

Abstract
PknB is an essential serine/threonine protein kinase required for mycobacterial cell division and cell-wall biosynthesis. Here we demonstrate that overexpression of the external PknB_PASTA domain in mycobacteria results in delayed regrowth, accumulation of elongated bacteria and increased sensitivity to β-lactam antibiotics. These changes are accompanied by altered production of certain enzymes involved in cell-wall biosynthesis as revealed by proteomics studies. The growth inhibition caused by overexpression of the PknB_PASTA domain is completely abolished by enhanced concentration of magnesium ions, but not muropeptides. Finally, we show that the addition of recombinant PASTA domain could prevent regrowth of Mycobacterium tuberculosis, and therefore offers an alternative opportunity to control replication of this pathogen. These results suggest that the PknB_PASTA domain is involved in regulation of peptidoglycan biosynthesis and maintenance of cell-wall architecture.
Keywords: Mycobacterium tuberculosis, serine/threonine protein kinase, magnesium, muropeptides, PknB, PASTA domain
1. Introduction
Serine/threonine protein kinases (STPKs) are widely distributed in Gram-positive and Gram-negative bacteria [1]. Mycobacterium tuberculosis, the causative agent of tuberculosis, possesses 11 STPKs [2] and two of them, PknA and PknB, are indispensable for growth in laboratory culture [3–5], while PknE [6], PknG [7,8] and PknH [9] have been implicated in M. tuberculosis virulence. The essential PknB kinase belongs to a distinct family of STPKs found only in Gram-positive bacteria [10]. The important feature of these kinases is the presence of the so-called PASTA (penicillin-binding protein and serine/threonine kinase associated) domains in the surface-exposed region [11]. In Firmicutes, PASTA-domain-containing kinases are not essential for growth. In Staphylococcus aureus, an Stk1 mutant was impaired in virulence and had higher resistance to Triton X-100 and fosmidomycin [12], while in S. pneumoniae StkP kinase was important for competence, biofilm formation and virulence [13]. More detailed investigation of the role of StkP in S. pneumoniae established that it has a crucial role in coordinating cell division and peptidoglycan synthesis during growth [14]. By contrast, PrkC in Bacillus subtilis was shown to be important for survival in stationary phase. A prkC deletion mutant had a significantly lower optical density in stationary phase compared with the wild-type bacilli [15]. However, in later studies it was demonstrated that PrkC regulated a novel muropeptide-mediated germination pathway [16] and possibly remodelling of the cell wall via controlling expression of YocH muralytic enzyme [17].
In Streptomyces coelicolor, a PASTA domain containing kinase PknB regulates carbon flux and antibiotic production, and is not essential for growth [18]. In Corynebacterium glutamicum, PknB is also dispensable for growth, and a significant change in replication and cell shape could only be detected in mutants missing several STPKs [19]. In this bacterium, PknB apparently regulates polymerization of FtsZ; however, the precise mechanism and biological significance of this observation require further investigation.
Mycobacteria appear to be a unique bacterial group in which PknB is essential for growth [3–5]. Its overexpression or partial depletion in M. smegmatis and M. bovis BCG caused dramatic alterations of cellular morphology and growth inhibition; the enzymatic activity of PknB was demonstrated to be essential for the observed effects [4,5]. Over the past decade, great progress has been achieved in the identification of PknB substrates [10]. They include proteins from various functional categories: cell-wall enzymes—InhA [20], PbpA [21] and MabA [22]; regulatory proteins—SigH [23], GarA [24] and FhaA [25]; and proteins involved in cell division—Wag31 [4] and MviN [26]. Also, PknB regulates an ‘oxygen-mediated replication switch’, and hence transition to dormancy and resuscitation [27]. Therefore, it is not surprising that PknB has been considered as a critical drug target for development of anti-tuberculosis antimicrobials. Possible application of PknB kinase inhibitors for killing of M. tuberculosis was investigated. Yet only limited success has been achieved, mainly because of poor penetration of these agents into mycobacterial cells [28].
PknB consists of several domains (figure 1); functions for some of them have been established, while the precise role of others remains unknown [29]. The components of PknB include a conserved catalytic kinase domain, a juxtamembrane part attached to a membrane-spanning region and surface-exposed sensory component, consisting of PASTA, designated as PknB_PASTA domain [30,31]. PASTA domains have been proposed to recognize growing strands of nascent peptidoglycan and activate STPKs. Structural studies showed that B. subtilis PASTA domain binds synthetic muropeptides at relatively high affinity, with the presence of the diaminopimelic acid in the muropeptide being crucial for this binding [32]. The PknB_PASTA domain from mycobacteria can also bind synthetic muropeptides; however, it remains unclear whether this binding influences activation of PknB, bacterial growth and resuscitation [33].
Figure 1.

Schematic representation of PknB constructs and strains generated in this study. Kinase, N-terminal kinase domain (aa 1–279); TM, transmembrane region (aa 331–354); TM-PASTA, penicillin and serine or threonine kinase-associated domain (aa 331–627); MYC-HIS, His-Tag; PASTA (aa 354–627).
In this study, we investigated whether the extracellular PknB_PASTA domain itself plays a distinct role in mycobacterial growth, and therefore can be potentially used as a drug target or a novel chemotherapeutic agent. The results presented here suggest that the PASTA domain probably recognizes growing peptidoglycan strands, and regulates production of penicillin-binding proteins and distribution of proteins involved in division and generation of septum. Furthermore, our data indicate that the interactions of the PknB_PASTA domain with the cell wall are important, and that its abundance may serve as a controlling mechanism for the integrity of the cell wall and bacterial growth.
2. Material and methods
2.1. Bacterial strains and growth media
Mycobacterium smegmatis mc2155, M. bovis BCG Glaxo strain and M. tuberculosis H37Rv were grown in Sauton's medium or Middlebrook 7H9 (Becton, Dickinson and Company) medium supplemented with Albumin-Dextrose Complex (designated as supplemented 7H9 medium). M. smegmatis was also grown in Lysogeny broth (LB) with addition of 0.05% (w/v) Tween 80 to prevent bacterial aggregation. All bacterial strains were cultivated at 37°C with shaking in BD Falcon conical tubes or in conical flasks. Where needed, antimicrobials were added at the following concentrations (µg ml−1): hygromycin 50; kanamycin 50; tetracycline 0.02.
Bacterial growth was followed by measurement of absorbance at 580 nm in a Jenway spectrophotometer. Growth of M. smegmatis in various media was performed using a Bioscreen Growth Analyser as described previously [34]. Presented Bioscreen data are mean values of two independent experiments done in quintuplicate. Three independent transformants were used for all growth experiments. Viability was assayed by estimation of colony-forming unit (CFU) on 7H10 agar (Becton, Dickinson and Company). Supernatants were prepared from logarithmic phase cultures of M. tuberculosis (OD580 = 0.6–0.8) grown in Sauton's medium, filter sterilized twice and used for experiments immediately. For dose-dependency experiments culture supernatants were freeze-dried and reconstituted in sterile water. For low-magnesium control experiments concentration of MgSO4 in Sauton's medium was 0.2 mM.
2.2. Microscopy
Mycobacterium smegmatis bacilli were mounted on PBS and observed by phase contrast microscopy using a Diphot 300 inverted microscope with a 100 W mercury light source. Images were recorded using a 12/10 bit, high-speed Peltier-cooled CCD camera (FDI, Photonic Science) using Image-Pro Plus (Media Cybernetics) software. Labelling of nascent peptidoglycan was done as described previously [35]. Briefly, mycobacteria were grown in Sauton's medium in the presence of 20 ng ml−1 of tetracycline to exponential growth phase (OD600 ∼ 0.5). A mixture of Vancomycin-BODIPY (Life Technologies) and unlabelled vancomycin (1 : 1) was added to cultures to the final concentration of 2 µg ml−1, followed by a 2 h incubation at 37°C with shaking. Bacteria were washed twice with PBS containing 0.1% Tween 80 and used for fluorescence microscopy.
2.3. Generation PknB constructs for overexpression studies
The pknB gene and its variants (electronic supplementary material, table S1) were amplified from M. tuberculosis H37Rv DNA and cloned into the BamHI and SpeI sites of the pMind plasmid [36] (for details of primers see electronic supplementary material, table S2). Constructs were confirmed by sequencing and electroporated into M. smegmatis or M. tuberculosis. Overexpression of pknB or its variants was induced by the addition of tetracycline and confirmed by qRT-PCR.
2.4. Preparation of muropeptides
Peptidoglycan from Escherichia coli and mycobacteria was isolated and purified as described previously [37,38]. For growth assays peptidoglycan (2 mg) was digested with mutanolysin, lysozyme, RpfB or MltA at 37°C for up to 72 h. Muramidases were used at the final concentration of 100 μg ml−1. Peptidoglycan digestion with mutanolysin and MltA was carried out in buffer containing 80 mM NaH2PO4, pH 4.8; with lysozyme in 25 mM NaH2PO4, pH 6.0; and with RpfB in 40 mM sodium citrate, pH 6.5. The presence of muropeptides was confirmed by HPLC. In separate experiments, peptidoglycan was sonicated three times for 30 s, briefly spun down at 100g to remove big fragments and used for growth assays. Synthetic muropeptides were prepared as described previously [39].
