

Abstract
The APETALA2/Ethylene-Responsive Factor (AP2/ERF) superfamily of transcription factors (TFs) regulates physiological, developmental and stress responses. Most of the AP2/ERF TFs belong to the ERF family in both dicotyledonous and monocotyledonous plants. ERFs are implicated in the responses to both biotic and abiotic stress and occasionally impart multiple stress tolerance. Studies have revealed that ERF gene function is conserved in dicots and monocots. Moreover, successful stress tolerance phenotypes are observed on expression in heterologous systems, making ERFs promising candidates for engineering stress tolerance in plants. In this review, we summarize the role of ERFs in general stress tolerance, including responses to biotic and abiotic stress factors, and endeavor to understand the cascade of ERF regulation resulting in successful signal-to-response translation in monocotyledonous plants.
Keywords: AP2/ERFs, ethylene-responsive factors, biotic stress, abiotic stress, monocot
Introduction
The APETALA2/Ethylene-Responsive Factor (AP2/ERF) superfamily of transcription factors (TFs) regulates diverse plant responses, from light acclimation (Vogel et al., 2014) to developmental responses such as flower pedicel abscission (Nakano et al., 2014), leaf senescence (Koyama et al., 2013), cell proliferation and shoot branching (Mehrnia et al., 2013). Other responses regulated by this superfamily include responses to biotic and abiotic stress, and overlapping responses to multiple stress factors (Jung et al., 2007). Originally considered to be plant-specific (Riechmann et al., 2000; Gutterson and Reuber, 2004), the AP2/ERF superfamily is now thought to have evolved through lateral gene transfer from bacteria and viruses into plants (Licausi et al., 2013).
The AP2/ERF superfamily is characterized by the AP2 domain, a conserved 60–70 amino acid DNA binding domain (DBD), which was initially identified in homeotic genes regulating flower development in Arabidopsis thaliana (Jofuku et al., 1994). Based on the number of AP2 domains, exon-intron structure and additional conserved protein motifs in Arabidopsis and rice, the AP2/ERF superfamily is subdivided into the following three families: ERF, AP2, and RAV (Nakano et al., 2006). Other reviews (Mizoi et al., 2012; Licausi et al., 2013) summarize the basis of this classification in detail. Unless stated otherwise, ERFs in this review refer to members of the ERF family.
The ERF family is further subdivided into the ERF and the C-repeat-binding factor/dehydration-responsive-element–binding protein (CBF/DREB) subfamilies based on different conserved amino acid residues within their respective AP2 domains (Sakuma et al., 2002; Nakano et al., 2006; Lata and Prasad, 2011). The amino acid sequence variation in the AP2 domains of ERF and DREB subfamily members reflects the differences in the DNA binding specificities of these TFs and forms the basis for conditional responses regulating different sets of stress-responsive genes. In dicotyledonous plants, CBF/DREB proteins bind to the C-repeat/dehydration-responsive-element (CRT/DRE; A/GCCGAC) cis-acting element in the promoters of target genes and are usually associated with responses to abiotic stress and the phytohormone abscisic acid (ABA; Sakuma et al., 2002; Licausi et al., 2013). ERF subfamily members bind to the GCC-box (AGCCGCC) and are associated with biotic stress responses, responses to the phytohormones jasmonic acid (JA) and ethylene (ET), wounding, and development (Sakuma et al., 2002). Several exceptions to this broad generalization exist. Arabidopsis thaliana (At) ERF1 (ERF-subfamily group III A-4) and AtTINY (DREB subfamily group IX B3; Nakano et al., 2006) are each capable of binding to both GCC and CRT/DRE elements, binding to the GCC box at the promoters of biotic stress-inducible genes and to the CRT/DRE elements of genes associated with abiotic stress tolerance thereby possibly balancing responses to biotic and abiotic stress conditions (Sun et al., 2008; Cheng et al., 2013). Also, ERF TFs of the subgroup IXA of the ERF subfamily in Arabidopsis, Nicotiana tabacum, and Catharanthus roseus contain fully conserved AP2 domains, but recognize distinct DNA sequences (Shoji et al., 2013). Taken together, although promoter motifs defining ERF and CBF/DREB-specific binding have been identified, both divergence and crosstalk of binding capacities exist, possibly promoting plasticity in stress tolerance.
In monocotyledonous plants, studies on the ERF family conducted over the last decade have mostly concentrated on the heterologous expression of Arabidopsis family members in crop plants and vice versa. Interestingly, most of these reports reveal augmented stress tolerance under transgenic conditions in comparison with wild-type plants. These findings suggest that the mechanisms for the translation of ERF function and their downstream targets are conserved between dicots and monocots. Here, we summarize current knowledge on the regulation and biological roles of members of the ERF family of AP2/ERF TFs in monocotyledonous plants.
The Monocot ERF Regulatome and Multistep Control of Stress Responses
The number of AP2/ERF TF genes that has been identified in monocot genomes ranges from 53 in barley (Hordeum vulgare; Hv) to 184 in maize (Zea mays; Zm; Table 1; Zhuang et al., 2011a; Du et al., 2014). Similar to AP2/ERFs in Arabidopsis, immense diversification of the AP2/ERF superfamily in monocots is observed with a clear dominance of the ERF family (Table 1). Bioinformatic studies on the chromosomal clustering of genes in monocots showed that segmental and tandem duplication events have led to the evolution and extensive diversification of the AP2/ERF superfamily (Sharoni et al., 2011; Lata et al., 2014), while polyploidy adds to increased complexity in wheat (Zhuang et al., 2011b). Moreover, extensive synteny exists within monocots, which is restricted by comparison in Arabidopsis (Rashid et al., 2012).
