

Abstract
The sperm maturation and storage functions of the epididymis are important determinants of ejaculate quality, and perhaps provide an avenue to male contraception. In the last 50 years, the creation of epididymal fertility profiles in laboratory animals was followed by recognition of new sperm maturation-related parameters (organization of the acrosome, of the sperm plasmalemma, and –S–S– -based structural change) which made it possible to confirm that a similar pattern of sperm maturation obtains in man. The novel sperm storage function of the cauda epididymidis in therian mammals is regulated by androgen, usually in conjunction with the low temperature of the scrotum. The temperature-dependence of the scrotal cauda is reflected in the secretory and ion transport functions of the epithelium, in its duct dimensions and so in sperm storage capacity. Moreover, a variety of indirect evidence suggests that an elevated temperature of the cauda created by clothing may be compromising its function in man. The pattern of change in the sperm plasmalemma involving sterols, and also glycosylphosphatidylinositol-linked macromolecules as spermatozoa enter the cauda region, may underlie the need for their capacitation subsequently in the female tract. Further, in a variety of taxa the anatomy of the scrotum, together with the U-shaped configuration of the epididymis/vas deferens, suggests that the cauda's storage function may also underlie the evolution of the scrotum. Finally, despite the still relative paucity of comparative evidence, we can consider now why the epididymis has come to be organized as it is.
At the Worcester Foundation for Experimental Biology, Shrewsbury, Massachusetts, USA in 1959, I joined the laboratory of MC Chang, which was focused on in vitro fertilization, cross-species fertilization, sperm capacitation, sperm transport in the female, the Fallopian tube, and implantation of the embryo. At that time, Chang's group had no specific interest in the epididymis, which was something of a scientific backwater compared to the focus then on the testis. I first became involved with the epididymis in asking whether, as spermatozoa are gaining the ability to fertilize there, they require the same period of capacitation as those from the cauda region – a point answered later for the pig, whose upper-corpus spermatozoa are capacitated more rapidly than those from the cauda.1 However, that question led me to others having to do with sperm maturation and sperm storage in the epididymis.
SPERM MATURATION IN THE EPIDIDYMIS
Development of fertilizing ability
In the early 1960s, specific information as to the fertilizing ability of spermatozoa in the epididymis was limited to the guinea pig,2 and rat,3 but there was none for the rabbit - my sperm capacitation model. I therefore created a fertility profile of the rabbit epididymis by tubal insemination of sperm aliquots from its successive segments via a flank incision. On the basis of fertilization rates and sperm numbers on the eggs, this revealed an exponential development of fertilizing ability beginning in the mid-corpus region, completed by the time spermatozoa reach the proximal cauda.4 Orgebin-Crist5 drew similar conclusions after insemination via the uterine route. However, the profile that obtains in the rabbit is not universal. It is skewed further towards the cauda epididymidis in the hamster,6 whereas some pig spermatozoa first gain the ability to fertilize in the upper corpus region.1
Some years later, by the use of X-irradiation as a marker,7 the relative competence of the spermatozoa in different regions of the rabbit cauda was assessed by mixed vaginal insemination of two equal populations. Spermatozoa from the upper cauda fertilized 64% of eggs in competition with those from the lower cauda and 82% in competition with equal numbers of vas deferens’ spermatozoa.8 Yet the superior population in the upper cauda is not immediately available for ejaculation. In accordance with the observation that stimulation of the (sympathetic) vas deferential nerve elicits tubular contractions only of the vas and main body of the cauda,9 repeated mating in the rat almost emptied these two regions - but with no change in sperm numbers in the upper cauda.8 As a further point, after vaginal insemination rabbit cauda spermatozoa competed equally with ejaculated spermatozoa in terms of their transport in the female and of fertilization success.10
As a sequitur to establishment of a fertility profile, the role of specific epididymal regions was examined by ligation of the epididymis (Figure 1a). Although this approach proved less successful in the hamster,6 rabbit spermatozoa became fertile throughout the epididymis up to the distal limb of the caput flexure when withheld for 8–12 days above ligatures in the corpus region.11,12 On the other hand, while many spermatozoa released after 12+ days after ligation from distended tubules of the caput flexure (Figure 1b) displayed highly progressive motility in culture for >24 h, these did not adhere to or penetrate the tubal eggs. This failure indicated that the head of the rabbit epididymis has an essential role in the development of the sperm's ability to fertilize oocytes. Secondly, the fact that highly motile spermatozoa from the proximal caput of the ligated epididymis could not do so pointed to the involvement, beside motility, of other facets of the sperm cell in this maturation. Subsequent observations demonstrated that development of the sperm's fertilizing ability in the epididymis also involves change in, (a) the sperm plasmalemma, (b) the acrosome, and (c) the structure of organelles in the sperm head and tail.
Figure 1.
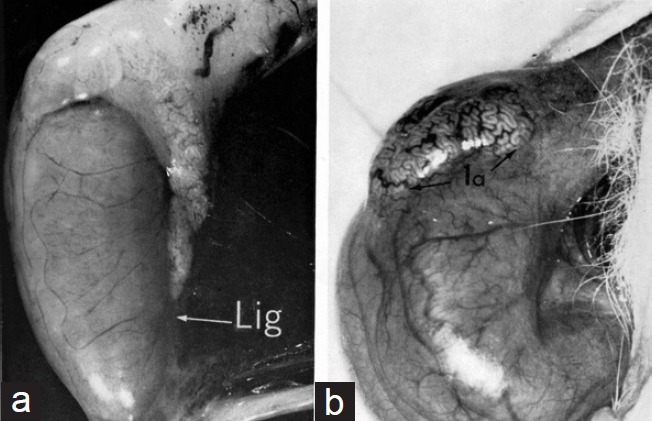
(a) Rabbit epididymis ligated 12 days previously in the lower part of the corpus epididymidis. Note the distended tubules proximal to the point of ligation. (b) Distended tubules in the initial segment of the caput epididymidis 12 or more days after ligation of the low corpus region. Spermatozoa released from these tubules were often highly motile but could not fertilize oocytes; nor did they display the tendency for head-head agglutination seen typically in mature spermatozoa.
