

Abstract
Certain ciliates of the subclass Scuticociliatia (scuticociliates) are facultative parasites of fishes in which they cause a suite of diseases collectively termed scuticociliatosis. Hitherto, comparatively little was known about genetics and genomics of scuticociliates or the mechanism of scuticociliatosis. In this study, a laboratory culture of the facultatively pathogenic scuticociliate Pseudocohnilembus persalinus was established and its genome sequenced, giving the first genome of a marine ciliate. Genome-wide horizontal gene transfer (HGT) analysis showed P. persalinus has acquired many unique prokaryote-derived genes that potentially contribute to the virulence of this organism, including cell adhesion, hemolysis and heme utilization genes. These findings give new insights into our understanding of the pathology of scuticociliates.
Scuticociliatosis, caused by certain scuticociliates1, is one of the most important parasitological problems in marine aquaculture worldwide2. In recent years there have been many reports of fatal outbreaks of infection by several scuticociliates species including Pseudocohnilembus persalinus, Uronema marinum, and Philasterides dicentrarchi in east Asia (Korea, Japan, China), Europe (Spain, Portugal) and other regions of the world, which have led to serious economic losses because of high mortalities of many fish species, in particular olive flounder and turbot1,3,4,5,6. Most studies of scuticociliatosis have focused on species identification, histopathology and immunology with little attention paid to molecular mechanisms of pathogenicity, mainly due to the lack of basic research on topics such as the life cycle, genetics and genome. Unlike the hymenostome ciliate Ichthyophthirius multifiliis, an obligate parasite of freshwater fish with a typical parasitic life cycle and distinct polymorphism7,8, scuticociliates are generally free-living in limnetic and marine ecosystems, feeding on other microorganisms such as bacteria, microalgae, protozoa etc9. Under certain circumstances, however, these ciliates may become opportunistic histophagous parasites, actively feeding on cells and tissue residues of a host organism, living and reproducing within the host tissues without observable morphological change.
Scuticociliates belong to the Scuticociliatia, one of six subclasses of the class Oligohymenophorea, the others being Peniculia, Hymenostomatia, Astomatia, Apostomatia, and Peritrichia10. Some oligohymenophoreans are the best-known of all ciliates and are commonly used as model laboratory organisms. These include the hymenostome species Tetrahymena thermophila11,12 and I. multifiliis and the peniculian species Paramecium tetraurelia13,14. Genomes of these ciliates have been sequenced and comparative genomics analyses have provided a comprehensive understanding of the unique features of these free-living (T. thermophila and P. tetraurelia) and typical parasitic (I. multifiliis) ciliates. Thus, it is expected that the genomes of parasitic scuticociliates will provide new insights and better understanding of their mechanisms of pathogenicity.
The typical Pseudocohnilembus species, P. persalinus has been reported previously as a free-living marine ciliate15,16,17. Since Kim etc. (2004) isolated P. persalinus from diseased olive flounder in Korea, it has become recognized as an important facultative parasite causing scuticociliatosis in commercially important fishes such as rainbow trout18 and olive flounder19. Pseudocohnilembus persalinus can be cultured in the laboratory as a free-living form by feeding with bacteria, which enables sufficient DNA/RNA to be collected for genome/transcriptome sequencing. Here, we report the genome of P. persalinus, the first to be sequenced among scuticociliates and, following comparative genomic analyses with its close relatives, investigate the possibility that the acquisition of its virulence may be via horizontal gene transfer (HGT) from bacteria.
Results and Discussion
Facultative parasitism of P. persalinus - a genomic view
Like most other ciliates, P. persalinus has two types of nucleus, a macronucleus (MAC) and a micronucleus (MIC), the former controlling the physiological and biochemical functions of the cell and the latter as a germ-line reserve. The first ciliate genome to be sequenced was the MAC genome of T. thermophila12 and this was achieved by first physically separating the MAC from the MIC. However, such a method has not been established for P. persalinus. In most ciliates the MIC is either haploid or diploid whereas the MAC is polyploid. In T. thermophila, for example, the MAC has an average ploidy up to 45C11, and in I. multifiliis, the MAC has an estimated ploidy up to 12,000C20. Therefore, we anticipated that the MAC of P. persalinus also has a high ploidy level and utilized this natural enrichment of MAC in order to perform MAC genome sequencing as had previously been achieved in I. multifiliis20. By employing this simple sequencing strategy, a preliminary assembly of about 63.6 Megabase (Mb) sequences with 2403 scaffolds was constructed. The GC content distribution of the preliminary assembly showed that there are three peaks of 19%, 41% and 67% (Fig. 1, red), indicating the presence of contaminants. Compared to the second and third peaks, the first peak contained a majority of sequences (sequence length, about 87%, Fig. 1, red) with a small population of scaffolds with low GC content (scaffold number, Fig. 1, blue). Published oligohymenophorean ciliate genomes usually show a low GC content (22% in T. thermophila, 15% in I. multifiliis and 28% in P. tetraurelia, Table 1), therefore sequences represented by the first peak probably derive from P. persalinus. In order to verify this, BLASTX searches were performed for all preliminary assembled sequences against the NCBI non-redundant protein sequences database. The results support the suggestion that sequences in the first peak belong to P. persalinus, whereas sequences in the second and the third peaks probably derive from bacterial sources. In order to exclude bacterial sequences, two steps were applied: 1) based on their distribution like in I. multifiliis20, filtering all scaffolds with GC content higher than 25%; 2) filtering those scaffolds with more than 50% BLASTX hits in the remaining sequences after first step, these most likely being of prokaryote origin. In practice, almost all the filtered scaffolds were removed in the first step, with only two additional short scaffolds (1.5 and 1.6 Kb, respectively) being removed at the second step. This suggests that the GC content is a good index for discriminating ciliate from bacterial sequences. Finally, about 55.5 Mb P. persalinus genome sequences with 288 scaffolds were obtained (Table 1). The N50 of the P. persalinus genome is about 368 Kb which is comparable to the genome assemblies of T. thermophila (521 Kb) and P. tetraurelia (413 Kb) (Table 1).
Figure 1. GC content distribution of preliminary genome assembly.
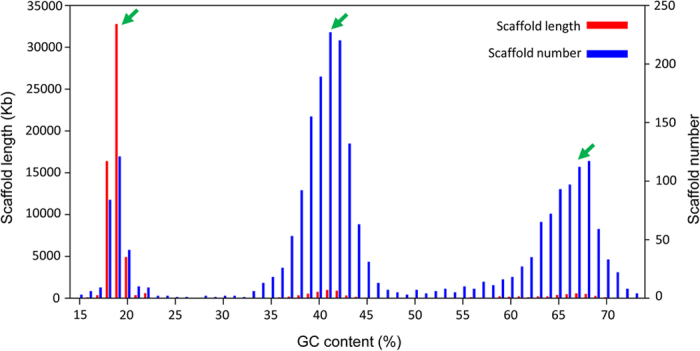
Red, scaffold length versus GC content; blue, scaffold numbers versus different GC content. Green arrows, GC peaks by scaffold length.
Table 1. Statistics of Pseudocohnilembus persalinus genome and comparison to three other sequenced oligohymenophorean ciliates.
| Species | P. persalinus | I. multifiliis | T. thermophila | P. tetraurelia |
|---|---|---|---|---|
| Genome size (Mb) | 55.5 | 47.8 | 103.0 | 72.1 |
| N50 (Kb) | 368 | 66 | 521 | 413 |
| Scaffold number | 288 | 1375 | 1148 | 697 |
| Longest scaffold (Mb) | 2.0 | 0.4 | 2.2 | 1.0 |
| Sequencing method/platform | Illumina | Sanger/454 | Sanger | Sanger |
| Average GC content | 19% | 16% | 22% | 28% |
| Gene number | 13186 | 8062 | 26460 | 39642 |
| Gene density (genes/Mb) | 238 | 169 | 256 | 548 |
P. persalinus: Pseudocohnilembus persalinus; I. multifiliis: Ichthyophthirius multifiliis; T. thermophila: Tetrahymena thermophila; P. tetraurelia: Paramecium tetraurelia.