TABLE 1.
Summary of ERF family members in monocots.
| Plant | Total number of AP2/ERF TFs identified | ERF family | Percentage ERF family | DREB subfamily members of the ERF family | ERF subfamily members of the ERF family | References |
|---|---|---|---|---|---|---|
| Rice | 164 | 113 | 68.9 | 52 | 79 | Nakano et al. (2006) |
| Wheat | 117 | 104 | 88.8 | 57 | 47 | Zhuang et al. (2011b) |
| Maize | 184 | 158 | 85.8 | 51 | 107 | Du et al. (2014) |
| Foxtail millet | 171 | 138 | 80.7 | 48 | 90 | Lata et al. (2014) |
| Sorghum | 126 | 105 | 83.3 | 52 | 53 | Yan et al. (2013) |
| Barley | 53 | 40 | 75.4 | 18 | 22 | Zhuang et al. (2011a) |
Despite being etymologically characterized as ET-responsive, ERFs can also be regulated in a manner independent of ET, by stress or by phytohormones including JA and ABA. In addition, transcriptional activators of the ET signaling pathway, including EHTYLENE INSENSITIVE2 (EIN2) may induce the expression of ERFs. For example, ET induces the expression of the rice (Oryza sativa; Os) OsERF063 and OsERF073 in an OsEIN2-dependent manner (Figure 1A; Ma et al., 2013). In general, the molecular regulation of gene expression relies on various steps of transcriptional activation/repression and post transcriptional and post translational control. While some information of transcriptional/post transcriptional control of ERFs in monocots is available (Figure 1), the involvement of post-translation control of ERFs in monocots remains mostly to be elucidated. The only clear evidence concerns the activation of rice SALT-RESPONSIVE-ERF1 (OsSERF1) upon its phosphorylation by MITOGEN-ACTIVATED PROTEIN KINASE5 (OsMAPK5). OsSERF1 phosphorylation enhances the transcription of downstream targets that include OsMAPK5 and OsSERF1 in a positive feedback loop (Schmidt et al., 2013; Figure 1B).
FIGURE 1.
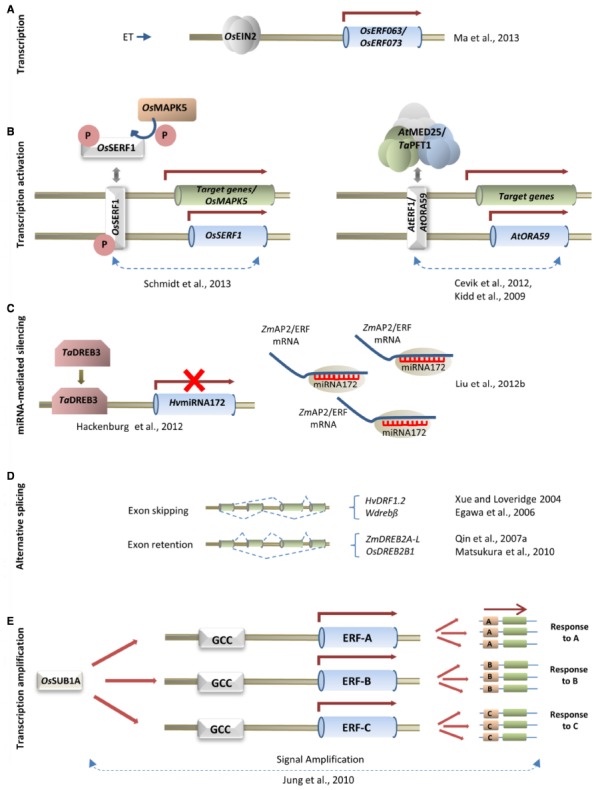
ERF regulation at multiple levels to confer stress tolerance in monocots. (A) Transcription: OsERF063 and OsERF073 transcript accumulation in rice is induced by ethylene (ET) in an OsEIN2-dependent manner. (B) Transcription activation. On left: Stress-induced phosphorylation of OsSERF1 by OsMAPK5 in rice produces the active ERF conformation facilitating DNA binding and thereby transcriptional activation of downstream genes, including OsMAPK5, and the ERF OsSERF1 itself in a positive feedback loop as indicated by the dotted line. On right: ERF transcription might be activated by the concerted action of the mediator complex (AtMED25/TaPFT1) binding to both AtERF1 and AtORA59 triggering target gene expression as well as AtORA59 transcript accumulation in a positive feedback loop that is indicated by the dotted line. (C) miRNA-mediated silencing: barley miRNA172 is repressed by TaDREB3 while miRNA172 in maize targets at least two AP2/ERF transcripts; the data suggest that miRNA172 acts both downstream and upstream of AP2/ERFs to regulate AP2/ERF transcript accumulation. (D) Alternative splicing: mRNA abundance of different DREB2 isoforms in barley, wheat, maize and rice is controlled by stress-induced alternative splicing. (E) Transcription amplification: Signal amplification by (OsSUB1A) that induces downstream ERFs (Jung et al., 2010) possibly by binding to upstream GCC box in the promoters of the other ERF genes. This in turn leads to the expression of specific stress responsive downstream targets (A–C). Abbreviations: miRNA, microRNA; TaDREB3, Triticum aestivum Dehydration Responsive Element Binding 3; AP2/ERF, APETALA2/ETHYLENE RESPONSIVE FACTOR; OsERF, Oryza sativa Ethylene Responsive Factor; OsEIN2, Oryza sativa Ethylene Insensitive 2; AtMED25, Arabidopsis thaliana MEDIATOR 25; TaPFT1, Triticum aestivum PHYTOCHROME AND FLOWERING TIME 1; AtERF1, Arabidopsis thaliana Ethylene Responsive Factor 1; AtORA59, Arabidopsis thaliana OCTADECANOID-RESPONSIVE ARABIDOPSIS APETALA2/ETHYLENE RESPONSE FACTOR domain protein59; OsSERF1, Oryza sativa SALT-RESPONSIVE-ERF1; OsMAPK5, Oryza sativa MITOGEN-ACTIVATED PROTEIN KINASE5, OsSUB1A, Oryza sativa SUBMERGENCE1 locus gene A; GCC box, (AGCCGCC) cis-acting element; TFs, Transcription factors.
