

Abstract
Sporopollenin is the main constituent of the exine layer of spore and pollen walls. The anther-specific chalcone synthase-like (ASCL) enzyme of Physcomitrella patens, PpASCL, has previously been implicated in the biosynthesis of sporopollenin, the main constituent of exine and perine, the two outermost layers of the moss spore cell wall. We made targeted knockouts of the corresponding gene, PpASCL, and phenotypically characterized ascl sporophytes and spores at different developmental stages. Ascl plants developed normally until late in sporophytic development, when the spores produced were structurally aberrant and inviable. The development of the ascl spore cell wall appeared to be arrested early in microspore development, resulting in small, collapsed spores with altered surface morphology. The typical stratification of the spore cell wall was absent with only an abnormal perine recognisable above an amorphous layer possibly representing remnants of compromised intine and/or exine. Equivalent resistance of the spore walls of ascl mutants and the control strain to acetolysis suggests the presence of chemically inert, defective sporopollenin in the mutants. Anatomical abnormalities of late-stage ascl sporophytes include a persistent large columella and an air space incompletely filled with spores. Our results indicate that the evolutionarily conserved PpASCL gene is needed for proper construction of the spore wall and for normal maturation and viability of moss spores.
Introduction
The transition of plants from aquatic to terrestrial environments about 500 million years ago is one of the most important events in the evolution of life on earth. On land, plants encountered new challenges, which included surviving exposure to higher fluxes of damaging UV irradiation and desiccation. Plants successfully adapted to these new environments by, among other things, producing novel secondary metabolites: phenylpropanoids for protection from UV rays and extracellular matrices of sporopollenin and cutin to counter desiccation [1, 2]. Discovering additional information about evolution of the biosynthetic pathways of these protective compounds will contribute significantly to our comprehension of the colonization of land by early plants.
Sporopollenin is the main polymeric component of the outer exine layer of spore and pollen walls, and consists of medium- to long-chain fatty acids and oxygenated aromatic compounds [3]. These constituents are coupled via extensive ester and ether linkages, resulting in a robust polymer that enables spore and pollen grains to tolerate physical abrasion, desiccation and UV-B irradiation [4, 5]. Recently, several genes have been shown to be involved in the biosynthesis of sporopollenin (Fig 1) [6]. The MALE STERILITY2 (MS2) gene encodes a fatty acyl reductase that reduces very long chain fatty acyl-CoA or acyl-acyl carrier protein to fatty alcohol [7, 8]. The cytochrome P450 genes, CYP703A2 and CYP704B1, encode fatty acid hydroxylases that catalyze in-chain and ω-hydroxylation respectively of mid- to long-chain fatty acids [9, 10]. Fatty alcohols and hydroxyfatty acids produced by these gene products, along with phenylpropanoids (e.g. p-coumaric acid and ferulic acid) [11], may serve as building blocks and provide oxygen atoms for ester and ether linkages in sporopollenin. In addition, the hydroxylated alkylpyrones generated by sequential actions of acyl-CoA synthetase, anther-specific chalcone synthase-like enzyme (ASCL) and tetraketide α-pyrone reductase are either direct sporopollenin monomers or incorporated into sporopollenin after further modifications [12, 13, 14]. ASCL is a type III polyketide synthase (PKS), and produces in vitro hydroxyalkyl α-pyrones by condensing hydroxyfatty acyl-CoA esters with malonyl-CoA molecules [14]. In Arabidopsis, PKSA and PKSB, encoding two paralogous ASCLs [15], are specifically expressed in anther tapetal cells during microspore development. A double pksa pksb knockout mutant was male sterile and produced defective pollen grains with no apparent exine, providing evidence for the involvement of ASCL in sporopollenin biosynthesis [14].
Fig 1. Reaction sequence for the biosynthesis of hydroxylated alkylpyrones as sporopollenin building blocks.
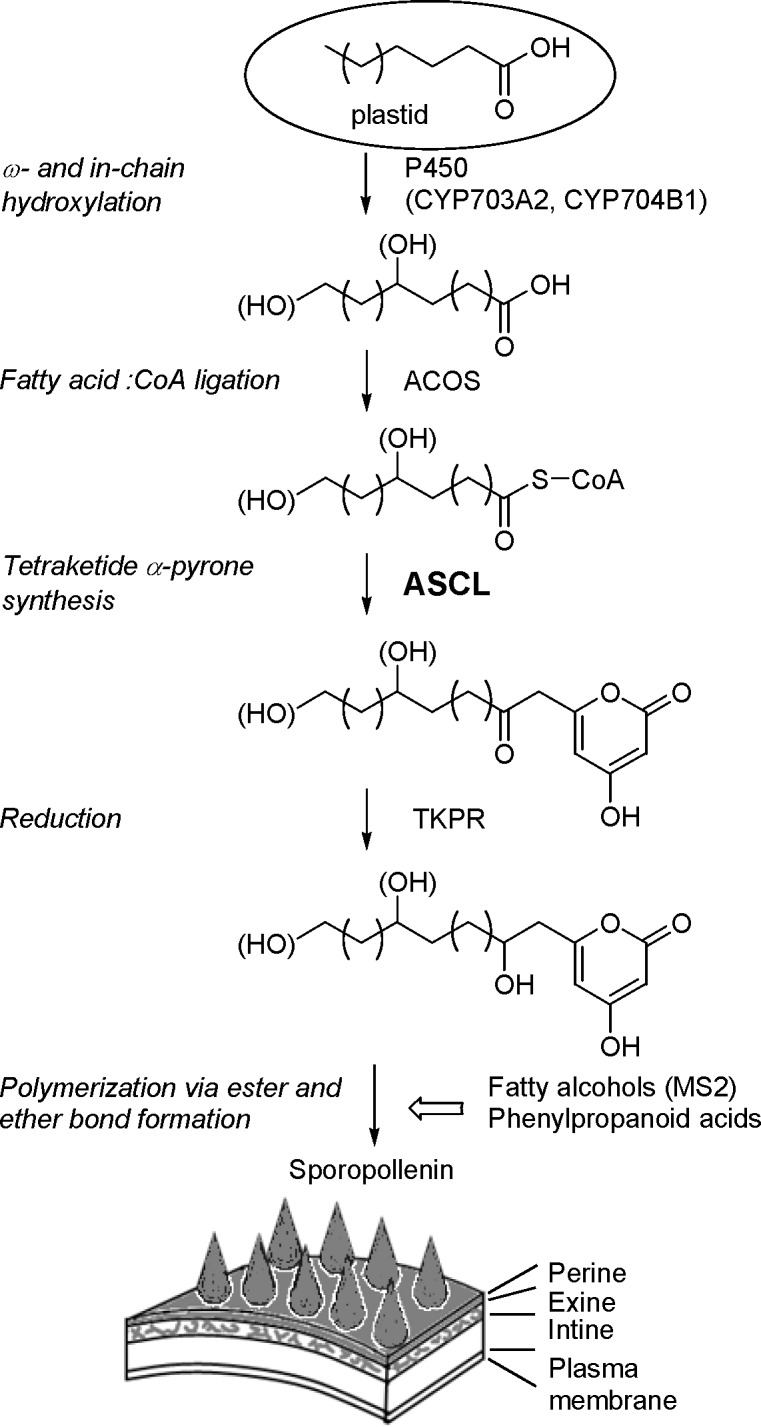
Medium- to long-chain fatty acids are produced in plastids and then translocated out to be used for the consecutive action of enzymes in sporopollenin biosynthesis. This proposed pathway produces sporopollenin building blocks that are polymerized along with fatty alcohols and phenylpropanoid acids on the surface of the spore or pollen wall by the formation of ester and ether linkages. Enzymes are listed to the right of the arrows with their corresponding reactions on the left. ACOS, acyl-CoA synthetase; ASCL, anther-specific chalcone synthase-like enzyme; MS2, Male Sterility 2; TKPR, tetraketide α-pyrone reductase.