The genome of P. persalinus provides evidence of parasitism: 1) its genome size is similar to another ciliate fish pathogen, I. multifiliis (47.8 Mb), and is significantly smaller than those of free-living ciliates such as T. thermophila (103 Mb) and P. tetraurelia (72.1 Mb) (Table 1); 2) the P. persalinus genome harbors 13,186 predicted protein coding genes, which is about two-fold less than the free-living T. thermophila (Table 1); 3) protein domain analysis showed very similar domain composition between P. persalinus and I. multifiliis, not only in the types but also the number (Figure S1); 4) Pseudocohnilembus persalinus and I. multifiliis have a similar fraction of parasitic lifestyle-relevant gene families and a relatively high percentage of proteases compared to free-living ciliates (Table S1). Compared to I. multifiliis, P. persalinus has more proteases especially in the cysteine and serine classes (Table S1) which may be key to various functions of a parasitic lifestyle including immunoevasion, excystment/encystment, ex-sheathing, and cell and tissue invasion21,22. Besides the proteases, 106 P. persalinus-specific transporters (Table S2) were found when compared to T. thermophila, and these transporters were significantly enriched in the sodium ion, zinc ion, calcium ion, ammonium and phosphate transmembrane transport systems (Figure S2). Some of these transport systems may play important roles in unique aspects of parasite biology. Calcium, for example, is an important factor in the invasion of erythrocytes9, and has been shown to help the secretion of parasite proteins during the invasion process of Toxoplasma gondii23,24. Thus, calcium transporters may be involved in the invasion process in P. persalinus. Furthermore, we compared the gene compositions of P. persalinus to the well-known parasitic ciliate I. multifiliis and the free-living T. thermophila. The results showed that very few unique orthologs (74) shared in I. multifiliis and P. persalinus (two parasites) are absent in T. thermophila (Figure S3), suggesting P. persalinus has a distinct mode of parasitism compared to I. multifiliis.
Scuticociliates infect aquatic organisms opportunistically and occurrences of scuticociliatosis seem to be influenced by environmental conditions, such as temperature and salinity, and weakening of the host due to bacterial infection25. The processes and mechanisms of the transition from free-living to parasitic lifestyle are unknown. By sequencing the genome of P. persalinus, this study revealed a number of features associated with parasitism.
Horizontal gene transfer (HGT) genes may play an important role in the virulence of P. persalinus
HGT, the transfer of genetic material between species26, was discovered half of a century ago27, but it is the current wealth of genomic sequence data that is revealing its real impact on evolution. Genes acquired by HGT can sometimes be associated with important evolutionary adaptations, including parasitism and pathogenicity28. The first evidence for the role of HGT in the acquisition of virulence determinants was between pneumococci in infected mice29. Subsequently, a number of studies have reported horizontal transfer of virulence genes (i) between prokaryotic pathogens, (ii) from prokaryotic to eukaryotic pathogens, and (iii) between eukaryotic pathogens28,30,31,32,33,34,35. In the obligate ciliate parasite I. multifiliis, which harbors an endosymbiotic bacterium, relatively few (10) HGT genes were predicted20. Using a phylogenetic approach (see Materials and Methods section), 74, 5, and 54 putative HGT genes were identified in T. thermophila, I. multifiliis and P. persalinus, respectively. The 54 putative HGT genes in P. persalinus were dispersed among 42 different assembled scaffolds (Table 2 and Supplementary Information), and had similar GC contents to the rest of the genes (Fig. 2A). PCR analysis of DNA also verified that these genes are present in the ciliate genome (Figure S4). In addition, 52 of these putative HGT genes were predicted to contain introns (Fig. 2B) which is a main feature of eukaryotic genes. Analysis of transcriptome (RNA-Seq) data reveals that 85% (44) of the putative HGT genes contain at least one intron (Fig. 2C). It is unlikely that the acquisition of introns by these HGT genes was for generating a more complicated proteome through alternative splicing because no alternative splicing was found in the RNA-Seq data. It is possible that the intron gains were the result of adaptation of the transferred gene to its new host cell machinery36. The presence of introns strongly suggest the origin of the putative HGT genes is the P. persalinus genome itself rather than bacterial contaminants, and that the HGT events occurred long ago in evolutionary time.
Table 2. 54 HGT genes in Pseudocohnilembus persalinus genome.
| Gene ID | Best hit in NCBI | E-vaule | Best hit species | Species category | Function category by annotation |
|---|---|---|---|---|---|
| PPERSA_00056740 | Ig family protein | 6.00E-31 | Pectobacterium wasabia | Proteobacteria | Cell adhesion |
| PPERSA_00029910 | Ig family protein | 8.00E-23 | Dickeya zeae | Proteobacteria | Cell adhesion |
| PPERSA_00047700 | Lysophospholipase L1 | 8.00E-34 | Amycolatopsis azurea | Actinobacteria | Hemolysis related protein |
| PPERSA_00098980 | phosphatidylinositol-specific phospholipase C1 like protein | 3.00E-06 | Flavobacterium sp. | Bacteroidetes | Hemolysis related protein |
| PPERSA_00002080 | phosphatidylinositol-specific phospholipase C1like protein | 8.00E-10 | Streptomyces thermolilacinus | Actinobacteria | Hemolysis related protein |
| PPERSA_00035610 | hemolysin III family channel protein | 1.00E-14 | Gordonia alkanivorans | Actinobacteria | Hemolysis related protein |
| PPERSA_00117390 | hemopexin repeat-containing protein | 4.00E-39 | Flavobacterium beibuense | Bacteroidetes | Heme related protein |
| PPERSA_00079580 | hemopexin repeat-containing protein | 5.00E-45 | Flavobacterium beibuense | Bacteroidetes | Heme related protein |
| PPERSA_00031570 | hemin receptor | 1.00E-06 | Mycobacterium chubuense | Actinobacteria | Heme related protein |
| PPERSA_00130810 | 2OG-Fe(II) oxygenase | 3.00E-143 | Aeromonas hydrophila | Proteobacteria | oxidoreductase |