2.5. Determination of minimum inhibitory concentration for selected antimicrobials
Tests were performed in 96-well microtitre plates using a dilution method [40]. Briefly, M. smegmatis (1 × 105 CFU) was inoculated in 100 µl of Sauton's medium supplemented with kanamycin and tetracycline, and a range of concentrations of antimicrobial tested. Plates were incubated for three days at 37°C. MIC was determined as the lowest concentration at which no visible growth was detected after 3 days of incubation. Fresh antimicrobial stocks were prepared for each experiment; four independent experiments were done.
2.6. Effect of recombinant PASTA protein on growth of Mycobacterium tuberculosis
Recombinant PASTA was purified as described previously [31] and filter sterilized prior to growth experiments. Mycobacteria (103 cells ml−1) were inoculated in 16-well microtitre plates containing supplemented 7H9 medium with different concentration of sterile rPknB_PASTA. For each concentration four replicates were inoculated. Sealed plates were incubated at 37°C without shaking. Optical density was measured after 15 and 25 days of incubation. The experiment was repeated twice. Similar results were obtained when mycobacteria were grown in Falcon tubes or conical plastic flasks. M. tuberculosis stationary phase culture was stored at 4°C for two months and 2 × 104 cells ml−1 were inoculated in conical flasks. The flasks were incubated at 37°C for up to 12 weeks. Optical density was measured at regular intervals using a Jenway spectrophotometer. The assay was performed in triplicate, three times.
2.7. Transcriptional profiling
Total RNA was isolated from 10 ml of mycobacterial cultures from exponential growth phase using the TRIzol method [41]. DNA contamination was removed with Turbo DNA-free DNAase (Ambion) before cDNA was generated using Superscript Reverse Transcriptase II (Invitrogen) and gene-specific primers (electronic supplementary material, table S2). Q-PCR was performed in a Corbett Rotor Gene 6000 real-time thermocycler using Absolute QPCR SYBR Green mix (Thermo) and gene-specific primers. Levels of expression were normalized to 16 s rRNA [42].
2.8. Protein electrophoresis and Western blot
Mycobacteria were collected by centrifugation, washed in PBS and resuspended in buffer containing 50 mM Tris–HCl, pH 8.0 and 150 mM NaCl. Bacteria were lysed in FastPrep-24 Instrument (MP Biomedicals, UK) using glass beads. Lysates were centrifuged at 14 000g for 20 min to separate soluble and insoluble fractions. Proteins were separated, using 12% SDS PAGE and transferred on nitrocellulose membrane. Primary antibodies were anti-polyhistidine antibodies (Sigma); alkaline-phosphatase conjugate anti-mouse antibodies were used as secondary antibodies. Sigma Fast BCIP/NBT was used as a substrate to visualize recognized proteins.
2.9. Isolation of membrane protein fraction for proteomics analysis
Membrane fractions were prepared as described previously [43]. Briefly, M. smegmatis TMP and MIND strains were inoculated in 2 l conical flasks containing 500 ml Sauton's medium. The cultures were incubated at 37°C with shaking (200 r.p.m.) for 3 h. Cultures from two flasks were used for preparation of membrane fractions of each strain. The bacteria were harvested by centrifugation at 6000g for 20 min and washed with PBS. The pellets were resuspended in 10 ml of extraction buffer containing 50 mM Tris–HCl, pH 8.0, 150 mM NaCl, 10 mM MgCl2 and stirred on ice for 10 min. The cell suspension was sonicated before adding 100 µg of DNase I per gram of cells and stirring on ice for 5 min to decrease viscosity. The suspension was centrifuged at 25 000g for 25 min at 4°C. The pellet was discarded and the supernatant centrifuged at 100 000g for 60 min at 4°C. After discarding supernatant the pellet was carefully resuspended in 100 mM sodium carbonate buffer, pH 11.5 and centrifuged at 100 000g for 60 min. The centrifugation and resuspension in carbonate buffer steps were repeated a minimum of three times before a final resuspension and centrifugation step using sterile MilliQ water. The membrane fractions were resuspended in 1 ml sterile deionized water and freeze-dried.
2.10. Analysis of membrane proteins
Proteomics was carried out by the University of Leicester Proteomics Facility (PNACL, University of Leicester, http://www2.le.ac.uk/colleges/medbiopsych/facilities-and-services/cbs/protein-and-dna-facility/pnacl). Trypsin digestion of membrane proteins was performed using a filter-aided sample preparation method [44]. LC–MS/MS was carried out using an RSLCnano HPLC system (Dionex, UK) and an LTQ-Orbitrap-Velos mass spectrometer (Thermo Scientific). Peptides were eluted from the trap column at a flow rate of 0.3 µl min−1 and through a reverse-phase PicoFrit capillary column (75 μm i.d. × 400 mm) containing Symmetry C18 100 Å media (Waters, UK) that was packed in-house using a high-pressure device (Proxeon Biosystems, Denmark) over a period of 4 h, with the output of the column sprayed directly into the nanospray ion source of the LTQ-Orbitrap-Velos mass spectrometer.
The raw data file obtained from each LC–MS/MS acquisition was processed using Proteome Discoverer (v. 1.4.0.288, Thermo Scientific), searching each file in turn using Mascot [45] (v. 2.2.04, Matrix Science) against the UniProtKB-Swissprot database. The peptide tolerance was set to 10 ppm and the MS/MS tolerance was set to 0.02 Da. Fixed modifications were set as carbamidomethyl (C) and variable modifications set as oxidation (M). A decoy database search was performed. The output from Proteome Discoverer was further processed using Scaffold Q + S4 (v. 4.0.5, Proteome Software, Portland, OR, USA). For quantitative experiments the digested peptides were labelled with the TMTsixplex Label Reagent kit (Life Sciences) according to the manufacturer's instructions. Scaffold Q+ (v. 4.3.4, Proteome Software) was used to quantify Label Based Quantitation (iTRAQ, TMT, SILAC) peptide and protein identifications. Both protein and peptide identifications were accepted if they could be established at greater than 95.0% probability and contained at least two identified peptides. Peptide probabilities from X! Tandem were assigned by the Peptide Prophet algorithm [46] with Scaffold delta-mass correction. Peptide probabilities from Mascot were assigned by the Scaffold Local FDR algorithm. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifiers PXD002120 and 10.6019/PXD002120.
2.11. Analytical gel-filtration
Fifty microlitres of PknB_PASTA at a concentration of 50 µM were injected on a S75 10/300 column (GE Life Science) at a 0.4 ml min−1 flow rate. The column was equilibrated with 25 mM Tris–HCl (pH 8.5), and 100 mM NaCl with or without 25 mM MgSO4. For the injection in the column in the buffer without MgSO4, the protein was pre-incubated with 5 mM EDTA. In the case of gel-filtration in the presence of MgSO4, the protein was pre-incubated with 25 mM MgSO4.
2.12. Nuclear magnetic resonance experiments
All nuclear magnetic resonance (NMR) experiments were carried out at 20°C on Bruker Avance III 700 (1H–15N double resonance experiments) or Avance III 500 (1H–13C–15N triple-resonance experiments) spectrometers equipped with 5 mm z-gradient TCI cryoprobe, using the standard pulse sequences. NMR samples consist of approximately 50 µM 15N-labelled protein dissolved in 10 mM phosphate buffer (pH 6.8), 100 mM NaCl, with 5% D2O for the lock prepared as described elsewhere [31].
3. Results
3.1. The external PASTA domain is important for PknB function
It has been previously shown that overexpression of enzymatically active PknB in mycobacteria results in growth inhibition and alteration of bacterial shape [4]. Additionally, the external PknB_PASTA domain is indispensable for functional complementation of PknB conditional mutants of M. smegmatis and M. tuberculosis [47]. However, the precise role of the PknB_PASTA domain in regulation of mycobacterial growth and peptidoglycan biosynthesis remains unclear. In our initial experiments, we investigated growth patterns of M. smegmatis overexpressing pknB.
A full-length pknB or its truncated form missing the region encoding the external PASTA domain (pknB-ΔPASTA) was cloned into pMind plasmid [36], containing a tetracycline-regulated promoter (figure 1; electronic supplementary material, table S1). The plasmids were transformed into M. smegmatis and the resultant strains, designated as PknB1 (pknB), PknB5 (pknB-ΔPASTA) and MIND (empty plasmid), grew similarly in all media tested in the absence of the inducer tetracycline (data not shown). However, addition of tetracycline to the PknB1 or PknB5 strains affected growth in liquid medium (figure 2). This inhibitory effect was probably caused by the induction of expression of pknB and pknB-ΔPASTA from the pMind plasmid. When grown in the presence of tetracycline, PknB1 and PknB5 expressed the pknB versions from pMind at relatively high level, corresponding to 1% (±0.3%) of all 16s-rRNA transcripts as judged by quantitative RT-PCR; no expression of the target gene was detected in mycobacteria in the absence of tetracycline. Interestingly, overexpressing the pknB versions resulted in different growth characteristics. In all media, the PknB1 strain had a longer apparent lag phase (time to detectable turbidity) and slower growth rate (figure 2; electronic supplementary material, table S3), indicating that overexpression of the truncated PknB missing the external PASTA domain was less toxic than overexpression of the full-length kinase.
Figure 2.