Bryophytes (liverworts, mosses, and hornworts) are the simplest and earliest-diverging lineages of land plants. The phylogeny of land plants supports the hypothesis that early land plants were bryophyte-like and thus bryophytes offer unique windows into the early evolution of land plants [16]. The moss Physcomitrella patens has emerged as a model bryophyte [17]. Physcomitrella is unique among plants in that targeted gene knockouts can be efficiently produced by homologous recombination [18]. Loss-of-function phenotypes can be readily screened at all stages of development after mutated protoplasts derived from haploid protonemal cells are regenerated. These characteristics render Physcomitrella an excellent system for studying plant physiology and early land plant evolution with reverse genetics approaches. Consequently, the genome of Physcomitrella has been sequenced, and large-scale gene expression profiling data are available from different tissues of the plant [19, 20].
The presence of sporopollenin in the outer cell wall layers of pollen and spores of land plants indicates that the development of this resistant biopolymer occurred early in land plant evolution. The genome of Physcomitrella contains putative orthologues of the sporopollenin biosynthesis genes, suggesting that the biosynthetic pathway of sporopollenin may be conserved in land plant lineages [21, 22]. PpASCL is the putative Physcomitrella orthologue of Arabidopsis PKSA and PKSB. EST abundance data and gene expression analysis by microarrays suggested that PpASCL is specifically expressed in the green sporophyte stage of development. Similar to PKSA and PKSB, recombinant PpASCL preferred hydroxylated fatty acyl-CoA esters as the starter substrate and produced hydroxyalkyl α-pyrones in vitro [21]. These results suggested that PpASCL may also play a role in the production of sporopollenin in Physcomitrella. An ascl loss-of-function mutant is then expected to have altered spore wall formation, similar to the pollen phenotype of pksa and pksb single and double mutants of Arabidopsis [14]. In this study, targeted PpASCL knockout transformants were made to elucidate the in planta function of PpASCL. Phenotypic characterization of the Physcomitrella ascl mutants supports a role for PpASCL in spore wall development, in agreement with its proposed participation in the evolutionarily conserved sporopollenin biosynthetic pathway.
Materials and Methods
Plant material, media and culture conditions
The Physcomitrella lines used in this study were characterized by an isogenic genetic background. Transgenic strains were derived from the pabB4 [23] control strain of P. patens (Hedw.) Bruch, Schimp & W. Gümbel. When grown on medium containing adequate (1.8 and 18 μM for gametophytes and sporophytes respectively) p-aminobenzoic acid (paba), pabB4 is phenotypically indistinguishable from the original Gransden wild type strain from which it was derived. It produces abundant sporophytes containing viable spores with normal spore coat ornamentation and completes its life-cycle in 2–3 months [24].
Plants were grown axenically on solid ABC medium [25] supplemented with paba (1.8 μM) at 22–25°C. Tissues for DNA and protoplast isolation were grown on medium including diammonium (+)-tartrate (5 mM) and overlaid with cellophane discs [26]. Filter-sterilized G418 (50 μg ml-1) was added as necessary to autoclaved medium. Continuous light culture conditions, described previously [27], were utilised with a single modification: Petri plates were covered with one layer of resin filter [Roscolux No. 114 (Hamburg frost), MacPhon Industries, Calgary, AB, Canada].
Preparation of linear PpASCL knockout construct
The knockout construct was derived using pTN182 [28]. This plasmid has a resistance cassette with a neomycin phosphotransferase II gene (nptII), which confers resistance to a range of aminoglycoside antibiotics including G418. 5′ and 3′ regions of PpASCL were PCR-amplified from Physcomitrella genomic DNA (gDNA) using the 5ʹF- and 5ʹR-ASCL-ClaI primer pair and 3ʹF- and 3ʹR-ASCL-NdeI primer pair respectively. The 995 bp 5' locus-specific DNA fragment comprised 549 bp of 5' untranslated region and 446 bp of protein coding sequence while the 757 bp 3′ locus-specific DNA fragment entirely comprised of coding DNA (Fig 2A). The primers used in this study are listed in S1 Table. The two PCR products were appropriately restricted and consecutively ligated into ClaI- and NdeI-restricted pTN182 plasmid to produce the knockout vector, pTN182-PpASCL-KO. Restriction and PCR analyses, as well as sequencing, were used to confirm the desired insertion and orientation of the two locus-specific regions of PpASCL.
Fig 2. Strategy for targeted knockout of PpASCL and genotyping of the resulting stable transformants by PCR.

(a) Schematic diagram of insertion of the linear knockout construct into the PpASCL locus via double homologous recombination. 35S-P, CaMV 35S promoter; nptII, neomycin phosphotransferase II gene; 35S-T, CaMV 35S transcription termination signal. (b) Schematic diagram of recombined gene locus after successful insertion. Expected PCR product sizes based on sequence information are shown. Single-headed arrows denote the locations of primers specific to PpASCL (Primers 1 and 2, which bind to genomic DNA sequences located outside the locus-specific regions used for homologous recombination) or to the nptII resistance cassette (Primers 3 and 4) used in the PCR analyses. Primer 1, ASCL-gDNA-F; 2, ASCL-gDNA-R; 3, pTN182-5ʹ-R; 4, pTN182-3ʹ-F. Primer sequences are provided in S1 Table. (c) PCR products using locus-specific primers 1 and 2 with DNA from untransformed control and each of three stable putative PpASCL knockout lines: ascl-1, -2 and -3. (d) PCR products, indicative of 5′ and 3′ recombination between the knockout vector and homologous DNA in the PpASCL locus, using primers 1 plus 3 (5ʹ recombination) and primers 2 plus 4 (3ʹ recombination). Amplified DNA products were resolved electrophoretically on 1.2% agarose gels and visualized by ethidium bromide fluorescence.
Linear knockout DNA was PCR-amplified from pTN182-PpASCL-KO using 5′F-ASCL-ClaI and 3′R-ASCL-NdeI. The amplified construct was gel purified, ethanol precipitated, resuspended in sterile water and then used directly for protoplast transformation.
