7 and its impact on viral replication. J Biol Chem. 2007;282:13456–13467. doi: 10.1074/jbc.M610546200. [DOI] [PubMed] [Google Scholar]
- 29.Christ F, Thys W, De Rijck J, Gijsbers R, Albanese A, Arosio D, Emiliani S, Rain JC, Benarous R, Cereseto A, et al. Transportin-SR2 imports HIV into the nucleus. Curr Biol. 2008;18:1192–1202. doi: 10.1016/j.cub.2008.07.079. [DOI] [PubMed] [Google Scholar]
- 30.Schröder AR, Shinn P, Chen H, Berry C, Ecker JR, Bushman F. HIV-1 integration in the human genome favors active genes and local hotspots. Cell. 2002;110:521–529. doi: 10.1016/s0092-8674(02)00864-4. [DOI] [PubMed] [Google Scholar]
- 31.Wu X, Li Y, Crise B, Burgess SM, Munroe DJ. Weak palindromic consensus sequences are a common feature found at the integration target sites of many retroviruses. J Virol. 2005;79:5211–5214. doi: 10.1128/JVI.79.8.5211-5214.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Serrao E, Ballandras-Colas A, Cherepanov P, Maertens GN, Engelman AN. Key determinants of target DNA recognition by retroviral intasomes. Retrovirology. 2015;12:39. doi: 10.1186/s12977-015-0167-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wu X, Li Y, Crise B, Burgess SM. Transcription start regions in the human genome are favored targets for MLV integration. Science. 2003;300:1749–1751. doi: 10.1126/science.1083413. [DOI] [PubMed] [Google Scholar]
- 34.Mitchell RS, Beitzel BF, Schroder AR, Shinn P, Chen H, Berry CC, Ecker JR, Bushman FD. Retroviral DNA integration: ASLV, HIV, and MLV show distinct target site preferences. PLoS Biol. 2004;2:E234. doi: 10.1371/journal.pbio.0020234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Narezkina A, Taganov KD, Litwin S, Stoyanova R, Hayashi J, Seeger C, Skalka AM, Katz RA. Genome-wide analyses of avian sarcoma virus integration sites. J Virol. 2004;78:11656–11663. doi: 10.1128/JVI.78.21.11656-11663.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bushman FD. Integration site selection by lentiviruses: biology and possible control. Curr Top Microbiol Immunol. 2002;261:165–177. doi: 10.1007/978-3-642-56114-6_8. [DOI] [PubMed] [Google Scholar]
- 37.Bushman F, Lewinski M, Ciuffi A, Barr S, Leipzig J, Hannenhalli S, Hoffmann C. Genome-wide analysis of retroviral DNA integration. Nat Rev Microbiol. 2005;3:848–858. doi: 10.1038/nrmicro1263. [DOI] [PubMed] [Google Scholar]
- 38.Christ F, Debyser Z. The LEDGF/p75 integrase interaction, a novel target for anti-HIV therapy. Virology. 2013;435:102–109. doi: 10.1016/j.virol.2012.09.033. [DOI] [PubMed] [Google Scholar]
- 39.Kvaratskhelia M, Sharma A, Larue RC, Serrao E, Engelman A. Molecular mechanisms of retroviral integration site selection. Nucleic Acids Res. 2014;42:10209–10225. doi: 10.1093/nar/gku769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ciuffi A, Llano M, Poeschla E, Hoffmann C, Leipzig J, Shinn P, Ecker JR, Bushman F. A role for LEDGF/p75 in targeting HIV DNA integration. Nat Med. 2005;11:1287–1289. doi: 10.1038/nm1329. [DOI] [PubMed] [Google Scholar]
- 41.Desimmie BA, Weydert C, Schrijvers R, Vets S, Demeulemeester J, Proost P, Paron I, De Rijck J, Mast J, Bannert N, et al. HIV-1 IN/Pol recruits LEDGF/p75 into viral particles. Retrovirology. 2015;12:16. doi: 10.1186/s12977-014-0134-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Marini B, Kertesz-Farkas A, Ali H, Lucic B, Lisek K, Manganaro L, Pongor S, Luzzati R, Recchia A, Mavilio F, et al. Nuclear architecture dictates HIV-1 integration site selection. Nature. 2015;521:227–231. doi: 10.1038/nature14226. [DOI] [PubMed] [Google Scholar]