| PPERSA_00076120 | 2OG-Fe(II) oxygenase | 8.00E-47 | Kordia algicida | Bacteroidetes | oxidoreductase |
| PPERSA_00055830 | D-amino acid dehydrogenase small subunit DadA | 4.00E-23 | Janthinobacterium sp. | Proteobacteria | oxidoreductase |
| PPERSA_00113410 | FAD-dependent pyridine nucleotide-disulfide oxidoreductase | 0.00E + 00 | Mahella australiensis | Firmicutes | oxidoreductase |
| PPERSA_00109590 | amine oxidase | 6.00E-37 | Microcystis aeruginosa | Cyanobacteria | oxidoreductase |
| PPERSA_00073400 | NAD-dependent epimerase/dehydratase | 3.00E-111 | Nitrosomonas sp. | Proteobacteria | oxidoreductase |
| PPERSA_00069080 | NAD-dependent epimerase/dehydratase | 1.00E-69 | Geobacter uraniireducens | Proteobacteria | oxidoreductase |
| PPERSA_00041150 | arsenate reductase | 2.00E-18 | Pasteurella multocida | Proteobacteria | oxidoreductase |
| PPERSA_00069770 | glutathione S-transferase | 7.00E-21 | Leeia oryzae | Proteobacteria | / |
| PPERSA_00059730 | major facilitator superfamily MFS_1 | 2.00E-34 | Clostridium carboxidivorans | Firmicutes | / |
| PPERSA_00059750 | major facilitator superfamily MFS_1 | 1.00E-14 | Clostridium drakei | Firmicutes | / |
| PPERSA_00084980 | magnesium transporter | 2.00E-10 | Fischerella muscicola | Cyanobacteria | / |
| PPERSA_00107980 | MFS-type transporter | 6.00E-45 | bacillus massiliosenegalensis | Firmicutes | / |
| PPERSA_00125230 | Beta-lactamase | 6.00E-37 | Paenibacillus sp. | Firmicutes | / |
| PPERSA_00036040 | glyoxalase/bleomycin resistance protein/dioxygenase | 3.00E-36 | Sphingopyxis sp. | Proteobacteria | / |
| PPERSA_00036460 | thioesterase | 2.00E-14 | Desulfomonile tiedjei | Proteobacteria | / |
| PPERSA_00036470 | thioesterase | 3.00E-15 | Alcanivorax hongdengensis | Proteobacteria | / |
| PPERSA_00021250 | beta-lactamase | 5.00E-50 | Cyanothece sp. | Cyanobacteria | / |
| PPERSA_00131510 | 3-oxoacyl-ACP synthase | 4.00E-13 | Chlorogloeopsis | Proteobacteria | / |
| PPERSA_00117970 | DNA polymerase III subunit epsilon | 4.00E-18 | Gammaproteobacteria bacterium SCGC AAA003-E02 | Proteobacteria | / |
| PPERSA_00073390 | 2-amino-3-ketobutyrate CoA ligase | 0.00E + 00 | Candidatus Cloacamonas | Cloacimonetes | / |
| PPERSA_00011350 | formyl transferase domain protein | 3.00E-16 | Streptomyces natalensis | Actinobacteria | / |
| PPERSA_00089970 | inosine/uridine-preferring nucleoside hydrolase | 7.00E-37 | Legionella wadsworthii | Proteobacteria | / |
| PPERSA_00125930 | bifunctional GMP synthase/glutamine amidotransferase protein | 0.00E + 00 | Lentisphaera araneosa | Chlamydiae | / |
| PPERSA_00035440 | Rhodanese domain protein | 5.00E-08 | Flavobacterium sp. | Bacteroidetes | / |
| PPERSA_00043810 | rhodanese-related sulfurtransferase | 1.00E-117 | Endozoicomonas elysicola | Proteobacteria | / |
| PPERSA_00057430 | SH3 protein, type 3 | 7.00E-58 | Gemmata obscuriglobus | Planctomycetes | / |
| PPERSA_00050710 | hypothetical protein | 6.00E-12 | Microscilla marina | Bacteroidetes | / |
| PPERSA_00036150 | putative phosphatase | 1.00E-12 | Photobacterium halotolerans | Proteobacteria | / |
| PPERSA_00010290 | PF08002 family protein | 3.00E-13 | Bacteroidetes bacterium | Bacteroidetes | / |
| PPERSA_00050920 | photopyrone synthase | 5.00E-14 | Photorhabdus luminescens | Proteobacteria | / |
| PPERSA_00042620 | primase | 5.00E-07 | Methanosarcina acetivorans | Methanomicrobia | / |
| PPERSA_00086310 | 3-oxoacyl-ACP synthase | 7.00E-11 | Cupriavidus sp. | Proteobacteria | / |
| PPERSA_00086300 | 3-oxoacyl-ACP synthase | 2.00E-10 | Pseudanabaena sp. | Cyanobacteria | / |
| PPERSA_00045220 | 3-oxoacyl-ACP synthase | 2.00E-14 | Granulicella mallensis | Fibrobacteres | / |
| PPERSA_00054440 | acid phosphatase | 1.00E-06 | Flavobacterium sp. | Bacteroidetes | / |
| PPERSA_00098990 | acid phosphatase | 2.00E-06 | Flavobacterium sp. | Bacteroidetes | / |
| PPERSA_00121720 | rRNA adenine methyltransferase | 1.00E-42 | Planktothrix | Cyanobacteria | / |
| PPERSA_00076020 | radical SAM domain protein | 8.00E-81 | Zavarzinella formosa | Planctomycetes | / |
| PPERSA_00125500 | cytidine deaminase | 8.00E-14 | Methanoculleus sp. | Methanomicrobia | / |
| PPERSA_00032590 | 2-hydroxy-6-oxo-6-phenylhexa-2,4-dienoate hydrolase | 6.00E-09 | Bordetella avium | Proteobacteria | / |
| PPERSA_00117680 | membrane-associated protein in eicosanoid and glutathione metabolism (mapeg) | 1.00E-16 | Luteimonas mephitis | Proteobacteria | / |
| PPERSA_00009640 | TPR repeat | 4.00E-12 | Microscilla marina | Bacteroidetes | / |
| PPERSA_00037920 | DTW domain protein | 1.00E-30 | Photobacterium damselae | Proteobacteria | / |
| PPERSA_00083530 | 2-nitropropane dioxygenase | 5.00E-63 | Kyrpidia tusciae | Firmicutes | / |
Figure 2. GC content and intron number distribution of the 54 HGT genes.
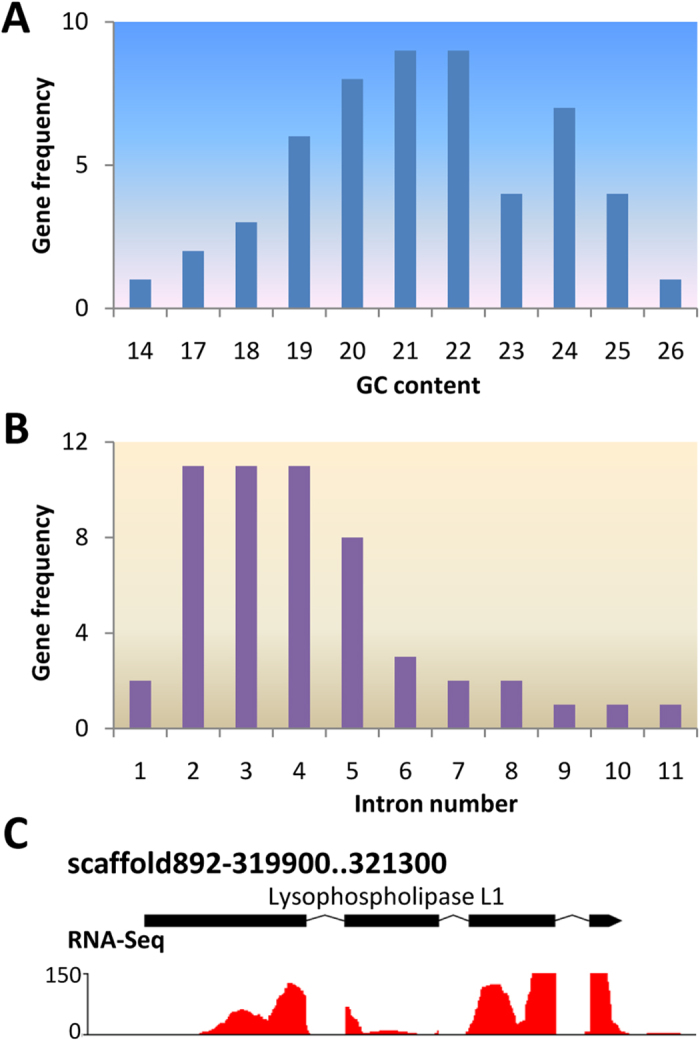
(A) the GC content distribution, similar to the GC content distribution of the assembled scaffolds; (B) the distribution of the predicted intron numbers, only two genes lack introns. (C) a RNA-Seq supported intron-containing HGT gene (PPERSA_00047700). These results suggest that the 54 HGTs belong to the P. persalinus genome rather than to bacterial contaminants.
The number of HGT genes in P. persalinus is similar to that in the free-living species T. thermophila, and far higher than the obligate parasite I. multifiliis. Therefore, the HGT genes in P. persalinus and T. thermophila were compared. In T. thermophila, 15 HGT genes are homologs of chemotaxis proteins (Table S3) which are related to the movement of an organism in response to a chemical stimulus such as the presence of food37. Fourteen genes are tetratricopeptide (TPR) repeat family homologs (Table S3) which have a variety of functions. Six are protein kinases (Table S3), the kinase families being extensively expanded in Tetrahymena compared to other organisms12. In P. persalinus, a gene ontology enrichment analysis suggested that the HGT genes are significantly enriched in functions such as oxidoreductase activity and iron ion binding, which clearly differ from the HGT genes in T. thermophila (Figure S5). Therefore, we carefully checked the functional annotations of HGT genes in P. persalinus and found a number of HGT genes (approximately 20%) may play important role in its virulence.