Transformation and selection of transgenic plants
Direct transfer of linear knockout constructs (approx. 20 μg in each case) into pabB4 protoplasts and selection of stable transformants were performed as previously described [29] except for the nature of transforming DNA used. Stable transformants were defined as those that retained their transformed phenotype (resistance to G418) after two successive rounds of culturing for two weeks each on non-selective medium before re-culturing in the presence of the selective agent [24].
Molecular analysis of transgenic plants: PCR genotyping, Southern blot and RT-PCR
gDNA was extracted from 7–14 day old protonemal tissue using a DNeasy Plant kit (Qiagen Inc., Toronto, ON, Canada). Insertion of knockout DNA into PpASCL in stable transformants was shown by PCR analysis with the primer pair, ASCL-gDNA-F and ASCL-gDNA-R, which bind to gDNA sequences located outside the locus-specific regions used for homologous recombination (Primers 1 and 2 in Fig 2A and 2B). Further verification of successful PpASCL allele replacement was obtained by using PpASCL- plus pTN182-specific PCR primer pairs to demonstrate the occurrence of both 5ʹ- and 3ʹ-targeted insertion (TI): ASCL-gDNA-F plus pTN182-5ʹ-R for 5ʹ TI and pTN182-3ʹ-F plus ASCL-gDNA-R for 3ʹ TI (Fig 2B).
For Southern blot analysis, AclI-restricted gDNA (10 μg) was electrophoretically resolved using a 1.2% agarose gel, transferred to Amersham Hybond-N+ membrane (GE Healthcare, Mississauga, ON, Canada) and UV cross-linked to the membrane (UV Stratalinker, Stratagene, La Jolla, CA, USA). A 743 bp long nptII-specific probe was made by PCR using the primer pair, pTN-G418-F and pTN-G418-R (S1 Table), with pTN182. Probe digoxigenin (DIG)-labelling, Southern hybridization and immunological detection were performed using the DIG High Prime DNA Labeling and Detection Starter Kit II (Roche, Mannheim, Germany) according to the manufacturer’s instructions.
For reverse transcription (RT)-PCR, protonemal (100 mg), gametophoric (100 mg) and sporophytic tissues were ground to fine powder in liquid nitrogen with sterile mortar and pestle. For sporophytes, up to 1000 sporophytes (10–100 mg) from different developmental stages were harvested. Total RNA was extracted using an RNeasy Plant Mini kit (Qiagen), and each RNA sample (0.2–4 μg) was reverse transcribed using an Omniscript RT kit (Qiagen). Gene-specific and intron-spanning primers (S1 Table) were used to amplify corresponding cDNA molecules using a touchdown PCR program. The PCR program started at 95°C for 4 min, followed by 5 cycles of 94°C for 15 s, 65°C for 30 s and 70°C for 1.5 min. The annealing temperature was 60°C for the next 5 cycles, 55°C for a further 5 cycles and 50°C for the final 23 cycles. The use of intron-spanning primers provided assurance that amplicons had been derived from processed RNA transcripts and not from gDNA or unprocessed transcripts. Physcomitrella actin3 (AY382283) was used as the internal reference.
Phenotypic analysis of sporophytes
Gametangia were induced on two to four month old gametophytes grown in glass culture tubes by lowering the temperature to 16°C [30]. Fertilization and sporophyte formation were achieved by irrigating the gametophytes with 5 ml of sterile water containing 18 μM paba followed by further incubation at 16°C [31]. Developing sporophytes were visible within one week after irrigation. For observation of sporophytes, gametophores with sporophytes were removed from culture tubes and placed in sterile water in a Petri dish. Sporophytes were detached from the gametophores, counted and their developmental stage recorded. To release spores, sporophytes were gently ruptured in a drop of sterile water using fine forceps.
Acetolysis
Acetolysis of spores was performed similarly to Dobritsa et al. [10]. Spore suspensions were centrifuged and decanted into a minimal volume of water. A large excess volume of freshly prepared 9:1 (v/v) acetic anhydride and sulphuric acid solution was added to the spores, which were then incubated at 70°C for 20 min. Treated spores were washed twice with sterile water and pipetted on to a slide. The slide was overlaid with a coverslip and the edges sealed with nail polish.
Staining and light microscopy
Staining with Simplified Alexander’s stain, a stain used to distinguish between aborted and non-aborted spores [32], was performed according to Peterson et al. [33].
Sporophytes for cryosectioning were fixed with 4% paraformaldehyde overnight at 4°C. Fixed sporophytes were suspended in a tissue freezing medium (VWR International, Edmonton, AB, Canada), frozen on dry ice, and then mounted to microtome chuck. pabB4 and ascl-2 sporophytes were sectioned to a thickness of 20 μm and 30 μm respectively and placed on to positively charged slides. Sections were then stained with 1% toluidine blue O (TBO) and excess stain was removed with H2O. Images of slide mounted samples were taken with a Nikon Eclipse 80i compound light microscope equipped with a DS-Ri1 digital camera. All other light microscopy images were taken using a DS-Fi1 digital camera, mounted to a Nikon SMZ1500 stereoscopic microscope.
Scanning electron microscopy
Untreated spores were fixed with formaldehyde and air dried on a coverslip overnight. Acetolysed spores were first washed with increasing concentrations of ethanol and then air dried on a coverslip overnight. The individual coverslips were mounted to SEM stubs with double-sided carbon tape and the samples were grounded with copper tape. They were then sputter-coated with gold for 2 min, with argon under vacuum, at a current of 7–8 mA (SC7620 Sputter Coater, Quorum Technologies, Guelph, ON, Canada). Gold coated samples were examined with the secondary electron detector fitted to a Jeol JSM-6360 scanning electron microscope. Samples were subjected to high vacuum during both sample preparation (gold-coating) and observation with the SEM. To determine the best conditions for operation, several spores were imaged at a range of beam currents (varied by increasing or decreasing the spot size), utilizing a smaller beam aperture and variable kV. Optimum conditions that provided the clearest and sharpest images at high magnification but that did not lead to sample charging or sample degradation were found to be 5 kV and a spot size of 25.