The sperm plasmalemma
In the course of epididymal ligation experiments, it became apparent that highly motile rabbit spermatozoa from the proximal caput did not undergo the head-to-head auto-agglutination typical of mature spermatozoa of most eutherian mammals when suspended in a serum-containing medium (with man a notable exception in this regard). That hint of change in the sperm plasmalemma was confirmed by the differential electrophoretic behavior of viable spermatozoa cooled to immotility at 4°C13 (Figure 2). Most caput spermatozoa displayed a relatively slow electrophoretic mobility and required about 4 min to adopt a head-to-anode orientation. By contrast, cauda spermatozoa not only displayed a greater electrophoretic mobility but assumed a tail-to-anode configuration within ca. 1 min – clearly indicating redistribution and an increase of net negative charge over the sperm surface as a correlate of epididymal maturation.13 That sperm surface change was confirmed using cationized ferric colloid as a marker in other species, including a macaque and man, among others.14 Subsequently, the use of fluorescinated lectins and other techniques established that epididymal maturation of the sperm plasmalemma involves an exchange of both glycosylated proteins, in some cases transferred by epididymosomes,15 and of sterols.16 However, sperm membrane cholesterol/phospholipid ratios illustrate the important point that the molecular details of this aspect of maturation may vary widely according to species.17
Figure 2.
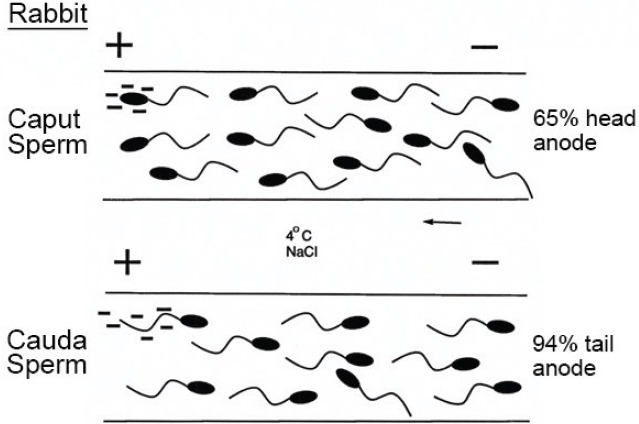
Diagram of the electrophoretic behavior of rabbit epididymal sperm populations from the caput and cauda regions, respectively. Not only do cauda spermatozoa exhibit a superior electrophoretic mobility, but the great majority assumes a tail-to-anode orientation in about 1 min, whereas caput spermatozoa slowly adopt a head-to-anode orientation in about 4 minutes.
Early investigations of epididymal change in the sperm surface also brought two misconceptions in this regard. Relying on cationized ferric colloid and fluorescinated lectins as surface markers, I first concluded that spermatozoa of monotremes, birds (excepting passerines) and reptiles do not undergo surface change during Wolffian duct passage.18 However, it was later shown using immunocytochemical methods that Wolffian duct proteins bind to maturing spermatozoa in a lizard,19 the chicken20 and a snake.21 In all, the current evidence suggests that sperm surface change in such subtherian vertebrates does not involve glycosylated elements, whereas these seem important for therian spermatozoa.22 Rather than fertilizing ability per se, posttesticular sperm surface changes in reptiles and birds may prove to relate primarily to the challenge of spermatozoa survival in the female tract.
A second misinterpretation was viewing epididymal change in the sperm plasmalemma solely in terms of fertilizing ability, the current picture suggesting that elements of this relate very much also to sperm storage in the cauda. For example, the acquisition of HIS protein23 and CD5224 as rat spermatozoa transition into the upper cauda was considered to be ‘maturation-related’. Yet some rat spermatozoa develop the ability to fertilize in the lower corpus region before any such surface modification. It seems most likely that the late sperm surface acquisition of HIS protein and CD52 relate rather to the longevity of sperm survival in the cauda. That the receptor for HIS protein evidently first appears on spermatozoa in the upper regions of the epididymis,23 is a further indication that some aspects of sperm surface change during epididymal transit relate not to fertilizing ability per se but rather to prolongation of the life of spermatozoa in the cauda region.
The acrosome
The evidence points to the therian acrosome as undergoing epididymal maturation-related changes at two levels – in its morphology, and in the organization of its matrix. Initially, Rose-Bengal stained spermatozoa gave me an impression that the acrosomes of rabbit spermatozoa from the proximal caput were somewhat “fatter” than those of cauda spermatozoa. Measurement of sperm head dimensions using a camera lucida confirmed this,25 with electron microscopy showing that an elongated margin of the acrosome in immature spermatozoa then retracts to form the asymmetrical bulbous border seen in mature rabbit spermatozoa.26 Coincidentally, Fawcett et al. illustrated a more dramatic change in the morphology of guinea and chinchilla acrosomes,27,28 and our later studies revealed varying degrees of this change in a macaque monkey, hyrax, white-tailed rat (Mystromys albicaudatus), and the prosimian bush baby (Galago senegalensis), with the most striking examples reported in the marsupial brush-tailed possum29,30 and tammar wallaby.30 On the other hand, there is no visible postspermiation change in the acrosome of the laboratory rat, the tree shrew (Tupaia glis), the lemur (Lemur fulvus)31 nor in man,32 and so this facet of maturation is not universal. A further and more subtle aspect - a reorganization of components within the acrosomal matrix during epididymal passage - has been described in such species as the hamster, guinea pig and boar.33,34,35
Sperm structure
In attempting to understand what determines its expansion within the egg, Harold Calvin and I observed that the sperm nucleus remains intact if exposed in vitro to sodium dodecyl sulfate (SDS) or to dithiothreitol (DTT) alone, but soon disassociates in SDS + DTT at pH 9.0. By building on this, exposure of spermatozoa from successive regions of the epididymis to SDS + DTT then showed that the nucleus, perinuclear material and also various tail structures become stabilized by –S–S– -crosslinks during epididymal passage.36 However, while an –S–S– -based stability of sperm tail elements is seen in some nonmammalian species,37 the keratinoid quality of the eutherian sperm head is highly unusual.38 It has often been suggested that this quality serves to protect the sperm genome, but the eutherian sperm head is not subjected to unusual “stresses” during epididymal maturation or in the female tract. Rather, the example of a buckled rabbit sperm head within the zona pellucida (ZP) (Figure 3) points to –S–S– -stiffening of the nucleus and perinuclear matrix as an adaptation to the challenge of penetrating an unusually robust egg coat.
