

Abstract
NMDA receptors (NMDAR) play an important role in neural plasticity including long-term potentiation and long-term depression, which are likely to explain their importance for learning and memory. Cognitive decline is a major problem facing an ageing human population, so much so that its reversal has become an important goal for scientific research and pharmaceutical development. Enhancement of NMDAR function is a core strategy toward this goal. In this review we indicate some of the major ways of potentiating NMDAR function by both direct and indirect modulation. There is good evidence that both positive and negative modulation can enhance function suggesting that a subtle approach correcting imbalances in particular clinical situations will be required. Excessive activation and the resultant deleterious effects will need to be carefully avoided. Finally we describe some novel positive allosteric modulators of NMDARs, with some subunit selectivity, and show initial evidence of their ability to affect NMDAR mediated events.
Keywords: Synaptic plasticity, NMDA receptor, cognitive enhancement, LTP, synaptic transmission, hippocampus, positive allosteric modulators
1. Introduction
The NMDA receptor (NMDAR) is a prime target for cognitive enhancement since it is centrally involved in cognitive processes. Approximately 30 years ago, it was shown that the transient activation of NMDARs is the trigger for the induction of long-term potentiation (LTP) at synapses made between CA3 and CA1 pyramidal neurons in the hippocampus (Collingridge et al., 1983). Shortly afterwards direct evidence was provided that NMDARs are also required for forms of hippocampus dependent learning and memory (Morris et al., 1986). These findings have led to numerous studies into the role of NMDARs in synaptic plasticity, learning and memory and have placed the NMDAR at the heart of cognition. Since NMDARs are required for these processes the simple notion is that boosting NMDAR function should enhance cognition and, indeed, there is evidence that this may be true under certain circumstances (Tang et al., 1999). We will commence our discussion on this assumption: that NMDAR activation leads to LTP and that this equates with learning and memory and consequently enhancing NMDAR function is good for cognition. This is, of course, a gross over-simplification. Most importantly, NMDAR activation can result in pathological conditions, such as epilepsy (Croucher et al, 1982), neuronal cell death (Simon et al, 1984) and hyperalgesia (Davies & Lodge, 1987). Therefore, too much activation of the NMDAR is detrimental. The key is to boost the physiological function without promoting the tendency for pathological consequences.
NMDARs are obligate heterotetramers formed from assemblies of GluN1 subunits with GluN2A-D and GluN3A/B. In addition, GluN3A can assemble with GluN1 (without other GluN2 subunits) to form excitatory, Ca2+-impermeant glycine receptors. Eight possible variations of the GluN1 subunit arise by alternative splicing of a single gene transcript. The presence of one splice cassette at the N-terminal region of GluN1 and two independent consecutive splice variants at the C terminus have been identified. Therefore, a large number of different NMDARs with differing functional and pharmacological properties exist in different parts of the brain or at different stages in development (Molnár, 2008). Unusually for the ionotropic glutamate receptors (iGluRs), L-glutamate is not the only agonist for the NMDAR. Glycine and D-serine, two neutral endogenous amino acids, are co-agonists and the presence of one or other along with glutamate are needed for the receptor to function. The binding sites for glutamate and glycine/D-serine are found on different subunits -glycine binds to the GluN1 (and GluN3) subunits while glutamate binds to the GluN2 subunits. Consequently, both subunit types are required to generate a fully functioning NMDAR.
The NMDAR has several unique properties that are important for its function. Foremost, it is sensitive to block by low micromolar concentrations of magnesium ions (Mg2+) (Ault et al., 1980) in a manner that is highly voltage-dependent (Nowak et al., 1984; Mayer et al., 1984). The consequence of this block is such that at normal resting membrane potentials (typically between -50 and -75 mV) the NMDAR is largely blocked by Mg2+ from the synaptic cleft. Depolarization greatly reduces the Mg2+ block so that the participation of NMDARs in the synaptic response becomes substantially greater (Collingridge et al., 1988). This property explains the “Hebbian” nature of synaptic plasticity, whereby the NMDAR senses the co-incidence between presynaptic activity (which releases L-glutamate to bind to the NMDAR) and postsynaptic activity (defined as enough depolarization to reduce the Mg2+ block sufficiently to trigger the induction of plasticity). We shall refer to this depolarization as “Hebbian depolarization”. The NMDAR is also directly permeable to Ca2+ and this is extremely relevant for both its physiological and pathological actions.
Due to the complex molecular organization, functional and pharmacological properties of NMDARs, the design of agents to boost cognition via the regulation of NMDAR function needs to take account of many factors. In the present article, we discuss ways in which NMDAR function can be regulated. Broadly speaking, compounds that regulate NMDAR function do so in one of two ways. First, they may interact with other proteins that then regulate NMDAR function indirectly. Second, they may bind directly to the NMDAR to regulate its function. In the present article we discuss some of the ways in which NMDAR function may be regulated and describe some recently reported NMDAR positive allosteric modulators (PAMs).
2. Indirect modulation
The properties of the NMDAR enables many forms of indirect modulation, many of which are probably utilized physiologically for cognitive purposes and can be exploited, in principle, for the design of cognitive enhancing compounds. Some of the more important indirect modulators are described below and illustrated schematically in Fig. 1.
Fig. 1.

Indirect modulation of NMDARs. (A) Schematic representation of some ways in which NMDAR function can be regulated indirectly. Neurotransmitters, and other neuronal regulators, can facilitate NMDAR function by augmenting the “Hebbian depolarization” and by intracellular regulation. NMDARs are important for (B) synaptic transmission (C) the induction of LTP and (D) the induction of LTD. Note that NMDARs contribute considerably to the synaptic response during high frequency synaptic transmission; in this example the NMDAR-EPSP has summated with the AMPAR-EPSPs (shaded yellow) to fire several action potentials (adapted from Herron et al, 1986). LTP is induced by a brief period of high frequency stimulation whilst LTD is induced by a prolonged period of low frequency stimulation. Key: Different types of receptor populations are shown by a colour-coded symbol. Inward current via AMPARs and NMDARs (carried mainly by Na+) contributes to the Hebbian depolarization and is shown by a red arrow. Outward current (carried mainly by the movement of K+ (GABAB) out of the cell or Cl- (GABAA) into the cell) opposes the Hebbian depolarization and is depicted by a blue arrow. Ca2+ entry is shown by the grey arrow and Mg2+ by a black circle.
2.1. AMPARs
During the induction of LTP, Hebbian depolarization is provided in part by the temporal summation of AMPAR-mediated EPSPs (Collingridge, 1985). Therefore one way, in theory, of boosting NMDAR function is to enhance the depolarization provided by the synaptic activation of AMPARs. This is one of the ideas behind the use of positive allosteric modulators of AMPARs (AMPAR PAMs), compounds that bind to the AMPAR itself to enhance its function. Following the initial descriptions of aniracetam (Ito et al., 1990), diazoxides and thiazides (Yamada and Rothman, 1992), including cyclothiazide (Palmer and Lodge, 1993; Patneau et al. 1993) and benzamides (Arai et al., 1994), AMPAR potentiators were found to limit receptor desensitization and deactivation (Partin et al., 1996). Such AMPAR PAMs were shown to potentiate LTP presumably by indirect enhancement of NMDARs (Staubli et al., 1994b), as demonstrated in vivo (Vandergriff et al., 2001). In parallel with these electrophysiological studies, AMPAR PAMs were soon shown to enhance learning and memory (Staubli et al., 1994a). Since then, many other structural classes have been described (Ward and Harries, 2010; Pirotte et al., 2010) and their positive effects on cognition in laboratory animals and human patients have been extensively reported and reviewed (Morrow et al., 2006; Arai and Kessler, 2007; O'Neill and Dix, 2007; Cleva et al., 2010; Lynch et al., 2011). The potential site of action of AMPAR PAMs, together with other cognitive enhancing agents that may act at the glutamatergic synapse, is shown schematically in Fig. 2.
Fig. 2.

Potential sites of action of cognitive enhancers at glutamatergic synapses and structures of some compounds that potentiate NMDAR function. The key is the same as in Fig. 1.
2.2. GABARs
GABA receptors (GABARs) provide a powerful physiological regulation of NMDARs. During low frequency transmission the synaptic activation of GABARs prevents NMDARs from contributing appreciably to the synaptic response by hyperpolarizing the neuron and thereby intensifying the Mg2+ block (Herron et al., 1985; Dingledine et al., 1985). GABAARs are activated rapidly whereas GABABRs are activated after a delay of around 20 ms but provide a longer lasting hyperpolarization (Davies et al., 1990). Together, these two inhibitory synaptic responses effectively limit the synaptic activation of NMDARs throughout its time-course. Consequently, blocking either GABAA or GABAB receptors may lead to the enhanced synaptic activation of NMDARs (Davies and Collingridge, 1996). Since the GABAAR mediated inhibitory postsynaptic potential (IPSP) coincides with the peak NMDAR synaptic conductance, this is likely to have the most dramatic effect. During low frequency synaptic transmission, a GABAAR antagonist enables a noticeable activation of NMDARs (Herron et al., 1985; Dingledine et al., 1985) and the effect is magnified during high frequency transmission, since it facilitates the temporal summation of NMDAR-EPSPs to generate a larger Hebbian depolarization. This effect can be sufficient to enhance the induction of LTP (Abraham et al., 1986).
GABABRs provide a more complex regulation of NMDARs. The postsynaptic GABABR IPSPs helps limit the synaptic activation of NMDARs and so its selective blockade is able to enhance the induction of LTP (Olpe et al, 1993). However, GABABRs are also located presynaptically where they function as both autoreceptors, inhibiting GABA release (Davies et al., 1990), and heteroreceptors, inhibiting glutamate release (Davies et al., 1993; Isaacson et al., 1993). The autoreceptor function is important for the induction of LTP by theta/priming patterns of activity (Davies et al., 1991), which are a more physiologically relevant pattern of activation than a conventional tetanus (Larson et al., 1986; Diamond et al., 1988). This is because theta frequencies are optimally tuned for the suppression of GABAR-mediated IPSPs, via the autoreceptor mechanism, and this promotes the synaptic activation of NMDARs by facilitating the Hebbian depolarization (Davies and Collingridge, 1993). Antagonism of GABABR autoreceptors therefore inhibits the induction of LTP when theta patterns of activity are used, by limiting the synaptic activation of NMDARs. However, when longer trains are used to induce LTP (i.e, a tetanus) GABABRs are no longer required to suppress GABAR-IPSPs and so GABABR antagonists no longer inhibit the induction of LTP. Whether the regulation of GABABRs can be exploited to enhance cognition is not known. The prediction from these LTP experiments is that the selective antagonism of postsynaptic GABABRs might have the desired effect. This might be possible since a greater occupancy of postsynaptic GABABRs is required to elicit a response and so this response is the more sensitive to antagonism (Seabrook et al., 1990).
2.3. Acetylcholine receptors
The crucial role of acetylcholine in cognition has long been recognized (Sarter and Parikh, 2005). There is good evidence that some of its actions might be mediated via the regulation of NMDARs. In particular, the stimulation of muscarinic AChRs (mAChRs) is known to facilitate the activation of NMDARs (Markram and Segal, 1990). This probably occurs via multiple mechanisms. Activation of mAChRs leads to depolarization of neurons and so would be expected to facilitate the Hebbian depolarization. Since mAChRs depolarize neurons via the inhibition of K+ channels, the associated increase in membrane resistance will also facilitate the Hebbian depolarization. A particularly interesting regulation is via the inhibition of SK channels that are located at synapses and activated by Ca2+ entry during the synaptic activation of NMDARs (Buchanan et al., 2010). In addition, it is likely that the activation of mAChRs facilitates the activation of NMDARs via mechanisms independent of K+ channels. Low concentrations of a mAChR agonist, carbachol, below those that appreciably affect K+ channels, can exert a powerful regulation of NMDARs, via a pathway that has not been fully delineated but seems to be independent of PKC and the release of Ca2+ from stores (Harvey et al., 1993). Activation of mAChRs can also lead to a long-lasting depression of NMDAR mediated synaptic transmission, via the induction of a form of synaptic plasticity. This effect is due to the internalization of NMDARs and is triggered by IP3 receptor-mediated Ca2+ release from stores (Jo et al., 2010). Clearly, the mAChR regulation of NMDARs is multifaceted and the direction of the regulation (enhancement or depression) depends on a variety of factors.
2.4. Metabotropic glutamate receptors
Group I mGluRs (mGlu1 and mGlu5), which like muscarinic M1 receptors couple to Gq11, are also able to regulate the activation of NMDARs in multiple ways. For example, it was found that activation of group I mGluRs, using 3,5-dihydroxyphenylglycine (DHPG) (Fitzjohn et al., 1996), or more specifically mGlu5 receptors, using 2-chloro-5-hydroxyphenylglycine (CHPG) (Doherty et al., 1997; Fig. 2), is able to directly potentiate the depolarization of CA1 neurons induced by NMDA. This effect is very robust and has been exploited in attempts to boost NMDAR function, though again its mechanism has not been fully elucidated. The mGlu5 receptor and NMDARs can work in concert for induction of LTP (Bortolotto et al., 2005; Lante et al., 2006) with mGlu5 playing an increasing role with age (Lante et al., 2006). An mGlu5 receptor antagonist blocked LTP in the dentate gyrus and reduced performance on a radial maze task (Manahan-Vaughan et al., 2004). A series of positive allosteric modulators (PAMs) of mGlu5 receptors have been developed and shown to facilitate the activation of NMDARs (see Vinson and Conn 2011 for review) and enhance LTP (Ayala et al., 2009; Kroker et al., 2011). These compounds have been found to increase behavioural flexibility in a set-shifting paradigm (Darrah et al., 2008) to enhance novel object recognition and to reduce impulsivity in a five choice serial reaction time test (Liu et al., 2008), to reverse cognitive deficits induced by MK-801 (Vales et al. 2010) and to improve learning deficits in the methylazoxymethanol acetate (MAM) model of schizophrenia (Gastambide et al., 2012; and see Vinson and Conn, 2011 for a review).