Transmission electron microscopy
Spores were fixed in 2.5% glutaraldehyde in PBS solution for 30 min, washed and resuspended in 0.1 M sodium phosphate buffer, pH 7.4. Spores were pelleted by centrifugation and washed twice with the buffer. Spores were post-fixed with osmium tetroxide, washed three times with H2O and then pelleted. An equal volume of molten low melting point agarose was added, the spores resuspended and centrifuged again to pellet them in the tip of a microcentrifuge tube. Once the agarose had solidified, the embedded spores were fixed with the formaldehyde-glutaraldehyde mixture [34]. The tip was subsequently cut, and the spores were fixed a second time using a Pelco 3441 Laboratory Microwave System (Ted Pella, Redding, CA, USA). The fixation was carried out for 2 min at 100 W, stopped for 2 min, and repeated for another 2 min at 100 W. Fixed spores were decanted, then washed three times in H2O and cut into smaller pieces before dehydrating in a graded ethanol series (30, 40, 50, 70, 80, 90, 95, three times with 100%). A transition step of 1:1 ethanol:acetone was followed by a brief 100% acetone step prior to infiltration with Spurr’s epoxy resin. A dropwise method up to 25% resin in acetone was used, followed by microwave infiltration (18 in Hg vacuum, 3 min PL3) at 50% and 75% resin in acetone. One overnight step in 100% resin was followed by microwave infiltration (twice with 100% resin). Samples were embedded in flat molds and polymerized for 18 h at 65°C. Blocks were thin sectioned at 70 nm (Leica UC7) using a diamond knife (Diatome), mounted on uncoated 200 mesh copper grids and stained with 2% uranyl acetate and Reynold’s lead citrate for 12 and 5 min respectively. Imaging was performed at 80 kV with a Hitachi H7600 TEM.
Results
Molecular analysis of stable transformants
Stable transformants were tested by PCR for the occurrence of PpASCL allele replacement (Fig 2). This resulted in amplification of a larger DNA fragment (4319 bp) from the recombinant locus in the three stable transformants, ascl1–3, than from PpASCL in the untransformed control strain (2482 bp) (Fig 2C). Additional PCRs demonstrated both 5ʹ- and 3ʹ-TI in each of the three ascl transgenic strains. Thus, PCRs with ASCL-gDNA-F plus pTN182-5ʹ-R (for testing 5ʹ TI) and pTN182-3ʹ-F plus ASCL-gDNA-R (for testing 3ʹ TI) yielded amplification products of sizes consistent with those predicted, 1361 and 1188 bp respectively, when gDNA from ascl mutants was used (Fig 2B and 2D). Conversely, no DNA fragments were amplified from control gDNA. In summary, these data confirmed 249 bp of PpASCL coding sequence had been replaced in ascl mutants with the aminoglycoside resistance cassette from pTN182. Consequently, these transgenic strains should lack ASCL enzyme activity.
One ascl mutant, ascl-2, was also subjected to Southern hybridization to ascertain whether or not it possessed any ectopic copies of the transgenic DNA as a result of illegitimate recombination. Hybridization of AclI-restricted ascl-2 gDNA with nptII-specific DIG-labelled probe produced a discrete Southern signal corresponding to a single, approx. 6 kb DNA fragment, which accorded closely with the predicted size, 5991 bp, of the expected AclI restriction fragment containing a recombinant, i.e. mutant, PpASCL allele. No signal was detected with the control gDNA (S1 Fig).
RT-PCR was used to demonstrate that PpASCL is expressed in a tissue- and developmental stage-specific manner. It was highly expressed in control sporophytes at the “expanding capsule” (E) and “green” (G) developmental stages but no expression was detected in “orange” sporophytes (O), protonemata or gametophores (Fig 3A). Failure to produce the 1115 bp amplicon when RT-PCR was performed using ascl-2 E + G sporophytes provided further confirmation that PpASCL had been successfully knocked out in this mutant. To test whether the PpASCL promoter is under the same spatio-temporal regulation in the control and ascl-2, we performed additional RT-PCRs with pabB4 E sporophytes (left panel, Fig 3B) and ascl-2 E + G sporophytes (right panel, Fig 3B), targeting separately the 3' and 5' ends of their respective normal and mutant PpASCL transcripts. The primer pair designed to amplify a 3′ region common to both transcripts yielded the predicted 536 bp amplicon in each case. Similarly, the primer pair targeting a 5′ region common to both transcripts produced the expected 332 bp amplicon. However, in the latter case, a larger amplicon was also generated in ascl-2 (lane 5′, right, Fig 3B). The size of this extra amplicon is consistent with its including the 121 nt long intron present towards the 5ʹ end of unprocessed pre-mRNA (intron 1 of PpASCL). Since the 3ʹ end of this intron is only 63 nucleotides upstream from the portion of PpASCL replaced by a selection cassette in ascl-2, a plausible explanation is that a splicing enhancer was located in the section of PpASCL replaced during gene knockout and its loss has resulted in less efficient removal of intron 1. It is unlikely to have resulted from contaminating gDNA since, if that had been the case, a larger amplicon incorporating intron 2 (107 nts long) would have been produced with the other primer pair. Also, no larger amplicons were generated from control sporophytes subjected to the same experimental procedures. Thus, transcriptional regulation appears to be identical for the normal and mutant PpASCL alleles.
Fig 3. RT-PCR analysis of (a) PpASCL expression in Physcomitrella at successive developmental stages and (b) PpASCL promoter activity in ascl-2.
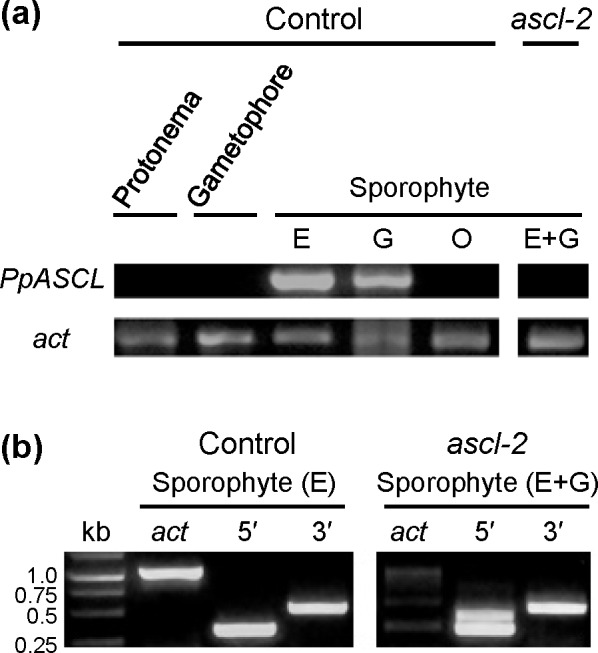
(a) The primers, ASCL-RT-F and ASCL-RT-R (S1 Table), were used to amplify 1115 nucleotides, including 1108 nts from the protein coding region of the PpASCL transcript. In the untransformed pabB4 control, PpASCL expression was detected in sporophytes at the expanding (E) and green (G) capsule stages, but not in the mature orange (O) stage nor in protonemata and gametophores. RT-PCR using ascl-2 sporophytes, comprising a minority at the expanding and a majority at the green capsule stages respectively (E+G), failed to yield the 1115 bp amplicon thus providing additional evidence that PpASCL has been knocked out in this mutant. (b) PpASCL promoter activity was examined in pabB4 E sporophytes and in ascl-2 E+G sporophytes. The primer pair, ASCL-RT-F and 5'R-ASCL-ClaI (S1 Table), was used to amplify a 5′ region predicted to be 332 nucleotides long in mature PpASCL transcripts. Similarly, the primer pair, 3'F-ASCL-NdeI and ASCL-RT-F, was used to amplify a 3′ region predicted to be 536 nucleotides long in mature PpASCL transcripts. Both expected amplicons were generated with sporophytes of both strains. An additional 5ʹ amplicon (upper band) was produced with ascl-2 that, based on its size, we attribute to inefficient splicing of intron 1 from the mutant pre-mRNA. Expression of the Physcomitrella actin3 gene (act) was used as a reference.
