

Abstract
The establishment of multipotent pancreas progenitors (MPP) should have a significant impact not only on the ontology of the pancreas, but also for the translational research of glucose-responding endocrine β-cells. Deficiency of the latter may lead to the pandemic type 1 or type 2 diabetes mellitus, a metabolic disorder. An ideal treatment of which would potentially be the replacement of destroyed or failed β-cells, by restoring function of endogenous pancreatic endocrine cells or by transplantation of donor islets or in vitro generated insulin-secreting cells. Thus, considerable research efforts have been devoted to identify MPP candidates in the pre- and post-natal pancreas for the endogenous neogenesis or regeneration of endocrine insulin-secreting cells. In order to advance this inconclusive but critical field, we here review the emerging concepts, recent literature and newest developments of potential MPP and propose measures that would assist its forward progression.
Keywords: Multipotent pancreas progenitors, Regeneration, Self-renewal, Clonogenesis, Differentiation
Core tip: Diabetes mellitus is a pandemic health problem that currently affects approximately 400 million people worldwide and its incidence is increasing by 2%-3% per year. At present, insulin deficiency in diabetes is treated by exogenous insulin given as either multiple daily injections or continuous subcutaneous infusion (pump), which is associated with acute, potentially life-threatening metabolic disturbances as well as chronic, vascular complications with significant morbidity and mortality. The ultimate solution would therefore be regenerative therapies by which lost β-cells in disease processes could be restored/replaced by surrogate insulin-secreting cells including those derived from multipotent pancreas progenitors.
INTRODUCTION
In contrast to what we would expect, modern humans encounter unprecedented health challenges, including the pandemic diabetes mellitus (DM), which is a major metabolic disorder worldwide with a progressively climbing incidence. This disease currently affects 387 million individuals, with one dying in every 7 s due to severe complications (http://www.idf.org/diabetesatlas). Among them, approximately 10% suffer from type 1 DM (T1DM), due to the absolute lack of glucose-responding β-cells destroyed by the patient’s own immune system. Provided autodestruction of β-cells is under control, a permanent replacement approach may therefore be an ideal solution for T1DM, through regeneration in situ of endogenous β-cells, or by the replacement with donated glucose-responding islets, or of in vitro produced insulin-secreting β-cells from stem cell sources such as pluripotent stem cells (PSCs). The other 90% diabetic subjects are currently affected by type 2 DM (T2DM), resulting from the inability to react to insulin regulation by key metabolic tissues, the inability to regulate the generation of glucose from the liver and the dysfunction of endocrine β-cells[1]. The latter is typically believed to be cause by the increase of β-cell death[2]. Recently, accumulating evidence has suggested that β-cell dysfunction in T2DM is also caused by the dedifferentiation of glucose-regulating β-cells[3,4]. Thus, the ability to restore function of failed endocrine cells would provide a novel redifferentiative treatment for T2DM. In order to develop regenerative medicine therapies to T1DM and T2DM, the interest to both stem cells and progenitors in the pancreas has recently been progressively increased.
In general, the concept of multipotent progenitors is exchangeable with that of stem cells. The latter are usually used to define undifferentiated primitive cells that have the capacity for self-renewal themselves as well as of differentiation into terminal functional cells. For example embryonic stem cells (ESC) and induced pluripotent stem cells (iPSC, both together known as PSC) are able to produce all types of 210 cells that build up the body. Debatably, progenitors with multipotency are stem cells. Progenitors could be multipotent, oligopotent or unipotent based on their developmental potentials. Multipotent progenitors/stem cells are of fundamental importance to normal physiology and to regeneration during disease/injury repair. Multipotent pancreas progenitors (MPP) would be a rare subset present in developing and adult pancreas, and have a capacity for regeneration when required, even though their location and origin have not yet been completely established and are still controversial. In this article, we will summarize knowledge on the candidate MPP along the natural route of endocrine pancreas development and in three functional components of the pancreas. We will also propose future research perspectives on the potential MPP.
CLASSICAL EMBRYOLOGY
The pancreas is an important digestive and endocrine organ originating from the endoderm of posterior foregut. The latter along the other two germ layers is derived from the inner cell mass and would develop into the pancreas primordia when a localized area of thickened columnar epithelia along the dorsal and ventral posterior foregut appears. The columnar endodermal layer evaginates into the neighboring mesenchymal tissues of mesoderm origin and gives rise to the dorsal and ventral buds of the pancreas, respectively. Whilst continuously proliferating and branching, two buds gradually fuse together due to the gut rotation movement. Subsequently, the primordial pancreas continue to expand, transform and, finally, differentiate into the mature organ. The mature pancreas is composed of the acinar compartment that secretes digestive enzymes and fluid, that are drained into the intestinal lumen by the pancreatic ducts and, the endocrine pancreas that secrete hormones (glucagon, insulin, somatostatin, ghrelin or pancreatic polypeptide) responsible for maintaining normal glucose metabolism.
The key developmental events in the human pancreas are different from what we know about from developing pancreas in mice[5]. For instance, human dorsal bud is detectable at 26 d post conception (dpc), which is an equivalent age of embryonic day (E)9.5 in mouse embryos. The cellular developmental sequences in humans also differ from that in mice. Although they are not visible until 52 dpc, approximately 2 wk later than the equivalent stage at which they can be detected in mice, insulin-positive β-cells in humans appear precedent, unlike that in mice, to glucagon-positive α-cells at 8-10 wk of gestation[6]. All islet cells are detectable at the end of the first trimester in humans[6], but at later stages (E17.5) in mice[7]. Finally, the dynamics of gene expression during embryonic development and in pathophysiological conditions also differ between the two species[8]. Readers are referred to more reviews of human pancreas development elsewhere[9-12].
POTENTIAL MPP ALONG THE DEVELOPMENT OF ENDOCRINE PANCREAS
Remarkably, PSC can be guided to differentiate into definitive endoderm (DE)-like progenitors ex vivo by applying knowledge of in vivo developmental mechanisms (Figure 1). For example, this has been achieved in culture by supplementing with a pharmacological level of activin A, a growth factor (GF) of the superfamily of the transforming GF β. Furthermore, human PSC-derived self-renewable DE-like cells have been reported to expand under the stimuli of four GFs[13]. These DE-like endodermal progenitors have proliferated over 24 passages with an astonishing increase in cell numbers. To establish their reliability and utility in developmental biology, drug screening and regenerative medicine, PSC-derived DE-like cells should be biologically and transcriptomically compared to embryo-derived DE cells. Although further research is required, these endodermal progenitors may indeed function as pre-MPP in addition to possessing an incredible capacity for expansion.
Figure 1.
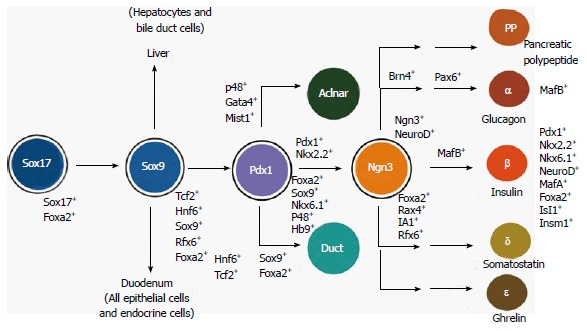
Multiple fate selections allow the development of the pancreas islet lineages. Committed from one of three germ layers (the ectoderm, mesoderm and endoderm) during gastrulation, the definitive endodermal cells (DE) are marked by the expression of Sox17 (the Sry-related HMG box transcription factor 17) and foxhead homeobox 2a (Foxa2). Along the anterior-posterior axis the DE is divided into foregut (giving rise to the lung, thyroid and esophagus), posterior foregut (PF), marked by the expression of the transcription factor hepatocyte nuclear factor 4a (Hnf4a) and hindgut (committing the intestine and colon). In vitro, retinoid acid would direct the DE cells to PF cells. Largely to the liver and duodenum, a fraction of the PF cells give rise to pancreatic progenitors (PP, marked by the expression of the transcription factor Pdx1). Mostly to the exocrine and ductal tissues, the PP commits to progenitors of the endocrine islet lineages [IP, marked by the expression of Neurog3, as well as neural differentiation 1 (NeuroD ), insulinoma associated 1 (IA1), Islet 1 (Isl1), paired box factor 6 (Pax6) and Rfx6]. The IP then differentiates into at least five types of islet cells [α, β, δ (somatostatin), pancreatic polypeptide (PP) and ε (ghrelin)].