Figure 3.
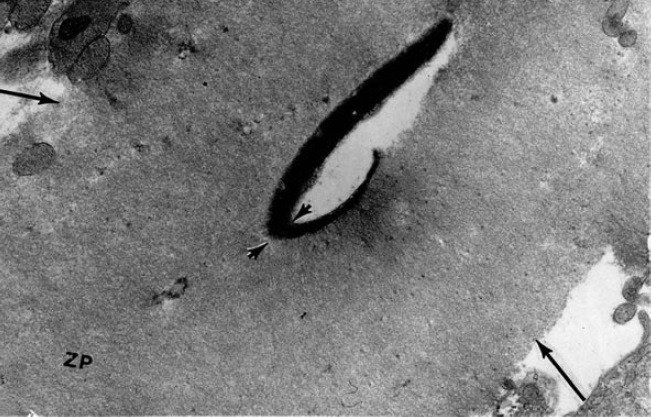
Electron micrograph of a rabbit sperm head in the act of penetrating the zona pellucida (ZP). In this example, the sperm head has buckled at the point marked by arrow heads.
SPERM MATURATION IN THE HUMAN EPIDIDYMIS
By the early 1970s, recognition of four parameters related to sperm maturation in the epididymis (motility, acrosomal modification, sperm structure, sperm plasmalemma), made it possible to probe the status of spermatozoa in the human epididymis. At that time, there was only the early report by Belonoschkin39 that human spermatozoa acquire the capacity for progressive motility there. Moreover, evidence of the success of high epididymo-vasostomy,40 and fertile spermatozoa withdrawn from a caput cyst41 cast some doubt as to whether epididymal maturation in man resembles that in the animals so far examined. In fact, the changes human spermatozoa undergo in traversing the epididymis prove to be comparable.
Of the epididymides donated by men undergoing orchiectomy for prostate cancer, only 16 (<10%) were chosen on the basis of their normal appearance. Within the epididymal population, the morphology of the sperm head was highly variable as a consequence of defects arising during spermiogenesis, but the human acrosome did not exhibit any change in its form after spermiation. On the other hand, we observed that human spermatozoa transiting the epididymis clearly undergo maturation-associated changes in the plasmalemma, in the structural character of the sperm head and tail, and in the capacity for motility.32 In regard to the sperm surface, the binding pattern of cationized ferric colloid gave evidence of maturation-associated change in the plasmalemma. Modification of the human sperm surface by epididymal secretions was later confirmed by the use of immunocytochemical techniques.42 In investigating the sperm's structural quality, –S–S– -related stabilization of the human sperm nucleus and structural tail organelles during epididymal passage were very evident on treatment of successive populations with SDS and DTT. As for their motility, spermatozoa released from the caput were immotile or displayed only a weak wide thrashing movement that produced little or no forward progression. Progressively motile spermatozoa, characterized by a stiff tail beat of limited sweep, were first seen in the population released from the lower corpus region, and then more so in cauda spermatozoa. However, unlike the picture coming from animals, many cauda spermatozoa displayed poor forward motility or none.32 This point was codified by Yeung et al.43 who observed that, in man, the percentage of motile spermatozoa in the corpus region exceeds that in the cauda population.
Although human spermatozoa thus undergo an epididymal maturation broadly comparable to that in animals, to what extent this depends on any one region of the epididymis is not yet clear, the functional evidence being derived necessarily from clinical situations. Cases involving epididymo-vasostomy showed higher pregnancy rates the lower the anastomosis along the epididymal duct.44,45,46 However, the essential role of the epididymis for sperm maturation in man was cast into some doubt by fertilization in vitro with spermatozoa recovered from the vasa efferentia of the obstructed epididymis,47 and with spermatozoa produced after anastomosis of a seminiferous tubule directly to the vas deferens.48 In the latter case, however, the vas deferens may have contributed key maturation factors since this expresses several genes in common with the epididymis.49
SPERM STORAGE IN THE CAUDA EPIDIDYMIDIS
In discussing the enigma of scrotal evolution with medical students in 1971, I asked for suggestions as to the adaptive significance of this. One student suggested that this development may relate not to the testis, but rather to sperm storage in the epididymis. That idea struck an immediate chord, in view of our focus then on the white-tailed rat (M. albicaudatus), in which the thick scrotal fur ceases abruptly at the lower poles of the testes, with a discrete bald patch overlying the cauda (Figure 4a). Although the scrotal anatomy varies among mammals, a similar pattern occurs in other groups including the bushbaby, G. senegalensis (Figure 4b), and this would seem to preferentially promote cooling of the cauda. A further example is seen in the laboratory rat, and others are not uncommon among widely different taxa,50 lending credence to the possibility that the cauda's sperm storage function was a major determinant of scrotal evolution.
Figure 4.
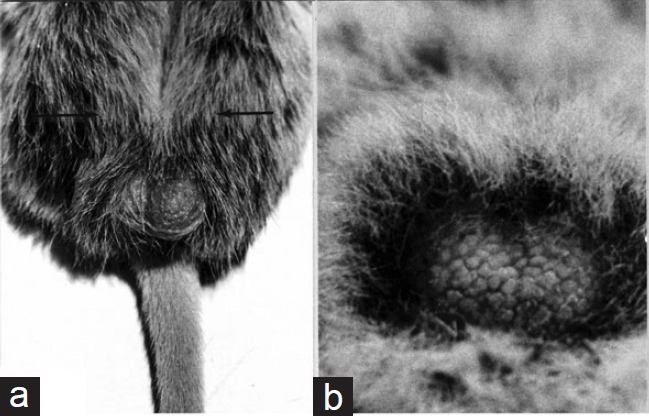
(a) Scrotum of the white-tailed rat, Mystromys albicaudatus. This view illustrates the localized fur-free segment overlying the cauda epididymidis. Arrows mark the furred region occupied by the testes. (b) Scrotum of an adult male bushbaby, Galago crassicaudatus. The scrotal surface over the testes is heavily furred, whereas the carunculated region overlying the cauda is bare.
