

Abstract
Pigment epithelium-derived factor (PEDF) is a broadly expressed multifunctional member of the serine proteinase inhibitor (serpin) family. This widely studied protein plays critical roles in many physiological and pathophysiological processes, including neuroprotection, angiogenesis, fibrogenesis and inflammation. The present review summarizes the temporal and spatial distribution patterns of PEDF in a variety of developing and adult organs, and discusses its functions in maintaining physiological homoeostasis. The major focus of the present review is to discuss the implication of PEDF in diabetic and hypoxia-induced angiogenesis, and the pathways mediating PEDF's effects under these conditions. Furthermore, the regulatory mechanisms of PEDF expression, function and degradation are also reviewed. Finally, the therapeutic potential of PEDF as an anti-angiogenic drug is briefly summarized.
Keywords: angiogenesis, diabetes, hypoxia, pigment epithelium-derived factor (PEDF), PEDF receptor, serpin, tissue distribution
Abbreviations: AGE, advanced glycation end-product; ATGL, adipose triglyceride lipase; ATRA, all-trans retinoic acid; bFGF, basic fibroblast growth factor; c-FLIP, cellular FLICE-like inhibitory protein; CK2, casein kinase 2; DR, diabetic retinopathy, E, embryonic day; ERK, extracellular-signal-regulated kinase; ER, oestrogen receptor; FasL, Fas ligand; Flt-1, FMS-like tyrosine kinase 1; HIF-1, hypoxia-inducible factor 1; HUVEC, human umbilical vein endothelial cell; IκB, inhibitor of κB kinase; IGF, insulin-like growth factor; JNK, c-Jun N-terminal kinases; KDR, kinase insert domain receptor; LR, laminin receptor; LRP, low-density lipoprotein receptor-related protein; MAPK, mitogen-activated protein kinase; MEK5, MAPK/ERK kinase 5; MMP, matrix metalloproteinase; NFAT, nuclear factor of activated T-cells; NFATc2, nuclear factor of activated T-cells cytoplasmic 2; NF-κB, nuclear factor κB; OIR, oxygen-induced retinopathy; PAI-1, plaminogen activator inhibitor-1; PEDF, pigment epithelium-derived factor; PEDF-R, PEDF receptor; PEDF-tg, transgenic overexpression of PEDF; PDGF, platelet-derived growth factor; PKA, protein kinase A; PPAR-γ, peroxisome proliferator-activated receptor γ; RAR, retinoic acid receptor; RARE, retinoic acid-response element; RCL, reactive centre loop; RPE, retinal pigment epithelial; RXR, retinoid X receptor; serpin, serine proteinase inhibitor; VEGF, vascular endothelial growth factor
INTRODUCTION
Pigment epithelium-derived factor (PEDF), also known as early population doubling level cDNA-1 [1,2], was originally isolated from the conditioned medium of cultured human fetal retinal pigment epithelial cells [3] and found to possess neuronal differentiation properties [4]. It is a highly conserved glycoprotein in mammals [5] and possesses a reactive centre loop (RCL) [5] that is a common structural characteristic of the serine proteinase inhibitor (serpin) family. Cleavage within the RCL by chymotrypsin does not impair PEDF's functions [6], suggesting that PEDF is a non-inhibitory serpin [6,7]. A decade after PEDF's identification, Dawson et al. [8] demonstrated PEDF as a potent endogenous anti-angiogenic factor. This opened a new era for the exploration of PEDF's functions in angiogenic diseases, especially in diabetes [9–11] and cancer [12–14]. PEDF levels were found to decline in angiogenic tissues/organs, such as the vitreous, aqueous humors and retinas from patients with proliferative diabetic retinopathy (DR) and in tumours from cancer patients [10,15–23]. Interestingly, circulating PEDF levels in proliferative DR patients are increased relative to diabetic patients without proliferative DR or non-diabetic controls [24–27]; this observation possibly reflects a systemic compensatory response to angiogenesis in proliferative DR. Thus restoration of PEDF levels within angiogenic sites could be a promising strategy for the treatment of angiogenesis-related diseases.
The PEDF protein plays fundamental roles in organogenesis [12,28–30] and homoeostatic maintenance of adult tissues/organs [12,14,31–33]. Defects or deficiencies of PEDF expression are closely associated with progression of angiogenic diseases [10,15–18,25,27,34,35]. The present review summarizes the temporal and spatial distribution of PEDF in multiple organs during developmental stages and adulthood, and discusses its implication in diabetic and hypoxia-induced angiogenic diseases. In addition, we review PEDF's anti-angiogenic mechanisms under these two types of acquired angiogenic conditions. Finally, the regulatory mechanisms of PEDF expression, function and degradation are reviewed, and the therapeutic potential of PEDF in angiogenic diseases is briefly discussed.
TISSUE DISTRIBUTION AND PHYSIOLOGICAL FUNCTIONS OF PEDF
The mRNA encoding the human PEDF (SERPINF1 mRNA) is expressed in most organs, including the liver, adipose tissue, eye, heart, kidney, ovary, testis, spleen, skeletal muscle, brain and bone [36]. Deficient or defective expression of PEDF leads to abnormal organ development. It was reported that human patients with undetectable circulating PEDF develop osteogenesis imperfecta type VI [28,37]. In mouse, global PEDF deficiency does not affect viability and fertility [12], but leads to pancreatic and prostatic hyperplasia [12], hepatic steatosis [31] and bone defects similar to human patients with osteogenesis imperfecta type VI [29], indicating that PEDF participates in important physiological events in both humans and animals.
PEDF in the liver
SERPINF1 mRNA is highly expressed in human fetal and adult livers [36]. In normal human and mouse livers, hepatocytes are the predominant PEDF-expressing cells [38,39]. In mouse embryonic livers, PEDF protein is detected as early as embryonic day (E) 12.5, with its expression continuously increasing during organogenesis and remaining at high levels in adult livers [39]. PEDF regulates lipid metabolism and maintains physiological homoeostasis in mouse livers. Significant accumulation of neutral lipid and triglyceride is observed in hepatocytes of 1-month-old PEDF-deficient mice, and is continuously increased with age, whereas restoration of PEDF decreases triglyceride content in PEDF-deficient hepatocytes [31]. The regulatory effect of PEDF on lipid metabolism is mediated by adipose triglyceride lipase (ATGL) [40]. PEDF activates ATGL to promote adipose lipolysis, which may contribute to insulin resistance in obese subjects [41] as ATGL-deficient mice do not develop PEDF-induced insulin resistance [40]. PEDF deficiency in the liver also results in hepatic steatosis as observed in adult PEDF-deficient mice even under normal feeding conditions [31]. When fed an alcohol-containing liquid diet, enhanced expression of the fibrotic marker α-smooth muscle actin is detected in the hepatic perisinusoidal space of PEDF-deficient mice compared with wild-type controls [38]. Consistently, in the livers from patients and animal models with hepatic steatosis, PEDF expression is dramatically down-regulated [38,42]. Overexpression of PEDF reverses liver fibrosis [42]. Clinical studies indicate a positive correlation between circulating PEDF levels and insulin resistance in patients who are morbidly obese [43] or diabetic [44–47]. It is known that insulin resistance leads to compromised hepatic glycogenesis and gluconeogenesis in both diabetic patients [48,49] and diabetic animals [50]; however, it is currently unknown whether the insulin resistance-induced aberrant gluconeogenesis and glycogenesis in the liver is due to elevated PEDF levels in obesity and diabetes.
PEDF in the pancreas
The SERPINF1 mRNA is weakly detected in the human pancreas [36]. Higher levels of human PEDF protein are detected in centroacinar cells and islet cells in comparison with most acinar cells and ductal cells [51]. PEDF regulates pancreatic vasculature development, and PEDF deficiency in the pancreas leads to a 2.6-fold increase in microvessel density and more dilated and thicker walled blood vessels [12]. In addition, PEDF deficiency can result in atypical hyperplastic phenotypes in the pancreas, such as enlarged pancreas, more exocrine glands, less differentiated acinar epithelial cells, and increased proliferating epithelial cells [12]. Furthermore, PEDF deficiency makes the pancreas more prone to fibrosis as PEDF-deficient pancreases display significantly increased basal transcription levels of collagen αI, transforming growth factor β1 and platelet-derived growth factor (PDGF) [32]. Enhanced expression of α-smooth muscle actin is also detected in both of the pancreatic vessels and ducts of PEDF-deficient mice [12,32].
The EL-KrasG12D mouse is a transgenic model that spontaneously develops pancreatic lesions and non-invasive cystic papillary neoplasms. At the age of 6–12 months, EL-KrasG12D/PEDF-deficient mice develop more severe acinar ductal metaplasia and cystic papillary neoplasm compared with age-matched EL-KrasG12D mice. Moreover, relative to aged controls, aged EL-KrasG12D/PEDF-deficient mice exhibit more severe pathological changes such as increased ductal metaplasia, augmented papillary neoplasm frequency, enlarged adipocytes, elevated tubular complexes and remarkable multifocal dysplastic changes [52]. Although PEDF is known to affect pancreatic vasculature, morphology and function, PEDF's expression and roles in the pancreas under diabetic conditions remain to be elucidated.
PEDF in the lung
The SERPINF1 mRNA is modestly detected in the human fetal lung, and persists throughout adulthood [36]. PEDF protein is expressed in human pulmonary epithelia, pulmonary fibroblasts/myofibroblasts, alveolar interstitium and bronchoalveolar lavage fluid [53]. PEDF has been suggested to be an important regulator of pulmonary angiogenesis and fibrosis. Lung-derived endothelial cells from PEDF-deficient mice exhibit enhanced migratory capacity and adherent ability compared with cells from wild-type mice, which may be ascribed to decreased cell surface integrins and increased vascular endothelial growth factor (VEGF) secretion [33]. Moreover, both arteriole and venule walls in PEDF-deficient lungs are thicker [33].
Wild-type mice exposed to hyperoxia from postnatal day 5–13 develop impaired alveolarization, which is accompanied by increased PEDF levels mainly in alveolar epithelia [54]. However, hyperoxia-compromised alveolarization is not detected in aged-matched PEDF-deficient mice [54], further implying that increased PEDF levels are the primary causative factor for reduced alveolarization. In idiopathic pulmonary fibrosis that is recently believed to be characterized by impaired angiogenesis instead of enhanced angiogenesis [55], pulmonary PEDF levels are dramatically increased and inversely correlated with VEGF levels and pulmonary microvascular density [53]. It is believed that increased PEDF levels prohibit pulmonary angiogenesis, thus exacerbating idiopathic pulmonary fibrosis [53].
PEDF in the bone
SERPINF1 mRNA is highly expressed in adult human bone marrow [36]. In mice, high levels of PEDF protein are detected in the epiphyseal cartilage (resting, proliferating and upper hypertrophic chondrocytes) and the periosteum (active osteoblasts and chondrocytes), suggesting that PEDF is spatially expressed in areas of endochondral ossification and bone remodelling [56]. Interestingly, PEDF protein is distributed in similar spatial patterns in newborn mice and 5- and 9-week-old mice [56]. The principal cell types that express and secrete PEDF are osteoblasts [57] and chondrocytes [56]. PEDF affects bone formation and resorption by the regulation of osteoblastic and osteoclastic differentiation [58–60]. PEDF promotes osteoblastic differentiation by inducing the expression of osteoblastic genes [58,60]. On the other hand, PEDF impedes osteoclastic differentiation by up-regulating osteoclast-repressive genes [59]. PEDF also affects bone mineralization [29,58,60]. PEDF treatment in human mesenchymal stem cells increases alkaline phosphatase activity that is critical in the mineralization process [60]. Interestingly, mesenchymal stem cells isolated from PEDF-deficient mice show reduced alkaline phosphatase expression [61]. Moreover, both PEDF protein and PEDF-derived peptides (residues 40–64, residues 78–102 and residues 90–114 corresponding to the full-length immature human PEDF sequence) enhance bone mineralization [58,62].
In patients with osteogenesis imperfecta type VI, a disease characterized by low bone mass and reduced bone strength, a homozygous frame-shift mutation in exon 4 or a termination mutation in exon 8 in the SERPINF1 gene is detected, resulting in undetectable levels of PEDF protein in the circulation [28]. Clinically, the circulating PEDF level is a critical biomarker for the screening of this disease [63]. Bogan et al. [29] reported that PEDF-deficient mice exhibited symptoms of osteogenesis imperfecta type VI patients with significant increases in osteoid maturation time, unmineralized bone matrix, mineral/matrix ratio and bone fragility. Notably, patients with heterozygous null mutated SERPINF1 gene possess normal and functional bones, although their circulating PEDF is significantly lower than that of control patients with a fully functional SERPINF1 gene [64].
However, circulating PEDF levels should be interpreted with caution, and not used as the sole biomarker for this disease. Recently, an atypical case of osteogenesis imperfecta type VI was reported in which the patient had normal serum PEDF levels and correct SERPINF1 gene sequence, but a heterozygous single point mutation in the IFITM5 gene [65]. The single point mutation of IFITM5 does not affect its own transcript, instead, it reduces the expression and secretion of PEDF in fibroblasts and osteoblasts isolated from the patient [65]. Why this mutation in IFITM5 affects PEDF expression and secretion and how it leads to osteogenesis imperfecta type VI remains unknown.
The Wnt/β-catenin pathway plays a pivotal role in bone formation and homoeostasis [66]. In bone formation, PEDF and Wnt/β-catenin play similar roles: increased PEDF or activation of Wnt/β-catenin signalling leads to high bone mass, whereas decreased PEDF or attenuation of the Wnt/β-catenin signalling results in low bone mass [29,66]. A recent study reported PEDF as an agonist of the Wnt/β-catenin pathway in human mesenchymal stem cells [61]. Another study also reported that treatment of PEDF in osteocytes induced phosphorylation of glycogen synthase kinase 3β (GSK-3β) and total β-catenin levels [58]. Our group and another group demonstrated that PEDF is an inhibitor of the Wnt/β-catenin pathway in an angiogenic eye model [67] and a wound-healing model [68]. Discrepancy between these studies is noted, yet conclusive explanation is unavailable at this moment and further studies are needed. Notably, in bone-derived cells, whether PEDF's stimulative effect on the Wnt/β-catenin pathway is mediated through low-density lipoprotein receptor-related protein 6 (LRP6) is not established. In addition, PEDF strongly inhibits the expression of sclerostin, an inhibitor of Wnt/β-catenin signalling [69], which might contribute to the activation of Wnt/β-catenin signalling observed in osteocytes [58].
PEDF in the kidney
SERPINF1 mRNA is moderately expressed in human fetal kidneys, and its level declines in adult human kidneys [36]. In human kidneys, PEDF protein is predominantly detected in tubules and with less intensity in glomeruli [70]. In mouse kidneys, PEDF protein localizes in the renal vasculature, interstitial spaces, glomeruli and tubules [71]. In rat kidneys, PEDF protein levels continuously increase with age [72]. In neonatal rats, PEDF protein is restricted to glomerular and capillary mesenchyme and endothelial cells of the nephrogenic zones [72]. By postnatal day 7, PEDF is also detected in the Bowman's capsules of maturing glomeruli locating at the inner cortical region [72]. The first detection of PEDF in podocytes close to the innermost region is at postnatal day 14 [72]. In adult rats, high levels of PEDF are predominantly detected in glomerular podocytes and endothelial cells, and vascular endothelial cells [72,73]. It is noteworthy that PEDF protein expression persists in all types of blood vessels in the kidneys throughout all fetal stages and adulthood. The significance of PEDF in renal vascular development has been well demonstrated, as renal microvessel density of PEDF-deficient mice is greatly increased compared with that in wild-type controls [12,14]. In a diabetic rat model, renal PEDF levels are significantly decreased [73], although PEDF levels in the kidneys of diabetic patients have not been measured. Clinical studies have shown that PEDF levels are closely correlated with vascular dysfunction-associated renal diseases. In patients with chronic kidney disease, plasma PEDF levels are significantly increased compared with those of patients without chronic kidney disease [74]. Moreover, in both Type 1 and Type 2 diabetes, serum PEDF levels correlate positively with serum creatinine concentrations [44,45,47], and inversely with glomerular filtration rate [44,47,75]. It is still unclear what roles PEDF plays in these renal diseases and whether its roles are related to its effects in the vasculature.
PEDF in the eye
The PEDF protein is detected in the human eye from early embryonic stages [76]. In very early stage of human fetal development, PEDF is restricted to the granules of retinal pigment epithelial (RPE) cells, the ganglion cell layer and sporadic cells in the neuroblastic layer [76]. In later human fetal stages, PEDF protein is found in RPE cells, corneal epithelia and endothelia, ciliary body non-pigmented epithelia and muscle, horizontal cells in the outer part of the inner nuclear layer, differentiating photoreceptors and apical cytoplasm of differentiating cones [76]. In adult human retinas, PEDF protein localizes in rods and cones, the inner nuclear layer, ganglion cell layer and inner plexiform layer. The choroid, corneal epithelia and endothelia, ciliary body non-pigmented epithelia and muscle, RPE cells and photoreceptors are also PEDF-positive [76]. In the macula of monkeys at all ages, the SERPINF1 mRNA is detected in the ganglion cell layer and RPE cells, with the highest level present at the fovea [77]. Abundant PEDF protein is also found in the monkey interphotoreceptor matrix surrounding rod and cone outer segments, with lower levels detected in the vitreous and aqueous humors due to the polarized and directional secretion of PEDF towards the neural retina by RPE cells [78]. A number of studies have reported very high levels of PEDF expression in RPE cells [76–78], and polarized PEDF secretion is closely associated with the polarization and maturation of RPE cells [78–80]. As high levels of PEDF are found in the interphotoreceptor matrix, vitreous and aqueous humors in species such as the cow [81–83] and human [76,83], purification from these sources was a common way to obtain PEDF before the eukaryotic expression system of PEDF was established. In mouse embryonic stages, PEDF protein is firstly detected at E14.5 in the inner plexiform layer and RPE cells, followed by detection at E18.5 in the ganglion cell layer, inner nuclear layer, ciliary body and choroid [84]. In rats, Serpinf1 mRNA is localized in the lens epithelia, RPE cells, ciliary epithelia, retinal ganglion cells and sporadic cells of the inner nuclear layer, and PEDF protein is identified in RPE cells, corneal epithelia and endothelia, ciliary epithelia, the nerve fibre layer, ganglion cell layer and inner and outer plexiform [85].