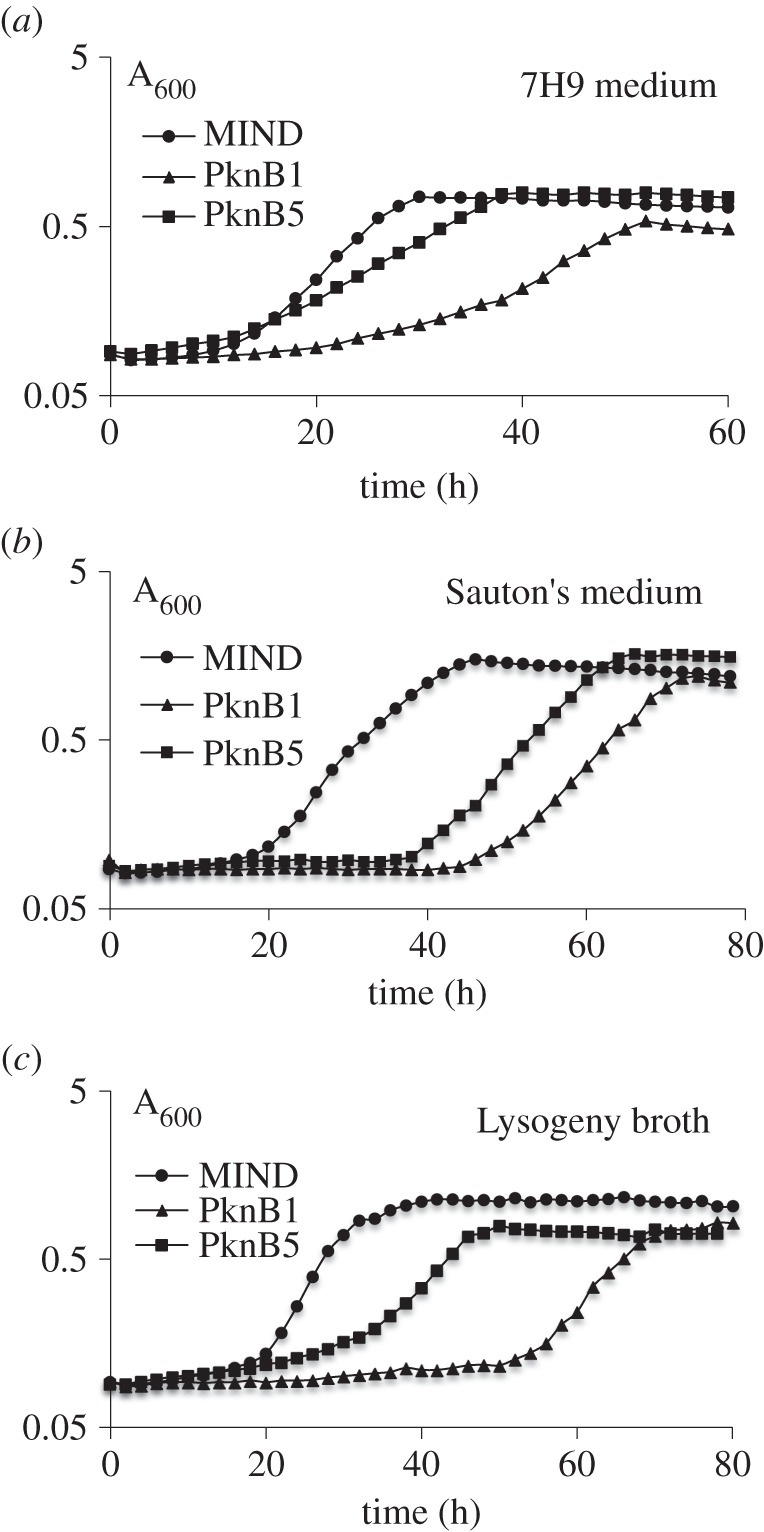
Effect of pknB and pknB-ΔPASTA overexpression on growth of M. smegmatis. Mycobacteria (5 × 106) from early stationary phase were inoculated in 100-well honeycomb plates, containing various media and incubated in Bioscreen Growth analyser at 37°C with shaking for 5 days in the presence of tetracycline. Presented data are mean values of 5 replicates from two independent experiments. Standard deviations were 10% or less of average values and are not shown for clarity.
To further investigate the role of the PknB_PASTA domain in bacterial growth directly, we cloned the pknB region encoding the membrane-anchored external domain of M. tuberculosis (designated as tmp, figure 1) into the pMind plasmid and transformed this plasmid into M. smegmatis. In the absence of tetracycline, both MIND and TMP strains grew similarly (figure 3a, filled symbols) and their cDNA analysed by q-RT-PCR contained similar levels of tmp transcripts. However, addition of tetracycline led to overexpression of the tmp transcript (reaching 9.6 ± 0.8% from the total 16 s rRNA transcripts) in the TMP strain, which resulted in growth inhibition, especially in Sauton's medium (electronic supplementary material, table S3; figure 3a, open symbols). The TMP strain required a longer time to initiate bacterial growth, but its growth rate was identical to the one observed in the MIND control strain. This phenotype was highly reproducible and did not change after passage in vitro (electronic supplementary material, figure S1). Expression of the PASTA domain without the transmembrane domain (PknB13), the transmembrane domain on its own (PknB9) or the transmembrane domain with one PASTA unit (PknB10; figure 1) had no significant effect on bacterial growth (figure 3b). Strains overexpressing the PASTA domain with two or three PASTA units (PknB11 and PknB12, respectively) displayed a marginal growth defect. All PASTA variants were expressed at similar levels according to qRT-PCR (data not shown). We generated Myc-6xHis-tagged versions of TMP (designated as TMPH strain; electronic supplementary material, table S1) and of the soluble PASTA domain (designated as PknB14) and investigated their localization by western blot. TMPH was detected in the envelope fraction, while PknB14 was found in the cytoplasm M. smegmatis (figure 3c,d). Importantly, TMPH and TMP strains showed no difference in growth under all conditions described here (data not shown). These results confirm that only the surface-exposed domain containing four PASTA units significantly impaired mycobacterial growth.
Figure 3.

Effect of PknB_PASTA overexpression on M. smegmatis growth. (a) Comparative growth kinetics of M. smegmatis strains in Sauton's medium. The same inoculum (approx. 2 × 106 cells per well) was used to seed induced (+tetracycline) and control cultures, respectively. (b) Overexpression of non-secreted PASTA domains or transmembrane domain on its own does not affect mycobacterial growth. (c) Detection of PknB_TM-PASTA-6 × HIS in membrane fraction of M. smegmatis strains by western blotting. (d) Detection of the PASTA domain missing the transmembrane region in cytoplasmic fraction of M. smegmatis strains by Western blotting.
Overproduction of the PknB_PASTA domain also influenced cellular morphology. Microscopic examination of the strains revealed accumulation of elongated TMP cells of irregular shape (figure 4b, left panel), similar to cells with partially depleted PknB [4], suggesting that the overexpressed PASTA domain possibly interfered with the PknB function. Furthermore, Vancomycin-BODIPY labelling revealed anomalous distribution of nascent peptidoglycan in TMP mycobacteria (figure 4b, middle panel). While in control MIND cells newly synthesized peptidoglycan was mainly localized at the poles and mid-cell (figure 4a), TMP cells displayed a diffused staining across the entire cell surface (figure 4b). We also observed significant proportions of unlabelled or weakly labelled TMP cells (figure 4b). TMP mycobacteria tend to aggregate, suggesting a modified cell surface, delayed cell separation or increased lysis.
Figure 4.
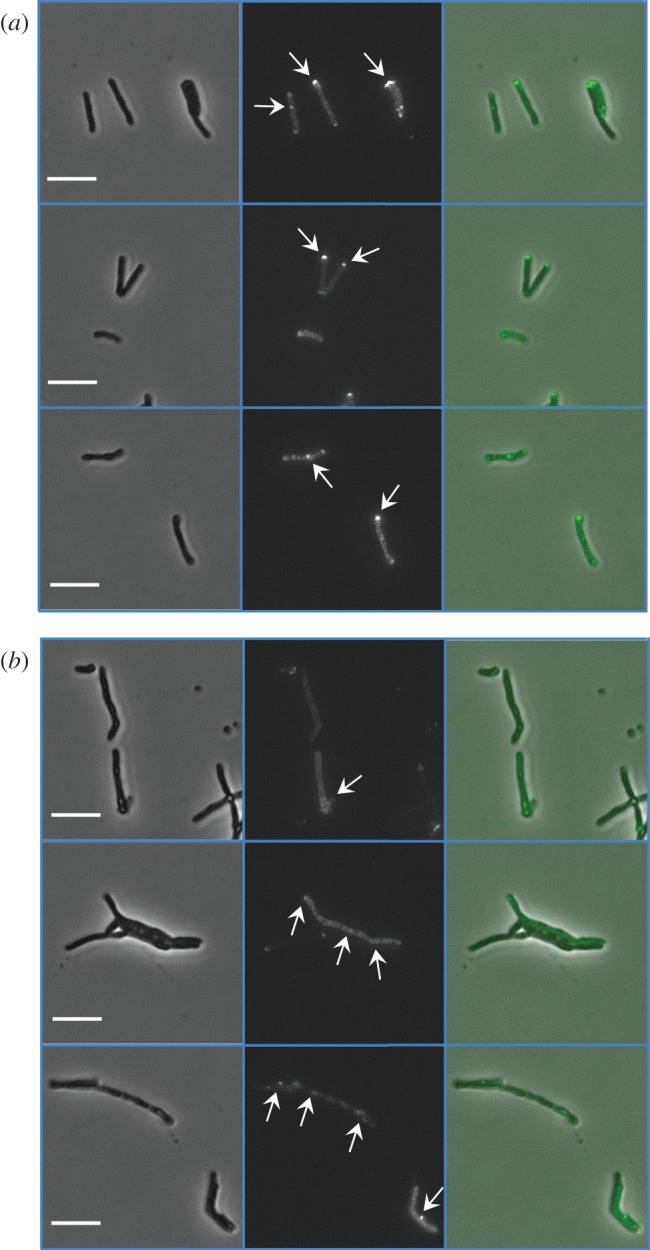
Overexpression of PknB_PASTA results in alteration of bacterial morphology and vancomycin labelling. Mycobacteria from logarithmic growth phase were labelled with Vancomycin-BODIPY conjugate, washed and mounted in PBS and examined using a Diphot 300 inverted microscope. Three independent fields for each strain are shown. Bars indicate 5 μm. (a) MIND and (b) TMP. Left panel, phase contrast; middle panel, fluorescence microscopy; right panel, merged images. Arrows indicate labelled peptidoglycan.