It has long been known that body temperature suppresses sperm storage in the cauda.51 However, following the idea that temperature-dependent promotion of sperm storage was the prime mover in scrotal evolution, the effect of an elevated temperature was studied by reflecting one or both epididymides to the abdomen, leaving normally-functioning testes in the scrotum.52 Such bilaterally cryptepididymal males remained potentially fertile–laying to rest any suggestion that sperm maturation is vulnerable to deep body temperature. At the same time, this model revealed that its higher temperature brings major change in the transepithelial transport of water, Na+, K+ and Cl− in the cauda,53 and also the disappearance of several proteins typically present in its secretions54,55 – undoubtedly both major factors in the temperature-induced collapse of regulated sperm storage there. Among other significant consequences, the time of sperm transit in cryptepididymal rabbits was reduced to 2–5 days from ca. 9 to 10 days in normal males.52 In addition, hamster spermatozoa recovered from the cryptepididymal cauda were capacitated in vitro significantly sooner than those from the scrotal cauda.56
A further outcome of the imposition of body temperature on the epididymis was a reduction of the diameter and length of the duct in the cauda region57 (Figure 5), and so in its storage capacity. Consequently, over a series, cryptepididymal rats ejaculated only ca. 25% of the number of spermatozoa produced by normal males (Figure 6), despite a comparable sperm output by the testis, and sperm numbers in the caput region. Although such cryptepididymal males fathered litters,52 this picture raises an issue as to the impact of sperm numbers in a natural setting. In the case of the rat at least, as more spermatozoa were inseminated, more reached the oviduct ampulla and fertilization occurred sooner.58 This was seen in the outcome of one versus two ejaculates (Figure 7).
Figure 5.
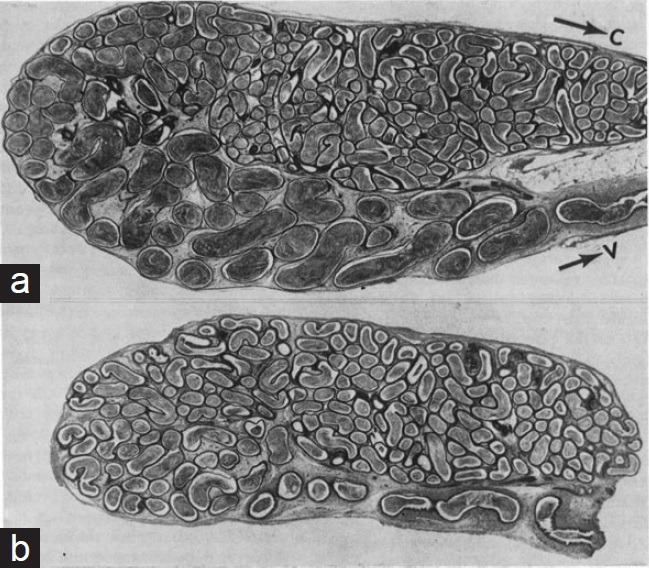
(a) Histological section of the scrotal cauda epididymidis of a rat. (b) A comparable section of the ipsilateral cauda reflected the abdomen 3 months previously. In (b) the diameter and length of the distal segment are reduced. Note that the epididymis in (b) remained in continuity with a normal scrotal testis, and had a normal sperm number in the caput. C: The lower region of the corpus epididymidis. V: the vas deferens.
Figure 6.
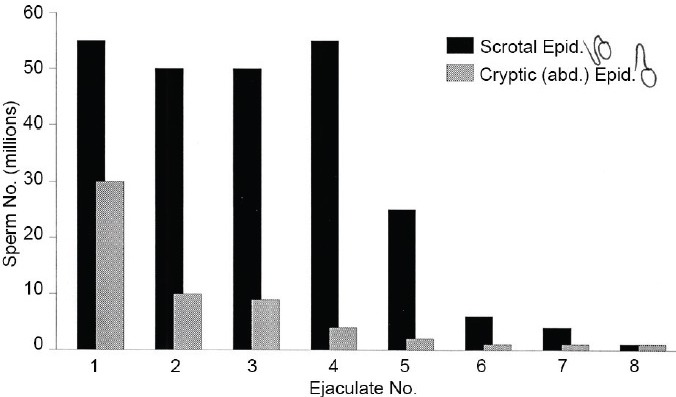
Histogram of the mean number of spermatozoa produced at ca. 15 min intervals in sequential ejaculates, by normal (n = 13) versus 3–4 weeks cryptepididymal (n = 8) rats. Notwithstanding a normal sperm production by the testes and numbers present in the caput, cryptepididymal males ejaculated only <25% of the spermatozoa produced by males with scrotal epididymides.
Figure 7.
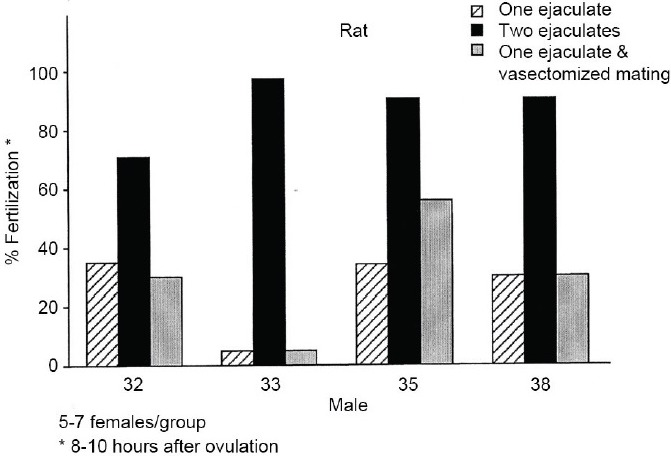
Histogram, for four males, of the percentage rat eggs fertilized by 8–10 h postovulation after either one or two ejaculations, or one ejaculation followed by mating with a vasectomized male. The sperm numbers contributed by a second ejaculate clearly favor earlier fertilization.