Cell adhesion proteins
Two of the Ig family of proteins were found in the HGT genes of P. persalinus. Domain analysis of these genes showed that both contain cadherin-like domains (Figure S6). Cadherins are a family of transmembrane proteins that play important roles in cell adhesion, forming adherens junctions to bind together cells within tissues. They are dependent on calcium ions (Ca2+) to function hence their name. Cadherins have been shown as important adhesins and invasins of pathogenic bacteria38. Comparing the transporters between the facultatively parasitic P. persalinus and the free-living T. thermophila showed that the P. persalinus-specific transporter is significantly enriched in calcium ion transport (Figure S2 and Table S2). One of the two Ig proteins HGT genes in P. persalinus contain a He_pig domain (PF05345) which contains a conserved core region of about 90 residue repeats found in several haemagglutinins and other cell-surface proteins (http://www.ncbi.nlm.nih.gov/Structure/cdd/cddsrv.cgi?uid=pfam05345), indicating that this gene may contribute to cell (e.g. erythrocyte cell) adhesion (Fig. 3).
Figure 3. Potential contributions of HGT genes in virulence of P. persalinus.
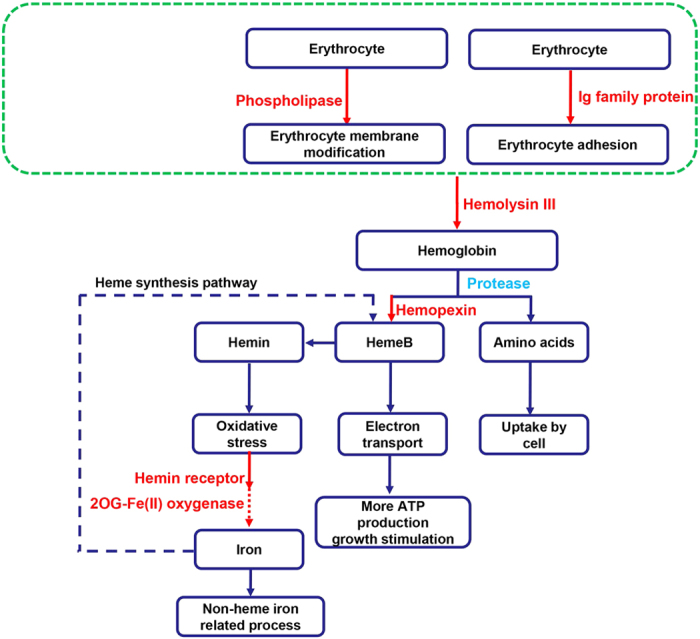
Red, HGT genes and their putative contribution to virulence; solid red arrow, processes and genes involved, supported by published studies. Red dashed arrow, potential function of HGT genes according to protein domain and gene regulatory information. Green dashed box, biological process in which the two Ig family proteins may be involved, although it is not known whether this process is necessary to the hemolysis pathway.
Hemolysis related proteins
It is well known that many bacterial pathogens induce hemolysis of host erythrocytes39. These bacteria could produce proteins, usually called hemolysin, that cause lysis of erythrocytes by destroying their cell membrane. Many proteins have been identified as hemolysins including phospholipase40 and hemolysin III41. Interestingly, three HGT phospholipase-related genes and a hemolysin III homolog were found in the P. persalinus genome (Table S2).
Bacterial phospholipases are a large group of enzymes that have a wide range of effects on host cells from minor alterations of cell membrane composition to increased vascular permeability and lethality, even at low concentrations42. In the scuticociliate Uronema marinum, it has been suggested that phospholipase C could acts as virulence factor that serves to actively disrupt host defense mechanisms43. Among the three phospholipases in P. persalinus, two (PPERSA_00098980 and PPERSA_00002080) were identified as phosphatidylinositol-specific phospholipase C (PI-PLC) and the other one (PPERSA_00047700) was identified as lysophospholipase (Table S2). Phospholipase has been reported to function in erythrocyte membrane modification and hemolysis, for example the PI-PLC, which could release acetylcholinesterase linked to phosphatidylinositol44,45,46,47. Thus, phospholipases may help P. persalinus utilize the host erythrocytes by disrupting their cell membrane (Fig. 3).
A hemolysin III family protein (Table 2, PPERSA_00035610) was identified among the HGT genes of P. persalinus. This gene is located in an assembled scaffold with 1.2 Mb in length and contains a RNA-Seq data-supported intron (Figure S7A). The coded protein contains a HlyIII domain (PF03006) and six transmembrane helices (Figure S7B). It has been shown that hemolysin III produced by the bacteria Bacillus cereus and Vibrio vulnificus is capable of hemolytic activity48. The P. persalinus hemolysin III gene closely resembles its B. cereus homolog (Figure S7C), especially in the functional domain region, suggesting that the protein for which this gene codes may play a role in the lysis of host erythrocytes (Fig. 3).
Heme related proteins
Hemolysis is the rupturing of the erythrocyte cell membrane and the release of its cytoplasm into surrounding tissue. By this process pathogens can acquire the erythrocyte cell contents and utilize it for their own metabolism. Erythrocyte cytoplasm is rich in hemoglobin which includes the iron-containing heme whose function is to bind and transport oxygen49. Hemoglobin can be digested by a series of protease enzymes, releasing its amino acids and heme. Amino acids can be directly intercepted by the pathogen, whereas free heme generates oxidative stress known to damage cells if not utilized or transformed50. In malaria, merozoites of Plasmodium invade erythrocytes, ingest host hemoglobin enclosing it in an acidic food vacuole51 and digest it using proteases52. The released heme is then incorporated into haemozoin53. Due to its abundance in the host, heme is a valuable source of iron for invading micro-organisms during hemolysis, and makes the host dramatically more susceptible to infections and their complications. Pseudocohnilembus persalinus has two hemopexin repeat protein homologs (PPERSA_00117390 and PPERSA_00079580) that appear to be of bacterial origin acquired by HGT (Table 2). These proteins are reported to have a high binding affinity for heme and are probably heme carriers50. The existence of these two HGT genes suggests that P. persalinus can uptake and utilize host iron in a similar way to bacteria (Fig. 3).
For many ciliates, e.g. Tetrahymena, the inclusion of inorganic iron salts in a culture medium produces a dramatic acceleration of growth and marked alterations in metabolism54. Iron supplementation could be used in heme synthesis and lead to an increased concentration or activity of certain heme enzymes, particularly in the electron transport chain which plays an important role in ATP synthesis thus stimulating cell growth55. In P. persalinus, the same heme de novo synthesis pathway was found as in Tetrahymena (Figure S8), suggesting that P. persalinus may also synthesize heme from iron in order to enhance growth. Therefore, it is reasonable to speculate that direct uptake of heme from the host could stimulate the reproduction of P. persalinus during infection (Fig. 3).
Hemin is the Fe(III) oxidation product of heme. An excess of hemin can interact with the cell membrane resulting in formation of reactive oxygen species (oxidative stress) and causing cellular injury56. For the host, the generation of hemin is a double-edged sword since it not only lyses pathogens57, but also induces hemolysis of erythrocytes58. Bacteria such as Yersinia enterocolitica have evolved hemin utilization systems that enable them to acquire iron by intercepting hemin using hemin receptor proteins59. A hemin receptor gene of bacterial origin (PPERSA_00031570) has been acquired by P. persalinus (Table 2), indicating that the ciliate has the ability to use hemin. In addition to being a source of iron, the binding of free hemin by the hemin receptor may help to reduce the oxidative stress for the ciliate.
Heme, or its Fe(III) oxidation product hemin, is catalytically broken down by heme oxygenase to carbon monoxide, bilirubin and iron. The iron can then be used by non-heme iron enzymes or participate the de novo heme synthesis pathway. However, no homolog of heme oxygenase was found in P. persalinus, suggesting the presence of an alternative hemin utilization system. In the bacterium Ralstonia metallidurans CH34, for example, some hemin-related proteins share a transcription factor binding site (potential operon) with 2OG-Fe(II) oxygenase (http://regprecise.lbl.gov/RegPrecise/regulon.jsp?regulon_id=9937), indicating that 2OG-Fe(II) oxygenase may be involved in the hemin utilization process, although its function was not determined. In P. persalinus, two 2OG-Fe(II) oxygenase (PPERSA_00130810 and PPERSA_00076120) were found as HGT genes (Table 2), raising the possibility that 2OG-Fe(II) oxygenase could function as heme oxygenase, cleaving the hemin ring to release the iron (Fig. 3). Thus, it appears that P. persalinus acquired by HGT almost the whole pathway for hemolysis and the utilization of heme.
