

Abstract
The urinary tract environment provides many conditions that deter colonization by microorganisms. D-serine is thought to be one of these stressors and is present at high concentrations in urine. D-serine interferes with L-serine and pantothenate metabolism and is bacteriostatic to many species. Uropathogenic Escherichia coli commonly possess the dsdCXA genetic locus, which allows them to use D-serine as a sole carbon, nitrogen, and energy source. It was previously reported that in the model UPEC strain CFT073, a dsdA mutant outcompetes wild type in the murine model of urinary tract infection. This “hypercolonization” was used to propose a model whereby UPEC strains sense D-serine in the urinary tract and subsequently up-regulate genes necessary for pathogenesis. Here, we show that inactivation of dsdA does not lead to hypercolonization. We suggest that this previously observed effect is due to an unrecognized secondary mutation in rpoS and that some D-serine specific effects described in other studies may be affected by the rpoS status of the strains used. Inactivation of dsdA in the original clinical isolate of CFT073 gives CFT073 ΔdsdA a growth defect in human urine and renders it unable to grow on minimal medium containing D-serine as the sole carbon source. However, CFT073 ΔdsdA is able to colonize the urinary tracts of CBA/J mice indistinguishably from wild type. These findings indicate that D-serine catabolism, though it may play role(s) during urinary tract infection, does not affect the ability of uropathogenic E. coli to colonize the murine urinary tract.
Introduction
Urinary tract infections (UTIs) represent a major public health concern, as they are among the most common human bacterial infections. Forty percent of women and 12% of men will experience a UTI in their lifetimes [1, 2]. Uropathogenic Escherichia coli (UPEC) is the most common causative agent of UTIs, representing up to 95% of reported community-acquired cases [3]. UPEC are members of the gut microbiota and gain access to the urinary tract via an ascending route [4]. During this ascent, the bacteria first colonize the periurethral area and traverse the urethra into the bladder, where they cause cystitis. Cystitis can progress to pyelonephritis after the bacteria ascend the ureters into the kidneys of the infected individual.
Although UTIs are common, many conditions in the urinary tract discourage the colonization and growth of microorganisms. Among these stresses are host immune components, physical deterrents, and the chemical composition of urine [5]. D-serine is one of the most abundant amino acids in mammalian urine, where it is present at concentrations from 3–115 μg/ml [6, 7]. D-serine is bacteriostatic, presumably by interfering with pantothenate and L-serine metabolism [8]. High concentrations of D-serine (0.22–14.7 μg/g) are also found in the brain [9], where D-serine acts as a co-agonist for N-methyl-D-aspartate receptors, which are involved in neural transmission and cognitive function [10, 11].
UPEC strains and neonatal meningitis E. coli (NMEC) strains frequently have the dsdCXA locus [12], which allows E. coli to use D-serine as the sole carbon, nitrogen, and energy source [13]. DsdC is a LysR-type transcriptional regulator [14] that positively affects the transcription of dsdXA in the presence of D-serine but in the absence of D-serine, dsdC transcription is autorepressed [15–18]. DsdX is a D-serine transporter [19] that belongs to the gluconate permease family of transport proteins [20]. In addition to DsdX, the D-alanine and glycine transporter, CycA, is capable of transporting D-serine [21, 22] and no other D-serine transporters are known in E. coli. DsdA is a pyridoxal 5’-phosphate-requiring deaminase that is capable of deaminating the D-amino acids D-serine, D-threonine, and D-allothreonine, though its activity is highest towards D-serine and it shows very little activity towards L-enantiomers [23–27].
Based on its commonality in UPEC/NMEC and the ability of DsdC, DsdX, and DsdA to sense, transport, and metabolize D-serine, the dsdCXA locus was hypothesized to be important for UPEC/NMEC pathogenesis [12]. Counterintuitively, mutational inactivation of dsdA gives rise to a hypercolonization phenotype in UPEC strain CFT073, where a dsdA mutant is recovered more frequently than wild type in the bladders and kidneys of CBA/J mice at 48 hours post infection of a 1:1 ratio of these strains [28]. This hypercolonization was used to propose a model whereby UPEC senses D-serine as a urinary tract-specific signal to up-regulate genes needed for pathogenesis. Indeed, UPEC use flagella-mediated motility to ascend the urinary tract during infection [29] and the dsdA mutant expresses more flagella and is more motile relative to wild type [28]. Additionally, many other virulence determinants including pap pili and hemolysin, are differentially regulated between these strains during UTI [30].
Other effectors of serine metabolism were described to affect urinary tract colonization by CFT073. In addition to a dsdA mutant, dsdC mutants hypercolonize relative to wild type, suggesting that there are mechanisms of D-serine dependent virulence gene expression that are independent of DsdC [31]. SdaA and SdaB are L-serine deaminases made by E. coli [32]. A dsdA sdaAB triple mutant is outcompeted by wild type in the bladders and kidneys during murine model UTI, suggesting that DL-serine catabolism is important for colonization of the urinary tract [31]. D-serine transport affects colonization by CFT073 as well: a cycA dsdXA triple mutant, which is unable to transport or degrade D-serine, shows a competitive defect in the kidneys of CBA/J mice. This suggests that the expression of traits needed for the colonization of kidneys is aided by the uptake of D-serine [31].
D-serine metabolism is also important for UTI by Staphylococcus saprophyticus [33]. Unlike UPEC isolates, S. saprophyticus does not have the dsdCXA locus. It has dsdA, which is apparently under the control of its own promoter. S. saprophyticus ΔdsdA is outcompeted by wild type in the bladders and kidneys of C3H/HeN mice. Furthermore, in S. saprophyticus, D-serine can be used as the sole carbon source during growth, D-serine metabolism provides a growth advantage in vitro, and the expression of the lipase ssp [34], is up-regulated in the presence of D-serine. Although UPEC and S. saprophyticus both have dsdA, it appears that the accumulation of D-serine in each of these uropathogens during infection has largely different effects. Aside from UPEC and S. saprophyticus, D-serine metabolism has not been characterized in any other uropathogens.