2.5. Other modulators
There are numerous modulators that can regulate NMDAR function via depolarization (to relieve the Mg2+ block and thereby facilitate the Hebbian depolarization). Regulation can also be via intracellular signaling pathways, as exemplified by mGluRs and mAChRs. The former is a regulation that can, in principle, occur during the synaptic release of L-glutamate and, as such, can contribute to the property of cooperativity. Acetylcholine represents one of potentially many neurotransmitters and neuronal regulators that may regulate NMDAR function in an associative manner. As discussed later (see 3.6.1), other G-protein-coupled receptors (GPCRs; e.g. pituitary adenylate cyclase activated peptide 1 receptors, PAC1Rs) and dopamine 1 receptors (D1Rs) have also been implicated in the subtype-specific modulation of NMDARs via Src family kinases (Yang et al., 2012). These properties may therefore be exploited physiologically during the execution of cognitive processes.
3. Direct modulation
3.1. Antagonists
Consistent with the idea that NMDAR-LTP is important for learning and memory, NMDAR antagonists have been found, in numerous investigations, to impair these processes. However, NMDAR antagonists can also be cognitively enhancing under certain circumstances. The most notable example of this is memantine, a substance that is used in the treatment of Alzheimer's disease where it has a modest effect in delaying the decline in cognitive function (Danysz and Parsons, 2003).
One way in which NMDAR antagonists may be able to enhance cognition is by selectively inhibiting the “pathological activation” while preserving the “physiological activation” of NMDARs. This principle was first demonstrated in a simple slice experiment in which Mg2+ was removed from the perfusing solution (Coan et al., 1989). This treatment led to the inhibition of LTP. Under these conditions, addition of the specific competitive NMDAR antagonist AP5 was able to fully restore the ability to induce LTP. This was because AP5 was able to block the aberrant activation of NMDARs caused by the removal of Mg2+ but during tetanic stimulation sufficient glutamate release occurred to outcompete the AP5 to activate NMDARs appropriately to enable the induction of LTP. These results are re-plotted and represented schematically in Fig. 3.
Fig. 3.

Inappropriate activation of NMDARs inhibits LTP. The panels show data (left panel; replotted from Coan et al, 1989) and schematics during baseline (centre) and a tetanus (right) under four experimental conditions (from top to bottom): in 1 mM Mg2+ (grey shading and black circles), following perfusion with Mg2+-free medium, and following the addition of either 20 μM or 200 μM D-AP5 in Mg2+-free medium (green shading and circles). Calibration bar is 4 mV and 5 min. Optimal conditions for LTP requires minimal activation of NMDARs except during the induction stimulus (time of delivery of the tetanus is indicated by an arrow). By removing Mg2+, NMDAR activation in enhanced throughout the recording period and this inhibits the generation of LTP. A low concentration of D-AP5 normalizes the situation by inhibiting spurious NMDAR activation, but is outcompeted by L-glutamate during high frequency stimulation. However, a high concentration of D-AP5 inhibits NMDARs during high frequency stimulation.
This mechanism (block of pathological but not physiological NMDAR activation) is the rationale behind the cognitive enhancing effects of memantine. Indeed, it was shown that memantine could, like AP5, restore the loss of LTP resulting from treatment with a low Mg2+ solution (Frankiewicz and Parsons, 1999). With memantine the mechanism is slightly different since it is a fast, voltage-dependent channel blocker (Bresink et al., 1996; Frankiewicz et al., 1996). Therefore memantine suppresses the pathological activation of NMDARs by occupying the channel but its block is relieved by the Hebbian depolarization during the induction of LTP (Fitzjohn et al., 2008).
Along the same lines, the pathological activation of NMDARs may be limited, without greatly affecting its physiological activation, by increasing the Mg2+ concentration. This strategy has also proven effective and so Mg2+ can be considered a cognitive enhancing agent (Slutsky et al., 2010). This principle can, of course, be extended to other types of NMDAR antagonists and compounds that regulate NMDAR function indirectly, such as mGlu5 receptor negative allosteric modulators. With respect to NMDAR antagonists, there is evidence to suggest that compounds selective for GluN2B may enhance cognitive and their possible use in Alzheimer's disease is under investigation (see Mony et al, 2009).
3.2. Glycine site
The co-agonist role of glycine or a glycine-like substance, such as D-serine, for the NMDAR channel complex was discovered by Johnson and Ascher (1987) who initially hypothesized an allosteric site that positively modulates the probability of channel opening. This initial observation was quickly followed by demonstrations of selective functional antagonists at the glycine site (HA-966: Fletcher and Lodge, 1988; Foster and Kemp, 1989), (7-chlorokynurenic acid: Kemp et al., 1988) and (1-aminocyclobutane-1-carboxylate: Hood et al., 1989). The absolute requirement for occupation of the glycine-site was confirmed by Kleckner and Dingledine (1988) by meticulous elimination of glycine from the extracellular medium. Full activation of NMDARs requires agonist binding at two glycine and two glutamate receptors on the tetrameric complex (Beneviste and Mayer, 1991; Clements and Westbrook, 1991). Site directed mutagenesis of the GluN1 subunit determined GluN1 as the site of glycine's action (Kuryatov et al., 1994: Wafford et al., 1995). Interestingly, the GluN3 subunits are also activated by glycine rather than by glutamate so that tetrameric GluN1/GluN3 receptors are putative excitatory glycine receptors, not requiring the presence of glutamate (Chatterton et al., 2002; Madry et al., 2007)
The apparent high affinity of glycine for this site (Johnson and Ascher, 1987; Kleckner and Dingledine, 1988) and several experimental studies at the time (Fletcher and Lodge, 1988; Kemp et al., 1988; but see Thomson et al., 1989) indicated that the glycine site was likely to be fully occupied in vivo either by glycine itself or by D-serine which had been originally described as mimicking the actions of glycine (Johnson and Ascher, 1987). Further studies (e.g. Berger et al., 1998) suggested that at some locations in the central nervous system (CNS) at least and, as a result of the activity of high affinity glycine transporters (GlyT-1; Supplison and Bergman, 1997), the glycine site is not fully saturated by glycine.
As a result of this incomplete occupancy, several potential strategies for enhancing NMDAR function, and hence putatively improving cognition, via the glycine site have emerged (Fig. 2). Most obvious is exogenous administration of agonists and partial agonists. Access of exogenous glycine to synapses utilising NMDARs is, however, limited by GlyT-1 and inhibition of this transporter is, therefore, more likely to be successful in increasing glycine levels at the NMDAR (Supplison and Bergman, 1997; Berger et al., 1998; Chen et al., 2003; Bergeron et al., 1998; Martina et al., 2004; Stevens et al., 2010). GlyT-1 inhibition has been shown to enhance LTP (Martina et al., 2004; Manahan-Vaughan et al., 2008) and cognition in both a social recognition test (Shimazaki et al., 2010) and an attentional set-shifting task (Nikifurik et al., 2011). Some preliminary studies in schizophrenics indicate improved cognitive performance (Tsai et al., 1998, 2004; Lane et al., 2005; Heresco-Levy et al., 1996) but the danger of side effects due to activation of strychnine-sensitive inhibitory glycine receptors needs attention (Kopac et al., 2010).
Increasing extracellular levels of D-serine, which does not activate the glycine inhibitory receptor, offers several approaches. Exogenous D-serine has been shown to enable LTP, when the glycine site is not fully occupied by glycine (Oliver at al., 1990; Bashir et al., 1990; Watanabe et al., 1992; Duffy et al., 2008). Endogenous D-serine, produced in astrocytes by serine racemase from L-serine and glycine, has been shown to be necessary for both LTP and LTD (Yang et al., 2003; Yang et al., 2005; Mothet et al., 2006; Henneberger et al., 2010; Fossat et al., 2012). Age-related or genetic deficiencies in serine racemase lead to reduced LTP and memory disruption which can be reversed by exogenous D-serine administration (Mothet et al., 2006; DeVito et al., 2011).
A further potential means of enhancing D-serine levels is by inhibition of its catabolising enzyme, D-amino acid oxidase (DAAO). This strategy has received a boost from the finding that mutant mice lacking DAAO show enhanced hippocampal LTP and improved Morris water maze performance compared with normal mice (Maekawa et al., 2005). Furthermore there are growing genetic associations between DAAO and schizophrenia, (for example Chumakov et al., 2002). Several DAAO inhibitors have been synthesised and assessed in models of schizophrenia (see Ferraris and Tsukamoto, 2011) but it seems that DAAO inhibition alone does not raise D-serine levels as efficiently as exogenous administration of D-serine (see Smith et al. 2010). In support of this, Strick et al. (2011) found that DAAO inhibition did not significantly affect either brain levels of D-serine, except in the cerebellum, or performance in cognitive behavioural assays although there was an increase in hippocampal theta rhythm.
Enhancement of NMDA activity via the glycine site remains an attractive therapeutic possibility. As occupancy of this site decreases with old age, some combination of exogenous administration of D-serine, inhibition of GlyT-1 and inhibition of DAAO may prove therapeutically successful.
3.3 Polyamines
Endogenous polyamines, such as spermine, have been shown to potentiate the activity of agonists on NMDARs (Williams et al., 1994). This potentiation is thought to be mediated by (i) a glycine-dependent, (ii) a voltage-dependent and (iii) a glycine-and voltage-independent effects of spermine. The glycine-dependent effect is observed in GluN2A or GluN2B containing NMDARs and is thought to be due to an increase in the affinity of glycine for the receptor. The presence of exon 5 in the N-terminal domain (NTD) of GluN1 does not affect the glycine-dependent effect nor does extracellular pH (Williams, 1997). The glycine-independent effect refers to the ability of spermine to enhance NMDAR currents evoked by saturating concentrations of glycine and glutamate. Tonic inhibition of NMDARs by protons can be relieved by stimulation by spermine (Traynelis et al., 1995). This effect is only observed in GluN1/GluN2B containing NMDARs and only if the NTD of the GluN1 subunit does not contain the exon-5 insert. Spermine has a weaker potentiating effect at subsaturating concentrations of glutamate due its effect of lowering sensitivity of GluN1/GluN2B-containing NMDARs to glutamate (Williams, 1994a).
3.4. Neurosteroids
Endogenous neurosteroids have numerous actions in the brain including modulation of GABARs and NMDARs (Korinek et al., 2011). Pregnanolone and allopregnanolone, like progesterone, potentiate the actions of GABARs with minimal inhibitory effects on NMDAR function, which may underlie their anaesthetic actions (Selye, 1941). Pregnanolone sulfate weakly inhibits both GABAR and NMDAR actions whereas pregnenolone sulfate (PS) strongly potentiates NMDAR function (Fig. 2) and weakly inhibits GABAergic activity. Other neurosteroids related to pregnenolone have similar but in general less efficacious profiles.
PS, which is synthesized in brain tissue (Corpechot et al., 1983), is then the neurosteroid of interest for potentiating NMDAR function. PS, first described as a positive and selective allosteric modulator of NMDARs on chick spinal neurons (Wu et al., 1991) and hippocampal neurons (Bowlby et al., 1993) was shown to enhance learning and memory (Flood et al., 1992; Mayo et al., 1993). It now appears that its actions are quite complex with both inhibitory and facilitatory sites on NMDARs (Park-Chung et al., 1997; Horak et al., 2006; Kotsakis et al., 2011). In GluN2A or GluN2B subunit containing NMDARs, the facilitatory site predominates whereas, on those with GluN2C or GluN2D, inhibition by PS predominates (Kotsakis et al., 2011). An earlier study suggested selective effects of PS and other neurosteroids (but not anabolic androgenic steroids) on GluN2B rather than GluN2A containing receptors (see Elfverson et al., 2011). These PS binding sites are likely to be in the M3-M4 extracellular loop (S2 domain) of the GluN2 subunits, close to the proton site (Kotsakis et al., 2011). A further complication is that positive modulation of NMDARs by PS has been reported to be phosphorylation state dependent (Petrovic et al., 2009).
Enhanced NMDAR function underlies enhanced LTP at CA1 synapses in the presence of PS (Sliwinski et al., 2004; Sabeti et al., 2007) and PS alone is able to induce long lasting potentiation (LLP) of synaptic efficacy at granule cell synapses (Chen et al., 2007). In this PS-induced LLP, the LTD-LTP induction curve is shifted to the left so that lower stimulus frequencies are needed to induce these two forms of plasticity (Chen et al., 2010). Levels of this endogenous neurosteroid declines with age in parallel with cognitive decline (Flood et al., 1995; Vallee et al., 1997). This decline in performance in the water maze task can be reversed by exogenous administration of PS (Vallee et al., 1997). PS has also been claimed to reduce the amnesic effects of stress (Reddy et al., 1998; Urani et al., 2002).
Dehydroepiandrosterone sulfate (DHEAS), itself synthesized from PS, also potentiates NMDARs and enhances LTP (Randall et al., 1995; Chen et al., 2006; for review see, Dubrovsky, 2005). Interestingly, inhibition of the synthesis of PS and DHEAS in hippocampal slices reduces NMDAR function and LTP induction, suggesting a local ongoing synthesis of these neurosteroids (Tanaka and Sokabe, 2012).
3.5. Histamine and ATP
Histamine has been shown to potentiate agonist stimulated effects on recombinant NMDARs containing GluN1/GluN2B but not those containing GluN2A or GluN2C subunits (Williams, 1994). The effect was dependent on high agonist concentrations and did not occur in NMDARs containing GluN1 subunits with the exon 5 insert in the NTD.
Adenosine triphosphate (ATP) inhibits recombinant NMDARs containing GluN1/GluN2A or GluN1/GluN2B subunits at low agonist concentrations but at saturating agonist concentrations acts as a potentiator (Kloda et al., 2004). In contrast, ATP potentiates NMDARs containing GluN1/GluN2C even at nonsaturating agonist concentrations. ATP has been proposed to compete with glutamate for its binding site and this may explain why high agonist concentrations are required to reveal the potentiating effect of ATP on GluN1/GluN2A or GluN1/GluN2B.
3.6. Intracellular modulation of NMDARs
The function of NMDARs, like other iGluRs, is also regulated by posttranslational modifications (e.g. phosphorylation, palmitoylation, ubiquitination, proteolytic cleavage by calpain) and by protein binding partners, Here we overview some of the complex intracellular pathways that are key modulators of NMDAR function. These regulatory mechanisms could in principle be targeted pharmacologically.