PEDF affects retinal vasculature via its anti-angiogenic properties. Compared with wild-type controls, the retinas of PEDF-deficient mice display the following changes: a faster vascular expansion rate from postnatal day 3 to 7, significantly increased deep vascular plexi at postnatal day 10 and a slightly increased endothelial cell/pericyte ratio at postnatal day 21 [30]. Adult PEDF-deficient mice at the age of 3 months also exhibit a 5.2-fold increase of retinal microvessel density and malpositioned vessels [12]. In addition, PEDF deficiency results in more severe vessel obliteration in the oxygen-induced retinopathy (OIR) model [30]. In a mouse model of DR, loss of PEDF results in a 2-fold increase of acellular capillaries [86]. In contrast, genetic overexpression of PEDF in mice significantly suppresses retinal neovascularization in the OIR model [87]. Similarly, adeno-associated virus overexpressing PEDF dramatically represses retinal neovascularization in an insulin-like growth factor 1 (IGF-1) transgenic mouse model [88]. In addition to the retina, PEDF and PEDF-derived peptides have also been demonstrated to strongly inhibit choroidal neovascularization [87,89–91] and corneal neovascularization [8,92–94]. Taken together, these studies indicate that PEDF is indeed a potent endogenous angiogenic inhibitor in the eye.
Lack of PEDF has subtle impacts on mouse retinal development, differentiation and function under normal conditions [30]. Yet PEDF plays an important supporting role in promoting retinal differentiation and maintaining retinal function. PEDF exhibits a pro-cone trophic action in a chicken rosetted retinal spheroids model by inducing cone opsin expression and decreasing rod numbers [95]. PEDF also promotes the differentiation and maturation of RPE cells [96]. In cultured heterogenic cell clusters from rat retinas, the RPE cell population is dramatically increased by PEDF accompanied with the following changes: increase in size, loss of vacuolization, acquisition of a more epithelium-like appearance, enhanced cell adhesion property and increased mature pigment granules [96]. Moreover, overexpression of PEDF causes bone marrow stromal cells to differentiate to RPE cells [97]. In addition, the neurotrophic function of PEDF promotes the survival of retinal neurons [98,99], retinal progenitor cells [80], RPE cells [100] and photoreceptors [98,100–103]. Notably, the cytoprotective effect of PEDF in retinal cells is reported to be mediated via the PEDF receptor (PEDF-R) [104]. Thus PEDF is indeed a demonstrable retinal protective factor.
IMPLICATION OF PEDF IN DIABETIC AND HYPOXIA-INDUCED ANGIOGENIC DISEASES
Angiogenesis is involved in many physiological and pathological processes, and is stimulated by angiogenic factors [105]. Angiogenic stages are composed of endothelial activation, sprouting, regression and maturation [106]. PEDF exerts its anti-angiogenic effects primarily by targeting endothelial cells. PEDF's functions in tumour angiogenesis have been reviewed by Hoshina et al. [107], Manolo et al. [108] and Becerra and Notario [109]. Rychlic et al. [110] and Liu et al. [111] have reviewed PEDF's roles in cardiovascular angiogenesis. In the present review, we focus on PEDF's functions in two major types of acquired angiogenesis: diabetic angiogenesis and hypoxia-induced angiogenesis.
PEDF-Rs in angiogenic regulation
PEDF-R (also known as ATGL/desnutrin/iPLA2-ζ/TTS2.2), encoded by the PNPLA2 gene in humans, is the first identified receptor for PEDF [112]. PEDF-R is expressed in human ocular tissues (fetal and adult RPE layers and retinas) [112], human Y-79 cells [112,113], RPE cells (including ARPE-19 and hTERT) [112,114], 661W (a mouse photoreceptor cell line) [115], human adipose tissue and 3T3-L1 cell line [112], bovine retina and RPE cells [114,116], rat R28 (a photoreceptor precursor cell line) [112], and a RGC-derived cell line [112,114,117]. PEDF-R mediates multiple activities of PEDF [40,104,118–123]. There is no direct evidence showing that PEDF's anti-angiogenic activities are mediated through PEDF-R in endothelial cells; however, it has been demonstrated that PEDF induces the expression of apoptotic Fas ligand (FasL) by regulating nuclear factor κB (NF-κB) in endothelial cells [124], and that the regulation of NF-κB by PEDF is PEDF-R-dependent [118]; thus it may be possible that PEDF exerts its anti-angiogenic activities through PEDF-R in endothelial cells.
Laminin receptor (LR) is another receptor identified to mediate PEDF's anti-angiogenic effects [125]. Knockdown of LR attenuates PEDF-induced endothelial cell apoptosis and migration [125]. A triple phosphomimetic mutant of PEDF, possessing more potent anti-proliferative, anti-migratory and pro-apoptotic effects in endothelial cells, exhibits higher binding affinity to LR, but not to PEDF-R compared with wild-type PEDF, suggesting that enhanced anti-angiogenic properties of the PEDF triple phosphomimetic mutant are mediated through LR [126].
F1-ATP synthase was also found to convey PEDF's anti-angiogenic activities. In endothelial cells, PEDF binds to the β-subunit of the F1-ATP synthase in a specific, reversible and saturable manner (Kd=1.51 nM in real-time surface plasmon resonance assay, Kd=3.04–4.97 nM in bovine retinal endothelial cells), resulting in a reduced F1-ATP synthesis activity and subsequently lower ATP levels, which impedes the energy supply for angiogenic events in endothelial cells [127].
Some serpins are known to bind to LRPs: SERPINE1 [128], SERPINE2 [129] and SERPINA1 [130] bind to LRP1; SERPINE1 [128] and SERPINA3 [130] bind to LRP2; and SERPINA3K binds to LRP6 [131]. Our previous study also demonstrated that PEDF binds to LRP6 [67], a co-receptor of the Wnt/β-catenin pathway that plays important pathogenic roles in retinal inflammation, neovascularization and vascular leakage in angiogenic eye models [67,87,131–133]. PEDF-deficient OIR retinas show enhanced activation of the Wnt/β-catenin pathway compared with wild-type OIR retinas [67], whereas transgenic overexpression of PEDF in mouse attenuates the Wnt/β-catenin signalling and neovascularization in the OIR and laser-induced choroidal neovascularization models [87].
An additional group of atypical non-canonical ‘receptors’, also known as extracellular proteins, has been reported to affect PEDF's angiostatic functions. PEDF binds to collagen I, which might modulate the integrin–collagen I interaction, thus affecting endothelial cell adhesion and docking [134,135]. In addition, PEDF binds to collagen II [134], collagen III [136] and glycoaminoglycans [137–140], which are also very likely to facilitate PEDF's anti-angiogenic functions.
Role of PEDF in diabetic angiogenesis
DR and diabetic nephropathy are diabetes-related angiogenic diseases. Proliferative DR is characterized by increased formation of immature vessels in the retina and vitreous, which can ultimately lead to intra-retinal or pre-retinal haemorrhage [141]. In the kidneys from 50-day-old diabetic rats, average capillary areas per glomerulus, capillary length and capillary numbers are significantly increased [142]. Diabetic angiogenesis is induced by factors including hyperglycemia, hypoxia, imbalanced redox states, elevated non-enzymatic glycosylation and nitration, etc. [141]. Notably, compared with either non-diabetic individuals or diabetic patients with non-proliferative DR, PEDF levels are markedly decreased in the vitreous, retinas and aqueous humors of diabetic patients with proliferative DR [10,15,16,18,143–149]. PEDF levels are also decreased in the kidneys from Type 1 diabetic mice and rats [47,71,73,150]. In contrast, PEDF levels in the circulation are significantly increased in diabetic patients relative to non-diabetic patients [24,25,27,44,45,47,151], and positively correlated with the severity of diabetic complications [24,27,44,47,151].
PEDF counteracts angiogenesis in both proliferative DR [152–154] and diabetic nephropathy [155]. Compared with other endogenous anti-angiogenic factors such as thrombospondin, endostatin and angiostatin, PEDF is more effective in inhibiting endothelial cell migration [8]. Activation of the p38 mitogen-activated protein kinase (p38 MAPK) mediates PEDF's anti-migratory effect in bovine aorta endothelia cells [126]. The receptor that transmits PEDF's effect to p38 MAPK was reported to be LR [126]. Matrix metalloproteinases 2/9 (MMP-2/9) promote angiogenesis by degrading the extracellular matrix, thus mobilizing endothelial cells [156]. PEDF down-regulates the activities of MMP-2/9 in the aqueous humor of a proliferative DR model [88].
In addition, PEDF inhibits the proliferation of endothelial cells by regulating the MAPK/extracellular-signal-regulated kinase (ERK) pathways [126,153,157–159]. The Wnt/β-catenin pathway is activated in diabetic angiogenesis [160] and promotes endothelial cell migration and proliferation via its targets VEGF [161] and MMPs [162–164]. As demonstrated by ligand-binding assays and co-immunoprecipitation, PEDF binds to LRP6, a co-receptor in the Wnt/β-catenin pathway, and suppresses Wnt signalling in ARPE-19 and Müller cells [67].
Quenching oxidative stress is another mechanism by which PEDF inhibits diabetic angiogenesis. PEDF suppresses nicotinamide adenine dinucleotide phosphate oxidase activity in the retinas of diabetic rats [152] and rats with retinal hyper-permeability induced by advanced glycation end-products (AGEs) [165]. Similarly, in ex vivo endothelial cell cultures, PEDF directly suppresses reactive oxygen species generation by inhibiting NADPH oxidase activity elicited by AGEs [165,166], tumour necrosis factor α [167] and angiotensin II [168]. Moreover, in bovine retinal endothelial cells, PEDF activates peroxisome-proliferator-activated receptor γ (PPAR-γ), which then up-regulates uncoupling protein 2 and subsequently decreases mitochondria-derived reactive oxygen species [169].
In addition to endothelial cells, pericyte loss has been well established to contribute to the angiogenic progression in DR [170]. PEDF protects pericytes against apoptosis induced by high glucose, H2O2, AGEs and oxidized low-density lipoprotein through its antioxidant and anti-inflammatory activities [171–173]. In addition, survival and proliferation of pericytes require endothelium-derived PDGF-BB [174]. PEDF promotes the proliferation of pericytes via up-regulating the expression of PDGF-BB [175].
Role of PEDF in hypoxia-induced angiogenesis
In hypoxic disease states, the activation of hypoxia-inducible factor 1 (HIF-1) precedes the occurrence of angiogenesis. Hypoxia stabilizes HIF-1α to form a heterodimer with HIF-1β [176], which then activates the transcription of pro-angiogenic genes [177,178]. OIR is a widely used ischaemia-induced retinal angiogenesis model [179]. In OIR mouse retinas, HIF-1 and VEGF are up-regulated [180] and trigger the occurrence and progression of retinal angiogenesis that peaks between postnatal day 17–21 [179]. In contrast, PEDF is down-regulated in the choroid and RPE cells of this model from postnatal day 13–17 [181]. Similarly, in the retinas of OIR rats, PEDF levels are decreased with its lowest levels detected at postnatal day 16, which is coincidental with the peak expression of VEGF in the retinas of the same model [11]. To provide in vivo evidence of PEDF's anti-angiogenic activities in the OIR model, we generated PEDF transgenic (PEDF-tg) mice that overexpressed PEDF. Compared with wild-type OIR retinas, PEDF-tg OIR retinas display a significant reduction in retinal neovasculature [87]. In contrast, PEDF-deficient OIR retinas exhibit more prominent VEGF overexpression [30,67] and more severe angiogenesis relative to wild-type OIR retinas [30]. Laser-induced choroid neovascularization is another hypoxia-triggered angiogenic model [182]. Our group and other groups found that overexpression or delivery of PEDF or PEDF-derived peptide dramatically suppressed choroidal neovascularization [87,90,91,183,184].
PEDF suppresses hypoxia-induced angiogenesis by either directly targeting HIF-1 or regulating the expression or the signalling cascades of HIF-1's target genes. It is reported that PEDF blocks HIF-1 nuclear translocation and represses VEGF promoter activity under hypoxic conditions in retinal capillary endothelial cells [153]. However, the mechanism by which PEDF inhibits HIF-1 nuclear translocation remains unknown. Regulatory effects of PEDF on the expression of HIF-1 target genes and their signalling are summarized below.
VEGF is a target gene of HIF-1, with a hypoxia-response element located at its promoter region [185,186]. Our group and others demonstrated that PEDF decreased the expression of hypoxia-induced VEGF in retinal capillary endothelial cells, Müller cells and an angiogenic eye model [93,153]. VEGF/kinase insert domain receptor (KDR) is a crucial angiogenic pathway, and PEDF was reported to disrupt this pathway. Our previous study has shown that PEDF competes with VEGF for binding to KDR in retinal capillary endothelial cells, muting the angiogenic events of the VEGF/KDR pathway [153]. In addition, VEGF/FMS-like tyrosine kinase 1 (Flt-1) signalling was demonstrated to be essential for the survival of human dermal microvascular endothelial cells [187]. PEDF treatment activates γ-secretase in bovine retinal microvascular endothelial cells, which then triggers the cleavage of Flt-1 and mutes the phosphorylation of Flt-1 [188], leading to suppressed VEGF/Flt-1 signalling which is required for the viability [187] and tube formation ability of endothelial cells [188]. In addition, increased concentrations of the extracellular domain of Flt-1 trap and sequester VEGF in the extracellular matrix [188,189], resulting in less available VEGF and subsequent attenuated VEGF/KDR signalling.
Basic fibroblast growth factor (bFGF) is a potent angiogenic factor regulated by HIF-1 [190,191]. bFGF activates nuclear factor of activated T-cells (NFAT), an angiogenic transcription factor, to promote angiogenic events in human microvascular endothelial cells and human umbilical vein endothelial cells (HUVECs) [192]. In response to PEDF treatment, the association between c-Jun N-terminal kinase 2 (JNK-2) and NFAT cytoplasmic 2 (NFATc2) is significantly increased in endothelial cells, which leads to elevated cytoplasmic retention of NFATc2 and decreased nuclear levels of NFAT to promote angiogenesis [192]. In addition, bFGF induces bovine aorta endothelial cell migration, whereas PEDF treatment activates p38 to counteract bFGF-stimulated endothelial cell migration [126].
MMP-2 expression is modulated by HIF-1 in human somatic endothelial cells and HUVECs [193–195]. In addition, hypoxia affects the activation [196,197] and expression [197,198] of MMP-9 in mouse cerebral microvasculature, in retinal cells and in HUVECs. PEDF was demonstrated to suppress the expression and activities of MMP-2/9 in the retinas with severe neovascularization [88]. Interestingly, PEDF's regulatory effect on MMP-2/9 expression is also reported in a spontaneous pancreatic carcinoma model [52].
Plaminogen activator inhibitor-1 (PAI-1) is another angiogenic factor under the regulation of HIF-1, with hypoxia-response elements located at its promoter region [199–202]. PEDF suppresses PAI-1 transcription in HUVECs [203]. Interestingly, even under normal conditions, PEDF significantly decreases mRNA levels of PAI-1 in HUVECs [203]. Furthermore, the activity of plasma-derived PAI-1 is also substantially reduced by PEDF in rats [204].
Common mechanisms for the anti-angiogenic activities of PEDF in diabetic and hypoxic conditions
In addition to the anti-angiogenic pathways mentioned above, a number of studies have demonstrated that under diabetic and hypoxic conditions, the common mechanisms by which PEDF inhibits angiogenesis are via promotion of endothelial cell death [9,124–126,159,192,205–214]. Both full-length PEDF [9,124,126,159,192,205–211] and PEDF-derived peptides [125,212–214] were reported to induce apoptosis of endothelial cells in in vitro culture or in angiogenic animal models. The apoptotic pathways in endothelial cells stimulated by PEDF are summarized as below (Figure 1).
Figure 1. Molecular mechanisms for the anti-angiogenic activities of PEDF.
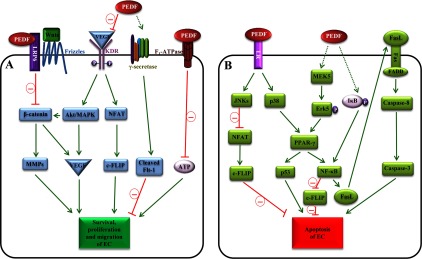
(A) PEDF blocks the survival, proliferation and migration of endothelial cells (ECs). PEDF binds to LRP6, an essential co-receptor of the Wnt/β-catenin pathway, which attenuates β-catenin nuclear translocation and subsequently the expression of angiogenic genes such as VEGF and MMPs. PEDF competes with VEGF for binding to KDR, inhibiting the downstream angiogenic Akt/MAPK and NFAT/c-FLIP pathways. The VEGF/Flt-1 signalling is critical for EC survival. PEDF elicits γ-secretase to cleave Flt-1, resulting in suppressed survival signalling of ECs. PEDF also binds to the β-subunit of F1-ATP synthase and inhibits the production of ATP, which is indispensable for EC angiogenic events. (B) PEDF promotes the apoptosis of ECs. PEDF activates JNKs through LR. Activated JNKs in the cytosol have higher binding affinity to NFATc2, resulting in cytosolic retention and thus less NFAT in the nucleus as a transcription factor that is required to promote the expression of an anti-apoptotic factor c-FLIP. PEDF also binds to LR to activate p38, which leads to activation of PPAR-γ. PEDF also activates PPAR-γ via the mediation of MEK5/Erk5. Activated PPAR-γ then stimulates p53 expression to induce EC apoptosis. NF-κB is another target gene of PPAR-γ. Increased NF-κB is able to up-regulate FasL expression to promote EC apoptosis via the Fas/FasL pathway. Moreover, NF-κB displaces NFAT and binds to the promoter of the c-FLIP gene, resulting in reduced levels of c-FLIP. Interestingly, PEDF also stimulates NF-κB via a PPAR-γ-independent pathway, i.e. by inducing IκB degradation. Red lines represent inhibition of pathways whereas continuous green arrows show activation of pathways. Broken green arrows illustrate activations of pathways whose detailed signalling cascades are currently not fully understood. Erk5, orphan MAPK; UCP-2, uncoupling protein 2.