3.2. Elevated Mg2+ ion level eliminates TMP-mediated growth inhibition
The results described above support the functional importance of the PknB_PASTA domain for PknB activity. The structural studies of the PknB_PASTA suggested that this domain could dimerize beacuse of conformational flexibility between the different PASTA domains [31]. It was proposed that the binding of putative ligands could induce the dimerization of kinase domains and consequent trans-activation of PknB [31]. Thus, the PASTA domain could serve as a receptor for signalling molecules (e.g. muropeptides) and activate bacterial resuscitation and growth, employing the mechanism previously proposed for B. subtilis spore germination [16,39]. The PknB_PASTA domain missing the functional kinase domain may compete with the native PknB for the ligands and dysregulate the PknB-mediated signalling pathways. Therefore, we investigated whether culture supernatant from growing mycobacteria (potentially containing the putative ligand) would eliminate the growth defect caused by TMP overexpression. Indeed, the growth inhibition was abolished by the addition of culture supernatant in a dose-dependent manner (figure 5a). Further tests showed that the active compound was a low-molecular-weight chemical resistant to heating and it was present in Sauton's medium prior to exposure to bacteria (data not shown). Eventually, we demonstrated that the active entity was Mg2+ from MgSO4 or MgCl2. Elevated concentration of Mg2+ could completely abolish the inhibition by PASTA-domain overexpression (figure 5b) and the dose-dependent effect of conditioned medium was largely because of increased Mg2+ concentrations (data not shown). Supernatants obtained from cultures grown in lower-MgSO4 (0.2 mM) medium had only a moderate growth-stimulatory effect on both control and TMP strains (figure 5c). The increased Mg2+ concentration did not influence expression of tmp (p > 0.05, t-test) and the production of TMPH, a His-tagged version of TM-PASTA (data not shown). Magnesium also improved growth of PknB1 and PknB5 strains (data not shown).
Figure 5.

Culture supernatant and high concentration of Mg2+ abolish the inhibitory effect caused by PknB-PASTA overexpression. (a) Culture supernatants abolish growth-inhibitory effect of PknB_TM-PASTA overexpression in a dose-dependent manner. Relative concentration of culture supernatant added is expressed as ‘fold concentration’. A value of onefold corresponds to undiluted culture supernatant. The apparent lag phase was calculated as a period of time when culture reached OD600 nm 0.1. (b) Mg2+ (10 mM) relieves growth inhibition of TMP in a manner comparable with undiluted culture supernatant. (c) Effect of low Mg2+ supernatant on apparent lag phase of TMP strain. (d) Effect of synthetic muropeptides on growth of M. smegmatis. Mycobacteria from early stationary phase were washed in Sauton's medium twice and 5 × 106 bacteria were inoculated in Sauton's medium, containing culture supernatant or synthetic muropeptides at final concentration of 10 µM. MPP, MurNAc-pentapeptide; DPP, GlcNAc-MurNAc-peptapeptide; ADPP, GlcNAc-1,6 anhydromuramyl-pentapeptide; TPP, tetra-saccharide-peptide; SN, 50% (v/v) culture supernatant. GlcNAc, N-acetylglucosamine; MurNAc, N-acetylmuramic acid; pentapeptide, l-Ala-γ-d-Glu-m-DAP-d-Ala-d-Ala. (b,d) Presented data are mean values of 5 replicates from two independent experiments. Standard deviations were 10% or less of average values and are not shown for clarity. SN, media were supplemented with culture supernatant.
To investigate whether the PknB_PASTA domain was able to bind Mg2+ ions, we used NMR methodology [31]. A 15N-labelled sample of recombinant PknB_PASTA (50 µM) was titrated with increasing amounts of MgCl2 (or MgSO4) up to 50 mM. The ion binding with either a lateral chain or the protein backbone would generate chemical-shift perturbation on the 1H–15N resonance measured in the HSQC experiment [48]. However, no chemical-shift perturbation was observed in the PknB_PASTA upon addition of MgCl2 or EDTA, suggesting a lack of interaction between the protein and Mg2+ ions (electronic supplementary material, figure S2), and the addition of MgSO4 to recombinant PASTA did not change the oligomerization state of recombinant PASTA (electronic supplementary material, figure S3). These results show that the ameliorating effect of Mg2+ on TMP-mediated growth inhibition was indirect.
3.3. Muropeptides do not eliminate TMP-mediated growth inhibition
In separate experiments, we tested the effects of muropeptides on the growth inhibition in the TMP strain. We used peptidoglycan from E. coli digested with various muramidases, sonicated or RpfB-digested peptidoglycan from M. smegmatis, and synthetic muropeptides, N-acetylglucosamine, N-acetylmuramic acid and N-acetylglucosaminyl–N-acetylmuramic acid (electronic supplementary material, table S4). We only observed a minor reduction in the duration of lag phase of TMP strain from 65 to 52 h in the presence of relatively high concentrations of digested or sonicated peptidoglycan (0.5 mg ml−1 and higher). The observed reduction in lag phase did not depend on the type of peptidoglycan used. Synthetic muropeptides, previously employed for germination of B. subtilis spores [39], did not reduce growth inhibition at any concentration tested (10 pM–50 mM) and did not stimulate growth of M. smegmatis (figure 5d). Among other divalent metals tested Ca2+ improved the TMP growth; however, it could not be used at concentrations above 5 mM because of poor solubility. Since Mg2+ ions are known to stabilize the cell wall and are frequently used for cultivation of bacteria with defective cell walls [49], we investigated whether the TMP strain was more susceptible to various antimicrobials compared with control.
3.4. Overexpression of PknB-PASTA increases sensitivity of Mycobacterium smegmatis to β-lactam antibiotics and alters the abundance of cell-wall proteins
We tested minimum inhibitory concentrations (MIC) of various antimicrobial agents for PknB1, TMP and MIND strains. MIC of rifampicin (inhibition of transcription), ethambutol (arabinogalactan biosynthesis) and streptomycin (protein biosynthesis) were identical in all the strains tested (table 1). However, the TMP strain was remarkably more sensitive to several β-lactam antibiotics: meropenem, ampicillin and clavulanate, an inhibitor of β-lactamase. In the presence of sub-inhibitory concentration of clavulanate (100 μM), MIC of meropenem for the TMP strain decreased below 0.15 μM compared with 6.5 μM in the control strain. Interestingly, overexpression of PknB did not alter antimicrobial susceptibility, suggesting that the PknB-PASTA domain on its own may perturb peptidoglycan synthesis, causing higher sensitivity to β-lactams. The perturbation of peptidoglycan synthesis could also explain the prolonged lag phase observed in the TMP strain. To address this possibility, membrane proteins isolated from the MIND and TMP strains in lag phase were investigated.
Table 1.
Effect of PknB-PASTA overexpression on antimicrobial susceptibility of M. smegmatis.
| strain | minimum inhibitory concentration against antibiotic (µM) |
||||||
|---|---|---|---|---|---|---|---|
| rifampicin | streptomycin | ethambutol | meropenem | clavulanate | meropenem+clavulanatea | ampicillin | |
| MIND | 1.2 | 0.4 | 2.5 | 13–19.5 | >1000 | 6.5–13.0 | 140–170 |
| PknB1 | 1.2 | 0.4 | 2.5 | 13–19.5 | >1000 | 6.5–13.0 | 140–170 |
| TMP | 1.2 | 0.4 | 2.5 | 6.5 | 150–200 | 0.13 | 30–45 |
aClavulanate was added at a concentration of 100 µM.