Sox9-expressing MPP
Cystic epithelial colonies expressing E-cadherin first were generated by our group (Figure 2). After dissociation, fetal pancreatic cells produced cystic colonies containing β-cells in the presence of the basement membrane glycoprotein laminin 1, 1, 1 and a bone morphogenetic protein[14,15]. Interestingly, spherical but not cystic colonies are generated in the presence of epidermal GF[16], suggesting these GFs play different roles during development. We have not investigated whether all or only some fractions of cystoid epithelial cells stochastically commits to different lineages, though it seems possible that these cystic colonies originated from multipotent progenitors that are Sox9+ (see below).
Figure 2.
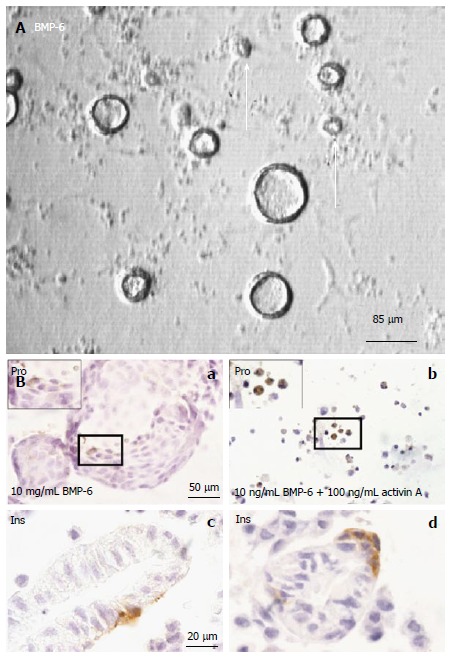
Cystic colony formation from dissociated fetal mouse pancreas cells. A: Phase contrast image showing that BMP-6 promotes colony formation. Open arrows indicate colonies ≤ 30 μm; B: Immunocytochemical analyses: a: Proinsulin staining. Fixed colonies were stained with proinsulin antibody (brown); b: Activin A antagonizes colony formation; c, d: Insulin staining. Histological sections of harvested colonies were stained with anti-insulin antibody (brown). Adapted and modified from ref.[14].
Sox genes encode versatile regulators of stem/progenitor cell fate[17], belonging to members of transcription factor family that contain the Sry (sex determining region Y) box-related high-mobility group. Sox9 for example critically regulates the development of multiple embryonic organs including the pancreas. This pivotal transcription factor is first detectable at E10.5 in the dorsal and ventral pancreases[18] and at E13.5, Sox9-expressing progenitors (Figure 3) develop into the exocrine and endocrine lineages in the pancreas[19]. However, by E16.5 the expression of Sox9 is progressively restricted to pancreatic ductal cells[19]. Genetic tracing studies reveal that Sox9 is also expressed in organs of other posterior foregut origin such as the liver, the bile duct and the duodenum. For example, it is present in bile ductal cells adjacent to the portal vein from E16.5. Sox9 also is broadly detectable in the intestinal epithelia at E13.5 but confined to the crypt region from E18.5[19]. Thus All Sox9-expressing cells in the posterior foregut region could be MPP. Supportive to these analyses, Sox9-expressing (Sox9+) multipotent progenitors purified from E11.5 Sox9-eGFP embryos generate expandable cystoid colonies that contain hormone-expressing cells in a laminin 1, 1, 1-enriched Matrigel-coated culture condition[20].
Figure 3.
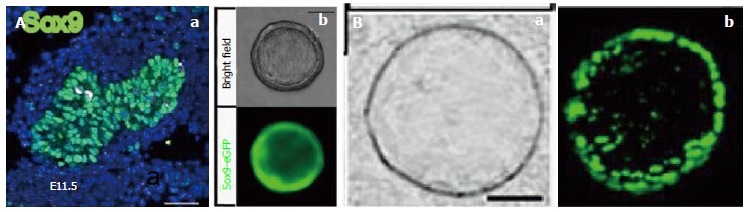
Cystic colonies generated from Sox9-tagged cells. A: Embryonic Sox9+ progenitors in the pancreas capable of generating cystic colonies; a: Sox9 is expressed in most ductal progenitors in E11.5 mouse pancreas; b: Cystic colonies are formed from purified Sox9-eGFP+ progenitors in E11.5 mouse pancreas; B: Purified Sox9+ cells in adult mouse pancreas capable of generating cystic colonies under a phase contrast (a) or a fluorescence microscope (b). Adapted and modified from[20,21].
Sox9+CD133+ ductal cells generates duct-like “ring/dense” colonies (1/5) in the culture of the Matrigel-containing methylcellulose-based semisolid medium. With the addition of the roof plate-specific spondin 1, a Wnt agonist, these ring/dense colony-forming cells can be serially dissociated and replated in the presence of Matrigel with an expansion of more than 100000-fold[21]. In a laminin-containing hydrogel, the Sox9+CD133+ (cluster differentiation 133) cells are able to give rise to acinar/endocrine colonies[21]. Further investigations are needed to ascertain whether Sox9+CD133+ cells are identical to, or distinct from, the colony-forming Lgr5 [leucine-rich repeat-containing G-protein coupled receptor 5, also known as GPR49 or GPR67 (G-protein coupled receptor 49 or 67)]-expressing cells described below. However, Sox9-positive ductal and centroacinar cells are unable to produce β-cells in a few experiments for regeneration or trans differentiation, such as with cerulean-induced acute pancreatitis, an in vitro culture experiment, pancreatic duct ligation (PDL), partial pancreatectomy and a streptozotocin-induced diabetic model[19,22]. Originated from the posterior foregut endoderm, a human bile duct progenitor population known as “biliary tree-derived cells”[23] may have a MPP potential and give rise to islet lineages. However, the purified population, molecular profile and detailed developmental potential of these “tree-derived cells” require further investigations.
Pdx1-expressing MPP
A cluster of unique cells in the thickened DE epithelium along the dorsal and ventral surfaces of the posterior foregut in E9.0-9.5 mouse embryos expresses the homeobox gene named Pdx1 (pancreas and duodenum transcription factor 1). The latter encodes a parahox homeobox-containing factor, critical for the establishment of primitive pancreas[24] as well as the maintenance of functional β-cells[25,26].
Studies have demonstrated that Pdx1-expressing (Pdx1+) progenitors are MPP because they give rise to acinar, ductal and islet tissues of the pancreas[27]. Genetic lineage tracing studies revealed that these MPP reside in the termini of the tree-like branching ducts in the developing pancreas that also express the acinar transcription factor known as Ptf1a and the exocrine cell marker known as Cpa1[28]. The Pdx1+ cells are capable of taking up the thymidine analogue bromodeoxyuridine (BrdU) and incorporating the latter into their genome during mitosis[29], revealing that these cells are proliferative.
Using developmental biological knowledge, PSC have been manipulated to generate Pdx1+ cells that have been expanded for 16-fold co-cultured with pancreas-derived mesenchymal cells[30]. To verify the proliferation of Pdx1+ cells, independent confirmation of this report will be required. We also need to ascertain whether these PSC-derived Pdx1+ cells share all or partial characteristics of embryonic pancreatic progenitors, because Pdx1 is also expressed in extra-pancreatic tissues including other endoderm-derived non-pancreas organs[31].
In developing human pancreas, numerous PDX1+ progenitors are present between 8 and 21 wk of gestation[32,33]. These PDX1+ progenitors are frequently expressed SOX9 and highly proliferative[34], supporting the notion that pancreatic progenitors are committed from SOX9+ multipotent progenitors. The progressive increase of PDX1+ cells that co-express insulin or somatostatin is observed in this period of gestation[32]. Further research is required to investigate whether the PDX1+ progenitor pool is established by its self-renewal or by direct differentiation of the posterior foregut progenitors or both.