THE HUMAN CAUDA EPIDIDYMIDIS
Given that clothing raises the human scrotal temperature by 4°C or more depending on the situation; it is perhaps not surprising to find that much of the picture seen in cryptepididymal animals is echoed in man.59 As reflected in the ratio of cauda sperm number/daily testis sperm production, the relative storage capacity of the cauda is the lowest in man among commonly studied mammals, being akin to that of the cryptepididymal rat. In the latter [Figure 6] and in man alike [Figure 8], the cauda's minimal capacity is reflected in a modest first ejaculate and then a precipitous fall in the sperm numbers produced in subsequent ejaculates. In addition, the percentage motility in sperm populations released from the human cauda is lower than that from the corpus.43 That the human cauda does not support sperm viability is indicated as well by the outcome of abstinence. Unlike the minimal consequence of this in animals, prolonged abstinence in man resulted in a major reduction in the motility index of the first ejaculate produced thereafter.59,60 That the passage of many spermatozoa through the human epididymis is relatively brief,61,62 and that some are capacitated rapidly in vitro, adds to the evidence that in modern societies the human cauda functions in a temperature-suppressed state.
Figure 8.
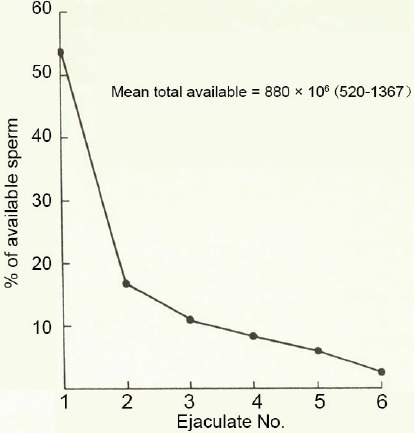
The pattern of mean sperm output in successive ejaculates produced over 24 h by 23–27 years old men (n = 10). The figure for each ejaculate is expressed as the percentage of the total produced.
COMMENT
While much has emerged about the epididymis over the last 50 years, puzzles remain at several levels. In particular, the sperm maturation and sperm storage functions of the therian epididymis raise a fundamental question as to why it is organized as it is, given the absence of these functions where fertilization is external; e.g., in the cyclostomes, teleosts and anuran amphibians studied.18 It has been difficult to develop a balanced perspective in this regard not least because research has focused primarily on therian and particularly eutherian mammals. The few observations on subtherian vertebrates indicate that where fertilization is internal, as in elasmobranchs, reptiles, birds and monotremes, a form of sperm maturation appears in the excurrent duct. However, this seems to be characterized only by development of the capacity for progressive motility (dependent on glycolysis as well as oxidative phosphorylation), and by sperm surface acquisition of protein moieties secreted by the duct.19,20,21 It is not yet clear for sub-therian vertebrates whether such surface change bears ultimately on the sperm's fertilizing ability per se,63 or whether in subtherian taxa these act only to foster the survival of spermatozoa during what is often a prolonged stint in the female tract.
At the same time, events in the epididymis appear to have become more complex in therian mammals. As judged by spermatozoa withheld in the testis by ligation of the efferent ducts, the capacity for sustained progressive motility has become epididymis-dependent. In addition, remodeling of the sperm plasmalemma involves an exchange of both sterols and glycosylated proteins, the details showing considerable variation from species to species. A change in the morphology of the acrosome seen in some species may parallel and even reflect a reorganization of its matrix. However, as seen in the rat, modifications of the matrix may also occur during epididymal passage in the absence of any coincident morphological change in the acrosome.64 It seems likely that one key to these acrosomal changes has been evolutionary change in the egg vestment that created novel demands on spermatozoa at fertilization.65 For instance, rather than immediate induction of the acrosome reaction and penetration seen in many invertebrates, fertilization in therian mammals appears to involve a period of preliminary sperm binding to the ZP that may involve not only glycosylated elements at the sperm surface but also others in the matrix exposed by the acrosome reaction. Not least, the unique –S–S– -cross-linking, which stiffens the eutherian sperm head, likely facilitates its penetration through a thick, resilient egg coat – a step that appears to have selected in turn for gamete fusion via the equatorial segment of the sperm head.65
The other epididymal function – sperm storage based on differentiation of the cauda region – was not evident in the few subtherians studied in this regard,18 notwithstanding a final bunching of spermatozoa into fasces in monotremes.66 In considering why the cauda has evolved as a regulated sperm storage site in therian mammals, several factors suggest themselves. Firstly, once they are mature, spermatozoa of laboratory mammals survive for only a few days if retained upstream of the special environment of the cauda, yet those of some snakes can remain viable for months throughout the Wolffian duct, even during testicular regression. This single comparison introduces the possibility that certain characteristics of mature therian spermatozoa make them relatively vulnerable without the support of the special environment created within the cauda. Secondly, whereas spermatozoa pass through the Wolffian duct in 24 or more hours in some nonpasserine birds,67 this requires around 10 days in eutherian mammals, regardless of huge differences in duct length, spermatozoa being wafted back and forth at each level before slowly moving on to the next. Allied to a relatively modest sperm production,68 these factors compromise the potential for rapid replacement by younger spermatozoa after ejaculation. This view seems consistent with the picture in passerine birds, which also have evolved such a sperm store within a scrotum-like protuberance.69 For in a prototypical passerine, Melopspizia melodia, modest numbers pass through a ciliated Wolffian duct, to complete what appears as a more complex maturation and then storage in the terminal glomus.18
Finally, a case can be made, (a) that the state in which sperm are stored in the cauda underlies the need for their capacitation subsequently in the female tract, and (b) that the storage function of the cauda initially drove the evolution of the scrotum.65 But what benefit does the cauda provide? As noted, this not only serves to prolong the life of mature spermatozoa, but the relative passivity of its more capacious duct allows them to accumulate, making significantly more available in an ejaculate series over a limited time period. As well as the implications of this for polygyny or for sperm competition, in the rat at least the sperm number made available by this organization of the cauda seems to favour sperm numbers reaching the oocytes and thus early fertilization58 [Figure 7]. The important role of sperm numbers is also suggested from studies in large animals. For example, in horses better pregnancy rates followed multiple ejaculations,70 and in sheep it appears that there can often be too few spermatozoa in a first ejaculate to ensure an optimal rate of conception.71,72 It is thus very possible that what appears as a suppressed state of the cauda epididymidis in man may underlie the fact that, on average, the establishment of a pregnancy requires unprotected intercourse for several months.