The Fas/FasL interaction leads to programmed cell death [215]. Fas is constitutively expressed at low levels on the cell surface of quiescent endothelial cells. In angiogenic states, high concentrations of angiogenic stimulators increase cell surface presentation of Fas in activated endothelial cells [209,216]. PEDF induces the transcription and cell surface display of FasL in human dermal microvascular endothelial cells [209], leading to enhanced Fas/FasL interaction and subsequent activation of caspase 8-dependent apoptotic signalling. This action of PEDF on FasL has been suggested to be through activation of NF-κB [124]. In a bFGF-induced corneal neovascularization model, PEDF inhibits new vessel growth in wild-type mice, but not in FasL-deficient or Fas-deficient mice [209], further demonstrating that Fas/FasL mediates PEDF's anti-angiogenic effects. However, Fas/FasL/caspase-8 is not the only pathway to mediate PEDF's apoptotic effects in endothelial cells. It has been reported that deficiency of FasL or Fas does not attenuate the inhibitory effect of PEDF on hypoxia-induced angiogenesis in OIR retinas [217]. Moreover, a FasL-neutralizing antibody does not abolish PEDF-induced apoptosis in HUVECs [208], implying that alternative pathway(s) is(are) mediating PEDF's pro-apoptotic effects under these circumstances.
Another PEDF-elicited apoptotic pathway is the LR/JNK/NFAT/cellular FLICE-like inhibitory protein (c-FLIP)/caspase 8 signalling pathway. Both wild-type PEDF [126,192] and a triple phosphomimetic mutant of PEDF [126] activate JNKs in bovine aortic endothelial cells and HUVECs. The receptor that transmits PEDF's effect to JNKs is probably LR, as activation of JNKs is positively correlated with binding affinities of PEDF to LR, but not with those to PEDF-R [126]. Activated JNKs directly bind to NFATc2, resulting in cytoplasmic retention of NFATc2. Cytoplasmic retention of NFATc2 compromises the availability of NFAT in the nucleus as a transcription factor, which in turn leads to insufficient transcription of c-FLIP. As c-FLIP is an endogenous inhibitor of caspase 8, attenuated expression of c-FLIP results in enhanced activity of caspase 8 and subsequent endothelial cell apoptosis [192].
The LR/p38/PPAR-γ/apoptosis pathway also mediates PEDF's pro-apoptotic effects in endothelial cells. PEDF binds to LR [126] to activate p38 in bovine aortic endothelial cells and HUVECs [126,208,210,218]. Ho et al. [208,210] demonstrated that PEDF-stimulated p38 induces the expression and activity of PPAR-γ in HUVECs. Activation of PPAR-γ then induces p53 expression in HUVECs [208,210] to induce endothelial cell apoptosis [208,210]. In addition, PEDF also activates PPAR-γ via the MAPK/ERK kinase 5 (MEK5)/ERK5/PPAR-γ pathway [159]. PEDF induces Erk5 phosphorylation via MEK5, which then activates PPAR-γ [159]. PEDF-stimulated PPAR-γ not only induces the expression of p53 in HUVECs [208,210], but also promotes the expression and activity of NF-κB in human dermal capillary endothelial cells to suppress angiogenesis [159]. Interestingly, PPAR-γ-independent activation of NF-κB by PEDF was also reported: PEDF can induce the phosphorylation and degradation of inhibitor of κB kinase (IκB) in HUVECs [124]. As a result, activated NF-κB binds to the FasL promoter and initiates FasL transcription [124]. Moreover, PEDF-elicited NF-κB also displaces NFAT and binds to the promoter region of c-FLIP, resulting in decreased transcription of c-FLIP and enhanced endothelial cell apoptosis [124].
REGULATION OF PEDF EXPRESSION, FUNCTION AND DEGRADATION
PEDF levels decline in angiogenic tissues/organs [10,15–18,34,35]. In contrast, circulating PEDF levels increase in both Type 1 and Type 2 diabetes relative to patients without diabetes [24–27,44,45,47], which might be indicative of a systemic compensatory response to the decreased expression of PEDF in angiogenic tissues/organs. Understanding how PEDF levels are regulated may shed light on its roles in physiological and pathophysiological conditions. In the present review we discuss documented pathways that regulate PEDF levels (Figure 2), however, other mechanisms may also participate in the regulation of PEDF expression, function and degradation.
Figure 2. Regulation of PEDF expression, function and degradation.
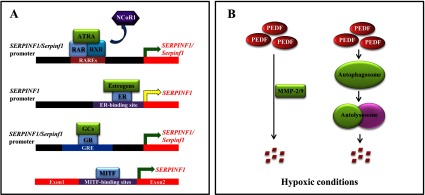
(A) Transcriptional regulation of human SERPINF1/rodent Serpinf1. Six RAREs are located at −1000 to −1 bp of the SERPINF1/Serpinf1 promoter region. In the presence of ATRA, RAR/RXR heterodimers bind to RAREs, dissociate from NCoR1, and recruit co-activators to activate SERPINF1/Serpinf1 transcription. In the absence of agonists, RAR/RXR heterodimers associate with NCoR1, which suppresses SERPINF1/Serpinf1 transcription. At least one ER-binding site is located at −864/+63 bp of the SERPINF1 promoter. Oestrogens induce ERs to form either heterodimers or homodimers to bind to the ER-binding site, triggering/suppressing SERPINF1 transcription depending on cell types and tissues. In addition, at least one GRE is found at −1721/+38 bp of the SERPINF1/Serpinf1 promoter. GCs bind to GRs and promote GR nuclear translocation, which then initiates SERPINF1/Serpinf1 transcription via the promoter region GRE. Three micropthalmia-associated transcription factor (MITF)-binding regions are identified within the first intron of the SERPINF1 gene. MITF binds to the MITF-binding sites and up-regulates the transcription of SERPINF1. (B) Down-regulation of PEDF by hypoxia. Under hypoxic conditions, expression and activities of MMP-2/9 are increased, which promote the degradation of PEDF protein. In addition, a HIF-1-independent pathway to degrade PEDF was also reported. Hypoxia stimulates the autophagosome to down-regulate PEDF levels. Continuous green arrows represent gene transcription activation, whereas broken yellow arrows illustrate either activation or suppression of gene transcription depending on context. GCs, glucocorticoid/glucocorticoid analogues; GR, glucocorticoid receptor; GRE, glucocorticoid response element; NCoR1, nuclear receptor co-repressor 1.
Transcriptional regulation
Transcription of human SERPINF1/rodent Serpinf1 genes is regulated by the binding of transcription factors to corresponding promoters. At −1000 to −1 bp of the human SERPINF1 promoter region, six retinoic acid-response elements (RAREs) have been identified [219]. In the presence of all-trans retinoic acid (ATRA), heterodimers formed by retinoic acid receptors (RARs) and retinoid X receptors (RXRs) bind to RAREs, dissociate from co-repressors and recruit co-activators [219], leading to active transcription of the SERPINF1/Serpinf1 gene. ATRA-induced expression of the SERPINF1/Serpinf1 gene was reported in human retinal pigment epithelia and cancer cells [220,221], bovine retinal endothelial cells [220], mouse and rat neurons and neuron-derived cancer cells [220]. On the other hand, when agonists are absent, the nuclear receptor co-repressor 1 associates with the RAR/RXR heterodimer and recruits repressive complex to the RAREs, resulting in reduced transcription of the SERPINF1 gene [219]. In addition, the SERPINF1 promoter contains at least one oestrogen receptor (ER)-binding site located at −864/+63 bp [222,223]. ER has two isoforms that are recognized as ERα and ERβ. In the presence of agonists, ERs form either heterodimers or homodimers and then translocate into the nucleus, functioning as transcription factors. Oestrogen agonists, such as 17β-oestradiol and ginsenoside Rb1, significantly induce the transcription of SERPINF1 in human Müller cells [224] and HUVECs [223]. 17β-Oestradiol-induced suppression of SERPINF1 transcription was also reported in human ovarian epithelial cells [222], human endometrial cells [225] and rhesus retinal capillary endothelial cells [226]. Activation or repression of SERPINF1 transcription probably depends on differential binding of ER heterodimers/homodimers to the promoter in different cell types or tissues. Corticosteroids and analogues, such as dexamethasone or triamcinolone, also regulate SERPINF1/Serpinf1 expression via the glucocorticoid-response element located at the promoter region. It has been reported that there is at least one dexamethasone-binding site located at −1721/+38 bp of the human SERPINF1 gene [227]. Up-regulation of SERPINF1/Serpinf1 by corticosteriod analogues has been reported in a variety of cell types: HUVECs [228], primary human trabecular meshwork cells [229], human ARPE-19 cells [228], mouse 3T3-L1 cells [227], mouse Müller glial cells [220] and rat glioma cells [220]. Morever, three micropthalmia-associated transcription factor-binding regions are identified within the first intron of the human SERPINF1 gene [230]. Up-regulation of PEDF by microthalmia-associated transcription factor is observed in human melanoma cell lines [230], primary melanocytes [230] and human RPE cells [231].
Post-translational modification
Mature PEDF protein can undergo post-translational modifications such as N-terminal pyroglutamate blocking [232], phosphorylation [233,234] and glycosylation [81,232,235]. PEDF contains a conserved glycosylation motif N-X-L across species [5]. An N-linked glycosylation site is found in both human and bovine PEDF [81,232,235]. However, the role of N-linked glycosylation in PEDF function is yet to be investigated. It is reported that human PEDF protein could be phosphorylated by casein kinase 2 (CK2) and protein kinase A (PKA) [233]. CK2 phosphorylates PEDF at Ser24 and Ser114, whereas PKA phosphorylates Ser227 to a lesser extent [233]. The PEDF mutant S24E/S114E mimicking the CK2 phosphorylation exhibits enhanced anti-angiogenic effects, but decreased neurotrophic activities. In addition, the PEDF mutant S227E, which mimics the PKA phosphorylation, displays attenuated anti-angiogenic effects and intact neurotrophic activities [233,234]. PEDF isoforms with different molecular masses have been identified [83,235–237]. These isoforms display differential biological activities [235,236]. The machinery for PEDF isoform production and the mechanisms for the different activities of these isoforms of PEDF are presently unknown.
Secretory regulation
The physiological concentration of PEDF in the human plasma is approximately 5 μg/ml (100 nM) [232]. The C-terminus (residues 415–418), RCL (residues 373–380), and hydrophobic core (Asn391–Thr403) corresponding to β-sheet B were found to be essential for human PEDF secretion [238]. The C-terminus truncated PEDF that lacks Arg416-Gly-Pro418 is secreted to a lesser extent compared with wild-type PEDF, and removal of Pro415-Arg-Gly-Pro418 completely abolishes PEDF secretion [238]. The transcription levels of these SERPINF1 truncated mutants are similar to that of wild-type SERPINF1, but the PEDF mutant protein lacking Pro415 is only detected in the endoplasmic reticulum, whereas wild-type PEDF protein localizes to both the endoplasmic reticulum and the Golgi, suggesting that Pro415 at the C-terminus is essential for transporting PEDF protein from the endoplasmic reticulum to the Golgi [238]. Removal of RCL (∆373–380) also results in insufficient secretion due to impaired PEDF protein transport from the endoplasmic reticulum to the Golgi [238]. In addition, replacement of Gly376 and Leu377 with alanine within the RCL completely abolishes PEDF secretion, suggesting that the RCL plays an important role in the interaction of PEDF with the quality control mechinary within the endoplasmic reticulum [238]. Moreover, amino acid mutations of the hydrophobic core of β-sheet B results in no PEDF secretion, which is also due to compromised protein transport from the endoplasmic reticulum to the Golgi [238]. Interestingly, secretion of PEDF is predominantly apical in polarized RPE cells, which is an indicator of RPE polarization and full function [78–80]. Polarized and directional secretion of PEDF towards the neural retina by RPE cells results in abundant accumulation of PEDF in the interphotoreceptor matrix, and vitreous and aqueous humors [76,81–83]. The mechanism for polarized PEDF secretion in RPE cells is unknown.
Degradative regulation
PEDF levels decrease in many disease conditions relative to non-disease conditions. Two degradative pathways regulating PEDF levels have been identified. Under hypoxia, the transcription factor HIF-1 is activated and induces the expression and activities of MMP-2/9 [193–198]. MMP-2/9 have been reported to proteolyze PEDF in a variety of cells including retinal cells [38,181,239,240]. A HIF-1-independent pathway to degrade PEDF was also reported, where hypoxia stimulated the autophagosome to down-regulate PEDF levels [241].
POTENTIAL CLNICAL APPLICATIONS OF PEDF AS AN ANGIOGENIC INHIBITOR
On the basis of clinical correlations between circulating PEDF levels and diabetic angiogenesis, PEDF has been proposed as a biomarker for the assessment of angiogenesis progression in diabetic patients [24,27,44,47]. A Phase I clinical trial identified that PEDF had therapeutic effects in wet age-related macular degeneration, and the effect of PEDF as an anti-angiogenic agent in this model was promising [242]. Still, the potential of PEDF as an anti-angiogenic drug awaits further support from future clinical trials. In the present review, we provide a summary of possible strategies that may help explore the therapeutic potential of PEDF as an angiostatic factor.
Prior studies suggest that modification of PEDF protein may be a feasible therapeutic strategy. PEDF phosphomimetic mutants S24E/S114E [233] and S24E/S114E/S227E [234] exhibit enhanced anti-angiogenic effects compared with wild-type PEDF. The mature human PEDF protein (not including the 20-amino-acid signal peptide) contains 36 serine, 10 tyrosine and 29 threonine residues that can potentially be modified by phosphomimetics. Whether phosphomimetics at those sites will enhance the anti-angiogenic effects of PEDF in humans remains to be investigated. It was reported that binding to collagen I [134,135] potentiates PEDF's anti-angiogenic activities. PEDF is also capable of binding to collagen II [134], collagen III [136] and glycoaminoglycans [137–140]. Therefore modifications to enhance PEDF binding affinities to collagens and glycosoaminoglycans may be an additional strategy to improve PEDF's anti-angiogenic activity.
Development of PEDF isoforms or variants with enhanced anti-angiogenic properties is another alternative method. PEDF isoforms are found to exhibit different activities [235,236]. PEDF isoforms can result from different post-translational modifications, such as glycosylation by different oligosaccharides [232,235,243], N-terminal pyroglutamate blockage [232,243] and multiple phosphorylations [233,234]. Generation of PEDF isoforms via post-translational modifications provides another way to enhance the anti-angiogenic effects of PEDF.
Another strategy is to develop PEDF-derived peptides that possess potent anti-angiogenic activities. Peptide-based therapeutics are expected to be superior to full-length protein-based medicine in the following aspects: improved water solubility, higher production yield and purity by chemical synthesis, and lower immunogenicity [244]. Nevertheless, peptide-based medicines have their own disadvantages such as short half-lives. As summarized in Table 1, many functional PEDF-derived peptides possess anti-angiogenic activities, whereas the rest display biological activities that might be beneficial for the organism under diabetic and hypoxia-induced angiogenesis.
Table 1. Properties and amino acid positions of human PEDF-derived peptides in relation to angiogenesis.
Note: numbering of amino acids corresponds to the full-length human PEDF sequence, i.e. 418 amino acids including the 20-amino-acid signal peptide.
| Properties | Positions and references | Cell or animal models used |
|---|---|---|
| Binding to PEDF receptor | Residues 44–77 [119,125,213,245], 44–121 [113], 46–70 [125], 78–121 [113,116,213] | Human Y-79, HuBMECs, T24 human urinary bladder carcinoma cells, HUVECs, human PC-3 cells and bovine retina plasma membrane |
| Binding to extracellular matrix | Residues 44–418 [134] | Not assayed |
| Neuron differentiation/neurotrophy | Residues 23–381 [6], 32–380 [6], 44–121 [6], 44–229 [6], 44–267 [6], 44–418 [246], 78–121 [113,213,247,248], 98–114 [213] | Human Y-79, embryonic rat motor neurons and human PC-3 cells |
| Neuroprotection | Residues 78–121 [154,247,249], 82–121 [250] | Human Y-79, embryonic rat motor neurons, diabetic retinal ganglion layer cells, rat organotypic spinal cord culture, mouse hypoxic retinal RGC layer, diabetic/ischaemic mouse inner plexiform layer and quinolinic acid-induced neurotoxicity in rat model |
| Anti-inflammation | Residues 60–77 [154], 78–121 [71,154], 82–121 [250] | Mouse retinal glial cells, mouse microglia, diabetic mouse vitreous and streptozotocin-induced diabetic mouse kidney |
| Anti-angiogenesis/anti-vasopermeability (anti-migration/anti-proliferation) | Residues 36–46 [213], 44–77 [91,212,213,251,252], 46–70 [125], 59–77 [212], 60–77 [154], 63–77 [212], 78–121 [154,253], 90–114 [62], 98–114 [213], 195–418 [254], 387–411 [62], 388–393 [93], 394–400 [93] | HuBMECs, b-FGF-induced corneal angiogenesis model, corneal micropocket assay, DIVAA, HUVECs, BRCEC, murine endothelial cells SVEC-4-10, mouse cornea, Akita mouse retina, matrigel plug assay on nude mouse, mouse corneal angiogenesis, PC-3 prostate cancer xenografts, Renca RCC exograft, VEGF-induced retinal vascular permeability mouse model, chicken embryo chorioallantoic membrane model OIR mouse model and laser-induced choroidal neovascularization rat model |
| Pro-apoptosis (in endothelial cells and tumour cells) | Residues 36–46 [213], 44–77 [212,213,245,251], 46–70 [125], 54–77 [212], 59–77 [212], 78–121 [213], 98–114 [213] | HuBMECs, bFGF-induced corneal angiogenesis model, BRCEC, human microvascular EC, T24 human urinary bladder carcinoma cells, HUVECs, mouse cornea, PC-3 cells, PC-3 prostate tumor xenograft and Renca RCC exograft |
Overall, the anti-angiogenic effects and other beneficial properties of PEDF make it an attractive candidate as a clinical therapeutic agent for angiogenesis. However, PEDF-induced insulin resistance and inflammation remain as potential concerns for its therapeutic applications. A previous study by Crowe et al. [41] showed that acute PEDF treatment in lean mice produced compromised insulin sensitivity in the skeletal muscle and liver. More importantly, prolonged systemic PEDF administration resulted in diabetogenic effects, including increased lipolysis and subsequent ectopic lipid deposition in the skeletal muscle and liver [41]. In addition, some studies reported that PEDF promoted the expression of inflammatory factors and cytokines in rat microglia [255,256] and neonatal rat astrocytes [257]. Furthermore, the anti-angiogenic effects of PEDF are not always favourable in diabetic complications. For instance, elevated PEDF levels in the circulation of patients with Type 1 [27,44] and Type 2 diabetes [26,45,47,258] may lead to deficient peripheral angiogenesis and defective wound healing, which might worsen diabetic symptoms such as diabetic foot ulcers. This concept is supported by a recently published report [68] that increased plasma PEDF levels are detected in Type 2 diabetic patients with diabetic foot ulcers compared with diabetic patients without diabetic foot ulcer, and neutralizing PEDF in diabetic mice accelerates wound healing by increasing angiogenesis. It is currently unclear whether increased PEDF levels in the circulation play a pathogenic role, or are a compensatory response to angiogenic diseases. To circumvent the systemic side effects, direct administration of PEDF into the angiogenic tissues/organs for the treatment of proliferative DR or other angiogenic diseases is a potential delivery approach. Further efforts are needed to confirm the potential clinical application of PEDF.