3.6.1. Functionally significant key phosphorylation sites of NMDAR subunits
The function and subcellular distribution of NMDARs are differentially regulated by phosphorylation of specific serine (S) / threonine (T) and tyrosine (Y) amino acid residues in the intracellular C-terminal domains of various subunit proteins (Salter et al., 2009; Traynelis et al., 2010). Protein kinases that catalyze phosphorylation and phosphoprotein phosphatases that catalyze dephosphorylation are recruited to NMDARs via interactions with postsynaptic density protein 95 (PSD95), A-kinase anchoring proteins (AKAP79/150) and yotiao (Colledge et al., 2000; Klauck et al., 1996; Salter et al., 2009; Westphal et al., 1999). This compartmentalisation is likely to increase the selectivity, efficiency and speed of phosphorylation and dephosphorylation events. In this section, we highlight some of the main functional changes produced by the phosphorylation of key sites in core NMDAR subunit proteins. See recent reviews for a more extensive list of possible, but not fully verified NMDAR phosphorylation sites (Salter et al., 2009; Traynelis et al., 2010).
In the alternatively spliced C-terminal C1-cassette of GluN1, protein kinase C (PKC) phosphorylates serine residues GluN1-S890 and GluN1-S896 (Tingley et al., 1997) (Fig. 4). In contrast, the neighbouring GluN1-S897 is phosphorylated by cyclic AMP-dependent protein kinase A (PKA; Tingley et al., 1997) (Fig. 4). Phosphorylation of these sites regulates cell surface expression and clustering of NMDARs (Crump et al., 2001; Ehlers et al., 1995; Fong et al., 2002; Scott et al., 2003; Tingley et al., 1997) and may affect channel function by modulating the inhibitory interaction between GluN1 and calmodulin (Ehlers et al., 1996; Hisatsune et al., 1997). GluN1-S890 and GluN1-S896 are preferentially phosphorylated by PKCγ and PKCα, respectively (Sánchez-Pérez and Felipo, 2005) (Fig. 4). Phosphorylation of the GluN1-S890 but not GluN1-S896 or GluN1-S897 residues facilitates rapid dispersal of synaptic NMDARs (Tingley et al., 1997). Activation of group I mGluRs by DHPG increases GluN1-S890 but not GluN1-S896 phosphorylation. Surface expressed GluN1 proteins are phosphorylated at S890 but not at S896 (Sánchez-Pérez and Felipo, 2005). The dual PKC/PKA phosphorylation of GluN1-S896/GluN1-S897 promotes NMDARs trafficking from the endoplasmic reticulum to the cell surface (Scott et al., 2001, 2003).
Fig. 4.

Sites of intracellular modulation of NMDARs. Schematic representation of the distribution of selected posttranslational regulatory sites on the intracellular C-terminal domains of GluN1, GluN2A and GluN2B NMDAR subunits. Serine (S) phosphorylation sites: GluN1-S890, GluN1-S896, GluN1-S897, GluN2B-S1303, GluN2B-S1323, GluN2B-S1480 (Chung et al., 2004; Leonard and Hell, 1997; Liao et al., 2001; Liu et al., 2006; Sanchez-Perez and Felipo, 2005; Sanz-Clemente et al., 2010; Scott et al., 2001, 2003; Tingley et al., 1997). Tyrosine (Y) phosphorylation sites: GluN1-Y837, GluN2A-Y842 GluN2A-Y1336, GluN2A-Y1387, GluN2B-Y1336, GluN2B-Y1472 (Lau and Huganir, 1995; Moon et al., 1994; Nakazawa et al., 2001; Vissel et al., 2001; Yang and Leonard, 2001). Cysteine (C) palmitoylation sites: GluN2A-C848, GluN2A-C853, GluN2A-C870, GluN2A-C1214, GluN2A-C1217, GluN2A-C1236, GluN2A-C1239, GluN2B-C849, GluN2B-C854, GluN2B-C871, GluN2B-C1215, GluN2B-C1218, GluN2B-C1242, GluN2B-C1239, GluN2B-C1245 (Hayashi et al., 2009). Calpain cleavage sites: GluN2A-1279, GluN2A-1330, GluN2B∼1030 (approximately) (Dong et al., 2006; Guttmann et al., 2001; Simpkins et al., 2003; Doshi and Lynch, 2009). See text for further details (3.6. Intracellular modulation of NMDARs).
The C-terminal domains of GluN2A and GluN2B are substrates for Ca2+/calmodulin-dependent protein kinase (CaMKII; Omkumar et al., 1996), PKC and PKA (Leonard and Hell, 1997; Liao et al., 2001; Fig 4). NMDAR-PSD95/SAP102 interactions are disrupted by casein kinase II (CK2)-mediated GluN2B-S1480 phosphorylation (Chung et al., 2004). This activity-dependent process is thought to be an important component of the developmental switch from GluN2B- to GluN2A-containing NMDARs at synapses (Sanz-Clemente et al., 2010). Phosphorylation of GluN2B-S1303 and GluN2B-S1323 by PKC potentiates GluN2B-containing NMDAR current (Liao et al., 2001; Fig 4). GluN2B-S1303 may also be phosphorylated by CaMKII, which affects the receptor-kinase interaction (Liu et al., 2006). In GluN2C, S1230 is phosphorylated by both PKA and PKC (Chen et al., 2006). Phosphomimetic mutation of GluN2C-S1230 accelerates channel kinetics by increasing the speed of both the rise and decay of NMDA-evoked currents (Chen et al., 2006). Increased PKA activity can facilitate the induction of LTP by increasing the Ca2+ permeability of NMDARs in dendritic spines (Skeberdis et al., 2006).
Several potential sites of tyrosine kinase phosphorylation have been identified in GluN1, GluN2A and GluN2B (Moon et al., 1994; Lau and Huganir 1995; Fig. 4). Disruption of GluN1-Y837 and GluN2A-Y842, by site-directed mutagenesis, prevented use-dependent desensitization of GluN1/GluN2A NMDARs (Vissel et al., 2001). However, the GluN1 intracellular C-terminus does not appear to be tyrosine phosphorylated in neurons (Lau and Huganir, 1995). GluN2A-Y1387 and GluN2B-Y1472 are major sites of phosphorylation by Src-family kinases (Nakazawa et al., 2001; Yang and Leonard, 2001; Fig 4). Tyrosine phosphorylation potentiates the NMDAR ion channel resulting in increased Ca2+ currents (Ali and Salter, 2001) and has been implicated in the regulation of the internalization of NMDARs (Vissel et al., 2001; Li et al., 2002). Increased GluN2B-Y1472 phosphorylation promotes the synaptic targeting of NMDARs (Prybylowski et al., 2005). Furthermore, Src-mediated upregulation of NMDARs is thought to play an important role in LTP of CA1 neurons (Groveman et al., 2012; Ohnishi et al., 2011; Trepanier et al., 2012). This is supported by the finding that the level of GluN2B-Y1472 phosphorylation increases following tetanic stimulation in the CA1 region of the hippocampus (Nakazawa et al., 2001). The striatal-enriched tyrosine phosphatase (STEP) dephosphorylates GluN2B-Y1472 (Braithwaite et al., 2006; Pelkey et al., 2002; Snyder et al., 2005) and also inactivates Fyn (Nguyen et al., 2002), therefore both directly and indirectly downregulates synaptic NMDAR expression. Furthermore, synaptic and extrasynaptic NMDARs are differentially phosphorylated at GluN2B-Y1472 and GluN2B-Y1336, respectively (Goebel-Goody et al., 2009), suggesting that modulation of NMDAR tyrosine phosphorylation affects receptor retention and translocation between synaptic and extrasynaptic sites (Gladding and Raymond, 2011).
A recent study raised the intriguing possibility that the direction of synaptic plasticity in CA1 neurons is determined by different classes of GPCRs that differentially target tyrosine phosphorylation sites in GluN2A and GluN2B NMDAR subunits (Fig. 4) via selective activation of Src and Fyn kinases (Yang et al., 2012). The Gαq-coupled pituitary adenylate cyclise activating peptide 1 receptors (PAC1Rs) selectively enhanced the activity of GluN2A-containing NMDARs through the activation of Src kinase. In contrast, the Gαs-coupled dopamine 1 receptors (D1R) enhanced GluN2B-containing NMDARs via selective activation of Fyn (Yang et al., 2012). While PAC1R lowered the threshold for LTP, D1R enhanced LTD indicating that NMDAR-mediated metaplasticity is gated by GPCRs (Yang et al., 2012). These findings are consistent with the notion that the balance between the activities of GluN2A- and GluN2B-containing NMDARs is a key determinant of the direction of synaptic plasticity (Cho et al., 2009; Fox et al., 2006; Liu et al., 2004; Massey et al, 2004) and GPCRs can provide a mechanism by which other neuromodulators affect NMDAR function (section 2. Indirect modulation). Compounds acting via GPCRs to alter the tyrosine phosphorylation status of NMDARs, offer a wide spectrum of possibilities for modulating cognitive function.
3.6.2. Palmitoylation of NMDARs
Palmitatic acid is a saturated fatty acid that is highly abundant in the CNS. Palmitate forms a covalent attachment to proteins via thioester bonds at cysteine (C) residues. This modification is labile, reversible and dynamically regulated by neuronal activity (Hayashi et al., 2009). GluN2A and GluN2B have two potential palmitoylation sites in their C-terminal domains (Cys clusters I and II; Fig 4, Hayashi et al., 2009). Palmitoylation of Cys cluster I controls stable synaptic expression and constitutive internalization of surface NMDARs. De-palmitoylation of Cys cluster II regulates surface delivery of NMDARs (Hayashi et al., 2009). Decreased GluN2B palmitoylation at both clusters is likely to reduce synaptic NMDAR population and increase extrasynaptic NMDAR numbers (Gladding and Raymond, 2011). Therefore, palmitoylation of GluN2 subunits also contributes to regulation of NMDAR trafficking and affects brain function.
3.6.3. Ubiquitination of NMDARs
Ubiquitin is a small (76 amino acid containing) protein that covalently attaches to specific lysine (K) residues in substrate proteins in an ATP-dependent, sequential action of three classes of enzymes, E1-3 (Mabb and Ehlers, 2010). The number and subunit composition of synaptic NMDARs are regulated by activity-dependent protein degradation through the ubiquitin-proteasome system (Ehlers, 2003). Increased synaptic activity leads to upregulation of GluN2A, PSD95 and Homer protein expression and downregulation of GluN1, GluN2B and Shank (Ehlers, 2003). These changes are blocked by proteosomal inhibitors (Ehlers, 2003). Mind bomb-2 (Mib2) E3 ubiquitin ligase interacts with and ubiquitinates the GluN2B NMDAR subunit in a Fyn phosphorylation-dependent manner (Jurd et al., 2008). These findings indicate that ubiquitination is an important mechanism for the removal and degradation of NMDARs that results in dynamic regulation of synaptic strength in response to activity.
3.6.4. Proteolytic cleavage of NMDAR subunits by calpain
The Ca2+-activated protease calpain cleaves the C-terminal domains of GluN2A-C subunits, but not GluN1 (Dong et al., 2006; Guttmann et al., 2001; Simpkins et al., 2003; Doshi and Lynch, 2009; Fig. 4). While the proteolytic truncation of NMDAR subunits removes regulatory and protein-protein interaction sites and reduces synaptic activity, basic ion channel gating and key pharmacological properties are not affected (Guttmann et al., 2001; Simkins et al., 2003). Therefore, it is plausible that calpain cleaved NMDARs remain functional on the cell surface at extrasynaptic sites (Gladding and Raymond, 2011).
3.6.5. Protein binding partners of NMDARs
Like other iGluRs, NMDARs also interact with a wide range of cytoskeletal, scaffolding and signalling proteins (e.g. α-actin-2, AP2, calmodulin, CaMKII, CARPI, COPII, GPS2, LIN7, MAP1S, PACSIN1, plectin, PSD95, RACK1, SALM1, SAP97, SAP102, S-SCAM; Traynelis et al. 2010). An auxiliary subunit, Neto1, has also been described for NMDARs (Ng et al., 2009). Neto1 interacts with an extracellular domain of GluN2 as well as through an intracellular interaction with PSD95. Loss of Neto1 in transgenic mice preferentially results in a loss of synaptic GluN2A expression, with only a modest impact on GluN2B expression, which leads to impaired hippocampal LTP and hippocampal-dependent learning and memory (Ng et al., 2009).
At synapses, NMDARs are stabilised by interaction with PSD95 (Roche et al., 2001; Li et al., 2003). The association of NMDARs with PSD95 and their subsequent endocytosis is regulated by tyrosine phosphorylation. Phosphorylation of GluN2B-Y1472 interferes with binding to PSD95 and promotes clathrin-dependent endocytosis by promoting the binding of AP2 to GluN2B. Dephosphorylation of GluN1-Y837 and GluN2A-Y842 might affect AP2 binding, promoting clathrin-dependent endocytosis in a similar way (Vissel et al., 2001). There is evidence that the synaptic activation of mAChRs leads to a reduction in the surface expression of synaptic NMDARs via the recruitment of hippocalcin, which triggers the exchange of PSD-95 for AP2 to promote endocytosis (Jo et al, 2010).
Intradendritic trafficking of GluN2B subunits requires them to associate with the motor protein KIF17 through LIN10 (Guillaud et al., 2003). Within the PSD, NMDARs are linked by α-actin and spectrin to f-actin, which associates with myosin and might be dynamically regulated by MLCK (myosin light chain kinase; Lei et al., 2001). Synaptic GluN2A containing NMDARs seem to be important for LTP and extrasynaptic GluN2B-containing receptors are involved in LTD (Cull-Candy et al., 2001). In adult neocortical slices de novo LTD induction is enhanced by blockage of glutamate uptake, indicating that the diffusion of glutamate to extrasynaptic GluN2B containing NMDARs triggers LTD (Massey et al., 2004). Furthermore, LTD can be induced after blockade of synaptic NMDARs (Massey et al., 2004). Therefore, NMDAR interactions, localisation and targeting are crucial determinants of synaptic plasticity and consequently cognition.
4. Recently discovered NMDAR PAMs
4.1 Phenanthrene derivatives
A series of 9-substituted phenanthrene-3-carboxylic acid derivatives has been reported to potentiate NMDAR activity with different patterns of GluN2 subunit selectivity (Costa et al., 2010). In an electrophysiological assay on GluN1/GluN2 NMDAR subtypes expressed in Xenopus oocytes, the 9-iodo derivative, UBP512, weakly potentiated GluN2A, had little or no effect on GluN2B and inhibited GluN2C- and GluN2D-containing NMDARs. When tested in the presence of 30-fold higher concentrations of glycine and glutamate UBP512 displayed an enhanced potentiating effect on GluN2A. Potentiation of GluN1/GluN2A by UBP512 due to chelation of Zn2+, resulting in reversal of Zn2+ inhibition, was ruled out in this study (Costa et al., 2010). The 9-cyclopropyl derivative, UBP710 (Fig. 2), potentiated GluN2A and GluN2B but had inhibitory activity on GluN2C or GluN2D-containing NMDARs, when tested at higher concentrations. The 9-i-hexyl derivative, UBP646, however, acted as a universal potentiator on recombinant NMDARs containing GluN2A-D, displaying the greatest potentiating effect on GluN2D.