Collectively, these observations indicated that ascl-2 was a knocked out transgenic PpASCL mutant, which had resulted from single-copy allele replacement unaccompanied by insertion of ectopic copies of the replacement vector.
Ascl-2 became the strain of choice for further experiments.
The developmental phenotype of ascl-2
Untransformed pabB4 control and ascl-2 gametophytes appeared to be morphologically and developmentally identical (S2 Fig). The formation of gametangia and fertilization were unaffected in ascl-2 as were the early stages of sporophytic morphogenesis. However, during late stages of sporophytic maturation, there were marked differences between these strains.
Morphogenesis of ascl-2 sporophytes
In this study, the sporophytic developmental stages are defined according to the sporophytic maturation and general colouration of the spores [35] as the initial development, expanding capsule, green, yellow and orange stages. Detailed descriptions of pabB4 sporophytes at different developmental stages are provided in S2 Table.
Growth and morphogenesis of ascl-2 and pabB4 control sporophytes were indistinguishable up to and including the mid green stage (Fig 4A–4F, 4a–4f). The earliest differences between control and ascl-2 sporophytes appeared at the late green stage of development (Fig 4G and 4H,4g,4h). In ascl-2, the developing spore mass exhibited a less well defined outline and did not fill the space within the capsule wall to the same extent as in the control (Fig 4I, 4K and 4M,4i,4k,4m, white arrowheads). This appeared to be due to a difference in size of the spores of the respective strains. Whereas control spores increased in size as they matured, ascl-2 spores did not. The general colouration of the spores did not appear to be different in the two strains (Fig 4K–4N, 4k–4n). While the overall developmental timing of ascl-2 sporophytic stages coincided with that of the control, a unique brown stage was observed in ascl-2, which formed from a subset of orange sporophytes after day 48 (Fig 4O and 4P).
Fig 4. Developmental timelines for pabB4, the untransformed control strain, and ascl-2 sporophytes and spores.

Photomicrographs of the typical morphologies of sporophytes and spores of control (A–N) and ascl-2 (a–p) at successive developmental stages. The number of days after irrigation of the cultures is shown with the names assigned to each sporophytic stage. Some stages have been subdivided to allow more detailed description of changes in spore development. White arrowheads denote the outlines of the spore masses within capsules. No spores are seen during the initial growth and expanding capsule stages. The control did not reach the brown sporophytic stage during the observation period. Sporophyte scale bars = 500 μm; Spore scale bars = 10 μm.
Free spores were almost never released from ascl-2 sporophytes. Disintegration of the outer walls of aged, orange and brown, ascl-2 capsules typically did not result in spore release. Some of these capsules contained an amorphous material that caused the spores to aggregate and prevented their dispersal. A similar sticky extracellular material has been described in Arabidopsis ASCL and TETRAKETIDE α-PYRONE REDUCTASE mutant anthers [13, 14, 36, 37].
Further differences were revealed when cross sections of control and ascl-2 sporophytes were compared (Fig 5). Firstly, while mature spores filled the locules comprising the air-space of yellow and orange control sporophytes (Fig 5A, 5B and 5D), the smaller spores of ascl-2 aggregated, leaving much of the air-space empty (Fig 5F, 5G and 5I). Secondly, the central, column-shaped columella of control capsules atrophied by the yellow stage and became noticeably smaller than in ascl-2 capsules in which it filled much of the inner capsular space from apex to base (Fig 5B and 5G). Thirdly, a tapetal layer, which stained darkly and formed a lining around the air-space, was more prominent in ascl-2 capsules (Fig 5I and 5J). Lastly, in ascl-2 capsules, sporopollenin orbicules (stained blue) were present around spores throughout the locules of the air-space and next to the tapetal layers, while the distribution of those in control capsules appeared to be more localized and mainly next to the tapetum (Fig 5E and 5J).
Fig 5. Photomicrographs of cryosectioned pabB4 control and ascl-2 sporophytes.

Cross sections of control (20 μm, a–e) and ascl-2 (30 μm, f–j) sporophytes were taken at the yellow (a,f) and orange (b–e, g–j) stages. Images were saved before (a,b,f,g) and after toluidine blue O staining (c–e, h–j). Orbicules present in locules of the air-space and on the tapetum wall surface are indicated with red arrows in (e) and (j). Sections (e) and (j) are magnified images of red-boxed areas of (d) and (i), respectively. Co, columella; E, epidermis; Lo, locule; T, tapetum. Scale bars = 100 μm.
Spore morphology
Control spores are spherical with an echinate surface composed of perine (perispore) elements, each with a tapering end and taller than 1 μm [38]. The abnormal phenotype of ascl-2 spores was apparent in the free microspores produced by dissociation of tetrads of meiospores and present at the late green sporophytic developmental stage (Fig 4H). For meaningful comparison of spores from different strains, spores were isolated in all cases from the orange sporophytic stage. All three stable ascl mutants made spores, which were smaller than those of the untransformed control and had a distinctive, abnormal morphology (Fig 6 and S3 Fig). The cell wall of mutant spores appeared to have collapsed resulting in many of the spores exhibiting a biconvex outline. The diameter of control spores, excluding ornamentation, was 28 ± 2.0 μm (n = 132, mean ± S.D.), whereas the longest dimension of ascl-2 spores was 18 ± 1.9 μm (n = 146). The mutant spores had a highly irregular surface with globular, granulose protrusions, and lacked the characteristic perine spines found on the spore wall of the control. The morphological differences were clearest in SEM images, which revealed that the typical perine spines of the control were replaced with granulose protrusions (Fig 6G and S3J–S3L Fig). Interestingly, SEM revealed an end-to-end fissure in ascl spore walls, which was also observed occasionally with light microscopy (S3B–S3D Fig). Fissuring does not appear to be an artefact of the SEM imaging process since it was not observed in the SEM images of control spores obtained under the identical conditions (Fig 6B and S3E Fig).
Fig 6. Morphological comparison of orange stage pabB4 control and ascl-2 spores.