CONCLUSIONS
PEDF is a multifunctional serpin present in almost all tissues/organs and is involved in the maintenance of a variety of physiological functions. PEDF deficiency is known to play pathogenic roles in a number of diseased processes. Notably, PEDF levels are changed in diabetic and hypoxia-induced angiogenic diseases, which are believed to exacerbate the diseases. With broad activities and functions, PEDF has great clinical potential for disease diagnosis, treatment and prognosis prediction. However, its clinical application, especially its potential to combat pathological angiogenesis, remains to be explored.
ACKNOWLEDGEMENTS
We greatly appreciate Dr. Elizabeth Pearsall Moran and Dr. Junping Chen at the University of Oklahoma Health Sciences Center for their review and editing of this paper before submission.
References
- 1.Pignolo R.J., Cristofalo V.J., Rotenberg M.O. Senescent WI-38 cells fail to express EPC-1, a gene induced in young cells upon entry into the G0 state. J. Biol. Chem. 1993;268:8949–8957. [PubMed] [Google Scholar]
- 2.Pignolo R.J., Rotenberg M.O., Cristofalo V.J. Analysis of EPC-1 growth state-dependent expression, specificity, and conservation of related sequences. J. Cell Physiol. 1995;162:110–118. doi: 10.1002/jcp.1041620113. [DOI] [PubMed] [Google Scholar]
- 3.Tombran-Tink J., Johnson L.V. Neuronal differentiation of retinoblastoma cells induced by medium conditioned by human RPE cells. Invest. Ophthalmol. Vis. Sci. 1989;30:1700–1707. [PubMed] [Google Scholar]
- 4.Tombran-Tink J., Chader G.G., Johnson L.V. PEDF: a pigment epithelium-derived factor with potent neuronal differentiative activity. Exp. Eye Res. 1991;53:411–414. doi: 10.1016/0014-4835(91)90248-D. [DOI] [PubMed] [Google Scholar]
- 5.Tombran-Tink J., Aparicio S., Xu X., Tink A.R., Lara N., Sawant S., Barnstable C.J., Zhang S.S. PEDF and the serpins: phylogeny, sequence conservation, and functional domains. J. Struct. Biol. 2005;151:130–150. doi: 10.1016/j.jsb.2005.05.005. [DOI] [PubMed] [Google Scholar]
- 6.Becerra S.P., Sagasti A., Spinella P., Notario V. Pigment epithelium-derived factor behaves like a noninhibitory serpin. Neurotrophic activity does not require the serpin reactive loop. J. Biol. Chem. 1995;270:25992–25999. doi: 10.1074/jbc.270.43.25992. [DOI] [PubMed] [Google Scholar]
- 7.Becerra S.P. Structure-function studies on PEDF. A noninhibitory serpin with neurotrophic activity. Adv. Exp. Med. Biol. 1997;425:223–237. doi: 10.1007/978-1-4615-5391-5. [DOI] [PubMed] [Google Scholar]
- 8.Dawson D.W., Volpert O.V., Gillis P., Crawford S.E., Xu H., Benedict W., Bouck N.P. Pigment epithelium-derived factor: a potent inhibitor of angiogenesis. Science. 1999;285:245–248. doi: 10.1126/science.285.5425.245. [DOI] [PubMed] [Google Scholar]
- 9.Stellmach V., Crawford S.E., Zhou W., Bouck N. Prevention of ischemia-induced retinopathy by the natural ocular antiangiogenic agent pigment epithelium-derived factor. Proc. Natl. Acad. Sci. U.S.A. 2001;98:2593–2597. doi: 10.1073/pnas.031252398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Spranger J., Osterhoff M., Reimann M., Mohlig M., Ristow M., Francis M.K., Cristofalo V., Hammes H.P., Smith G., Boulton M., Pfeiffer A.F. Loss of the antiangiogenic pigment epithelium-derived factor in patients with angiogenic eye disease. Diabetes. 2001;50:2641–2645. doi: 10.2337/diabetes.50.12.2641. [DOI] [PubMed] [Google Scholar]
- 11.Gao G., Li Y., Fant J., Crosson C.E., Becerra S.P., Ma J.X. Difference in ischemic regulation of vascular endothelial growth factor and pigment epithelium-derived factor in brown Norway and Sprague Dawley rats contributing to different susceptibilities to retinal neovascularization. Diabetes. 2002;51:1218–1225. doi: 10.2337/diabetes.51.4.1218. [DOI] [PubMed] [Google Scholar]
- 12.Doll J.A., Stellmach V.M., Bouck N.P., Bergh A.R., Lee C., Abramson L.P., Cornwell M.L., Pins M.R., Borensztajn J., Crawford S.E. Pigment epithelium-derived factor regulates the vasculature and mass of the prostate and pancreas. Nat. Med. 2003;9:774–780. doi: 10.1038/nm870. [DOI] [PubMed] [Google Scholar]
- 13.Crawford S.E., Stellmach V., Ranalli M., Huang X., Huang L., Volpert O., De Vries G.H., Abramson L.P., Bouck N. Pigment epithelium-derived factor (PEDF) in neuroblastoma: a multifunctional mediator of Schwann cell antitumor activity. J. Cell. Sci. 2001;114:4421–4428. doi: 10.1242/jcs.114.24.4421. [DOI] [PubMed] [Google Scholar]
- 14.Abramson L.P., Stellmach V., Doll J.A., Cornwell M., Arensman R.M., Crawford S.E. Wilms’ tumor growth is suppressed by antiangiogenic pigment epithelium-derived factor in a xenograft model. J. Pediatr. Surg. 2003;38:336–342. doi: 10.1053/jpsu.2003.50104. [DOI] [PubMed] [Google Scholar]
- 15.Yokoi M., Yamagishi S., Saito A., Yoshida Y., Matsui T., Saito W., Hirose S., Ohgami K., Kase M., Ohno S. Positive association of pigment epithelium-derived factor with total antioxidant capacity in the vitreous fluid of patients with proliferative diabetic retinopathy. Br. J. Ophthalmol. 2007;91:885–887. doi: 10.1136/bjo.2006.110890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Boehm B.O., Lang G., Feldmann B., Kurkhaus A., Rosinger S., Volpert O., Lang G.K., Bouck N. Proliferative diabetic retinopathy is associated with a low level of the natural ocular anti-angiogenic agent pigment epithelium-derived factor (PEDF) in aqueous humor. a pilot study. Horm. Metab. Res. 2003;35:382–386. doi: 10.1055/s-2003-41362. [DOI] [PubMed] [Google Scholar]
- 17.Boehm B.O., Lang G., Volpert O., Jehle P.M., Kurkhaus A., Rosinger S., Lang G.K., Bouck N. Low content of the natural ocular anti-angiogenic agent pigment epithelium-derived factor (PEDF) in aqueous humor predicts progression of diabetic retinopathy. Diabetologia. 2003;46:394–400. doi: 10.1007/s00125-003-1040-9. [DOI] [PubMed] [Google Scholar]
- 18.Ogata N., Nishikawa M., Nishimura T., Mitsuma Y., Matsumura M. Unbalanced vitreous levels of pigment epithelium-derived factor and vascular endothelial growth factor in diabetic retinopathy. Am. J. Ophthalmol. 2002;134:348–353. doi: 10.1016/S0002-9394(02)01568-4. [DOI] [PubMed] [Google Scholar]
- 19.Guan M., Yam H.F., Su B., Chan K.P., Pang C.P., Liu W.W., Zhang W.Z., Lu Y. Loss of pigment epithelium derived factor expression in glioma progression. J. Clin. Pathol. 2003;56:277–282. doi: 10.1136/jcp.56.4.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Uehara H., Miyamoto M., Kato K., Ebihara Y., Kaneko H., Hashimoto H., Murakami Y., Hase R., Takahashi R., Mega S., et al. Expression of pigment epithelium-derived factor decreases liver metastasis and correlates with favorable prognosis for patients with ductal pancreatic adenocarcinoma. Cancer Res. 2004;64:3533–3537. doi: 10.1158/0008-5472.CAN-03-3725. [DOI] [PubMed] [Google Scholar]
- 21.Zhang L., Chen J., Ke Y., Mansel R.E., Jiang W.G. Expression of pigment epithelial derived factor is reduced in non-small cell lung cancer and is linked to clinical outcome. Int. J. Mol. Med. 2006;17:937–944. [PubMed] [Google Scholar]
- 22.Cai J., Parr C., Watkins G., Jiang W.G., Boulton M. Decreased pigment epithelium-derived factor expression in human breast cancer progression. Clin. Cancer Res. 2006;12:3510–3517. doi: 10.1158/1078-0432.CCR-06-0094. [DOI] [PubMed] [Google Scholar]
- 23.Holekamp N.M., Bouck N., Volpert O. Pigment epithelium-derived factor is deficient in the vitreous of patients with choroidal neovascularization due to age-related macular degeneration. Am. J. Ophthalmol. 2002;134:220–227. doi: 10.1016/S0002-9394(02)01549-0. [DOI] [PubMed] [Google Scholar]
- 24.Matsuyama K., Ogata N., Matsuoka M., Shima C., Wada M., Jo N., Matsumura M. Relationship between pigment epithelium-derived factor (PEDF) and renal function in patients with diabetic retinopathy. Mol. Vis. 2008;14:992–996. [PMC free article] [PubMed] [Google Scholar]
- 25.Ogata N., Matsuoka M., Matsuyama K., Shima C., Tajika A., Nishiyama T., Wada M., Jo N., Higuchi A., Minamino K., et al. Plasma concentration of pigment epithelium-derived factor in patients with diabetic retinopathy. J. Clin. Endocrinol. Metab. 2007;92:1176–1179. doi: 10.1210/jc.2006-2249. [DOI] [PubMed] [Google Scholar]
- 26.Chen H.B., Jia W.P., Lu J.X., Bao Y.Q., Li Q., Lu F.D., Lu W., Yu H.Y., Xiang K.S. Change and significance of serum pigment epithelium-derived factor in type 2 diabetic nephropathy. Zhonghua Yi Xue Za Zhi. 2007;87:1230–1233. [PubMed] [Google Scholar]
- 27.Katakami N., Kaneto H., Yamasaki Y., Matsuhisa M. Increased serum pigment epithelium-derived factor levels in type 1 diabetic patients with diabetic retinopathy. Diabetes Res. Clin. Pract. 2008;81:e4–e7. doi: 10.1016/j.diabres.2008.03.009. [DOI] [PubMed] [Google Scholar]
- 28.Venturi G., Gandini A., Monti E., Dalle Carbonare L., Corradi M., Vincenzi M., Valenti M.T., Valli M., Pelilli E., Boner A., et al. Lack of expression of SERPINF1, the gene coding for pigment epithelium-derived factor, causes progressively deforming osteogenesis imperfecta with normal type I collagen. J. Bone Miner. Res. 2012;27:723–728. doi: 10.1002/jbmr.1480. [DOI] [PubMed] [Google Scholar]
- 29.Bogan R., Riddle R.C., Li Z., Kumar S., Nandal A., Faugere M.C., Boskey A., Crawford S.E., Clemens T.L. A mouse model for human osteogenesis imperfecta type VI. J. Bone Miner. Res. 2013;28:1531–1536. doi: 10.1002/jbmr.1892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Huang Q., Wang S., Sorenson C.M., Sheibani N. PEDF-deficient mice exhibit an enhanced rate of retinal vascular expansion and are more sensitive to hyperoxia-mediated vessel obliteration. Exp. Eye Res. 2008;87:226–241. doi: 10.1016/j.exer.2008.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chung C., Doll J.A., Gattu A.K., Shugrue C., Cornwell M., Fitchev P., Crawford S.E. Anti-angiogenic pigment epithelium-derived factor regulates hepatocyte triglyceride content through adipose triglyceride lipase (ATGL) J. Hepatol. 2008;48:471–478. doi: 10.1016/j.jhep.2007.10.012. [DOI] [PubMed] [Google Scholar]
- 32.Schmitz J.C., Protiva P., Gattu A.K., Utsumi T., Iwakiri Y., Neto A.G., Quinn M., Cornwell M.L., Fitchev P., Lugea A., et al. Pigment epithelium-derived factor regulates early pancreatic fibrotic responses and suppresses the profibrotic cytokine thrombospondin-1. Am. J. Pathol. 2011;179:2990–2999. doi: 10.1016/j.ajpath.2011.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shin E.S., Sorenson C.M., Sheibani N. PEDF expression regulates the pro-angiogenic and pro-inflammatory phenotype of the lung endothelium. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013;306:620–634. doi: 10.1152/ajplung.00188.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Duh E.J., Yang H.S., Haller J.A., De Juan E., Humayun M.S., Gehlbach P., Melia M., Pieramici D., Harlan J.B., Campochiaro P.A., Zack D.J. Vitreous levels of pigment epithelium-derived factor and vascular endothelial growth factor: implications for ocular angiogenesis. Am. J. Ophthalmol. 2004;137:668–674. doi: 10.1016/j.ajo.2003.11.015. [DOI] [PubMed] [Google Scholar]
- 35.Yoshida Y., Yamagishi S., Matsui T., Nakamura K., Imaizumi T., Yoshimura K., Yamakawa R. Positive correlation of pigment epithelium-derived factor and total antioxidant capacity in aqueous humour of patients with uveitis and proliferative diabetic retinopathy. Br. J. Ophthalmol. 2007;91:1133–1134. doi: 10.1136/bjo.2007.115188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tombran-Tink J., Mazuruk K., Rodriguez I.R., Chung D., Linker T., Englander E., Chader G.J. Organization, evolutionary conservation, expression and unusual Alu density of the human gene for pigment epithelium-derived factor, a unique neurotrophic serpin. Mol. Vis. 1996;2:11. [PubMed] [Google Scholar]
- 37.Becker J., Semler O., Gilissen C., Li Y., Bolz H.J., Giunta C., Bergmann C., Rohrbach M., Koerber F., Zimmermann K., et al. Exome sequencing identifies truncating mutations in human SERPINF1 in autosomal-recessive osteogenesis imperfecta. Am. J. Hum. Genet. 2011;88:362–371. doi: 10.1016/j.ajhg.2011.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chung C., Shugrue C., Nagar A., Doll J.A., Cornwell M., Gattu A., Kolodecik T., Pandol S.J., Gorelick F. Ethanol exposure depletes hepatic pigment epithelium-derived factor, a novel lipid regulator. Gastroenterology. 2009;136:331–340. doi: 10.1053/j.gastro.2008.09.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sawant S., Aparicio S., Tink A.R., Lara N., Barnstable C.J., Tombran-Tink J. Regulation of factors controlling angiogenesis in liver development: a role for PEDF in the formation and maintenance of normal vasculature. Biochem. Biophys. Res. Commun. 2004;325:408–413. doi: 10.1016/j.bbrc.2004.10.041. [DOI] [PubMed] [Google Scholar]
- 40.Borg M.L., Andrews Z.B., Duh E.J., Zechner R., Meikle P.J., Watt M.J. Pigment epithelium-derived factor regulates lipid metabolism via adipose triglyceride lipase. Diabetes. 2011;60:1458–1466. doi: 10.2337/db10-0845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Crowe S., Wu L.E., Economou C., Turpin S.M., Matzaris M., Hoehn K.L., Hevener A.L., James D.E., Duh E.J., Watt M.J. Pigment epithelium-derived factor contributes to insulin resistance in obesity. Cell Metab. 2009;10:40–47. doi: 10.1016/j.cmet.2009.06.001. [DOI] [PubMed] [Google Scholar]
- 42.Ho T.C., Chen S.L., Shih S.C., Wu J.Y., Han W.H., Cheng H.C., Yang S.L., Tsao Y.P. Pigment epithelium-derived factor is an intrinsic antifibrosis factor targeting hepatic stellate cells. Am. J. Pathol. 2010;177:1798–1811. doi: 10.2353/ajpath.2010.091085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gattu A.K., Birkenfeld A.L., Jornayvaz F., Dziura J., Li F., Crawford S.E., Chu X., Still C.D., Gerhard G.S., Chung C., Samuel V. Insulin resistance is associated with elevated serum pigment epithelium-derived factor (PEDF) levels in morbidly obese patients. Acta Diabetol. 2012;49(Suppl. 1):S161–S169. doi: 10.1007/s00592-012-0397-y. [DOI] [PubMed] [Google Scholar]
- 44.Jenkins A.J., Zhang S.X., Rowley K.G., Karschimkus C.S., Nelson C.L., Chung J.S., O’Neal D.N., Januszewski A.S., Croft K.D., Mori T.A., et al. Increased serum pigment epithelium-derived factor is associated with microvascular complications, vascular stiffness and inflammation in Type 1 diabetes. Diabet. Med. 2007;24:1345–1351. doi: 10.1111/j.1464-5491.2007.02281.x. [DOI] [PubMed] [Google Scholar]
- 45.Jenkins A., Zhang S.X., Gosmanova A., Aston C., Dashti A., Baker M.Z., Lyons T., Ma J.X. Increased serum pigment epithelium derived factor levels in Type 2 diabetes patients. Diabetes Res. Clin. Pract. 2008;82:e5–e7. doi: 10.1016/j.diabres.2008.06.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nakamura K., Yamagishi S., Adachi H., Kurita-Nakamura Y., Matsui T., Inoue H. Serum levels of pigment epithelium-derived factor (PEDF) are positively associated with visceral adiposity in Japanese patients with type 2 diabetes. Diabetes Metab. Res. Rev. 2009;25:52–56. doi: 10.1002/dmrr.820. [DOI] [PubMed] [Google Scholar]
- 47.Jenkins A.J., Fu D., Azar M., Stoner J.A., Kaufman D.G., Zhang S., Klein R.L., Lopes-Virella M.F., Ma J.X., Lyons T.J., Vatd Investigators Clinical correlates of serum pigment epithelium-derived factor in type 2 diabetes patients. J. Diabetes Complications. 2014;28:353–359. doi: 10.1016/j.jdiacomp.2014.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Basu R., Chandramouli V., Dicke B., Landau B., Rizza R. Obesity and type 2 diabetes impair insulin-induced suppression of glycogenolysis as well as gluconeogenesis. Diabetes. 2005;54:1942–1948. doi: 10.2337/diabetes.54.7.1942. [DOI] [PubMed] [Google Scholar]