The NTD is not necessary for the potentiating effect of UBP512 or UBP710, as these compounds still caused potentiation when tested on GluN1/GluN2A receptors without NTDs (Costa et al., 2010). Chimeric receptor studies were used to investigate whether the S1 and S2 regions of the ligand binding domain (LBD) of GluN2A were involved in the NMDAR potentiating activity of UBP512 and UBP710 (Costa et al., 2010). These studies relied on the different modes of action of UBP512 and UBP710 on GluN2 subunits, i.e. the potentiation of GluN2A and inhibition of GluN2C. Swapping the S1 domain of GluN2A with the S1 domain of GluN2C did not affect the potentiating effect of UBP512 and UBP710. However, swapping the S2 domain of GluN2A with the S2 domain of GluN2C converted the compounds to inhibitors, suggesting that they may be binding to the S2 domain or that the S2 domain is involved in the transduction of the potentiating effect.
Examination of the crystal structure of the LBDs of GluN1/GluN2A suggested that Y535 in GluN1 can play a positive allosteric modulatory role by interacting with hydrophobic residues in the dimer interface, thereby stabilizing the dimer interface and slowing deactivation (Furukawa et al., 2005). In support of a role for Y535 in controlling deactivation, the Y535L mutant showed a modest increase in the rates of glycine and glutamate deactivation, whereas the Y535F mutant showed slightly slower deactivation rates (Furukawa et al., 2005). Given that the S2 domain of GluN2A is required for the potentiating effect of UBP512 and UBP710 on GluN2A, it is reasonable to suggest that these compounds are binding at the GluN1/GluN2A dimer interface to block desensitization and/or slow deactivation, perhaps by stabilizing the interaction of Y535 with the hydrophobic site in the dimer interface or by contributing additional stabilization to that provided by Y535. However, it is also possible that these compounds are binding within or in close proximity to the transmembrane region leading to stabilization of the open channel conformation. More work is required to identify the precise binding site(s) and mechanism of action of the UBP compounds.
4.2 Naphthalene derivatives
A naphthalene derivative, 3,5-dihydroxynaphthalene-2-carboxylic acid, UBP551 showed a selective potentiating effect on GluN2D and inhibitory activity on GluN2A-C (Costa et al., 2010). The concentration response curve for the potentiating activity of UBP551 on GluN2D is bell shaped, greatest potentiation was observed at 30 μM and potentiation was reduced at higher concentrations. UBP551 is unique amongst the recently reported potentiators in that it has differential activity on GluN2C and GluN2D. The mechanism underlying the potentiating effect of UBP551 on GluN2D is unknown but it may differ from that of the structurally dissimilar phenanthrene based potentiators.
4.3 Coumarin derivatives
The coumarin derivative, 6-bromo-4-methylcoumarin-3-carboxylic acid, UBP714 (Fig. 2), has been shown to have a weak potentiating effect on recombinant GluN2A-, GluN2B- and GluN2D-containing NMDARs (Fig. 5 and Irvine et al., 2012). UBP714 potentiated field excitatory postsynaptic potentials (fEPSPs) mediated by NMDARs but not those due to AMPARs in the CA1 region of the hippocampus (Fig. 5 and Irvine et al., 2012). Interestingly, an analogue of UBP714, without the 4-methyl group, 6-bromocoumarin-3-carboxylic acid, UBP608, was a moderately potent inhibitor of GluN1/GluN2A with an IC50 value 18.6 μM, suggesting that the methyl group of UBP714 is necessary for potentiating activity (Costa et al., 2010; Irvine et al., 2012).
Fig. 5.
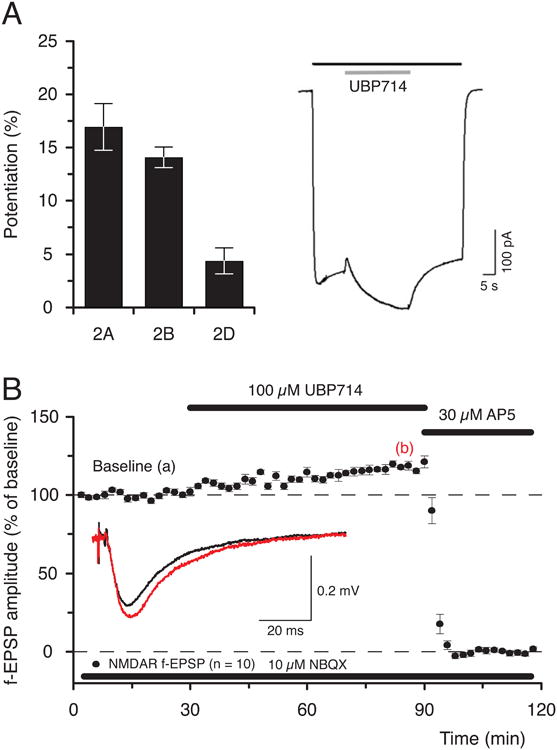
UBP714 potentiates NMDAR responses A. Data (n = 4, mean ± S.E.M.), showing that UBP714 potentiates NMDAR-mediated GluN1/GluN2A (17 ± 2%, 2A), GluN1/GluN2B (14 ± 1%, 2B) and GluN1/GluN2D (4 ± 1%, 2D) responses in Xenopus laevis oocytes (reprinted from Irvine et al, 2012). The trace to the right shows that 100 μM UBP714 (gray bar) potentiates GluN1/GluN2A NMDAR response, which was evoked by applying 10 μM glutamate and 10 μM glycine (black bar). B. Data (n =10) showing that UBP714 potentiates pharmacologically isolated NMDAR-mediated f-EPSPs (inset) in hippocampal slices from adult rat (19 ± 2%, 1-h after the start of application of 100 μM UBP714, reprinted from Irvine et al, 2012).
4.4 Isoquinoline derivatives
A novel structural class of NMDAR potentiator, CIQ ((3-chlorophenyl)(6,7-dimethoxy-1-((4-methoxyphenoxy)methyl)-3,4-dihydroisoquinolin-2-(1H)-yl)methanone, Fig. 2) has been reported to selectively potentiate NMDARs containing GluN2C or GluN2D subunits (Mullasseril et al., 2010). CIQ was found to potentiate recombinant triheteromeric GluN1/GluN2A/GluN2C or GluN1/GluN2A/GluN2D NMDARs, suggesting that only one GluN2C or GluN2D subunit is required for the potentiating effect. Single channel analysis of the effect of CIQ on GluN1/GluN2D suggested that the potentiating effect was due to an increase in channel opening frequency, without altering mean open time. CIQ also potentiated NMDAR currents mediated by GluN2D expressed in subthalamic neurons. CIQ appears to have different structural requirements for its potentiating effect on NMDARs compared to the UBP compounds and PS. Chimeric receptor and point mutation studies suggest that the linker between the NTD and the LBD and T592 in the M1 region of GluN2D are required for the potentiating effect of CIQ. This, coupled with the minimal effect of CIQ on NMDAR deactivation, suggests that the dimer interface is not a likely site for CIQ binding and that the potentiating effect is occurring by a different mechanism to that observed for UBP compounds.
5. Concluding remarks
This review indicates a number of ways in which NMDAR function could be enhanced. NMDARs are ubiquitously expressed throughout the CNS and as such are involved in all functions of neural circuits of the brain and spinal cord. The complexity and multiplicity of NMDARs, its heteromeric structure with intracellular sites for modulation of function and its subtypes offer numerous opportunities for therapeutic interventions. Because of the widely accepted role of NMDARs in plasticity and memory, the obvious approach for cognitive enhancement is potentiation of NMDAR function. However, given the complex roles of NMDARs in synaptic transmission and bidirectional synaptic plasticity the normalization of function could be a better strategy. Putative compounds for consideration will need to have subtle effects; over-stimulation of NMDARs will inter alia likely lead to exacerbation of pain, hyperexcitability and neurodegeneration. Subunit selectivity and limited efficacy may therefore be desirable properties. New compounds with direct but allosteric and specific effects on the NMDAR subunits may offer the most fruitful approach
Highlights.
Cognitive decline is a major problem facing an ageing human population.
NMDA receptors play an important role in neural plasticity and learning and memory.
Excessive activation of NMDA receptors results in deleterious effects.
Subtle fine-tuning of NMDAR function is needed for control of cognitive decline.
Positive allosteric modulators of NMDARs can affect NMDAR mediated events.
Acknowledgments
This work was supported by the MRC (Grants G0601509 and G061812), BBSRC (Grants BB/F012519/1 and BB/J015938/1) and the NIH (Grant MH60252).
Abbreviations
- AChR
acetylcholine receptor
- AKAP79/150
A-kinase anchoring proteins
- AMPAR
AMPA receptor
- CaMKII
Ca2+/calmodulin-dependent protein kinase
- CK2
casein kinase II
- D1R
dopamine 1 receptor
- EPSC
excitatory postsynaptic current
- EPSP
excitatory postsynaptic potential
- fEPSP
field excitatory postsynaptic potentials
- GABAAR
GABAA receptor
- GABABR
GABAB receptor
- GPCR
G-protein-coupled receptors
- iGluR
ionotropic glutamate receptor
- IPSP
inhibitory postsynaptic potential
- LTP
long-term potentiation
- mAChR
muscarinic acetylcholine receptor
- mGluR
metabotropic glutamate receptor
- NMDAR
NMDA receptor
- NMDAR-LTP
NMDA receptor dependent long-term potentiation
- PAC1R
pituitary adenylate cyclase activated peptide 1 receptor
- PKA
protein kinase A
- PKC
protein kinase C
- PSD95
postsynaptic density protein 95
- STEP
striatal-enriched tyrosine phosphatase
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Abraham WC, Gustafsson B, Wigström H. Single high strength afferent volleys can produce long-term potentiation in the hippocampus in vitro. Neuroscience Letters. 1986;70:217–222. doi: 10.1016/0304-3940(86)90466-0. [DOI] [PubMed] [Google Scholar]
- Ali DW, Salter MW. NMDA receptor regulation by Src kinase signaling in excitatory synaptic transmission and plasticity. Curr Opin Neurobiol. 2001;11:336–342. doi: 10.1016/s0959-4388(00)00216-6. [DOI] [PubMed] [Google Scholar]
- Arai AC, Kessler M. Pharmacology of ampakine modulators: from AMPA receptors to synapses and behavior. Curr Drug Targets. 2007;8:583–602. doi: 10.2174/138945007780618490. [DOI] [PubMed] [Google Scholar]
- Arai A, Kessler M, Xiao P, Ambros-Ingerson J, Rogers G, Lynch G. A centrally active drug that modulates AMPA receptor gated currents. Brain Res. 1994;638:343–346. doi: 10.1016/0006-8993(94)90669-6. [DOI] [PubMed] [Google Scholar]
- Ault B, Evans RH, Francis AA, Oakes DJ, Watkins JC. Selective depression of excitatory amino acid induced depolarizations by magnesium ions in isolated spinal cord preparations. J Physiol. 1980;307:413–428. doi: 10.1113/jphysiol.1980.sp013443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayala JE, Chen Y, Banko JL, Sheffler DJ, Williams R, Telk AN, Watson NL, Xiang Z, Zhang Y, Jones PJ, Lindsley CW, Olive MF, Conn PJ. mGluR5 positive allosteric modulators facilitate both hippocampal LTP and LTD and enhance spatial learning. Neuropsychopharmacology. 2009;34:2057–2071. doi: 10.1038/npp.2009.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bashir ZI, Tam B, Collingridge GL. Activation of the glycine site in the NMDA receptor is necessary for the induction of LTP. Neurosci Lett. 1990;108:261–266. doi: 10.1016/0304-3940(90)90651-o. [DOI] [PubMed] [Google Scholar]
- Benveniste M, Mayer ML. Kinetic analysis of antag- onist action at N-methyl-o-aspartic acid receptors. Two binding sites each for glutamate and glycine. Biophys J. 1991;59:560–573. doi: 10.1016/S0006-3495(91)82272-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger AJ, Dieudonné S, Ascher P. Glycine uptake governs glycine site occupancy at NMDA receptors of excitatory synapses. J Neurophysiol. 1998;80:3336–3340. doi: 10.1152/jn.1998.80.6.3336. [DOI] [PubMed] [Google Scholar]
- Bergeron R, Meyer TM, Coyle JT, Greene RW. Modulation of N-methyl-D-aspartate receptor function by glycine transport. Proc Natl Acad Sci U S A. 1998;95:15730–15734. doi: 10.1073/pnas.95.26.15730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bortolotto ZA, Collett VJ, Conquet F, Jia Z, van der Putten H, Collingridge GL. The regulation of hippocampal LTP by the molecular switch, a form of metaplasticity, requires mGlu5 receptors. Neuropharmacology. 2005;49:13–25. doi: 10.1016/j.neuropharm.2005.05.020. [DOI] [PubMed] [Google Scholar]
- Bowlby MR. Pregnenolone sulfate potentiation of N-methyl-D-aspartate receptor channels in hippocampal neurons. Mol Pharmacol. 1993;43:813–819. [PubMed] [Google Scholar]
- Braithwaite SP, Adkisson M, Leung J, Nava A, Masterson B, Urfer R, Oksenberg D, Nikolich K. Regulation of NMDA receptor trafficking and function by striatal-enriched tyrosine phosphatase (STEP) Eur J Neurosci. 2006;23:2847–2856. doi: 10.1111/j.1460-9568.2006.04837.x. [DOI] [PubMed] [Google Scholar]