Transcription in E. coli is catalyzed by RNA polymerase holoenzyme, which is comprised of core polymerase and a dissociable sigma factor. E. coli has one housekeeping sigma factor (σ70) and six alternative sigma factors [35]. Core polymerase is unable to initiate transcription alone and requires an associated sigma factor to define promoter specificity and initiate transcription. RpoS (σS), the best studied of the alternative sigma factors, affects the expression of ~10% of genes in E. coli K-12, either directly or indirectly [36]. Mutations in rpoS have been demonstrated in accordance with the transfer of E. coli strains between laboratories, which is likely due to the growth advantage in stationary phase (GASP) phenotype afforded by loss of function mutations in rpoS [37–39].
Here, we show that the strains used in the characterization of the dsdA hypercolonization phenotype in CFT073 are not isogenic for rpoS. In the original clinical isolate of CFT073, where rpoS is intact, mutational inactivation of dsdA has no effect on the ability of CFT073 to colonize the murine urinary tract. We propose additional experiments that will allow for a better understanding of the role that D-serine plays in UPEC pathogenesis.
Results
The sequenced CFT073 strain has a frameshift mutation in rpoS
We previously reported the genome sequence of UPEC strain CFT073 [40], which was isolated from the blood of a pyelonephritis patient in the late 1980s [41]. Upon examination of the CFT073 genome sequence, we observed that this isolate of CFT073 has no annotated rpoS gene. However, two annotated ORFs from this strain (c3307 and c3306) each share 99% nucleotide sequence identity with rpoS from K-12. Upon further examination of these ORFs, we noticed that there is a 5 base-pair duplication (TAGAG) at the 3’ end of c3307, which leads to a frameshift (amber mutation) in the rpoS gene of this strain. Herein, we refer to the sequenced CFT073 isolate as CFT073 rpoS am to reflect this amber mutation.
We determined the nucleotide sequence of the rpoS allele from CFT073 rpoS am and from a spontaneous nalidixic acid resistant mutant of CFT073 (CFT073 gyrA S83L) [42] via Sanger sequencing. Because CFT073 gyrA S83L colonizes mice indistinguishably from its nalS parent [42], CFT073 gyrA S83L strain was previously used as wild type to facilitate enumeration of wild type/mutant ratios during competitive murine model UTI [28, 43]. We found that CFT073 rpoS am, but not CFT073 gyrA S83L, has the defective rpoS am allele (Fig 1A).
Fig 1. A frameshift mutation is present in rpoS from CFT073 rpoS am and strain derivatives thereof.

(A) The rpoS allele from CFT073 (WAM4505), CFT073 gyrA S83L (WAM2266), and CFT073 rpoS am (WAM2267) was sequenced via Sanger sequencing using the “rpoS sequencing F” and “rpoS sequencing R” primers listed in Table 1 and an alignment of the rpoS coding regions from these strains was performed using the ClustalW function in MacVector 9.0.2. (B) Dendrogram showing relevant strains used in this study (Also listed in Table 1) and their applicable characteristics. Strains mentioned in Panel A are outlined with a double black box.
Because rpoS mutations arise after prolonged culture in stationary phase and passage between laboratories, we wanted to determine if the original patient isolate of CFT073 has a functional rpoS allele. We obtained this isolate from Harry L. T. Mobley (University of Michigan) and showed that rpoS is intact in this strain (Fig 1A). We have since deposited the original rpoS + patient isolate of CFT073 in the American Type Culture Collection as ATCC# BAA-2503. This strain is intended to replace ATCC# 700928 (CFT073 rpoS am) as wild type in all future research. Fig 1B shows a dendrogram of strains used in this study, created based on differences relative to the original patient isolate.
Hypercolonization is independent of dsdA
Because the dsdA mutant and wild type strains used to describe the dsdA hypercolonization phenotype are not isogenic for rpoS, we investigated whether rpoS affects the dsdA hypercolonization phenotype. To this end, we subjected CBA/J mice to competitive UTI using strains containing dsdA and rpoS am mutations. We then analyzed bacterial burdens in their bladders and kidneys at 48 hpi (Fig 2). First, we showed that CFT073 ΔdsdA rpoS am competed equally well with an rpoS am strain in the bladders of these animals. As was observed previously [28], CFT073 dsdA Δ445bp gyrA S83L outcompeted an rpoS am strain in the bladders (Fig 2A). However, the dsdA + parental strain (CFT073 gyrA S83L) also outcompeted an rpoS am strain in the bladders and kidneys (Fig 2). CFT073 dsdA Δ445bp gyrA S83L did not outcompete an rpoS am strain in the kidneys. However, kidney RCIs from the two competitive infection experiments using CFT073 ΔdsdA rpoS am and CFT073 gyrA S83L demonstrate that the lack of dsdA does not provide a fitness advantage in this niche (Fig 2B).
Fig 2. Inactivation of dsdA does not engender a competitive advantage in CBA/J mice during experimental UTI.

CFT073 ΔlacZYA rpoS am and CFT073 ΔdsdA rpoS am or CFT073 dsdA Δ445bp gyrA S83L or CFT073 gyrA S83L were co-inoculated at a 1:1 ratio into CBA/J mice (n = 16, 8, and 18, respectively). Mice were sacrificed at 48hpi. Bacteria from (A) bladder and (B) kidney homogenates were enumerated on MacConkey’s medium plus lactose. Several mice from the co-infections had no detectable bacteria in their kidneys (4 of 16, 5 of 8, and 3 of 18, respectively). Lines are drawn at the geometric mean relative competitive index (RCI). Statistical Significance was assessed by a Wilcoxon signed-rank test relative to a hypothetical RCI of 1.
We deleted dsdA in the original patient isolate of CFT073 and subjected it to analysis via murine model UTI as above (Fig 3A). CFT073 ΔdsdA does not outcompete wild type in the bladders or kidneys of CBA/J mice at 48hpi. Additionally, there is no difference in bladder or kidney burdens in mice subjected to single-strain challenge with CFT073 or CFT073 ΔdsdA (Fig 3B).
Fig 3. CFT073 ΔdsdA colonizes the murine urinary tract indistinguishably from wild type.
