

Abstract
Tendons and ligaments are extracellular matrix (ECM)-rich structures that interconnect muscles and bones. Recent work has shown how tendon fibroblasts (tenocytes) interact with muscles via the ECM to establish connectivity and strengthen attachments under tension. Similarly, ECM-dependent interactions between tenocytes and cartilage/bone ensure that tendon-bone attachments form with the appropriate strength for the force required. Recent studies have also established a close lineal relationship between tenocytes and skeletal progenitors, highlighting the fact that defects in signals modulated by the ECM can alter the balance between these fates, as occurs in calcifying tendinopathies associated with aging. The dynamic fine-tuning of tendon ECM composition and assembly thus gives rise to the remarkable characteristics of this unique tissue type. Here, we provide an overview of the functions of the ECM in tendon formation and maturation that attempts to integrate findings from developmental genetics with those of matrix biology.
KEY WORDS: Tendon, Ligament, Tenocyte, Extracellular matrix
Summary: This Review discusses how intrinsic regulatory mechanisms as well as extrinsic factors, such as mechanical force, regulate the extracellular matrix to control tendon development and maturation.
Introduction
Tendons and ligaments are connective tissues that transmit mechanical forces between muscles and bones. Tendons attach muscle to skeleton, whereas ligaments attach skeletal elements to each other and stabilize skeletal joints. Vertebrates have evolved a remarkable variety of tendons and ligaments to accommodate their distinct modes of locomotion as well as their dramatic variations in body size and strength. These range from broad sheets to highly elastic cables, such as those of the Achilles’ tendon and the cruciate ligaments of the knee. Because of their structural roles, injuries to tendons and ligaments are extremely common and often debilitating. Thus, a fundamental question in musculoskeletal biology is how these connective tissue structures develop in the correct locations and acquire the strength necessary to translate contractions of muscles into skeletal movements.
Both tendons and ligaments contain fibroblasts (termed tenocytes and ligamentocytes, respectively) embedded in a unique extracellular matrix (ECM) that is composed mainly of collagen fibril arrays capable of withstanding incredibly strong tensile forces. These fibrils are crosslinked to one another and wrapped in a tendon sheath (Banos et al., 2008; Kannus et al., 1998; Ros et al., 1995). This basic structure is shared among tendons and ligaments, and the fibroblasts that produce the ECM develop from common progenitors with similar gene expression profiles (Juneja and Veillette, 2013; Sugimoto et al., 2013; Tozer and Duprez, 2005; Yang et al., 2013). However, each tendon or ligament differs in its precise ECM composition, size and strength (Birch et al., 2013). How are these differences established? The answer to this question has important implications for understanding how diseases or damage to tendons and ligaments arise and for developing better treatment strategies.
Despite their pivotal roles in musculoskeletal connectivity and functional stability, the mechanisms that control tendon development have received much less attention than the processes of myogenesis or skeletogenesis. Only a handful of factors are known to help specify tenocyte progenitor cells (TPCs) at muscle attachments, induce them to differentiate, and maintain and repair them in the adult (Aslan et al., 2008; Huang et al., 2015; Liu et al., 2012, 2014; Schweitzer et al., 2010; Yang et al., 2013). Studies in animal models (e.g. fly, fish, chick and mouse embryos), in particular those focusing on the formation of myotendinous junctions (MTJs; the major sites of force transmission), have revealed a crucial link between developing TPCs and the dynamic ECM that surrounds these cells. Indeed, ECM proteins (e.g. collagens, laminins, thrombospondins) initially guide myofibers to their sites of attachment, but also mediate signaling between TPCs and muscles, regulate the maturation of MTJs, and maintain tendons in response to mechanical force (Kjaer, 2004; Schwartz et al., 2013; Snow and Henry, 2009). Here, we review recent genetic studies that have identified crucial roles for the ECM in tendon development, and we discuss the emerging nexus between the transcriptional control of tenocyte differentiation and the organization of the ECM associated with muscle fibers (myomatrix), MTJs and tendons (Fig. 1).
Fig. 1.
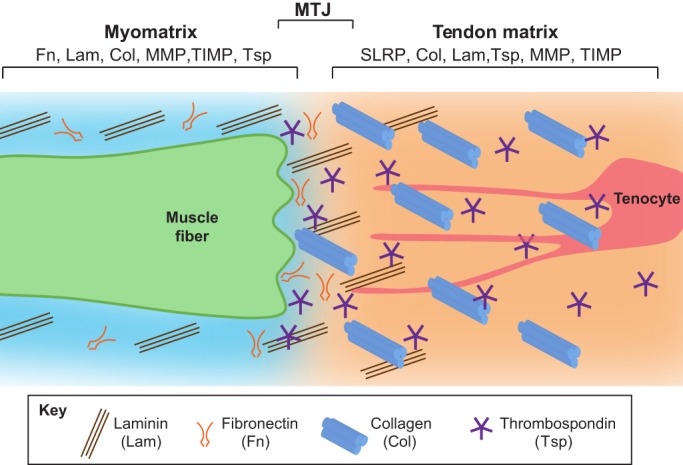
Composition of the ECM surrounding muscle, tendon and myotendinous junctions. A muscle fiber (green) secretes ECM components into its surroundings (the myomatrix). Some of these components overlap with those of the tendon ECM, which is secreted by tenocytes (red). Myomatrix is primarily composed of Lam trimers and Fn. By contrast, the tendon matrix is rich in Col1a trimers and thrombospondin pentamers. The myotendinous junction (MTJ) is the narrow zone in which ECM components of tendon and muscle interact.
ECM production and regulation in developing tendons
Collagens (predominantly Col1a) constitute the bulk of mature mammalian tendon ECM and MTJs, whereas laminins (Lams) and many other non-collagenous ECM components comprise the remainder (Kannus et al., 1998; Kannus, 2000; Kjaer, 2004; Birch et al., 2013; Thorpe et al., 2013). Which cells secrete these ECM proteins, how are they produced in the correct proportions, and how do they assemble?
The transcriptional regulation of ECM production by TPCs
Strikingly, all of the key transcription factors known to function in TPC development directly regulate the transcription of genes encoding ECM proteins (Fig. 2A,B). The best studied of these is Scleraxis (Scx), a basic helix-loop-helix transcription factor, and the earliest known marker of TPCs. Scx is first induced through the interplay of sonic hedgehog (Shh) and fibroblast growth factor (FGF) signaling in the ‘syndetome’ compartment of somites (the region of the sclerotome adjacent to the myotome) and by transforming growth factor beta (TGFβ) signaling in the limbs of mice at embryonic day (E) 10.5 (Schweitzer et al., 2001; Brent et al., 2003; Havis et al., 2014). In vitro, Scx overexpression is sufficient to transform mesenchymal stem cells (MSCs) and human embryonic stem cells (hESCs) into tenocytes (Fig. 2A) (Alberton et al., 2012; Chen et al., 2012; Li et al., 2015). Thus, considerable effort has been made to elucidate the functions of Scx in the tenocyte lineage and to identify its downstream targets.
Fig. 2.

Transcriptional regulation of tenocyte specification and tendon ECM production. (A) A mesenchymal stem cell (MSC, purple), the common progenitor for skeletal and tenocyte progenitors, becomes a skeletogenic progenitor cell (SPC, blue) if it is exposed to high levels of BMP signaling, then it expresses Sox5/6/9 followed by Runx2 during its differentiation into an osteoblast. By contrast, an MSC becomes a tendon progenitor cell (TPC, pink) if it receives high levels of Shh, FGF and TGFβ signaling, then expresses scleraxis (Scx) followed by Mkx, Egr1 and Tnmd during its transition into a tenocyte. The fate of the progenitor cells is determined by the level of Sox9 and Scx. The plasticity of progenitor cell fate at this stage is represented by the double-headed gray arrow. (B) The transcription factors involved in tenocyte specification also regulate the transcription of genes encoding ECM proteins. Direct (black) and indirect (gray) transcriptional target genes regulated by Scx, Egr1, Egr2 and Mkx in TPCs are indicated. Note that Mkx also represses the expression of factors involved in myogenic and skeletogenic progenitor formation. (C) Transcription factors expressed in skeletogenic progenitor cells (e.g. Sox9 and Runx2) directly regulate the transcription of a distinct set of ECM target genes.
The loss of Scx (i.e. as in Scx−/− mutant mice) disrupts tenocyte differentiation leading to atrophy of force-transmitting tendons and a disorganized tendon ECM (Murchison et al., 2007). The expression of the major structural collagens, Col1a1, Col1a2, Col3a1 and Col14a1, is strongly reduced in Scx−/− mutants. Scx directly controls Col1a1 and Col1a2 transcription (Fig. 2B) (Espira et al., 2009; Léjard et al., 2007). Numerous other tendon regulators, such as the glycoprotein tenomodulin (Tnmd), are also downregulated in Scx−/− mutants (Murchison et al., 2007; Shukunami et al., 2006). At the ultrastructural level, the loss of Scx disrupts the sheaths that surround each fascicle of collagen fibrils as well as cellular processes, which normally encircle the fibrils (Murchison et al., 2007). These results demonstrate that Scx controls tendon ECM production, which is essential for effective force transmission.
It should be noted that TPCs still develop in Scx−/− mutant mice, suggesting that other genes are required for the initial steps of TPC specification. In Drosophila, the transcription factor Stripe (Sr) specifies TPCs in epidermal segment border cells (Volk and VijayRaghavan, 1994). Flies lacking Sr function fail to form TPCs and display disrupted muscle patterning and attachments, whereas Sr overexpression transforms ectodermal progenitors into TPCs (Becker et al., 1997). Embryonic TPCs in mice express orthologs of Sr – Egr1 and Egr2 (Lejard et al., 2011) – and Egr1 is sufficient to induce Scx expression and specify MSCs as tenocytes in vitro (Guerquin et al., 2013). However, like Scx, both Egr1 and Egr2 appear to be dispensable for tenocyte specification, as Egr1−/−/Egr2−/− double mutant mice are viable. Instead they regulate the tendon ECM and MTJ, binding to tendon-specific enhancer elements of Col1a1 and Col1a2 that are also bound by Scx (Fig. 2B) (Léjard et al., 2007, 2011; Guerquin et al., 2013). Egr1−/− mutant mice also downregulate Tnmd and are slow to heal tendon injuries as adults (Guerquin et al., 2013). Thus, in contrast to flies, vertebrate Egrs function in tendon ECM production rather than TPC specification.
Another potential TPC ‘specifier’ is the TALE family atypical Iroquois-like homeodomain protein Mohawk (Mkx). Like Scx and Egr1, Mkx can drive bone marrow-derived MSCs towards a tenocyte fate in vitro (Liu et al., 2015; Otabe et al., 2015). However, in mouse embryos the expression of Mkx begins in developing tenocytes later than that of Scx or Egr1/2 (at E13.5-14.5), becoming restricted to tendon sheath cells and collateral ligaments (which stabilize joints) in the limbs by E16.5 (Anderson et al., 2006). Mkx−/− mutant mice are viable, fertile, and form normal tendons at first with no defects in Scx expression but later exhibit reduced levels of Col1a1, Col1a2, Tnmd, fibromodulin (Fmod) and decorin (Dcn) as well as thinning of collagen fibrils (Ito et al., 2010; Kimura et al., 2011; Liu et al., 2010). Similar to Scx, Mkx can function as a transcriptional activator when complexed with Smad2/3 to promote Col1a1, Col1a2, Tnmd and Dcn expression as well as TGFβ2 expression in murine MSCs (Liu et al., 2015). However, at other promoters it interacts with the Sin3A/histone deacetylase (HDAC) complex to repress gene expression, including that of key myogenic factors such as MyoD (Myod1), Sox6 and the cartilage determinant Sox9 (Fig. 2B) (Anderson et al., 2009; Anderson et al., 2012; Berthet et al., 2013; Chuang et al., 2014). Thus, like Scx and Egr1/2, Mkx controls tendon maturation and ECM production and might function, in part, to maintain tenocytes by preventing them from acquiring myogenic or skeletogenic fates.
Both Scx and Mkx interact with Smad3, an essential transcriptional mediator of TGFβ signaling, to regulate tendon ECM production (Berthet et al., 2013; Hosokawa et al., 2010; Katzel et al., 2011; Oka et al., 2008; Pryce et al., 2009). Accordingly, Tgfb2−/−/Tgfb3−/− conditional double mutant mice, or conditional mutant mice lacking Tgfβr2 receptors in limb mesenchyme, initially form TPCs in the limbs but lose them by E14.5, suggesting a role for TGFβ signaling in tendon maintenance (Pryce et al., 2009). Tendon defects in Smad3−/− mutants are much less severe, with transient reductions in Col1a1, Col1a2 and Tnmd expression in their limbs. Like Mkx, Smad3 can also inhibit the expression and activity of MyoD as well as that of skeletogenic factors such as Runx2 in vitro (Fig. 2A, Fig. 3) (Alliston et al., 2001; Kang et al., 2005; Liu et al., 2001). However, unlike Mkx, which represses MyoD transcription, Smad3 acts post-translationally by binding E-box sites in MyoD and sequestering it away from its targets (Chuang et al., 2014). These results hint at a dynamic network involving TGFβ signaling, Scx and Mkx to achieve and maintain the tenocyte fate.