One of the most salient clinical manifestations of scuticociliatosis is haemorrhagic lesions60. Histopathological observations typically show many scuticociliates in the blood vessels, gills, fins, skin muscle, brain and lamina propria of the digestive tract, accompanied by necrosis and haemorrhages61. Like Uronema marinum which destroys host tissue by proteases62, P. persalinus may also utilize proteases to break the skin–blood barrier and gain entry to internal organs. Virulence HGT genes may thus contribute to the subsequent destruction of red blood cells and the acquisition of host-derived nutrients and energy for ciliate cell proliferation.
It has been reported that scuticociliatosis often accompanies bacterial disease (e.g. vibriosis), and the increased bacterial load probably helps ciliates to thrive and proliferate during the initial phase of infection60. Therefore it can be speculated that the synergistic invasion by pathogenic bacteria and scuticociliates, and the presence of both in a shared environment, may provide the opportunity for the transfer of genetic materials from the former to the latter. Alternatively, many ciliates harbor bacterial symbionts63 which may also provide the conditions for HGT. Evidence for the presence of endosymbiotic bacteria in P. persalinus comes from the preliminary genome assembly, the second and third peaks with 41% and 67% GC (Fig. 1) representing two bacterial species. The homology search results showed these could be species closely related to Pseudoalteromonas and Halomonas, respectively. They are very likely bacterial symbionts harbored in P. persalinus because sequences of Escherichia coli DH5 alpha, the only food organism supplied to P. persalinus cultured in laboratory, were not found. The 54 HGT genes in P. persalinus were not, however, included in these two peaks. Although homology searches showed that these 54 HGT genes do not have an enriched bacterial source, many of the best homologs occur in two large bacterial classes: Actinobacteria and Gammaproteobacteria (Table 2). Therefore, it is more likely that the HGT genes in P. persalinus have multiple independent origins.
Based on infection experiments, some researchers have concluded that the scuticociliates P. persalinus, P. hargisi and U. marinum are not primary pathogens of oysters, rather they are non-pathogenic, free-living, bacteriophagous and/or saprophagous ciliates that opportunistically attached to lesions on the host that are originally produced by bacterial infection or some other cause61. However, the identification here of virulence HGT genes in P. persalinus provides evidence that there is a molecular basis for its pathogenicity. Recent attempts to develop vaccines targeting antigens such as those responsible for host cell immobilization, proton-translocating inorganic pyrophosphatases, cathepsin L-like cysteine protease, etc. have met with limited success because of different levels of virulence and serotype-specific protection among species/strains of pathogens64,65,66,67. It is anticipated that the virulence HGT genes identified here will help us to gain a better understanding of the pathology of scuticociliatosis and provide potential antigen candidates for the development of anti-scuticociliatosis vaccines.
Conclusions
In summary, we report the genome of scuticociliate Pseudocohnilembus persalinus, the first marine ciliate genome as far as we know. The genome of P. persalinus genome is 55.5 Mb, i.e., about half the size of the model free-living ciliate Tetrahymena thermiphila. The P. persalinus genome harbors many prokaryote-derived horizontally transferred genes; function analysis showed that many of these HGT genes are potential virulence factors. These findings help to increase our knowledge and understanding of the mechanism of the common fish disease, scuticociliatosis.
Methods
Pseudocohnilembus persalinus culture, total DNA and total RNA extraction
Pseudocohnilembus persalinus was isolated from water in a shrimp-farming pond (36°08′N,120°43′E; water temperature 27 °C; salinity 20%; pH 7.5) in Qingdao, China68. The species was identified by its morphology, morphogenesis68 and 18S rDNA marker (Figure S9). In order to obtain sufficient DNA and RNA material for sequencing, P. persalinus cells were cultured in the laboratory using sterilized sea water with Escherichia coli DH5 alpha as a food source. Contamination by bacteria was prevented by treating the cell cultures with lysozyme (200 μg/ml for 2 hours at 28 °C ) before the DNA and RNA extraction. The total DNA was extracted using the method described for Tetrahymena69, and the total RNA was extracted using the RNeasy Protect Cell Mini Kit (Qiagen, Valencia, CA) according to protocol in TetraFGD70.
Pseudocohnilembus persalinus genome and transcriptome sequencing
The P. persalinus genome was sequenced using the Illumina platform. Paired-end (about 190 bp insert size) and mate-pair (about 2 Kb insert size) libraries were constructed and sequenced using the standard protocol of Illumina (https://icom.illumina.com/). Briefly, genomic DNA was fragmented and fragments of appropriate size (see above) were selected. For mate-pair library construction, fragment ends were biotinylated and circularized, and the fragments were then enriched using biotin. Fragment ends were then repaired, A-tailed and ligated with sequencing adaptors. Adaptor-ligated fragments were PCR amplified using Phusion polymerase, denatured with sodium hydroxide and diluted in hybridization buffer. The prepared libraries were loaded onto the flowcell and sequenced.
For transcriptome sequencing, Poly-A mRNAs were isolated using Dynal magnetic beads (Invitrogen) and fragmented by heating to 94 °C. First strand cDNAs were synthesized with reverse transcriptase and random hexamer primers, and then the second strands were synthesized with DNA polymerase and random hexamer primers. Double strand cDNAs were end-repaired and a single adenosine moiety was added. Illumina adapters were ligated and gel-electrophoresis was used to select DNA fragments about 200 bp size. Libraries were PCR-amplified using Phusion polymerase. Cluster formation, primer hybridization and pair-end sequencing were performed using proprietary reagents according to manufacturer-recommended protocols (https://icom.illumina.com/).
Genome assembly and bacterial contamination exclusion
The paired-end reads and mate-paired reads of the Illumina sequencing were assembled using SOAPdenovo software71, which uses the de Bruijn graph data structure to construct contigs. A series of K-mer values (from 33 to 79) were used to assemble the P. persalinus genome, and the assembly with the longest N50 length was chosen by deleting scaffolds shorter than 1 Kb. Bacterial contaminants were excluded in two stages, sample preparation (see above) and bioinformatics analysis. In the bioinformatics analysis stage, bacterial contaminants were first excluded using the GC content, any scaffolds with a GC content more than 25% being discarded. The remaining scaffolds were then BLAST searched against the NCBI non-redundant protein database; any scaffolds with more than 50% hits belonging to the prokaryotes were excluded. The remaining scaffolds were regarded as the P. persalinus genome assembly sequences.
Gene prediction and annotation
Using the RNA-Seq data, transcripts were both de novo assembled using Trinity72, and reference-guided assembled using the Tophat73 and Cufflinks74 pipeline. A combination of de novo and reference-guided assembled transcripts were validated by aligning the putative transcripts onto the assembled genome using PASA75. A set of the so-called best models generated by PASA was used to train the gene prediction software Augustus76 and GlimmerHMM77. The training parameters were then used by the two programs to de novo predict the gene models. The Augustus software could accept the cDNA or protein evidence, therefore the assembled transcripts were also used as the cDNA evidence for Augustus. Finally, an integrated set of gene models was created using Evidence Modeler78 by merging all of the predicted gene models.
Homologs of P. persalinus genes were BLAST searched against the NCBI non-redundant protein database. The KEGG pathway information was annotated using the KAAS server (http://www.genome.jp/kegg/kaas/). Protein domain information was annotated using the Pfam database79. Gene ontology (GO) information was annotated using the Goanna server (http://agbase.msstate.edu/cgi-bin/tools/GOanna.cgi). For each gene set, GO enrichment analysis was also carried out using BinGO80. FDR correction was used to control the false positive rate. If a GO term in a test gene set showed a corrected p value less than 0.05 compared with the reference set (all the GO annotated genes), the GO term (function) was determined to be significantly overrepresented in the test gene set. To annotate the proteases, a batch BLAST (http://merops.sanger.ac.uk/cgi-bin/batch_blast) was performed against the MEROPS database using all predicted genes. Genes with BLAST hit E-values less than 1e-10 were regarded as proteases, and the best hit in the MEROPS database was used to assign the protease classes. To annotate the membrane transporters, all the genes were BLAST searched against the TCDB transporters database81, and the transmembrane helices were predicted using SCAMPI82, Toppred83 and TMHMM84. A gene was regarded as a membrane transporter if it had a BLAST hit in the TCDB transporters database with an E-value less than 1e-03 and showed at least one transmembrane helix in all three programs.