- 43.Studamire B, Goff SP. Host proteins interacting with the Moloney murine leukemia virus integrase: multiple transcriptional regulators and chromatin binding factors. Retrovirology. 2008;5:48. doi: 10.1186/1742-4690-5-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.De Rijck J, de Kogel C, Demeulemeester J, Vets S, El Ashkar S, Malani N, Bushman FD, Landuyt B, Husson SJ, Busschots K, et al. The BET family of proteins targets moloney murine leukemia virus integration near transcription start sites. Cell Rep. 2013;5:886–894. doi: 10.1016/j.celrep.2013.09.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sharma A, Larue RC, Plumb MR, Malani N, Male F, Slaughter A, Kessl JJ, Shkriabai N, Coward E, Aiyer SS, et al. BET proteins promote efficient murine leukemia virus integration at transcription start sites. Proc Natl Acad Sci USA. 2013;110:12036–12041. doi: 10.1073/pnas.1307157110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Knaus RJ, Hippenmeyer PJ, Misra TK, Grandgenett DP, Müller UR, Fitch WM. Avian retrovirus pp32 DNA binding protein. Preferential binding to the promoter region of long terminal repeat DNA. Biochemistry. 1984;23:350–359. doi: 10.1021/bi00297a026. [DOI] [PubMed] [Google Scholar]
- 47.Engelman A, Cherepanov P. Retroviral Integrase Structure and DNA Recombination Mechanism. Microbiol Spectr. 2014;2:1–22. doi: 10.1128/microbiolspec.MDNA3-0024-2014.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Valkov E, Gupta SS, Hare S, Helander A, Roversi P, McClure M, Cherepanov P. Functional and structural characterization of the integrase from the prototype foamy virus. Nucleic Acids Res. 2009;37:243–255. doi: 10.1093/nar/gkn938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gupta K, Curtis JE, Krueger S, Hwang Y, Cherepanov P, Bushman FD, Van Duyne GD. Solution conformations of prototype foamy virus integrase and its stable synaptic complex with U5 viral DNA. Structure. 2012;20:1918–1928. doi: 10.1016/j.str.2012.08.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Pandey KK, Bera S, Korolev S, Campbell M, Yin Z, Aihara H, Grandgenett DP. Rous sarcoma virus synaptic complex capable of concerted integration is kinetically trapped by human immunodeficiency virus integrase strand transfer inhibitors. J Biol Chem. 2014;289:19648–19658. doi: 10.1074/jbc.M114.573311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Shi K, Pandey KK, Bera S, Vora AC, Grandgenett DP, Aihara H. A possible role for the asymmetric C-terminal domain dimer of Rous sarcoma virus integrase in viral DNA binding. PLoS One. 2013;8:e56892. doi: 10.1371/journal.pone.0056892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bojja RS, Andrake MD, Weigand S, Merkel G, Yarychkivska O, Henderson A, Kummerling M, Skalka AM. Architecture of a full-length retroviral integrase monomer and dimer, revealed by small angle X-ray scattering and chemical cross-linking. J Biol Chem. 2011;286:17047–17059. doi: 10.1074/jbc.M110.212571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pandey KK, Bera S, Grandgenett DP. The HIV-1 integrase monomer induces a specific interaction with LTR DNA for concerted integration. Biochemistry. 2011;50:9788–9796. doi: 10.1021/bi201247f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lee SP, Xiao J, Knutson JR, Lewis MS, Han MK. Zn2+ promotes the self-association of human immunodeficiency virus type-1 integrase in vitro. Biochemistry. 1997;36:173–180. doi: 10.1021/bi961849o. [DOI] [PubMed] [Google Scholar]