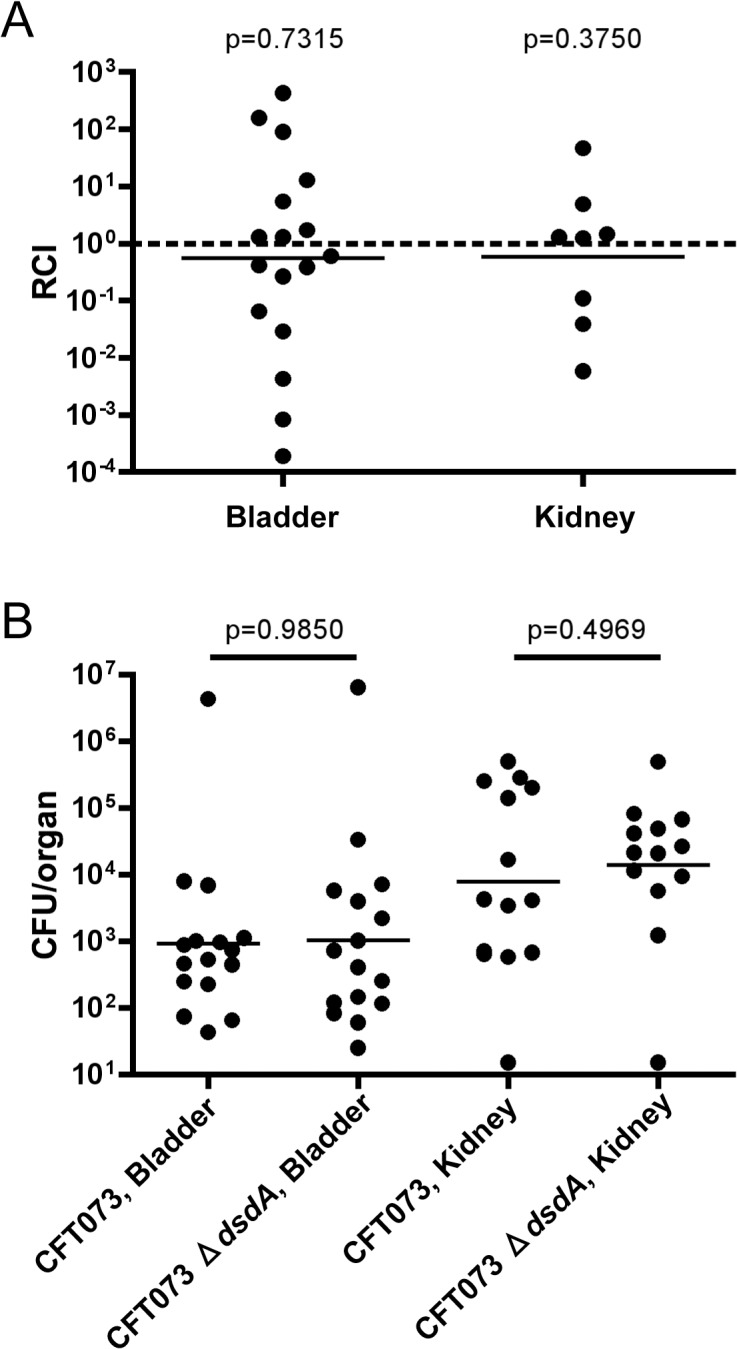
(A) CFT073 ΔdsdA and CFT073 ΔlacZYA were co-inoculated at a 1:1 ratio into CBA/J mice (n = 17). Bacteria from bladder and kidney homogenates were enumerated on MacConkey’s medium plus lactose. Lines are drawn at the geometric mean relative competitive index (RCI). Nine mice had no detectable bacteria in their kidneys. Statistical Significance was assessed by a Wilcoxon signed-rank test relative to a hypothetical RCI of 1. (B) Manipulations were carried out as described for panel A except that single strains were used (n = 16 for each). Two mice infected with CFT073 had no detectable bacteria in their kidneys and three mice infected with CFT073 ΔdsdA had no detectable bacteria in their kidneys. Lines are drawn at the geometric mean CFU/organ. Statistical significance was assessed by the Mann-Whitney U test.
dsdA does not affect motility in CFT073
We analyzed the motility of the dsdA mutants and their parental strains by measuring diameters of migration on Adler’s motility medium (S1 Fig), as described in Materials and Methods. Zones of migration from five independent spots of each dsdA + strain (CFT073, CFT073 gyrA S83L, and CFT073 rpoS am) and each dsdA - strain (CFT073 ΔdsdA, CFT073 dsdA Δ445bp gyrA S83L, and CFT073 ΔdsdA rpoS am) were measured. It was determined that dsdA does not affect motility under these conditions. Under the conditions tested, strains containing the rpoS am allele are significantly less motile than strains that are wild type for rpoS (S1 Fig).
dsdA affects the growth of CFT073 in human urine
It was previously shown that a dsdA mutant has a prolonged lag phase relative to an rpoS mutant when grown in human urine. This lag phase was associated with a significant reduction in viable bacterial counts. However, an increased growth rate was observed at 10–12 hours post inoculation, with equal viable CFUs between strains at 11 hours post inoculation [28]. To reproduce the growth defect that dsdA - strains have when grown in urine, we grew dsdA + strains (CFT073, CFT073 gyrA S83L, and CFT073 rpoS am) and dsdA - strains (CFT073 ΔdsdA, CFT073 dsdA Δ445bp gyrA S83L, and CFT073 ΔdsdA rpoS am) in filter sterilized human urine. All dsdA - strains had comparably reduced steady-state growth rate but achieved culture density indistinguishable from that of dsdA + strains after 5 hours (Fig 4). Additionally, the dsdA - strains were unable to grow on MOPS minimal medium containing D-serine as the sole carbon source (S2 Fig).
Fig 4. dsdA - strains have a growth defect in human urine.

Bacteria were grown overnight in pooled, filter sterilized human urine. Bacteria were washed 2x in phosphate buffered saline and the OD600 of each cell suspension was normalized to OD600 = 1.5. Bacteria were then inoculated into fresh, pre-warmed urine to OD600 = 0.03. Bacteria were allowed to grow for 12 hours. OD600 (Panel A) and viable counts (Panel B) were measured at the time points indicated above. dsdA + strains used in this analysis are CFT073, CFT073 gyrA S83L, and CFT073 rpoS am, and dsdA - strains are CFT073 ΔdsdA, CFT073 dsdA Δ445bp gyrA S83L, and CFT073 ΔdsdA rpoS am. Data points represent mean OD600 and mean viable counts for each strain type where applicable and error bars are drawn to represent standard error of the mean (±SEM).