- 49.Boden G., Cheung P., Homko C. Effects of acute insulin excess and deficiency on gluconeogenesis and glycogenolysis in type 1 diabetes. Diabetes. 2003;52:133–137. doi: 10.2337/diabetes.52.1.133. [DOI] [PubMed] [Google Scholar]
- 50.Torres T.P., Fujimoto Y., Donahue E.P., Printz R.L., Houseknecht K.L., Treadway J.L., Shiota M. Defective glycogenesis contributes toward the inability to suppress hepatic glucose production in response to hyperglycemia and hyperinsulinemia in zucker diabetic fatty rats. Diabetes. 2011;60:2225–2233. doi: 10.2337/db09-1156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Samkharadze T., Erkan M., Reiser-Erkan C., Demir I.E., Kong B., Ceyhan G.O., Michalski C.W., Esposito I., Friess H., Kleeff J. Pigment epithelium-derived factor associates with neuropathy and fibrosis in pancreatic cancer. Am. J. Gastroenterol. 2011;106:968–980. doi: 10.1038/ajg.2010.479. [DOI] [PubMed] [Google Scholar]
- 52.Grippo P.J., Fitchev P.S., Bentrem D.J., Melstrom L.G., Dangi-Garimella S., Krantz S.B., Heiferman M.J., Chung C., Adrian K., Cornwell M.L., et al. Concurrent PEDF deficiency and Kras mutation induce invasive pancreatic cancer and adipose-rich stroma in mice. Gut. 2012;61:1454–1464. doi: 10.1136/gutjnl-2011-300821. [DOI] [PubMed] [Google Scholar]
- 53.Cosgrove G.P., Brown K.K., Schiemann W.P., Serls A.E., Parr J.E., Geraci M.W., Schwarz M.I., Cool C.D., Worthen G.S. Pigment epithelium-derived factor in idiopathic pulmonary fibrosis: a role in aberrant angiogenesis. Am. J. Respir. Crit. Care Med. 2004;170:242–251. doi: 10.1164/rccm.200308-1151OC. [DOI] [PubMed] [Google Scholar]
- 54.Chetty A., Bennett M., Dang L., Nakamura D., Cao G.J., Mujahid S., Volpe M., Herman I., Becerra S.P., Nielsen H.C. Pigment epithelium-derived factor mediates impaired lung vascular development in neonatal hyperoxia. Am. J. Respir. Cell Mol. Biol. 2014 doi: 10.1165/rcmb.2013-0229OC. doi:10.1165/rcmb.2013-0229OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hanumegowda C., Farkas L., Kolb M. Angiogenesis in pulmonary fibrosis: too much or not enough? Chest. 2012;142:200–207. doi: 10.1378/chest.11-1962. [DOI] [PubMed] [Google Scholar]
- 56.Quan G.M., Ojaimi J., Li Y., Kartsogiannis V., Zhou H., Choong P.F. Localization of pigment epithelium-derived factor in growing mouse bone. Calcif. Tissue Int. 2005;76:146–153. doi: 10.1007/s00223-004-0068-2. [DOI] [PubMed] [Google Scholar]
- 57.Tombran-Tink J., Barnstable C.J. Osteoblasts and osteoclasts express PEDF, VEGF-A isoforms, and VEGF receptors: possible mediators of angiogenesis and matrix remodeling in the bone. Biochem. Biophys. Res. Commun. 2004;316:573–579. doi: 10.1016/j.bbrc.2004.02.076. [DOI] [PubMed] [Google Scholar]
- 58.Li F., Song N., Tombran-Tink J., Niyibizi C. Pigment epithelium derived factor suppresses expression of Sost/sclerostin by osteocytes: implication for its role in bone matrix mineralization. J. Cell Physiol. 2014 doi: 10.1002/jcp.24859. doi: 10.1002/jcp.24859. [DOI] [PubMed] [Google Scholar]
- 59.Akiyama T., Dass C.R., Shinoda Y., Kawano H., Tanaka S., Choong P.F. PEDF regulates osteoclasts via osteoprotegerin and RANKL. Biochem. Biophys. Res. Commun. 2010;391:789–794. doi: 10.1016/j.bbrc.2009.11.139. [DOI] [PubMed] [Google Scholar]
- 60.Li F., Song N., Tombran-Tink J., Niyibizi C. Pigment epithelium derived factor enhances differentiation and mineral deposition of human mesenchymal stem cells. Stem Cells. 2013;31:2714–2723. doi: 10.1002/stem.1505. [DOI] [PubMed] [Google Scholar]
- 61.Gattu A.K., Swenson E.S., Iwakiri Y., Samuel V.T., Troiano N., Berry R., Church C.D., Rodeheffer M.S., Carpenter T.O., Chung C. Determination of mesenchymal stem cell fate by pigment epithelium-derived factor (PEDF) results in increased adiposity and reduced bone mineral content. FASEB J. 2013;27:4384–4394. doi: 10.1096/fj.13-232900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ek E.T., Dass C.R., Contreras K.G., Choong P.F. PEDF-derived synthetic peptides exhibit antitumor activity in an orthotopic model of human osteosarcoma. J. Orthop. Res. 2007;25:1671–1680. doi: 10.1002/jor.20434. [DOI] [PubMed] [Google Scholar]
- 63.Rauch F., Husseini A., Roughley P., Glorieux F.H., Moffatt P. Lack of circulating pigment epithelium-derived factor is a marker of osteogenesis imperfecta type VI. J. Clin. Endocrinol. Metab. 2012;97:E1550–E1556. doi: 10.1210/jc.2012-1827. [DOI] [PubMed] [Google Scholar]
- 64.Al-Jallad H., Palomo T., Moffatt P., Roughley P., Glorieux F.H., Rauch F. Normal bone density and fat mass in heterozygous SERPINF1 mutation carriers. J. Clin. Endocrinol. Metab. 2014;99:E2446–E2450. doi: 10.1210/jc.2014-2505. [DOI] [PubMed] [Google Scholar]
- 65.Farber C.R., Reich A., Barnes A.M., Becerra P., Rauch F., Cabral W.A., Bae A., Quinlan A., Glorieux F.H., Clemens T.L., Marini J.C. A novel IFITM5 mutation in severe atypical osteogenesis imperfecta type VI impairs osteoblast production of pigment epithelium-derived factor. J. Bone Miner. Res. 2014;29:1402–1411. doi: 10.1002/jbmr.2173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Baron R., Kneissel M. WNT signaling in bone homeostasis and disease: from human mutations to treatments. Nat. Med. 2013;19:179–192. doi: 10.1038/nm.3074. [DOI] [PubMed] [Google Scholar]
- 67.Park K., Lee K., Zhang B., Zhou T., He X., Gao G., Murray A.R., Ma J.X. Identification of a novel inhibitor of the canonical Wnt pathway. Mol. Cell Biol. 2011;31:3038–3051. doi: 10.1128/MCB.01211-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Qi W., Yang C., Dai Z., Che D., Feng J., Mao Y., Cheng R., Wang Z., He X., Zhou T., et al. High levels of pigment epithelium-derived factor in diabetes impair wound healing through suppression of Wnt signaling. Diabetes. 2014 doi: 10.2337/db14-1111. doi:10.2337/db14-1111. [DOI] [PubMed] [Google Scholar]
- 69.Li X., Zhang Y., Kang H., Liu W., Liu P., Zhang J., Harris S.E., Wu D. Sclerostin binds to LRP5/6 and antagonizes canonical Wnt signaling. J. Biol. Chem. 2005;280:19883–19887. doi: 10.1074/jbc.M413274200. [DOI] [PubMed] [Google Scholar]
- 70.Abramson L.P., Browne M., Stellmach V., Doll J., Cornwell M., Reynolds M., Arensman R.M., Crawford S.E. Pigment epithelium-derived factor targets endothelial and epithelial cells in Wilms’ tumor. J. Pediatr. Surg. 2006;41:1351–1356. doi: 10.1016/j.jpedsurg.2006.04.038. [DOI] [PubMed] [Google Scholar]
- 71.Awad A.S., Gao T., Gvritishvili A., You H., Liu Y., Cooper T.K., Reeves W.B., Tombran-Tink J. Protective role of small pigment epithelium-derived factor (PEDF) peptide in diabetic renal injury. Am. J. Physiol. Renal Physiol. 2013;305:F891–F900. doi: 10.1152/ajprenal.00149.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Pina A.L., Kubitza M., Brawanski A., Tombran-Tink J., Kloth S. Expression of pigment-epithelium-derived factor during kidney development and aging. Cell Tissue Res. 2007;329:329–338. doi: 10.1007/s00441-007-0420-8. [DOI] [PubMed] [Google Scholar]
- 73.Wang J.J., Zhang S.X., Lu K., Chen Y., Mott R., Sato S., Ma J.X. Decreased expression of pigment epithelium-derived factor is involved in the pathogenesis of diabetic nephropathy. Diabetes. 2005;54:243–250. doi: 10.2337/diabetes.54.1.243. [DOI] [PubMed] [Google Scholar]
- 74.Shiga Y., Miura S., Mitsutake R., Yamagishi S., Saku K. Significance of plasma levels of pigment epithelium-derived factor as determined by multidetector row computed tomography in patients with mild chronic kidney disease and/or coronary artery disease. J. Int. Med. Res. 2011;39:880–890. doi: 10.1177/147323001103900322. [DOI] [PubMed] [Google Scholar]
- 75.Hui E., Yeung C.Y., Lee P.C., Woo Y.C., Fong C.H., Chow W.S., Xu A., Lam K.S. Elevated circulating pigment epithelium-derived factor predicts the progression of diabetic nephropathy in patients with type 2 diabetes. J. Clin. Endocrinol. Metab. 2014;99:E2169–E2177. doi: 10.1210/jc.2014-2235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Karakousis P.C., John S.K., Behling K.C., Surace E.M., Smith J.E., Hendrickson A., Tang W.X., Bennett J., Milam A.H. Localization of pigment epithelium derived factor (PEDF) in developing and adult human ocular tissues. Mol. Vis. 2001;7:154–163. [PubMed] [Google Scholar]
- 77.Kozulin P., Natoli R., Bumsted O’Brien K.M., Madigan M.C., Provis J.M. The cellular expression of antiangiogenic factors in fetal primate macula. Invest. Ophthalmol. Vis. Sci. 2010;51:4298–4306. doi: 10.1167/iovs.09-4905. [DOI] [PubMed] [Google Scholar]
- 78.Becerra S.P., Fariss R.N., Wu Y.Q., Montuenga L.M., Wong P., Pfeffer B.A. Pigment epithelium-derived factor in the monkey retinal pigment epithelium and interphotoreceptor matrix: apical secretion and distribution. Exp. Eye Res. 2004;78:223–234. doi: 10.1016/j.exer.2003.10.013. [DOI] [PubMed] [Google Scholar]
- 79.Sonoda S., Sreekumar P.G., Kase S., Spee C., Ryan S.J., Kannan R., Hinton D.R. Attainment of polarity promotes growth factor secretion by retinal pigment epithelial cells: relevance to age-related macular degeneration. Aging (Albany NY) 2010;2:28–42. doi: 10.18632/aging.100111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zhu D., Deng X., Spee C., Sonoda S., Hsieh C.L., Barron E., Pera M., Hinton D.R. Polarized secretion of PEDF from human embryonic stem cell-derived RPE promotes retinal progenitor cell survival. Invest. Ophthalmol. Vis. Sci. 2011;52:1573–1585. doi: 10.1167/iovs.10-6413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Wu Y.Q., Notario V., Chader G.J., Becerra S.P. Identification of pigment epithelium-derived factor in the interphotoreceptor matrix of bovine eyes. Protein Expr. Purif. 1995;6:447–456. doi: 10.1006/prep.1995.1060. [DOI] [PubMed] [Google Scholar]
- 82.Wu Y.Q., Becerra S.P. Proteolytic activity directed toward pigment epithelium-derived factor in vitreous of bovine eyes. Implications of proteolytic processing. Invest. Ophthalmol. Vis. Sci. 1996;37:1984–1993. [PubMed] [Google Scholar]
- 83.Tombran-Tink J., Shivaram S.M., Chader G.J., Johnson L.V., Bok D. Expression, secretion, and age-related downregulation of pigment epithelium-derived factor, a serpin with neurotrophic activity. J. Neurosci. 1995;15:4992–5003. doi: 10.1523/JNEUROSCI.15-07-04992.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Behling K.C., Surace E.M., Bennett J. Pigment epithelium-derived factor expression in the developing mouse eye. Mol. Vis. 2002;8:449–454. [PubMed] [Google Scholar]
- 85.Ogata N., Wada M., Otsuji T., Jo N., Tombran-Tink J., Matsumura M. Expression of pigment epithelium-derived factor in normal adult rat eye and experimental choroidal neovascularization. Invest. Ophthalmol. Vis. Sci. 2002;43:1168–1175. [PubMed] [Google Scholar]
- 86.Sorenson C.M., Wang S., Gendron R., Paradis H., Sheibani N. Thrombospondin-1 deficiency exacerbates the pathogenesis of diabetic retinopathy. J. Diabetes Metab. 2013 doi: 10.4172/2155-6156.S12-005. doi:10.4172/2155-6156.S12-005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Park K., Jin J., Hu Y., Zhou K., Ma J.X. Overexpression of pigment epithelium-derived factor inhibits retinal inflammation and neovascularization. Am. J. Pathol. 2011;178:688–698. doi: 10.1016/j.ajpath.2010.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Haurigot V., Villacampa P., Ribera A., Bosch A., Ramos D., Ruberte J., Bosch F. Long-term retinal PEDF overexpression prevents neovascularization in a murine adult model of retinopathy. PLoS ONE. 2012;7:e41511. doi: 10.1371/journal.pone.0041511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Mori K., Duh E., Gehlbach P., Ando A., Takahashi K., Pearlman J., Mori K., Yang H.S., Zack D.J., Ettyreddy D., et al. Pigment epithelium-derived factor inhibits retinal and choroidal neovascularization. J. Cell Physiol. 2001;188:253–263. doi: 10.1002/jcp.1114. [DOI] [PubMed] [Google Scholar]
- 90.Saishin Y., Silva R.L., Saishin Y., Kachi S., Aslam S., Gong Y.Y., Lai H., Carrion M., Harris B., Hamilton M., et al. Periocular gene transfer of pigment epithelium-derived factor inhibits choroidal neovascularization in a human-sized eye. Hum. Gene Ther. 2005;16:473–478. doi: 10.1089/hum.2005.16.473. [DOI] [PubMed] [Google Scholar]
- 91.Amaral J., Becerra S.P. Effects of human recombinant PEDF protein and PEDF-derived peptide 34-mer on choroidal neovascularization. Invest. Ophthalmol. Vis. Sci. 2010;51:1318–1326. doi: 10.1167/iovs.09-4455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kuo C.N., Chen C.Y., Chen S.N., Yang L.C., Lai L.J., Lai C.H., Chen M.F., Hung C.H., Chen C.H. Inhibition of corneal neovascularization with the combination of bevacizumab and plasmid pigment epithelium-derived factor-synthetic amphiphile interaction-18 (p-PEDF-SAINT-18) vector in a rat corneal experimental angiogenesis model. Int. J. Mol. Sci. 2013;14:8291–8305. doi: 10.3390/ijms14048291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Matsui T., Nishino Y., Maeda S., Yamagishi S. PEDF-derived peptide inhibits corneal angiogenesis by suppressing VEGF expression. Microvasc. Res. 2012;84:105–108. doi: 10.1016/j.mvr.2012.02.006. [DOI] [PubMed] [Google Scholar]
- 94.Kuo C.N., Yang L.C., Yang C.T., Lai C.H., Chen M.F., Chen C.Y., Chen C.H., Wu P.C., Kou H.K., Chen Y.J., et al. Inhibition of corneal neovascularization with plasmid pigment epithelium-derived factor (p-PEDF) delivered by synthetic amphiphile interaction-18 (SAINT-18) vector in an experimental model of rat corneal angiogenesis. Exp. Eye Res. 2009;89:678–685. doi: 10.1016/j.exer.2009.06.021. [DOI] [PubMed] [Google Scholar]
- 95.Volpert K.N., Tombran-Tink J., Barnstable C., Layer P.G. PEDF and GDNF are key regulators of photoreceptor development and retinal neurogenesis in reaggregates from chick embryonic retina. J. Ocul. Biol. Dis. Infor. 2009;2:1–11. doi: 10.1007/s12177-009-9014-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Malchiodi-Albedi F., Feher J., Caiazza S., Formisano G., Perilli R., Falchi M., Petrucci T.C., Scorcia G., Tombran-Tink J. PEDF (pigment epithelium-derived factor) promotes increase and maturation of pigment granules in pigment epithelial cells in neonatal albino rat retinal cultures. Int. J. Dev. Neurosci. 1998;16:423–432. doi: 10.1016/S0736-5748(98)00014-8. [DOI] [PubMed] [Google Scholar]
- 97.Arnhold S., Heiduschka P., Klein H., Absenger Y., Basnaoglu S., Kreppel F., Henke-Fahle S., Kochanek S., Bartz-Schmidt K.U., Addicks K., Schraermeyer U. Adenovirally transduced bone marrow stromal cells differentiate into pigment epithelial cells and induce rescue effects in RCS rats. Invest. Ophthalmol. Vis. Sci. 2006;47:4121–4129. doi: 10.1167/iovs.04-1501. [DOI] [PubMed] [Google Scholar]
- 98.Cayouette M., Smith S.B., Becerra S.P., Gravel C. Pigment epithelium-derived factor delays the death of photoreceptors in mouse models of inherited retinal degenerations. Neurobiol. Dis. 1999;6:523–532. doi: 10.1006/nbdi.1999.0263. [DOI] [PubMed] [Google Scholar]