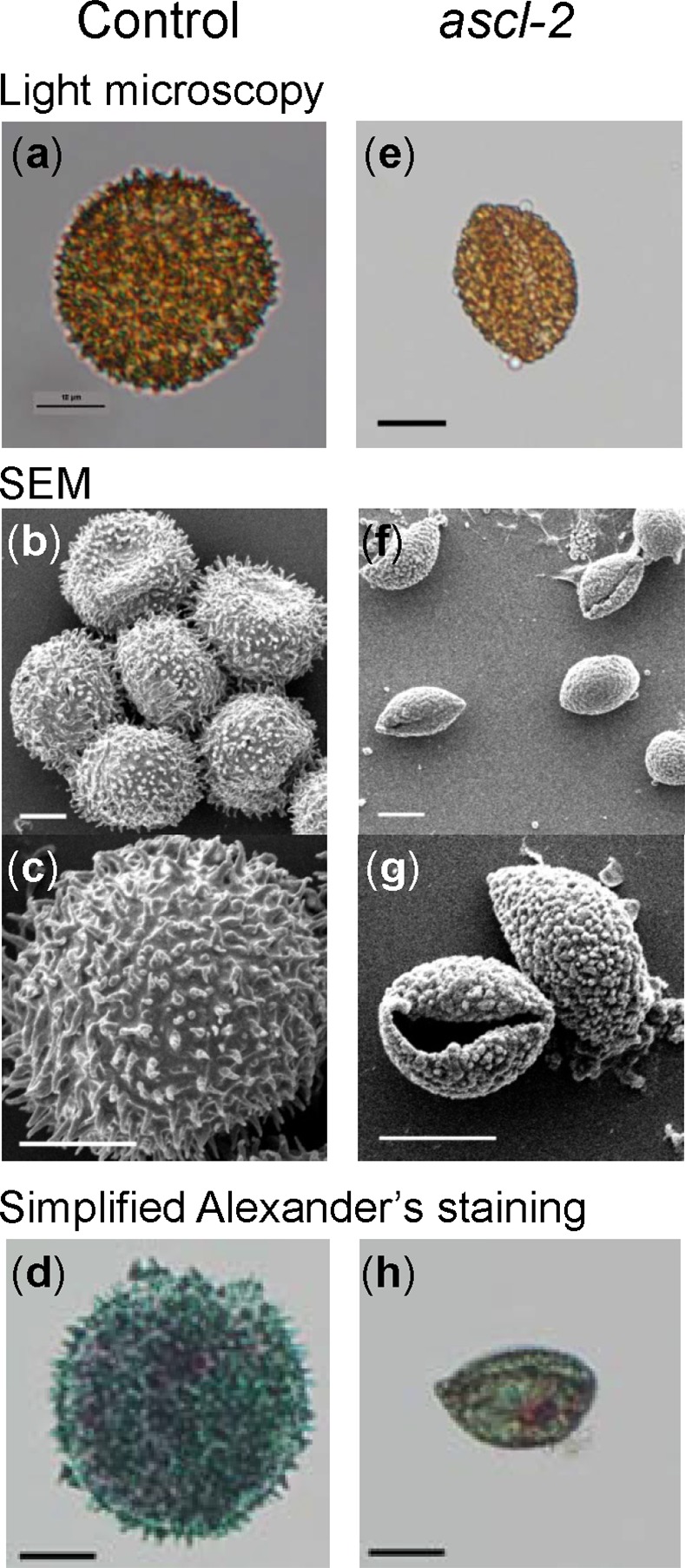
Images of typical spores isolated from mature orange sporophytes were acquired using light microscopy (a,e) and SEM (b,c,f,g). Spores from the control (a–c) and ascl-2 (e–g) are shown. Light microscopy images of control (d) and ascl-2 (h) spores after treatment with simplified Alexander’s stain are also shown. Scale bars = 10 μm.
TEM of control spores clearly depicted the typical stratification of the spore wall comprising the innermost, fibrillar intine, a more electron dense exine (exospore) and the outermost perine layer with spines (Fig 7A) [39]. In ascl-2 spores, the only seemingly identifiable layer appeared to be the perine surrounding the spore and comprising the abnormal ornamentation on its surface (Fig 7B). An irregular, amorphous layer was present below much of the outer perine that may represent remnants of the intine (or exine) acting as the foundation for perine deposition (indicated by an arrow in Fig 7B). As mentioned above, the integrity of the spore wall in ascl-2 was compromised by an opening or fissure in the wall and the ascl-2 spores were only partially filled with cytoplasm, presumably because of leakage of cytoplasm from the spores via the fissure in the mutant spore wall. There also appeared to be uneven deposition of perine or perine-like material at the surface of the remaining cytoplasm within the ascl-2 spores (Fig 7B).
Fig 7. Transmission electron micrographs of pabB4 control and ascl-2 spores.
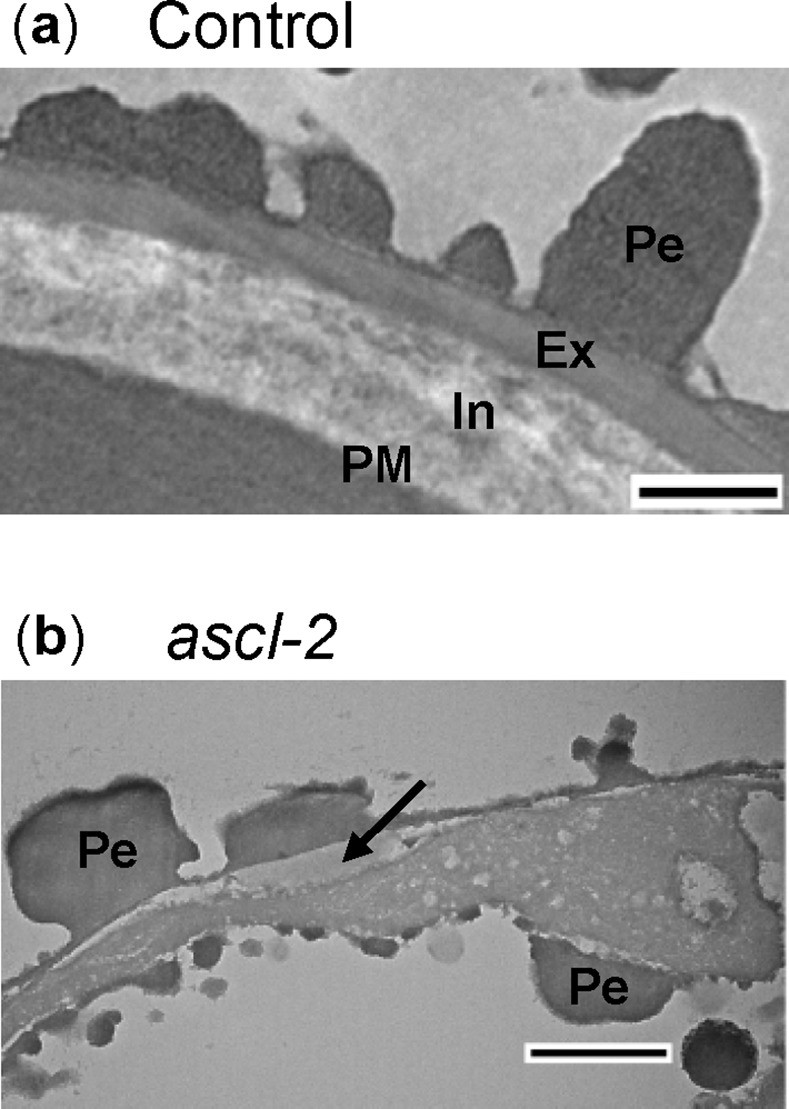
Cross sections of spores from mature orange control (a) and ascl-2 (b) sporophytes were examined with TEM. An amorphous layer found below much of the perine is indicated by an arrow in (b). Ex, exine; In, intine; Pe, perine; PM, plasma membrane. Scale bars = 500 nm.