Additional effects of D-serine transport and DL-serine metabolism on colonization during murine model UTI
Previously, Anfora et al. reported that D-serine transport and DL-serine catabolism were important for colonization of the murine urinary tract [31]. First, in the rpoS am background, a strain unable to transport or degrade D-serine showed a competitive defect in the kidneys of CBA/J mice relative to the parent strain. It was hypothesized that the expression of traits needed for the colonization of kidneys is aided by the uptake of D-serine. To further address this hypothesis in the wild type CFT073 background, we co-challenged CBA/J mice with CFT073 ΔcycA ΔdsdX and CFT073 ΔlacZYA. At 48 hpi, CFT073 ΔcycA ΔdsdX is outcompeted by wild type 1.67-fold but there is no difference in colonization of the kidneys in these animals (S3 Fig). Anfora et al. also reported in that study that a strain unable to catabolize DL-serine is outcompeted by an rpoS mutant, suggesting that DL-serine catabolism is needed by UPEC during UTI. We therefore addressed the importance of DL-serine catabolism in the wild type CFT073 background where, as is shown in S3 Fig, a strain defective for DL-serine catabolism shows a competitive defect in the bladders and kidneys of these animals during experimental UTI.
Discussion
We discovered that the “wild type” CFT073 strains previously used in our laboratory were either rpoS + (CFT073 gyrA S83L and derivatives) or had a frameshift mutation in rpoS (CFT073 rpoS am and derivatives) and that the original patient isolate of CFT073 has a functional rpoS allele (Fig 1). Subsequently, using strains isogenic for rpoS, we were unable to reproduce the dsdA hypercolonization phenotype that was observed in our laboratory by Roesch et al. [28]. Using murine model UTI, we determined that dsdA neither positively nor negatively affects urinary tract colonization by CFT073 (Figs 2 and 3). We also show that dsdA does not affect motility (S1 Fig) and that dsdA - strains have a growth defect in urine relative to dsdA + strains (Fig 4). The urine-specific growth defect that we observe here is not as pronounced as what was described previously. We suspect that this is due to the inconsistent composition of urine between collections. Indeed, there is inter- and intra-personal variation in the chemical composition of urine with respect to D-serine concentrations (3–115 μg/ml) and for other urine components [6, 7, 44]. We suggest here that the findings from the previous study by Roesch et al. are due to defects by a strain with the non-functional rpoS am allele. Additionally, Anfora et al. showed that CFT073 mutants unable to sense or metabolize D-serine have a competitive advantage over a CFT073 rpoS am strain at 48hpi in murine model UTI [31]. We expect that the hypercolonization by these strains is also due to rpoS and not the lack of dsdC or dsdA.
Our findings affect the interpretation of work by Haugen et al., where genes needed for hypercolonization by a dsdA mutant over an rpoS mutant were identified [30]. Because the strains used in that study were not isogenic for rpoS ([30] and Fig 1), it is unclear which genes are differentially regulated in a σS-dependent, DsdA-dependent, or σS/DsdA co-dependent fashion. Interestingly, our data suggest that a σS-dependent mechanism of mitigating the bacteriostatic effects of D-serine is utilized by UPEC during pyelonephritis. Specifically, in Figs 2B and 3A, we demonstrate that a dsdA is important for kidney infection, exclusively in an rpoS mutant background. As such, a comparison of the transcriptomes of CFT073, CFT073 ΔrpoS, and CFT073 ΔdsdA during experimental UTI will allow for a better understanding of these effects.
The roles played by DL-serine metabolism and D-serine transport during UTI were also investigated by Anfora et al. [31] using strains not isogenic for rpoS. There, it was found that intracellular accumulation of DL-serine, but not the accumulation D-serine or L-serine alone, negatively affects UPEC fitness during UTI. Using strains that are isogenic for rpoS, we demonstrate that a dsdA sdaAB triple mutant is outcompeted by wild type in the bladders and kidneys of CBA/J mice (S3 Fig), further supporting the findings that DL-serine catabolism, but not D-serine catabolism alone is needed during UTI [31]. Also, using a pair of strains both containing the non-functional rpoS am allele, it was shown that a strain unable to transport D-serine [19] competed equally well with its parent strain in the bladders but was at a competitive defect in the kidneys of CBA/J mice at 48hpi. These results suggested that the expression of traits needed for kidney colonization was aided by the uptake of D-serine. However, we demonstrate that CFT073 strains containing the functional rpoS allele, the ability to transport D-serine is dispensable for colonization of bladders and kidneys during competitive murine UTI (S3 Fig). This provides evidence that the previously observed defect in kidney colonization was influenced by the rpoS am allele and it supports our hypothesis that a σS-dependent mechanism of mitigating the effects of D-serine is utilized by UPEC during pyelonephritis.
This study directly addresses phenotypes reported by Roesch et al. in 2003 and by Anfora et al. in 2007 [28, 31] using the original clinical isolate of CFT073 and isogenic mutants. We extend our findings to inform further commentary on the studies by Anfora et al. and Haugen et al. [30, 31]. We provide evidence that many of the conclusions drawn in these studies are more accurately described in the context of the rpoS status of the strains used. As functional σS is needed during UTI [45], we suggest that the previously described hypercolonization phenotype of a dsdA - strain is more accurately described as a colonization defect by an rpoS - strain. The results of this work led us to study the roles and regulation of rpoS, which we continue to investigate in UPEC and other E. coli pathotypes [46].
Materials and Methods
Ethics Statement
This study was done in strict agreement with the recommendations found in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The murine model UTI protocol was approved by the UW-Madison Animal Care and Use Committee (Permit Number: M00450-0-07-08). All efforts were made to minimize suffering. Human urine samples were collected from volunteers who gave written consent. Permission for collection of human urine was obtained from the University of Wisconsin Health Sciences Institutional Review Board (IRB). All samples were obtained and used at the University of Wisconsin-Madison.