- 55.Li M, Craigie R. Processing of viral DNA ends channels the HIV-1 integration reaction to concerted integration. J Biol Chem. 2005;280:29334–29339. doi: 10.1074/jbc.M505367200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Bera S, Pandey KK, Vora AC, Grandgenett DP. Molecular Interactions between HIV-1 integrase and the two viral DNA ends within the synaptic complex that mediates concerted integration. J Mol Biol. 2009;389:183–198. doi: 10.1016/j.jmb.2009.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hare S, Gupta SS, Valkov E, Engelman A, Cherepanov P. Retroviral intasome assembly and inhibition of DNA strand transfer. Nature. 2010;464:232–236. doi: 10.1038/nature08784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Nowotny M. Retroviral integrase superfamily: the structural perspective. EMBO Rep. 2009;10:144–151. doi: 10.1038/embor.2008.256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Montaño SP, Pigli YZ, Rice PA. The μ transpososome structure sheds light on DDE recombinase evolution. Nature. 2012;491:413–417. doi: 10.1038/nature11602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Maertens GN, Hare S, Cherepanov P. The mechanism of retroviral integration from X-ray structures of its key intermediates. Nature. 2010;468:326–329. doi: 10.1038/nature09517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Davies DR, Goryshin IY, Reznikoff WS, Rayment I. Three-dimensional structure of the Tn5 synaptic complex transposition intermediate. Science. 2000;289:77–85. doi: 10.1126/science.289.5476.77. [DOI] [PubMed] [Google Scholar]
- 62.Hare S, Maertens GN, Cherepanov P. 3’-processing and strand transfer catalysed by retroviral integrase in crystallo. EMBO J. 2012;31:3020–3028. doi: 10.1038/emboj.2012.118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Yin Z, Lapkouski M, Yang W, Craigie R. Assembly of prototype foamy virus strand transfer complexes on product DNA bypassing catalysis of integration. Protein Sci. 2012;21:1849–1857. doi: 10.1002/pro.2166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Espeseth AS, Felock P, Wolfe A, Witmer M, Grobler J, Anthony N, Egbertson M, Melamed JY, Young S, Hamill T, et al. HIV-1 integrase inhibitors that compete with the target DNA substrate define a unique strand transfer conformation for integrase. Proc Natl Acad Sci USA. 2000;97:11244–11249. doi: 10.1073/pnas.200139397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Grobler JA, Stillmock K, Hu B, Witmer M, Felock P, Espeseth AS, Wolfe A, Egbertson M, Bourgeois M, Melamed J, et al. Diketo acid inhibitor mechanism and HIV-1 integrase: implications for metal binding in the active site of phosphotransferase enzymes. Proc Natl Acad Sci USA. 2002;99:6661–6666. doi: 10.1073/pnas.092056199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Hare S, Vos AM, Clayton RF, Thuring JW, Cummings MD, Cherepanov P. Molecular mechanisms of retroviral integrase inhibition and the evolution of viral resistance. Proc Natl Acad Sci USA. 2010;107:20057–20062. doi: 10.1073/pnas.1010246107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Cherepanov P, Maertens GN, Hare S. Structural insights into the retroviral DNA integration apparatus. Curr Opin Struct Biol. 2011;21:249–256. doi: 10.1016/j.sbi.2010.12.005. [DOI] [PubMed] [Google Scholar]
- 68.Pommier Y, Johnson AA, Marchand C. Integrase inhibitors to treat HIV/AIDS. Nat Rev Drug Discov. 2005;4:236–248. doi: 10.1038/nrd1660. [DOI] [PubMed] [Google Scholar]
- 69.Pommier Y, Marchand C. Interfacial inhibitors: targeting macromolecular complexes. Nat Rev Drug Discov. 2012;11:25–36. doi: 10.1038/nrd3404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Neamati N. HIV-1 integrase inhibitor design: Overview and historial perspectives. In HIV-1 Integrase, Mechanism and Inhibitor Design. New Jersey: John Wiley & Sons, Inc; 2011. pp. 165–196. [Google Scholar]