Acetolysis showed that both control and ascl-2 spore walls are resistant to this chemical treatment. After acetolysis, a slightly yellow-coloured remnant of the spore wall remained intact, comprising the spines of the control and the irregular surface ornamentation of ascl-2 (Fig 8A and 8F). SEM images of acetolysed spores from the early and late orange sporophytic stages revealed the intact echinate and granulose surface patterns of control and ascl-2 spores, respectively (Fig 8E and 8J). Untreated control spores, observed by SEM with and without fixation, were comparable, with none of the spores collapsing (Fig 6B and 6C). By contrast, acetolysed, unfixed control spores collapsed in the vacuum of the SEM (Fig 8B–8E), while those of ascl-2, which had a compromised and fractured cell wall, were unaffected (Fig 8G–8J). In some cases, the collapsed control spores exhibited a triradiate ridge indicative of their tetrad origin (Fig 8B and 8C).
Fig 8. Light microscopy and SEM images of acetolysed spores.

Photomicrographs of spores of pabB4 control (a) and ascl-2 (f) and SEM images of gold coated spores from control (b–e) and ascl-2 (g–j) after acetolysis. Spores were isolated from the orange sporophytic developmental stage. Triradiate ridges are visible in some collapsed control spores (b,c). Scale bars (a,b,d,f,g,i) = 10 μm; (c,e,h,j) = 5 μm.
Spore viability
PabB4 control spores from the yellow and orange sporophytic stages, plated on solid ABC medium supplemented with paba and incubated in continuous white light, began germinating after 4 days. By the fourth day, a large percentage of orange spores had germinated and most of them had germinated after 7 days. Under identical conditions, no ascl-2 spores from any stage of sporophytic development had germinated. Simplified Alexander’s stain revealed that cytoplasm uniformly filled most of the interior of control spores whereas the majority of stained ascl-2 spores exhibited a comparatively irregular distribution of cytoplasm (Fig 6D and 6H). Thus, compared to control spores, ascl-2 spores were depleted in cytoplasm and were nonviable.
Discussion
Evolutionarily conserved genes involved in exine formation
The Physcomitrella genome contains orthologues of Arabidopsis genes involved in exine formation, more specifically in the biosynthesis of sporopollenin. These moss orthologues were first identified by phylogenetic and EST abundance analyses [9, 21, 22], and their expression in Physcomitrella spores was later supported by a genome-wide microarray expression study [20]. Also, they were shown to be up-regulated in early stage sporophytes compared to gametophytes [40]. However, direct evidence of the involvement of the moss orthologues in spore wall formation was initially lacking. Ara et al. reported that PpCYP703B2 partially rescued a defect in the exine layer of the pollen wall of the rice mutant, cyp703a3 [41]. When the Physcomitrella orthologue encoding GAMYB, a transciption factor that regulates CYP703 expression, was knocked out, the resulting Ppgamyb2 spores had severely defective perine and outer exine layers [41]. These results are in agreement with the proposed role of CYP703B2 in sporopollenin biosynthesis [9, 21]. More importantly, Wallace et al. [8] created knockout lines of PpMS2-1, a Physcomitrella orthologue of Arabidopsis MS2, and showed that PpMS2-1 is essential for normal spore wall development, providing the first in planta evidence for conserved functions of Arabidopsis and Physcomitrella orthologues implicated in sporopollenin biosynthesis. Our study shows that PpASCL, the putative Physcomitrella orthologue of Arabidopsis PKSA and PKSB [21], is required for proper exine and intine formation and also for the construction of a normal perine layer. The abnormal perine projections of ascl-2 and Ppms2-1 spores are similar in appearance and, in both cases, the distinction between intine and exine layers has been lost or blurred and the combined width of these two layers has been reduced when compared to the trilaminate spore wall of control strains. Interestingly, spores of both mutants are smaller than control spores. These combined observations strengthen the contention that sporopollenin biosynthesis is an ancient adaptation to conditions in a terrestrial environment that has been conserved throughout subsequent land plant evolution.
The Physcomitrella ascl mutant phenotype is both similar to and distinct from that of the Arabidopsis pksa pksb double mutant
In Physcomitrella and other mosses belonging to the Bryopsida class, exine formation and sporopollenin deposition are thought to occur during the tetrad stage of spore development [22]. Consistent with this, PpASCL expression was detected exclusively in the early stages of spore development. Similarly, expression of Arabidopsis PKSA and PKSB is tightly regulated both temporally and spatially, occurring primarily during the tetrad and/or free microspore stages of pollen development [14].
ASCL is a hydroxyalkyl α-pyrone synthase in vitro and hydroxyalkyl α-pyrones are presumed to be in planta sporopollenin precursors. Based on spectroscopic data, Fraser et al. [42] proposed that the structure of sporopollenin is the same in all land plants. Sporopollenin is the major constituent of the exine layer of both spore and pollen walls. Both ascl-2 spores and pksa pksb pollen lack a recognisable exine layer and are inviable. However, the abnormal phenotype of pksa pksb double mutants in Arabidopsis is more severe than that of ascl single mutants in Physcomitrella. The majority of pksa pksb double mutants fail to produce pollen grains apparently because their microspores atrophy [14]. This suggests that Arabidopsis is unable to produce robust, albeit defective, sporopollenin without ASCL products. Conversely, degeneration of defective ascl-2 spores did not occur and aborted spores remained intact even after several months. While bryophytes have a single-copy ASCL, angiosperm species possess at least two PKS paralogues, which form separate clades in phylogenetic trees, suggesting functional diversification [15]. Collectively, these results indicate that ASCL products are components of sporopollenin in bryophytes and angiosperms but their contributions to sporopollenin integrity in these two plant groups may be different. Arabidopsis PKSA and PKSB and their products (presumed to be tri- and tetraketide α-pyrones) appear to have a more integral role in exine formation than does Physcomitrella PpASCL. This may result partly from differences between spores and pollen with respect to the process of exine formation, especially the mode of sporopollenin deposition, during wall formation [8, 22].