Strains, plasmids, and oligonucleotides
Strains, plasmids, and oligonucleotides used in this study are listed in Table 1. PCR amplification of DNA was done using GoTaq (Promega) according to the manufacturer’s specifications. CFT073 ΔdsdA was constructed using the Lambda Red mutagnesis protocol [47], which was modified to incorporate a generalized transduction step using ΦEB49 [48] prior to pCP20-mediated antibiotic resistance cassette removal, as described previously [45]. CFT073 ΔcycA ΔdsdX was constructed by ΦEB49-mediated generalized transduction of cycA::KmR from CFT073 cycA::KmR rpoS am and dsdX::CmR from CFT073 dsdX::CmR rpoS am into CFT073, followed by pCP20-mediated antibiotic cassette removal. CFT073 ΔdsdA sdaA::CmR sdaB::KmR was constructed by ΦEB49-mediated generalized transduction of sdaA::CmR and sdaB::KmR from CFT073 sdaA::CmR sdaB::KmR rpoS am into CFT073 ΔdsdA.
Table 1. Strains, plasmids, and oligonucleotides used in this study.
| Strain, plasmid, or oligo | Relevant characteristics or sequence | Source |
|---|---|---|
| WAM2266 | CFT073 gyrA S83L, nalR | H. Mobley |
| WAM2267 | CFT073 rpoS am | H. Mobley |
| WAM2615 | WAM2266 derivative, dsdA Δ445bp gyrA S83L, nalR | [28] |
| WAM2880 | WAM2267 derivative, CFT074 ΔlacZYA rpoS am | [50] |
| WAM2914 | WAM2267 derivative, CFT073 ΔdsdA rpoS am | P. Roesch |
| WAM2966 | WAM2267 derivative, CFT073 dsdX::CmR rpoS am | A. Anfora |
| WAM3707 | WAM2267 derivative, CFT073 sdaA::CmR sdaB::KmR rpoS am | A. Anfora |
| WAM4248 | WAM2267 derivative, CFT073 cycA::KmR rpoS am | G. Baisa |
| WAM4505 | CFT073, original patient isolate | H. Mobley |
| WAM4520 | WAM4505 derivative, CFT073 ΔlacZYA | [45] |
| WAM4567 | WAM4505 derivative, CFT073 ΔdsdA | This study |
| WAM5311 | WAM4505 derivative, CFT073 ΔdsdA sdaA::CmR sdaB::KmR | This study |
| WAM5314 | WAM4505 derivative, CFT073 ΔcycA ΔdsdX | This study |
| ΦEB49 | Generalized transducing phage | [48] |
| pKD46 | Expresses λ RED recombination functions, ApR | [47] |
| pKD3 | Template for λ RED-mediated replacement, CmR | [47] |
| pKD4 | Template for λ RED-mediated replacement, KmR | [47] |
| pCP20 | Encodes FLP recombinase, ApR | [47] |
| cycA F Check | 5’-GAC TAT CCC GCA GGA ACT GG-3’ | Our laboratory |
| cycA R Check | 5’-CGG CAT TAA TGA ACT GAT TGA TGA C-3’ | Our laboratory |
| dsdA λ RED F | 5’-CCT GCT GTC ATT TAT CAT CTA AGC GCA AAG AGACGT ACT TGT GTA GGC TGG AGC TGC TTC G-3’ | This study |
| dsdA λ RED R | 5’-CAC CCA GGG AAA GGA TGG CGA TGC TGC GTT GAA ACG TTA CAT ATG AAT ATC CTC CTT AG-3’ | This study |
| dsdA F Check | 5’-GGT TCC GGT GCG ATT GGC TGC-3’ | This study |
| dsdA R Check | 5’-GGA TGG CGA TGC TGC GTT G-3’ | This study |
| dsdX F Check | 5’-GAA TTG GTC TGA CAC TTC AAC GCT GC-3’ | Our laboratory |
| dsdX R Check | 5’-GCA ACC AGT TCT GAT TCA ATA ATC CCC-3’ | Our laboratory |
| rpoS sequencing F | 5’-CTG AGT GCC TAC GCC CAT AAC GAC-3’ | This study |
| rpoS sequencing R | 5’-CAA TTA CTG TGC GCT TAA AAT GAT GAT TG-3’ | This study |
| sdaA F Check | 5’-CGG GAA ACC CTA AAT CAT CGT CAG G-3’ | Our laboratory |
| sdaA R Check | 5’-GGT TGC GGA AGG GAA TCT ACC A-3’ | Our laboratory |
| sdaB F Check | 5’-GCG TGG CAA TCA CCA ATA CAG TTG A-3’ | Our laboratory |
| sdaB R Check | 5’-CGC TAG CCG CGT CTT ATC CG-3’ | Our laboratory |
Media and bacterial growth conditions
Strains were grown using the following liquid or solid media: Luria-Bertani (LB) broth or agar, MacConkey’s medium plus lactose, filter sterilized human urine, and 3-(N-morpholino)propanesulphonic acid (MOPS) minimal medium [49] supplemented with either glycerol (0.4%) or D-serine (500 μg/ml) as the sole carbon source. Urine was collected and pooled from healthy human volunteers (n = 3) with no recent history of antibiotic use. All strains were grown aerobically at 37°C and supplemented with antibiotics as applicable: kanamycin (Km, 40 μg/ml), carbenicillin (Cb, 250 μg/ml), and chloramphenicol (Cm, 20 μg/ml). In vitro growth analysis in urine was carried out essentially as described previously [28]: bacteria were grown overnight in pooled, filter sterilized human urine at 37°C with aeration, washed twice in phosphate buffered saline (PBS), and added to filter sterilized human urine to an OD600 = 0.03. Viable bacterial counts were measured by plating onto LB agar and OD600 readings were taken. For growth on minimal media, bacteria were grown overnight on MOPS glycerol plates and single colonies of each strain were swabbed onto both MOPS glycerol and MOPS D-serine minimal media and allowed to grow for 24 hours and 120 hours, respectively.