Strategies for proliferation, self-renewal and differentiation of pancreatic progenitors will be of importance in developmental biology and regenerative medicine. Interestingly, reserpine and tetrabenazine that inhibit the vesicular monoamine transporter-2 are shown to direct PSC-derived Pdx1+ cells to produce cells that express neurogenin 3 (Neurog3, also known as Ngn3)[35], a DNA-binding protein, belonging to the transcription factor family of the basic helix-loop-helix category. However, as a positive control, Pdx1+ progenitors purified from embryonic pancreas would have been tested with these two molecules to examine their capacity and efficiency to give rise to islet progenitors. Furthermore, caution needs to be exercised for the use of genetic lineage tracing in PSC differentiation in vitro because temporospatial cues are essential for the success of in vivo lineage tracing studies (see review by[36]). To enrich PSC-derived genuine pancreatic progenitors for further expansion and differentiation, identification of their specific markers would be highly valuable.
Neurog3-expressing pancreatic endocrine progenitors
Approximately in E9.5 mouse embryos, within the thickened posterior foregut DE epithelium there is a small cluster of cells that express a high level of the Neurog3, an endocrine determinant[27,37,38]. The cells that express Neurog3 highly are the progenitors of endocrine pancreas as they develop into all islet lineage cells in vivo. Several studies verify the critical role of Neurog3 in the ontogenesis of endocrine pancreas: Islet cells are not observed in Neurog3 targeted mouse pancreas[37]; genetic tracing demonstrates that Neurog3+ progenitors differentiate into all five types of islet cells[27]; isolated adult Neurog3+ cells reappeared after PDL can, after inoculation into an embryonic pancreas ex vivo, give rise to five types of endocrine cells[38]. Nevertheless, a few Neurog3+ cells are observed to coexpress insulin in the dual fluorescence-tagged developing mouse pancreas[39]. Another laboratory reported that PDL allows the activation of Neurog3 expression but the Neurog3-expressing cells are not able to complete the entire β-cell regeneration program[22]. Furthermore a recent study found that β-cell mass and insulin content are totally unchanged after PDL-induced injury[40], unsupportive to the conclusion of an active β-cell regeneration. The reason for these inconsistencies is unknown and future investigation is warranted to confirm or refute the conclusion.
Whereas the expression of mouse Neurog3 mRNA in the developing pancreas plateaus approximately E15.5[41] (approximately week 9 of gestation in humans), that of human NEUROG3 is low prior to 9 wk, but increases sharply onward and remains very high until 17 wk[33]. Furthermore some cells coexpress both NEUROG3 and insulin in the embryonic pancreas from 10 to 21 wk[32]. Although earlier studies showed that Neurog3+ cells could proliferate[42,43], clonal assays by “mosaic analysis with double marker” (MADM) have confirmed that Neurog3+ cells are quiescent and commit to only one cell type of the endocrine pancreas[44]. A recent study consistently demonstrated that the activation of Neurog3 itself inhibits cell division by the activation of cyclin-dependent kinase inhibitor 1a[45]. The inconsistencies between the previous and recent reports require future research to reconcile. Again, it is formally possible that PSC-derived Neurog3-expressing cells are not completely bona fide islet progenitors as Neurog3 is expressed in endoderm-derived non-pancreas organs.
Insulin+ multipotent progenitors
Embryonic insulin-expressing cells may be endocrine progenitors in the pancreas as they can give rise to other islet cell types in addition to β-cells at least during development[46]. By employing fluorescence-tagged tracing strategies, multipotent progenitors that are insulin+ (arguably to be MPP) in the pancreatic tissues are believed to be originated from the Pdx1+ progenitors, instead from the ectoderm-derived neural crest. These MPP-like cells express several islet progenitor markers and are able to differentiate into all endocrine cells in vivo. The MPP express a low concentration of insulin and low copies or complete lack of glucose transporter-2, clearly different from functional mature β-cells[47]. Nevertheless, studies have demonstrated that the expression of insulin gene is activated in Pdx1+ MPP, via Neurog3+ precursors to adult β-cells[48,49], thus future investigation is needed to establish what stages at which insulin+ MPP are present: The embryonic, fetal and/or adult endocrine pancreas.
Human insulin+ multipotent progenitors have also been described with a similar developmental potential to the mouse ones. These cells, isolated from either mice or humans, could, after transplantation into diabetic mice, develop into functional cells to ameliorate hyperglycemia. Surprisingly, however, these insulin+ multipotent progenitors also gave rise to neural lineage cells in vivo[47].
Ghrelin (ε)-expressing progenitors?
Ghrelin-expressing ε-cells are the fifth cell type in the endocrine pancreas and are first discovered in the stomach. Ghrelin is a polypeptide hormone composed of 28-amino acid residuals and known to negatively regulate insulin secretion from mouse, rat and human islets[50]. Now it is well established that pancreaticε-cells are detectable in mid-gestation in mice and humans and their number plateaus during the perinatal period[51]. However, there are a substantial number of ε-cells in only human but not mouse and rat adult islets[51], raising the possibility thatε-cells could participate in the regulation of glucose homeostasis.
Deletion of the Arx gene, encoding an α-cell transcription factor, drastically reduces the number of ε-cells[52]. In contrast, knockout of Nkx2.2, Pax4 or Pax6 significantly increases the number of these cells at the expense of reducing other pancreatic endocrine cell types[53,54]. Intriguingly, genetic studies of lineage tracing demonstrated that ε-cells give rise to α, PP and, to a lesser extent, β-cells in adult mouse pancreas[55], suggesting that these cells have a unusual plasticity for trans differentiation towards, and may even be progenitors of, other islet cells. However, whether ε-cells would act as MPP of functional islets remains to be established.
Can islet β-cells behave like unipotent “progenitors”?
Functional duplication of insulin-secreting β-cells is first described by Dor et al[56] by using RIP (rat insulin promoter)-driving genetic lineage tracing. They revealed that mouse β-cells in the endocrine pancreas could be reproduced by RIP-expressing cells under the physiological condition or after partial pancreatectomy. This report however did not preclude the presence of and the role of MPP by assuming that all RIP-expressing cells are functional β-cells. Similarly, in a transgenic model using the RIP to govern diphtheria toxin expression resulted in 70%-80% β-cell apoptosis, disrupted architecture of endocrine pancreas and eventually in the development of diabetes. Withdrawing the expression of diphtheria resulted in a significant recovery of β-cell mass, islet architecture and of normoglycemia[57]. Further analysis revealed that a subset of 20%-30% surviving “β”-cells played a major role in the β-cell regeneration and recovery of euglycemia[57]. These studies suggest that islet β-cells are indeed facultative unipotent progenitor cells.
In the sophisticated double transgenic MADM (designated RIP-CreER; Rosa26GR/Rosa26RG) mouse pancreas, each RIP-expressing clone is made up of only 5 cells after one month of chase, slightly increasing to 8 cells by two months[58]. The clusters that express RIP have been viewed as supportive evidence of regeneration of functional cells, but this should be treated with caution as discussed hereafter. A further study with a reductionist approach on the gene Hnf4a (hepatocyte nuclear factor 4a) suggested that the β-cell regeneration may be associated with the signaling cascade of the Ras/Erk pathway[59] and even be modulated by cell cycle regulators such as cyclin D2[60]. Collectively, it is critical to establish and identify the well-known RIP-expressing cells either in self-duplication or in dedifferentiation both in situ and ex vivo, as the latter may be a key MPP candidate for a T1DM replacement solution.
Furthermore, lineage tracing labeled with a thymidine analogue showed that β-cells are produced within an islet by rare self-renewable cells that have a long duplication-refractory time. Under stress conditions such as during pregnancy or after partial pancreatectomy, the number of self-renewable cells is dramatically elevated[61]. However the molecular nature of these rare cells and the replication-refractory length have not been established. Future studies should characterize their molecular identity and reveal whether these self-renewable replicating cells are fully functional. Due to the ethical barrier and technical difficulties, similar studies are not possible to be undertaken to label human endocrine pancreas in vivo, but a similar research should be repeated with larger mammals.