In total, we identified 1071 proteins in the membrane fractions of both strains using LTQ-Orbitrap-Velos mass spectrometry. The TMPH strain had 90 proteins specific for this sample and 95 proteins were detected in the MIND strain only. The majority of proteins (886) were identified in both strains. Importantly, production of the PknB_PASTA domain in the membrane fraction was confirmed by detection of 22 unique fragments with 86% peptide coverage in the TMPH strain (electronic supplementary material, table S5). Most of the proteins detected (705) were assigned as hypothetical with unknown function, while 267 were predicted as membrane or cell-surface proteins. This result confirms that the extraction procedure selectively enriched the membrane proteins. A high number of membrane proteins implicated in transport of metals, nutrients and enzymes was found in the membrane fractions of both strains. Next, peptides were labelled using the TMPsixplex kit to compare the relative abundance of membrane proteins in TMP and MIND (electronic supplementary material, table S5). We focused our analysis on cell division proteins and enzymes involved in cell-wall biosynthesis. Most cell division proteins were equally present in the TMP and MIND strains (data not shown). However, CwsA was more abundant in the TMP strain (table 2). The precise function of CwsA is unknown but it has been shown to interact with CrgA, which is annotated as an inhibitor of septum formation [50,51]. CrgA was detected only in the membrane fractions of TMP but not in those of MIND, and its relative abundance could not be calculated. Another CrgA interaction partner, Wag31, was detected at similar levels in both strains. Interestingly, the TMP strain had more enzymes involved in biosynthesis of mycolyl-arabinogalactan (table 2; electronic supplementary material, table S5), including several mycolyl- and galactofuranosyl-transferases. Regarding peptidoglycan-related enzymes, the levels of two metallo-β-lactamases, l,d-transpeptidase LdtB and d-alanyl-d-alanine carboxypeptidase DacB1, were increased in the TMP membrane fraction (table 2). Complete data are available via ProteomeXchange with identifier PXD002120. Collectively these results may suggest that the overexpression of the PASTA domain influences the biosynthesis and architecture of mycobacterial cell wall.
Table 2.
Cell-wall enzymes and proteins differently abundant in TMP compared with MIND.
| M. tuberculosis protein | M. smegmatis protein | protein description | ratio TMP/MIND log2-fold |
|---|---|---|---|
| Rv0014c | MSMEG_0028 | His-Tagged TM-PASTA | 5.9 |
| Rv0008c | MSMEG_0023 | cell-wall protein CwsA | 1.0 |
| Rv3804c | MSMEG_6398 | FbpA mycolyl transferase | 1.0 |
| Rv3790 | MSMEG_6382 | DprE1 decaprenylphosphoryl-β-d-ribose 2′oxidase | 0.9 |
| Rv3577 | MSMEG_6071 | metallo-β-lactamase | 0.9 |
| Rv0129c | MSMEG_3580 | FbpB mycolyl transferase | 0.6 |
| Rv3782 | MSMEG_6367 | galactofuranosyl transferase | 0.8 |
| Rv3808c | MSMEG_6403 | galactofuranosyl transferase | 0.8 |
| Rv3265 | MSMEG_1826 | dTDP-RhA:a-d-GlcNAc-diphosphoryl polyprenol, a-3-l-rhamnosyl transferase | 0.7 |
| Rv0906 | MSMEG_5638 | metallo-β-lactamase | 0.6 |
| Rv2518c | MSMEG_4745 | LdtB l,d-transpeptidase | 0.6 |
| Rv0237 | MSMEG_0361 | glycosyl hydrolase | 0.6 |
| Rv3330 | MSMEG_1661 | DacB1D-alanyl-d-alanine carboxypeptidase | 0.5 |
| Rv2748c | MSMEG_2690 | DNA translocase FtsK | 0.5 |
| Rv2171 | MSMEG_4239 | conserved lipoprotein | −0.8 |
| Rv2721c | MSMEG_2739 | transmembrane alanine and glycine-rich protein | −0.5 |
3.5. The addition of recombinant PASTA protein abolishes growth of Mycobacterium tuberculosis
The results described above suggest that overexpression of the PknB_PASTA domain can be employed for growth inhibition and increase of susceptibility to antimicrobials in medically important pathogen M. tuberculosis. We therefore investigated whether the PknB_PASTA domain would have a similar effect in M. tuberculosis. We transformed pMind pknB7 and empty pMind plasmids (electronic supplementary material, table S1) into M. tuberculosis H37Rv and investigated growth of the resultant strains (TMP and MIND) in the presence of tetracycline. As shown in figure 6a, overexpression of the external membrane attached PASTA (TMP) indeed inhibited initiation of M. tuberculosis growth. We next studied if addition of recombinant PASTA protein containing PASTA units only (residues 354–626, designated as rPASTA) may alter the growth. We noted that the addition of rPASTA to M. tuberculosis resulted in growth defect at concentration of 10 µg ml−1 (figure 6b) and increased apparent lag phase from 5 to 14 days. This growth inhibition was temporary and the cultures eventually produced normal growth, while addition of recombinant PASTA to logarithmic phase culture had no effect on growth (data not shown). However, the growth-inhibitory effect of rPASTA was more pronounced in experiments when mycobacteria were stored at 4°C for two months (figure 6c). We found that these cells retained culturability and produced normal growth both in liquid and solid media. Nevertheless, their growth initiation was significantly delayed by addition of rPASTA at concentration 10 µg ml−1. The mycobacteria were not killed by rPASTA and after two-month incubation generated normal stationary phase culture (figure 6c). In all experiments, control buffer did not have any inhibitory effect. These results suggest that the PASTA domain may directly interfere with mycobacterial regrowth and, therefore, presents a plausible target for the design of specific drugs altering bacterial regrowth and resuscitation.
Figure 6.

PknB_PASTA reduces growth of Mycobacterium tuberculosis. (a) Overexpression of PknB_PASTA domain delays growth initiation. Mycobacteria (approx. 1 × 106) were inoculated in supplemented 7H9 medium containing hygromycin and tetrocycline. (b) Effect of different concentrations of rPASTA on M. tuberculosis growth. Mycobacteria (103 cells ml−1) were inoculated in supplemented 7H9 medium containing different concentration of sterile rPASTA. The optical density was measured after 21 days of incubation. (c) Regrowth of stored M. tuberculosis was delayed by the addition of rPASTA. M. tuberculosis (2 × 104 cells ml−1) was inoculated in supplemented 7H9 medium. Buffer was used as control and rPASTA was added at final concentration of 10 µg ml−1. (b,c) Each data point is an average value of 3 biological replicates, error bars indicate standard deviations. Experiments were performed three times and results of one typical experiment are shown.
4. Discussion and conclusion
PknB has been shown to regulate bacterial division, growth and cell-wall biosynthesis [10]. The data presented in this study suggest that the PknB-mediated signalling pathway can be targeted through the external PknB_PASTA domain. Overexpression of the PknB_PASTA domain delayed the regrowth of both M. tuberculosis and M. smegmatis. The growth inhibition was ameliorated by addition of high concentrations of Mg2+ but not by muropeptides. Mg2+ ions are crucial for all living organisms, and the concentration may vary, reaching as high as 20 mM in eukaryotic [52] and a massive 100 mM in prokaryotic cells [53]. In Gram-negative bacteria magnesium ions stabilize the outer membrane [54] and are found in high concentrations in the cell wall of Gram-positive bacteria, which bind divalent ions to their teichoic acids [54]. Mycobacteria do not possess teichoic acids and the binding of Mg+2 ions has not been studied in the mycobacterial cell wall in detail, although Mg2+ ions improved growth of mycobacteria in media with low pH [55]. Growth of B. subtilis mutants missing certain peptidoglycan binding proteins was improved at high Mg2+ levels, suggesting that divalent metals may compensate growth defects due to altered peptidoglycan biosynthesis [56].
The precise mechanism of the magnesium effect on growth of the TMP strain is unknown. Mg2+ ions may stabilize the cellular membrane and protein complexes associated with the membrane disturbed by overexpression of PknB_PASTA domain, although it is also possible that Mg2+ ions directly bind to the growing strands of stems of peptidoglycan and promotes their polymerization. Further dissection of molecular mechanisms underlying the inhibitory effect of PknB_PASTA overexpression will facilitate our understanding of the biological function of PknB and its extracellular domain, as well as the roles of Mg2+ ions in cell-wall architecture. Three possible scenarios may explain the observed effects associated with overexpression of PknB_PASTA (figure 7). (i) The overexpressed PknB_PASTA competes for the ligand (peptidoglycan) with a native PknB, and therefore interferes with the PknB-mediated signalling pathway (figure 7a). (ii) The PknB_PASTA domain interacts with penicillin-binding proteins and dysregulates peptidoglycan biosynthesis (figure 7b). (iii) Finally, highly abundant PknB_PASTA may directly disrupt the structure of peptidoglycan by binding to the growing strand, and physically prevent extension and cross-linking (figure 7c).
Figure 7.