Fig. 3.
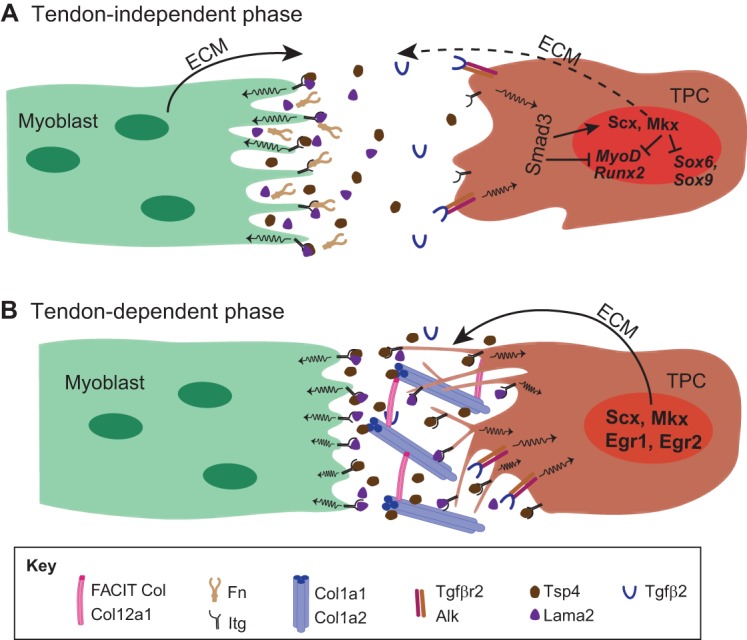
Myoblast-tenocyte interactions and ECM production. (A,B) The formation of myotendinous junctions can be considered as a two-step process. In the initial tendon-independent phase (A) in vertebrates (shown here for zebrafish trunk muscles), myoblasts (green) synthesize a ‘pre-tendon’ ECM that includes the integrin ligands Tsp4 and Lama2. This ECM accumulates in the absence of TPCs (brown). Mechanotransduction coupled with TGFβ signaling (through Tgfβ2 and Tgfβr2) leads to the Smad3-dependent expression of Scx and Mkx in TPCs, which in turn leads to the expression of tendon-selective ECM genes. Smad3 and Mkx also repress the activity of MyoD, Sox9 and Runx2 to repress myogenic and skeletogenic fates during tenocyte differentiation. A later tendon-dependent phase (B) relies on the production of ECM, particularly Col1a1, Col1a2, Col12a1 and Col14a1, by more mature TPCs, which extend processes into the ECM.
In summary, to date no single factor fits the bill as being both necessary and sufficient for TPC specification. Rather, transcriptional regulators of tenocytes share functions in the production of tendon ECM and MTJ assembly and in the repression of other mesenchymal fates. This is important not only in the context of tenocyte development but also in the regulation of MSCs, where the balance between these factors determines if a cell becomes a TPC or a skeletogenic (or myogenic) progenitor (Fig. 2A). An attractive model is one in which Scx (in concert with TGFβ signaling) shifts the fate of MSCs towards TPCs and initiates tendon ECM production, whereas factors expressed later in development, such as Mkx and Egr1, supplement the role of Scx by inducing the expression of ECM proteins as well as by repressing myogenic and skeletogenic fates. Furthermore, signaling mediated by mechanical forces upregulates expression of Scx, Mkx and Smad3, stimulating more ECM production, thereby providing positive feedback for fine-tuning tendon strength, as we discuss further below (Eliasson et al., 2008; Maeda et al., 2011).
ECM production and function during tenocyte morphogenesis and MTJ formation
Although the migration of muscle progenitors has been well studied in both invertebrates and vertebrates, very little is known about the role of ECM in morphogenesis of TPCs and the establishment of the MTJ. In Drosophila, myoblasts migrate to sites of attachment and interact with tenocytes located at fixed sites at segment borders in the epidermis (Volk and VijayRaghavan, 1994; Schweitzer et al., 2010). These myoblasts recognize tenocytes through multiple signals including Thrombospondin (Tsp) in the ECM, which binds muscle integrins (Itgs) (Subramanian et al., 2007). Similarly, vertebrate myoblasts in the trunk and limbs elongate and attach via TPCs already localized to future muscle attachment sites. How do migrating vertebrate myofibers and TPCs interact, and how do these interactions differ between the trunk, limb and head? In the chick, early progenitor pools of limb TPCs condense and split to form individual tendons (Kardon, 1998). By contrast, cranial TPCs that arise in the neural crest migrate to the locations of future MTJs (Grenier et al., 2009; Noden, 1988; Noden and Trainor, 2005). Trunk TPCs arise from the syndetome of somites, whereas limb TPCs are thought to originate from lateral plate mesoderm (Brent et al., 2003; Kardon, 1998). These distinct embryonic origins and modes of tendon morphogenesis raise the question of where the patterning information for muscle connectivity lies, within the TPCs or within the muscles themselves? Chick-quail chimera studies suggests that, at least for cranial and limb muscles, the TPCs and tendon matrix determine the pattern of attachments (Kardon, 1998; Kieny and Chevallier, 1979; Noden, 1988). Do muscles and the myomatrix play any role in TPC formation or maintenance? In the limbs of chick embryos in which muscle progenitors have been surgically removed, tenocytes initially develop in the correct locations but later degenerate (Kardon, 1998). Limb tendons also degenerate in MyoD−/− and Pax3−/− mutant mice that lack the entire limb musculature (Bonnin et al., 2005; Brent et al., 2005). Cranial tendons in myod1−/−/myf5−/− double mutant zebrafish show similar defects (Chen and Galloway, 2014). These results suggest a dependence on muscles and the myomatrix for tenocyte maintenance but not specification. As discussed below, these phenotypes could be due to a lack of mechanical forces transmitted via the ECM.
What are the roles of specific ECM components of the myomatrix or tendon ECM during the initial establishment of contact between myoblasts and TPCs at attachment sites? Studies of axial muscles in the zebrafish trunk have provided insights into this process (Snow and Henry, 2009). These muscles attach to intersegmental boundaries (ISBs) during embryogenesis before the appearance of TPCs. ISBs are anatomically distinct from later tendons, but contain many of the same ECM components and serve analogous functions in bearing the forces of muscle contraction. During this ‘tendon-independent’ phase of development (Fig. 3A), fibronectin (Fn) and laminin-alpha2 (Lama2) in the myomatrix are highly enriched at ISBs as the myoblasts elongate and are required for embryonic muscle attachments (Koshida et al., 2005; Snow et al., 2008). Mammalian muscles also require Fn and Lam for migration and attachment (Turner et al., 1983; Bajanca et al., 2006; Vaz et al., 2012). Fn and Lam bind to Itg and dystrophin/dystroglycan complexes on muscle cell surfaces. In zebrafish, this leads to localized phosphorylation of focal adhesion kinase (pFAK; Ptk2ab – Zebrafish Information Network) at the ends of myofibers where they attach to ISBs, which stabilizes myotome boundaries (Bassett et al., 2003; Henry et al., 2005; Parsons et al., 2002; Snow et al., 2008). TPCs only appear later along the ISBs, where they contribute additional ECM to strengthen existing attachments in what we refer to here as a ‘tendon-dependent’ phase (Fig. 3B) (Charvet et al., 2011, 2013; Chen and Galloway, 2014; Subramanian and Schilling, 2014). Thus, in zebrafish, somitic muscles attach via the myomatrix at ISBs prior to the appearance of TPCs. This may help explain how other types of attachments, such as ‘fleshy insertions’ of mammalian muscles, develop.
Another ECM protein recently shown to be crucial for tendon development is thrombospondin 4 (Tsp4; also known as Thbs4). Like Fn and Lam, zebrafish Tsp4b is initially produced by myoblasts and accumulates at ISBs prior to muscle attachment (Fig. 3A) (Subramanian and Schilling, 2014). Tsp4b maintains muscle attachments at the ISB, and its depletion leads to detachment upon contraction. The transplantation of wild-type myoblasts into Tsp4b-deficient embryos locally rescues muscle attachments at ISBs, consistent with a role for Tsp4b in the myomatrix during the tendon-independent phase of attachment (Subramanian and Schilling, 2014). Interestingly, tsp4b mRNA abruptly disappears from differentiating myofibers as they attach, suggesting a feedback mechanism that regulates tsp4b transcription. Mammalian myoblasts also express Tsp4, and human TSP4 (THBS4) expression increases in pathological conditions such as Duchenne muscular dystrophy and alpha-sarcoglycanopathies (Chen et al., 2000; Jelinsky et al., 2010). Thus, Tsp4 in the myomatrix helps muscles attach in the absence of TPCs and might also facilitate subsequent tendon maintenance and response to damage.
ECM and collagen fibril assembly during tendon and MTJ maturation
As MTJs mature, tenocytes secrete the bulk of the MTJ/tendon ECM, particularly the many proteins and proteoglycans that make up the core functional units, the collagen fibrils (Fig. 3B). These interact with one another and align into groups of fibrils or fascicles during progressive phases of muscle attachment (Fig. 4). This fibrillar organization is essential for tendons to bear the stress of muscle contraction and prevent bone detachment (avulsion fractures) by controlling force distribution (Birch et al., 2013; Pan et al., 2013; Schwartz et al., 2013; Pingel et al., 2014). The fibrillar network of proteins includes: (1) core force-transmitting, structural collagens (particularly Col1a1, Col1a2, Col2a1 and Col3a1); (2) scaffolding proteins [e.g. Tsp2 (Thbs2), Tsp4, Comp, Lama2]; and (3) specialized crosslinking collagens (e.g. Col6a1, Col12a1, Col14a1 and Col22a1) and various crosslinking factors [e.g. Dcn, Fmod, biglycan (Bgn)], which hold fibrils together to distribute forces efficiently and reduce friction (Figs 3, 4) (Wang et al., 2012; Dunkman et al., 2013). Although the integration of structural collagens into fibrils has been well documented, recent studies have provided insight into the functions of scaffolding proteins and specialized collagens during fibril assembly. These studies suggest that similar to the early ECM at developing MTJs, ECM proteins of the maturing tendon provide continuous feedback in response to mechanical force (Choi et al., 2014; Popov et al., 2015; Wall and Banes, 2005; Zhang and Wang, 2010).
Fig. 4.

Maturation and assembly of the tendon ECM. Diagram illustrating progressive changes in the ECM at an MTJ as it matures. (A) In the early attachment phase, myoblasts (green) first extend towards a cartilage condensation (blue) and reorganize the local ECM by secreting Tsp4 (red), which interacts with Fn and Lam. A magnified view (right) of the boxed area illustrates how Tsp4 pentamers assemble Fn, Lam and Dcn and facilitate binding to Itgs on both muscle and cartilage cell surfaces, thereby promoting adhesion. (B) Following this, in the mid-attachment phase, linear collagen fibrils (Col1a1 trimers, dark blue) form, tenocytes (red) invade, and Sox9+/Scx+ progenitors (dark blue and orange) become detected at the future attachment site on the cartilage, the enthesis. The magnified view illustrates how Col1a1 trimers begin to align perpendicular to skeletal cells (enthesis, dark blue and orange). Dystrophin (DMD) complexes appear on muscle surfaces. (C) In the final late attachment phase, collagen fibrils become crosslinked into a lattice, with tenocytes (red cells) extending processes to surround fibrils, and entheses chondrifying (purple). The magnified view shows Col1a1 trimers becoming crosslinked by FACIT collagens and surrounded by tenocyte (red) processes, stabilizing the ECM and its interactions with Itgs on muscle and cartilage cells.