The above described investigations of β-cell self-renewal as a regenerative mechanism have drawn considerable interest in last several years, due to its promise as a way to increase β-cell mass for the treatment of diabetes. However, these studies have not excluded whether the insulin transcript (as controlled, for instance, by the RIP transgene) is only expressed in functional β-cells. There is increasing evidence suggesting that is not the case. First, the expression of insulin gene is detectable in the Pdx1+ progenitors, dramatically increasing in Neurog3+ precursors and peaking in mature islet β-cells[48,49]. Second, the demonstration of insulin+ multipotent progenitors[47] precludes insulin as an exclusive marker of functional β-cells. Third, insulin protein has been detectable in some mouse and human islet precursors as described above.
Taken together, the expression of insulin gene is clearly not exclusive for functional β-cells. It is formally possible β-cell populations in adults are maintained not only by the self-replication of functional glucose-regulating cells but also by the self-duplication and development of MPP. In order to demonstrate that MPP are indeed present in the adult endocrine pancreas, their clonogenesis must be established with isolated single cell MPP candidates ex vivo, with data generated from the intermediary stages of the clonogenesis, and with demonstrable ability to give rise to at least non-β endocrine cells in vivo.
MPP PRESENT IN THREE TYPES OF PANCREAS TISSUES
Accumulating evidence generated in ex vivo studies has suggested that MPP are present in three major tissues of the pancreas: The pancreatic ductal[62-64], acinar and islet compartments[65,66]. For instance, the application of flow cytometry has identified a potential MPP population in the mouse pancreas both in development and in adults[67]. These candidates are characterized by the exhibition of the receptor c-met for hepatocyte GF, without the presence of hematopoietic lineage markers including CD45, c-Kit (stem cell factor receptor), Flk-1 and TER119. The purified population is able to give rise to several types of pancreatic cells ex vivo and generate pancreatic endocrine and exocrine cells in situ after transplantation[67]. Nevertheless, the spatial localization and the molecular natures of these c-met positive cells are completely unknown and the clonogenesis has not been established at the single cell level.
MPP likely present in the ductal epithelium
Many studies suggest that MPP are present in the ductal epithelium. Bonner-Weir et al[68] first reported that human ductal epithelial cells in adult pancreas are able to differentiate into islet-like clusters containing insulin-secreting β-cells. Ramiya et al[63] showed that insulin-secreting islet-like clusters generated in vitro from mouse MPP are capable of ameliorating diabetes after being grafted under the kidney capsule. In cultures of human “pancreatic ductal cell aggregates” after isolation of the islets for transplantation, cells with fibroblast-like morphology appear known as pancreatic “mesenchymal stem cells (MSC)”. These cells were passaged over 12 times and expressed an array of markers of bone marrow MSC such as CD13, CD29, CD44, CD49f (also known as α6 integrin subunit), CD54, CD90 (also known as Thy1) and CD105. The pancreatic MSC are shown to be able to give rise to cross-germ layer cells such as endoderm-originated non-pancreas lineages[69]. Utilizing culture protocols suitable for producing the neurospheres of ectoderm origin, ductal cells from mouse pancreas are shown to give rise to neurosphere-like clusters that subsequently give rise to a few types of endocrine cells, such as insulin-secreting β cells[66]. Nevertheless, the exact nature of the special cells has not been established. A major limitation of the forementioned reports is the use of unenriched/unpurified cell preparations in addition to not demonstrating single-cell self-renewability.
Furthermore, numerous CK19+ ductal cells and islet cells are detected after PDL[38]. The lineage relationship between the ductal cells and islet cells has further been addressed using the genetic lineage tracing of the Cre-loxP system. Using the system, the Cre governs the promoter activity of carbonic anhydrase II gene, encoding a marker of adult ductal epithelia and controls the removal of the stop sequence (Rosa-loxP-stop-loxP-lacZ) in the reporter Rosa26 (R26R) mice. This resulted in Cre-driving β-galactosidase expression in ductal epithelia. After 28 d in normal or PDL pancreas, the activity of β-galactosidase is detectable in numerous ductal epithelia, localized acinar tissue and in up to 40% islet cells[70,71]. These data have been viewed as further evidence that the ductal cells expressing carbonic anhydrase II are able to regenerate mouse endocrine cells. It remains unclear, nevertheless, how many cells that express carbonic anhydrase II have this regenerative capacity, how many potential MPP are present or whether a trans differentiation process also takes place.
Nevertheless, once an exon of the ductal epithelial marker gene Tcf2 (T cell factor 2, also known as Hnf1b, hepatocyte nuclear factor 1b) is exchanged with the transgene containing Cre, the lineage tracing has demonstrated that the Tcf2+ cells in the postnatal duct cannot develop into endocrine cells in both normal and PDL pancreas[72]. However, a complicating factor in this report is that one allele of Tcf2 was non-functional, leading to reducing by half the transcription of Tcf2. As heterozygous Tcf2 mutant does not support pancreas development in mice and humans[73,74], inactivation of one allele of Tcf2 in the lineage tracing studies may have led to haploinsufficiency and affected the differentiation of potential MPP into functional islet cells.
Fbw7 (F-box and WD-40 domain protein 7), an ubiquitin ligase, is expressed in embryonic and adult ductal epithelial cells. Deletion of this gene stabilizes the heavily ubiquitinated Neurog3, and reprograms the ductal cells to α, β and somatostatin-producing δ-cells[75]. This study suggests that pancreatic ductal cells are a latent MPP and Fbw7 is a critical cell-fate regulator. Nevertheless, this report did not describe whether all or a small fraction of the ductal cells express Fbw7 and what the frequency of the reprogramming event is; both are critical to assess whether this might become a viable strategy to regenerate islet cells by suppressing the Fbw7 signaling. A fundamental assay on reprogrammed cells to ameliorate diabetes has also not been reported.
Interestingly, PDL robustly activates the Wnt signaling pathway and allows the regenerating ducts to express Lgr5[76], a Wnt target which marks actively dividing stem cells such as those present in the intestine[77]. Purified ductal Lgr5-expressing cells are also responsive to spondin 1 and form clonal 3D pancreatic organoids within the gel-forming Matrigel that generate ductal as well as endocrine lineages upon transplantation[76].
MPP likely present in acinar tissue of the pancreas
In the clinic with experimental transplantation of donated islets from the cadaver’s pancreas, the acinar cells are normally un-used. The discovery of a scientific and practical value for the acinar cells has hence drawn considerable attention. Cotransplantation with pancreatic cells from fetal mice under the kidney capsule in the immunocompromised mice, the acinar cells are reported to give rise to islet cells with undetectable β-cell division or cell fusion[78]. The results imply that MPP or progenitor cells might be present in the acinar tissue of human adult pancreas. Moreover, the Cre-loxP lineage tracing analysis showed that acinar cells expressing amylase and elastase gave rise to insulin-producing cells in a rotating culture system[79]. Nevertheless, a self-renewable assay at the single-cell level and its intermediate steps need to be demonstrated. Additionally, these studies have not excluded the possibility that some pancreatic acinar cells might have trans differentiated into surrogate insulin-expressing cells[80,81]. This likelihood was supported by a previous study that showed that mouse acinar cells could be directly re-programmed in vivo to β-like cells by forced expression of three DNA-binding genes, namely, Pdx1, Neurog3 as well as MafA[82]. In addition, as the acinar cells from cadavers[78] were not isolated by FACS, residual ductal or even islet cells present in the system may have compromised their conclusion. Supporting this view, a lineage tracing study in vivo did not provide evidence that the mouse acinar cells would give rise to insulin-secreting β-cells after 70%-80% pancreatectomy[83]. Thus these inconsistencies remain to be addressed more carefully.