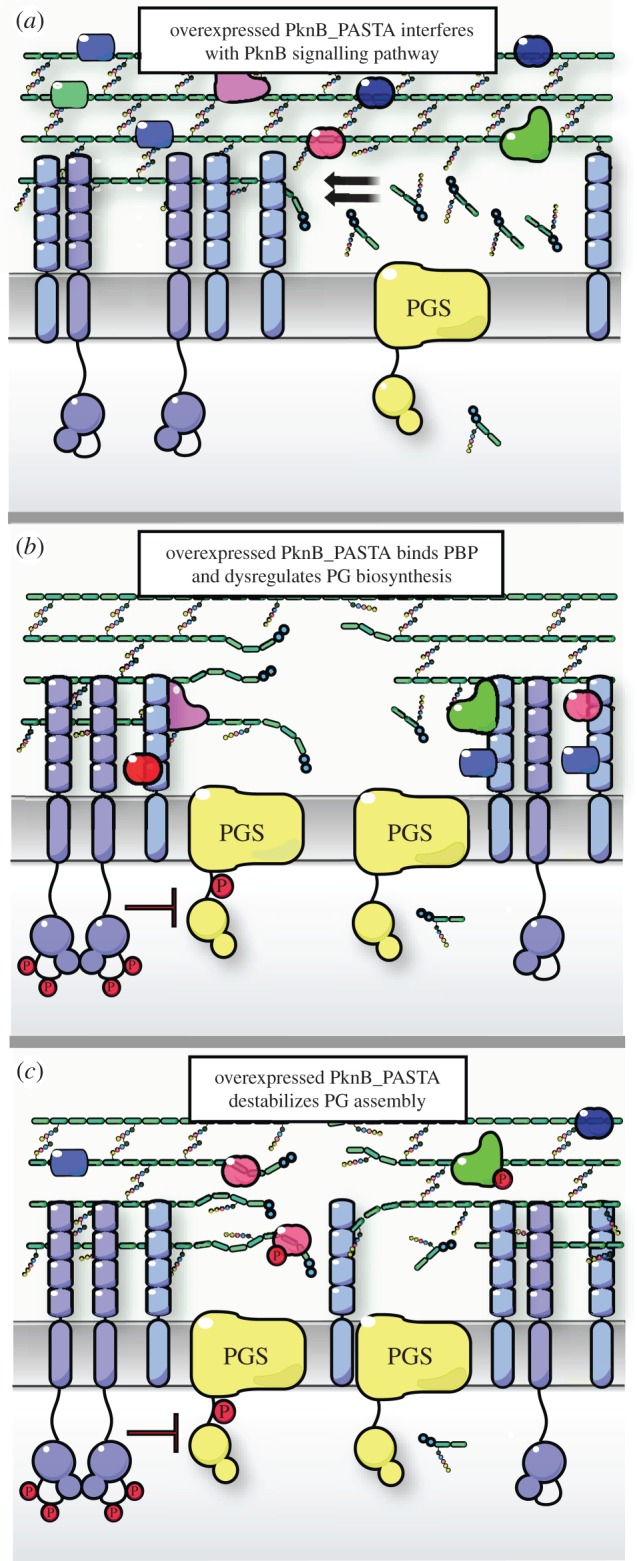
Schematic representation of possible mechanisms of PknB_PASTA-mediated growth inhibition. (a) In the TMP strain the overexpressed PknB_PASTA competes with native PknB for peptidoglycan ligands and interferes with PknB-mediated signalling. (b) Overexpressed PknB_PASTA binds penicillin-binding proteins and dysregulates peptidoglycan polymerization. (c) Overexpressed PknB_PASTA binds to peptidoglycan and destabilizes its structure. PGS, peptidoglycan synthesis complex.
The regulatory function of PknB (phosphorylation of cell-wall enzymes and division factors) is evolved to coordinate cell-wall biosynthesis and division. In fact, a recently published study established that PknB coordinated assembly of peptidoglycan synthesizing complex via recruiting FhaA and phosphorylating MviN, a protein essential for mycobacterial growth [26].
In Listeria monocytogenes the inactivation of a PASTA-containing kinase, a homologue of PknB, resulted in increased susceptibility to β-lactam antimicrobials [57], as in the TMP strain, which presumably shows high susceptibility to β-lactam antimicrobials because of partial inactivation of PknB function. However, while the PknB mutant lyses and is not able to grow, the TMP strain only shows an initial impairment of growth in the form of a prolonged lag phase, but is able to grow normally in exponential phase. Thus, other factors apart from interference with PknB function may contribute to the growth phenotype of the TMP strain.
A possible direct interaction of PknB_PASTA with penicillin-binding proteins is in accordance with recent findings that the PASTA domain of StkP kinase from S. pneumoniae was able to bind penicillin-binding protein 2× [58]. We also cannot exclude that the PASTA domain changes the architecture of the cell wall by interacting directly with peptidoglycan (figure 7c), as was previously observed in vitro [16,59]. The results presented in this study may suggest that the overexpressed PknB_PASTA domain interacts with growing strands of peptidoglycan and possibly physically interferes with its cross linking (figure 7c), culminating in higher susceptibility of the overexpressing strain to meropenem, clavulanate and ampicillin. Meropenem has been shown to inhibit both dd-carboxypeptidase and ld-transpeptidase activities [60], and in combination with clavulanate it is highly active against growing and persisting mycobacteria [61].
Our findings are consistent with the previously proposed peptidoglycan-sensing role of PknB_PASTA [11,30]. Further investigation is required to clarify whether the PknB_PASTA domain recognizes the specific structures present in the region of peptidoglycan growth and therefore activates dimerization of the catalytic domains, or whether it plays a more structural role in supporting growing peptidoglycan and ensuring proper localization of PknB. Strains overexpressing truncated forms of the PASTA domain did not have pronounced growth defect (figure 3b), suggesting that four PASTA units and the transmembrane region are required for PknB functionality. Removal of the transmembrane part resulted in mislocalization of the PASTA domain. Although the precise role of the individual PASTA units awaits elucidation, our data are consistent with the previous observation that the truncated PknB missing one or more PASTA domains was not able to complement the conditional pknB mutant [47]. It is also remains unclear whether the PASTA domain can sense short muropeptides released by mycobacteria during growth. In our experiments, we obtained a very modest effect of peptidoglycan on the growth inhibition caused by PASTA overproduction and no stimulation of growth or resuscitation by externally added muropeptides. It is possible that specific muropeptides are required for activation of PknB and subsequent resuscitation, as muropeptides have been shown to possess resuscitation-stimulatory activity [62]. It is conceivable that the PASTA domain may be directly involved in resuscitation by sensing in vivo rearrangement of peptidoglycan, possibly because of the action of resuscitation-promoting factors [34,63] and other muramidases, and regulation of cell-wall biosynthesis. Additional experiments on monitoring rearrangement of peptidoglycan and interaction between the external PASTA domain and stem peptidoglycan, using solid-state NMR [64] and probes for microscopy of living bacteria such as highly sensitive atomic force microscopy cantilevers [65] will allow us to establish the precise PknB-mediated sensing mechanism.
Supplementary Material
Supplementary Material
Supplementary Material
Acknowledgements
We are grateful to Roger Buxton for critical reading of this manuscript and to Brian Robertson for the pMind plasmid. Mtb DNA was provided by Colorado State University (Contract No.HHSN266200400091C; NIH, NIAID N01-AI-40091). We acknowledge the Centre for Core Biotechnology Services at the University of Leicester for support with Containment Level 3 experiments and the University of Leicester Proteomics Facility for analysis of membrane proteins.
Data accessibility
The datasets supporting this article have been uploaded as part of the electronic supplementary material.
Authors' contributions
O.T. conceived of the study, carried out molecular work and growth experiments, analysed data and drafted the manuscript; J.L. carried out molecular work, prepared peptidoglycan and muropeptides; C.H.J. carried out molecular work and prepared samples for proteomics study; P.B. carried out protein purification and NMR experiments; D.M. performed and analysed growth experiments; F.F. and D.G. contributed to molecular work and helped draft the manuscript; D.H. and M.L. synthesized and purified muropeptides; A.B. carried out proteomics experiments and analysed proteomics data; W.V. coordinated peptidoglycan and muropeptide experiments, provided muropeptides and helped draft the manuscript; S.M. coordinated synthetic muropeptide experiments, provided synthetic muropeptides and helped draft the manuscript; M.C.-G. conceived of the study, designed the study and coordinated protein work; G.V.M. conceived of the study, designed the study, analysed data, coordinated the study and drafted the manuscript.
Competing interests
The authors declare that they have no conflict of interest.
Funding
This study was supported by BBSRC grant BB/H008586/1 (O.T. and G.V.M.), MRC-DTG (J.L.), the National Institutes of Health grant AI090348 (D.H., M.L. and S.M.) and the EC DIVINOCELL project (W.V.).