- 71.Métifiot M, Marchand C, Pommier Y. HIV integrase inhibitors: 20-year landmark and challenges. Adv Pharmacol. 2013;67:75–105. doi: 10.1016/B978-0-12-405880-4.00003-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Di Santo R. Inhibiting the HIV integration process: past, present, and the future. J Med Chem. 2014;57:539–566. doi: 10.1021/jm400674a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Craigie R. The road to HIV-1 integrase inhibitors: the case for supporting basic research. Future Virol. 2014;9:899–903. doi: 10.2217/fvl.14.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Pandey KK, Bera S, Vora AC, Grandgenett DP. Physical trapping of HIV-1 synaptic complex by different structural classes of integrase strand transfer inhibitors. Biochemistry. 2010;49:8376–8387. doi: 10.1021/bi100514s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Bera S, Pandey KK, Vora AC, Grandgenett DP. HIV-1 integrase strand transfer inhibitors stabilize an integrase-single blunt-ended DNA complex. J Mol Biol. 2011;410:831–846. doi: 10.1016/j.jmb.2011.01.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Wang JY, Ling H, Yang W, Craigie R. Structure of a two-domain fragment of HIV-1 integrase: implications for domain organization in the intact protein. EMBO J. 2001;20:7333–7343. doi: 10.1093/emboj/20.24.7333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Yang ZN, Mueser TC, Bushman FD, Hyde CC. Crystal structure of an active two-domain derivative of Rous sarcoma virus integrase. J Mol Biol. 2000;296:535–548. doi: 10.1006/jmbi.1999.3463. [DOI] [PubMed] [Google Scholar]
- 78.Hightower KE, Wang R, Deanda F, Johns BA, Weaver K, Shen Y, Tomberlin GH, Carter HL, Broderick T, Sigethy S, et al. Dolutegravir (S/GSK1349572) exhibits significantly slower dissociation than raltegravir and elvitegravir from wild-type and integrase inhibitor-resistant HIV-1 integrase-DNA complexes. Antimicrob Agents Chemother. 2011;55:4552–4559. doi: 10.1128/AAC.00157-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Grobler J, McKemma PM, Ly S, Stillmock KA, Bahnck CM, Danovich RM, Dornadula G, Hazuda D, Miller MD. HIV integrase inhibitor dissociation rates correlate with efficacy in vitro. Antiviral Ther. 2009;14 Suppl 1:A27. [Google Scholar]
- 80.Messiaen P, Wensing AM, Fun A, Nijhuis M, Brusselaers N, Vandekerckhove L. Clinical use of HIV integrase inhibitors: a systematic review and meta-analysis. PLoS One. 2013;8:e52562. doi: 10.1371/journal.pone.0052562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.White KL, Raffi F, Miller MD. Resistance analyses of integrase strand transfer inhibitors within phase 3 clinical trials of treatment-naive patients. Viruses. 2014;6:2858–2879. doi: 10.3390/v6072858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Rawson JM, Mansky LM. Retroviral vectors for analysis of viral mutagenesis and recombination. Viruses. 2014;6:3612–3642. doi: 10.3390/v6093612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Llibre JM, Pulido F, García F, García Deltoro M, Blanco JL, Delgado R. Genetic barrier to resistance for dolutegravir. AIDS Rev. 2015;17:56–64. [PubMed] [Google Scholar]
- 84.Mesplède T, Wainberg MA. Is resistance to dolutegravir possible when this drug is used in first-line therapy? Viruses. 2014;6:3377–3385. doi: 10.3390/v6093377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Miller MM, Liedtke MD, Lockhart SM, Rathbun RC. The role of dolutegravir in the management of HIV infection. Infect Drug Resist. 2015;8:19–29. doi: 10.2147/IDR.S58706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.DeAnda F, Hightower KE, Nolte RT, Hattori K, Yoshinaga T, Kawasuji T, Underwood MR. Dolutegravir interactions with HIV-1 integrase-DNA: structural rationale for drug resistance and dissociation kinetics. PLoS One. 2013;8:e77448. doi: 10.1371/journal.pone.0077448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Anstett K, Mesplede T, Oliveira M, Cutillas V, Wainberg MA. Dolutegravir resistance mutation R263K cannot coexist in combination with many classical integrase inhibitor resistance substitutions. J Virol. 