Col12a1, Col14a1 and Col22a1 belong to the class of fibril-associated collagens with interrupted triple helices (FACITs), which localize to muscle attachments in avian and mouse tendons, and are also expressed in human tendon fibroblasts (Fig. 3B; Fig. 4) (Koch et al., 2004; Wälchli et al., 1994). Studies of FACITs at zebrafish ISBs have been informative for understanding their functions as MTJs mature. In zebrafish, muscles first express Col12a1 and Col22a1 at larval stages and these progressively align into the orthogonal fibril arrays of mature MTJ/tendon ECM (Charvet et al., 2011, 2013). Col12a1 is expressed earlier than Col22a1 and colocalizes with Lama2 throughout muscle fiber attachment (Bader et al., 2009). Col22a1 maintains attachments under tension, and its expression increases in tendinopathies (see Box 1) in humans (Charvet et al., 2013; Jelinsky et al., 2011). Recently, mutations in the human COL12A1 gene that disrupt COL12A1 secretion have been linked to a form of Bethlem myopathy (Bushby et al., 2014; Schessl et al., 2006); other forms of this myopathy are caused by mutations in COL6A1, COL6A2 and COL6A3, which also show elevated expression in human tendinopathies (Bönnemann, 2011; Jelinsky et al., 2011). In mice, Col12a1 interacts with tenascin C (Tnc) and helps crosslink other collagens during fibril maturation by interacting with Dcn (Veit et al., 2006). Transmission electron microscopy studies also suggest that Col12a1 forms complexes with structural collagens (e.g. Col1a1, Col1a2), as well as other scaffolding and crosslinking proteins (Dcn, Fmod, thrombospondins) (Font et al., 1996). Thus the FACIT proteins are highly conserved regulators of tendon ultrastructure and elasticity.
Box 1. Tendinopathies
The term ‘tendinopathy’ refers to a diverse set of tendon disorders (overlapping in some cases both in genetic and molecular terms with myopathies) that are caused either by genetic mutations in ECM components of MTJs or by mechanical stress that leads to tendon ECM damage. Hallmarks of tendinopathies include COL1A1, COL1A2, COL4A1 and COL4A2 overexpression, fibril disorganization, increased collagen crosslinking, reduced tissue inhibition of MMPs (i.e. TIMP activity), and elevated expression of MMP2, MMP14 and MMP19 as well as of versican, biglycan and Dcn (Jelinsky et al., 2011; Parkinson et al., 2011; Dunkman et al., 2013; Zhou et al., 2014). Recent studies have also identified COL6A1/2/3 and COL12A1 as genes underlying one form of human ‘myotendinopathy’, which affects both myomatrix structure and tendon matrix structure (Bönnemann, 2011; Pan et al., 2013).
Other proteins involved in fibril assembly include the small leucine-rich proteoglycans (SLRPs) such as Dcn (Fig. 4), Fmod, biglycan and lumican. These are found in relatively small amounts in tendons, yet loss-of-function mutations in the genes encoding these proteins disrupt collagen fibrillogenesis (Chakravarti, 2002; Corsi et al., 2002; Zhang et al., 2006). SLRPs mainly crosslink collagen fibrils, but they also appear to cross-regulate one another's transcription through as yet unknown mechanisms (Yoon and Halper, 2005; Zhang et al., 2006; Dunkman et al., 2013). This might involve feedback regulation of Scx and Mkx expression through the tendon ECM, and recent studies have suggested that Dcn and Fmod are regulated by Mkx (Fig. 2B) (Ito et al., 2010; Liu et al., 2010; Alberton et al., 2012). These results point to a system by which the dynamics of fibril ultrastructure feedback on ECM production to modify tendon strength continuously.
Tendon ECM and collagen fibrils are also continuously remodeled in response to mechanical forces, at least around the circumference of a tendon fascicle (Frolova et al., 2014; Heinemeier et al., 2012; Herchenhan et al., 2013; Kjaer, 2004; Pingel et al., 2014). TPCs in culture subjected to moderate mechanical forces show an increase in collagen fibril diameter, though diameter decreases with excessive mechanical force (De Almeida et al., 2010; Pingel et al., 2014). This remodeling occurs primarily through the activities of matrix metalloproteinases (MMPs) and their corresponding tissue inhibitors (TIMPs), as well as disintegrin and metalloprotease with thrombospondin repeats (ADAMTS) proteases (Bedi et al., 2010; Gotoh et al., 2013; Jones et al., 2006; Maeda et al., 2013). Almost all of the 23 MMPs and 19 ADAMTS proteins known to be expressed in vertebrates are detectable in adult tendon tissue and play a variety of both positive and negative roles in establishing a functional MTJ and tendon ECM (Davis et al., 2013; Spanoudes et al., 2014). MMPs are zinc-dependent endopeptidases that bind and unwind the triple helix of collagen monomers. Collagenases such as Mmp1 target the structural collagens for degradation, whereas gelatinases such as Mmp2 and Mmp9 and membrane-bound MMPs, such as Mmp14, degrade smaller network collagens. Importantly, recent studies have shown that the expression and activity of MMPs are regulated by signals activated in response to mechanical forces, such as Itg and Tgfβ (Yu and Stamenkovic, 2000; Farhat et al., 2015). MMP misregulation also occurs upon tendon inflammation, and recent studies suggest that MMP inhibition can improve tendon repair (Bedi et al., 2010; Jelinsky et al., 2011; Farhat et al., 2012; Davis et al., 2013).
ECM-mediated signaling during tendon and MTJ formation, maturation and repair
Multiple signaling pathways involving the ECM influence both the formation of muscle attachments and the maturation of tenocytes. Important players in muscle cells include Itgs and dystrophin, which interact with ECM components at the MTJ. Furthermore, in both muscles and tendons, mechanical forces are thought to have a role in dynamic remodeling of the ECM.
Integrin signaling
Many collagens and laminins at developing MTJs directly bind Itg complexes that are present in the membranes of muscle cells and tenocytes (Fig. 5) (Docheva et al., 2014; Mayer et al., 1997; Pan et al., 2013; Rooney et al., 2006, 2012). In muscle, these continuous structural links between ECM proteins, the sarcolemma and the cytoskeleton, maintain fiber integrity and modulate adhesion and gene expression. Not surprisingly, several types of human tendinopathies (see Box 1) are associated with changes in the expression of Itgs and their ligands (Bönnemann, 2011; Jelinsky et al., 2011; Schessl et al., 2006). It is thus important to determine the specific roles of Itg signaling during MTJ maturation and tendon repair after injury.
Fig. 5.

Model for ECM-mediated feedback from mechanical force and its effects on tenocyte gene expression. A tenocyte (orange) synthesizes the tendon ECM, including Lama2, Lama4 (brown), Tsp4b (red pentagons), Col1a (blue) and FACIT Col (purple), all of which signal through Itg receptors (dark blue) on muscle and tenocyte cell surfaces in response to mechanical stress (gray arrows). In addition, stress causes the ECM to release TGFβ (yellow) from the TGFβ large latent complex (LLC) (gray dotted arrows). Itg and TGFβ signaling in tenocytes feedback to regulate Scx-, Egr1/2- and Mkx-induced transcription (dashed arrows) of the same Itg ligands as well as of other ECM components to modulate tendon stiffness. Smad3 also interacts with Scx and Mkx to activate target ECM genes. The muscle fiber also contributes to the tendon matrix by secreting FACIT Col22a1.
Different Itg heterodimer combinations lend specificity for different ligands. For example, laminins (Lama2, Lama4) bind Itga7/b1 in the muscle basement membrane (Fig. 5) (Yurchenco et al., 2004; Durbeej, 2010; Carmignac and Durbeej, 2012), and defects in LAMA2 have been associated with merosin-deficient muscular dystrophy in humans (Tomé et al., 1994; Rooney et al., 2012). Furthermore, mutations that disrupt genes encoding crosslinking collagens (e.g. COL12A1 in the case of Ehlers–Danlos syndrome), which bind Itga1/b1 or Itga2/b1, cause widespread defects in skin, bones and tendons (Mayer et al., 1997; Zou et al., 2014).
Which other Itg ligands control MTJ formation and maturation? In Drosophila, Sr promotes the transcription of the single fly thrombospondin (Tsp) gene in tenocytes. Tsp binds αPS2 (If)/βPS (Mys) Itg heterodimers on fly muscle and tendon cell surfaces and patterns muscle attachments (Chanana et al., 2007; Subramanian et al., 2007). Vertebrates have at least five thrombospondins, with diverse functions in cell migration, vasculogenesis, wound healing and cancer (Adams and Lawler, 2004; Bornstein et al., 2004; Kyriakides et al., 1999; Mustonen et al., 2012). Among these, zebrafish Tsp4b is first secreted by myoblasts (Fig. 3A) and later by TPCs (Fig. 3B), and is essential for muscle attachment (Subramanian and Schilling, 2014). Accordingly, zebrafish embryos depleted of Tsp4b have defects in laminin localization and FAK phosphorylation (indicating reduced Itg signaling) at developing ISBs, and their muscles detach under tension. These findings suggest that, in zebrafish, Tsp4 plays key roles in organizing the ECM of both muscle and tendon, particularly those components essential for Itg signaling. This requirement for Tsp4 is at least partially conserved, as Tsp4−/− mutant mice show defects in ECM deposition in developing tendons (Fig. 4) (Frolova et al., 2014; Subramanian and Schilling, 2014). Tsp4 and other subtype B thrombospondins form pentamers that directly bind to collagens, laminins and other ECM proteins (Hauser et al., 1995). Surprisingly, human TSP4 protein microinjected into the ECM surrounding Tsp4b-deficient myofibers in zebrafish localizes to ISBs and locally rescues laminin localization, Itg signaling and muscle attachments, suggesting that Tsp4 could function as a scaffold for other ECM proteins during their assembly at muscle attachments. Consistent with this model, the ability of microinjected zebrafish tsp4b mRNA to rescue Tsp4b-deficient attachments requires Itg binding and pentamerization; mutation of either the Itg-binding (KGD) domain or the pentamerization (CQAC) domain of Tsp4b disrupts its localization to tendons and eliminates its ability to rescue muscle attachments in Tsp4b-deficient larvae (Subramanian and Schilling, 2014). These results, along with a potentially conserved requirement for Tsp4 in mice, suggest that TSP4 defects could contribute to human tendinopathies, highlighting TSP4 as an attractive therapeutic target for strengthening the tendon ECM. They also highlight the close association between the structural and signaling roles of the ECM.
TGFβ signaling
TGFβ signaling provides another striking example of the relationship between ECM structure and signaling in tendons. Genetic studies in mice have revealed crucial roles for TGFβ at multiple steps in tendon development, maturation, maintenance and repair (Figs 2, 3, 5). Removing the functions of both TGFβ2 and TGFβ3 ligands, or of TGFβr2, eliminates most if not all differentiated tendons, whereas exogenous TGFβ is sufficient to induce the expression of Scx and Col1a1 (Fig. 2A) (Chuang et al., 2014; Pryce et al., 2009). Tgfb2−/−/Tgfb3−/− mutant mice lose Scx expression in TPCs between E11.5 and E12.5, suggesting that TGFβ is required for TPC maintenance (Pryce et al., 2009). Signaling through TGFβr2 phosphorylates Smad2 and Smad3, which translocate to the nucleus and activate target genes, thereby maintaining differentiated tenocytes (Figs 3 and 5). Mouse Smad3−/− mutants have reduced tendon tensile strength and increased spacing between collagen fascicles as well as reduced Mkx and increased Mmp9 expression (Berthet et al., 2013; Katzel et al., 2011).
Interestingly, the most likely source of TGFβ ligand at muscle attachments is the ECM. A recent in vitro study has shown that during tenocyte differentiation, Mkx activates the expression of TGFβ in differentiating MSCs (Fig. 5) (Liu et al., 2015). TGFβs are secreted bound to latent TGFβ-binding proteins (LTBPs), which form part of the large latency complex (LLC) in the ECM (Wipff et al., 2007; Maeda et al., 2011). They are also secreted along with latency-associated peptides (LAPs), which block association with TGFβ receptors, and along with other proteins of the LLC, they become incorporated into ECM via interactions between LTBPs and Fn, fibrillin or Dcn (Isogai et al., 2003; Rifkin, 2005; Farhat et al., 2012). In this manner, TGFβs are stored in the ECM and must be released from the LLC and LAPs in order to be ‘activated’ and available to interact with cognate receptors (Horiguchi et al., 2012). Activation may occur by release of TGFβ stimulated by shearing forces, LLC degradation by proteases, interactions with Itgs through RGD motifs on LTBPs themselves, or by the activity of Mmp2, Mmp9 and Bmp1 proteases (Munger and Sheppard, 2011). Few studies have addressed these mechanisms of TGFβ activation specifically for the ECM of tendons or MTJs. A recent transcriptomic analysis of Scx-expressing tenocytes from mouse limbs reveals that both TGFβ and MAPK signaling are strongly upregulated, but that only TGFβ upregulates Tsp2, Tsp4 and LTBP components of the ECM and promotes the tenocyte cell fate (Havis et al., 2014).