References
- 1.Pereira SFF, Goss L, Dworkin J. 2011. Eukaryote-like serine/threonine kinases and phosphatases in bacteria. Microbiol. Mol. Biol. R 75, 192–212. (doi:10.1128/MMBR.00042-10) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chao J, Wong D, Zheng X, Poirier V, Bach H, Hmama Z, Av-Gay Y. 2010. Protein kinase and phosphatase signaling in Mycobacterium tuberculosis physiology and pathogenesis. Biochim. Biophys. Acta 1804, 620–627. (doi:10.1016/j.bbapap.2009.09.008) [DOI] [PubMed] [Google Scholar]
- 3.Fernandez P, Saint-Joanis B, Barilone N, Jackson M, Gicquel B, Cole ST, Alzari PM. 2006. The Ser/Thr protein kinase PknB is essential for sustaining mycobacterial growth. J. Bacteriol. 188, 7778–7784. (doi:10.1128/JB.00963-06) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kang CM, Abbott DW, Park ST, Dascher CC, Cantley LC, Husson RN. 2005. The Mycobacterium tuberculosis serine/threonine kinases PknA and PknB: substrate identification and regulation of cell shape. Genes Dev. 19, 1692–1704. (doi:10.1101/gad.1311105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Forti F, Crosta A, Ghisotti D. 2009. Pristinamycin-inducible gene regulation in mycobacteria. J. Biotechnol. 140, 270–277. (doi:10.1016/j.jbiotec.2009.02.001) [DOI] [PubMed] [Google Scholar]
- 6.Kumar D, Narayanan S. 2012. PknE, a serine/threonine kinase of Mycobacterium tuberculosis modulates multiple apoptotic paradigms. Infect. Genet. Evol. 12, 737–747. (doi:10.1016/j.meegid.2011.09.008) [DOI] [PubMed] [Google Scholar]
- 7.Walburger A. et al. 2004. Protein kinase G from pathogenic mycobacteria promotes survival within macrophages. Science 304, 1800–1804. (doi:10.1126/science.1099384) [DOI] [PubMed] [Google Scholar]
- 8.Cowley S, et al. 2004. The Mycobacterium tuberculosis protein serine/threonine kinase PknG is linked to cellular glutamate/glutamine levels and is important for growth in vivo. Mol. Microbiol. 52, 1691–1702. (doi:10.1111/j.1365-2958.2004.04085.x) [DOI] [PubMed] [Google Scholar]
- 9.Gómez-Velasco A, Bach H, Rana AK, Cox LR, Bhatt A, Besra GS, Av-Gay Y. 2013. Disruption of the serine/threonine protein kinase H affects phthiocerol dimycocerosates synthesis in Mycobacterium tuberculosis. Microbiology 159, 726–736. (doi:10.1099/mic.0.062067-0) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Molle V, Kremer L. 2010. Division and cell envelope regulation by Ser/Thr phosphorylation: mycobacterium shows the way. Mol. Microbiol. 75, 1064–1077. (doi:10.1111/j.1365-2958.2009.07041) [DOI] [PubMed] [Google Scholar]
- 11.Jones G, Dyson P. 2006. Evolution of transmembrane protein kinases implicated in coordinating remodeling of gram-positive peptidoglycan: inside versus outside. J. Bacteriol. 188, 7470–7476. (doi:10.1128/JB.00800-06) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Debarbouille M, Dramsi S, Dusserget O, Nahori MA, Vaganay E, Jouvion G, Gazzone A, Msadek T, Duclos B. 2009. Characterization of a serine/threonine kinase involved in virulence of Staphylococcus aureus. J. Bacteriol. 191, 4070–4081. (doi:10.1128/JB.01813-08) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Banu LD, Conrads G, Rehrauer H, Hussain H, Allan E, van der Ploeg JR. 2010. The Streptococcus mutans serine/threonine kinase, PknB, regulates competence development, bacteriocin production, and cell wall metabolism. Infect. Immun. 78, 2209–2220. (doi:10.1128/IAI.01167-09) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Beilharz K, Nováková L, Fadda D, Branny P, Massidda O, Veening JW. 2012. Control of cell division in Streptococcus pneumoniae by the conserved Ser/Thr protein kinase StkP. Proc. Natl Acad. Sci. USA 109, E905–E913. (doi:10.1073/pnas.1119172109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gaidenko TM, Kim TJ, Price CW. 2002. The PrpC serine-threonine phosphatase and PrkC kinase have opposing physiological roles in stationary-phase Bacillus subtilis cells. J. Bacteriol. 184, 6109–6114. (doi:10.1128/JB.184.22.6109-6114.2002) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shah IM, Laaberki MH, Popham DL, Dworkin J. 2008. A eukaryotic-like Ser/Thr kinase signals bacteria to exit dormancy in response to peptidoglycan fragments. Cell 135, 486–496. (doi:10.1016/j.cell.2008.08.039) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shah IM, Dworkin J. 2010. Induction and regulation of a secreted peptidoglycan hydrolase by a membrane Ser/Thr kinase that detects muropeptides. Mol. Microbiol. 75, 1232–1243. (doi:10.1111/j.1365-2958.2010.07046.x) [DOI] [PubMed] [Google Scholar]
- 18.Jones G, Del Sol R, Dudley E, Dyson P. 2011. Forkhead-associated proteins genetically linked to the serine/threonine kinase PknB regulate carbon flux towards antibiotic biosynthesis in Streptomyces coelicolor. Microbial. Biotechnol. 4, 263–274. (doi:10.1111/j.1751-7915.2010.00237.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schultz C, Niebisch A, Schwaiger A, Viets U, Metzger S, Bramkamp M, Bott M. 2009. Genetic and biochemical analysis of the serine/threonine protein kinases PknA, PknB, PknG and Pknl of Corynebacterium glutamicum: evidence for non-essentiality and for phosphorylation of OdhI and FtsZ by multiple kinases. Mol. Microbiol. 74, 724–741. (doi:10.1111/j.1365-2958.2009.06897.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Molle V, Gulten G, Vilchèze C, Veyron-Churlet R, Zanella-Cléon I, Sacchettini JC, Jacobs WR, Jr, Kremer L. 2010. Phosphorylation of InhA inhibits mycolic acid biosynthesis and growth of Mycobacterium tuberculosis. Mol. Microbiol. 78, 1591–1605. (doi:10.1111/j.1365-2958.2010.07446.x) [DOI] [PubMed] [Google Scholar]
- 21.Dasgupta A, Datta P, Kundu M, Basu J. 2006. The serine/threonine kinase PknB of Mycobacterium tuberculosis phosphorylates PBPA, a penicillin-binding protein required for cell division. Microbiology 152, 493–504. (doi:10.1099/mic.0.28630-0) [DOI] [PubMed] [Google Scholar]
- 22.Veyron-Churlet R, Zanella-Cleon I, Cohen-Gonsaud M, Molle V, Kremer L. 2010. Phosphorylation of the Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein reductase MabA regulates mycolic acid biosynthesis. J. Biol. Chem. 285, 12 714–12 725. (doi:10.1074/jbc.M110.105189) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Park ST, Kang CM, Husson RN. 2008. Regulation of the SigH stress response regulon by an essential protein kinase in Mycobacterium tuberculosis. Proc. Natl Acad. Sci. USA 105, 13 105–13 110. (doi:10.1073/pnas.0801143105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ventura M, et al. 2013. GarA is an essential regulator of metabolism in Mycobacterium tuberculosis. Mol. Microbiol. 90, 356–366. (doi:10.1111/mmi.12368) [DOI] [PubMed] [Google Scholar]
- 25.Roumestand C, Leiba J, Galophe N, Margeat E, Padilla A, Bessin Y, Barthe P, Molle V, Cohen-Gonsaud M. 2011. Structural insight into the Mycobacterium tuberculosis Rv0020c protein and its interaction with the PknB kinase. Structure 19, 1525–1534. (doi:10.1016/j.str.2011.07.011) [DOI] [PubMed] [Google Scholar]
- 26.Gee CL, et al. 2012. A phosphorylated pseudokinase complex controls cell wall synthesis in mycobacteria. Sci. Signal. 5, ra7 (doi:10.1126/scisignal.2002525) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ortega C, Liao R, Anderson LN, Rustad T, Ollodart AR, Wright AT, Sherman DR, Grundner C. 2014. Mycobacterium tuberculosis Ser/Thr protein kinase B mediates an oxygen-dependent replication switch. PLoS Biol. 12, e1001746 (doi:10.1371/journal.pbio.1001746) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lougheed KE, et al. 2011. Effective inhibitors of the essential kinase PknB and their potential as anti-mycobacterial agents. Tuberculosis 91, 277–286. (doi:10.1016/j.tube.2011.03.005) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mieczkowski C, Iavarone AT, Alber T. 2008. Auto-activation mechanism of the Mycobacterium tuberculosis PknB receptor Ser/Thr kinase. EMBO J. 27, 3186–3197. (doi:10.1038/emboj.2008.236) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yeats C, Finn RD, Bateman A. 2002. The PASTA domain: a β-lactam-binding domain. Trends Biochem. Sci. 27, 438–440. (doi:10.1016/S0968-0004(02)02164-3) [DOI] [PubMed] [Google Scholar]
- 31.Barthe P, Mukamolova GV, Roumestand C, Cohen-Gonsaud M. 2010. The structure of PknB extracellular PASTA domain from Mycobacterium tuberculosis suggests a ligand-dependent kinase activation. Structure 18, 606–615. (doi:10.1016/j.str.2010.02.013) [DOI] [PubMed] [Google Scholar]