Murine Model UTI
Six- to nine-week old female CBA/J mice were used for all experiments described herein. Mice were purchased from Harlan Laboratories (competitive infections) or Jackson Laboratories (single-strain challenge). Mice were either inoculated with a single strain (single infection) or a 1:1 ratio of two strains (competitive infection) as described previously [45] and were sacrificed at 48 hours post infection (hpi). Bladders were homogenized in PBS + 0.0025% Triton X-100 and 10-fold serial dilutions in PBS were plated onto MacConkey’s medium plus lactose. In competitive infections, either CFT073 ΔlacZYA rpoS am or CFT073 ΔlacZYA was used to facilitate enumeration of wild type/mutant ratios, where applicable. CFT073 ΔlacZYA mutants colonize the murine urinary tract indistinguishly from the parental strain [50].
Motility of dsdA mutant strains
Overnight cultures of dsdA mutants and their parent strains were plated onto Adler motility medium as described previously [28, 51] with two modifications. First, overnight cultures were washed two times in PBS and OD600 was normalized to 1.0 prior to plate inoculation. Second, the diameter of migration from five independent spots was measured after 20 hours of incubation at room temperature. The difference in motility among strains was assessed by one-way analysis of variance (ANOVA) with Tukey’s Multiple Comparison Test to assess statistical significance.
Statistical analyses
All statistical analyses were carried out using Prism 4.0 (GraphPad, Inc.). Statistical significance was determined by Mann-Whitney U test or Wilcoxon signed-rank test for log distributed data where applicable and was determined by unpaired T test or one-way ANOVA for normally distributed data where applicable. P values ≤0.05 represent statistically significant differences between data sets.
Supporting Information
dsdA + strains (CFT073, CFT073 gyrA S83L, and CFT073 rpoS am) and dsdA - strains (CFT073 ΔdsdA, CFT073 dsdA Δ445bp gyrA S83L, and CFT073 ΔdsdA rpoS am) were inoculated onto Adler’s motility medium and zones of migration were measured as described in Materials and Methods. Bars represent mean diameter of migration and error bars represent standard error of the mean (±SEM). The differences in motility between strains were assessed by one-way analysis of variance (ANOVA) with Tukey’s Multiple Comparison Test to assess statistical significance.
(TIF)
dsdA + strains (CFT073, CFT073 gyrA S83L, and CFT073 rpoS am) and dsdA - strains (CFT073 ΔdsdA, CFT073 dsdA Δ445bp gyrA S83L, and CFT073 ΔdsdA rpoS am) were swabbed onto (A) MOPS glycerol minimal medium and (B) MOPS D-serine minimal medium and were incubated aerobically at 37°C for 24 and 120 hours, respectively.
(TIF)
CFT073 ΔlacZYA was co-inocualted with either CFT073 ΔdsdA sdaA::CmR sdaB::KmR or CFT073 ΔcycA ΔdsdX at a 1:1 ratio into CBA/J mice (n = 10 each). Mice were sacrificed at 48hpi. Bacteria from (A) bladder and (B) kidney homogenates were enumerated on MacConkey’s medium plus lactose. Lines are drawn at the geometric mean relative competitive index (RCI). Statistical significance was assessed by a Wilcoxon signed-rank test relative to a hypothetical RCI of 1. One of the CFT073 ΔdsdA sdaA::CmR sdaB::KmR/ CFT073 ΔlacZYA co-infected animals had no detectable bacteria in her bladder and one of the CFT073 ΔcycA ΔdsdX / CFT073 ΔlacZYA co-infected animals had no detectable bacteria in her kidneys.
(TIF)
Acknowledgments
We thank Eric Battaglioli and Erica Raterman for constructive comments on experiments and Justin Lemke for constructive comments on this manuscript.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the National Institutes of Health (http://www.nih.gov/) Grant R01DK063250, awarded to RAW. The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Foxman B. Recurring urinary tract infection: incidence and risk factors. American Journal of Public Health. 1990;80:331–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. O'Hanley P. Prospects for urinary tract infection vaccines In: Mobley HL, Warren JW, editors. Urinary Tract Infections: molecular pathogenesis and clinical management. Washington DC: ASM Press; 1996. [Google Scholar]
- 3. Kucheria R, Dasgupta P, Sacks SH, Khan MS, Sheerin NS. Urinary tract infections: new insights into a common problem. Postgraduate medical journal. 2005;81(952):83–6. Epub 2005/02/11. 10.1136/pgmj.2004.023036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Foxman B. The epidemiology of urinary tract infection. Nature Reviews Urology. 2010;7:653–60. 10.1038/nrurol.2010.190 [DOI] [PubMed] [Google Scholar]
- 5. Sivick KE, Mobley HL. Waging war against uropathogenic Escherichia coli: winning back the urinary tract. Infection and Immunity. 2010;78(2):568–85. Epub 2009/11/18. 10.1128/IAI.01000-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Huang Y, Nishikawa T, Satoh K, Iwata T, Fukushima T, Santa T, et al. Urinary Excretion of D-Serine in Human: Comparison of Different Ages and Species. Biological and Pharmaceutical Bulletin. 1998;21(2):156–62. [DOI] [PubMed] [Google Scholar]
- 7. Patzold R, Schieber A, Bruckner H. Gas chromatographic quantification of free D-amino acids in higher vertebrates. Biomedical chromatography. 2005;19(6):466–73. Epub 2005/07/23. 10.1002/bmc.515 . [DOI] [PubMed] [Google Scholar]
- 8. Cosloy SD, McFall E. Metabolism of D-serine in Escherichia coli K-12: mechanism of growth inhibition. Journal of Bacteriology. 1973;114(2):685–94. Epub 1973/05/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Kumashiro S, Hashimoto A, Nishikawa T. Free D-serine in post-mortem brains and spinal cords of individuals with and without neuropsychiatric diseases. Brain Research. 1995;681(1–2):117–25. Epub 1995/05/29. . [DOI] [PubMed] [Google Scholar]
- 10. Danysz W, Parsons CG. Glycine and N-methyl-D-aspartate receptors: physiological significance and possible therapeutic applications. Pharmacological Reviews. 1998;50(4):597–664. Epub 1998/12/22. . [PubMed] [Google Scholar]