REFERENCES
- 1.Hunter RH, Holtz W, Henfrey PJ. Epididymal function in the boar in relation to the fertilizing ability of spermatozoa. J Reprod Fertil. 1976;46:463–6. doi: 10.1530/jrf.0.0460463. [DOI] [PubMed] [Google Scholar]
- 2.Young WC. A study of the function of the epididymis. III Functional changes undergone by spermatozoa during their passage through the epididymis and vas deferens in the guineas pig. J Exp Biol. 1931;8:151–62. [Google Scholar]
- 3.Blandau RJ, Rumery RE. Fertilizing capacity of spermatozoa recovered from various segments of the epididymis. Anat Rec. 1964;139:202. [Google Scholar]
- 4.Bedford JM. Development of the fertilizing ability of spermatozoa in the epididymis of the rabbit. J Exp Zool. 1966;163:319–30. [Google Scholar]
- 5.Orgebin-Crist MC. Sperm maturation in rabbit epididymis. Nature. 1967;216:816–8. doi: 10.1038/216816a0. [DOI] [PubMed] [Google Scholar]
- 6.Horan AH, Bedford JM. Development of the fertilizing ability of spermatozoa in the epididymis of the Syrian hamster. J Reprod Fertil. 1972;30:417–23. doi: 10.1530/jrf.0.0300417. [DOI] [PubMed] [Google Scholar]
- 7.Bedford JM, Hunter RH. The influence of X-irradiation of rabbit spermatozoa on fertilization and early cleavage. J Reprod Fertil. 1968;17:49–57. doi: 10.1530/jrf.0.0170049. [DOI] [PubMed] [Google Scholar]
- 8.Bedford JM. Sperm dynamics in the epididymis. In: Asch RH, Balmaceda JP, Johnston I, editors. Gamete Physiology. Norwell, Massachusetts: Serono Symposia USA; 1990. pp. 53–67. [Google Scholar]
- 9.Cross BA, Glover TD. The hypothalamus and seminal emission. J Endocrinol. 1958;16:385–95. doi: 10.1677/joe.0.0160385. [DOI] [PubMed] [Google Scholar]
- 10.Overstreet JW, Bedford JM. Transport, capacitation and fertilizing ability of epididymal spermatozoa. J Exp Zool. 1974;89:203–14. doi: 10.1002/jez.1401890208. [DOI] [PubMed] [Google Scholar]
- 11.Bedford JM. Effects of duct ligation on the fertilizing ability of spermatozoa from different regions of the rabbit epididymis. J Exp Zool. 1967;166:271–81. doi: 10.1002/jez.1401660210. [DOI] [PubMed] [Google Scholar]
- 12.Orgebin-Crist MC. Studies on the function of the epididymis. Biol Reprod. 1969;1(Suppl 1):155–75. doi: 10.1095/biolreprod1.supplement_1.155. [DOI] [PubMed] [Google Scholar]
- 13.Bedford JM. Changes in the electrophoretic properties of rabbit spermatozoa during passage through the epididymis. Nature. 1963;200:1178–80. doi: 10.1038/2001178a0. [DOI] [PubMed] [Google Scholar]
- 14.Bedford JM. Biology of primate spermatozoa. Contrib Primatol. 1974;3:97–139. [PubMed] [Google Scholar]
- 15.Sullivan R, Saez F. Epididymosomes, prostasomes, and liposomes: their roles in mammalian male reproductive physiology. Reproduction. 2013;146:R21–35. doi: 10.1530/REP-13-0058. [DOI] [PubMed] [Google Scholar]
- 16.Jones R. Plasma membrane structure and remodelling during sperm maturation in the epididymis. J Reprod Fertil Suppl. 1998;53:73–84. [PubMed] [Google Scholar]
- 17.Cross NL. Role of cholesterol in sperm capacitation. Biol Reprod. 1998;59:7–11. doi: 10.1095/biolreprod59.1.7. [DOI] [PubMed] [Google Scholar]
- 18.Bedford JM. Evolution of the sperm maturation and sperm storage functions of the epididymis. In: Fawcett DW, Bedford JM, editors. The Spermatozoon – Maturation, Motility, Surface Properties and Comparative Aspects. Baltimore: Urban and Schwarzenberg; 1979. pp. 7–22. [Google Scholar]
- 19.Depeiges A, Dufaure JP. Binding to spermatozoa of a major soluble protein secreted by the epididymis of the lizard, Lacerta vivipara. Gamete Res. 1983;7:401–6. [Google Scholar]
- 20.Esponda P, Bedford JM. Surface of the rooster spermatozoon changes in passing through the Wolffian duct. J Exp Zool. 1985;234:441–9. doi: 10.1002/jez.1402340311. [DOI] [PubMed] [Google Scholar]
- 21.Esponda P, Bedford JM. Post-testicular change in the reptile sperm surface with particular reference to the snake, Natrix fasciata. J Exp Zool. 1987;241:123–32. doi: 10.1002/jez.1402410115. [DOI] [PubMed] [Google Scholar]
- 22.Tulsiani DR. Glycan-modifying enzymes in luminal fluid of the mammalian epididymis: an overview of their potential role in sperm maturation. Mol Cell Endocrinol. 2006;250:58–65. doi: 10.1016/j.mce.2005.12.025. [DOI] [PubMed] [Google Scholar]
- 23.Rifkin JM, Olson GE. Characterization of maturation-dependent extrinsic proteins of the rat sperm surface. J Cell Biol. 1985;100:1582–91. doi: 10.1083/jcb.100.5.1582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Derr P, Yeung CH, Cooper TG, Kirchhoff C. Synthesis and glycosylation of CD52, the major ‘maturation-associated’ antigen of rat spermatozoa, in the cauda epididymidis. Reproduction. 2001;121:435–46. doi: 10.1530/rep.0.1210435. [DOI] [PubMed] [Google Scholar]
- 25.Bedford JM. Morphological changes in rabbit spermatozoa during passage through the epididymis. J Reprod Fertil. 1963;5:169–77. doi: 10.1530/jrf.0.0050169. [DOI] [PubMed] [Google Scholar]