- Bresink I, Benke TA, Collett VJ, Seal AJ, Parsons CG, Henley JM, Collingridge GL. Effects of memantine on recombinant rat NMDA receptors expressed in HEK 293 cells. Br J Pharmacol. 1996;119:195–204. doi: 10.1111/j.1476-5381.1996.tb15971.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchanan KA, Petrovic MM, Chamberlain SEL, Marrion NV, Mellor JR. Facilitation of long-term potentiation by muscarinic M(1) receptors is mediated by inhibition of SK channels. Neuron. 2010;68:948–963. doi: 10.1016/j.neuron.2010.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cameron K, Bartle E, Roark R, Fanelli D, Pham M, Pollard B, Borkowski B, Rhoads S, Kim J, Rocha M, Kahlson M, Kangala M, Gentile L. Neurosteroid binding to the amino terminal and glutamate binding domains of ionotropic glutamate receptors. Steroids. 2012 doi: 10.1016/j.steroids.2012.03.011. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- Chatterton JE, Awobuluyi M, Premkumar LS, Takahashi H, Talantova M, Shin y, Cui J, Tu S, Sevarino KA, Nakanishi N, Tong G, Lipton SA, Zhang D. Excitatory glycine receptors containing the NR3 family of NMDA receptor subunits. Nature. 2002;415:793–798. doi: 10.1038/nature715. [DOI] [PubMed] [Google Scholar]
- Chen L, Cai W, Chen L, Zhou R, Furuya K, Sokabe M. Modulatory metaplasticity induced by pregnenolone sulfate in the rat hippocampus: a leftward shift in LTP/LTD-frequency curve. Hippocampus. 2009;20:499–512. doi: 10.1002/hipo.20649. [DOI] [PubMed] [Google Scholar]
- Chen L, Miyamoto Y, Furuya K, Dai XN, Mori N, Sokabe M. Chronic DHEAS administration facilitates hippocampal long-term potentiation via an amplification of Src-dependent NMDA receptor signaling. Neuropharmacology. 2006;51:659–670. doi: 10.1016/j.neuropharm.2006.05.011. [DOI] [PubMed] [Google Scholar]
- Chen L, Muhlhauser M, Yang CR. Glycine tranporter-1 blockade potentiates NMDA-mediated responses in rat prefrontal cortical neurons in vitro and in vivo. J Neurophysiol. 2003;89:691–703. doi: 10.1152/jn.00680.2002. [DOI] [PubMed] [Google Scholar]
- Cho KK, Khibnik L, Philpot BD, Bear MF. The ratio of NR2A/B NMDA receptor subunits determines the qualities of ocular dominance plasticity in visual cortex. Proc Natl Acad Sci USA. 2009;106:5377–5382. doi: 10.1073/pnas.0808104106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chumakov I, Blumenfeld M, Guerassimenko O, Cavarec L, Palicio M, Abderrahim H, Bougueleret L, Barry C, Tanaka H, La Rosa P, Puech A, Tahri N, Cohen-Akenine A, Delabrosse S, Lissarrague S, Picard FP, Maurice K, Essioux L, Millasseau P, Grel P, Debailleul V, Simon AM, Caterina D, Dufaure I, Malekzadeh K, Belova M, Luan JJ, Bouillot M, Sambucy JL, Primas G, Saumier M, Boubkiri N, Martin-Saumier S, Nasroune M, Peixoto H, Delaye A, Pinchot V, Bastucci M, Guillou S, Chevillon M, Sainz-Fuertes R, Meguenni S, Aurich-Costa J, Cherif D, Gimalac A, Van Duijn C, Gauvreau D, Ouellette G, Fortier I, Raelson J, Sherbatich T, Riazanskaia N, Rogaev E, Raeymaekers P, Aerssens J, Konings F, Luyten W, Macciardi F, Sham PC, Straub RE, Weinberger DR, Cohen N, Cohen D. Genetic and physiological data implicating the new human gene G72 and the gene for D-amino acid oxidase in schizophrenia. Proc Natl Acad Sci USA. 2002;99:13675–13680. doi: 10.1073/pnas.182412499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung HJ, Huang YH, Lau LF, Huganir RL. Regulation of the NMDA receptor complex and trafficking by activity-dependent phosphorylation of the NR2B subunit PDZ ligand. J Neurosci. 2004;24:10248–10259. doi: 10.1523/JNEUROSCI.0546-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clements JD, Westbrook GL. Activation kinetics reveal the number of glutamate and glycine binding sites on the N-methyl-D-aspartate receptor. Neuron. 1991;7:605–613. doi: 10.1016/0896-6273(91)90373-8. [DOI] [PubMed] [Google Scholar]
- Cleva RM, Gass JT, Widholm JJ, Olive MF. Glutamatergic targets for enhancing extinction learning in drug addiction. Curr Neuropharmacol. 2010;8:394–408. doi: 10.2174/157015910793358169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coan EJ, Irving AJ, Collingridge GL. Low-frequency activation of the NMDA receptor system can prevent the induction of LTP. Neuroscience Letters. 1989;105:205–210. doi: 10.1016/0304-3940(89)90038-4. [DOI] [PubMed] [Google Scholar]
- Colledge M, Dean RA, Scott GK, Langeberg LK, Huganir RL, Scott JD. Targeting of PKA to glutamate receptors through a MAGUK-AKAP complex. Neuron. 2000;27:107–119. doi: 10.1016/s0896-6273(00)00013-1. [DOI] [PubMed] [Google Scholar]
- Collingridge GL. Long-term potentiation in the hippocampus - mechanisms of initiation and modulation by neurotransmitters. Trends in Pharmacological Sciences. 1985;10:407–411. [Google Scholar]
- Collingridge GL, Herron CE, Lester RA. Synaptic activation of N-methyl-D-aspartate receptors in the Schaffer collateral-commissural pathway of rat hippocampus. The Journal of Physiology. 1988;399:283–300. doi: 10.1113/jphysiol.1988.sp017080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collingridge GL, Kehl SJ, McLennan H. Excitatory amino acids in synaptic transmission in the Schaffer collateral-commissural pathway of the rat hippocampus. The Journal of Physiology. 1983;334:33–46. doi: 10.1113/jphysiol.1983.sp014478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corpéchot C, Synguelakis M, Talha S, Axelson M, Sjodvall J, Vihko R, Baulieu EE, Robel P. Pregnenolone and its sulfate ester in the rat brain. Brain Res. 1983;270:119–125. doi: 10.1016/0006-8993(83)90797-7. [DOI] [PubMed] [Google Scholar]
- Costa BM, Irvine MW, Fang G, Eaves RJ, Mayo-Martin MB, Skifter DA, Jane DE, Monaghan DT. A novel family of negative and positive allosteric modulators of NMDA receptors. J Pharmacol Exp Ther. 2010;335(3):614–621. doi: 10.1124/jpet.110.174144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croucher MJ, Collins JF, Meldrum BS. Anticonvulsant action of excitatory amino acid antagonists. Science. 1982;216:899–901. doi: 10.1126/science.7079744. [DOI] [PubMed] [Google Scholar]
- Crump FT, Dillman KS, Craig AM. cAMP-dependent protein kinase mediates activity-regulated synaptic targeting of NMDA receptors. J Neurosci. 2001;21:5079–5088. doi: 10.1523/JNEUROSCI.21-14-05079.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cull-Candy S, Brickley S, Farrant M. NMDA receptor subunits: diversity, development and disease, Curr. Opin Neurobiol. 2001;11:327–335. doi: 10.1016/s0959-4388(00)00215-4. [DOI] [PubMed] [Google Scholar]
- Danysz W, Parsons CG. The NMDA receptor antagonist memantine as a symptomatological and neuroprotective treatment for Alzheimer's disease: preclinical evidence. Int J Geriat Psychiatry. 2003;18:S23–S32. doi: 10.1002/gps.938. [DOI] [PubMed] [Google Scholar]
- Darrah JM, Stefani MR, Moghaddam B. Interaction of N-methyl-D-aspartate and group 5 metabotropic glutamate receptors on behavioral flexibility using a novel operant set-shift paradigm. Behav Pharmacol. 2008;19:225–234. doi: 10.1097/FBP.0b013e3282feb0ac. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies CH, Collingridge GL. The physiological regulation of synaptic inhibition by GABAB autoreceptors in rat hippocampus. The Journal of Physiology. 1993;472:245–265. doi: 10.1113/jphysiol.1993.sp019945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies CH, Collingridge GL. Regulation of EPSPs by the synaptic activation of GABAB autoreceptors in rat hippocampus. The Journal of Physiology. 1996;496(Pt 2):451–470. doi: 10.1113/jphysiol.1996.sp021698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies CH, Davies SN, Collingridge GL. Paired-pulse depression of monosynaptic GABA-mediated inhibitory postsynaptic responses in rat hippocampus. The Journal of Physiology. 1990;424:513–531. doi: 10.1113/jphysiol.1990.sp018080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies SN, Lodge D. Evidence for involvement of N-methylaspartate receptors in ‘wind-up’ of class 2 neurones in the dorsal horn of the rat. Brain Res. 1987;424:402–406. doi: 10.1016/0006-8993(87)91487-9. [DOI] [PubMed] [Google Scholar]
- Davies CH, Pozza MF, Collingridge GL. CGP 55845A: a potent antagonist of GABAB receptors in the CA1 region of rat hippocampus. Neuropharmacology. 1993;32:1071–1073. doi: 10.1016/0028-3908(93)90073-c. [DOI] [PubMed] [Google Scholar]
- Davies CH, Starkey SJ, Pozza MF, Collingridge GL. GABA autoreceptors regulate the induction of LTP. Nature. 1991;349:609–611. doi: 10.1038/349609a0. [DOI] [PubMed] [Google Scholar]
- DeVito LM, Balu DT, Kanter BR, Lykken C, Basu AC, Coyle JT, Eichenbaum H. Serine racemase deletion disrupts memory for order and alters cortical dendritic morphology. Genes Brain Behav. 2011;10:210–222. doi: 10.1111/j.1601-183X.2010.00656.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diamond DM, Dunwiddie TV, Rose GM. Characteristics of hippocampal primed burst potentiation in vitro and in the awake rat. J Neurosci. 1988;8:4079–4088. doi: 10.1523/JNEUROSCI.08-11-04079.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dingledine R, Hynes MA, King GL. Involvement of N-methyl-D-aspartate receptors in epileptiform bursting in the rat hippocampal slice. J Physiol. 1986;380:175–189. doi: 10.1113/jphysiol.1986.sp016279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doherty AJ, Palmer MJ, Henley JM, Collingridge GL, Jane DE. (RS)-2-chloro-5-hydroxyphenylglycine (CHPG) activates mGlu5, but no mGlu1, receptors expressed in CHO cells and potentiates NMDA responses in the hippocampus. Neuropharmacology. 1997;36:265–267. doi: 10.1016/s0028-3908(97)00001-4. [DOI] [PubMed] [Google Scholar]
- Dong YN, Wu HY, Hsu FC, Coulter DA, Lynch DR. Developmental and cell-selective variations in N-methyl-D-aspartate receptor degradation by calpain. J Neurochem. 2006;99:206–217. doi: 10.1111/j.1471-4159.2006.04096.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doshi S, Lynch DR. Calpain and the glutamatergic synapse. Front Biosci (Schol Ed) 2009;1:466–476. doi: 10.2741/s38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dubrovsky BO. Steroids, neuroactive steroids and neurosteroids in psychopathology. Prog Neuropsychopharmacol Biol Psychiatry. 2005;29:169–192. doi: 10.1016/j.pnpbp.2004.11.001. [DOI] [PubMed] [Google Scholar]
- Duffy S, Labrie V, Roder JC. D-serine augments NMDA-NR2B receptor- dependent hippocampal long-term depression and spatial reversal learning. Neuropsychopharmacology. 2008;33:1004–1018. doi: 10.1038/sj.npp.1301486. [DOI] [PubMed] [Google Scholar]
- Ehlers MD. Activity level controls postsynaptic composition and signaling via the ubiquitin-proteasome system. Nat Neurosci. 2003;6:231–242. doi: 10.1038/nn1013. [DOI] [PubMed] [Google Scholar]
- Ehlers MD, Tingley WG, Huganir RL. Regulated subcellular distribution of the NR1 subunit of the NMDA receptor. Science. 1995;269:1734–1737. doi: 10.1126/science.7569904. [DOI] [PubMed] [Google Scholar]
- Ehlers MD, Zhang S, Bernhadt JP, Huganir RL. Inactivation of NMDA receptors by direct interaction of calmodulin with the NR1 subunit. Cell. 1996;84:745–755. doi: 10.1016/s0092-8674(00)81052-1. [DOI] [PubMed] [Google Scholar]
- Elfverson M, Johansson T, Zhou Q, Le Grevès P, Nyberg F. Chronic administration of the anabolic androgenic steroid nandrolone alters neurosteroid action at the sigma-1 receptor but not at the sigma-2 or NMDA receptors. Neuropharmacology. 2011;61:1172–1181. doi: 10.1016/j.neuropharm.2011.01.005. [DOI] [PubMed] [Google Scholar]
- Ferraris DV, Tsukamoto T. Recent advances in the discovery of D-amino acid oxidase inhibitors and their therapeutic utility in schizophrenia. Curr Pharm Des. 2011;17:103–111. doi: 10.2174/138161211795049633. [DOI] [PubMed] [Google Scholar]
- Fitzjohn SM, Doherty AJ, Collingridge GL. The use of the hippocampal slice preparation in the study of Alzheimer's disease. European Journal of Pharmacology. 2008;585:50–59. doi: 10.1016/j.ejphar.2008.02.077. [DOI] [PubMed] [Google Scholar]
- Fitzjohn SM, Irving AJ, Palmer MJ, Harvey J, Lodge D, Collingridge GL. Activation of group I mGluRs potentiates NMDA responses in rat hippocampal slices. Neuroscience Letters. 1996;203:211–213. doi: 10.1016/0304-3940(96)12301-6. [DOI] [PubMed] [Google Scholar]
- Fletcher EJ, Lodge D. Glycine reverses antagonism of N-methyl-D-aspartate (NMDA) by 1-hydroxy-3-aminopyrrolidone-2 (HA-966) but not by D-2-amino-5-phosphonovalerate (D-AP5) on rat cortical slices. Eur J Pharmacol. 1988;151:161–162. doi: 10.1016/0014-2999(88)90711-x. [DOI] [PubMed] [Google Scholar]