A recently identified component of the transcriptional machinery, the mediator complex is a large multi-protein complex that forms a central transcriptional co-regulator (Bäckström et al., 2007). It binds to gene promoter motifs forming a bridge to facilitate binding of TFs and RNA polymerase II. The mediator complex consists of about 20–30 subunits and is conserved in eukaryotes from yeast to plants and humans. The Arabidopsis MEDIATOR25 (AtMED25) functions as a positive regulator of JA-dependent gene expression (Kidd et al., 2009; Cevik et al., 2012). It interacts with four group-IX ERFs, including AtERF1 and Arabidopsis OCTADECANOID-RESPONSIVE ARABIDOPSIS APETALA2/ETHYLENE RESPONSE FACTOR domain protein59 (AtORA59), at the promoters of these genes. AtERF1 and AtORA59, in turn, bind to the AtORA59 promoter (Zander et al., 2014) resulting in amplified expression of the downstream targets of AtORA59 (Cevik et al., 2012; Zander et al., 2014). The wheat (Triticum aestivum; Ta) MED25 homolog PHYTOCHROME AND FLOWERING TIME 1 (TaPFT1), when expressed constitutively in Arabidopsis, fully complements the Arabidopsis Atmed25 phenotype. This result suggests a conserved function for MED25 in Arabidopsis and wheat (Kidd et al., 2009; Figure 1B).
Other studies suggest a role for miRNAs (20–24 nucleotide non-coding RNAs that post-transcriptionally repress gene expression) in the regulation of AP2/ERFs. Maize Zm-miRNA28 is associated with germination and predicted to target the ERF-like gene TC313437 (Ding et al., 2013). Another example in maize is related to abiotic stress. Waterlogging due to floods or sudden rains causes hypoxia of roots leading to anaerobic stress-induced metabolic changes and the accumulation of reactive oxygen species. Studies on short term waterlogging in two maize lines revealed the up regulation of Zm-microRNA172 (Zm-miRNA172), which targets the repression of the AP2/ERF TFs ZM5G862109 and ZM2G076602 (Liu et al., 2012b). Interestingly, miRNA172 was also identified in barley among miRNAs that were down-regulated by the heterologous expression of wheat DREB3 (TaDREB3; Hackenberg et al., 2012). TaDREB3 overexpression in barley resulted in robust drought-tolerant plants in comparison to severely wilted wild type plants on drought stress. The TaDREB3-dependent downregulation of Hv-miRNA172 has been associated with the regulation of the expression of AP2/ERFs of the AP2 subfamily (Hackenberg et al., 2012). Taken together, miRNA172 represents a versatile regulatory node both targeting the expression of AP2/ERFs or itself being targeted by AP2/ERF TFs such as TaDREB3 (Figure 1C).
Alternative splicing has been reported to play a major role in the stress-induced transcriptome of plants, with housekeeping genes undergoing normal splicing and genes encoding stress-related proteins over-represented in the alternatively spliced fraction (Li et al., 2013). Interestingly, the DREB2-like barley gene DEHYDRATION RESPONSIVE FACTOR1 (HvDRF1) and its orthologs in wheat (TaDRF1/Wdreb2), maize (ZmDREB2A) and rice (OsDREB2B) undergo alternative splicing by differential exon usage (Xue and Loveridge, 2004; Egawa et al., 2006; Qin et al., 2007a; Matsukura et al., 2010). While the inactive forms of HvDRF1 (HvDRF1.2) and Wdreb2 (Wdrebß) originate due to exon skipping causing a premature termination before the AP2 sequence (Xue and Loveridge, 2004; Egawa et al., 2006), the inactive forms of OsDREB2B (OsDREB2B1) and ZmDREB2A (ZmDREB2A-L) are similarly terminated prematurely due to frameshift caused by a 53 bp exon retention (Qin et al., 2007a; Matsukura et al., 2010; Figure 1D). HvDRF1 produces two active ERF isoforms (HvDRF1.1 and HvDRF1.3) associated with ABA dependent drought tolerance. The wheat gene Wdreb2 produces three isoforms, Wdrebα, Wdrebß, and Wdrebγ, which are homologous to HvDRF1.1, HvDRF1.2, and HvDRF1.3, respectively. The accumulation of Wdrebα and Wdrebγ transcripts is transient and more pronounced during drought/salt stress, while all three forms are induced by low temperature (Egawa et al., 2006). Likewise, the active isoform of the maize gene ZmDREB2A, ZmDREB2A-S, is significantly induced by drought and heat stress (Qin et al., 2007a). In rice, only the functionally active splice isoform of OsDREB2B (OsDREB2B2) is induced by low temperature stress (Matsukura et al., 2010). Taken together, alternative splicing allows isoform-specific transcript level adjustments regulating responses to normal and stress-induced conditions (Xue and Loveridge, 2004).