MPP likely present in the endocrine pancreas
A large body of evidence suggests that the islets of Langerhans harbor the MPP. The potential MPP are nestin+ and hormone- present in both rat and human islets. These nestin+ cells are reported to expand in culture for approximately 8 mo and differentiate into cells that exhibit markers of liver (α-fetoprotein) and pancreas lineages (including amylase, CK19, PDX1, glucagon, insulin and neural-specific cell adhesion molecule). These nestin+ MPP candidates may thus have been involved in the generation of new pancreatic islet cells[65], potentially modulated by the incretin hormone GLP1, a processed product from the polypeptide proglucagon[84]. It, however, remains unclear whether these nestin+ cells are either islet or duct-originated multipotent progenitors that transform into neurosphere-like structures as well as differentiated into β-like cells of the endocrine pancreas[66], or the outgrown fibroblast-like cells that expand readily and differentiate in vitro into aggregates of non-typical hormone-expressing endocrine cells[85]. Human endocrine cells transduced with a RIP-controlling transgene dedifferentiated into fibroblast-like cells that proliferate up to 16 population doublings without detectable insulin expression[86]. Nevertheless, in vivo Nestin+ cells are mostly present in non-endodermal-derived compartments[87,88]. Governed by their promoters, Pdx1- and RIP-expressing cells have not committed significantly ex vivo to the observed cells with fibroblast-like morphology[89]. These discrepancies require clarifications in the future studies.
CONCLUSION
The presence of MPP remains to be conclusively established. Future development of the field needs to: establish essential criteria for MPP; screen and select cell surface antigens that can be used to generate antibodies for purification of candidate MPP and establish a simple, effective and robust in vitro assays and in vivo experimental protocols for the examination of the multipotency and lineage commitments of isolated MPP candidates.
The following minimal criteria have to be satisfied: (1) enriched or purified cell population by FACS or other technologies should form colonies at the single cell level; (2) single cells after purification should be studied ex vivo with their self-renewability; (3) colony cells would be able to differentiate in vitro into multiple functional cellular lineages; and (4) the clonogenic cells post transplanted into a recipient should be able to give rise in vivo to different terminally differentiated lineages.
Thus, embryonic mouse Sox9+ multipotent progenitors and probably adult Sox9+ ductal cells satisfy some but not all criteria of MPP. Research into MPP would therefore be highly valuable for two reasons. First, MPP differentiation and trans differentiation of non-β-cell types in the pancreas may provide an important source for surrogate β-cells. Second, as there is a significant difference in regeneration capability of islets from mice, rats to humans, we should not extrapolate directly from rodent regenerative data to humans.
Future MPP research should apply integrated approaches, different from many previous in situ or ex vivo studies that did not target defined cell populations. Application of flow cytometry and cell surface markers would allow the separation of interested subsets for demonstrating in vitro the capacity of self-duplication, clonogenesis and differentiation. The latest technologies such as single-cell RNA-seq, single-cell genomics and single-cell epigenomics[90-92] should be applied to examine their developmental dynamics, differentiation pathways, gene interactions and genetic heterogeneity, and along with genetic studies to characterize their growth pattern, biological potential and lineage commitment in vivo. Lastly cross-sector, cross-institutional and global collaborations as well as the involvement of the biotechnological and drug companies will eventually deepen our understanding of MPP that assists the establishment of a platform towards a regenerative therapy for both T1DM and T2DM.
Footnotes
Supported by Telethon Perth Child Health Research Foundation; the Diabetes Research Foundation of Western Australia; the University of Western Australia; and the National Health and Medical Research Council Program, No. 53000400.
Conflict-of-interest statement: Authors do not have conflict of interest to declare.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: May 23, 2015
First decision: August 4, 2015
Article in press: November 11, 2015
P- Reviewer: Sumi S, Yu R S- Editor: Ji FF L- Editor: A E- Editor: Wu HL
References
- 1.Halban PA, Polonsky KS, Bowden DW, Hawkins MA, Ling C, Mather KJ, Powers AC, Rhodes CJ, Sussel L, Weir GC. β-cell failure in type 2 diabetes: postulated mechanisms and prospects for prevention and treatment. J Clin Endocrinol Metab. 2014;99:1983–1992. doi: 10.1210/jc.2014-1425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Butler PC, Meier JJ, Butler AE, Bhushan A. The replication of beta cells in normal physiology, in disease and for therapy. Nat Clin Pract Endocrinol Metab. 2007;3:758–768. doi: 10.1038/ncpendmet0647. [DOI] [PubMed] [Google Scholar]
- 3.Hanley SC, Pilotte A, Massie B, Rosenberg L. Cellular origins of adult human islet in vitro dedifferentiation. Lab Invest. 2008;88:761–772. doi: 10.1038/labinvest.2008.41. [DOI] [PubMed] [Google Scholar]
- 4.Talchai C, Xuan S, Lin HV, Sussel L, Accili D. Pancreatic β cell dedifferentiation as a mechanism of diabetic β cell failure. Cell. 2012;150:1223–1234. doi: 10.1016/j.cell.2012.07.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Richardson MK, Hanken J, Gooneratne ML, Pieau C, Raynaud A, Selwood L, Wright GM. There is no highly conserved embryonic stage in the vertebrates: implications for current theories of evolution and development. Anat Embryol (Berl) 1997;196:91–106. doi: 10.1007/s004290050082. [DOI] [PubMed] [Google Scholar]
- 6.Piper K, Brickwood S, Turnpenny LW, Cameron IT, Ball SG, Wilson DI, Hanley NA. Beta cell differentiation during early human pancreas development. J Endocrinol. 2004;181:11–23. doi: 10.1677/joe.0.1810011. [DOI] [PubMed] [Google Scholar]
- 7.Herrera PL, Huarte J, Sanvito F, Meda P, Orci L, Vassalli JD. Embryogenesis of the murine endocrine pancreas; early expression of pancreatic polypeptide gene. Development. 1991;113:1257–1265. doi: 10.1242/dev.113.4.1257. [DOI] [PubMed] [Google Scholar]
- 8.Fougerousse F, Bullen P, Herasse M, Lindsay S, Richard I, Wilson D, Suel L, Durand M, Robson S, Abitbol M, et al. Human-mouse differences in the embryonic expression patterns of developmental control genes and disease genes. Hum Mol Genet. 2000;9:165–173. doi: 10.1093/hmg/9.2.165. [DOI] [PubMed] [Google Scholar]
- 9.Lukinius A, Ericsson JL, Grimelius L, Korsgren O. Ultrastructural studies of the ontogeny of fetal human and porcine endocrine pancreas, with special reference to colocalization of the four major islet hormones. Dev Biol. 1992;153:376–385. doi: 10.1016/0012-1606(92)90122-w. [DOI] [PubMed] [Google Scholar]