- 99.Cao W., Tombran-Tink J., Chen W., Mrazek D., Elias R., McGinnis J.F. Pigment epithelium-derived factor protects cultured retinal neurons against hydrogen peroxide-induced cell death. J. Neurosci. Res. 1999;57:789–800. doi: 10.1002/(SICI)1097-4547(19990915)57:6<789::AID-JNR4>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- 100.Wang Y., Subramanian P., Shen D., Tuo J., Becerra S.P., Chan C.C. Pigment epithelium-derived factor reduces apoptosis and pro-inflammatory cytokine gene expression in a murine model of focal retinal degeneration. ASN NEURO. 2013;5:e00126. doi: 10.1042/AN20130028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Cao W., Tombran-Tink J., Elias R., Sezate S., Mrazek D., McGinnis J.F. In vivo protection of photoreceptors from light damage by pigment epithelium-derived factor. Invest. Ophthalmol. Vis. Sci. 2001;42:1646–1652. [PubMed] [Google Scholar]
- 102.Imai D., Yoneya S., Gehlbach P.L., Wei L.L., Mori K. Intraocular gene transfer of pigment epithelium-derived factor rescues photoreceptors from light-induced cell death. J. Cell Physiol. 2005;202:570–578. doi: 10.1002/jcp.20155. [DOI] [PubMed] [Google Scholar]
- 103.Jablonski M.M., Tombran-Tink J., Mrazek D.A., Iannaccone A. Pigment epithelium-derived factor supports normal development of photoreceptor neurons and opsin expression after retinal pigment epithelium removal. J. Neurosci. 2000;20:7149–7157. doi: 10.1523/JNEUROSCI.20-19-07149.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Subramanian P., Locatelli-Hoops S., Kenealey J., DesJardin J., Notari L., Becerra S.P. Pigment epithelium-derived factor (PEDF) prevents retinal cell death via PEDF receptor (PEDF-R): identification of a functional ligand binding site. J. Biol. Chem. 2013;288:23928–23942. doi: 10.1074/jbc.M113.487884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Folkman J., Klagsbrun M. Angiogenic factors. Science. 1987;235:442–447. doi: 10.1126/science.2432664. [DOI] [PubMed] [Google Scholar]
- 106.Wietecha M.S., Cerny W.L., DiPietro L.A. Mechanisms of vessel regression: toward an understanding of the resolution of angiogenesis. Curr. Top Microbiol. Immunol. 2013;367:3–32. doi: 10.1007/82_2012_287. [DOI] [PubMed] [Google Scholar]
- 107.Hoshina D., Abe R., Yamagishi S.I., Shimizu H. The role of PEDF in tumor growth and metastasis. Curr. Mol. Med. 2010;10:292–295. doi: 10.2174/156652410791065327. [DOI] [PubMed] [Google Scholar]
- 108.Manalo K.B., Choong P.F., Becerra S.P., Dass C.R. Pigment epithelium-derived factor as an anticancer drug and new treatment methods following the discovery of its receptors: a patent perspective. Expert. Opin. Ther. Pat. 2011;21:121–130. doi: 10.1517/13543776.2011.545347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Becerra S.P., Notario V. The effects of PEDF on cancer biology: mechanisms of action and therapeutic potential. Nat. Rev. Cancer. 2013;13:258–271. doi: 10.1038/nrc3484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Rychli K., Huber K., Wojta J. Pigment epithelium-derived factor (PEDF) as a therapeutic target in cardiovascular disease. Expert Opin. Ther. Targets. 2009;13:1295–1302. doi: 10.1517/14728220903241641. [DOI] [PubMed] [Google Scholar]
- 111.Liu J.T., Chen Y.L., Chen W.C., Chen H.Y., Lin Y.W., Wang S.H., Man K.M., Wan H.M., Yin W.H., Liu P.L., Chen Y.H. Role of pigment epithelium-derived factor in stem/progenitor cell-associated neovascularization. J. Biomed. Biotechnol. 2012;2012:871272. doi: 10.1155/2012/871272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Notari L., Baladron V., Aroca-Aguilar J.D., Balko N., Heredia R., Meyer C., Notario P.M., Saravanamuthu S., Nueda M.L., Sanchez-Sanchez F., et al. Identification of a lipase-linked cell membrane receptor for pigment epithelium-derived factor. J. Biol. Chem. 2006;281:38022–38037. doi: 10.1074/jbc.M600353200. [DOI] [PubMed] [Google Scholar]
- 113.Alberdi E., Aymerich M.S., Becerra S.P. Binding of pigment epithelium-derived factor (PEDF) to retinoblastoma cells and cerebellar granule neurons. Evidence for a PEDF receptor. J. Biol. Chem. 1999;274:31605–31612. doi: 10.1074/jbc.274.44.31605. [DOI] [PubMed] [Google Scholar]
- 114.Subramanian P., Notario P.M., Becerra S.P. Pigment epithelium-derived factor receptor (PEDF-R): a plasma membrane-linked phospholipase with PEDF binding affinity. Adv. Exp. Med. Biol. 2010;664:29–37. doi: 10.1007/978-1-4419-1399-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Desjardin J.T., Becerra S.P., Subramanian P. Searching for alternatively spliced variants of phospholipase domain-containing 2 (Pnpla2), a novel gene in the retina. J. Clin. Exp. Ophthalmol. 2013;4:295. doi: 10.4172/2155-9570.1000295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Aymerich M.S., Alberdi E.M., Martinez A., Becerra S.P. Evidence for pigment epithelium-derived factor receptors in the neural retina. Invest. Ophthalmol. Vis. Sci. 2001;42:3287–3293. [PubMed] [Google Scholar]
- 117.Subramanian P., Rapp M., Becerra S.P. Identification of pigment epithelium-derived factor receptor (PEDF-R) antibody epitopes. Adv. Exp. Med. Biol. 2012;723:799–805. doi: 10.1007/978-1-4614-0631-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Hirsch J., Johnson C.L., Nelius T., Kennedy R., Riese W., Filleur S. PEDF inhibits IL8 production in prostate cancer cells through PEDF receptor/phospholipase A2 and regulation of NFkappaB and PPARgamma. Cytokine. 2011;55:202–210. doi: 10.1016/j.cyto.2011.04.010. [DOI] [PubMed] [Google Scholar]
- 119.Ladhani O., Sanchez-Martinez C., Orgaz J.L., Jimenez B., Volpert O.V. Pigment epithelium-derived factor blocks tumor extravasation by suppressing amoeboid morphology and mesenchymal proteolysis. Neoplasia. 2011;13:633–642. doi: 10.1593/neo.11446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Chavan S.S., Hudson L.K., Li J.H., Ochani M., Harris Y., Patel N.B., Katz D., Scheinerman J.A., Pavlov V.A., Tracey K.J. Identification of pigment epithelium-derived factor as an adipocyte-derived inflammatory factor. Mol. Med. 2012;18:1161–1168. doi: 10.2119/molmed.2012.00156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Moreno-Navarrete J.M., Touskova V., Sabater M., Mraz M., Drapalova J., Ortega F., Serrano M., Catalan V., Gomez-Ambrosi J., Ortiz M.R., et al. Liver, but not adipose tissue PEDF gene expression is associated with insulin resistance. Int. J. Obes. (Lond). 2013;37:1230–1237. doi: 10.1038/ijo.2012.223. [DOI] [PubMed] [Google Scholar]
- 122.Gonzalez R., Jennings L.L., Knuth M., Orth A.P., Klock H.E., Ou W., Feuerhelm J., Hull M.V., Koesema E., Wang Y., et al. Screening the mammalian extracellular proteome for regulators of embryonic human stem cell pluripotency. Proc. Natl. Acad. Sci. U.S.A. 2010;107:3552–3557. doi: 10.1073/pnas.0914019107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Rapp M., Woo G., Al-Ubaidi M.R., Becerra S.P., Subramanian P. Pigment epithelium-derived factor protects cone photoreceptor-derived 661W cells from light damage through Akt activation. Adv. Exp. Med. Biol. 2014;801:813–820. doi: 10.1007/978-1-4614-3209-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Aurora A.B., Biyashev D., Mirochnik Y., Zaichuk T.A., Sanchez-Martinez C., Renault M.A., Losordo D., Volpert O.V. NF-kappaB balances vascular regression and angiogenesis via chromatin remodeling and NFAT displacement. Blood. 2010;116:475–484. doi: 10.1182/blood-2009-07-232132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Bernard A., Gao-Li J., Franco C.A., Bouceba T., Huet A., Li Z. Laminin receptor involvement in the anti-angiogenic activity of pigment epithelium-derived factor. J. Biol. Chem. 2009;284:10480–10490. doi: 10.1074/jbc.M809259200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Konson A., Pradeep S., D’Acunto C.W., Seger R. Pigment epithelium-derived factor and its phosphomimetic mutant induce JNK-dependent apoptosis and p38-mediated migration arrest. J. Biol. Chem. 2011;286:3540–3551. doi: 10.1074/jbc.M110.151548. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 127.Notari L., Arakaki N., Mueller D., Meier S., Amaral J., Becerra S.P. Pigment epithelium-derived factor binds to cell-surface F(1)-ATP synthase. FEBS J. 2010;277:2192–2205. doi: 10.1111/j.1742-4658.2010.07641.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Willnow T.E., Goldstein J.L., Orth K., Brown M.S., Herz J. Low density lipoprotein receptor-related protein and gp330 bind similar ligands, including plasminogen activator-inhibitor complexes and lactoferrin, an inhibitor of chylomicron remnant clearance. J. Biol. Chem. 1992;267:26172–26180. [PubMed] [Google Scholar]
- 129.Conese M., Olson D., Blasi F. Protease nexin-1-urokinase complexes are internalized and degraded through a mechanism that requires both urokinase receptor and alpha 2-macroglobulin receptor. J. Biol. Chem. 1994;269:17886–17892. [PubMed] [Google Scholar]
- 130.Poller W., Willnow T.E., Hilpert J., Herz J. Differential recognition of alpha 1-antitrypsin-elastase and alpha 1-antichymotrypsin-cathepsin G complexes by the low density lipoprotein receptor-related protein. J. Biol. Chem. 1995;270:2841–2845. doi: 10.1074/jbc.270.6.2841. [DOI] [PubMed] [Google Scholar]
- 131.Zhang B., Zhou K.K., Ma J.X. Inhibition of connective tissue growth factor overexpression in diabetic retinopathy by SERPINA3K via blocking the WNT/beta-catenin pathway. Diabetes. 2010;59:1809–1816. doi: 10.2337/db09-1056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Hu Y., Chen Y., Lin M., Lee K., Mott R.A., Ma J.X. Pathogenic role of the Wnt signaling pathway activation in laser-induced choroidal neovascularization. Invest. Ophthalmol. Vis. Sci. 2013;54:141–154. doi: 10.1167/iovs.12-10281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Lee K., Hu Y., Ding L., Chen Y., Takahashi Y., Mott R., Ma J.X. Therapeutic potential of a monoclonal antibody blocking the Wnt pathway in diabetic retinopathy. Diabetes. 2012;61:2948–2957. doi: 10.2337/db11-0300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Meyer C., Notari L., Becerra S.P. Mapping the type I collagen-binding site on pigment epithelium-derived factor. Implications for its antiangiogenic activity. J. Biol. Chem. 2002;277:45400–45407. doi: 10.1074/jbc.M208339200. [DOI] [PubMed] [Google Scholar]
- 135.Hosomichi J., Yasui N., Koide T., Soma K., Morita I. Involvement of the collagen I-binding motif in the anti-angiogenic activity of pigment epithelium-derived factor. Biochem. Biophys. Res. Commun. 2005;335:756–761. doi: 10.1016/j.bbrc.2005.07.140. [DOI] [PubMed] [Google Scholar]
- 136.Kozaki K., Miyaishi O., Koiwai O., Yasui Y., Kashiwai A., Nishikawa Y., Shimizu S., Saga S. Isolation, purification, and characterization of a collagen-associated serpin, caspin, produced by murine colon adenocarcinoma cells. J. Biol. Chem. 1998;273:15125–15130. doi: 10.1074/jbc.273.24.15125. [DOI] [PubMed] [Google Scholar]
- 137.Alberdi E., Hyde C.C., Becerra S.P. Pigment epithelium-derived factor (PEDF) binds to glycosaminoglycans: analysis of the binding site. Biochemistry. 1998;37:10643–10652. doi: 10.1021/bi9802317. [DOI] [PubMed] [Google Scholar]
- 138.Becerra S.P., Perez-Mediavilla L.A., Weldon J.E., Locatelli-Hoops S., Senanayake P., Notari L., Notario V., Hollyfield J.G. Pigment epithelium-derived factor binds to hyaluronan. Mapping of a hyaluronan binding site. J. Biol. Chem. 2008;283:33310–33320. doi: 10.1074/jbc.M801287200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Valnickova Z., Petersen S.V., Nielsen S.B., Otzen D.E., Enghild J.J. Heparin binding induces a conformational change in pigment epithelium-derived factor. J. Biol. Chem. 2007;282:6661–6667. doi: 10.1074/jbc.M610471200. [DOI] [PubMed] [Google Scholar]
- 140.Alberdi E.M., Weldon J.E., Becerra S.P. Glycosaminoglycans in human retinoblastoma cells: heparan sulfate, a modulator of the pigment epithelium-derived factor-receptor interactions. BMC Biochem. 2003;4:1. doi: 10.1186/1471-2091-4-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Martin A., Komada M.R., Sane D.C. Abnormal angiogenesis in diabetes mellitus. Med. Res. Rev. 2003;23:117–145. doi: 10.1002/med.10024. [DOI] [PubMed] [Google Scholar]
- 142.Nyengaard J.R., Rasch R. The impact of experimental diabetes mellitus in rats on glomerular capillary number and sizes. Diabetologia. 1993;36:189–194. doi: 10.1007/BF00399948. [DOI] [PubMed] [Google Scholar]
- 143.Mohan N., Monickaraj F., Balasubramanyam M., Rema M., Mohan V. Imbalanced levels of angiogenic and angiostatic factors in vitreous, plasma and postmortem retinal tissue of patients with proliferative diabetic retinopathy. J. Diabetes Complications. 2012;26:435–441. doi: 10.1016/j.jdiacomp.2012.05.005. [DOI] [PubMed] [Google Scholar]
- 144.Wang H., Feng L., Hu J.W., Xie C.L., Wang F. Characterisation of the vitreous proteome in proliferative diabetic retinopathy. Proteome. Sci. 2012;10:15. doi: 10.1186/1477-5956-10-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Huber M., Wachtlin J. Vitreous levels of proteins implicated in angiogenesis are modulated in patients with retinal or choroidal neovascularization. Ophthalmologica. 2012;228:188–193. doi: 10.1159/000339952. [DOI] [PubMed] [Google Scholar]
- 146.Zheng B., Li T., Chen H., Xu X., Zheng Z. Correlation between ficolin-3 and vascular endothelial growth factor-to-pigment epithelium-derived factor ratio in the vitreous of eyes with proliferative diabetic retinopathy. Am. J. Ophthalmol. 2011;152:1039–1043. doi: 10.1016/j.ajo.2011.05.022. [DOI] [PubMed] [Google Scholar]
- 147.Matsunaga N., Chikaraishi Y., Izuta H., Ogata N., Shimazawa M., Matsumura M., Hara H. Role of soluble vascular endothelial growth factor receptor-1 in the vitreous in proliferative diabetic retinopathy. Ophthalmology. 2008;115:1916–1922. doi: 10.1016/j.ophtha.2008.06.025. [DOI] [PubMed] [Google Scholar]
- 148.Murugeswari P., Shukla D., Rajendran A., Kim R., Namperumalsamy P., Muthukkaruppan V. Proinflammatory cytokines and angiogenic and anti-angiogenic factors in vitreous of patients with proliferative diabetic retinopathy and eales’ disease. Retina. 2008;28:817–824. doi: 10.1097/IAE.0b013e31816576d5. [DOI] [PubMed] [Google Scholar]
- 149.Garcia-Ramirez M., Canals F., Hernandez C., Colome N., Ferrer C., Carrasco E., Garcia-Arumi J., Simo R. Proteomic analysis of human vitreous fluid by fluorescence-based difference gel electrophoresis (DIGE): a new strategy for identifying potential candidates in the pathogenesis of proliferative diabetic retinopathy. Diabetologia. 2007;50:1294–1303. doi: 10.1007/s00125-007-0627-y. [DOI] [PubMed] [Google Scholar]
- 150.Wang J.J., Zhang S.X., Mott R., Knapp R.R., Cao W., Lau K., Ma J.X. Salutary effect of pigment epithelium-derived factor in diabetic nephropathy: evidence for antifibrogenic activities. Diabetes. 2006;55:1678–1685. doi: 10.2337/db05-1448. [DOI] [PubMed] [Google Scholar]
- 151.Arimura T., Miura S., Sugihara M., Iwata A., Yamagishi S., Saku K. Association between plasma levels of pigment epithelium-derived factor and renal dysfunction in patients with coronary artery disease. Cardiol. J. 2011;18:515–520. doi: 10.5603/CJ.2011.0006. [DOI] [PubMed] [Google Scholar]
- 152.Yoshida Y., Yamagishi S., Matsui T., Jinnouchi Y., Fukami K., Imaizumi T., Yamakawa R. Protective role of pigment epithelium-derived factor (PEDF) in early phase of experimental diabetic retinopathy. Diabetes Metab. Res. Rev. 2009;25:678–686. doi: 10.1002/dmrr.1007. [DOI] [PubMed] [Google Scholar]