- 26.Bedford JM. Changes in fine structure of the rabbit sperm head during passage through the epididymis. J Anat. 1965;99:891–906. [PMC free article] [PubMed] [Google Scholar]
- 27.Fawcett DW, Hollenberg RD. Changes in the acrosome of the guinea pig spermatozoa during passage through the epididymis. Z Zellforsch Mikrosk Anat. 1963;60:276–92. doi: 10.1007/BF00350481. [DOI] [PubMed] [Google Scholar]
- 28.Fawcett DW, Phillips DH. Observations on the release of spermatozoa and on changes in the head during passage through the epididymis. J Reprod Fertil. 1969;Suppl 6:405–18. [Google Scholar]
- 29.Temple-Smith PD, Bedford JM. The features of sperm maturation in the epididymis of a marsupial, the brushtailed possum Trichosurus vulpecula. Am J Anat. 1976;147:471–99. doi: 10.1002/aja.1001470407. [DOI] [PubMed] [Google Scholar]
- 30.Lin M, Rodger JC. Acrosome formation during sperm transit through the epididymis in two marsupials, the tammar wallaby (Macropus eugenii) and the brushtail possum (Trichosurus vulpecula) J Anat. 1999;194(Pt 2):223–32. doi: 10.1046/j.1469-7580.1999.19420223.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bedford JM. Unpublished observations [Google Scholar]
- 32.Bedford JM, Calvin H, Cooper GW. The maturation of spermatozoa in the human epididymis. J Reprod Fertil Suppl. 1973;18:199–213. [PubMed] [Google Scholar]
- 33.Olson GE, Winfrey VP, Bi M, Hardy DM, NagDas SK. Zonadhesin assembly into the hamster sperm acrosomal matrix occurs by distinct targeting strategies during spermiogenesis and maturation in the epididymis. Biol Reprod. 2004;71:1128–34. doi: 10.1095/biolreprod.104.029975. [DOI] [PubMed] [Google Scholar]
- 34.Buffone MG, Foster JA, Gerton GL. The role of the acrosomal matrix in fertilization. Int J Dev Biol. 2008;52:511–22. doi: 10.1387/ijdb.072532mb. [DOI] [PubMed] [Google Scholar]
- 35.Puigmule M, Fabrega A, Yeste M, Bonet S, Pinart E. Study of the proacrosin/acrosin system in epididymal, ejaculated and in vitro-capacitated boar spermatozoa. Reprod Fertil Dev. 2011;23:837–45. doi: 10.1071/RD10345. [DOI] [PubMed] [Google Scholar]
- 36.Calvin HI, Bedford JM. Formation of disulphide bonds in the nucleus and accessory structures of mammalian spermatozoa during maturation in the epididymis. J Reprod Fertil Suppl. 1971;13(Suppl 13):65–75. [PubMed] [Google Scholar]
- 37.Bedford JM, Calvin HI. Changes in -S-S- linked structures of the sperm tail during epididymal maturation, with comparative observations in sub-mammalian species. J Exp Zool. 1974;187:181–204. doi: 10.1002/jez.1401870202. [DOI] [PubMed] [Google Scholar]
- 38.Bedford JM, Calvin HI. The occurrence and possible functional significance of -S-S- crosslinks in sperm heads, with particular reference to eutherian mammals. J Exp Zool. 1974;188:137–55. doi: 10.1002/jez.1401880203. [DOI] [PubMed] [Google Scholar]
- 39.Belonoschkin B. Biology of spermatozoa in the human testis and epididymis. Arch Gynecol. 1942;174:357–63. [Google Scholar]
- 40.Young D. Surgical treatment of male infertility. J Reprod Fertil. 1970;23:541–2. doi: 10.1530/jrf.0.0230541. [DOI] [PubMed] [Google Scholar]
- 41.Hanley HG. Pregnancy following artificial insemination from epididymal cyst. Proc Soc Study Fertil. 1956;8:20–1. [PubMed] [Google Scholar]
- 42.Tezón JG, Ramella E, Cameo MS, Vazquez MH, Blaquier JA. Immunochemical localization of secretory antigens in the human epididymis and their association with spermatozoa. Biol Reprod. 1985;32:591–7. doi: 10.1095/biolreprod32.3.591. [DOI] [PubMed] [Google Scholar]
- 43.Yeung CH, Cooper TG, Oberpenning F, Schultze H, Nieschlag E. Changes in movement characteristics of human sperm along the length of the epididymis. Biol Reprod. 1993;49:274–80. doi: 10.1095/biolreprod49.2.274. [DOI] [PubMed] [Google Scholar]
- 44.Schoysman RJ, Bedford JM. The role of the human epididymis in sperm maturation and sperm storage as reflected in the consequences of epididymovasostomy. Fertil Steril. 1986;46:293–9. doi: 10.1016/s0015-0282(16)49528-2. [DOI] [PubMed] [Google Scholar]
- 45.Silber SJ. Role of epididymis in sperm maturation. Urology. 1989;33:47–51. doi: 10.1016/0090-4295(89)90066-6. [DOI] [PubMed] [Google Scholar]
- 46.Patrizio P, Ord T, Silber SJ, Asch RH. Correlation between epididymal length and fertilization rate in men with congenital absence of the vas deferens. Fertil Steril. 1994;61:265–8. doi: 10.1016/s0015-0282(16)56515-7. [DOI] [PubMed] [Google Scholar]
- 47.Silber SJ. Pregnancy caused by sperm from vasa efferentia. Fertil Steril. 1988;49:373–5. doi: 10.1016/s0015-0282(16)59733-7. [DOI] [PubMed] [Google Scholar]
- 48.Schoysman R. Clinical situations challenging the established concept of epididymal physiology in the human. Acta Eur Fertil. 1993;24:55–60. [PubMed] [Google Scholar]
- 49.Krull N, Ivell R, Osterhoff C, Kirchhoff C. Region-specific variation of gene expression in the human epididymis as revealed by in situ hybridization with tissue-specific cDNAs. Mol Reprod Dev. 1993;34:16–24. doi: 10.1002/mrd.1080340104. [DOI] [PubMed] [Google Scholar]