- 32.Squeglia F, et al. 2011. Chemical basis of peptidoglycan discrimination by PrkC, a key kinase involved in bacterial resuscitation from dormancy. J. Am. Chem. Soc. 133, 20 676–20 679. (doi:10.1021/ja208080r) [DOI] [PubMed] [Google Scholar]
- 33.Mir M, Asong J, Li X, Cardot J, Boons GJ, Husson RN. 2011. The extracytoplasmic domain of the Mycobacterium tuberculosis Ser/Thr kinase PknB binds specific muropeptides and is required for PknB localization. PLoS Pathog. 7, e1002182 (doi:10.1371/journal.ppat.1002182) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mukamolova GV, Turapov OA, Young DI, Kaprelyants AS, Kell DB, Young M. 2002. A family of autocrine growth factors in Mycobacterium tuberculosis. Mol. Microbiol. 46, 623–635. (doi:10.1046/j.1365-2958.2002.03184.x35) [DOI] [PubMed] [Google Scholar]
- 35.Joyce G, Williams KJ, Robb M, Noens E, Tizzano B, Shahrezaei V, Robertson BD. 2012. Cell division site placement and asymmetric growth in mycobacteria. PLoS ONE 7, e44582 (doi:10.1371/journal.pone.0044582) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Blokpoel MC, Murphy HN, O'Toole R, Wiles S, Runn ES, Stewart GR, Young DB, Robertson BD. 2005. Tetracycline-inducible gene regulation in mycobacteria. Nucleic Acids Res. 33, e22 (doi:10.1093/nar/gni023) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Glauner B, Holtje JV, Schwarz U. 1988. The composition of the murein of Escherichia coli. J. Biol. Chem. 263, 10 088–10 095. [PubMed] [Google Scholar]
- 38.Mahapatra S, Crick DC, McNeil MR, Brennan PJ. 2008. Unique structural features of the peptidoglycan of Mycobacterium leprae. J. Bacterio.l 190, 655–661. (doi:10.1128/JB.00982-07) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lee M, Hesek D, Shah IM, Oliver AG, Dworkin J, Mobashery S. 2010. Synthetic peptidoglycan motifs for germination of bacterial spores. ChemBioChem 11, 2525–2529. (doi:10.1002/cbic.201000626) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Jorgensen JH, Turnidges JD, Washington JA. 1999. Antibacterial susceptibility tests: dilution and disk diffusion methods. In Manual of clinical microbiology (eds Murray PR, Baron EJ, Pfaller MA, Tenover FC, Yolken RH.), pp. 1526–1543. Washington, DC: ASM. [Google Scholar]
- 41.Waddell SJ, Butcher PD. 2010. Use of DNA arrays to study transcriptional responses to antimycobacterial compounds. Methods Mol. Biol. 642, 75–91. (doi:10.1007/978-1-60327-279-7_6) [DOI] [PubMed] [Google Scholar]
- 42.Turapov O, et al. 2014. Antimicrobial treatment improves mycobacterial survival in non-permissive growth conditions. Antimicrobial Agents Chemother. 58, 2798–2806. (doi:10.1128/AAC.02774-13) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lee BY, Hefta SA, Brennan PJ. 1992. Characterization of the major membrane protein of virulent Mycobacterium tuberculosis. Infect. Immun. 60, 2066–2074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wisniewski JR, Zougman A, Nagaraj N, Mann M. 2009. Universal sample preparation method for proteome analysis. Nat. Methods 6, 359–362. (doi:10.1038/nmeth.1322) [DOI] [PubMed] [Google Scholar]
- 45.Perkins DN, Pappin DJ, Creasy DM, Cottrell JS. 1999. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 20, 3551–3567. (doi:10.1002/(SICI)1522-2683(19991201)20:18<3551::AID-ELPS3551>3.0.CO;2-2) [DOI] [PubMed] [Google Scholar]
- 46.Keller A, Nesvizhskii AI, Kolker E, Aebersold R. 2002. Empirical statistical model to estimate the accuracy of peptide identification made by MS/MS and database search. Anal. Chem. 74, 5383–5392. (doi:10.1021/ac025747h) [DOI] [PubMed] [Google Scholar]
- 47.Chawla Y, Upadhyay S, Khan S, Nagarajan SN, Forti F, Nandicoori VK. 2014. Protein kinase B (PknB) of Mycobacterium tuberculosis is essential for growth of the pathogen in vitro as well as for survival within the host. J. Biol. Chem. 289, 13 858–13 875. (doi:10.1074/jbc.M114.563536) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ohashi W, Inouye S, Yamazaki T, Hirota H. 2005. NMR analysis of the Mg2+-binding properties of aequorin, a Ca2+-binding photoprotein. J. Biochem. 138, 613–620. (doi:10.1093/jb/mvi164) [DOI] [PubMed] [Google Scholar]
- 49.Horwitz AH, Casida LE., Jr 1978. Survival and reversion of a stable L form in soil. Can. J. Microbiol. 24, 50–55. (doi:10.1139/m78-009) [DOI] [PubMed] [Google Scholar]
- 50.Del Sol R, Mullins JG, Grantcharova N, Flardh K, Dyson P. 2006. Influence of CrgA on assembly of the cell division protein FtsZ during development of Streptomyces coelicolor. J. Bacteriol. 188, 1540–1550. (doi:10.1128/JB.188.4.1540-1550.2006) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Plocinski P, et al. 2012. Mycobacterium tuberculosis CwsA interacts with CrgA and Wag31, and the CrgA-CwsA complex is involved in peptidoglycan synthesis and cell shape determination. J. Bacteriol. 194, 6398–6409. (doi:10.1128/JB.01005-12) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jahnen-Dechent W, Kettler M. 2012. Magnesium basics. Clin. Kidney J 5, i3–i14. (doi:10.1093/ndtplus/sfr163) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Moncany ML, Kellenberger E. 1981. High magnesium content of Escherichia coli B. Experientia 37, 846–847. (doi:10.1007/BF01985672) [DOI] [PubMed] [Google Scholar]
- 54.Yamaguchi A, Yanai M, Tomiyama N, Sawai T. 1986. Effects of magnesium and sodium ions on the outer membrane permeability of cephalosporins in Escherichia coli. FEBS Lett. 208, 43–47. (doi:10.1016/0014-5793(86)81528-9) [DOI] [PubMed] [Google Scholar]
- 55.Piddington DL, Kashkouli A, Buchmeier NA. 2000. Growth of Mycobacterium tuberculosis in a defined medium is very restricted by acid pH and Mg2+ levels. Infect. Immun. 68, 4518–4522. (doi:10.1128/IAI.68.8.4518-4522.2000) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Murray T, Popham DL, Setlow P. 1998. Bacillus subtilis cells lacking penicillin-binding protein 1 require increased levels of divalent cations for growth. J. Bacteriol. 180, 4555–4563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Pensinger DA, et al. 2014. Selective pharmacologic inhibition of a PASTA kinase increases Listeria monocytogenes susceptibility to β-lactam antibiotics. Antimicrobial Agents Chemother. 58, 4486–4494. (doi:10.1128/AAC.02396-14) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Morlot C, Bayle L, Jacq M, Fleurie A, Tourcier G, Galisson F, Vernet T, Grangeasse C, Di Guilmi AM. 2013. Interaction of Penicillin-Binding Protein 2x and Ser/Thr protein kinase StkP, two key players in Streptococcus pneumoniae R6 morphogenesis. Mol. Microbiol. 90, 88–102. (doi:10.1111/mmi.12348) [DOI] [PubMed] [Google Scholar]
- 59.Maestro B, Novaková L, Hesek D, Lee M, Leyva E, Mobashery S, Sanz JM, Branny P. 2011. Recognition of peptidoglycan and β-lactam antibiotics by the extracellular domain of the Ser/Thr protein kinase StkP from Streptococcus pneumoniae. FEBS Lett. 585, 357–363. (doi:10.1016/j.febslet.2010.12.016) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kumar P, Arora K, Lloyd JR, Lee IY, Nair V, Fischer E, Boshoff HI, Barry CE., III 2012. Meropenem inhibits d,d-carboxypeptidase activity in Mycobacterium tuberculosis. Mol. Microbiol. 86, 367–381. (doi:10.1111/j.1365-2958.2012.08199.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.England K, Boshoff HI, Arora K, Weiner D, Dayao E, Schimel D, Via LE, Barry CE., III 2012. Meropenem-clavulanic acid shows activity against Mycobacterium tuberculosis in vivo. Antimicrobial Agents Chemother. 56, 3384–3387. (doi:10.1128/AAC.05690-11) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Nikitushkin VD, Demina GR, Shleeva MO, Kaprelyants AS. 2013. Peptidoglycan fragments stimulate resuscitation of ‘non-culturable’ mycobacteria. Antonie van Leeuwenhoek 103, 37–46. (doi:10.1007/s10482-012-9784-1) [DOI] [PubMed] [Google Scholar]
- 63.Kana BD, Mizrahi V. 2010. Resuscitation-promoting factors as lytic enzymes for bacterial growth and signaling. FEMS Immunol. Med. Microbiol. 58, 39–50. (doi:10.1111/j.1574-695X.2009.00606.x) [DOI] [PubMed] [Google Scholar]
- 64.Kern T, et al. 2010. Dynamics characterization of fully hydrated bacterial cell walls by solid-state NMR: evidence for cooperative binding of metal ions. J. Am. Chem. Soc. 132, 10 911–10 919. (doi:10.1021/ja104533w) [DOI] [PubMed] [Google Scholar]
- 65.Longo G, Alonso-Sarduy L, Rio LM, Bizzini A, Trampuz A, Notz J, Dietler G, Kasas S. 2013. Rapid detection of bacterial resistance to antibiotics using AFM cantilevers as nanomechanical sensors. Nat. Nanotechnol. 8, 522–526. (doi:10.1038/nnano.2013.120) [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets supporting this article have been uploaded as part of the electronic supplementary material.