- 10.Polak M, Bouchareb-Banaei L, Scharfmann R, Czernichow P. Early pattern of differentiation in the human pancreas. Diabetes. 2000;49:225–232. doi: 10.2337/diabetes.49.2.225. [DOI] [PubMed] [Google Scholar]
- 11.Pan FC, Brissova M. Pancreas development in humans. Curr Opin Endocrinol Diabetes Obes. 2014;21:77–82. doi: 10.1097/MED.0000000000000047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.De Krijger RR, Aanstoot HJ, Kranenburg G, Reinhard M, Visser WJ, Bruining GJ. The midgestational human fetal pancreas contains cells coexpressing islet hormones. Dev Biol. 1992;153:368–375. doi: 10.1016/0012-1606(92)90121-v. [DOI] [PubMed] [Google Scholar]
- 13.Cheng X, Ying L, Lu L, Galvão AM, Mills JA, Lin HC, Kotton DN, Shen SS, Nostro MC, Choi JK, et al. Self-renewing endodermal progenitor lines generated from human pluripotent stem cells. Cell Stem Cell. 2012;10:371–384. doi: 10.1016/j.stem.2012.02.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jiang FX, Stanley EG, Gonez LJ, Harrison LC. Bone morphogenetic proteins promote development of fetal pancreas epithelial colonies containing insulin-positive cells. J Cell Sci. 2002;115:753–760. doi: 10.1242/jcs.115.4.753. [DOI] [PubMed] [Google Scholar]
- 15.Jiang FX, Harrison LC. Convergence of bone morphogenetic protein and laminin-1 signaling pathways promotes proliferation and colony formation by fetal mouse pancreatic cells. Exp Cell Res. 2005;308:114–122. doi: 10.1016/j.yexcr.2005.03.041. [DOI] [PubMed] [Google Scholar]
- 16.Jiang FX, Harrison LC. Laminin-1 and epidermal growth factor family members co-stimulate fetal pancreas cell proliferation and colony formation. Differentiation. 2005;73:45–49. doi: 10.1111/j.1432-0436.2005.07301002.x. [DOI] [PubMed] [Google Scholar]
- 17.Sarkar A, Hochedlinger K. The sox family of transcription factors: versatile regulators of stem and progenitor cell fate. Cell Stem Cell. 2013;12:15–30. doi: 10.1016/j.stem.2012.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lynn FC, Smith SB, Wilson ME, Yang KY, Nekrep N, German MS. Sox9 coordinates a transcriptional network in pancreatic progenitor cells. Proc Natl Acad Sci USA. 2007;104:10500–10505. doi: 10.1073/pnas.0704054104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Furuyama K, Kawaguchi Y, Akiyama H, Horiguchi M, Kodama S, Kuhara T, Hosokawa S, Elbahrawy A, Soeda T, Koizumi M, et al. Continuous cell supply from a Sox9-expressing progenitor zone in adult liver, exocrine pancreas and intestine. Nat Genet. 2011;43:34–41. doi: 10.1038/ng.722. [DOI] [PubMed] [Google Scholar]
- 20.Sugiyama T, Benitez CM, Ghodasara A, Liu L, McLean GW, Lee J, Blauwkamp TA, Nusse R, Wright CV, Gu G, et al. Reconstituting pancreas development from purified progenitor cells reveals genes essential for islet differentiation. Proc Natl Acad Sci USA. 2013;110:12691–12696. doi: 10.1073/pnas.1304507110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jin L, Feng T, Shih HP, Zerda R, Luo A, Hsu J, Mahdavi A, Sander M, Tirrell DA, Riggs AD, et al. Colony-forming cells in the adult mouse pancreas are expandable in Matrigel and form endocrine/acinar colonies in laminin hydrogel. Proc Natl Acad Sci USA. 2013;110:3907–3912. doi: 10.1073/pnas.1301889110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kopp JL, Dubois CL, Schaffer AE, Hao E, Shih HP, Seymour PA, Ma J, Sander M. Sox9+ ductal cells are multipotent progenitors throughout development but do not produce new endocrine cells in the normal or injured adult pancreas. Development. 2011;138:653–665. doi: 10.1242/dev.056499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang Y, Lanzoni G, Carpino G, Cui CB, Dominguez-Bendala J, Wauthier E, Cardinale V, Oikawa T, Pileggi A, Gerber D, et al. Biliary tree stem cells, precursors to pancreatic committed progenitors: evidence for possible life-long pancreatic organogenesis. Stem Cells. 2013;31:1966–1979. doi: 10.1002/stem.1460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jonsson J, Carlsson L, Edlund T, Edlund H. Insulin-promoter-factor 1 is required for pancreas development in mice. Nature. 1994;371:606–609. doi: 10.1038/371606a0. [DOI] [PubMed] [Google Scholar]
- 25.Ohneda K, Mirmira RG, Wang J, Johnson JD, German MS. The homeodomain of PDX-1 mediates multiple protein-protein interactions in the formation of a transcriptional activation complex on the insulin promoter. Mol Cell Biol. 2000;20:900–911. doi: 10.1128/mcb.20.3.900-911.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gao T, McKenna B, Li C, Reichert M, Nguyen J, Singh T, Yang C, Pannikar A, Doliba N, Zhang T, et al. Pdx1 maintains β cell identity and function by repressing an α cell program. Cell Metab. 2014;19:259–271. doi: 10.1016/j.cmet.2013.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gu G, Dubauskaite J, Melton DA. Direct evidence for the pancreatic lineage: NGN3+ cells are islet progenitors and are distinct from duct progenitors. Development. 2002;129:2447–2457. doi: 10.1242/dev.129.10.2447. [DOI] [PubMed] [Google Scholar]
- 28.Zhou Q, Law AC, Rajagopal J, Anderson WJ, Gray PA, Melton DA. A multipotent progenitor domain guides pancreatic organogenesis. Dev Cell. 2007;13:103–114. doi: 10.1016/j.devcel.2007.06.001. [DOI] [PubMed] [Google Scholar]
- 29.Seymour PA, Freude KK, Tran MN, Mayes EE, Jensen J, Kist R, Scherer G, Sander M. SOX9 is required for maintenance of the pancreatic progenitor cell pool. Proc Natl Acad Sci USA. 2007;104:1865–1870. doi: 10.1073/pnas.0609217104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sneddon JB, Borowiak M, Melton DA. Self-renewal of embryonic-stem-cell-derived progenitors by organ-matched mesenchyme. Nature. 2012;491:765–768. doi: 10.1038/nature11463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Holland AM, Góñez LJ, Naselli G, Macdonald RJ, Harrison LC. Conditional expression demonstrates the role of the homeodomain transcription factor Pdx1 in maintenance and regeneration of beta-cells in the adult pancreas. Diabetes. 2005;54:2586–2595. doi: 10.2337/diabetes.54.9.2586. [DOI] [PubMed] [Google Scholar]
- 32.Lyttle BM, Li J, Krishnamurthy M, Fellows F, Wheeler MB, Goodyer CG, Wang R. Transcription factor expression in the developing human fetal endocrine pancreas. Diabetologia. 2008;51:1169–1180. doi: 10.1007/s00125-008-1006-z. [DOI] [PubMed] [Google Scholar]
- 33.Jeon J, Correa-Medina M, Ricordi C, Edlund H, Diez JA. Endocrine cell clustering during human pancreas development. J Histochem Cytochem. 2009;57:811–824. doi: 10.1369/jhc.2009.953307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.McDonald E, Li J, Krishnamurthy M, Fellows GF, Goodyer CG, Wang R. SOX9 regulates endocrine cell differentiation during human fetal pancreas development. Int J Biochem Cell Biol. 2012;44:72–83. doi: 10.1016/j.biocel.2011.09.008. [DOI] [PubMed] [Google Scholar]
- 35.Sakano D, Shiraki N, Kikawa K, Yamazoe T, Kataoka M, Umeda K, Araki K, Mao D, Matsumoto S, Nakagata N, et al. VMAT2 identified as a regulator of late-stage β-cell differentiation. Nat Chem Biol. 2014;10:141–148. doi: 10.1038/nchembio.1410. [DOI] [PubMed] [Google Scholar]
- 36.Jiang FX, Morahan G. Directed differentiation of late stage islet lineages remains a knowledge gap in pancreatic endocrine development. J J Bone Stem Res. 2015;1:002. [Google Scholar]