- Flood JF, Morley JE, Roberts E. Memory-enhancing effects in male mice of pregnenolone and steroids metabolically derived from it. Proc Natl Acad Sci USA. 1992;89:1567–1571. doi: 10.1073/pnas.89.5.1567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flood JF, Morley JE, Roberts E. Pregnenolone sulfate enhances post-training memory processes when injected in very low doses into limbic system structures: the amygdala is by far the most sensitive. Proc Natl Acad Sci USA. 1995;92:10806–10810. doi: 10.1073/pnas.92.23.10806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fong DK, Rao A, Crump FT, Craig AM. Rapid synaptic remodeling by protein kinase C: reciprocal translocation of NMDA receptors and calcium/calmodulin-dependent kinase II. J Neurosci. 2002;22:2153–2164. doi: 10.1523/JNEUROSCI.22-06-02153.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fossat P, Turpin FR, Sacchi S, Dulong J, Shi T, Rivet JM, Sweedler JV, Pollegioni L, Millan MJ, Oliet SH, Mothet JP. Glial D-serine gates NMDA receptors at excitatory synapses in prefrontal cortex. Cereb Cortex. 2012;22:595–606. doi: 10.1093/cercor/bhr130. [DOI] [PubMed] [Google Scholar]
- Foster AC, Kemp JA. HA-966 antagonizes N-methyl-D-aspartate receptors through a selective interaction with the glycine modulatory site. J Neurosci. 1989;9:2191–2196. doi: 10.1523/JNEUROSCI.09-06-02191.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox CJ, Russell KI, Wang YT, Christie BR. Contribution of NR2A and NR2B NMDA subunits to bidirectional synaptic plasticity in the hippocampus in vivo. Hippocampus. 2006;16:907–915. doi: 10.1002/hipo.20230. [DOI] [PubMed] [Google Scholar]
- Frankiewicz T, Potier B, Bashir ZI, Collingridge GL, Parsons CG. Effects of memantine and MK-801 on NMDA-induced currents in cultured neurones and on synaptic transmission and LTP in area CA1 of rat hippocampal slices. Br J Pharmacol. 1996;117:689–697. doi: 10.1111/j.1476-5381.1996.tb15245.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furukawa H, Singh SK, Mancusso R, Gouaux E. Subunit arrangement and function in NMDA receptors. Nature. 2005;438:185–192. doi: 10.1038/nature04089. [DOI] [PubMed] [Google Scholar]
- Gastambide F, Cotel MC, Gilmour G, O'Neill MJ, Robbins TW, Tricklebank MD. Selective remediation of reversal learning deficits in the neurodevelopmental MAM model of schizophrenia by a novel mGlu5 positive allosteric modulator. Neuropsychopharmacology. 2012;37:1057–1066. doi: 10.1038/npp.2011.298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gladding CM, Raymond LA. Mechanisms underlying NMDA receptor synaptic/extrasynaptic distribution. Mol Cell Neurosci. 2011;48:308–320. doi: 10.1016/j.mcn.2011.05.001. [DOI] [PubMed] [Google Scholar]
- Gobel-Goody SM, Davies KD, Alvestad Linger RM, Freund RK, Browning MD. Phospho-regulation of synaptic and extrasynaptic N-methyl-D-aspartate receptors in adult hippocampal slices. Neuroscience. 2009;158:1446–1459. doi: 10.1016/j.neuroscience.2008.11.006. [DOI] [PubMed] [Google Scholar]
- Groveman BR, Feng S, Fang XQ, Pflueger M, Lin SX, Bienkiewicz EA, Yu X. The regulation of N-methyl-D-aspartate receptors by Src kinase. FEBS J. 2012;279:20–28. doi: 10.1111/j.1742-4658.2011.08413.x. [DOI] [PubMed] [Google Scholar]
- Guillaud L, Setou M, Hirokawa N. KIF17 dynamics and regulation of NR2B trafficking in hippocampal neurons. J Neurosci. 2003;23:131–140. doi: 10.1523/JNEUROSCI.23-01-00131.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guttmann RP, Baker DL, Seifert KM, Cohen AS, Coulter DA, Lynch DR. Specific proteolysis of the NR2 subunit at multiple sites by calpain. J Neurochem. 2001;78:1083–1093. doi: 10.1046/j.1471-4159.2001.00493.x. [DOI] [PubMed] [Google Scholar]
- Harvey J, Balasubramaniam R, Collingridge GL. Carbachol can potentiate N-methyl-D-aspartate responses in the rat hippocampus by a staurosporine and thapsigargin-insensitive mechanism. Neuroscience Letters. 1993;162:165–168. doi: 10.1016/0304-3940(93)90586-a. [DOI] [PubMed] [Google Scholar]
- Hayashi T, Thomas GM, Huganir RL. Dual palmitoylation of NR2 subunits regulates NMDA receptor trafficking. Neuron. 2009;64:213–226. doi: 10.1016/j.neuron.2009.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henneberger C, Papouin T, Oliet SH, Rusakov DA. Long-term potentiation depends on release of d-serine from astrocytes. Nature. 2010;463:232–236. doi: 10.1038/nature08673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heresco-Levy U, Javitt DC, Ermilov M, Mordel C, Horowitz A, Kelly D. Double-blind, placebo-controlled, crossover trial of glycine adjuvant therapy for treatment-resistant schizophrenia. Br J Psychiatry. 1996;169:610–617. doi: 10.1192/bjp.169.5.610. [DOI] [PubMed] [Google Scholar]
- Herron CE, Williamson R, Collingridge GL. A selective N-methyl-D-aspartate antagonist depresses epileptiform activity in rat hippocampal slices. Neurosci Lett. 1985;61:255–60. doi: 10.1016/0304-3940(85)90473-2. [DOI] [PubMed] [Google Scholar]
- Hisatsune C, Umemori H, Inoue T, Michikawa T, Kohda K, Mikoshiba K, Yamamoto T. Phosphorylation-dependent regulation of N-methyl-D-aspartate receptors by calmodulin. J Biol Chem. 1997;272:20805–20810. doi: 10.1074/jbc.272.33.20805. [DOI] [PubMed] [Google Scholar]
- Hood WF, Sun ET, Compton RP, Monahan JB. 1-Aminocyclobutane-1- carboxylate (ACBC): a specific antagonist of the N-methyl-D-aspartate receptor coupled glycine receptor. Eur J Pharmacol. 1989;161:281–282. doi: 10.1016/0014-2999(89)90861-3. [DOI] [PubMed] [Google Scholar]
- Horak M, Vlcek K, Petrovic M, Chodounska H, Vyklicky L., Jr Molecular mechanism of pregnenolone sulfate action at NR1/NR2B receptors. J Neurosci. 2004;24:10318–10325. doi: 10.1523/JNEUROSCI.2099-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horak M, Vlcek K, Chodounska H, Vyklicky L., Jr Subtype-dependence of N-methyl-D-aspartate receptor modulation by pregnenolone sulfate. Neuroscience. 2006;137:93–102. doi: 10.1016/j.neuroscience.2005.08.058. [DOI] [PubMed] [Google Scholar]
- Irvine MW, Costa BM, Volianskis A, Fang G, Ceolin L, Collingridge GL, Monaghan DT, Jane DE. Coumarin-3-carboxylic acid derivatives as inhibitors and potentiators of recombinant and native N-methyl-D-aspartate receptors. Neurochemistry International. 2012 doi: 10.1016/j.neuint.2011.12.020. In Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isaacson JS, Solis JM, Nicoll RA. Local and diffuse synaptic actions of GABA in the hippocampus. Neuron. 1993;10:165–175. doi: 10.1016/0896-6273(93)90308-e. [DOI] [PubMed] [Google Scholar]
- Ito I, Tanabe S, Kohda A, Sugiyama H. Allosteric potentiation of quisqualate receptors by a nootropic drug aniracetam. J Physiol. 1990;424:533–543. doi: 10.1113/jphysiol.1990.sp018081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang MK, Mierke DF, Russek SJ, Farb DH. A steroid modulatory domain on NR2B controls N-methyl-D-aspartate receptor proton sensitivity. Proc Natl Acad Sci USA. 2004;101:8198–8203. doi: 10.1073/pnas.0401838101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin R, Clark S, Weeks AM, Dudman JT, Gouaux E, Partin KM. Mechanism of positive allosteric modulators acting on AMPA receptors. J Neurosci. 2005;25:9027–9036. doi: 10.1523/JNEUROSCI.2567-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jo J, Son GH, Winters BL, Kim MJ, Whitcomb DJ, Dickinson BA, Lee YB, Futai K, Amici M, Sheng M, Collingridge GL, Cho K. Muscarinic receptors induce LTD of NMDAR EPSCs via a mechanism involving hippocalcin, AP2 and PSD-95. Nat Neurosci. 2010;13:1216–1224. doi: 10.1038/nn.2636. [DOI] [PubMed] [Google Scholar]
- Johnson JW, Ascher P. Glycine potentiates the NMDA response in cultured mouse brain neurons. Nature. 1987;325:529–531. doi: 10.1038/325529a0. [DOI] [PubMed] [Google Scholar]
- Jurd R, Thornton C, Wang J, Luong K, Phamluong K, Kharazia V, Gibb SL, Ron D. Mind bomb-2 is an E3 ligase that ubiquitinates the N-methyl-D-aspartate receptor NR2B subunit in a phosphorylation-dependent manner. J Biol Chem. 2008;283:301–310. doi: 10.1074/jbc.M705580200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemp JA, Foster AC, Leeson PD, Priestley T, Tridgett R, Iversen LL, Woodruff GN. 7-Chlorokynurenic acid is a selective antagonist at the glycine modulatory site of the N-methyl-D-aspartate receptor complex. Proc Natl Acad Sci USA. 1988;85:6547–6550. doi: 10.1073/pnas.85.17.6547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klauck TM, Faux MC, Labudda K, Langeberg LK, Jaken S, Scott JD. Coordination of three signaling enzymes by AKAP79, a mammalian scaffold protein. Science. 1996;271:1589–1592. doi: 10.1126/science.271.5255.1589. [DOI] [PubMed] [Google Scholar]
- Kleckner NW, Dingledine R. Requirement for glycine in activation of NMDA receptors expressed in Xenopus oocytes. Science. 1988;241:835–837. doi: 10.1126/science.2841759. [DOI] [PubMed] [Google Scholar]
- Kloda A, Clements JD, Lewis RJ, Adams DJ. Adenosine triphosphate acts as both a competitive antagonist and a positive allosteric modulator at recombinant N-methyl-D-aspartate receptors. Mol Pharmacol. 2004;65:1386–1396. doi: 10.1124/mol.65.6.1386. [DOI] [PubMed] [Google Scholar]
- Kopec K, Flood DG, Gasior M, McKenna BA, Zuvich E, Schreiber J, Salvino JM, Durkin JT, Ator MA, Marino MJ. Glycine transporter (GlyT1) inhibitors with reduced residence time increase prepulse inhibition without inducing hyperlocomotion in DBA/2 mice. Biochem Pharmacol. 2010;80:1407–1417. doi: 10.1016/j.bcp.2010.07.004. [DOI] [PubMed] [Google Scholar]
- Korinek M, Kapras V, Vyklicky V, Adamusova E, Borovska J, Vales K, Stuchlik A, Horak M, Chodounska H, Vyklicky L., Jr Neurosteroid modulation of N-methyl-D-aspartate receptors: molecular mechanism and behavioral effects. Steroids. 2011;76:1409–1418. doi: 10.1016/j.steroids.2011.09.002. [DOI] [PubMed] [Google Scholar]
- Kostakis E, Jang MK, Russek SJ, Gibbs TT, Farb DH. A steroid modulatory domain in NR2A collaborates with NR1 exon-5 to control NMDAR modulation by pregnenolone sulfate and protons. J Neurochem. 2011;119:486–496. doi: 10.1111/j.1471-4159.2011.07442.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroker KS, Rast G, Rosenbrock H. Differential effect of the mGlu5 receptor positive allosteric modulator ADX-47273 on early and late hippocampal LTP. Neuropharmacology. 2011;61:707–714. doi: 10.1016/j.neuropharm.2011.05.014. [DOI] [PubMed] [Google Scholar]
- Kuryatov A, Laube B, Betz H, Kuhse J. Mutational analysis of the glycine-binding site of the NMDA receptor: structural similarity with bacterial amino acid-binding proteins. Neuron. 1994;12:1291–300. doi: 10.1016/0896-6273(94)90445-6. [DOI] [PubMed] [Google Scholar]
- Larson J, Wong D, Lynch G. Patterned stimulation at the theta frequency is optimal for the induction of hippocampal long-term potentiation. Brain Res. 1986;368:347–350. doi: 10.1016/0006-8993(86)90579-2. [DOI] [PubMed] [Google Scholar]
- Lane HY, Chang YC, Liu YC, Chiu CC, Tsai GE. Sarcosine or D-serine add-on treatment for acute exacerbation of schizophrenia: a randomized, double-blind, placebo-controlled study. Arch Gen Psychiatry. 2005;62:1196–1204. doi: 10.1001/archpsyc.62.11.1196. [DOI] [PubMed] [Google Scholar]
- Lanté F, Cavalier M, Cohen-Solal C, Guiramand J, Vignes M. Developmental switch from LTD to LTP in low frequency-induced plasticity. Hippocampus. 2006;16:981–989. doi: 10.1002/hipo.20228. [DOI] [PubMed] [Google Scholar]
- Lau LF, Huganir RL. Differential tyrosine phosphorylation of N-methyl-D-aspartate receptor subunits. J Biol Chem. 1995;270:20036–20041. doi: 10.1074/jbc.270.34.20036. [DOI] [PubMed] [Google Scholar]
- Lei S, Czerwinska E, Czerwinski W, Walsh MP, McDonald JF. Regulation of NMDA receptor activity by F-actin and myosin light chain kinase. J Neurosci. 2001;21:8464–8472. doi: 10.1523/JNEUROSCI.21-21-08464.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonard AS, Hell JW. Cyclic AMP-dependent protein kinase and protein kinase C phosphorylate N-methyl-D-aspartate receptors at different sites. J Biol Chem. 1997;272:12107–12115. doi: 10.1074/jbc.272.18.12107. [DOI] [PubMed] [Google Scholar]
- Li B, Chen N, Luo T, Otsu Y, Murphy TH, Raymond LA. Differential regulation of synaptic and extra-synaptic NMDA receptors. Nat Neurosci. 2002;5:833–834. doi: 10.1038/nn912. [DOI] [PubMed] [Google Scholar]