The correct regulation of TGFβ activity is crucial not only for tendon development but also for healing injured tendons. Injuries lead to excessive release of TGFβ owing to mechanical force-mediated activation of TGFβ (discussed below) and can cause fibrotic scarring of the tendon, thereby disrupting its function (Farhat et al., 2015). In one model, elevated TGFβ might overactivate MMPs, which in turn promotes further release of active TGFβ from the ECM as well as activating expression of Scx and Mkx and driving further ECM production.
Mechanical forces and signaling
As alluded to throughout this Review, tenocytes actively sense mechanical force, leading to changes in gene expression, cytoskeletal organization and ECM secretion (Fig. 5) (Banos et al., 2008; Maeda et al., 2009, 2013). Such feedback must be extremely important for a tissue that constantly adjusts its stiffness to changing loads. It depends, at least in part, on signaling through gap junctional complexes localized to tenocyte processes (Kruegel and Miosge, 2010; Maeda et al., 2012). Indeed, rats subjected to running on treadmills have increased Tnmd and Col1a1 expression as well as TPC proliferation (Eliasson et al., 2009; Zhang and Wang, 2013). Elevated secretion of both Col4a1 and Col6a1 is also seen in developing chick tendons under stress, and this alters the crosslinking of fibrils (Marturano et al., 2014), thereby fine-tuning tendon strength and promoting repair (Bailey et al., 1998; Willett et al., 2010). What are the molecular mechanisms underlying these cellular responses?
TGFβ signaling is one such mechanosensitive pathway that could control the response of tendons to force. Mechanical force causes release of TGFβ1 from LTBPs in the ECM (Fig. 5) (Maeda et al., 2011; Wipff et al., 2007). Under normal loads, TGFβ signaling through Smad2/3 maintains Scx expression in tenocytes, whereas excessive loading disrupts TGFβ signaling, damages the ECM and leads to tenocyte cell death in mice (Maeda et al., 2009, 2010, 2011). TGFβ signaling in response to force also upregulates ITGA1 and ITGA2 expression in human TPC cultures (Popov et al., 2015). Tenocytes elevate expression of Tgfb1, Tgfbr2 and Smad7 in response to injury in mice (Guerquin et al., 2013). Thus, one attractive model is that force triggers TGFβ signaling leading to increased expression of Scx and Mkx, which in turn activates TGFβ expression, creating a positive-feedback loop that leads secondarily to remodeling/strengthening of ECM (Fig. 5) (Liu et al., 2015). Notably, TGFβ signaling in response to mechanical forces also controls MMP expression during tendon repair (Katzel et al., 2011; Farhat et al., 2015). Interestingly, injured tendons in Egr1−/− mutant mice fail to upregulate TGFβ and Scx or to repair their tendons efficiently (Guerquin et al., 2013).
The unique collagen fibril organization of tendons allows them to bear the stress of muscle contraction and prevents bone fractures (Davis et al., 2013; Pan et al., 2013; Schwartz et al., 2013; Zhang et al., 2006). How fibrils physically anchor to cells and how the dynamics of these anchors are regulated under load remain unclear. Biopsies of human Achilles’ tendons have shown that fibrils buckle in overloaded tendons (Pingel et al., 2014). In tenocytes cultured in collagen gels, mechanical force translates into cytoskeletal force through non-muscle myosin II bound to actin fibrils associated with focal adhesion complexes, which in turn associate with Itgs and other ECM receptors. The chemical inhibition of myosin II function (using blebbistatin) reduces cytoskeletal traction forces and leads to ECM remodeling (Maeda et al., 2013). Thus, in addition to inducing transcriptional changes, mechanical force can stimulate changes in cytoskeletal tension that reverberate back to the ECM to alter its ultrastructure.
MMPs have also been shown to modulate ECM structure in response to mechanical cues during MTJ maturation and after injury. For example, tenocytes in silicone micropillar gels elevate MMP expression levels in response to gel deformation (Maeda et al., 2013). In addition, humans show dramatic increases in the levels of MMP2, MMP9 and MMP14 in adult tendons following endurance exercise, suggesting that these proteins aid MTJ repair in response to mechanotransduction (Rullman et al., 2009). This effect depends on the timing of loading as, in cultured tendon fascicles, cells upregulate MMP2 and MMP13 after very short cycles of loading, but downregulate MMP1 after longer cycles (Maeda et al., 2009, 2013). Similarly, in vitro studies of mechanical loading on mouse tenocytes have shown that, whereas low levels of shear force lead to upregulation of Col1a and Tmnd, increasing the force leads to upregulation of Runx2 and Sox9 (Zhang and Wang, 2015). Understanding these dynamic responses to mechanical stimulation might lead to improved therapeutic interventions; the systemic inhibition of MMPs, for example, can reduce fibrotic scarring of muscle ECM (Farhat et al., 2015). MMP expression is also regulated by TGFβs (Yu and Stamenkovic, 2000; Ge and Greenspan, 2006; Farhat et al., 2015). In turn, MMP2, MMP9 and BMP1 proteases might specifically digest LLC and release active TGFβ, establishing a positive-feedback loop that could help fine-tune MMP levels, both during normal tendon function and in response to tendon injury or exercise (Fig. 5).
ECM functions at tendon-bone attachments
So far, we have emphasized the ECM associated with tendon-muscle attachments. But tendons also attach, at their other ends, to cartilage/bone under a unique set of mechanical constraints. Many tendons insert into bony protrusions known as osteotendinous junctions or ‘entheses’, such as the deltoid tuberosity on the humerus (Fig. 6A). A characteristic structural feature of entheses is the presence of ‘fibrocartilage’, a tissue with physical properties somewhere in between cartilage and tendon. Within an enthesis, a rapid transition from a more bone-like cellular and ECM structure to a more cartilage-like (less rigid) structure in regions closer to the tendon is observed. In addition, a unique mineralization gradient forms from the bony front to the point of tendon insertion, the width of which is constant with corresponding changes in cellular density (Schwartz et al., 2012). The enthesis ECM also shows more Dcn and Bgn localized to the tendon side, whereas Col2a1, Col9, Col10 and aggrecan localize to the bony side (Thomopoulos et al., 2003). These gradual transitions in ECM and rigidity are essential for the proper transmission of contractile forces to the bone to prevent avulsion fractures (Zelzer et al., 2014).
Fig. 6.

ECM functions at tendon-bone attachments. (A) Diagrams illustrating changes in cartilage at the developing humero-ulnar joint of the mouse forelimb in wild-type embryos (left) and in splotch delayed (spd, Pax3) mutants (right), which lack muscles. Proliferating chondrocytes express Col2a1 (light blue), whereas cells forming at the edges of the joint express Col2b (dark blue), and cells in the joint interzone secrete Gdf5 (green) into the joint region. The loss of muscles in spd mutants leads to loss of Gdf5 expression, disorganized Col2b+ interzone cells and joint fusion. (B) Diagrams illustrating changes in cartilage and tenocytes at a developing eminence. The primary field contains cells that form chondrocytes within the developing bone, whereas the secondary field consists of Sox9-positive progenitor cells that lie outside of the primary field. In wild-type embryos, three different subsets of Scx-expressing cells at a muscle insertion site of a developing long bone are found: Sox9+/Scx+, Scx+ or Scx+/Bmp4+. Loss of Tgfβr2 in limb mesenchyme or of Sox9 in tenocytes leads to a loss of the Sox9/Scx co-expressing and Sox9-expressing population in the secondary field, but not other tenocytes. Loss of Bmp4 signaling leads to a loss of both Sox9+/Scx+ and Scx+ populations in the secondary field. Dotted lines outline primary field. Dashed lines outline secondary field. lof, loss of function.
In the appendicular skeleton, entheses are established through specialized contours and protuberances on bones called eminences. Recent genetic studies in mice have begun to elucidate the signals that control the formation of these structures (Blitz et al., 2009, 2013; Zelzer et al., 2014). In mouse embryos, eminence development coincides with the formation of muscle/tendon attachments, suggesting that attachments impose physical changes on the bone. However, mouse mutants that lack muscles, such as splotch delayed (spd) or muscular dysgenesis (mdg) mutants, still form eminences like the deltoid tuberosity on the humerus, despite severe joint fusions (Fig. 6B) (Blitz et al., 2009).
Perhaps not surprisingly, TGFβ signaling also plays important roles in the formation of entheses. Loss of TGFβ signaling through ablation of Tgfβr2 in limb mesenchyme eliminates eminences in the limb, though this might be due to a broader or earlier role for TGFβ signaling in skeletal/tendon progenitors (Fig. 6B) (Blitz et al., 2009, 2013). Instead, evidence is building to suggest that the more crucial inducers of eminences are other members of the TGFβ superfamily, namely bone morphogenetic proteins (BMPs). The conditional deletion in mice of Bmp4 specifically in tenocytes using an Scx:Cre driver completely eliminates eminences (Blitz et al., 2009). Pulse-chase labeling studies show that eminences form from secondary fields of Scx/Sox9 co-expressing cells that lie immediately adjacent to major skeletal condensations (Fig. 6B) (Blitz et al., 2013); these cells form in mutants lacking BMP signaling, but do not differentiate. Early Sox9 expression is observed in both the skeletal condensation and the eminence, but the subset of these cells that express Scx are delayed in expression of Col2a1, thereby restricting chondrogenesis to the developing enthesis. Scx-expressing cells in developing entheses express Bmp4 and this is lost in tendons of Scx−/− mutant mice, which lack entheses. Thus, an attractive hypothesis is that Bmp4 secreted by tenocytes induces enthesis formation in adjacent skeletogenic mesenchyme (Fig. 6B). Interestingly, Sox9-expressing secondary fields of eminence progenitors still form in Bmpr1a−/− mutant mice but never differentiate, indicating a role for BMP signaling in maturation rather than specification. Taken together, these studies establish an early phase of eminence/enthesis development, which is independent of muscle development, and suggest that Scx-driven Bmp4 signaling non-autonomously regulates their formation (Fig. 6B) (Blitz et al., 2013; Murchison et al., 2007; Pryce et al., 2009). Such coordinated expression of Scx and Sox9 in tenocytes and chondrocytes is an emerging theme both during tendon development and in MSCs (Fig. 2A) (Asou et al., 2002; Soeda et al., 2010; Sugimoto et al., 2013). The close lineal relationship between tenocytes and chondrocytes/osteocytes (Fig. 2A) and their regulation at entheses also may help explain the ectopic ossification of tendons (see Box 2) that occurs normally in some species as well as during aging and disease (Magne and Bougault, 2015; Zhang and Wang, 2015).
Box 2. Ectopic tendon calcification
Ectopic ossification of tendons occurs normally in some species and is also observed during aging and in disease (Magne and Bougault, 2015; Zhang and Wang, 2015). Spondyloarthritis, for example, is an inflammatory enthesitis (Weinreb et al., 2014) that can lead to ectopic ossification spreading from the bone to the tendon or ligament. Ectopic ossification is also seen in calcifying tendinopathies, a common consequence of aging affecting as many as 1 in 5 adults over 50 years of age. Tendon ossification appears to be caused, in part, by tenocyte-dependent degradation of the tendon ECM (Magne and Bougault, 2015) as well as altered responses to mechanical forces and BMP/Smad signaling (Rui et al., 2013). The genetic inactivation of two small proteoglycans (Bgn and Fmod) of the ECM of cultured mouse TPCs leads to ectopic activation of BMP signaling and tendon ossification (Bi et al., 2007). Mechanical stretching of TPCs in vitro can also lead to BMP upregulation and abnormal ossification (Zhang and Wang, 2013, 2015). Surprisingly, BMP signaling oscillates in a circadian manner, and these cycles are deregulated in arrhythmic mutant mice, which correlates with increased tendon ossification (Yeung et al., 2014) Thus, tendons and ligaments are really tissues living on the edge with respect to their ECM composition and cellular constituents, presumably owing to their extreme responsiveness to feedback through mechanical force.
Finally, and not surprisingly, entheses are also extremely sensitive to mechanical forces. Mechanical forces have long been known to be important for the development of the bones to which muscles attach, as well as for the maintenance of skeletogenic cell populations (Shwartz et al., 2013). The paralysis of specific muscles leads to bone defects at sites of attachment and loss of joint progenitors (Kahn et al., 2009). The tendon ECM plays vital roles in these adaptations of muscles and bones to mechanical loading (Kjaer, 2004).
Conclusions and perspectives
Vertebrate tendons begin life similar to skeletal progenitors in the embryo, but rapidly establish unique identities and tissue organization, in large part through interactions with and assembly of the tendon ECM. This involves: (1) essential transcription factors expressed in tenocytes, such as Scx, Mkx and Egr1, that drive the expression of ECM proteins; (2) ECM components such as laminins that help establish muscle attachments in the absence of tenocytes and control tenocyte morphogenesis; (3) ECM components such as Tsp4 that drive the assembly of collagen fibrils at MTJs through their interactions with other ECM proteins and Itgs on muscle cell surfaces; and (4) the maintenance and repair of these ECM components in response to mechanical forces. In this Review, we have highlighted recent genetic studies that have provided insights into the molecular mechanisms underlying these different processes.