- 37.Gradwohl G, Dierich A, LeMeur M, Guillemot F. neurogenin3 is required for the development of the four endocrine cell lineages of the pancreas. Proc Natl Acad Sci USA. 2000;97:1607–1611. doi: 10.1073/pnas.97.4.1607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Xu X, D’Hoker J, Stangé G, Bonné S, De Leu N, Xiao X, Van de Casteele M, Mellitzer G, Ling Z, Pipeleers D, et al. Beta cells can be generated from endogenous progenitors in injured adult mouse pancreas. Cell. 2008;132:197–207. doi: 10.1016/j.cell.2007.12.015. [DOI] [PubMed] [Google Scholar]
- 39.Hara M, Dizon RF, Glick BS, Lee CS, Kaestner KH, Piston DW, Bindokas VP. Imaging pancreatic beta-cells in the intact pancreas. Am J Physiol Endocrinol Metab. 2006;290:E1041–E1047. doi: 10.1152/ajpendo.00365.2005. [DOI] [PubMed] [Google Scholar]
- 40.Rankin MM, Wilbur CJ, Rak K, Shields EJ, Granger A, Kushner JA. β-Cells are not generated in pancreatic duct ligation-induced injury in adult mice. Diabetes. 2013;62:1634–1645. doi: 10.2337/db12-0848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schwitzgebel VM, Scheel DW, Conners JR, Kalamaras J, Lee JE, Anderson DJ, Sussel L, Johnson JD, German MS. Expression of neurogenin3 reveals an islet cell precursor population in the pancreas. Development. 2000;127:3533–3542. doi: 10.1242/dev.127.16.3533. [DOI] [PubMed] [Google Scholar]
- 42.Jensen J, Heller RS, Funder-Nielsen T, Pedersen EE, Lindsell C, Weinmaster G, Madsen OD, Serup P. Independent development of pancreatic alpha- and beta-cells from neurogenin3-expressing precursors: a role for the notch pathway in repression of premature differentiation. Diabetes. 2000;49:163–176. doi: 10.2337/diabetes.49.2.163. [DOI] [PubMed] [Google Scholar]
- 43.Oliver-Krasinski JM, Kasner MT, Yang J, Crutchlow MF, Rustgi AK, Kaestner KH, Stoffers DA. The diabetes gene Pdx1 regulates the transcriptional network of pancreatic endocrine progenitor cells in mice. J Clin Invest. 2009;119:1888–1898. doi: 10.1172/JCI37028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Desgraz R, Herrera PL. Pancreatic neurogenin 3-expressing cells are unipotent islet precursors. Development. 2009;136:3567–3574. doi: 10.1242/dev.039214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Miyatsuka T, Kosaka Y, Kim H, German MS. Neurogenin3 inhibits proliferation in endocrine progenitors by inducing Cdkn1a. Proc Natl Acad Sci USA. 2011;108:185–190. doi: 10.1073/pnas.1004842108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Alpert S, Hanahan D, Teitelman G. Hybrid insulin genes reveal a developmental lineage for pancreatic endocrine cells and imply a relationship with neurons. Cell. 1988;53:295–308. doi: 10.1016/0092-8674(88)90391-1. [DOI] [PubMed] [Google Scholar]
- 47.Smukler SR, Arntfield ME, Razavi R, Bikopoulos G, Karpowicz P, Seaberg R, Dai F, Lee S, Ahrens R, Fraser PE, et al. The adult mouse and human pancreas contain rare multipotent stem cells that express insulin. Cell Stem Cell. 2011;8:281–293. doi: 10.1016/j.stem.2011.01.015. [DOI] [PubMed] [Google Scholar]
- 48.Jiang FX, Mehta M, Morahan G. Quantification of insulin gene expression during development of pancreatic islet cells. Pancreas. 2010;39:201–208. doi: 10.1097/MPA.0b013e3181bab68f. [DOI] [PubMed] [Google Scholar]
- 49.Jiang FX, Morahan G. Pancreatic stem cells: from possible to probable. Stem Cell Rev. 2012;8:647–657. doi: 10.1007/s12015-011-9333-8. [DOI] [PubMed] [Google Scholar]
- 50.Dezaki K. Ghrelin function in insulin release and glucose metabolism. Endocr Dev. 2013;25:135–143. doi: 10.1159/000346064. [DOI] [PubMed] [Google Scholar]
- 51.Wierup N, Sundler F, Heller RS. The islet ghrelin cell. J Mol Endocrinol. 2014;52:R35–R49. doi: 10.1530/JME-13-0122. [DOI] [PubMed] [Google Scholar]
- 52.Collombat P, Mansouri A, Hecksher-Sorensen J, Serup P, Krull J, Gradwohl G, Gruss P. Opposing actions of Arx and Pax4 in endocrine pancreas development. Genes Dev. 2003;17:2591–2603. doi: 10.1101/gad.269003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Prado CL, Pugh-Bernard AE, Elghazi L, Sosa-Pineda B, Sussel L. Ghrelin cells replace insulin-producing beta cells in two mouse models of pancreas development. Proc Natl Acad Sci USA. 2004;101:2924–2929. doi: 10.1073/pnas.0308604100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hill JT, Mastracci TL, Vinton C, Doyle ML, Anderson KR, Loomis ZL, Schrunk JM, Minic AD, Prabakar KR, Pugliese A, et al. Ghrelin is dispensable for embryonic pancreatic islet development and differentiation. Regul Pept. 2009;157:51–56. doi: 10.1016/j.regpep.2009.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Arnes L, Hill JT, Gross S, Magnuson MA, Sussel L. Ghrelin expression in the mouse pancreas defines a unique multipotent progenitor population. PLoS One. 2012;7:e52026. doi: 10.1371/journal.pone.0052026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Dor Y, Brown J, Martinez OI, Melton DA. Adult pancreatic beta-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 2004;429:41–46. doi: 10.1038/nature02520. [DOI] [PubMed] [Google Scholar]
- 57.Nir T, Melton DA, Dor Y. Recovery from diabetes in mice by beta cell regeneration. J Clin Invest. 2007;117:2553–2561. doi: 10.1172/JCI32959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Brennand K, Huangfu D, Melton D. All beta cells contribute equally to islet growth and maintenance. PLoS Biol. 2007;5:e163. doi: 10.1371/journal.pbio.0050163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gupta RK, Gao N, Gorski RK, White P, Hardy OT, Rafiq K, Brestelli JE, Chen G, Stoeckert CJ, Kaestner KH. Expansion of adult beta-cell mass in response to increased metabolic demand is dependent on HNF-4alpha. Genes Dev. 2007;21:756–769. doi: 10.1101/gad.1535507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Georgia S, Bhushan A. Beta cell replication is the primary mechanism for maintaining postnatal beta cell mass. J Clin Invest. 2004;114:963–968. doi: 10.1172/JCI22098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Teta M, Rankin MM, Long SY, Stein GM, Kushner JA. Growth and regeneration of adult beta cells does not involve specialized progenitors. Dev Cell. 2007;12:817–826. doi: 10.1016/j.devcel.2007.04.011. [DOI] [PubMed] [Google Scholar]
- 62.Cornelius JG, Tchernev V, Kao KJ, Peck AB. In vitro-generation of islets in long-term cultures of pluripotent stem cells from adult mouse pancreas. Horm Metab Res. 1997;29:271–277. doi: 10.1055/s-2007-979036. [DOI] [PubMed] [Google Scholar]
- 63.Ramiya VK, Maraist M, Arfors KE, Schatz DA, Peck AB, Cornelius JG. Reversal of insulin-dependent diabetes using islets generated in vitro from pancreatic stem cells. Nat Med. 2000;6:278–282. doi: 10.1038/73128. [DOI] [PubMed] [Google Scholar]
- 64.Suzuki A, Oyama K, Fukao K, Nakauchi H, Taniguchi H. Establishment of clonal colony-forming assay system for pancreatic stem/progenitor cells. Cell Transplant. 2002;11:451–453. [PubMed] [Google Scholar]
- 65.Zulewski H, Abraham EJ, Gerlach MJ, Daniel PB, Moritz W, Müller B, Vallejo M, Thomas MK, Habener JF. Multipotential nestin-positive stem cells isolated from adult pancreatic islets differentiate ex vivo into pancreatic endocrine, exocrine, and hepatic phenotypes. Diabetes. 2001;50:521–533. doi: 10.2337/diabetes.50.3.521. [DOI] [PubMed] [Google Scholar]