- 153.Zhang S.X., Wang J.J., Gao G., Parke K., Ma J.X. Pigment epithelium-derived factor downregulates vascular endothelial growth factor (VEGF) expression and inhibits VEGF-VEGF receptor 2 binding in diabetic retinopathy. J. Mol. Endocrinol. 2006;37:1–12. doi: 10.1677/jme.1.02008. [DOI] [PubMed] [Google Scholar]
- 154.Liu Y., Leo L.F., McGregor C., Grivitishvili A., Barnstable C.J., Tombran-Tink J. Pigment epithelium-derived factor (PEDF) peptide eye drops reduce inflammation, cell death and vascular leakage in diabetic retinopathy in Ins2(Akita) mice. Mol. Med. 2012;18:1387–1401. doi: 10.2119/molmed.2012.00008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Wang J.J., Zhang S.X., Mott R., Chen Y., Knapp R.R., Cao W., Ma J.X. Anti-inflammatory effects of pigment epithelium-derived factor in diabetic nephropathy. Am. J. Physiol. Renal Physiol. 2008;294:F1166–F1173. doi: 10.1152/ajprenal.00375.2007. [DOI] [PubMed] [Google Scholar]
- 156.Rundhaug J.E. Matrix metalloproteinases and angiogenesis. J. Cell. Mol. Med. 2005;9:267–285. doi: 10.1111/j.1582-4934.2005.tb00355.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Yafai Y., Lange J., Wiedemann P., Reichenbach A., Eichler W. Pigment epithelium-derived factor acts as an opponent of growth-stimulatory factors in retinal glial-endothelial cell interactions. Glia. 2007;55:642–651. doi: 10.1002/glia.20495. [DOI] [PubMed] [Google Scholar]
- 158.Hutchings H., Maitre-Boube M., Tombran-Tink J., Plouet J. Pigment epithelium-derived factor exerts opposite effects on endothelial cells of different phenotypes. Biochem. Biophys. Res. Commun. 2002;294:764–769. doi: 10.1016/S0006-291X(02)00548-X. [DOI] [PubMed] [Google Scholar]
- 159.Biyashev D., Veliceasa D., Kwiatek A., Sutanto M.M., Cohen R.N., Volpert O.V. Natural angiogenesis inhibitor signals through Erk5 activation of peroxisome proliferator-activated receptor gamma (PPARgamma) J. Biol. Chem. 2010;285:13517–13524. doi: 10.1074/jbc.M110.117374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Dejana E. The role of Wnt signaling in physiological and pathological angiogenesis. Circ. Res. 2010;107:943–952. doi: 10.1161/CIRCRESAHA.110.223750. [DOI] [PubMed] [Google Scholar]
- 161.Zhang X., Gaspard J.P., Chung D.C. Regulation of vascular endothelial growth factor by the Wnt and K-ras pathways in colonic neoplasia. Cancer Res. 2001;61:6050–6054. [PubMed] [Google Scholar]
- 162.Brabletz T., Jung A., Dag S., Hlubek F., Kirchner T. β-Catenin regulates the expression of the matrix metalloproteinase-7 in human colorectal cancer. Am. J. Pathol. 1999;155:1033–1038. doi: 10.1016/S0002-9440(10)65204-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Crawford H.C., Fingleton B.M., Rudolph-Owen L.A., Goss K.J., Rubinfeld B., Polakis P., Matrisian L.M. The metalloproteinase matrilysin is a target of β-catenin transactivation in intestinal tumors. Oncogene. 1999;18:2883–2891. doi: 10.1038/sj.onc.1202627. [DOI] [PubMed] [Google Scholar]
- 164.Wu B., Crampton S.P., Hughes C.C. Wnt signaling induces matrix metalloproteinase expression and regulates T cell transmigration. Immunity. 2007;26:227–239. doi: 10.1016/j.immuni.2006.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Yamagishi S., Nakamura K., Matsui T., Inagaki Y., Takenaka K., Jinnouchi Y., Yoshida Y., Matsuura T., Narama I., Motomiya Y., et al. Pigment epithelium-derived factor inhibits advanced glycation end product-induced retinal vascular hyperpermeability by blocking reactive oxygen species-mediated vascular endothelial growth factor expression. J. Biol. Chem. 2006;281:20213–20220. doi: 10.1074/jbc.M602110200. [DOI] [PubMed] [Google Scholar]
- 166.Yamagishi S., Matsui T., Nakamura K., Yoshida T., Takeuchi M., Inoue H., Yoshida Y., Imaizumi T. Pigment-epithelium-derived factor suppresses expression of receptor for advanced glycation end products in the eye of diabetic rats. Ophthalmic Res. 2007;39:92–97. doi: 10.1159/000099244. [DOI] [PubMed] [Google Scholar]
- 167.Yamagishi S., Inagaki Y., Nakamura K., Abe R., Shimizu T., Yoshimura A., Imaizumi T. Pigment epithelium-derived factor inhibits TNF-alpha-induced interleukin-6 expression in endothelial cells by suppressing NADPH oxidase-mediated reactive oxygen species generation. J. Mol. Cell Cardiol. 2004;37:497–506. doi: 10.1016/j.yjmcc.2004.04.007. [DOI] [PubMed] [Google Scholar]
- 168.Yamagishi S., Nakamura K., Ueda S., Kato S., Imaizumi T. Pigment epithelium-derived factor (PEDF) blocks angiotensin II signaling in endothelial cells via suppression of NADPH oxidase: a novel anti-oxidative mechanism of PEDF. Cell Tissue Res. 2005;320:437–445. doi: 10.1007/s00441-005-1094-8. [DOI] [PubMed] [Google Scholar]
- 169.Zheng Z., Chen H., Zhao H., Liu K., Luo D., Chen Y., Chen Y., Yang X., Gu Q., Xu X. Inhibition of JAK2/STAT3-mediated VEGF upregulation under high glucose conditions by PEDF through a mitochondrial ROS pathway in vitro. Invest. Ophthalmol. Vis. Sci. 2010;51:64–71. doi: 10.1167/iovs.09-3511. [DOI] [PubMed] [Google Scholar]
- 170.Ejaz S., Chekarova I., Ejaz A., Sohail A., Lim C.W. Importance of pericytes and mechanisms of pericyte loss during diabetes retinopathy. Diabetes Obes. Metab. 2008;10:53–63. doi: 10.1111/j.1463-1326.2007.00795.x. [DOI] [PubMed] [Google Scholar]
- 171.Zhang S.X., Wang J.J., Dashti A., Wilson K., Zou M.H., Szweda L., Ma J.X., Lyons T.J. Pigment epithelium-derived factor mitigates inflammation and oxidative stress in retinal pericytes exposed to oxidized low-density lipoprotein. J. Mol. Endocrinol. 2008;41:135–143. doi: 10.1677/JME-08-0011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Yamagishi S., Inagaki Y., Amano S., Okamoto T., Takeuchi M., Makita Z. Pigment epithelium-derived factor protects cultured retinal pericytes from advanced glycation end product-induced injury through its antioxidative properties. Biochem. Biophys. Res. Commun. 2002;296:877–882. doi: 10.1016/S0006-291X(02)00940-3. [DOI] [PubMed] [Google Scholar]
- 173.Amano S., Yamagishi S., Inagaki Y., Nakamura K., Takeuchi M., Inoue H., Imaizumi T. Pigment epithelium-derived factor inhibits oxidative stress-induced apoptosis and dysfunction of cultured retinal pericytes. Microvasc. Res. 2005;69:45–55. doi: 10.1016/j.mvr.2004.11.001. [DOI] [PubMed] [Google Scholar]
- 174.Enge M., Bjarnegard M., Gerhardt H., Gustafsson E., Kalen M., Asker N., Hammes H.P., Shani M., Fassler R., Betsholtz C. Endothelium-specific platelet-derived growth factor-B ablation mimics diabetic retinopathy. EMBO J. 2002;21:4307–4316. doi: 10.1093/emboj/cdf418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Yamagishi S., Nakamura K., Takenaka K., Matsui T., Jinnouchi Y., Imaizumi T. Pigment epithelium-derived factor (PEDF) promotes growth of pericytes through autocrine production of platelet-derived growth factor-B. Microvasc. Res. 2005;69:128–134. doi: 10.1016/j.mvr.2005.04.001. [DOI] [PubMed] [Google Scholar]
- 176.Wang G.L., Jiang B.H., Rue E.A., Semenza G.L. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. U.S.A. 1995;92:5510–5514. doi: 10.1073/pnas.92.12.5510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Pugh C.W., Ratcliffe P.J. Regulation of angiogenesis by hypoxia: role of the HIF system. Nat. Med. 2003;9:677–684. doi: 10.1038/nm0603-677. [DOI] [PubMed] [Google Scholar]
- 178.Ke Q., Costa M. Hypoxia-inducible factor-1 (HIF-1) Mol Pharmacol. 2006;70:1469–1480. doi: 10.1124/mol.106.027029. [DOI] [PubMed] [Google Scholar]
- 179.Smith L.E., Wesolowski E., McLellan A., Kostyk S.K., D’Amato R., Sullivan R., D’Amore P.A. Oxygen-induced retinopathy in the mouse. Invest. Ophthalmol. Vis. Sci. 1994;35:101–111. [PubMed] [Google Scholar]
- 180.Mowat F.M., Luhmann U.F., Smith A.J., Lange C., Duran Y., Harten S., Shukla D., Maxwell P.H., Ali R.R., Bainbridge J.W. HIF-1alpha and HIF-2alpha are differentially activated in distinct cell populations in retinal ischaemia. PLoS ONE. 2010;5:e11103. doi: 10.1371/journal.pone.0011103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Notari L., Miller A., Martinez A., Amaral J., Ju M., Robinson G., Smith L.E., Becerra S.P. Pigment epithelium-derived factor is a substrate for matrix metalloproteinase type 2 and type 9: implications for downregulation in hypoxia. Invest. Ophthalmol. Vis. Sci. 2005;46:2736–2747. doi: 10.1167/iovs.04-1489. [DOI] [PubMed] [Google Scholar]
- 182.Miller H., Miller B., Ishibashi T., Ryan S.J. Pathogenesis of laser-induced choroidal subretinal neovascularization. Invest. Ophthalmol. Vis. Sci. 1990;31:899–908. [PubMed] [Google Scholar]
- 183.Mori K., Gehlbach P., Yamamoto S., Duh E., Zack D.J., Li Q., Berns K.I., Raisler B.J., Hauswirth W.W., Campochiaro P.A. AAV-mediated gene transfer of pigment epithelium-derived factor inhibits choroidal neovascularization. Invest. Ophthalmol. Vis. Sci. 2002;43:1994–2000. [PubMed] [Google Scholar]
- 184.Hamilton M.M., Byrnes G.A., Gall J.G., Brough D.E., King C.R., Wei L.L. Alternate serotype adenovector provides long-term therapeutic gene expression in the eye. Mol. Vis. 2008;14:2535–2546. [PMC free article] [PubMed] [Google Scholar]
- 185.Liu Y., Cox S.R., Morita T., Kourembanas S. Hypoxia regulates vascular endothelial growth factor gene expression in endothelial cells. Identification of a 5′ enhancer. Circ. Res. 1995;77:638–643. doi: 10.1161/01.res.77.3.638. [DOI] [PubMed] [Google Scholar]
- 186.Shweiki D., Itin A., Soffer D., Keshet E. Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature. 1992;359:843–845. doi: 10.1038/359843a0. [DOI] [PubMed] [Google Scholar]
- 187.Zhang Z., Neiva K.G., Lingen M.W., Ellis L.M., Nor J.E. VEGF-dependent tumor angiogenesis requires inverse and reciprocal regulation of VEGFR1 and VEGFR2. Cell Death Differ. 2010;17:499–512. doi: 10.1038/cdd.2009.152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Cai J., Jiang W.G., Grant M.B., Boulton M. Pigment epithelium-derived factor inhibits angiogenesis via regulated intracellular proteolysis of vascular endothelial growth factor receptor 1. J. Biol. Chem. 2006;281:3604–3613. doi: 10.1074/jbc.M507401200. [DOI] [PubMed] [Google Scholar]
- 189.Kendall R.L., Thomas K.A. Inhibition of vascular endothelial cell growth factor activity by an endogenously encoded soluble receptor. Proc. Natl. Acad. Sci. U.S.A. 1993;90:10705–10709. doi: 10.1073/pnas.90.22.10705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Zou D., Zhang Z., He J., Zhang K., Ye D., Han W., Zhou J., Wang Y., Li Q., Liu X., et al. Blood vessel formation in the tissue-engineered bone with the constitutively active form of HIF-1alpha mediated BMSCs. Biomaterials. 2012;33:2097–2108. doi: 10.1016/j.biomaterials.2011.11.053. [DOI] [PubMed] [Google Scholar]
- 191.Calvani M., Rapisarda A., Uranchimeg B., Shoemaker R.H., Melillo G. Hypoxic induction of an HIF-1alpha-dependent bFGF autocrine loop drives angiogenesis in human endothelial cells. Blood. 2006;107:2705–2712. doi: 10.1182/blood-2005-09-3541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Zaichuk T.A., Shroff E.H., Emmanuel R., Filleur S., Nelius T., Volpert O.V. Nuclear factor of activated T cells balances angiogenesis activation and inhibition. J. Exp. Med. 2004;199:1513–1522. doi: 10.1084/jem.20040474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Ben-Yosef Y., Lahat N., Shapiro S., Bitterman H., Miller A. Regulation of endothelial matrix metalloproteinase-2 by hypoxia/reoxygenation. Circ. Res. 2002;90:784–791. doi: 10.1161/01.RES.0000015588.70132.DC. [DOI] [PubMed] [Google Scholar]
- 194.Rodrigues M., Xin X., Jee K., Babapoor-Farrokhran S., Kashiwabuchi F., Ma T., Bhutto I., Hassan S.J., Daoud Y., Baranano D., et al. VEGF secreted by hypoxic Muller cells induces MMP-2 expression and activity in endothelial cells to promote retinal neovascularization in proliferative diabetic retinopathy. Diabetes. 2013;62:3863–3873. doi: 10.2337/db13-0014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Cheng X.W., Kuzuya M., Kim W., Song H., Hu L., Inoue A., Nakamura K., Di Q., Sasaki T., Tsuzuki M., et al. Exercise training stimulates ischemia-induced neovascularization via phosphatidylinositol 3-kinase/Akt-dependent hypoxia-induced factor-1 alpha reactivation in mice of advanced age. Circulation. 2010;122:707–716. doi: 10.1161/CIRCULATIONAHA.109.909218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Yamamoto Y., Osanai T., Nishizaki F., Sukekawa T., Izumiyama K., Sagara S., Okumura K. Matrix metalloprotein-9 activation under cell-to-cell interaction between endothelial cells and monocytes: possible role of hypoxia and tumor necrosis factor-alpha. Heart Vessels. 2012;27:624–633. doi: 10.1007/s00380-011-0214-5. [DOI] [PubMed] [Google Scholar]
- 197.Bauer A.T., Burgers H.F., Rabie T., Marti H.H. Matrix metalloproteinase-9 mediates hypoxia-induced vascular leakage in the brain via tight junction rearrangement. J. Cereb. Blood Flow Metab. 2010;30:837–848. doi: 10.1038/jcbfm.2009.248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Hollborn M., Stathopoulos C., Steffen A., Wiedemann P., Kohen L., Bringmann A. Positive feedback regulation between MMP-9 and VEGF in human RPE cells. Invest. Ophthalmol. Vis. Sci. 2007;48:4360–4367. doi: 10.1167/iovs.06-1234. [DOI] [PubMed] [Google Scholar]
- 199.Kaluz S., Kaluzova M., Stanbridge E.J. Rational design of minimal hypoxia-inducible enhancers. Biochem. Biophys. Res. Commun. 2008;370:613–618. doi: 10.1016/j.bbrc.2008.03.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Patel N., Sundaram N., Yang M., Madigan C., Kalra V.K., Malik P. Placenta growth factor (PlGF), a novel inducer of plasminogen activator inhibitor-1 (PAI-1) in sickle cell disease (SCD) J. Biol. Chem. 2010;285:16713–16722. doi: 10.1074/jbc.M110.101691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Liao H., Hyman M.C., Lawrence D.A., Pinsky D.J. Molecular regulation of the PAI-1 gene by hypoxia: contributions of Egr-1, HIF-1alpha, and C/EBPalpha. FASEB J. 2007;21:935–949. doi: 10.1096/fj.06-6285com. [DOI] [PubMed] [Google Scholar]
- 202.Jung S.Y., Song H.S., Park S.Y., Chung S.H., Kim Y.J. Pyruvate promotes tumor angiogenesis through HIF-1-dependent PAI-1 expression. Int. J. Oncol. 2011;38:571–576. doi: 10.3892/ijo.2010.859. [DOI] [PubMed] [Google Scholar]
- 203.Matsui T., Higashimoto Y., Taira J., Yamagishi S. Pigment epithelium-derived factor (PEDF) binds to caveolin-1 and inhibits the pro-inflammatory effects of caveolin-1 in endothelial cells. Biochem. Biophys. Res. Commun. 2013;441:405–410. doi: 10.1016/j.bbrc.2013.10.074. [DOI] [PubMed] [Google Scholar]
- 204.Yamagishi S., Matsui T., Nakamura K., Takenaka K. Administration of pigment epithelium-derived factor prolongs bleeding time by suppressing plasminogen activator inhibitor-1 activity and platelet aggregation in rats. Clin. Exp. Med. 2009;9:73–76. doi: 10.1007/s10238-008-0010-4. [DOI] [PubMed] [Google Scholar]
- 205.Bai Y.J., Huang L.Z., Xu X.L., Du W., Zhou A.Y., Yu W.Z., Li X.X. Polyethylene glycol-modified pigment epithelial-derived factor: new prospects for treatment of retinal neovascularization. J. Pharmacol. Exp. Ther. 2012;342:131–139. doi: 10.1124/jpet.112.192575. [DOI] [PubMed] [Google Scholar]
- 206.Konson A., Pradeep S., Seger R. Phosphomimetic mutants of pigment epithelium-derived factor with enhanced antiangiogenic activity as potent anticancer agents. Cancer Res. 2010;70:6247–6257. doi: 10.1158/0008-5472.CAN-10-0434. [DOI] [PubMed] [Google Scholar]
- 207.Yang J., Chen S., Huang X., Han J., Wang Q., Shi D., Cheng R., Gao G., Yang X. Growth suppression of cervical carcinoma by pigment epithelium-derived factor via anti-angiogenesis. Cancer Biol. Ther. 2010;9:967–974. doi: 10.4161/cbt.9.12.11635. [DOI] [PubMed] [Google Scholar]