The discovery of Scx and Mkx has provided inroads into understanding the tendon/ligament gene regulatory program and revealed the close relationship between tenocytes and skeletal lineages. Scx is the earliest known marker of TPCs, and its overexpression transforms MSCs into tenocytes (Alberton et al., 2012), whereas Sox9 overexpression converts MSCs into cartilage (Fig. 2A) (Takimoto et al., 2012). However, unlike Sox9, which when eliminated leads to a failure to form cartilage, loss-of-function Scx or Mkx mutations in mice do not eliminate tenocytes, and mutants are viable although they have severe musculoskeletal deformities. This implies that tendon development involves multiple, partially redundant transcription factors and cell-cell signals, each playing a unique role in building and maintaining the complex network of ECM proteins at muscle attachments.
These studies also highlight the fact that the stability of muscle attachments is not pre-established during development but evolves through constant adaptation of the ECM to changing mechanical load. This occurs regionally within each tendon from its muscle origin to its bony insertion. Understanding the tendon/ligament gene regulatory program thus requires knowledge of how this dynamic network of ECM proteins self-assembles and feeds back upon transcriptional regulators in response to mechanical forces.
The musculoskeletal system and its associated ECM have also evolved to suit dramatically different modes of locomotion, feeding strategies and body sizes of different vertebrates. Changes in the interconnections between individual muscles and bones underlie many of the evolutionary differences between species. For example, mandibles of fish (Malawi cichlids), birds (Darwin's finches) and dogs can have very different functional morphologies depending on their feeding strategies, which notably have all been linked to changes in BMP signaling (Abzhanov et al., 2004; Albertson et al., 2005; Schoenebeck et al., 2012). These reflect coordinated changes in the regulation of cranial neural crest cells that form not only the craniofacial skeleton, but also cranial TPCs during evolution. Indeed, recent studies grafting neural crest cells between quail and duck embryos reveal that some distinct craniofacial morphologies have evolved through changes in cell-intrinsic mechanisms. Jaw cartilage differentiation, marked by Runx2 expression, occurs earlier in the duck embryo, whereas tendon differentiation, marked by Scx expression, occurs earlier in the quail (Tokita and Schneider, 2009). How these species-specific differences in timing of skeletal and tendon differentiation reflect changes in ECM organization and in mechanical stress to suit their adaptive functions is an important area for future investigation.
Understanding the genetic control of the development of tendons, their integration into the musculoskeletal system and their ECM organization also has important implications for regenerative medicine. Current efforts to treat tendon injuries or diseases focus either on ameliorating inflammation or driving MSCs towards the tenocyte fate with hopes of stem cell therapy (Yang et al., 2013). Both approaches leave out the crucial role of establishing the appropriate ECM for a specific tendon type and the loads that it needs to bear. Attaining the correct strength also relies on correct cellular responses to mechanical forces, which are transduced through the ECM. Thus, it is important to consider the ECM and its functions going forward in efforts to improve diagnosis and therapy design for tendon disorders.
Acknowledgements
We thank members of the Schilling lab and anonymous reviewers for critical reading of the manuscript.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Funding
The authors were supported by National Institutes of Health awards [R21 AR62792 and R01 DE13828 to T.F.S.]. Deposited in PMC for release after 12 months.
References
- Abzhanov A., Protas M., Grant B. R., Grant P. R. and Tabin C. J. (2004). Bmp4 and morphological variation of beaks in Darwin's finches. Science 305, 1462-1465. 10.1126/science.1098095 [DOI] [PubMed] [Google Scholar]
- Adams J. C. and Lawler J. (2004). The thrombospondins. Int. J. Biochem. Cell Biol. 36, 961-968. 10.1016/j.biocel.2004.01.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alberton P., Popov C., Prägert M., Kohler J., Shukunami C., Schieker M. and Docheva D. (2012). Conversion of human bone marrow-derived mesenchymal stem cells into tendon progenitor cells by ectopic expression of scleraxis. Stem Cells Dev. 21, 846-858. 10.1089/scd.2011.0150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albertson R. C., Streelman J. T., Kocher T. D. and Yelick P. C. (2005). Integration and evolution of the cichlid mandible: the molecular basis of alternate feeding strategies. Proc. Natl. Acad. Sci. USA 102, 16287-16292. 10.1073/pnas.0506649102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alliston T., Choy L., Ducy P., Karsenty G. and Derynck R. (2001). TGF-beta-induced repression of CBFA1 by Smad3 decreases cbfa1 and osteocalcin expression and inhibits osteoblast differentiation. EMBO J. 20, 2254-2272. 10.1093/emboj/20.9.2254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson D. M., Arredondo J., Hahn K., Valente G., Martin J. F., Wilson-Rawls J. and Rawls A. (2006). Mohawk is a novel homeobox gene expressed in the developing mouse embryo. Dev. Dyn. 235, 792-801. 10.1002/dvdy.20671 [DOI] [PubMed] [Google Scholar]
- Anderson D. M., Beres B. J., Wilson-Rawls J. and Rawls A. (2009). The homeobox gene Mohawk represses transcription by recruiting the sin3A/HDAC co-repressor complex. Dev. Dyn. 238, 572-580. 10.1002/dvdy.21873 [DOI] [PubMed] [Google Scholar]
- Anderson D. M., George R., Noyes M. B., Rowton M., Liu W., Jiang R., Wolfe S. A., Wilson-Rawls J. and Rawls A. (2012). Characterization of the DNA-binding properties of the Mohawk homeobox transcription factor. J. Biol. Chem. 287, 35351-35359. 10.1074/jbc.M112.399386 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aslan H., Kimelman-Bleich N., Pelled G. and Gazit D. (2008). Molecular targets for tendon neoformation. J. Clin. Invest. 118, 439-444. 10.1172/JCI33944 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asou Y., Nifuji A., Tsuji K., Shinomiya K., Olson E. N., Koopman P. and Noda M. (2002). Coordinated expression of scleraxis and Sox9 genes during embryonic development of tendons and cartilage. J. Orthop. Res. 20, 827-833. 10.1016/S0736-0266(01)00169-3 [DOI] [PubMed] [Google Scholar]
- Bader H. L., Keene D. R., Charvet B., Veit G., Driever W., Koch M. and Ruggiero F. (2009). Zebrafish collagen XII is present in embryonic connective tissue sheaths (fascia) and basement membranes. Matrix Biol. 28, 32-43. 10.1016/j.matbio.2008.09.580 [DOI] [PubMed] [Google Scholar]
- Bailey A. J., Paul R. G. and Knott L. (1998). Mechanisms of maturation and ageing of collagen. Mech. Ageing Dev. 106, 1-56. 10.1016/S0047-6374(98)00119-5 [DOI] [PubMed] [Google Scholar]
- Bajanca F., Luz M., Raymond K., Martins G. G., Sonnenberg A., Tajbakhsh S., Buckingham M. and Thorsteinsdóttir S. (2006). Integrin alpha6beta1-laminin interactions regulate early myotome formation in the mouse embryo. Development 133, 1635-1644. 10.1242/dev.02336 [DOI] [PubMed] [Google Scholar]
- Banos C. C., Thomas A. H. and Kuo C. K. (2008). Collagen fibrillogenesis in tendon development: current models and regulation of fibril assembly. Birth Defects Res. C. Embryo Today 84, 228-244. 10.1002/bdrc.20130 [DOI] [PubMed] [Google Scholar]
- Bassett D. I., Bryson-Richardson R. J., Daggett D. F., Gautier P., Keenan D. G. and Currie P. D. (2003). Dystrophin is required for the formation of stable muscle attachments in the zebrafish embryo. Development 130, 5851-5860. 10.1242/dev.00799 [DOI] [PubMed] [Google Scholar]
- Becker S., Pasca G., Strumpf D., Min L. and Volk T. (1997). Reciprocal signaling between Drosophila epidermal muscle attachment cells and their corresponding muscles. Development 124, 2615-2622. [DOI] [PubMed] [Google Scholar]
- Bedi A., Kovacevic D., Hettrich C., Gulotta L. V., Ehteshami J. R., Warren R. F. and Rodeo S. A. (2010). The effect of matrix metalloproteinase inhibition on tendon-to-bone healing in a rotator cuff repair model. J. Shoulder Elbow Surg. 19, 384-391. 10.1016/j.jse.2009.07.010 [DOI] [PubMed] [Google Scholar]
- Berthet E., Chen C., Butcher K., Schneider R. A., Alliston T. and Amirtharajah M. (2013). Smad3 binds Scleraxis and Mohawk and regulates tendon matrix organization. J. Orthop. Res. 31, 1475-1483. 10.1002/jor.22382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bi Y., Ehirchiou D., Kilts T. M., Inkson C. A., Embree M. C., Sonoyama W., Li L., Leet A. I., Seo B.-M., Zhang L. et al. (2007). Identification of tendon stem/progenitor cells and the role of the extracellular matrix in their niche. Nat. Med. 13, 1219-1227. 10.1038/nm1630 [DOI] [PubMed] [Google Scholar]
- Birch H. L., Thorpe C. T. and Rumian A. P. (2013). Specialization of extracellular matrix for function in tendons and ligaments. Muscles. Ligaments Tendons J. 3, 12-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blitz E., Viukov S., Sharir A., Shwartz Y., Galloway J. L., Pryce B. A., Johnson R. L., Tabin C. J., Schweitzer R. and Zelzer E. (2009). Bone ridge patterning during musculoskeletal assembly is mediated through SCX regulation of Bmp4 at the tendon-skeleton junction. Dev. Cell 17, 861-873. 10.1016/j.devcel.2009.10.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blitz E., Sharir A., Akiyama H. and Zelzer E. (2013). Tendon-bone attachment unit is formed modularly by a distinct pool of Scx- and Sox9-positive progenitors. Development 140, 2680-2690. 10.1242/dev.093906 [DOI] [PubMed] [Google Scholar]
- Bönnemann C. G. (2011). The collagen VI-related myopathies: muscle meets its matrix. Nat. Rev. Neurol. 7, 379-390. 10.1038/nrneurol.2011.81 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonnin M.-A., Laclef C., Blaise R., Eloy-Trinquet S., Relaix F., Maire P. and Duprez D. (2005). Six1 is not involved in limb tendon development, but is expressed in limb connective tissue under Shh regulation. Mech. Dev. 122, 573-585. 10.1016/j.mod.2004.11.005 [DOI] [PubMed] [Google Scholar]
- Bornstein P., Agah A. and Kyriakides T. R. (2004). The role of thrombospondins 1 and 2 in the regulation of cell–matrix interactions, collagen fibril formation, and the response to injury. Int. J. Biochem. Cell Biol. 36, 1115-1125. 10.1016/j.biocel.2004.01.012 [DOI] [PubMed] [Google Scholar]
- Brent A. E., Schweitzer R. and Tabin C. J. (2003). A somitic compartment of tendon progenitors. Cell 113, 235-248. 10.1016/S0092-8674(03)00268-X [DOI] [PubMed] [Google Scholar]
- Brent A. E., Braun T. and Tabin C. J. (2005). Genetic analysis of interactions between the somitic muscle, cartilage and tendon cell lineages during mouse development. Development 132, 515-528. 10.1242/dev.01605 [DOI] [PubMed] [Google Scholar]
- Bushby K. M. D., Collins J. and Hicks D. (2014). Collagen type VI myopathies. Adv. Exp. Med. Biol. 802, 185-199. 10.1007/978-94-007-7893-1_12 [DOI] [PubMed] [Google Scholar]
- Carmignac V. and Durbeej M. (2012). Cell-matrix interactions in muscle disease. J. Pathol. 226, 200-218. 10.1002/path.3020 [DOI] [PubMed] [Google Scholar]
- Chakravarti S. (2002). Functions of lumican and fibromodulin: lessons from knockout mice. Glycoconj. J. 19, 287-293. 10.1023/A:1025348417078 [DOI] [PubMed] [Google Scholar]