- Li B, Otzu Y, Murphy TH, Raymond LA. Developmental decrease in NMDA receptor desensitization associated with shift to synapse and tnteraction with postsynaptic density-95. J Neurosci. 2003;23:11244–11254. doi: 10.1523/JNEUROSCI.23-35-11244.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao GY, Wagner DA, Hsu MH, Leonard JP. Evidence for direct protein kinase C mediated modulation of N-methyl-D-aspartate receptor current. Mol Pharmacol. 2001;59:960–964. doi: 10.1124/mol.59.5.960. [DOI] [PubMed] [Google Scholar]
- Liu F, Grauer S, Kelley C, Navarra R, Graf R, Zhang G, Atkinson PJ, Popiolek M, Wantuch C, Khawaja X, Smith D, Olsen M, Kouranova E, Lai M, Pruthi F, Pulicicchio C, Day M, Gilbert A, Pausch MH, Brandon NJ, Beyer CE, Comery TA, Logue S, Rosenzweig-Lipson S, Marquis KL. ADX47273 [S-(4-fluoro-phenyl)-{3-[3-(4-fluoro-phenyl)-[1,2,4]-oxadiazol-5-yl]-piperidin-1-yl}-methanone]: a novel metabotropic glutamate receptor 5-selective positive allosteric modulator with preclinical antipsychotic-like and procognitive activities. J Pharmacol Exp Ther. 2008;327:827–839. doi: 10.1124/jpet.108.136580. [DOI] [PubMed] [Google Scholar]
- Liu XY, Chu XP, Mao LM, Wang M, Lan HX, Li MH, Zhang GC, Parelkar NK, Fibuch EE, Haines M, Neve KA, Liu F, Xiong ZG, Wang JQ. Modulation of D2R-NR2B interactions in response to cocaine. Neuron. 2006;52:897–909. doi: 10.1016/j.neuron.2006.10.011. [DOI] [PubMed] [Google Scholar]
- Lynch G, Palmer LC, Gall CM. The likelihood of cognitive enhancement. Pharmacol Biochem Behav. 2011;99:116–29. doi: 10.1016/j.pbb.2010.12.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mabb AM, Ehlers MD. Ubiquitin in postsynaptic function and plasticity. Annu Rev Cell Dev Biol. 2010;26:179–210. doi: 10.1146/annurev-cellbio-100109-104129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madry C, Mesic I, Betz H, Laube B. The N-terminal domains of both NR1 and NR2 subunits determine allosteric Zn2+ inhibition and glycine affinity of N-methyl-D-aspartate receptors. Mol Pharmacol. 2007;72:1535–44. doi: 10.1124/mol.107.040071. [DOI] [PubMed] [Google Scholar]
- Maekawa M, Watanabe M, Yamaguchi S, Konno SR, Hori Y. Spatial learning and long-term potentiation of mutant mice lacking d-amino-acid oxidase. Neurosci Res. 2005;53:34–38. doi: 10.1016/j.neures.2005.05.008. [DOI] [PubMed] [Google Scholar]
- Manahan-Vaughan D, Ngomba RT, Storto M, Kulla A, Catania MV, Chiechio S, Rampello L, Passarelli F, Capece A, Reymann KG, Nicoletti F. An increased expression of the mGlu5 receptor protein following LTP induction at the perforant path-dentate gyrus synapse in freely moving rats. Neuropharmacology. 2003;44:17–25. doi: 10.1016/s0028-3908(02)00342-8. [DOI] [PubMed] [Google Scholar]
- Manahan-Vaughan D, Wildförster V, Thomsen C. Rescue of hippocampal LTP and learning deficits in a rat model of psychosis by inhibition of glycine transporter-1 (GlyT1) Eur J Neurosci. 2008;28:1342–1350. doi: 10.1111/j.1460-9568.2008.06433.x. [DOI] [PubMed] [Google Scholar]
- Martina M, Gorfinkel Y, Halman S, Lowe JA, Periyalwar P, Schmidt CJ, Bergeron R. Glycine transporter type 1 blockade changes NMDA receptor-mediated responses and LTP in hippocampal CA1 pyramidal cells by altering extracellular glycine levels. J Physiol. 2004;557:489–500. doi: 10.1113/jphysiol.2004.063321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marx CE, Bradford DW, Hamer RM, Naylor JC, Allen TB, Lieberman JA, Strauss JL, Kilts JD. Pregnenolone as a novel therapeutic candidate in schizophrenia: emerging preclinical and clinical evidence. Neuroscience. 2011;191:78–90. doi: 10.1016/j.neuroscience.2011.06.076. [DOI] [PubMed] [Google Scholar]
- Massey PV, Johnson BE, Moult PR, Auberson YP, Brown MW, Molnar E, Collingridge GL, Bashir ZI. Differential roles of NR2A and NR2B-containing NMDA receptors in cortical long-term potentiation and long-term depression. J Neurosci. 2004;24:7821–7828. doi: 10.1523/JNEUROSCI.1697-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer ML, Westbrook GL, Guthrie PB. Voltage-dependent block by Mg2+ of NMDA responses in spinal cord neurones. Nature. 1984;309:261–3. doi: 10.1038/309261a0. [DOI] [PubMed] [Google Scholar]
- Mayo W, Dellu F, Robel P, Cherkaoui J, Le Moal M, Baulieu EE, Simon H. Infusion of neurosteroids into the nucleus basalis magnocellularis affects cognitive processes in the rat. Brain Res. 1993;607:324–328. doi: 10.1016/0006-8993(93)91524-v. [DOI] [PubMed] [Google Scholar]
- Molnár E. Molecular organization and regulation of glutamate receptors in developing and adult mammalian central nervous systems. In: Lajtha A, Vizi ES, editors. Handbook of Neurochemistry and Molecular Neurobiology: Neurotransmitter Systems. Third. Springer; 2008. pp. 415–441. Reference. [Google Scholar]
- Mony L, Kew JNC, Gunthorpe MJ, Paoletti P. Allosteric modulators of NR2B-containing NMDA receptors: molecular mechanisms and therapeutic potential. Br J Pharmacol. 2009;157:1301–1317. doi: 10.1111/j.1476-5381.2009.00304.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moon IS, Apperson ML, Kennedy MB. The major tyrosine-phosphorylated protein in the postsynaptic density fraction is N-methyl-D-aspartate receptor subunit 2B. Proc Natl Acad Sci USA. 1994;91:3954–3958. doi: 10.1073/pnas.91.9.3954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris RG, Anderson E, Lynch GS, Baudry M. Selective impairment of learning and blockade of long-term potentiation by an N-methyl-D-aspartate receptor antagonist, AP5. Nature. 1986;319:774–776. doi: 10.1038/319774a0. [DOI] [PubMed] [Google Scholar]
- Morrow JA, Maclean JK, Jamieson C. Recent advances in positive allosteric modulators of the AMPA receptor. Curr Opin Drug Discov Dev. 2006;9:571–579. [PubMed] [Google Scholar]
- Mothet JP, Rouaud E, Sinet PM, Potier B, Jouvenceau A, Dutar P, Videau C, Epelbaum J, Billard JM. A critical role for the glial-derived neuro- modulator d-serine in the age-related deficits of cellular mechanisms of learning and memory. Aging Cell. 2006;5:267–274. doi: 10.1111/j.1474-9726.2006.00216.x. [DOI] [PubMed] [Google Scholar]
- Mullasseril P, Hansen KB, Vance KM, Ogden KK, Yuan H, Kurtkaya NL, Santangelo R, Orr AG, Le P, Vellano KM, Liotta DC, Traynelis SF. A subunit-selective potentiator of NR2C- and NR2D-containing NMDA receptors. Nat Commun. 2010;1:1–8. doi: 10.1038/ncomms1085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakazawa T, Komai S, Tezuka T, Hisatsune C, Umemori H, Semba K, Mishina M, Manabe T, Yamamoto T. Characterization of Fyn-mediated tyrosine phosphorylation sites on GluR epsilon 2 (NR2B) subunit of the N-methyl-D-aspartate receptor. J Biol Chem. 2001;276:693–699. doi: 10.1074/jbc.M008085200. [DOI] [PubMed] [Google Scholar]
- Nikiforuk A, Kos T, Rafa D, Behl B, Bespalov A, Popik P. Blockade of glycine transporter 1 by SSR-504734 promotes cognitive flexibility in glycine/NMDA receptor-dependent manner. Neuropharmacology. 2011;61:262–267. doi: 10.1016/j.neuropharm.2011.04.010. [DOI] [PubMed] [Google Scholar]
- Ng D, Pitcher GM, Szilard RK, Sertié A, Kanisek M, Clapcote SJ, Lipina T, Kalia LV, Joo D, McKerlie C, Cortez M, Roder JC, Salter MW, McInnes RR. Neto1 is a novel CUB-domain NMDA receptor interacting protein required for synaptic plasticity and learning. PLoS Biol. 2009;7:e41. doi: 10.1371/journal.pbio.1000041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen TH, Liu J, Lombroso PJ. Striatal enriched phosphatise 61 dephosphorylates Fyn at phosphotyrosine 420. J Biol Chem. 2002;277:24274–24279. doi: 10.1074/jbc.M111683200. [DOI] [PubMed] [Google Scholar]
- Nowak L, Bregetovski P, Ascher P, Herbert A, Prochiantz A. Magnesium gates glutamate-activated channels in mouse central neurons. Nature. 1984;307:462–5. doi: 10.1038/307462a0. [DOI] [PubMed] [Google Scholar]
- Ohnishi H, Murata Y, Okazawa H, Matozaki T. Src family kinases: modulators of neurotransmitter receptor function and behavior. Trends Neurosci. 2011;34:629–637. doi: 10.1016/j.tins.2011.09.005. [DOI] [PubMed] [Google Scholar]
- Oliver MW, Larson J, Lynch G. Activation of the glycine site associated with the NMDA receptor is required for induction of LTP in neonatal hippocampus. Int J Dev Neurosci. 1990;8:417–24. doi: 10.1016/0736-5748(90)90074-c. [DOI] [PubMed] [Google Scholar]
- Olpe HR, Wörner W, Ferrat T. Stimulation parameters determine role of GABAB receptors in long-term potentiation. Experientia. 1993;49:542–6. doi: 10.1007/BF01955159. [DOI] [PubMed] [Google Scholar]
- Omkumar RV, Kiely MJ, Rosenstein AJ, Min KT, Kennedy MB. Identification of a phosphorylation site for calcium/calmodulin-dependent protein kinase II in the NR2B subunit of the N-methyl-D-aspartate receptor. J Biol Chem. 1996;271:31670–31678. doi: 10.1074/jbc.271.49.31670. [DOI] [PubMed] [Google Scholar]
- O'Neill MJ, Dix S. AMPA receptor potentiators as cognitive enhancers. IDrugs. 2007;10:185–192. [PubMed] [Google Scholar]
- Palmer AJ, Lodge D. Cyclothiazide reverses AMPA receptor antagonism of the 2,3-benzodiazepine, GYKI 53655. Eur J Pharmacol. 1993;244:193–204. doi: 10.1016/0922-4106(93)90027-7. ?? [DOI] [PubMed] [Google Scholar]
- Park-Chung M, Wu F, Purdy RH, Malayev AA, Gibbs TT, Farb DH. Distinct sites for inverse modulation of NMDA receptors by sulfated steroids. Mol Pharmacol. 1997;52:1113–1123. doi: 10.1124/mol.52.6.1113. [DOI] [PubMed] [Google Scholar]
- Partin KM, Fleck MW, Mayer ML. AMPA receptor flip/flop mutants affecting deactivation, desensitization, and modulation by cyclothiazide, aniracetam, and thiocyanate. J Neurosci. 1996;16:6634–47. doi: 10.1523/JNEUROSCI.16-21-06634.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patneau DK, Vyklicky L, Jr, Mayer ML. Hippocampal neurons exhibit cyclothiazide-sensitive rapidly desensitizing responses to kainate. J Neurosci. 1993;13:3496–509. doi: 10.1523/JNEUROSCI.13-08-03496.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelkey KA, Askalan R, Paul S, Kalia LV, Nguyen TH, Pitcher GM, Salter MW, Lombroso PJ. Tyrosine phosphatise STEP is a tonic brake on induction of long-term potentiation. Neuron. 2002;34:127–138. doi: 10.1016/s0896-6273(02)00633-5. [DOI] [PubMed] [Google Scholar]
- Petrovic M, Sedlacek M, Cais O, Horak M, Chodounska H, Vyklicky JR. Pregnenolone sulfate modulation of N-methyl-D-aspartate receptors is phosphorylation dependent. Neuroscience. 2009;160:616–628. doi: 10.1016/j.neuroscience.2009.02.052. [DOI] [PubMed] [Google Scholar]
- Pirotte B, Francotte P, Goffin E, Fraikin P, Danober L, Lesur B, Botez I, Caignard DH, Lestage P, de Tullio P. Ring-fused thiadiazines as core structures for the development of potent AMPA receptor potentiators. Curr Med Chem. 2010;17:3575–3582. doi: 10.2174/092986710792927859. [DOI] [PubMed] [Google Scholar]
- Prybylowski K, Chang K, Sans N, Kan L, Vicini S, Wenthold RJ. The synaptic localization of NR2B-containing NMDA receptors is controlled by interactions with PDZ proteins and AP-2. Neuron. 2005;47:845–857. doi: 10.1016/j.neuron.2005.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Randall RD, Lee SY, Meyer JH, Wittenberg GF, Gruol DL. Acute alcohol blocks neurosteroid modulation of synaptic transmission and long-term potentiation in the rat hippocampal slice. Brain Res. 1995;701:238–248. doi: 10.1016/0006-8993(95)01007-9. [DOI] [PubMed] [Google Scholar]
- Reddy DS, Kulkarni SK. The effects of neurosteroids on acquisition and retention of a modified passive-avoidance learning task in mice. Brain Res. 1998;791:108–16. doi: 10.1016/s0006-8993(98)00085-7. [DOI] [PubMed] [Google Scholar]
- Roche KW, Standley S, McCallum J, Dune L, Ehlers MD, Wenthold RJ. Molecular determinants of NMDA receptor internalization. Nat Neurosci. 2001;4:794–802. doi: 10.1038/90498. [DOI] [PubMed] [Google Scholar]
- Sabeti J, Nelson TE, Purdy RH, Gruol DL. Steroid pregnenolone sulfate enhances NMDA-receptor-independent long-term potentiation at hippocampal CA1 synapses: role for L-type calcium channels and sigma-receptors. Hippocampus. 2007;17:349–69. doi: 10.1002/hipo.20273. [DOI] [PubMed] [Google Scholar]