- 11. Baranano DE, Ferris CD, Snyder SH. Atypical neural messengers. Trends in Neurosciences. 2001;24(2):99–106. Epub 2001/02/13. . [DOI] [PubMed] [Google Scholar]
- 12. Moritz RL, Welch RA. The Escherichia coli argW-dsdCXA genetic island is highly variable, and E. coli K1 strains commonly possess two copies of dsdCXA . Journal of Clinical Microbiology. 2006;44(11):4038–48. Epub 2006/11/08. 10.1128/JCM.01172-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Bloom FR, McFall E. Isolation and characterization of D-serine deaminase constitutive mutants by utilization of D-serine as sole carbon or nitrogen source. Journal of Bacteriology. 1975;121(3):1078–84. Epub 1975/03/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Henikoff S, Haughn GW, Calvo JM, Wallace JC. A large family of bacterial activator proteins. Proceedings of the National Academy of Sciences of the United States of America. 1988;85(18):6602–6. Epub 1988/09/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Bloom FR, McFall E, Young MC, Carothers AM. Positive control in the D-serine deaminase system of Escherichia coli K-12. Journal of Bacteriology. 1975;121(3):1092–101. Epub 1975/03/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Heincz MC, Kelker NE, McFall E. Positive control of D-serine deaminase synthesis in vitro. Proceedings of the National Academy of Sciences of the United States of America. 1978;75(4):1695–9. Epub 1978/04/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Heincz MC, McFall E. Role of the dsdC activator in regulation of D-serine deaminase synthesis. Journal of Bacteriology. 1978;136(1):96–103. Epub 1978/10/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. McFall E, Heincz MC. Identification and control of synthesis of the DsdC activator protein. Journal of Bacteriology. 1983;153(2):872–7. Epub 1983/02/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Anfora AT, Welch RA. DsdX is the second D-serine transporter in uropathogenic Escherichia coli clinical isolate CFT073. Journal of Bacteriology. 2006;188(18):6622–8. Epub 2006/09/06. 10.1128/JB.00634-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Peekhaus N, Tong S, Reizer J, Saier MH Jr, Murray E, Conway T. Characterization of a novel transporter family that includes multiple Escherichia coli gluconate transporters and their homologues. FEMS Microbiology Letters. 1997;147(2):233–8. Epub 1997/02/15. . [DOI] [PubMed] [Google Scholar]
- 21. Cosloy SD. D-serine transport system in Escherichia coli K-12. Journal of Bacteriology. 1973;114(2):679–84. Epub 1973/05/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Wargel RJ, Shadur CA, Neuhaus FC. Mechanism of D-cycloserine action: transport systems for D-alanine, D-cycloserine, L-alanine, and glycine. Journal of Bacteriology. 1970;103(3):778–88. Epub 1970/09/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Dowhan W Jr, Snell EE. D-serine dehydratase from Escherichia coli. 3. Resolution of pyridoxal 5'-phosphate and coenzyme specificity. The Journal of Biological Chemistry. 1970;245(18):4629–35. Epub 1970/09/25. . [PubMed] [Google Scholar]
- 24. Dupourque D, Newton WA, Snell EE. Purification and properties of D-serine dehydrase from Escherichia coli . The Journal of Biological Chemistry. 1966;241(5):1233–8. Epub 1966/03/10. . [PubMed] [Google Scholar]
- 25. Federiuk CS, Bayer R, Shafer JA. Characterization of the catalytic pathway for D-serine dehydratase. Evidence for variation of the rate-determining step with substrate structure. The Journal of Biological Chemistry. 1983;258(9):5379–85. Epub 1983/05/10. . [PubMed] [Google Scholar]
- 26. Federiuk CS, Shafer JA. A reaction pathway for transimination of the pyridoxal 5'-phosphate in D-serine dehydratase by amino acids. The Journal of Biological Chemistry. 1983;258(9):5372–8. Epub 1983/05/10. . [PubMed] [Google Scholar]
- 27. Labow R, Robinson WG. Crystalline D-serine dehydrase. The Journal of Biological Chemistry. 1966;241(5):1239–43. Epub 1966/03/10. . [PubMed] [Google Scholar]
- 28. Roesch PL, Redford P, Batchelet S, Moritz RL, Pellett S, Haugen BJ, et al. Uropathogenic Escherichia coli use d-serine deaminase to modulate infection of the murine urinary tract. Molecular Microbiology. 2003;49(1):55–67. 10.1046/j.1365-2958.2003.03543.x [DOI] [PubMed] [Google Scholar]
- 29. Lane MC, Alteri CJ, Smith SN, Mobley HL. Expression of flagella is coincident with uropathogenic Escherichia coli ascension to the upper urinary tract. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(42):16669–74. Epub 2007/10/11. 10.1073/pnas.0607898104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Haugen BJ, Pellett S, Redford P, Hamilton HL, Roesch PL, Welch RA. In vivo gene expression analysis identifies genes required for enhanced colonization of the mouse urinary tract by uropathogenic Escherichia coli strain CFT073 dsdA . Infection and Immunity. 2007;75(1):278–89. Epub 2006/11/01. 10.1128/IAI.01319-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Anfora AT, Haugen BJ, Roesch P, Redford P, Welch RA. Roles of serine accumulation and catabolism in the colonization of the murine urinary tract by Escherichia coli CFT073. Infection and Immunity. 2007;75(11):5298–304. Epub 2007/09/06. 10.1128/IAI.00652-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Su H, Newman EB. A novel L-serine deaminase activity in Escherichia coli K-12. Journal of Bacteriology. 1991;173(8):2473–80. Epub 1991/04/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Korte-Berwanger M, Sakinc T, Kline K, Nielsen HV, Hultgren S, Gatermann SG. Significance of the D-serine-deaminase and D-serine metabolism of Staphylococcus saprophyticus for virulence. Infection and Immunity. 2013;81(12):4525–33. Epub 2013/10/02. 10.1128/iai.00599-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Sakinc T, Woznowski M, Ebsen M, Gatermann SG. The surface-associated protein of Staphylococcus saprophyticus is a lipase. Infection and Immunity. 2005;73(10):6419–28. Epub 2005/09/24. 10.1128/iai.73.10.6419-6428.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Gruber TM, Gross CA. Multiple Sigma Subunits and the Partitioning of Bacterial Transcription Space. Annual Review of Microbiology. 2003;57:441–66. [DOI] [PubMed] [Google Scholar]