- Chanana B., Graf R., Koledachkina T., Pflanz R. and Vorbrüggen G. (2007). AlphaPS2 integrin-mediated muscle attachment in Drosophila requires the ECM protein Thrombospondin. Mech. Dev. 124, 463-475. 10.1016/j.mod.2007.03.005 [DOI] [PubMed] [Google Scholar]
- Charvet B., Malbouyres M., Pagnon-Minot A., Ruggiero F. and Le Guellec D. (2011). Development of the zebrafish myoseptum with emphasis on the myotendinous junction. Cell Tissue Res. 346, 439-449. 10.1007/s00441-011-1266-7 [DOI] [PubMed] [Google Scholar]
- Charvet B., Guiraud A., Malbouyres M., Zwolanek D., Guillon E., Bretaud S., Monnot C., Schulze J., Bader H. L., Allard B. et al. (2013). Knockdown of col22a1 gene in zebrafish induces a muscular dystrophy by disruption of the myotendinous junction. Development 140, 4602-4613. 10.1242/dev.096024 [DOI] [PubMed] [Google Scholar]
- Chen J. W. and Galloway J. L. (2014). The development of zebrafish tendon and ligament progenitors. Development 141, 2035-2045. 10.1242/dev.104067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y.-W., Zhao P., Borup R. and Hoffman E. P. (2000). Expression profiling in the muscular dystrophies: identification of novel aspects of molecular pathophysiology. J. Cell Biol. 151, 1321-1336. 10.1083/jcb.151.6.1321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., Yin Z., Chen J. I., Shen W. l., Liu H. H., Tang Q., Fang Z., Lu L. R., Ji J. and Ouyang H. W. (2012). Force and scleraxis synergistically promote the commitment of human ES cells derived MSCs to tenocytes. Sci. Rep. 2, 977 10.1038/srep00977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi W. J., Park M. S., Park K. H., Courneya J.-P., Cho J. S., Schon L. C. and Lee J. W. (2014). Comparative analysis of gene expression in normal and degenerative human tendon cells: effects of cyclic strain. Foot Ankle Int. 35, 1045-1056. 10.1177/1071100714540885 [DOI] [PubMed] [Google Scholar]
- Chuang H.-N., Hsiao K.-M., Chang H.-Y., Wu C.-C. and Pan H. (2014). The homeobox transcription factor Irxl1 negatively regulates MyoD expression and myoblast differentiation. FEBS J. 281, 2990-3003. 10.1111/febs.12837 [DOI] [PubMed] [Google Scholar]
- Corsi A., Xu T., Chen X.-D., Boyde A., Liang J., Mankani M., Sommer B., Iozzo R. V., Eichstetter I., Robey P. G., et al. (2002). Phenotypic effects of biglycan deficiency are linked to collagen fibril abnormalities, are synergized by decorin deficiency, and mimic Ehlers-Danlos-like changes in bone and other connective tissues. J. Bone Miner. Res. 17, 1180-1189. 10.1359/jbmr.2002.17.7.1180 [DOI] [PubMed] [Google Scholar]
- Davis M. E., Gumucio J. P., Sugg K. B., Bedi A. and Mendias C. L. (2013). MMP inhibition as a potential method to augment the healing of skeletal muscle and tendon extracellular matrix. J. Appl. Physiol. 115, 884-891. 10.1152/japplphysiol.00137.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Almeida F. M., Tomiosso T. C., Biancalana A., Mattiello-Rosa S. M., Vidal B. C., Gomes L. and Pimentel E. R. (2010). Effects of stretching on morphological and biochemical aspects of the extracellular matrix of the rat calcaneal tendon. Cell Tissue Res. 342, 97-105. 10.1007/s00441-010-1027-z [DOI] [PubMed] [Google Scholar]
- Docheva D., Popov C., Alberton P. and Aszodi A. (2014). Integrin signaling in skeletal development and function. Birth Defects Res. C Embryo Today Rev. 102, 13-36. 10.1002/bdrc.21059 [DOI] [PubMed] [Google Scholar]
- Dunkman A. A., Buckley M. R., Mienaltowski M. J., Adams S. M., Thomas S. J., Satchell L., Kumar A., Pathmanathan L., Beason D. P., Iozzo R. V. et al. (2013). Decorin expression is important for age-related changes in tendon structure and mechanical properties. Matrix Biol. 32, 3-13. 10.1016/j.matbio.2012.11.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durbeej M. (2010). Laminins. Cell Tissue Res. 339, 259-268. 10.1007/s00441-009-0838-2 [DOI] [PubMed] [Google Scholar]
- Eliasson P., Fahlgren A. and Aspenberg P. (2008). Mechanical load and BMP signaling during tendon repair: a role for follistatin? Clin. Orthop. Relat. Res. 466, 1592-1597. 10.1007/s11999-008-0253-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eliasson P., Andersson T. and Aspenberg P. (2009). Rat Achilles tendon healing: mechanical loading and gene expression. J. Appl. Physiol. 107, 399-407. 10.1152/japplphysiol.91563.2008 [DOI] [PubMed] [Google Scholar]
- Espira L., Lamoureux L., Jones S. C., Gerard R. D., Dixon I. M. C. and Czubryt M. P. (2009). The basic helix–loop–helix transcription factor scleraxis regulates fibroblast collagen synthesis. J. Mol. Cell. Cardiol. 47, 188-195. 10.1016/j.yjmcc.2009.03.024 [DOI] [PubMed] [Google Scholar]
- Farhat Y. M., Al-Maliki A. A., Chen T., Juneja S. C., Schwarz E. M., O'Keefe R. J. and Awad H. A. (2012). Gene expression analysis of the pleiotropic effects of TGF-β1 in an in vitro model of flexor tendon healing. PLoS ONE 7, e51411 10.1371/journal.pone.0051411 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farhat Y. M., Al-Maliki A. A., Easa A., O'Keefe R. J., Schwarz E. M. and Awad H. A. (2015). TGF-β1 suppresses plasmin and MMP activity in flexor tendon cells via PAI-1: implications for scarless flexor tendon repair. J. Cell Physiol. 230, 318-326. 10.1002/jcp.24707 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Font B., Eichenberger D., Rosenberg L. M. and Van Der Rest M. (1996). Characterization of the interactions of type XII collagen with two small proteoglycans from fetal bovine tendon, decorin and fibromodulin. Matrix Biol. 15, 341-348. 10.1016/S0945-053X(96)90137-7 [DOI] [PubMed] [Google Scholar]
- Frolova E. G., Drazba J., Krukovets I., Kostenko V., Blech L., Harry C., Vasanji A., Drumm C., Sul P., Jenniskens G. J. et al. (2014). Control of organization and function of muscle and tendon by thrombospondin-4. Matrix Biol. 37, 35-48. 10.1016/j.matbio.2014.02.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ge G. and Greenspan D. S. (2006). BMP1 controls TGFβ1 activation via cleavage of latent TGFβ-binding protein. J. Cell Biol. 175, 111-120. 10.1083/jcb.200606058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gotoh M., Mitsui Y., Shibata H., Yamada T., Shirachi I., Nakama K., Okawa T., Higuchi F. and Nagata K. (2013). Increased matrix metalloprotease-3 gene expression in ruptured rotator cuff tendons is associated with postoperative tendon retear. Knee Surg. Sports Traumatol. Arthrosc. 21, 1807-1812. 10.1007/s00167-012-2209-x [DOI] [PubMed] [Google Scholar]
- Grenier J., Teillet M.-A., Grifone R., Kelly R. G. and Duprez D. (2009). Relationship between neural crest cells and cranial mesoderm during head muscle development. PLoS ONE 4, e4381 10.1371/journal.pone.0004381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guerquin M.-J., Charvet B., Nourissat G., Havis E., Ronsin O., Bonnin M.-A., Ruggiu M., Olivera-Martinez I., Robert N., Lu Y. et al. (2013). Transcription factor EGR1 directs tendon differentiation and promotes tendon repair. J. Clin. Invest. 123, 3564-3576. 10.1172/JCI67521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauser N., Paulsson M., Kale A. and DiCesare P. (1995). Tendon extracellular matrix contains pentameric thrombospondin-4 (TSP-4). FEBS Lett. 368, 307-310. 10.1016/0014-5793(95)00675-Y [DOI] [PubMed] [Google Scholar]
- Havis E., Bonnin M.-A., Olivera-Martinez I., Nazaret N., Ruggiu M., Weibel J., Durand C., Guerquin M.-J., Bonod-Bidaud C., Ruggiero F. et al. (2014). Transcriptomic analysis of mouse limb tendon cells during development. Development 141, 3683-3696. 10.1242/dev.108654 [DOI] [PubMed] [Google Scholar]
- Heinemeier K. M., Skovgaard D., Bayer M. L., Qvortrup K., Kjaer A., Kjaer M., Magnusson S. P. and Kongsgaard M. (2012). Uphill running improves rat Achilles tendon tissue mechanical properties and alters gene expression without inducing pathological changes. J. Appl. Physiol. 113, 827-836. 10.1152/japplphysiol.00401.2012 [DOI] [PubMed] [Google Scholar]
- Henry C. A., McNulty I. M., Durst W. A., Munchel S. E. and Amacher S. L. (2005). Interactions between muscle fibers and segment boundaries in zebrafish. Dev. Biol. 287, 346-360. 10.1016/j.ydbio.2005.08.049 [DOI] [PubMed] [Google Scholar]
- Herchenhan A., Bayer M. L., Svensson R. B., Magnusson S. P. and Kjaer M. (2013). In vitro tendon tissue development from human fibroblasts demonstrates collagen fibril diameter growth associated with a rise in mechanical strength. Dev. Dyn. 242, 2-8. 10.1002/dvdy.23896 [DOI] [PubMed] [Google Scholar]
- Horiguchi M., Ota M. and Rifkin D. B. (2012). Matrix control of transforming growth factor-β function. J. Biochem. 152, 321-329. 10.1093/jb/mvs089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosokawa R., Oka K., Yamaza T., Iwata J., Urata M., Xu X., Bringas P., Nonaka K. and Chai Y. (2010). TGF-beta mediated FGF10 signaling in cranial neural crest cells controls development of myogenic progenitor cells through tissue–tissue interactions during tongue morphogenesis. Dev. Biol. 341, 186-195. 10.1016/j.ydbio.2010.02.030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang A. H., Lu H. H. and Schweitzer R. (2015). Molecular regulation of tendon cell fate during development. J. Orthop. Res. 33, 800-812. 10.1002/jor.22834 [DOI] [PubMed] [Google Scholar]
- Isogai Z., Ono R. N., Ushiro S., Keene D. R., Chen Y., Mazzieri R., Charbonneau N. L., Reinhardt D. P., Rifkin D. B. and Sakai L. Y. (2003). Latent transforming growth factor beta-binding protein 1 interacts with fibrillin and is a microfibril-associated protein. J. Biol. Chem. 278, 2750-2757. 10.1074/jbc.M209256200 [DOI] [PubMed] [Google Scholar]
- Ito Y., Toriuchi N., Yoshitaka T., Ueno-Kudoh H., Sato T., Yokoyama S., Nishida K., Akimoto T., Takahashi M., Miyaki S. et al. (2010). The Mohawk homeobox gene is a critical regulator of tendon differentiation. Proc. Natl. Acad. Sci. USA 107, 10538-10542. 10.1073/pnas.1000525107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jelinsky S. A., Archambault J., Li L. and Seeherman H. (2010). Tendon-selective genes identified from rat and human musculoskeletal tissues. J. Orthop. Res. 28, 289-297. [DOI] [PubMed] [Google Scholar]
- Jelinsky S. A., Rodeo S. A., Li J., Gulotta L. V., Archambault J. M. and Seeherman H. J. (2011). Regulation of gene expression in human tendinopathy. BMC Musculoskelet. Disord. 12, 86 10.1186/1471-2474-12-86 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones G. C., Corps A. N., Pennington C. J., Clark I. M., Edwards D. R., Bradley M. M., Hazleman B. L. and Riley G. P. (2006). Expression profiling of metalloproteinases and tissue inhibitors of metalloproteinases in normal and degenerate human achilles tendon. Arthritis Rheum. 54, 832-842. 10.1002/art.21672 [DOI] [PubMed] [Google Scholar]
- Juneja S. C. and Veillette C. (2013). Defects in tendon, ligament, and enthesis in response to genetic alterations in key proteoglycans and glycoproteins: a review. Arthritis 2013, 154812 10.1155/2013/154812 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahn J., Shwartz Y., Blitz E., Krief S., Sharir A., Breitel D. A., Rattenbach R., Relaix F., Maire P., Rountree R. B. et al. (2009). Muscle contraction is necessary to maintain joint progenitor cell fate. Dev. Cell 16, 734-743. 10.1016/j.devcel.2009.04.013 [DOI] [PubMed] [Google Scholar]
- Kang J. S., Alliston T., Delston R. and Derynck R. (2005). Repression of Runx2 function by TGF-beta through recruitment of class II histone deacetylases by Smad3. EMBO J. 24, 2543-2555. 10.1038/sj.emboj.7600729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kannus P. (2000). Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 10, 312-320. 10.1034/j.1600-0838.2000.010006312.x [DOI] [PubMed] [Google Scholar]