Bacterial horizontal transferred genes identification
Pseudocohnilembus persalinus HGT genes were identified by two steps, similar to the strategy used in Ricard et al.85: Firstly, similarity searches were performed to screen the potential prokaryotic origin genes by using a BLASTP search against to the NCBI non-redundant database (Figure S10). In this step, the E-value 1E-05 was used as the cutoff, and if a P. persalinus gene had a best hit belonging to the prokaryotes, it was regarded as a candidate gene. Phylogenetic approaches are widely used to identify HGT genes85,86,87, so we also employed phylogenetic analyses to to further identify the P. persalinus HGT genes based on the screened candidates. All candidate genes retrieved from the first step were BLASTP searched (E-value: 1E-05) against both the prokaryote and eukaryote protein databases (generated from the Refseq data: ftp://ftp.ncbi.nlm.nih.gov/refseq/release/) in order to retrieve homologs in both eukaryotes and prokaryotes. For a protein in P. persalinus, if there were homologs (E-value: 1E-05) in both domains, the sequences were retrieved in order to construct phylogenetic trees. Two methods (programs) were used, namely FASTTREE88 and PHYML89. For FASTTREE, all the homologs with E-value less than 1E-05 were used to construct the tree; for PHYML, only the top ten homologs (if present) were used to construct the tree. Sequences alignments were performed using MUSCLE90. A gene clustered in the prokaryotic clade which had a eukaryotic outgroup was accepted as an HGT gene, a technique now widely used to identify HGT genes85,86,87,91. As shown in Figure S11, only gene in P. persalinus with this kind of tree topology was considered an HGT gene. If a P. persalinus gene only has homologs with E-values less than 1E-05 in prokaryotes, the phylogenetic analysis could not work. HGTs were determined if there were at least 5 prokaryotic hits and the E-value of the best hit in prokaryotes and eukaryotes differed by more than 5 orders of magnitude (i.e., the E-value of best hit in eukaryotes will be larger than 1 if the best prokaryotic hit E-value is 1E-05). Some HGT genes may have diverged significantly after the HGT event. For example, gene PPERSA_00031570 with an E-value 1E-06 to its best BLAST hit (prokaryotic protein), has a bacterial-like globin (Pfam domain: PF01152) and is therefore highly likely to be of prokaryotes origin. In such cases, an E-value 1E-05 was used if no homolog was found in the eukaryotes. In addition, to confirm the existence of HGT genes, PCR analysis was performed for 20 of 54 HGT genes identified by the bioinformatics (for primers, see Table S4), 100% of which were verified.
Data access
This Whole Genome Shotgun project has been deposited at DDBJ/EMBL/GenBank under the accession LDAU00000000. The version described in this paper is version LDAU01000000. The raw genome sequences reads have been deposited in Sequence Read Archive (SRA) under accession SRX883501 and SRX883503. Transcriptome data has also been deposited in SRA under accession SRX849480.
Additional Information
How to cite this article: Xiong, J. et al. Genome of the facultative scuticociliatosis pathogen Pseudocohnilembus persalinus provides insight into its virulence through horizontal gene transfer. Sci. Rep. 5, 15470; doi: 10.1038/srep15470 (2015).
Supplementary Material
Acknowledgments
We thank Prof. Weibo Song (Ocean University of China) for sharing the P. persalinus cells. We also thank the critical comments from Jun Zhou (Harvard University) and Chengjie Fu (Uppsala University). This work was supported by the Natural Science Foundation of China (No. 31301930) to JX, the Projects of International Cooperation and Exchanges, Ministry of Science and Technology of China (No. 2013DFG32390) and the Natural Science Foundation of China (No. 31372168) to WM.
Footnotes
Author Contributions W.M. and J.X. designed the project. M.T., X.P. and J.X. collected samples and prepared DNA and RNA. J.X., G.W., J.C., C.J. and D.Y. performed the analyses. J.X., W.M. and A.W. wrote the manuscript. All authors read and approved the final manuscript.
References
- Noga E. J. Fish disease: diagnosis and treatment (John Wiley & Sons, 2010). [Google Scholar]
- Cheung P. J., Nigrelli R. F. & Ruggieri G. D. Studies on the Morphology of Uronema-Marinum Dujardin (Ciliatea, Uronematidae) with a Description of the Histopathology of the Infection in Marine Fishes. Journal of Fish Diseases 3, 295–303 (1980). [Google Scholar]
- Kim S. M., Cho J. B., Kim S. K., Nam Y. K. & Kim K. H. Occurrence of scuticociliatosis in olive flounder Paralichthys olivaceus by Phiasterides dicentrarchi (Ciliophora : Scuticociliatida). Diseases of Aquatic Organisms 62, 233–238 (2004). [DOI] [PubMed] [Google Scholar]
- Jee B. Y., Kim Y. C. & Park M. S. Morphology and biology of parasite responsible for scuticociliatosis of cultured olive flounder Paralichthys olivaceus. Diseases of Aquatic Organisms 47, 49–55 (2001). [DOI] [PubMed] [Google Scholar]
- Iglesias R. et al. Philasterides dicentrarchi (Ciliophora, Scuticociliatida) as the causative agent of scuticociliatosis in farmed turbot Scophthalmus maximus in Galicia (NW Spain). Diseases of Aquatic Organisms 46, 47–55 (2001). [DOI] [PubMed] [Google Scholar]
- Puig L., Traveset R., Palenzuela O. & Padros F. Histopathology of experimental scuticociliatosis in turbot Scophthalmus maximus. Diseases of Aquatic Organisms 76, 131–140 (2007). [DOI] [PubMed] [Google Scholar]
- Clark T. G., Lin T.-L. & Dickerson H. W. Surface immobilization antigens of Ichthyophthirius multifiliis: their role in protective immunity. Annual Review of Fish Diseases 5, 113–131 (1995). [Google Scholar]
- Wang X. & Dickerson H. W. Surface immobilization antigen of the parasitic ciliate Ichthyophthirius multifiliis elicits protective immunity in channel catfish (Ictalurus punctatus). Clin Diagn Lab Immunol 9, 176–81 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan X., Al-Farraj S. A., Gao F. & Gu F. Morphological reports on two species of Dexiotricha (Ciliophora, Scuticociliatia), with a note on the phylogenetic position of the genus. Int J Syst Evol Microbiol 64, 680–8 (2014). [DOI] [PubMed] [Google Scholar]
- Lynn D. H. The ciliated protozoa: characterization, classification, and guide to the literature, (Springer, 2008). [Google Scholar]
- Asai D. J. & Editors F. J. Tetrahymena thermophila 580 (Academic press, Methods in Cell Biology, 2000). [Google Scholar]
- Eisen J. A. et al. Macronuclear genome sequence of the ciliate Tetrahymena thermophila, a model eukaryote. PLoS Biol 4, e286 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aury J. M. et al. Global trends of whole-genome duplications revealed by the ciliate Paramecium tetraurelia. Nature 444, 171–178 (2006). [DOI] [PubMed] [Google Scholar]
- Beisson J. et al. Paramecium tetraurelia: the renaissance of an early unicellular model. Cold Spring Harb Protoc 2010, pdb emo140 (2010). [DOI] [PubMed] [Google Scholar]
- Evans F. R. & Thompson J. C. Pseudocohnilembidae N. Fam. Hymenostome Ciliate Family Containing 1 Genus Pseudocohnilembus N. G. With 3 New Species. Journal of Protozoology 11, 344-& (1964). [Google Scholar]
- Weibo S. Morphological and taxonomical studies on some marine scuticociliates from China Sea, with description of two new species, Philasterides armatalis sp. n. and Cyclidium varibonneti sp. n.(Protozoa: Ciliophora: Scuticociliatida). Acta Protozool 39, 295–322 (2000). [Google Scholar]
- Pomp R. & Wilbert N. Taxononmic and ecological studies of ciliates from Australian saline soils: colpodids and hymenostomate ciliates. Marine and Freshwater Research 39, 479–495 (1988). [Google Scholar]