- 208.Ho T.C., Chen S.L., Yang Y.C., Liao C.L., Cheng H.C., Tsao Y.P. PEDF induces p53-mediated apoptosis through PPAR gamma signaling in human umbilical vein endothelial cells. Cardiovasc. Res. 2007;76:213–223. doi: 10.1016/j.cardiores.2007.06.032. [DOI] [PubMed] [Google Scholar]
- 209.Volpert O.V., Zaichuk T., Zhou W., Reiher F., Ferguson T.A., Stuart P.M., Amin M., Bouck N.P. Inducer-stimulated Fas targets activated endothelium for destruction by anti-angiogenic thrombospondin-1 and pigment epithelium-derived factor. Nat. Med. 2002;8:349–357. doi: 10.1038/nm0402-349. [DOI] [PubMed] [Google Scholar]
- 210.Ho T.C., Chen S.L., Yang Y.C., Lo T.H., Hsieh J.W., Cheng H.C., Tsao Y.P. Cytosolic phospholipase A2-α is an early apoptotic activator in PEDF-induced endothelial cell apoptosis. Am. J. Physiol. Cell Physiol. 2009;296:C273–C284. doi: 10.1152/ajpcell.00432.2008. [DOI] [PubMed] [Google Scholar]
- 211.Mori K., Gehlbach P., Ando A., McVey D., Wei L., Campochiaro P.A. Regression of ocular neovascularization in response to increased expression of pigment epithelium-derived factor. Invest. Ophthalmol. Vis. Sci. 2002;43:2428–2434. [PubMed] [Google Scholar]
- 212.Mirochnik Y., Aurora A., Schulze-Hoepfner F.T., Deabes A., Shifrin V., Beckmann R., Polsky C., Volpert O.V. Short pigment epithelial-derived factor-derived peptide inhibits angiogenesis and tumor growth. Clin. Cancer Res. 2009;15:1655–1663. doi: 10.1158/1078-0432.CCR-08-2113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Filleur S., Volz K., Nelius T., Mirochnik Y., Huang H., Zaichuk T.A., Aymerich M.S., Becerra S.P., Yap R., Veliceasa D., et al. Two functional epitopes of pigment epithelial-derived factor block angiogenesis and induce differentiation in prostate cancer. Cancer Res. 2005;65:5144–5152. doi: 10.1158/0008-5472.CAN-04-3744. [DOI] [PubMed] [Google Scholar]
- 214.Gong Q., Yang X., Cai W., Gao G., Yang Z. Expression and purification of functional epitope of pigment epithelium-derived factor in E. coli with inhibiting effect on endothelial cells. Protein J. 2010;29:167–173. doi: 10.1007/s10930-010-9236-6. [DOI] [PubMed] [Google Scholar]
- 215.Ju S.T., Panka D.J., Cui H., Ettinger R., el-Khatib M., Sherr D.H., Stanger B.Z., Marshak-Rothstein A. Fas(CD95)/FasL interactions required for programmed cell death after T-cell activation. Nature. 1995;373:444–448. doi: 10.1038/373444a0. [DOI] [PubMed] [Google Scholar]
- 216.Kaplan H.J., Leibole M.A., Tezel T., Ferguson T.A. Fas ligand (CD95 ligand) controls angiogenesis beneath the retina. Nat. Med. 1999;5:292–297. doi: 10.1038/6509. [DOI] [PubMed] [Google Scholar]
- 217.Barreiro R., Schadlu R., Herndon J., Kaplan H.J., Ferguson T.A. The role of Fas-FasL in the development and treatment of ischemic retinopathy. Invest. Ophthalmol. Vis. Sci. 2003;44:1282–1286. doi: 10.1167/iovs.02-0478. [DOI] [PubMed] [Google Scholar]
- 218.Chen L., Zhang S.S., Barnstable C.J., Tombran-Tink J. PEDF induces apoptosis in human endothelial cells by activating p38 MAP kinase dependent cleavage of multiple caspases. Biochem. Biophys. Res. Commun. 2006;348:1288–1295. doi: 10.1016/j.bbrc.2006.07.188. [DOI] [PubMed] [Google Scholar]
- 219.Doyon G., St-Jean S., Darsigny M., Asselin C., Boudreau F. Nuclear receptor co-repressor is required to maintain proliferation of normal intestinal epithelial cells in culture and down-modulates the expression of pigment epithelium-derived factor. J. Biol. Chem. 2009;284:25220–25229. doi: 10.1074/jbc.M109.022632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Tombran-Tink J., Lara N., Apricio S.E., Potluri P., Gee S., Ma J.X., Chader G., Barnstable C.J. Retinoic acid and dexamethasone regulate the expression of PEDF in retinal and endothelial cells. Exp. Eye Res. 2004;78:945–955. doi: 10.1016/j.exer.2003.12.013. [DOI] [PubMed] [Google Scholar]
- 221.Uchida H., Hayashi H., Kuroki M., Uno K., Yamada H., Yamashita Y., Tombran-Tink J., Kuroki M., Oshima K. Vitamin A up-regulates the expression of thrombospondin-1 and pigment epithelium-derived factor in retinal pigment epithelial cells. Exp. Eye Res. 2005;80:23–30. doi: 10.1016/j.exer.2004.08.004. [DOI] [PubMed] [Google Scholar]
- 222.Cheung L.W., Au S.C., Cheung A.N., Ngan H.Y., Tombran-Tink J., Auersperg N., Wong A.S. Pigment epithelium-derived factor is estrogen sensitive and inhibits the growth of human ovarian cancer and ovarian surface epithelial cells. Endocrinology. 2006;147:4179–4191. doi: 10.1210/en.2006-0168. [DOI] [PubMed] [Google Scholar]
- 223.Leung K.W., Cheung L.W., Pon Y.L., Wong R.N., Mak N.K., Fan T.P., Au S.C., Tombran-Tink J., Wong A.S. Ginsenoside Rb1 inhibits tube-like structure formation of endothelial cells by regulating pigment epithelium-derived factor through the oestrogen beta receptor. Br. J. Pharmacol. 2007;152:207–215. doi: 10.1038/sj.bjp.0707359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Li C., Tang Y., Li F., Turner S., Li K., Zhou X., Centola M., Yan X., Cao W. 17β-Estradiol (betaE2) protects human retinal Muller cell against oxidative stress in vitro: evaluation of its effects on gene expression by cDNA microarray. Glia. 2006;53:392–400. doi: 10.1002/glia.20291. [DOI] [PubMed] [Google Scholar]
- 225.Chuderland D., Ben-Ami I., Friedler S., Hasky N., Ninio-Many L., Goldberg K., Bar-Joseph H., Grossman H., Shalgi R. Hormonal regulation of pigment epithelium-derived factor (PEDF) expression in the endometrium. Mol. Cell Endocrinol. 2014;390:85–92. doi: 10.1016/j.mce.2014.04.006. [DOI] [PubMed] [Google Scholar]
- 226.Parvathaneni K., Grigsby J.G., Betts B.S., Tsin A.T. Estrogen-induced retinal endothelial cell proliferation: possible involvement of pigment epithelium-derived factor and phosphoinositide 3-kinase/mitogen-activated protein kinase pathways. J. Ocul. Pharmacol. Ther. 2013;29:27–32. doi: 10.1089/jop.2011.0252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Zhou Y., Xu F., Deng H., Bi Y., Sun W., Zhao Y., Chen Z., Weng J. PEDF expression is inhibited by insulin treatment in adipose tissue via suppressing 11beta-HSD1. PLoS ONE. 2013;8:e84016. doi: 10.1371/journal.pone.0084016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Tong J.P., Lam D.S., Chan W.M., Choy K.W., Chan K.P., Pang C.P. Effects of triamcinolone on the expression of VEGF and PEDF in human retinal pigment epithelial and human umbilical vein endothelial cells. Mol. Vis. 2006;12:1490–1495. [PubMed] [Google Scholar]
- 229.Perruccio E.M., Rowlette L.L., Comes N., Locatelli-Hoops S., Notari L., Becerra S.P., Borras T. Dexamethasone increases pigment epithelium-derived factor in perfused human eyes. Curr. Eye Res. 2008;33:507–515. doi: 10.1080/02713680802110208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Fernandez-Barral A., Orgaz J.L., Baquero P., Ali Z., Moreno A., Tiana M., Gomez V., Riveiro-Falkenbach E., Canadas C., Zazo S., et al. Regulatory and functional connection of microphthalmia-associated transcription factor and anti-metastatic pigment epithelium derived factor in melanoma. Neoplasia. 2014;16:529–542. doi: 10.1016/j.neo.2014.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Ma X., Pan L., Jin X., Dai X., Li H., Wen B., Chen Y., Ma A., Qu J., Hou L. Microphthalmia-associated transcription factor acts through PEDF to regulate RPE cell migration. Exp. Cell Res. 2012;318:251–261. doi: 10.1016/j.yexcr.2011.11.002. [DOI] [PubMed] [Google Scholar]
- 232.Petersen S.V., Valnickova Z., Enghild J.J. Pigment-epithelium-derived factor (PEDF) occurs at a physiologically relevant concentration in human blood: purification and characterization. Biochem. J. 2003;374:199–206. doi: 10.1042/BJ20030313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Maik-Rachline G., Shaltiel S., Seger R. Extracellular phosphorylation converts pigment epithelium-derived factor from a neurotrophic to an antiangiogenic factor. Blood. 2005;105:670–678. doi: 10.1182/blood-2004-04-1569. [DOI] [PubMed] [Google Scholar]
- 234.Maik-Rachline G., Seger R. Variable phosphorylation states of pigment-epithelium-derived factor differentially regulate its function. Blood. 2006;107:2745–2752. doi: 10.1182/blood-2005-06-2547. [DOI] [PubMed] [Google Scholar]
- 235.Duh E.J., Yang H.S., Suzuma I., Miyagi M., Youngman E., Mori K., Katai M., Yan L., Suzuma K., West K., et al. Pigment epithelium-derived factor suppresses ischemia-induced retinal neovascularization and VEGF-induced migration and growth. Invest. Ophthalmol. Vis. Sci. 2002;43:821–829. [PubMed] [Google Scholar]
- 236.Subramanian P., Deshpande M., Locatelli-Hoops S., Moghaddam-Taaheri S., Gutierrez D., Fitzgerald D.P., Guerrier S., Rapp M., Notario V., Becerra S.P. Identification of pigment epithelium-derived factor protein forms with distinct activities on tumor cell lines. J. Biomed. Biotechnol. 2012;2012:425907. doi: 10.1155/2012/425907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Wang S., Gottlieb J.L., Sorenson C.M., Sheibani N. Modulation of thrombospondin 1 and pigment epithelium-derived factor levels in vitreous fluid of patients with diabetes. Arch. Ophthalmol. 2009;127:507–513. doi: 10.1001/archophthalmol.2009.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Shao H., Schvartz I., Shaltiel S. Secretion of pigment epithelium-derived factor. Mutagenic study. Eur. J. Biochem. 2003;270:822–831. doi: 10.1046/j.1432-1033.2003.03374.x. [DOI] [PubMed] [Google Scholar]
- 239.Xiao Q., Zeng S., Lv M., Ling S. Small hairpin loop RNA targeting HIF-1alpha down-regulates VEGF and up-regulates PEDF in human retinal pigment epithelial cells under hypoxic condition. J. Huazhong. Univ. Sci. Technolog. Med. Sci. 2008;28:460–464. doi: 10.1007/s11596-008-0419-8. [DOI] [PubMed] [Google Scholar]
- 240.Bandyopadhyay M., Rohrer B. Matrix metalloproteinase activity creates pro-angiogenic environment in primary human retinal pigment epithelial cells exposed to complement. Invest. Ophthalmol. Vis. Sci. 2012;53:1953–1961. doi: 10.1167/iovs.11-8638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Fernandez-Barral A., Orgaz J.L., Gomez V., del Peso L., Calzada M.J., Jimenez B. Hypoxia negatively regulates antimetastatic PEDF in melanoma cells by a hypoxia inducible factor-independent, autophagy dependent mechanism. PLoS ONE. 2012;7:e32989. doi: 10.1371/journal.pone.0032989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Rasmussen H., Chu K.W., Campochiaro P., Gehlbach P.L., Haller J.A., Handa J.T., Nguyen Q.D., Sung J.U. Clinical protocol. An open-label, phase I, single administration, dose-escalation study of ADGVPEDF.11D (ADPEDF) in neovascular age-related macular degeneration (AMD) Hum. Gene. Ther. 2001;12:2029–2032. [PubMed] [Google Scholar]
- 243.Conti A., Ricchiuto P., Iannaccone S., Sferrazza B., Cattaneo A., Bachi A., Reggiani A., Beltramo M., Alessio M. Pigment epithelium-derived factor is differentially expressed in peripheral neuropathies. Proteomics. 2005;5:4558–4567. doi: 10.1002/pmic.200402088. [DOI] [PubMed] [Google Scholar]
- 244.Craik D.J., Fairlie D.P., Liras S., Price D. The future of peptide-based drugs. Chem. Biol. Drug Des. 2013;81:136–147. doi: 10.1111/cbdd.12055. [DOI] [PubMed] [Google Scholar]
- 245.Deshpande M., Notari L., Subramanian P., Notario V., Becerra S.P. Inhibition of tumor cell surface ATP synthesis by pigment epithelium-derived factor: implications for antitumor activity. Int. J. Oncol. 2012;41:219–227. doi: 10.3892/ijo.2012.1431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Becerra S.P., Palmer I., Kumar A., Steele F., Shiloach J., Notario V., Chader G.J. Overexpression of fetal human pigment epithelium-derived factor in Escherichia coli. A functionally active neurotrophic factor. J. Biol. Chem. 1993;268:23148–23156. [PubMed] [Google Scholar]
- 247.Bilak M.M., Becerra S.P., Vincent A.M., Moss B.H., Aymerich M.S., Kuncl R.W. Identification of the neuroprotective molecular region of pigment epithelium-derived factor and its binding sites on motor neurons. J. Neurosci. 2002;22:9378–9386. doi: 10.1523/JNEUROSCI.22-21-09378.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Smith N.D., Schulze-Hoepfner F.T., Veliceasa D., Filleur S., Shareef S., Huang L., Huang X.M., Volpert O.V. Pigment epithelium-derived factor and interleukin-6 control prostate neuroendocrine differentiation via feed-forward mechanism. J. Urol. 2008;179:2427–2434. doi: 10.1016/j.juro.2008.01.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Sanagi T., Yabe T., Yamada H. Adenoviral gene delivery of pigment epithelium-derived factor protects striatal neurons from quinolinic acid-induced excitotoxicity. J. Neuropathol. Exp. Neurol. 2010;69:224–233. doi: 10.1097/NEN.0b013e3181cfc46f. [DOI] [PubMed] [Google Scholar]
- 250.Li H., Tran V.V., Hu Y., Mark Saltzman W., Barnstable C.J., Tombran-Tink J. A PEDF N-terminal peptide protects the retina from ischemic injury when delivered in PLGA nanospheres. Exp. Eye Res. 2006;83:824–833. doi: 10.1016/j.exer.2006.04.014. [DOI] [PubMed] [Google Scholar]
- 251.Longeras R., Farjo K., Ihnat M., Ma J.X. A PEDF-derived peptide inhibits retinal neovascularization and blocks mobilization of bone marrow-derived endothelial progenitor cells. Exp. Diabetes Res. 2012;2012:518426. doi: 10.1155/2012/518426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Esipov R.S., Beirakhova K.A., Chupova L.A., Likhvantseva V.K., Stepanova E.V., Miroshnikov A.I. Recombinant fragment of pigment epithelium-derived factor (44–77) prevents pathological corneal neovascularization. Bioorg. Khim. 2012;38:78–85. doi: 10.1134/s1068162012010074. [DOI] [PubMed] [Google Scholar]
- 253.Liu H., Ren J.G., Cooper W.L., Hawkins C.E., Cowan M.R., Tong P.Y. Identification of the antivasopermeability effect of pigment epithelium-derived factor and its active site. Proc. Natl. Acad. Sci. U.S.A. 2004;101:6605–6610. doi: 10.1073/pnas.0308342101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Sanchez-Sanchez F., Aroca-Aguilar J.D., Segura I., Ramirez-Castillejo C., Riese H.H., Coca-Prados M., Escribano J. Expression and purification of functional recombinant human pigment epithelium-derived factor (PEDF) secreted by the yeast Pichia pastoris. J. Biotechnol. 2008;134:193–201. doi: 10.1016/j.jbiotec.2008.01.005. [DOI] [PubMed] [Google Scholar]
- 255.Sanagi T., Yabe T., Yamada H. The regulation of pro-inflammatory gene expression induced by pigment epithelium-derived factor in rat cultured microglial cells. Neurosci. Lett. 2005;380:105–110. doi: 10.1016/j.neulet.2005.01.035. [DOI] [PubMed] [Google Scholar]
- 256.Takanohashi A., Yabe T., Schwartz J.P. Pigment epithelium-derived factor induces the production of chemokines by rat microglia. Glia. 2005;51:266–278. doi: 10.1002/glia.20203. [DOI] [PubMed] [Google Scholar]
- 257.Yabe T., Sanagi T., Schwartz J.P., Yamada H. Pigment epithelium-derived factor induces pro-inflammatory genes in neonatal astrocytes through activation of NF-kappa B and CREB. Glia. 2005;50:223–234. doi: 10.1002/glia.20171. [DOI] [PubMed] [Google Scholar]
- 258.Li S., Fu X.A., Zhou X.F., Chen Y.Y., Chen W.Q. Angiogenesis-related cytokines in serum of proliferative diabetic retinopathy patients before and after vitrectomy. Int. J. Ophthalmol. 2012;5:726–730. doi: 10.3980/j.issn.2222-3959.2012.06.14. [DOI] [PMC free article] [PubMed] [Google Scholar]