ERFs in Monocotyledonous Plants: Convergent Biotic and Abiotic Stress-responsive Nodes
ERFs are versatile TFs that are involved in development as well as responses to different types of stresses. Often, ERF TFs affect cross talk between different responses acting at the interface between different signaling cascades. In Arabidopsis, for example, AtERF1 binds to different promoter elements to regulate responses to pathogen resistance (by binding to the GCC box) and to drought, salt and heat stress (by binding to the DRE/CRT element; Cheng et al., 2013). Similarly, TaERF1 binds to both the GCC box and the DRE/CRT elements, regulating responses to both biotic and abiotic stress in wheat (Xu et al., 2007). Several ERF subfamily members in monocots are associated with development and responses to biotic and abiotic stress, whereas CBF/DREB subfamily members to our present knowledge appear mostly related to responses to abiotic stress.
The ERF Subfamily: Stress and Developmental Responses
Several monocot ERFs of the ERF subfamily are induced in response to development and biotic stresses, occasionally imparting multiple stress tolerance upon heterologous expression. The ectopic expression of the barley ERF gene Hv ROOT ABUNDANT FACTOR (HvRAF) in Arabidopsis promotes resistance to Ralstonia solanacearum and enhances tolerance to salt stress (Jung et al., 2007). In addition, constitutive expression of the wheat ERF genes, TaERF1 in Arabidopsis and TaERF3 in wheat enhances tolerance to salt and drought stress (Xu et al., 2007; Rong et al., 2014), while the PATHOGEN-INDUCED ERF1 (TaPIE1) increases freezing stress tolerance in wheat (Zhu et al., 2014). Conversely, TaERF4 acts as a negative regulator of salinity tolerance when constitutively expressed in Arabidopsis (Dong et al., 2012). Furthermore, TaERF1 overexpression enhances tolerance to the necrotrophic pathogen Botrytis cinerea in Arabidopsis and to Pseudomonas syringae pathovar tabaci in tobacco compared to the respective wild type conditions (Xu et al., 2007). In response to the powdery mildew fungus Blumeria graminis, TaERF3 is induced in a SA-dependent manner in a powdery mildew resistant wheat cultivar (Zhang et al., 2007). The same gene is induced by methyl jasmonate/ET, which is correlated with resistance against the necrotrophic pathogens Fusarium graminearum and Rhizoctonia cerealis (Zhang et al., 2007). The involvement of ERFs in wheat resistance against R. cerealis was further supported by evidence of enhanced resistance to this pathogen in transgenic TaPIE1 over expressing wheat plants (Zhu et al., 2014).
Transcript abundance of ERFs may contribute to condition-specific responses. For example, the overexpression of the rice gene OsSERF1 in rice leads to salt tolerance (Schmidt et al., 2013) and knockdown of this gene leads to developmental responses such as grain filling. OsSERF1 positively regulates the transcription of target genes, including the MAP kinase kinase OsMAP3K6 and OsMAPK5 (Figure 1B) and other TFs, such as OsDREB2A and negatively regulates RICE PROLAMIN-BOX BINDING FACTOR (RPBF) which is responsible for grain filling (Schmidt et al., 2014). Several other rice ERFs contribute to developmental or biotic stress responses either positively or negatively. While overexpression of the rice ERF OsERF1 in Arabidopsis resulted in stunted growth and increased accumulation of ET responsive genes (Hu et al., 2008), overexpression of another ERF OsEATB (for Oryza sativa ERF protein associated with tillering and panicle branching) resulted in reduced plant height but higher tillering, thereby increasing yield potential (Qi et al., 2011). Also, overexpression of the SNORKEL1 (OsSK1) and SNORKEL2 (OsSK2) genes, which encode ERFs homologous to AtERF1, induces elongated internodes in an ET-dependent manner in rice (Hattori et al., 2009). Finally, OsERF922 is a negative regulator of defense against the rice blast fungus Magnaporthe oryzae and salt stress (Liu et al., 2012a). Interestingly, OsERF922 overexpression enhanced ABA levels in rice, which might be causative for the enhanced susceptibility to M. oryzae, as exogenous ABA treatment also induces susceptibility to this pathogen (Koga et al., 2004).
A few ERFs have been associated with long-distance leaf-to-leaf and root-to-leaf signaling in monocots to induce a state of potentiation/priming against future pathogen attack. Treatment of rice roots with the Pseudomonas isolate EA105 induces the expression of OsERF1 in distal uninfected leaves and enhances resistance to M. oryzae compared to non-infected plants (Spence et al., 2014). Similarly, we related two previously uncharacterized ERFs, HvERF-like and HvERF_44411, to leaf-to-leaf long-distance signaling in barley systemic immunity. Transcript accumulation of both of the ERFs identified was induced in barley leaves that were infected with Pseudomonas syringae pathovar japonica or Xanthomonas translucens pathovar cerealis and in systemic, uninfected leaves of the infected plants (Dey et al., 2014), possibly priming the systemic leaves for rapid transcriptional reprogramming upon further pathogen challenge.
The CBF/DREB Subfamily: Responses to Cold, Drought and Salinity
The abiotic stress-responsive CBF/DREBs highlight the plasticity of the ERF family in multiple stress tolerance. Divided into two subgroups, CBF/DREB1 and DREB2, the CBF/DREB1 members are broadly associated with freezing tolerance and the DREB2s with dehydration and salt stress responses (Choi et al., 2002; Mizoi et al., 2012). Cross talk between these responses has been observed (Oh et al., 2007; Matsukura et al., 2010), which may be partly attributed to a common subset of stress-inducible downstream targets in addition to TF-specific target genes depending on the initial stress response. Target genes of non-cold-inducible CBF/DREB1s, for example, impart drought tolerance and overlap with those that are induced by cold-inducible CBF/DREB1s (Mizoi et al., 2012). The barley cold-responsive genes HvCBF1 (Hordeum vulgare C-repeat-binding-factor 1), HvCBF2, and HvCBF3 are transcriptionally induced by chilling (Choi et al., 2002; Xue, 2003). Low temperature induces the active conformation of HvCBF2, thereby facilitating DNA binding. HvCBF2 thus represents a TF that undergoes temperature-sensitive changes that result in an increased TF-DNA binding affinity and the transcriptional activation of target genes (Xue, 2003). The rice genes OsDREB1A and OsDREB1B are induced by cold and OsDREB2A by dehydration and salinity. However, overexpression of OsDREB1A alone enhanced tolerance to drought, salinity and freezing in both Arabidopsis and rice (Dubouzet et al., 2003; Ito et al., 2006), while OsDREB1B1 increased tolerance to both high and low-temperature stress in transgenic Arabidopsis (Qin et al., 2007b). Individual ectopic expression of the wheat CBF/DREB1s TaCBF14 and TaCBF15, in barley improved tolerance to freezing (Soltész et al., 2013).