- Jones S. R. M., Prosperi-Porta G. & LaPatra S. E. First Isolation of Pseudocohnilembus persalinus (Ciliophora: Scuticociliatida) From Freshwater-Reared Rainbow Trout, Oncorhynchus mykiss. Journal of Parasitology 96, 1014–1016 (2010). [DOI] [PubMed] [Google Scholar]
- Kim S. M. et al. Pseudocohnilembus persalinus (Ciliophora : Scuticociitida) is an additional species causing scuticociliatosis in olive flounder Paralichthys olivaceus. Diseases of Aquatic Organisms 62, 239–244 (2004). [DOI] [PubMed] [Google Scholar]
- Coyne R. S. et al. Comparative genomics of the pathogenic ciliate Ichthyophthirius multifiliis, its free-living relatives and a host species provide insights into adoption of a parasitic lifestyle and prospects for disease control. Genome Biology 12 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sajid M. & McKerrow J. H. Cysteine proteases of parasitic organisms. Molecular and Biochemical Parasitology 120, 1–21 (2002). [DOI] [PubMed] [Google Scholar]
- Sakanari J. A., Staunton C. E., Eakin A. E., Craik C. S. & Mckerrow J. H. Serine Proteases from Nematode and Protozoan Parasites - Isolation of Sequence Homologs Using Generic Molecular Probes. Proceedings of the National Academy of Sciences of the United States of America 86, 4863–4867 (1989). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lovett J. L. & Sibley L. D. Intracellular calcium stores in Toxoplasma gondii govern invasion of host cells. Journal of Cell Science 116, 3009–3016 (2003). [DOI] [PubMed] [Google Scholar]
- Billker O., Lourido S. & Sibley L. D. Calcium-Dependent Signaling and Kinases in Apicomplexan Parasites. Cell Host & Microbe 5, 612–622 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crosbie P. B. & Munday B. L. Environmental factors and chemical agents affecting the growth of the pathogenic marine ciliate Uronema nigricans. Dis Aquat Organ 36, 213–9 (1999). [DOI] [PubMed] [Google Scholar]
- Syvanen M. Cross-Species Gene-Transfer - Implications for a New Theory of Evolution. Journal of Theoretical Biology 112, 333–343 (1985). [DOI] [PubMed] [Google Scholar]
- Akiba T., Koyama K., Ishiki Y., Kimura S. & Fukushima T. On the mechanism of the development of multiple-drug-resistant clones of Shigella. Jpn J Microbiol 4, 219–27 (1960). [DOI] [PubMed] [Google Scholar]
- Jones J. T., Furlanetto C. & Kikuchi T. Horizontal gene transfer from bacteria and fungi as a driving force in the evolution of plant parasitism in nematodes. Nematology 7, 641–646 (2005). [Google Scholar]
- Griffith F. The significance of pneumococcal types. Journal of Hygiene 27, 113–159 (1928). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore P. & Lindsay J. Genetic variation among hospital isolates of methicillin-sensitive Staphylococcus aureus: evidence for horizontal transfer of virulence genes. Journal of clinical microbiology 39, 2760–2767 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coburn P. S., Baghdayan A. S., Dolan G. T. & Shankar N. Horizontal transfer of virulence genes encoded on the Enterococcus faecalis pathogenicity island. Mol Microbiol 63, 530–44 (2007). [DOI] [PubMed] [Google Scholar]
- de la Cruz F. & Davies J. Horizontal gene transfer and the origin of species: lessons from bacteria. Trends Microbiol 8, 128–33 (2000). [DOI] [PubMed] [Google Scholar]
- Keeling P. J. & Palmer J. D. Horizontal gene transfer in eukaryotic evolution. Nat Rev Genet 9, 605–18 (2008). [DOI] [PubMed] [Google Scholar]
- Rosewich U. L. & Kistler H. C. Role of Horizontal Gene Transfer in the Evolution of Fungi. Annu Rev Phytopathol 38, 325–363 (2000). [DOI] [PubMed] [Google Scholar]
- de Wit P. J. Molecular characterization of gene-for-gene systems in plant-fungus interactions and the application of avirulence genes in control of plant pathogens. Annu Rev Phytopathol 30, 391–418 (1992). [DOI] [PubMed] [Google Scholar]
- Bruto M., Prigent-Combaret C., Luis P., Moenne-Loccoz Y. & Muller D. Frequent, independent transfers of a catabolic gene from bacteria to contrasted filamentous eukaryotes. Proceedings of the Royal Society B-Biological Sciences 281 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macnab R. M. & Koshland D. E. Jr. The gradient-sensing mechanism in bacterial chemotaxis. Proc Natl Acad Sci USA 69, 2509–12 (1972). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niemann H. H., Schubert W. D. & Heinz D. W. Adhesins and invasins of pathogenic bacteria: a structural view. Microbes Infect 6, 101–12 (2004). [DOI] [PubMed] [Google Scholar]
- Neter E. Bacterial hemagglutination and hemolysis. Bacteriol Rev 20, 166–88 (1956). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Titball R. W. Bacterial phospholipases. Symp Ser Soc Appl Microbiol 27, 127S–137S (1998). [PubMed] [Google Scholar]
- Baida G. E. & Kuzmin N. P. Mechanism of action of hemolysin III from Bacillus cereus. Biochimica Et Biophysica Acta-Biomembranes 1284, 122–124 (1996). [DOI] [PubMed] [Google Scholar]
- Songer J. G. Bacterial phospholipases and their role in virulence. Trends in Microbiology 5, 156–161 (1997). [DOI] [PubMed] [Google Scholar]
- Seo J. S. et al. Uronema marinum: Identification and biochemical characterization of phosphatidylcholine-hydrolyzing phospholipase C. Experimental Parasitology 110, 22–29 (2005). [DOI] [PubMed] [Google Scholar]
- Low M. G. & Finean J. B. Modification of Erythrocyte-Membranes by a Purified Phosphatidylinositol-Specific Phospholipase-C (Staphylococcus-Aureus). Biochemical Journal 162, 235–240 (1977). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Futerman A. H., Low M. G., Michaelson D. M. & Silman I. Solubilization of Membrane‐Bound Acetyicholinesterase by a Phosphatidylinositol‐Specific Phospholipase C. Journal of neurochemistry 45, 1487–1494 (1985). [DOI] [PubMed] [Google Scholar]
- Taguchi R., Suzuki K., Nakabayashi T. & Ikezawa H. Acetyicholinesterase Release from Mammalian Erythrocytes by Phosphatidylinositol-Specific Phospholipase C of Bacillus thuringiensis and Characterization of the Released Enzyme. Journal of biochemistry 96, 437–446 (1984). [DOI] [PubMed] [Google Scholar]
- Toutant J. P., Roberts W. L., Murray N. R. & Rosenberry T. L. Conversion of human erythrocyte acetylcholinesterase from an amphiphilic to a hydrophilic form by phosphatidylinositol‐specific phospholipase C and serum phospholipase D. European Journal of Biochemistry 180, 503–508 (1989). [DOI] [PubMed] [Google Scholar]
- Chen Y. C., Chang M. C., Chuang Y. C. & Jeang C. L. Characterization and virulence of hemolysin III from Vibrio vulnificus. Current Microbiology 49, 175–179 (2004). [DOI] [PubMed] [Google Scholar]
- Hendrickson W. A. Hemoglobin - Structure, Function, Evolution, and Pathology - Dickerson,Re, Geis,I. Nature 304, 193–193 (1983). [Google Scholar]
- Piccard H., Van Den Steen P. E. & Opdenakker G. Hemopexin domains as multifunctional liganding modules in matrix metal loproteinases and other proteins. Journal of Leukocyte Biology 81, 870–892 (2007). [DOI] [PubMed] [Google Scholar]
- Aikawa M., Huff C. G. & Sprinz H. Comparative Feeding Mechanisms of Avian and Primate Malarial Parasites. Military Medicine 131, 969-& (1966). [PubMed] [Google Scholar]
- Banerjee R. et al. Four plasmepsins are active in the Plasmodium falciparum food vacuole, including a protease with an active-site histidine. Proceedings of the National Academy of Sciences of the United States of America 99, 990–995 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egan T. J. et al. Fate of haem iron in the malaria parasite Plasmodium falciparum. Biochemical Journal 365, 343–347 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chipman A. D. et al. The First Myriapod Genome Sequence Reveals Conservative Arthropod Gene Content and Genome Organisation in the Centipede Strigamia maritima. Plos Biology 12 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shug A. L., Elson C. & Shrago E. Effect of iron on growth, cytochromes, glycogen and fatty acids of Tetrahymena pyriformis. J Nutr 99, 379–86 (1969). [DOI] [PubMed] [Google Scholar]