- 36. Weber H, Polen T, Heuveling J, Wendisch VF, Hengge R. Genome-wide analysis of the general stress response network in Escherichia coli: sigmaS-dependent genes, promoters, and sigma factor selectivity. Journal of Bacteriology. 2005;187(5):1591–603. Epub 2005/02/18. 10.1128/JB.187.5.1591-1603.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Snyder E, Gordon DM, Stoebel DM. Escherichia coli Lacking RpoS Are Rare in Natural Populations of Non-Pathogens. G3 (Bethesda, Md). 2012;2(11):1341–4. Epub 2012/11/23. 10.1534/g3.112.003855 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Spira B, de Almeida Toledo R, Maharjan RP, Ferenci T. The uncertain consequences of transferring bacterial strains between laboratories—rpoS instability as an example. BMC Microbiology. 2011;11:248 Epub 2011/11/10. 10.1186/1471-2180-11-248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Zambrano MM, Siegele DA, Almiron M, Tormo A, Kolter R. Microbial competition: Escherichia coli mutants that take over stationary phase cultures. Science. 1993;259(5102):1757–60. Epub 1993/03/19. . [DOI] [PubMed] [Google Scholar]
- 40. Welch RA, Burland V, Plunkett G 3rd, Redford P, Roesch P, Rasko D, et al. Extensive mosaic structure revealed by the complete genome sequence of uropathogenic Escherichia coli . Proceedings of the National Academy of Sciences of the United States of America. 2002;99(26):17020–4. Epub 2002/12/10. 10.1073/pnas.252529799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Mobley HL, Green DM, Trifillis AL, Johnson DE, Chippendale GR, Lockatell CV, et al. Pyelonephritiogenic Escherichia coli and Killing of Cultured Human Renal Proximal Tubular Epithelial Cells: Role of Hemolysin in Some Strains. Infection and Immunity. 1990;58(5):1281–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Mobley HL, Jarvis KG, Elwood JP, Whittle DI, Lockatell CV, Russel RG, et al. Isogenic P-fimbrial deletion mutants of pyelonephritogenic Escherichia coli: the role of alpha-gal(1–4)-beta-gal binding in virulence of a wild-type strain. Molecular Microbiology. 1993;10(1):143–55. [DOI] [PubMed] [Google Scholar]
- 43. Gunther NWt, Snyder JA, Lockatell V, Blomfield I, Johnson DE, Mobley HL. Assessment of virulence of uropathogenic Escherichia coli type 1 fimbrial mutants in which the invertible element is phase-locked on or off. Infection and Immunity. 2002;70(7):3344–54. Epub 2002/06/18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Putnam DF. Composition and Concentrative Properties of Human Urine. NASA Contractor Report. 1971.
- 45. Hryckowian AJ, Welch RA. RpoS contributes to phagocyte oxidase-mediated stress resistance during urinary tract infection by Escherichia coli CFT073. mBio. 2013;4(1):e00023–13. Epub 2013/02/14. 10.1128/mBio.00023-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Hryckowian AJ, Battesti A, Lemke JJ, Meyer ZC, Welch RA. IraL is an RssB anti-adaptor that stabilizes RpoS during logarithmic phase growth in Escherichia coli and Shigella . mBio. 2014;5(3):e01043–14. Epub 2014/05/29. 10.1128/mBio.01043-14 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(12):6640–5. Epub 2000/06/01. 10.1073/pnas.120163297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Battaglioli EJ, Baisa GA, Weeks AE, Schroll RA, Hryckowian AJ, Welch RA. Isolation of generalized transducing bacteriophages for uropathogenic strains of Escherichia coli . Applied and Environmental Microbiology. 2011;77(18):6630–5. Epub 2011/07/26. 10.1128/AEM.05307-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Neidhardt FC, Bloch PL, Smith DF. Culture medium for enterobacteria. Journal of Bacteriology. 1974;119(3):736–47. Epub 1974/09/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Redford P, Welch RA. Role of sigma E-regulated genes in Escherichia coli uropathogenesis. Infection and Immunity. 2006;74(7):4030–8. Epub 2006/06/23. 10.1128/IAI.01984-05 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Adler J. Chemotaxis in Bacteria. Science. 1966;153(3737):708–16. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
dsdA + strains (CFT073, CFT073 gyrA S83L, and CFT073 rpoS am) and dsdA - strains (CFT073 ΔdsdA, CFT073 dsdA Δ445bp gyrA S83L, and CFT073 ΔdsdA rpoS am) were inoculated onto Adler’s motility medium and zones of migration were measured as described in Materials and Methods. Bars represent mean diameter of migration and error bars represent standard error of the mean (±SEM). The differences in motility between strains were assessed by one-way analysis of variance (ANOVA) with Tukey’s Multiple Comparison Test to assess statistical significance.
(TIF)
dsdA + strains (CFT073, CFT073 gyrA S83L, and CFT073 rpoS am) and dsdA - strains (CFT073 ΔdsdA, CFT073 dsdA Δ445bp gyrA S83L, and CFT073 ΔdsdA rpoS am) were swabbed onto (A) MOPS glycerol minimal medium and (B) MOPS D-serine minimal medium and were incubated aerobically at 37°C for 24 and 120 hours, respectively.
(TIF)
CFT073 ΔlacZYA was co-inocualted with either CFT073 ΔdsdA sdaA::CmR sdaB::KmR or CFT073 ΔcycA ΔdsdX at a 1:1 ratio into CBA/J mice (n = 10 each). Mice were sacrificed at 48hpi. Bacteria from (A) bladder and (B) kidney homogenates were enumerated on MacConkey’s medium plus lactose. Lines are drawn at the geometric mean relative competitive index (RCI). Statistical significance was assessed by a Wilcoxon signed-rank test relative to a hypothetical RCI of 1. One of the CFT073 ΔdsdA sdaA::CmR sdaB::KmR/ CFT073 ΔlacZYA co-infected animals had no detectable bacteria in her bladder and one of the CFT073 ΔcycA ΔdsdX / CFT073 ΔlacZYA co-infected animals had no detectable bacteria in her kidneys.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.