ERFs: A Biotechnological Toolkit for Signal Amplification and Multiple Stress Tolerance
In several cases, the robust stress-tolerant phenotypes of plants that over express heterologous ERFs may result in part from signal amplification via the induction of further endogenous ERFs added to other stress-inducible genes. For example, the constitutive expression of TaDREB2 and of TaDREB3 in barley induced tolerance to freezing and drought stress and the endogenous expression of the barley CBF/DREB factors HvCBF1, HvCBF3, HvCBF6, HvCBF10A, HvCBF11, HvCBF15, and HvCBF16, in addition to other stress-responsive genes (Morran et al., 2011). The CRT/DRE elements in the promoter regions of CBF/DREB genes could provide an explanation for the robust amplification of stress signals (Soltész et al., 2013). The role of ERFs in the amplification/multiplicity of stress signals is also notable in rice (Jung et al., 2010; Wan et al., 2011; Zhang et al., 2013). DROUGHT-INDUCED ERF1 (OsDERF1) is induced by drought, ET and ABA, while rice plants overexpressing OsDERF1 show enhanced sensitivity to drought. An opposite phenotype is observed in knockdown plants, revealing that OsDERF1 is a negative regulator of drought tolerance in rice (Wan et al., 2011). Notably, the genes that are activated by OsDERF1 include two ERFs, OsERF3 and OsAP2-39 (Wan et al., 2011). The hierarchal concept of an upstream ERF regulating multiple other ERFs is also observed for SUBMERGENCE1A (OsSUB1A) that is one of three ERFs constituting the Submergence1 (Sub1) locus in rice. OsSUB1A plays a pivotal role in the recovery of submerged rice plants and is associated with negative regulation of gibberellic acid responsiveness and ET production. Overexpression of OsSUB1A results in increased ABA responsiveness and tolerance to oxidative stress (Fukao et al., 2011). Furthermore, comparative transcriptome analysis of submergence-intolerant and submergence-tolerant, OsSUB1A-containing rice lines revealed an OsSUB1A-dependent regulation of 12 target ERFs. Further analysis allowed the classification of these ERFs into three distinct functional categories, thereby controlling different biological aspects of stress tolerance (Jung et al., 2010; Figure 1E).
Conclusion and Perspectives
The TFs of the ERF family are among the most versatile stress-responsive TFs in plants. The extensive plasticity of ERFs in inducing multiple stress tolerance in heterologous plants is perhaps the greatest motivation for exploiting the family for plant biotechnology in light of the changing global climate. While ERFs from newly sequenced crop plants continue to be identified, it will be interesting to further characterize their role at the functional level. Moreover, large-scale transcriptome co-expression analyses may provide a basis for identifying upstream regulatory ‘hub’ ERFs. “Hub” ERFs might activate downstream TFs, including other ERFs, which may in turn lead to signal amplification and multiple stress tolerance. It will further be essential to correlate ERFs with their target cis-elements in different crop plants and to characterize specific and potentially non-specific gene expression changes upon ERF over expression and their effects on the robustness of the plant under stress. With this knowledge, new strategies may be designed to engineer multiple stress tolerance in crops.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Research on ERFs in barley in the lab of ACV is funded by the Deutsche Forschungsgemeinschaft as part of SFB924. We are grateful to Frank Gaupels for critical reading and fruitful comments. We apologize to colleagues whose work could not be cited due to space limitations.