- Kannus P., Jozsa L., Järvinen T. A. H., Järvinen T. L. N., Kvist M., Natri A. and Järvinen M. (1998). Location and distribution of non-collagenous matrix proteins in musculoskeletal tissues of rat. Histochem. J. 30, 799-810. 10.1023/A:1003448106673 [DOI] [PubMed] [Google Scholar]
- Kardon G. (1998). Muscle and tendon morphogenesis in the avian hind limb. Development 125, 4019-4032. [DOI] [PubMed] [Google Scholar]
- Katzel E. B., Wolenski M., Loiselle A. E., Basile P., Flick L. M., Langstein H. N., Hilton M. J., Awad H. A., Hammert W. C. and O'Keefe R. J. (2011). Impact of Smad3 loss of function on scarring and adhesion formation during tendon healing. J. Orthop. Res. 29, 684-693. 10.1002/jor.21235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kieny M. and Chevallier A. (1979). Autonomy of tendon development in the embryonic chick wing. J. Embryol. Exp. Morphol. 49, 153-165. [PubMed] [Google Scholar]
- Kimura W., Machii M., Xue X., Sultana N., Hikosaka K., Sharkar M. T. K., Uezato T., Matsuda M., Koseki H. and Miura N. (2011). Irxl1 mutant mice show reduced tendon differentiation and no patterning defects in musculoskeletal system development. Genesis 49, 2-9. 10.1002/dvg.20688 [DOI] [PubMed] [Google Scholar]
- Kjaer M. (2004). Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol. Rev. 84, 649-698. 10.1152/physrev.00031.2003 [DOI] [PubMed] [Google Scholar]
- Koch M., Schulze J., Hansen U., Ashwodt T., Keene D. R., Brunken W. J., Burgeson R. E., Bruckner P. and Bruckner-Tuderman L. (2004). A novel marker of tissue junctions, collagen XXII. J. Biol. Chem. 279, 22514-22521. 10.1074/jbc.M400536200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koshida S., Kishimoto Y., Ustumi H., Shimizu T., Furutani-Seiki M., Kondoh H. and Takada S. (2005). Integrin alpha5-dependent fibronectin accumulation for maintenance of somite boundaries in zebrafish embryos. Dev. Cell 8, 587-598. 10.1016/j.devcel.2005.03.006 [DOI] [PubMed] [Google Scholar]
- Kruegel J. and Miosge N. (2010). Basement membrane components are key players in specialized extracellular matrices. Cell. Mol. Life Sci. 67, 2879-2895. 10.1007/s00018-010-0367-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyriakides T. R., Leach K. J., Hoffman A. S., Ratner B. D. and Bornstein P. (1999). Mice that lack the angiogenesis inhibitor, thrombospondin 2, mount an altered foreign body reaction characterized by increased vascularity. Proc. Natl. Acad. Sci. USA 96, 4449-4454. 10.1073/pnas.96.8.4449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Léjard V., Brideau G., Blais F., Salingcarnboriboon R., Wagner G., Roehrl M. H. A., Noda M., Duprez D., Houillier P. and Rossert J. (2007). Scleraxis and NFATc regulate the expression of the pro-alpha1(I) collagen gene in tendon fibroblasts. J. Biol. Chem. 282, 17665-17675. 10.1074/jbc.M610113200 [DOI] [PubMed] [Google Scholar]
- Lejard V., Blais F., Guerquin M.-J., Bonnet A., Bonnin M.-A., Havis E., Malbouyres M., Bidaud C. B., Maro G., Gilardi-Hebenstreit P. et al. (2011). EGR1 and EGR2 involvement in vertebrate tendon differentiation. J. Biol. Chem. 286, 5855-5867. 10.1074/jbc.M110.153106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y., Ramcharan M., Zhou Z., Leong D. J., Akinbiyi T., Majeska R. J. and Sun H. B. (2015). The role of scleraxis in fate determination of mesenchymal stem cells for tenocyte differentiation. Sci. Rep. 5, 13149 10.1038/srep13149 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Liu D., Black B. L. and Derynck R. (2001). TGF-β inhibits muscle differentiation through functional repression of myogenic transcription factors by Smad3. Genes Dev. 15, 2950-2966. 10.1101/gad.925901 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu W., Watson S. S., Lan Y., Keene D. R., Ovitt C. E., Liu H., Schweitzer R. and Jiang R. (2010). The atypical homeodomain transcription factor Mohawk controls tendon morphogenesis. Mol. Cell. Biol. 30, 4797-4807. 10.1128/MCB.00207-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C.-F., Aschbacher-Smith L., Barthelery N. J., Dyment N., Butler D. and Wylie C. (2012). Spatial and temporal expression of molecular markers and cell signals during normal development of the mouse patellar tendon. Tissue Eng. A 18, 598-608. 10.1089/ten.tea.2011.0338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu H., Zhu S., Zhang C., Lu P., Hu J., Yin Z., Ma Y., Chen X. and OuYang H. (2014). Crucial transcription factors in tendon development and differentiation: their potential for tendon regeneration. Cell Tissue Res. 356, 287-298. 10.1007/s00441-014-1834-8 [DOI] [PubMed] [Google Scholar]
- Liu H., Zhang C., Zhu S., Lu P., Zhu T., Gong X., Zhang Z., Hu J., Yin Z., Heng B. C. et al. (2015). Mohawk promotes the tenogenesis of mesenchymal stem cells through activation of the TGFβ signaling pathway. Stem Cells 33, 443-455. 10.1002/stem.1866 [DOI] [PubMed] [Google Scholar]
- Maeda E., Shelton J. C., Bader D. L. and Lee D. A. (2009). Differential regulation of gene expression in isolated tendon fascicles exposed to cyclic tensile strain in vitro. J. Appl. Physiol. 106, 506-512. 10.1152/japplphysiol.90981.2008 [DOI] [PubMed] [Google Scholar]
- Maeda E., Fleischmann C., Mein C. A., Shelton J. C., Bader D. L. and Lee D. A. (2010). Functional analysis of tenocytes gene expression in tendon fascicles subjected to cyclic tensile strain. Connect. Tissue Res. 51, 434-444. 10.3109/03008201003597056 [DOI] [PubMed] [Google Scholar]
- Maeda T., Sakabe T., Sunaga A., Sakai K., Rivera A. L., Keene D. R., Sasaki T., Stavnezer E., Iannotti J., Schweitzer R. et al. (2011). Conversion of mechanical force into TGF-β-mediated biochemical signals. Curr. Biol. 21, 933-941. 10.1016/j.cub.2011.04.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeda E., Ye S., Wang W., Bader D. L., Knight M. M. and Lee D. A. (2012). Gap junction permeability between tenocytes within tendon fascicles is suppressed by tensile loading. Biomech. Model. Mechanobiol. 11, 439-447. 10.1007/s10237-011-0323-1 [DOI] [PubMed] [Google Scholar]
- Maeda E., Sugimoto M. and Ohashi T. (2013). Cytoskeletal tension modulates MMP-1 gene expression from tenocytes on micropillar substrates. J. Biomech. 46, 991-997. 10.1016/j.jbiomech.2012.11.056 [DOI] [PubMed] [Google Scholar]
- Magne D. and Bougault C. (2015). What understanding tendon cell differentiation can teach us about pathological tendon ossification. Histol. Histopathol. 30, 901-910. [DOI] [PubMed] [Google Scholar]
- Marturano J. E., Xylas J. F., Sridharan G. V., Georgakoudi I. and Kuo C. K. (2014). Lysyl oxidase-mediated collagen crosslinks may be assessed as markers of functional properties of tendon tissue formation. Acta Biomater. 10, 1370-1379. 10.1016/j.actbio.2013.11.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer U., Saher G., Fässler R., Bornemann A., Echtermeyer F., von der Mark H., Miosge N., Pöschl E. and von der Mark K. (1997). Absence of integrin alpha 7 causes a novel form of muscular dystrophy. Nat. Genet. 17, 318-323. 10.1038/ng1197-318 [DOI] [PubMed] [Google Scholar]
- Munger J. S. and Sheppard D. (2011). Cross talk among TGF-β signaling pathways, integrins, and the extracellular matrix. Cold Spring Harb. Perspect. Biol. 3, a005017 10.1101/cshperspect.a005017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murchison N. D., Price B. A., Conner D. A., Keene D. R., Olson E. N., Tabin C. J. and Schweitzer R. (2007). Regulation of tendon differentiation by scleraxis distinguishes force-transmitting tendons from muscle-anchoring tendons. Development 134, 2697-2708. 10.1242/dev.001933 [DOI] [PubMed] [Google Scholar]
- Mustonen E., Ruskoaho H. and Rysä J. (2012). Thrombospondin-4, tumour necrosis factor-like weak inducer of apoptosis (TWEAK) and its receptor Fn14: Novel extracellular matrix modulating factors in cardiac remodelling. Ann. Med. 44, 793-804. 10.3109/07853890.2011.614635 [DOI] [PubMed] [Google Scholar]
- Noden D. M. (1988). Interactions and fates of avian craniofacial mesenchyme. Development 103 Suppl., 121-140. [DOI] [PubMed] [Google Scholar]
- Noden D. M. and Trainor P. A. (2005). Relations and interactions between cranial mesoderm and neural crest populations. J. Anat. 207, 575-601. 10.1111/j.1469-7580.2005.00473.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oka K., Oka S., Hosokawa R., Bringas P., Brockhoff H. C., Nonaka K. and Chai Y. (2008). TGF-beta mediated Dlx5 signaling plays a crucial role in osteo-chondroprogenitor cell lineage determination during mandible development. Dev. Biol. 321, 303-309. 10.1016/j.ydbio.2008.03.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otabe K., Nakahara H., Hasegawa A., Matsukawa T., Ayabe F., Onizuka N., Inui M., Takada S., Ito Y., Sekiya I. et al. (2015). Transcription factor Mohawk controls tenogenic differentiation of bone marrow mesenchymal stem cells in vitro and in vivo. J. Orthop. Res. 33, 1-8. 10.1002/jor.22750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan T.-C., Zhang R.-Z., Markova D., Arita M., Zhang Y., Bogdanovich S., Khurana T. S., Bönnemann C. G., Birk D. E. and Chu M.-L. (2013). COL6A3 protein deficiency in mice leads to muscle and tendon defects similar to human collagen VI congenital muscular dystrophy. J. Biol. Chem. 288, 14320-14331. 10.1074/jbc.M112.433078 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parkinson J., Samiric T., Ilic M. Z., Cook J. and Handley C. J. (2011). Involvement of proteoglycans in tendinopathy. J. Musculoskelet. Neuronal Interact. 11, 86-93. [PubMed] [Google Scholar]
- Parsons M. J., Pollard S. M., Saúde L., Feldman B., Coutinho P., Hirst E. M. A. and Stemple D. L. (2002). Zebrafish mutants identify an essential role for laminins in notochord formation. Development 129, 3137-3146. [DOI] [PubMed] [Google Scholar]
- Pingel J., Lu Y., Starborg T., Fredberg U., Langberg H., Nedergaard A., Weis M., Eyre D., Kjaer M. and Kadler K. E. (2014). 3-D ultrastructure and collagen composition of healthy and overloaded human tendon: evidence of tenocyte and matrix buckling. J. Anat. 224, 548-555. 10.1111/joa.12164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popov C., Burggraf M., Kreja L., Ignatius A., Schieker M. and Docheva D. (2015). Mechanical stimulation of human tendon stem/progenitor cells results in upregulation of matrix proteins, integrins and MMPs, and activation of p38 and ERK1/2 kinases. BMC Mol. Biol. 16, 1219 10.1186/s12867-015-0036-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pryce B. A., Watson S. S., Murchison N. D., Staverosky J. A., Dünker N. and Schweitzer R. (2009). Recruitment and maintenance of tendon progenitors by TGFbeta signaling are essential for tendon formation. Development 136, 1351-1361. 10.1242/dev.027342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rifkin D. B. (2005). Latent transforming growth factor-beta (TGF-beta) binding proteins: orchestrators of TGF-beta availability. J. Biol. Chem. 280, 7409-7412. 10.1074/jbc.R400029200 [DOI] [PubMed] [Google Scholar]
- Rooney J. E., Welser J. V., Dechert M. A., Flintoff-Dye N. L., Kaufman S. J. and Burkin D. J. (2006). Severe muscular dystrophy in mice that lack dystrophin and alpha7 integrin. J. Cell Sci. 119, 2185-2195. 10.1242/jcs.02952 [DOI] [PubMed] [Google Scholar]