- Balla J. et al. Endothelial-Cell Heme Uptake from Heme-Proteins - Induction of Sensitization and Desensitization to Oxidant Damage. Proceedings of the National Academy of Sciences of the United States of America 90, 9285–9289 (1993). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orjih A. U., Banyal H. S., Chevli R. & Fitch C. D. Hemin Lyses Malaria Parasites. Science 214, 667–669 (1981). [DOI] [PubMed] [Google Scholar]
- Li S. D., Su Y. D., Li M. & Zou C. G. Hemin-mediated hemolysis in erythrocytes: Effects of ascorbic acid and glutathione. Acta Biochimica Et Biophysica Sinica 38, 63–69 (2006). [DOI] [PubMed] [Google Scholar]
- Stojiljkovic I. & Hantke K. Hemin Uptake System of Yersinia-Enterocolitica - Similarities with Other Tonb-Dependent Systems in Gram-Negative Bacteria. Embo Journal 11, 4359–4367 (1992). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azad I., Al-Marzouk A., James C., Almatar S. & Al-Gharabally H. Scuticociliatosis-associated mortalities and histopathology of natural infection in cultured silver pomfret (Pampus argenteus Euphrasen) in Kuwait. Aquaculture 262, 202–210 (2007). [Google Scholar]
- Song J. Y. et al. Pathogenicity of Miamiensis avidus (syn. Philasterides dicentrarchi), Pseudocohnilembus persalinus, Pseudocohnilembus hargisi and Uronema marinum (Ciliophora, Scuticociliatida). Diseases of Aquatic Organisms 83, 133–43 (2009). [DOI] [PubMed] [Google Scholar]
- Lee E. H., Kim C. S., Cho J. B., Ahn K. J. & Kim K. H. Measurement of protease activity of live Uronema marinun (Ciliata: Scuticociliatida) by fluorescence polarization. Dis Aquat Organ 54, 85–8 (2003). [DOI] [PubMed] [Google Scholar]
- Görtz H.-D. Symbiotic associations between ciliates and prokaryotes. in The prokaryotes 364–402 (Springer, 2006). [Google Scholar]
- Mallo N., Lamas J., Piazzon C. & Leiro J. M. Presence of a plant-like proton-translocating pyrophosphatase in a scuticociliate parasite and its role as a possible drug target. Parasitology. 1–14 (2014). [DOI] [PubMed] [Google Scholar]
- Shin S. P. et al. Expression and characterization of cathepsin L-like cysteine protease from Philasterides dicentrarchi. Parasitol Int 63, 359–65 (2014). [DOI] [PubMed] [Google Scholar]
- Leon-Rodriguez L., Luzardo-Alvarez A., Blanco-Mendez J., Lamas J. & Leiro J. Biodegradable microparticles covalently linked to surface antigens of the scuticociliate parasite P. dicentrarchi promote innate immune responses in vitro. Fish Shellfish Immunol 34, 236–43 (2013). [DOI] [PubMed] [Google Scholar]
- Leon-Rodriguez L., Luzardo-Alvarez A., Blanco-Mendez J., Lamas J. & Leiro J. A vaccine based on biodegradable microspheres induces protective immunity against scuticociliatosis without producing side effects in turbot. Fish Shellfish Immunol 33, 21–7 (2012). [DOI] [PubMed] [Google Scholar]
- Pan X., Ma H., Shao C., Lin X. & Hu X. Stomatogenesis and morphological redescription of Pseudocohnilembus persalinus (Ciliophora: Scuticociliatida). Acta Hydrobiologica Sinica 36(3), 489–494 (2012). [Google Scholar]
- Saveliev S. V. PCR-based detection of a rare linear DNA in cell culture. Biol Proced Online 4, 70–80 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiong J. et al. Tetrahymena functional genomics database (TetraFGD): an integrated resource for Tetrahymena functional genomics. Database (Oxford) 2013, bat008 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie Y. et al. SOAPdenovo-Trans: de novo transcriptome assembly with short RNA-Seq reads. Bioinformatics (2014). [DOI] [PubMed] [Google Scholar]
- Grabherr M. G. et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nature Biotechnology 29, 644–U130 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trapnell C., Pachter L. & Salzberg S. L. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trapnell C. et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nature Protocols 7, 562–578 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haas B. J. et al. Improving the Arabidopsis genome annotation using maximal transcript alignment assemblies. Nucleic Acids Research 31, 5654–5666 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanke M., Diekhans M., Baertsch R. & Haussler D. Using native and syntenically mapped cDNA alignments to improve de novo gene finding. Bioinformatics 24, 637–644 (2008). [DOI] [PubMed] [Google Scholar]
- Majoros W. H., Pertea M. & Salzberg S. L. TigrScan and GlimmerHMM: two open source ab initio eukaryotic gene-finders. Bioinformatics 20, 2878–2879 (2004). [DOI] [PubMed] [Google Scholar]
- Haas B. J. et al. Automated eukaryotic gene structure annotation using EVidenceModeler and the program to assemble spliced alignments. Genome Biology 9 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bateman A. et al. The Pfam protein families database. Nucleic Acids Research 28, 263–6 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maere S., Heymans K. & Kuiper M. BiNGO: a Cytoscape plugin to assess overrepresentation of Gene Ontology categories in Biological Networks. Bioinformatics 21, 3448–3449 (2005). [DOI] [PubMed] [Google Scholar]
- Saier M. H., Tran C. V. & Barabote R. D. TCDB: the Transporter Classification Database for membrane transport protein analyses and information. Nucleic Acids Research 34, D181–D186 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernsel A. et al. Prediction of membrane-protein topology from first principles. Proceedings of the National Academy of Sciences of the United States of America 105, 7177–7181 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vonheijne G. Membrane-Protein Structure Prediction - Hydrophobicity Analysis and the Positive-inside Rule. Journal of Molecular Biology 225, 487–494 (1992). [DOI] [PubMed] [Google Scholar]
- Sonnhammer E. L., Von Heijne G. & Krogh A. A hidden Markov model for predicting transmembrane helices in protein sequences. Proc Int Conf Intell Syst Mol Biol 6, 175–82 (1998). [PubMed] [Google Scholar]
- Ricard G. et al. Horizontal gene transfer from Bacteria to rumen Ciliates indicates adaptation to their anaerobic, carbohydrates-rich environment. Bmc Genomics 7 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Z. W., Shen Y. H., Xiang Z. H. & Zhang Z. Pathogen-origin horizontally transferred genes contribute to the evolution of Lepidopteran insects. Bmc Evolutionary Biology 11 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanhope M. J. et al. Phylogenetic analyses do not support horizontal gene transfers from bacteria to vertebrates. Nature 411, 940–944 (2001). [DOI] [PubMed] [Google Scholar]
- Price M. N., Dehal P. S. & Arkin A. P. FastTree 2-Approximately Maximum-Likelihood Trees for Large Alignments. Plos One 5 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guindon S., Dufayard J. F., Hordijk W., Lefort V. & Gascuel O. PhyML: Fast and Accurate Phylogeny Reconstruction by Maximum Likelihood. Infection Genetics and Evolution 9, 384–385 (2009). [Google Scholar]
- Edgar R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research 32, 1792–1797 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao H. et al. Host-to-Pathogen Gene Transfer Facilitated Infection of Insects by a Pathogenic Fungus. Plos Pathogens 10 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.