References
- Bäckström S., Elfving N., Nilsson R., Wingsle G., Björklund S. (2007). Purification of a plant mediator from Arabidopsis thaliana identifies PFT1 as the Med25 subunit. Mol. Cell 26, 717–729. 10.1016/j.molcel.2007.05.007 [DOI] [PubMed] [Google Scholar]
- Cevik V., Kidd B. N., Zhang P., Hills C. E., Kiddle S. J., Denby K. J., et al. (2012). MEDIATOR25 acts as an integrative hub for the regulation of jasmonate-responsive gene expression in Arabidopsis. 160, 541–555. 10.1104/pp.112.202697 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng M.-C., Liao P.-M., Kuo W.-W., Lin T.-P. (2013). The Arabidopsis ETHYLENE RESPONSE FACTOR1 regulates abiotic stress-responsive gene expression by binding to different cis-acting elements in response to different stress signals. Plant Physiol. 162, 1566–1582. 10.1104/pp.113.221911 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi D., Rodriguez E. M., Close T. J. (2002). Barley Cbf3 gene identification, expression pattern, and map location. Society 129, 1781–1787. 10.1104/pp.003046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dey S., Wenig M., Langen G., Sharma S., Kugler K. G., Knappe C., et al. (2014). Bacteria-triggered systemic immunity in barley is associated with WRKY and ETHYLENE RESPONSIVE FACTORs but not with salicylic acid. Plant Physiol. 166, 2133–2151. 10.1104/pp.114.249276 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding H., Gao J., Luo M., Peng H., Lin H., Yuan G., et al. (2013). Identification and functional analysis of miRNAs in developing kernels of a viviparous mutant in maize. Crop. J. 1, 115–126. 10.1016/j.cj.2013.07.013 [DOI] [Google Scholar]
- Dong W., Ai X., Xu F., Quan T., Liu S., Xia G. (2012). Isolation and characterization of a bread wheat salinity responsive ERF transcription factor. Gene 511, 38–45. 10.1016/j.gene.2012.09.039 [DOI] [PubMed] [Google Scholar]
- Dubouzet J. G., Sakuma Y., Ito Y., Kasuga M., Dubouzet E. G., Miura S., et al. (2003). OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought, high salt and cold responsive gene expression. Plant J. 33, 751–763. 10.1046/j.1365-313X.2003.01661.x [DOI] [PubMed] [Google Scholar]
- Du H., Huang M., Zhang Z. (2014). Genome-wide analysis of the AP2/ERF gene family in maize waterlogging stress response. Euphytica 198, 115–126. 10.1007/s10681-014-1088-2 [DOI] [Google Scholar]
- Egawa C., Kobayashi F., Ishibashi M., Nakamura T., Nakamura C., Takumi S. (2006). Differential regulation of transcript accumulation and alternative splicing of a DREB2 homolog under abiotic stress conditions in common wheat. Genes Genet. Syst. 81, 77–91. 10.1266/ggs.81.77 [DOI] [PubMed] [Google Scholar]
- Fukao T., Yeung E., Bailey-Serres J. (2011). The submergence tolerance regulator SUB1A mediates crosstalk between submergence and drought tolerance in rice. Plant Cell 23, 412–427. 10.1105/tpc.110.080325 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutterson N., Reuber T. L. (2004). Regulation of disease resistance pathways by AP2/ERF transcription factors. Curr. Opin. Plant Biol. 7, 465–471. 10.1016/j.pbi.2004.04.007 [DOI] [PubMed] [Google Scholar]
- Hackenberg M., Shi B. J., Gustafson P., Langridge P. (2012). A transgenic transcription factor (TaDREB3) in barley affects the expression of microRNAs and other small non-coding RNAs. PLoS ONE 7:e42030. 10.1371/journal.pone.0042030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hattori Y., Nagai K., Furukawa S., Song X.-J., Kawano R., Sakakibara H., et al. (2009). The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 460, 1026–1030. 10.1038/nature08258 [DOI] [PubMed] [Google Scholar]
- Hu Y., Zhao L., Chong K., Wang T. (2008). Overexpression of OsERF1, a novel rice ERF gene, up-regulates ethylene-responsive genes expression besides affects growth and development in Arabidopsis. J. Plant Physiol. 165, 1717–1725. 10.1016/j.jplph.2007.12.006 [DOI] [PubMed] [Google Scholar]
- Ito Y., Katsura K., Maruyama K., Taji T., Kobayashi M., Seki M., et al. (2006). Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol. 47, 141–153. 10.1093/pcp/pci230 [DOI] [PubMed] [Google Scholar]
- Jofuku K. D., den Boer B. G., Van Montagu M., Okamuro J. K. (1994). Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell 6, 1211–1225. 10.1105/tpc.6.9.1211 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung J., Won S. Y., Suh S. C., Kim H., Wing R., Jeong Y., et al. (2007). The barley ERF-type transcription factor HvRAF confers enhanced pathogen resistance and salt tolerance in Arabidopsis. Planta 225, 575–588. 10.1007/s00425-006-0373-2 [DOI] [PubMed] [Google Scholar]
- Jung K.-H., Seo Y.-S., Walia H., Cao P., Fukao T., Canlas P. E., et al. (2010). The submergence tolerance regulator Sub1A mediates stress-responsive expression of AP2/ERF transcription factors. Plant Physiol. 152, 1674–1692. 10.1104/pp.109.152157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kidd B. N., Edgar C. I., Kumar K. K., Aitken E. A., Schenk P. M., Manners J. M., et al. (2009). The mediator complex subunit PFT1 is a key regulator of jasmonate-dependent defense in Arabidopsis. Plant Cell 21, 2237–2252. 10.1105/tpc.109.066910 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koga H., Dohi K., Mori M. (2004). Abscisic acid and low temperatures suppress the whole plant-specific resistance reaction of rice plants to the infection of Magnaporthe grisea. Physiol. Mol. Plant Pathol. 65, 3–9. 10.1016/j.pmpp.2004.11.002 [DOI] [Google Scholar]