- Salter MW, Dong Y, Kalia LV, Liu XJ, Pitcher G. Regulation of NMDA receptors by kinases and phosphatises. In: Van Dongen AM, editor. Biology of the NMDA receptor. CRC Press; Boca Raton (FL): 2009. Chapter 7. [PubMed] [Google Scholar]
- Sánchez-Pérez AM, Felipo V. Serines 890 and 896 of the NMDA receptor subunit NR1 are differentially phosphorylated by protein kinase C isoforms. Neurochem Int. 2005;47:84–91. doi: 10.1016/j.neuint.2005.04.011. [DOI] [PubMed] [Google Scholar]
- Sanz-Clemente A, Matta JA, Isaac JT, Roche KW. Casein kinase 2 regulates the NR2 subunit composition of synaptic NMDA receptors. Neuron. 2010;67:984–996. doi: 10.1016/j.neuron.2010.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarter M, Parikh V. Choline transporters, cholinergic transmission and cognition. Nat Rev Neurosci. 2005;6:48–56. doi: 10.1038/nrn1588. [DOI] [PubMed] [Google Scholar]
- Scott DB, Blanpied TA, Swanson GT, Zhang C, Ehlers MD. An NMDA receptor ER retention signal regulated by phosphorylation and alternative splicing. J Neurosci. 2001;21:3063–3072. doi: 10.1523/JNEUROSCI.21-09-03063.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott DB, Blanpied TA, Ehlers MD. Coordinated PKA and PKC phosphorylation suppress RXR-mediated ER retention and regulates the surface delivery of NMDA receptors. Neuropharmacology. 2003;45:755–767. doi: 10.1016/s0028-3908(03)00250-8. [DOI] [PubMed] [Google Scholar]
- Seabrook GR, Howson W, Lacey MG. Electrophysiological characterization of potent agonists and antagonists at pre- and postsynaptic GABAB receptors on neurones in rat brain slices. Br J Pharmacol. 1990;101:949–57. doi: 10.1111/j.1476-5381.1990.tb14186.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selye H. Anesthetic effect of steroid hormones. Proc Soc Exp Biol Med. 1941;46:116–221. [Google Scholar]
- Shimazaki T, Kaku A, Chaki S. D-Serine and a glycine transporter-1 inhibitor enhance social memory in rats. Psychopharmacology (Berl) 2010;209:263–270. doi: 10.1007/s00213-010-1794-y. [DOI] [PubMed] [Google Scholar]
- Simpkins KL, Guttmann RP, Dong Y, Chen Z, Sokol S, Neumar RW, Lynch DR. Selective activation induced cleavage of the NR2B subunit by calpain. J Neurosci. 2003;23:11322–11331. doi: 10.1523/JNEUROSCI.23-36-11322.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon RP, Swan JH, Griffiths T, Meldrum BS. Blockade of N-methyl-D-aspartate receptors may protect against ischemic damage in the brain. Science. 1984;226(4676):850–852. doi: 10.1126/science.6093256. [DOI] [PubMed] [Google Scholar]
- Skeberdis VA, Chevaleyre V, Lau CG, Goldberg JH, Pettit DL, Suadicani SO, Lin Y, Bennett MV, Yuste R, Castillo PE, Zukin RS. Protein kinase A regulates calcium permeability of NMDA receptors. Nat Neurosci. 2006;9:501–510. doi: 10.1038/nn1664. [DOI] [PubMed] [Google Scholar]
- Sliwinski A, Monnet FP, Schumacher M, Morin-Surun MP. Pregnenolone sulfate enhances long-term potentiation in CA1 in rat hippocampus slices through the modulation of N-methyl-D-aspartate receptors. J Neurosci Res. 2004;78:691–701. doi: 10.1002/jnr.20332. [DOI] [PubMed] [Google Scholar]
- Slutsky I, Abumaria N, Wu LJ, Huang C, Zhang L, Li B, Zhao X, Govindarajan A, Zhao MG, Zhuo M, Tonegawa S, Liu G. Enhancement of learning and memory by elevating brain magnesium. Neuron. 2010;65:165–177. doi: 10.1016/j.neuron.2009.12.026. [DOI] [PubMed] [Google Scholar]
- Smith SM, Uslaner JM, Hutson PH. The Therapeutic Potential of D-Amino Acid Oxidase (DAAO) Inhibitors. Open Med Chem J. 2010;4:3–9. doi: 10.2174/1874104501004020003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snyder EM, Nong Y, Almeida CG, Paul S, Moran T, Choi EY, Nairn AC, Salter MW, Lombroso PJ, Gouras GK, Greengard P. Regulation of NMDA receptor trafficking by amyloid-beta. Nat Neurosci. 2005;8:1051–1058. doi: 10.1038/nn1503. [DOI] [PubMed] [Google Scholar]
- Stäubli U, Perez Y, Xu FB, Rogers G, Ingvar M, Stone-Elander S, Lynch G. Centrally active modulators of glutamate receptors facilitate the induction of long-term potentiation in vivo. Proc Natl Acad Sci USA. 1994b;91:11158–11162. doi: 10.1073/pnas.91.23.11158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staubli U, Rogers G, Lynch G. Facilitation of glutamate receptors enhances memory. Proc Natl Acad Sci USA. 1994a;91:777–781. doi: 10.1073/pnas.91.2.777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens ER, Gustafson EC, Miller RF. Glycine transport accounts for the differential role of glycine vs. d-serine at NMDA receptor coagonist sites in the salamander retina. Eur J Neurosci. 2010;31:808–816. doi: 10.1111/j.1460-9568.2010.07135.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strick CA, Li C, Scott L, Harvey B, Hajos M, Steyn SJ, Piotrowski MA, James LC, Downs JT, Rago B, Becker SL, El-Kattan A, Xu Y, Ganong AH, Tingley FD, III, Ramirez AD, Seymour PA, Guanowsky V, Majchrzak MJ, Fox CB, Schmidt CJ, Duplantier AJ. Modulation of NMDA receptor function by inhibition of D-amino acid oxidase in rodent brain. Neuropharmacology. 2011;61:1001–1015. doi: 10.1016/j.neuropharm.2011.06.029. [DOI] [PubMed] [Google Scholar]
- Sun Y, Olson R, Horning M, Armstrong N, Mayer M, Gouaux E. Mechanism of glutamate desensitization. Nature. 2002;417:245–253. doi: 10.1038/417245a. [DOI] [PubMed] [Google Scholar]
- Supplisson S, Bergman C. Control of NMDA receptor activation by a glycine transporter co-expressed in Xenopus oocytes. J Neurosci. 1997;17:4580–90. doi: 10.1523/JNEUROSCI.17-12-04580.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka M, Sokabe M. Continuous de novo synthesis of neurosteroids is required for normal synaptic transmission and plasticity in the dentate gyrus of the rat hippocampus. Neuropharmacology. 2012;62:2373–87. doi: 10.1016/j.neuropharm.2012.02.007. [DOI] [PubMed] [Google Scholar]
- Tang YP, Shimizu E, Dube GR, Rampon C, Kerchner GA, Zhuo M, Liu G, Tsien JZ. Genetic enhancement of learning and memory in mice. Nature. 1999;401:63–69. doi: 10.1038/43432. [DOI] [PubMed] [Google Scholar]
- Tingley WG, Ehlers MD, Kameyama K, Doherty C, Ptak JB, Riley CT, Huganir RL. Characterization of protein kinase A and protein kinase C phosphorylation of the N-methyl-D-aspartate receptor NR1 subunit using phosphorylation site-specific antibodies. J Biol Chem. 1997;272:5157–5166. doi: 10.1074/jbc.272.8.5157. [DOI] [PubMed] [Google Scholar]
- Thomson AM, Walker VE, Flynn DM. Glycine enhances NMDA-receptor mediated synaptic potentials in neocortical slices. Nature. 1989;338:422–424. doi: 10.1038/338422a0. [DOI] [PubMed] [Google Scholar]
- Traynelis SF, Hartley M, Heinemann SF. Control of proton sensitivity of the NMDAR by RNA splicing and polyamines. Science. 1995;268:873–876. doi: 10.1126/science.7754371. [DOI] [PubMed] [Google Scholar]
- Traynelis SF, Wollmuth LP, McBain CJ, Menniti FS, Vance KM, Ogden KK, Hansen KB, Yuan H, Myers SJ, Dingledine R. Glutamate receptor ion channels: structure, regulation, and function. Pharmacol Rev. 2010;62:405–496. doi: 10.1124/pr.109.002451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trepanier CH, Jackson MF, MacDonald JF. Regulation of NMDA receptors by the tyrosine kinase Fyn. FEBS J. 2012;279:12–19. doi: 10.1111/j.1742-4658.2011.08391.x. [DOI] [PubMed] [Google Scholar]
- Tsai G, Lane HY, Yang P, Chong MY, Lange N. Glycine transporter I inhibitor, N-methylglycine (sarcosine), added to antipsychotics for treatment of schizophrenia. Biol Psychiatry. 2004;55:452–456. doi: 10.1016/j.biopsych.2003.09.012. [DOI] [PubMed] [Google Scholar]
- Tsai G, Yang P, Chung LC, Lange N, Coyle JT. D-serine added to anti- psychotics for the treatment of schizophrenia. Biol Psychiatry. 1998;44:1081–1089. doi: 10.1016/s0006-3223(98)00279-0. [DOI] [PubMed] [Google Scholar]
- Vallée M, Mayo W, Darnaudéry M, Corpéchot C, Young J, Koehl M, Le Moal M, Baulieu EE, Robel P, Simon H. Neurosteroids: deficient cognitive performance in aged rats depends on low pregnenolone sulfate levels in the hippocampus. Proc Natl Acad Sci USA. 1997;94:14865–14870. doi: 10.1073/pnas.94.26.14865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vales K, Svoboda J, Benkovicova K, Bubenikova-Valesova V, Stuchlik A. The difference in effect of mGlu2/3 and mGlu5 receptor agonists on cognitive impairment induced by MK-801. Eur J Pharmacol. 2010;639:91–8. doi: 10.1016/j.ejphar.2009.11.067. [DOI] [PubMed] [Google Scholar]
- Vandergriff J, Huff K, Bond A, Lodge D. Potentiation of responses to AMPA on central neurones by LY392098 and LY404187 in vivo. Neuropharmacology. 2001;40:1003–9. doi: 10.1016/s0028-3908(01)00031-4. [DOI] [PubMed] [Google Scholar]
- Vinson PN, Conn PJ. Metabotropic glutamate receptors as therapeutic targets for schizophrenia. Neuropharmacology. 2012;62:1461–72. doi: 10.1016/j.neuropharm.2011.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vissel B, Krupp JJ, Heinemann SF, Westbrook GL. A use-dependent tyrosine dephosphorylation of NMDA receptors is independent of ion flux. Nat Neurosci. 2001;4:587–596. doi: 10.1038/88404. [DOI] [PubMed] [Google Scholar]
- Wafford KA, Kathoria M, Bain CJ, Marshall G, Le Bourdellès B, Kemp JA, Whiting PJ. Identification of amino acids in the N-methyl-D-aspartate receptor NR1 subunit that contribute to the glycine binding site. Mol Pharmacol. 1995;47:374–80. [PubMed] [Google Scholar]
- Watanabe Y, Saito H, Abe K. Effects of glycine and structurally related amino acids on generation of long-term potentiation in rat hippocampal slices. Eur J Pharmacol. 1992;223:179–84. doi: 10.1016/0014-2999(92)94837-l. [DOI] [PubMed] [Google Scholar]
- Ward SE, Harries M. Recent advances in the discovery of selective AMPA receptor positive allosteric modulators. Curr Med Chem. 2010;17:3503–13. doi: 10.2174/092986710792927840. [DOI] [PubMed] [Google Scholar]
- Westphal RS, Tavalin SJ, Lin JW, Alto NM, Fraser ID, Langeberg LK, Sheng M, Scott JD. Regulation of NMDA receptors by an associated phosphatise kinase signaling complex. Science. 1999;285:93–96. doi: 10.1126/science.285.5424.93. [DOI] [PubMed] [Google Scholar]
- Williams K. Mechanisms influencing stimulatory effects of spermine at recombinant N-methyl-D-aspartate receptors. Mol Pharmacol. 1994a;46:161–168. [PubMed] [Google Scholar]
- Williams K. Subunit-specific potentiation of recombinant N-methyl-D-aspartate receptors by histamine. Mol Pharmacol. 1994b;46:531–541. [PubMed] [Google Scholar]
- Williams K. Interactions of polyamines with ion channels. Biochem J. 1997;325:289–297. doi: 10.1042/bj3250289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu FS, Gibbs TT, Farb DH. Pregnenolone sulfate: a positive allosteric modulator at the N-methyl-D-aspartate receptor. Mol Pharmacol. 1991;40:333–336. [PubMed] [Google Scholar]
- Yamada KA, Rothman SM. Diazoxide reversibly blocks gluta- mate desensitization and prolongs excitatory postsynaptic currents in rat hippocampal neurons. J Phvsiol (Lond) 1992;458:385–407. doi: 10.1113/jphysiol.1992.sp019424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang M, Leonard JP. Identification of mouse NMDA receptor subunit NR2A C-terminal tyrosine sites phosphorylated by coexpression with v-Src. J Neurochem. 2001;77:580–588. doi: 10.1046/j.1471-4159.2001.00255.x. [DOI] [PubMed] [Google Scholar]
- Yang Y, Ge W, Chen Y, Zhang Z, Shen W, Wu C, Poo M, Duan S. Contribution of astrocytes to hippocampal long-term potentiation through release of D-serine. Proc Natl Acad Sci U S A. 2003;100:15194–15199. doi: 10.1073/pnas.2431073100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang K, Trepanier C, Sidhu B, Xie YF, Li H, Lei G, Salter MW, Orser BA, Nakazawa T, Yamamoto T, Jackson MF, MacDonald JF. Metaplasticity gated through differential regulation of GluN2A versus GluN2B receptors by Src family kinases. EMBO J. 2012;31:805–816. doi: 10.1038/emboj.2011.453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang S, Qiao H, Wen L, Zhou W, Zhang Y. D-serine enhances impaired long-term potentiation in CA1 subfield of hippocampal slices from aged senescence-accelerated mouse prone/8. Neurosci Lett. 2005;379:7–12. doi: 10.1016/j.neulet.2004.12.033. [DOI] [PubMed] [Google Scholar]