2015;89:4681–4684. doi: 10.1128/JVI.03485-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Andrews CD, Spreen WR, Mohri H, Moss L, Ford S, Gettie A, Russell-Lodrigue K, Bohm RP, Cheng-Mayer C, Hong Z, et al. Long-acting integrase inhibitor protects macaques from intrarectal simian/human immunodeficiency virus. Science. 2014;343:1151–1154. doi: 10.1126/science.1248707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Niederer HA, Bangham CR. Integration site and clonal expansion in human chronic retroviral infection and gene therapy. Viruses. 2014;6:4140–4164. doi: 10.3390/v6114140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Antoine C, Müller S, Cant A, Cavazzana-Calvo M, Veys P, Vossen J, Fasth A, Heilmann C, Wulffraat N, Seger R, et al. Long-term survival and transplantation of haemopoietic stem cells for immunodeficiencies: report of the European experience 1968-99. Lancet. 2003;361:553–560. doi: 10.1016/s0140-6736(03)12513-5. [DOI] [PubMed] [Google Scholar]
- 91.Cavazzana-Calvo M, Hacein-Bey S, de Saint Basile G, Gross F, Yvon E, Nusbaum P, Selz F, Hue C, Certain S, Casanova JL, et al. Gene therapy of human severe combined immunodeficiency (SCID)-X1 disease. Science. 2000;288:669–672. doi: 10.1126/science.288.5466.669. [DOI] [PubMed] [Google Scholar]
- 92.Gaspar HB, Parsley KL, Howe S, King D, Gilmour KC, Sinclair J, Brouns G, Schmidt M, Von Kalle C, Barington T, et al. Gene therapy of X-linked severe combined immunodeficiency by use of a pseudotyped gammaretroviral vector. Lancet. 2004;364:2181–2187. doi: 10.1016/S0140-6736(04)17590-9. [DOI] [PubMed] [Google Scholar]
- 93.Hacein-Bey-Abina S, Von Kalle C, Schmidt M, McCormack MP, Wulffraat N, Leboulch P, Lim A, Osborne CS, Pawliuk R, Morillon E, et al. LMO2-associated clonal T cell proliferation in two patients after gene therapy for SCID-X1. Science. 2003;302:415–419. doi: 10.1126/science.1088547. [DOI] [PubMed] [Google Scholar]
- 94.Howe SJ, Mansour MR, Schwarzwaelder K, Bartholomae C, Hubank M, Kempski H, Brugman MH, Pike-Overzet K, Chatters SJ, de Ridder D, et al. Insertional mutagenesis combined with acquired somatic mutations causes leukemogenesis following gene therapy of SCID-X1 patients. J Clin Invest. 2008;118:3143–3150. doi: 10.1172/JCI35798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Hacein-Bey-Abina S, Pai SY, Gaspar HB, Armant M, Berry CC, Blanche S, Bleesing J, Blondeau J, de Boer H, Buckland KF, et al. A modified γ-retrovirus vector for X-linked severe combined immunodeficiency. N Engl J Med. 2014;371:1407–1417. doi: 10.1056/NEJMoa1404588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Sakuma T, Barry MA, Ikeda Y. Lentiviral vectors: basic to translational. Biochem J. 2012;443:603–618. doi: 10.1042/BJ20120146. [DOI] [PubMed] [Google Scholar]
- 97.Suerth JD, Labenski V, Schambach A. Alpharetroviral vectors: from a cancer-causing agent to a useful tool for human gene therapy. Viruses. 2014;6:4811–4838. doi: 10.3390/v6124811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Moiani A, Suerth JD, Gandolfi F, Rizzi E, Severgnini M, De Bellis G, Schambach A, Mavilio F. Genome-wide analysis of alpharetroviral integration in human hematopoietic stem/progenitor cells. Genes (Basel) 2014;5:415–429. doi: 10.3390/genes5020415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Aiyer S, Rossi P, Malani N, Schneider WM, Chandar A, Bushman FD, Montelione GT, Roth MJ. Structural and sequencing analysis of local target DNA recognition by MLV integrase. Nucleic Acids Res. 2015;43:5647–5663. doi: 10.1093/nar/gkv410. [DOI] [PMC free article] [PubMed] [Google Scholar]