- Koyama T., Nii H., Mitsuda N., Ohta M., Kitajima S., Ohme-Takagi M., et al. (2013). A regulatory cascade involving class II ETHYLENE RESPONSE FACTOR transcriptional repressors operates in the progression of leaf senescence. Plant Physiol. 162, 991–1005. 10.1104/pp.113.218115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lata C., Mishra A. K., Muthamilarasan M., Bonthala V. S., Khan Y., Prasad M. (2014). Genome-wide investigation and expression profiling of AP2/ERF transcription factor superfamily in foxtail millet (Setaria italica L.). PLoS ONE 9:e113092. 10.1371/journal.pone.0113092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lata C., Prasad M. (2011). Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 62, 4731–4748. 10.1093/jxb/err210 [DOI] [PubMed] [Google Scholar]
- Licausi F., Ohme-Takagi M., Perata P. (2013). APETALA2/Ethylene responsive factor (AP2/ERF) transcription factors: mediators of stress responses and developmental programs. New Phytol. 199, 639–649. 10.1111/nph.12291 [DOI] [PubMed] [Google Scholar]
- Li W., Lin W.-D., Ray P., Lan P., Schmidt W. (2013). Genome-wide detection of condition-sensitive alternative splicing in Arabidopsis roots. Plant Physiol. 162, 1750–1763. 10.1104/pp.113.217778 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu D., Chen X., Liu J., Ye J., Guo Z. (2012a). The rice ERF transcription factor OsERF922 negatively regulates resistance to Magnaporthe oryzae and salt tolerance. J. Exp. Bot. 63, 3899–3912. 10.1093/jxb/ers079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Z., Kumari S., Zhang L., Zheng Y., Ware D. (2012b). Characterization of miRNAs in response to short-term waterlogging in three inbred lines of Zea mays. PLoS ONE 7:e39786. 10.1371/journal.pone.0039786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma B., He S. J., Duan K. X., Yin C. C., Chen H., Yang C., et al. (2013). Identification of rice ethylene-response mutants and characterization of MHZ7/OsEIN2 in distinct ethylene response and yield trait regulation. Mol. Plant 6, 1830–1848. 10.1093/mp/sst087 [DOI] [PubMed] [Google Scholar]
- Matsukura S., Mizoi J., Yoshida T., Todaka D., Ito Y., Maruyama K., et al. (2010). Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Mol. Genet. Genomics 283, 185–196. 10.1007/s00438-009-0506-y [DOI] [PubMed] [Google Scholar]
- Mehrnia M., Balazadeh S., Zanor M.-I., Mueller-Roeber B. (2013). EBE, an AP2/ERF transcription factor highly expressed in proliferating cells, affects shoot architecture in Arabidopsis. Plant Physiol. 162, 842–857. 10.1104/pp.113.214049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizoi J., Shinozaki K., Yamaguchi-Shinozaki K. (2012). AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta Gene Regul. Mech. 1819, 86–96. 10.1016/j.bbagrm.2011.08.004 [DOI] [PubMed] [Google Scholar]
- Morran S., Eini O., Pyvovarenko T., Parent B., Singh R., Ismagul A., et al. (2011). Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors. Plant Biotechnol. J. 9, 230–249. 10.1111/j.1467-7652.2010.00547.x [DOI] [PubMed] [Google Scholar]
- Nakano T., Fujisawa M., Shima Y., Ito Y. (2014). The AP2/ERF transcription factor SlERF52 functions in flower pedicel abscission in tomato. J. Exp. Bot. 65, 3111–3119. 10.1093/jxb/eru154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakano T., Suzuki K., Fujimura T., Shinshi H. (2006). Genome-wide analysis of the ERF gene family. Plant Physiol. 140, 411–432. 10.1104/pp.105.073783 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh S. J., Kwon C. W., Choi D. W., Song S. I., Kim J. K. (2007). Expression of barley HvCBF4 enhances tolerance to abiotic stress in transgenic rice. Plant Biotechnol. J. 5, 646–656. 10.1111/j.1467-7652.2007.00272.x [DOI] [PubMed] [Google Scholar]
- Qin F., Kakimoto M., Sakuma Y., Maruyama K., Osakabe Y., Tran L. S. P., et al. (2007a). Regulation and functional analysis of ZmDREB2A in response to drought and heat stresses in Zea mays L. Plant J. 50, 54–69. 10.1111/j.1365-313X.2007.03034.x [DOI] [PubMed] [Google Scholar]
- Qin Q., Liu J., Zhang Z., Peng R., Xiong A., Yao Q., et al. (2007b). Isolation, optimization, and functional analysis of the cDNA encoding transcription factor OsDREB1B in Oryza sativa L. Mol. Breed. 19, 329–340. 10.1007/s11032-006-9065-7 [DOI] [Google Scholar]
- Qi W., Sun F., Wang Q., Chen M., Huang Y., Feng Y.-Q., et al. (2011). Rice ethylene-response AP2/ERF factor OsEATB restricts internode elongation by down-regulating a gibberellin biosynthetic gene. Plant Physiol. 157, 216–228. 10.1104/pp.111.179945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rashid M., Guangyuan H., Guangxiao Y., Hussain J., Xu Y. (2012). AP2/ERF transcription factor in rice: genome-wide canvas and syntenic relationships between monocots and eudicots. Evol. Bioinforma. 2012, 321–355. 10.4137/EBO.S9369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riechmann J. L., Heard J., Martin G., Reuber L., Jiang C., Keddie J., et al. (2000). Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science 290, 2105–2110. 10.1126/science.290.5499.2105 [DOI] [PubMed] [Google Scholar]
- Rong W., Qi L., Wang A., Ye X., Du L., Liang H., et al. (2014). The ERF transcription factor TaERF3 promotes tolerance to salt and drought stresses in wheat. Plant Biotechnol. J. 12, 468–479. 10.1111/pbi.12153 [DOI] [PubMed] [Google Scholar]
- Sakuma Y., Liu Q., Dubouzet J. G., Abe H., Shinozaki K., Yamaguchi-Shinozaki K. (2002). DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 290, 998–1009. 10.1006/bbrc.2001.6299 [DOI] [PubMed] [Google Scholar]