- 66.Seaberg RM, Smukler SR, Kieffer TJ, Enikolopov G, Asghar Z, Wheeler MB, Korbutt G, van der Kooy D. Clonal identification of multipotent precursors from adult mouse pancreas that generate neural and pancreatic lineages. Nat Biotechnol. 2004;22:1115–1124. doi: 10.1038/nbt1004. [DOI] [PubMed] [Google Scholar]
- 67.Suzuki A, Nakauchi H, Taniguchi H. Prospective isolation of multipotent pancreatic progenitors using flow-cytometric cell sorting. Diabetes. 2004;53:2143–2152. doi: 10.2337/diabetes.53.8.2143. [DOI] [PubMed] [Google Scholar]
- 68.Bonner-Weir S, Taneja M, Weir GC, Tatarkiewicz K, Song KH, Sharma A, O’Neil JJ. In vitro cultivation of human islets from expanded ductal tissue. Proc Natl Acad Sci USA. 2000;97:7999–8004. doi: 10.1073/pnas.97.14.7999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Seeberger KL, Dufour JM, Shapiro AM, Lakey JR, Rajotte RV, Korbutt GS. Expansion of mesenchymal stem cells from human pancreatic ductal epithelium. Lab Invest. 2006;86:141–153. doi: 10.1038/labinvest.3700377. [DOI] [PubMed] [Google Scholar]
- 70.Bonner-Weir S, Inada A, Yatoh S, Li WC, Aye T, Toschi E, Sharma A. Transdifferentiation of pancreatic ductal cells to endocrine beta-cells. Biochem Soc Trans. 2008;36:353–356. doi: 10.1042/BST0360353. [DOI] [PubMed] [Google Scholar]
- 71.Inada A, Nienaber C, Katsuta H, Fujitani Y, Levine J, Morita R, Sharma A, Bonner-Weir S. Carbonic anhydrase II-positive pancreatic cells are progenitors for both endocrine and exocrine pancreas after birth. Proc Natl Acad Sci USA. 2008;105:19915–19919. doi: 10.1073/pnas.0805803105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Solar M, Cardalda C, Houbracken I, Martín M, Maestro MA, De Medts N, Xu X, Grau V, Heimberg H, Bouwens L, et al. Pancreatic exocrine duct cells give rise to insulin-producing beta cells during embryogenesis but not after birth. Dev Cell. 2009;17:849–860. doi: 10.1016/j.devcel.2009.11.003. [DOI] [PubMed] [Google Scholar]
- 73.Haumaitre C, Barbacci E, Jenny M, Ott MO, Gradwohl G, Cereghini S. Lack of TCF2/vHNF1 in mice leads to pancreas agenesis. Proc Natl Acad Sci USA. 2005;102:1490–1495. doi: 10.1073/pnas.0405776102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Haumaitre C, Fabre M, Cormier S, Baumann C, Delezoide AL, Cereghini S. Severe pancreas hypoplasia and multicystic renal dysplasia in two human fetuses carrying novel HNF1beta/MODY5 mutations. Hum Mol Genet. 2006;15:2363–2375. doi: 10.1093/hmg/ddl161. [DOI] [PubMed] [Google Scholar]
- 75.Sancho R, Gruber R, Gu G, Behrens A. Loss of Fbw7 reprograms adult pancreatic ductal cells into α, δ, and β cells. Cell Stem Cell. 2014;15:139–153. doi: 10.1016/j.stem.2014.06.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Huch M, Bonfanti P, Boj SF, Sato T, Loomans CJ, van de Wetering M, Sojoodi M, Li VS, Schuijers J, Gracanin A, et al. Unlimited in vitro expansion of adult bi-potent pancreas progenitors through the Lgr5/R-spondin axis. EMBO J. 2013;32:2708–2721. doi: 10.1038/emboj.2013.204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. doi: 10.1038/nature06196. [DOI] [PubMed] [Google Scholar]
- 78.Hao E, Tyrberg B, Itkin-Ansari P, Lakey JR, Geron I, Monosov EZ, Barcova M, Mercola M, Levine F. Beta-cell differentiation from nonendocrine epithelial cells of the adult human pancreas. Nat Med. 2006;12:310–316. doi: 10.1038/nm1367. [DOI] [PubMed] [Google Scholar]
- 79.Minami K, Okuno M, Miyawaki K, Okumachi A, Ishizaki K, Oyama K, Kawaguchi M, Ishizuka N, Iwanaga T, Seino S. Lineage tracing and characterization of insulin-secreting cells generated from adult pancreatic acinar cells. Proc Natl Acad Sci USA. 2005;102:15116–15121. doi: 10.1073/pnas.0507567102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Baeyens L, De Breuck S, Lardon J, Mfopou JK, Rooman I, Bouwens L. In vitro generation of insulin-producing beta cells from adult exocrine pancreatic cells. Diabetologia. 2005;48:49–57. doi: 10.1007/s00125-004-1606-1. [DOI] [PubMed] [Google Scholar]
- 81.Minami K, Okano H, Okumachi A, Seino S. Role of cadherin-mediated cell-cell adhesion in pancreatic exocrine-to-endocrine transdifferentiation. J Biol Chem. 2008;283:13753–13761. doi: 10.1074/jbc.M710034200. [DOI] [PubMed] [Google Scholar]
- 82.Zhou Q, Brown J, Kanarek A, Rajagopal J, Melton DA. In vivo reprogramming of adult pancreatic exocrine cells to beta-cells. Nature. 2008;455:627–632. doi: 10.1038/nature07314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Desai BM, Oliver-Krasinski J, De Leon DD, Farzad C, Hong N, Leach SD, Stoffers DA. Preexisting pancreatic acinar cells contribute to acinar cell, but not islet beta cell, regeneration. J Clin Invest. 2007;117:971–977. doi: 10.1172/JCI29988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Abraham EJ, Leech CA, Lin JC, Zulewski H, Habener JF. Insulinotropic hormone glucagon-like peptide-1 differentiation of human pancreatic islet-derived progenitor cells into insulin-producing cells. Endocrinology. 2002;143:3152–3161. doi: 10.1210/endo.143.8.8973. [DOI] [PubMed] [Google Scholar]
- 85.Gershengorn MC, Hardikar AA, Wei C, Geras-Raaka E, Marcus-Samuels B, Raaka BM. Epithelial-to-mesenchymal transition generates proliferative human islet precursor cells. Science. 2004;306:2261–2264. doi: 10.1126/science.1101968. [DOI] [PubMed] [Google Scholar]
- 86.Russ HA, Bar Y, Ravassard P, Efrat S. In vitro proliferation of cells derived from adult human beta-cells revealed by cell-lineage tracing. Diabetes. 2008;57:1575–1583. doi: 10.2337/db07-1283. [DOI] [PubMed] [Google Scholar]
- 87.Lardon J, Rooman I, Bouwens L. Nestin expression in pancreatic stellate cells and angiogenic endothelial cells. Histochem Cell Biol. 2002;117:535–540. doi: 10.1007/s00418-002-0412-4. [DOI] [PubMed] [Google Scholar]
- 88.Selander L, Edlund H. Nestin is expressed in mesenchymal and not epithelial cells of the developing mouse pancreas. Mech Dev. 2002;113:189–192. doi: 10.1016/s0925-4773(02)00023-0. [DOI] [PubMed] [Google Scholar]
- 89.Chase LG, Ulloa-Montoya F, Kidder BL, Verfaillie CM. Islet-derived fibroblast-like cells are not derived via epithelial-mesenchymal transition from Pdx-1 or insulin-positive cells. Diabetes. 2007;56:3–7. doi: 10.2337/db06-1165. [DOI] [PubMed] [Google Scholar]
- 90.Trapnell C, Cacchiarelli D, Grimsby J, Pokharel P, Li S, Morse M, Lennon NJ, Livak KJ, Mikkelsen TS, Rinn JL. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat Biotechnol. 2014;32:381–386. doi: 10.1038/nbt.2859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Shapiro E, Biezuner T, Linnarsson S. Single-cell sequencing-based technologies will revolutionize whole-organism science. Nat Rev Genet. 2013;14:618–630. doi: 10.1038/nrg3542. [DOI] [PubMed] [Google Scholar]
- 92.Macaulay IC, Haerty W, Kumar P, Li YI, Hu TX, Teng MJ, Goolam M, Saurat N, Coupland P, Shirley LM, et al. G& amp; T-seq: parallel sequencing of single-cell genomes and transcriptomes. Nat Methods. 2015;12:519–522. doi: 10.1038/nmeth.3370. [DOI] [PubMed] [Google Scholar]