- 50.Bedford JM. Anatomical evidence for the epididymis as the prime mover in the evolution of the scrotum. Am J Anat. 1978;152:483–507. doi: 10.1002/aja.1001520404. [DOI] [PubMed] [Google Scholar]
- 51.Cummins JM, Glover TD. Artificial cryptorchidism and fertility in the rabbit. J Reprod Fertil. 1970;23:423–33. doi: 10.1530/jrf.0.0230423. [DOI] [PubMed] [Google Scholar]
- 52.Bedford JM. Influence of abdominal temperature on epididymal function in the rat and rabbit. Am J Anat. 1978;152:509–21. doi: 10.1002/aja.1001520405. [DOI] [PubMed] [Google Scholar]
- 53.Wong PY, Au CL, Bedford JM. Biology of the scrotum. II. Suppression by abdominal temperature of transepithelial ion and water transport in the cauda epididymidis. Biol Reprod. 1982;26:683–9. doi: 10.1095/biolreprod26.4.683. [DOI] [PubMed] [Google Scholar]
- 54.Esponda P, Bedford JM. The influence of body temperature and castration on the protein composition of fluid in the rat cauda epididymidis. J Reprod Fertil. 1986;78:505–14. doi: 10.1530/jrf.0.0780505. [DOI] [PubMed] [Google Scholar]
- 55.Regalado F, Esponda P, Nieto A. Temperature and androgens regulate the biosynthesis of secretory proteins from rabbit cauda epididymidis. Mol Reprod Dev. 1993;36:448–53. doi: 10.1002/mrd.1080360407. [DOI] [PubMed] [Google Scholar]
- 56.Bedford JM, Yanagimachi R. Epididymal storage at abdominal temperature reduces the time required for capacitation of hamster spermatozoa. J Reprod Fertil. 1991;91:403–10. doi: 10.1530/jrf.0.0910403. [DOI] [PubMed] [Google Scholar]
- 57.Foldesy RG, Bedford JM. Biology of the scrotum I. Temperature and androgen as determinants of the sperm storage capacity of the rat cauda epididymidis. Biol Reprod. 1982;26:673–82. doi: 10.1095/biolreprod26.4.673. [DOI] [PubMed] [Google Scholar]
- 58.Bedford JM, Kim HH. Cumulus oophorus as a sperm sequestering device, in vivo. J Exp Zool. 1993;265:321–8. doi: 10.1002/jez.1402650314. [DOI] [PubMed] [Google Scholar]
- 59.Bedford JM. The status and the state of the human epididymis. Hum Reprod. 1994;9:2187–99. doi: 10.1093/oxfordjournals.humrep.a138416. [DOI] [PubMed] [Google Scholar]
- 60.Pellestor F, Girardet A, Andreo B. Effect of long abstinence periods on human sperm quality. Int J Fertil Menopausal Stud. 1994;39:278–82. [PubMed] [Google Scholar]
- 61.Rowley MJ, Teshima F, Heller CG. Duration of transit of spermatozoa through the human male ductular system. Fertil Steril. 1970;21:390–6. [PubMed] [Google Scholar]
- 62.Johnson L, Varner DD. Effect of daily spermatozoan production but not age on transit time of spermatozoa through the human epididymis. Biol Reprod. 1988;39:812–7. doi: 10.1095/biolreprod39.4.812. [DOI] [PubMed] [Google Scholar]
- 63.Morris SA, Howarth B, Jr, Crim JW, Rodriguez de Cordoba A, Esponda P, et al. Specificity of sperm-binding Wolffian duct proteins in the rooster and their persistence on spermatozoa in the female host glands. J Exp Zool. 1987;242:189–98. doi: 10.1002/jez.1402420210. [DOI] [PubMed] [Google Scholar]
- 64.Nagdas SK, Skudlarek MD, Orgebin-Crist MC, Tulsiani DR. Biochemical alterations in the proacrosin-acrosin system during epididymal maturation of the rat spermatozoa. J Androl. 1992;13:36–43. [PubMed] [Google Scholar]
- 65.Bedford JM. Singular features of fertilization and their impact on the male reproductive system in eutherian mammals. Reproduction. 2013;147:R43–52. doi: 10.1530/REP-13-0436. [DOI] [PubMed] [Google Scholar]
- 66.Nixon B, Ecroyd HW, Dacheux JL, Jones RC. Monotremes provide a key to understanding the evolutionary significance of epididymal sperm maturation. J Androl. 2011;32:665–71. doi: 10.2164/jandrol.110.012716. [DOI] [PubMed] [Google Scholar]
- 67.Clulow J, Jones RC. Production, transport, maturation, storage and survival of spermatozoa in the male Japanese quail, Coturnix coturnix. J Reprod Fertil. 1982;64:259–66. doi: 10.1530/jrf.0.0640259. [DOI] [PubMed] [Google Scholar]
- 68.Roosen-Runge EC. London: Cambridge University Press; 1977. The Process of Spermatogenesis in Animals. [Google Scholar]
- 69.Wolfson A. Sperm storage at lower-than-body temperature outside the body cavity in some passerine birds. Science. 1954;120:68–71. doi: 10.1126/science.120.3106.68. [DOI] [PubMed] [Google Scholar]
- 70.Voss JL, Squires EL, Pickett BW, Shideler RK, Eikenberry DJ. Effect of number and frequency of inseminations on fertility of mares. J Reprod Fertil Suppl. 1982;32:53–7. [PubMed] [Google Scholar]
- 71.Mattner PE, Braden AW. Studies in flock mating of sheep. 2. Fertilization and prenatal mortality. Aust J Exp Agric Anim Husbandry. 1967;7:110–6. [Google Scholar]
- 72.Fulkerson WJ, Synnott AL, Lindsay DR. Numbers of spermatozoa required to effect a normal rate of conception in naturally mated Merino ewes. J Reprod Fertil. 1982;66:129–32. doi: 10.1530/jrf.0.0660129. [DOI] [PubMed] [Google Scholar]