- Rooney J. E., Knapp J. R., Hodges B. L., Wuebbles R. D. and Burkin D. J. (2012). Laminin-111 protein therapy reduces muscle pathology and improves viability of a mouse model of merosin-deficient congenital muscular dystrophy. Am. J. Pathol. 180, 1593-1602. 10.1016/j.ajpath.2011.12.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ros M. A., Rivero F. B., Hinchliffe J. R. and Hurle J. M. (1995). Immunohistological and ultrastructural study of the developing tendons of the avian foot. Anat. Embryol. 192, 483-496. 10.1007/BF00187179 [DOI] [PubMed] [Google Scholar]
- Rui Y. F., Lui P. P. Y., Wong Y. M., Tan Q. and Chan K. M. (2013). Altered fate of tendon-derived stem cells isolated from a failed tendon-healing animal model of tendinopathy. Stem Cells Dev. 22, 1076-1085. 10.1089/scd.2012.0555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rullman E., Norrbom J., Strömberg A., Wågsäter D., Rundqvist H., Haas T. and Gustafsson T. (2009). Endurance exercise activates matrix metalloproteinases in human skeletal muscle. J. Appl. Physiol. 106, 804-812. 10.1152/japplphysiol.90872.2008 [DOI] [PubMed] [Google Scholar]
- Schessl J., Zou Y. and Bönnemann C. G. (2006). Congenital muscular dystrophies and the extracellular matrix. Semin. Pediatr. Neurol. 13, 80-89. 10.1016/j.spen.2006.06.003 [DOI] [PubMed] [Google Scholar]
- Schoenebeck J. J., Hutchinson S. A., Byers A., Beale H. C., Carrington B., Faden D. L., Rimbault M., Decker B., Kidd J. M., Sood R. et al. (2012). Variation of BMP3 contributes to dog breed skull diversity. PLoS Genet. 8, e1002849 10.1371/journal.pgen.1002849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz A. G., Pasteris J. D., Genin G. M., Daulton T. L. and Thomopoulos S. (2012). Mineral distributions at the developing tendon enthesis. PLoS ONE 7, e48630 10.1371/journal.pone.0048630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz A. G., Lipner J. H., Pasteris J. D., Genin G. M. and Thomopoulos S. (2013). Muscle loading is necessary for the formation of a functional tendon enthesis. Bone 55, 44-51. 10.1016/j.bone.2013.03.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schweitzer R., Chyung J. H., Murtaugh L. C., Brent A. E., Rosen V., Olson E. N., Lassar A. and Tabin C. J. (2001). Analysis of the tendon cell fate using Scleraxis, a specific marker for tendons and ligaments. Development 128, 3855-3866. [DOI] [PubMed] [Google Scholar]
- Schweitzer R., Zelzer E. and Volk T. (2010). Connecting muscles to tendons: tendons and musculoskeletal development in flies and vertebrates. Development 137, 2807-2817. 10.1242/dev.047498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shukunami C., Takimoto A., Oro M. and Hiraki Y. (2006). Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev. Biol. 298, 234-247. 10.1016/j.ydbio.2006.06.036 [DOI] [PubMed] [Google Scholar]
- Shwartz Y., Blitz E. and Zelzer E. (2013). One load to rule them all: mechanical control of the musculoskeletal system in development and aging. Differentiation 86, 104-111. 10.1016/j.diff.2013.07.003 [DOI] [PubMed] [Google Scholar]
- Snow C. J. and Henry C. A. (2009). Dynamic formation of microenvironments at the myotendinous junction correlates with muscle fiber morphogenesis in zebrafish. Gene Expr. Patterns 9, 37-42. 10.1016/j.gep.2008.08.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snow C. J., Peterson M. T., Khalil A. and Henry C. A. (2008). Muscle development is disrupted in zebrafish embryos deficient for fibronectin. Dev. Dyn. 237, 2542-2553. 10.1002/dvdy.21670 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soeda T., Deng J. M., de Crombrugghe B., Behringer R. R., Nakamura T. and Akiyama H. (2010). Sox9-expressing precursors are the cellular origin of the cruciate ligament of the knee joint and the limb tendons. Genesis 48, 635-644. 10.1002/dvg.20667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spanoudes K., Gaspar D., Pandit A. and Zeugolis D. I. (2014). The biophysical, biochemical, and biological toolbox for tenogenic phenotype maintenance in vitro. Trends Biotechnol. 32, 474-482. 10.1016/j.tibtech.2014.06.009 [DOI] [PubMed] [Google Scholar]
- Subramanian A. and Schilling T. F. (2014). Thrombospondin-4 controls matrix assembly during development and repair of myotendinous junctions. eLife 2014, e02372 10.7554/elife.02372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian A., Wayburn B., Bunch T. and Volk T. (2007). Thrombospondin-mediated adhesion is essential for the formation of the myotendinous junction in Drosophila. Development 134, 1269-1278. 10.1242/dev.000406 [DOI] [PubMed] [Google Scholar]
- Sugimoto Y., Takimoto A., Akiyama H., Kist R., Scherer G., Nakamura T., Hiraki Y. and Shukunami C. (2013). Scx+/Sox9+ progenitors contribute to the establishment of the junction between cartilage and tendon/ligament. Development 140, 2280-2288. 10.1242/dev.096354 [DOI] [PubMed] [Google Scholar]
- Takimoto A., Oro M., Hiraki Y. and Shukunami C. (2012). Direct conversion of tenocytes into chondrocytes by Sox9. Exp. Cell Res. 318, 1492-1507. 10.1016/j.yexcr.2012.04.002 [DOI] [PubMed] [Google Scholar]
- Thomopoulos S., Williams G. R., Gimbel J. A., Favata M. and Soslowsky L. J. (2003). Variation of biomechanical, structural, and compositional properties along the tendon to bone insertion site. J. Orthop. Res. 21, 413-419. 10.1016/S0736-0266(03)0057-3 [DOI] [PubMed] [Google Scholar]
- Thorpe C. T., Birch H. L., Clegg P. D. and Screen H. R. C. (2013). The role of the non-collagenous matrix in tendon function. Int. J. Exp. Pathol. 94, 248-259. 10.1111/iep.12027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tokita M. and Schneider R. A. (2009). Developmental origins of species-specific muscle pattern. Dev. Biol. 331, 311-325. 10.1016/j.ydbio.2009.05.548 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomé F. M., Evangelista T., Leclerc A., Sunada Y., Manole E., Estournet B., Barois A., Campbell K. P. and Fardeau M. (1994). Congenital muscular dystrophy with merosin deficiency. C. R. Acad. Sci. III. 317, 351-357. [PubMed] [Google Scholar]
- Tozer S. and Duprez D. (2005). Tendon and ligament: development, repair and disease. Birth Defects Res. C. Embryo Today 75, 226-236. 10.1002/bdrc.20049 [DOI] [PubMed] [Google Scholar]
- Turner D. C., Lawton J., Dollenmeier P., Ehrismann R. and Chiquet M. (1983). Guidance of myogenic cell migration by oriented deposits of fibronectin. Dev. Biol. 95, 497-504. 10.1016/0012-1606(83)90052-0 [DOI] [PubMed] [Google Scholar]
- Vaz R., Martins G. G., Thorsteinsdóttir S. and Rodrigues G. (2012). Fibronectin promotes migration, alignment and fusion in an in vitro myoblast cell model. Cell Tissue Res. 348, 569-578. 10.1007/s00441-012-1364-1 [DOI] [PubMed] [Google Scholar]
- Veit G., Hansen U., Keene D. R., Bruckner P., Chiquet-Ehrismann R., Chiquet M. and Koch M. (2006). Collagen XII interacts with avian tenascin-X through its NC3 domain. J. Biol. Chem. 281, 27461-27470. 10.1074/jbc.M603147200 [DOI] [PubMed] [Google Scholar]
- Volk T. and VijayRaghavan K. (1994). A central role for epidermal segment border cells in the induction of muscle patterning in the Drosophila embryo. Development 120, 59-70. [DOI] [PubMed] [Google Scholar]
- Wälchli C., Koch M., Chiquet M., Odermatt B. F. and Trueb B. (1994). Tissue-specific expression of the fibril-associated collagens XII and XIV. J. Cell Sci. 107, 669-681. [DOI] [PubMed] [Google Scholar]
- Wall M. E. and Banes A. J. (2005). Early responses to mechanical load in tendon: Role for calcium signaling, gap junctions and intercellular communication. J. Musculoskelet. Neuronal Interact. 5, 70-84. [PubMed] [Google Scholar]
- Wang J. H.-C., Guo Q. and Li B. (2012). Tendon biomechanics and mechanobiology – a minireview of basic concepts and recent advancements. J. Hand Ther. 25, 133-141. 10.1016/j.jht.2011.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinreb J. H., Sheth C., Apostolakos J., McCarthy M. B., Barden B., Cote M. P. and Mazzocca A. D. (2014). Tendon structure, disease, and imaging. Muscles. Ligaments Tendons J. 4, 66-73. 10.11138/mltj/2014.4.1.066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willett T. L., Labow R. S., Aldous I. G., Avery N. C. and Lee J. M. (2010). Changes in collagen with aging maintain molecular stability after overload: evidence from an in vitro tendon model. J. Biomech. Eng. 132, 031002 10.1115/1.4000933 [DOI] [PubMed] [Google Scholar]
- Wipff P.-J., Rifkin D. B., Meister J.-J. and Hinz B. (2007). Myofibroblast contraction activates latent TGF-beta1 from the extracellular matrix. J. Cell Biol. 179, 1311-1323. 10.1083/jcb.200704042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang G., Rothrauff B. B. and Tuan R. S. (2013). Tendon and ligament regeneration and repair: clinical relevance and developmental paradigm. Birth Defects Res. C. Embryo Today 99, 203-222. 10.1002/bdrc.21041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeung C.-Y. C., Gossan N., Lu Y., Hughes A., Hensman J. J., Bayer M. L., Kjær M., Kadler K. E. and Meng Q.-J. (2014). Gremlin-2 is a BMP antagonist that is regulated by the circadian clock. Sci. Rep. 4, 5183 10.1038/srep05183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon J. H. and Halper J. (2005). Tendon proteoglycans: biochemistry and function. J. Musculoskelet. Neuronal Interact. 5, 22-34. [PubMed] [Google Scholar]
- Yu Q. and Stamenkovic I. (2000). Cell surface-localized matrix mealloproteinase-9 proteolytically activates TGF-beta and promotes tumor invasion and angiogenesis. Genes Dev. 14, 163-176. [PMC free article] [PubMed] [Google Scholar]
- Yurchenco P. D., Amenta P. S. and Patton B. L. (2004). Basement membrane assembly, stability and activities observed through a developmental lens. Matrix Biol. 22, 521-538. 10.1016/j.matbio.2003.10.006 [DOI] [PubMed] [Google Scholar]
- Zelzer E., Blitz E., Killian M. L. and Thomopoulos S. (2014). Tendon-to-bone attachment: from development to maturity. Birth Defects Res. C Embryo Today 102, 101-112. 10.1002/bdrc.21056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J. and Wang J. H. C. (2010). Mechanobiological response of tendon stem cells: implications of tendon homeostasis and pathogenesis of tendinopathy. J. Orthop. Res. 28, 639-643. [DOI] [PubMed] [Google Scholar]
- Zhang J. and Wang J. H.-C. (2013). The effects of mechanical loading on tendons – an in vivo and in vitro model study. PLoS ONE 8, e71740 10.1371/journal.pone.0071740 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J. and Wang J. H.-C. (2015). Moderate exercise mitigates the detrimental effects of aging on tendon stem cells. PLoS ONE 10, e0130454 10.1371/journal.pone.0130454 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang G., Ezura Y., Chervoneva I., Robinson P. S., Beason D. P., Carine E. T., Soslowsky L. J., Iozzo R. V. and Birk D. E. (2006). Decorin regulates assembly of collagen fibrils and acquisition of biomechanical properties during tendon development. J. Cell Biochem. 98, 1436-1449. 10.1002/jcb.20776 [DOI] [PubMed] [Google Scholar]
- Zhou B., Zhou Y. and Tang K. (2014). An overview of structure, mechanical properties, and treatment for age-related tendinopathy. J. Nutr. Health Aging 18, 441-448. 10.1007/s12603-014-0026-2 [DOI] [PubMed] [Google Scholar]
- Zou Y., Zwolanek D., Izu Y., Gandhy S., Schreiber G., Brockmann K., Devoto M., Tian Z., Hu Y., Veit G. et al. (2014). Recessive and dominant mutations in COL12A1 cause a novel EDS/myopathy overlap syndrome in humans and mice. Hum. Mol. Genet. 23, 2339-2352. 10.1093/hmg/ddt627 [DOI] [PMC free article] [PubMed] [Google Scholar]