- Schmidt R., Mieulet D., Hubberten H., Obata T., Hoefgen R., Fernie A. R., et al. (2013). SALT-RESPONSIVE ERF1 regulates reactive oxygen species–dependent signaling during the initial response to salt stress in rice. Plant Cell 25, 1–18. 10.1105/tpc.113.113068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt R., Schippers J. H. M., Mieulet D., Watanabe M., Hoefgen R., Guiderdoni E., et al. (2014). SALT-RESPONSIVE ERF1 is a negative regulator of grain filling and gibberellin-mediated seedling establishment in rice. Mol. Plant 7, 404–421. 10.1093/mp/sst131 [DOI] [PubMed] [Google Scholar]
- Sharoni A. M., Nuruzzaman M., Satoh K., Shimizu T., Kondoh H., Sasaya T., et al. (2011). Gene structures, classification and expression models of the AP2/EREBP transcription factor family in rice. Plant Cell Physiol. 52, 344–360. 10.1093/pcp/pcq196 [DOI] [PubMed] [Google Scholar]
- Shoji T., Mishima M., Hashimoto T. (2013). Divergent DNA-binding specificities of a group of ETHYLENE RESPONSE FACTOR transcription factors involved in plant defense. Plant Physiol. 162, 977–990. 10.1104/pp.113.217455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soltész A., Smedley M., Vashegyi I., Galiba G., Harwood W., Vágújfalvi A. (2013). Transgenic barley lines prove the involvement of TaCBF14 and TaCBF15 in the cold acclimation process and in frost tolerance. J. Exp. Bot. 64, 1849–1862. 10.1093/jxb/ert050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spence C., Alff E., Johnson C., Ramos C., Donofrio N., Sundaresan V., et al. (2014). Natural rice rhizospheric microbes suppress rice blast infections. BMC Plant Biol. 14:130. 10.1186/1471-2229-14-130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun S., Yu J. P., Chen F., Zhao T. J., Fang X. H., Li Y. Q., et al. (2008). TINY, a dehydration-responsive element (DRE)-binding protein-like transcription factor connecting the DRE- and ethylene-responsive element-mediated signaling pathways in Arabidopsis. J. Biol. Chem. 283, 6261–6271. 10.1074/jbc.M706800200 [DOI] [PubMed] [Google Scholar]
- Vogel M. O., Moore M., König K., Pecher P., Alsharafa K., Lee J., et al. (2014). Fast retrograde signaling in response to high light involves metabolite export, MITOGEN-ACTIVATED PROTEIN KINASE6, and AP2/ERF transcription factors in Arabidopsis. Plant Cell 26, 1151–1165. 10.1105/tpc.113.121061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wan L., Zhang J., Zhang H., Zhang Z., Quan R., Zhou S., et al. (2011). Transcriptional activation of OsDERF1 in OsERF3 and OsAP2-39 negatively modulates ethylene synthesis and drought tolerance in rice. PLoS ONE 6:e25216. 10.1371/journal.pone.0025216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu Z. S., Xia L. Q., Chen M., Cheng X. G., Zhang R. Y., Li L. C., et al. (2007). Isolation and molecular characterization of the Triticum aestivum L. ethylene-responsive factor 1 (TaERF1) that increases multiple stress tolerance. Plant Mol. Biol. 65, 719–732. 10.1007/s11103-007-9237-9 [DOI] [PubMed] [Google Scholar]
- Xue G. P. (2003). The DNA-binding activity of an AP2 transcriptional activator HvCBF2 involved in regulation of low-temperature responsive genes in barley is modulated by temperature. Plant J. 33, 373–383. 10.1046/j.1365-313X.2003.01630.x [DOI] [PubMed] [Google Scholar]
- Xue G. P., Loveridge C. W. (2004). HvDRF1 is involved in abscisic acid-mediated gene regulation in barley and produces two forms of AP2 transcriptional activators, interacting preferably with a CT-rich element. Plant J. 37, 326–339. 10.1046/j.1365-313X.2003.01963.x [DOI] [PubMed] [Google Scholar]
- Yan H. W., Hong L., Zhou Y. Q., Jiang H. Y., Zhu S. W., Fan J., et al. (2013). A genome-wide analysis of the ERF gene family in sorghum. Genet. Mol. Res. 12, 2038–2055. 10.4238/2013.May.13.1 [DOI] [PubMed] [Google Scholar]
- Zander M., Thurow C., Gatz C. (2014). TGA transcription factors activate the salicylic acid-suppressible branch of the ethylene-induced defense program by regulating ORA59 expression. Plant Physiol. 165, 1671–1683. 10.1104/pp.114.243360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H., Zhang J., Quan R., Pan X., Wan L., Huang R. (2013). EAR motif mutation of rice OsERF3 alters the regulation of ethylene biosynthesis and drought tolerance. Planta 237, 1443–1451. 10.1007/s00425-013-1852-x [DOI] [PubMed] [Google Scholar]
- Zhang Z., Yao W., Dong N., Liang H., Liu H., Huang R. (2007). A novel ERF transcription activator in wheat and its induction kinetics after pathogen and hormone treatments. J. Exp. Bot. 58, 2993–3003. 10.1093/jxb/erm151 [DOI] [PubMed] [Google Scholar]
- Zhu X., Qi L., Liu X., Cai S., Xu H., Huang R., et al. (2014). The wheat ethylene response factor transcription factor pathogen-induced ERF1 mediates host responses to both the necrotrophic pathogen Rhizoctonia cerealis and freezing stresses. Plant Physiol. 164, 1499–514. 10.1104/pp.113.229575 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhuang J., Anyia A., Vidmar J., Xiong A. S., Zhang J. (2011a). Discovery and expression assessment of the AP2-like genes in Hordeum vulgare. Acta Physiol. Plant. 33, 1639–1649. 10.1007/s11738-010-0700-x [DOI] [Google Scholar]
- Zhuang J., Chen J. M., Yao Q. H., Xiong F., Sun C. C., Zhou X. R., et al. (2011b). Discovery and expression profile analysis of AP2/ERF family genes from Triticum aestivum. Mol. Biol. Rep. 38, 745–753. 10.1007/s11033-010-0162-7 [DOI] [PubMed] [Google Scholar]