It is not known which sporophytic cells express PpASCL. Most characterized angiosperm sporopollenin monomer biosynthetic genes are preferentially expressed in anther tapetal cells and a few are also expressed in developing microspores. CYP703A2, an in-chain hydroxylase, is present in the tapetum and developing microspores [9]. Cyp703a2 mutant pollen development is indistinguishable from that of the wild type until after release from the tetrad when two sizes of pollen are observed, one of similar size to wild type and another much smaller that appears to have been arrested in development soon after free microspore release [9]. PKSA and PKSB are also expressed in the tetrads of developing microspores but neither single nor double mutants exhibit a reduced pollen size, possibly because of the observed minimal tetrad gene expression [14]. The smaller size of ascl-2 and Ppms2-1 spores and cyp703a2 pollen is atypical. Most other mutants containing mutated sporopollenin biosynthesis genes produce normal-sized mature pollen grains. Smaller ascl-2 spore size may be the result of premature arrest of spore development and perhaps indicates earlier expression of PpASCL in spore development compared to the time of expression of most sporopollenin biosynthesis genes in Arabidopsis pollen.
Alexander’s or Simplified Alexander’s stain has been used to visualize the differences between aborted and non-aborted pollen [32, 33]. NtPKS1i, a Nicotiana tabacum RNAi line with knocked down expression of NtPKS1, a PpASCL ortholog, is male sterile. Pollen grains from mature NtPKS1i anthers do not contain cytoplasm as demonstrated by Alexander’s staining. Wang et al. [43] suggested that loss of the cytoplasm results from cytoplasmic leakage through a structurally compromised cell wall during microspore development. Similarly, ascl-2 spores are non-viable and Simplified Alexander’s staining revealed that the majority of them exhibit an irregular distribution of cytoplasm. The wall of ascl-2 spores may be compromised in a similar way to that of NtPKS1i pollen, early in free microspore development, causing developmental arrest and abortion of the spores from cytoplasmic leakage.
Since intine, which is substantially comprised of pectin, appears to be absent or greatly reduced in the wall of ascl-2 spores, we cannot be certain that PpASCL is involved exclusively in sporopollenin synthesis. However, we suggest that the most probable explanation of the lack of intine in this case is that the development of ascl spores is arrested prior to intine deposition as a result of compromised sporopollenin synthesis.
Defective sporopollenin is resistant to acetolysis
Acetolysis treatment is used to determine chemical resistance of the sporopollenin, and defects thereof, in spore and pollen walls. Acetolysis experiments with pollen from Arabidopsis strains mutated in MS2, CYP704B1, PKSA and PKSB reveal various levels of chemical sensitivity. MS2 pollen grains are moderately sensitive to acetolysis since most of them are lysed but retain remnants of pollen cell wall after this treatment [44]. Pollen grains from a cytochrome P450 mutant, cyp704b1, lack a distinguishable exine layer and are destroyed by acetolysis [10]. Single mutants of PKSA (LAP6) and PKSB (LAP5) produce pollen that are resistant to acetolysis, retaining their shape and overall structure [37]. The ascl-2 spore wall is as resistant to acetolysis as that of the control with the distinct spore morphologies of these strains being unaffected. Specifically, the spines of the control and the irregular granula of ascl-2 spores are readily discernible in SEM images of acetolysed spores. This indicates that, despite the structural integrity and organization of the ascl-2 spore cell wall being compromised, it contains a defective sporopollenin with chemical resistance equivalent to that of control sporopollenin. Similar resistance of a defective sporopollenin to acetolysis was observed in the Physcomitrella MS2 knockout line [8]. Such observations oblige a cautious interpretation of the level of acetolysis resistance as a means of assessing the condition of sporopollenin.
The structure of sporopollenin is poorly characterized compared to that of other biopolymers mainly due to its resistance to chemical and biochemical treatments. In particular, the chemical nature of in planta ASCL products and their contributions to sporopollenin are unclear. Studying the defective sporopollenin of ascl-2 and other knockout lines of sporopollenin biosynthesis genes should provide clarification.
In conclusion, the abnormal phenotype of ascl-2, an ASCL knockout mutant in the moss, Physcomitrella patens, is consistent with a role for PpASCL in exine and perine formation during spore development and provides additional support for an ancient, evolutionarily conserved pathway of sporopollenin biosynthesis. Finally, it seems reasonable to speculate that some or perhaps all the anatomical abnormalities of ascl sporophytes result secondarily from the early arrestment of spore development in ascl mutants.
Supporting Information
Southern blot of AclI restricted pabB4 control and ascl-2 gDNA, hybridized with a DIG-labelled probe specific to the npt-II resistance cassette. Probe hybridization is denoted by an arrowhead. The positions of DNA size markers are indicated at left. The expected size of the AclI restriction fragment from a recombinant, i.e. mutant, PpASCL allele is 5991 bp.
(PDF)
Six weeks old, vegetatively propagated colonies of pabB4 and ascl-2. Scale bars = 5 mm.
(PDF)
(a–d) Photomicrographs of spores isolated from mature sporophytes. Fractured ascl spore walls are clearly visible in (b) and (c). Ascl plants produced smaller spores. The diameter of control spores was 28 ± 2.0 μm (n = 132, mean ± S.D.); the longest dimension of ascl spores was 20 ± 1.7 μm (n = 63), 18 ± 1.9 (n = 146) and 19 ± 2.2 (n = 39) in ascl-1, ascl-2 and ascl-3, respectively. The ascl spores were significantly smaller than the control spores (ANOVA, p < 0.0001). (e–l) SEM images of fixed, gold-coated spores. All scale bars = 10 μm.
(PDF)
Cross sections of control (a) and ascl-2 (b,c) spores from orange sporophytes were examined by TEM. In the control, the spore wall is intact and the spore is filled with cytoplasm, which here shows little structural resolution except for the faint outlines of oil droplets. The integrity of the spore wall in ascl-2 has been compromised and the arrows in (b) and (c) indicate the position of an opening or fissure in the wall. The ascl-2 spores are only partially filled with cytoplasm, presumably because of leakage of cytoplasm from the spores via the fissure in the mutant spore wall. There also appears to be uneven deposition of perine or perine-like material at the surface of the remaining cytoplasm within the ascl-2 spores. The perine projections on the outside of ascl-2 spores are smoother and less pointed than those on the outside of control spores. Magnified images of areas outlined in red in (a) and (c) are shown in Fig 7A and 7B, respectively. Scale bars = 2 μm.
(PDF)
(PDF)
(PDF)
The number of sporophytes at each developmental stage is given as a percentage of the total number in a culture tube.
(PDF)
Acknowledgments
We thank Drs. Tomoaki Nishiyama and Mitsuyasu Hasebe (NIBB, Japan) for providing the pTN182 vector.
Data Availability
All relevant data are within the paper and its Supporting Information file.
Funding Statement
This work was funded by the Natural Sciences and Engineering Research Council of Canada (262038-2013, www.nserc-crsng.gc.ca, to DYS) and Natural Sciences and Engineering Research Council of Canada (2982-2008, www.nserc-crsng.gc.ca, to NWA). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Kenrick P, Crane PR. The origin and early evolution of plants on land. Nature. 1997; 389:33−39. [Google Scholar]
- 2.Bowman JL, Floyd SK, Sakakibara K. Green genes–comparative genomics of the green branch of life. Cell. 2007; 129:229−234. [DOI] [PubMed] [Google Scholar]
- 3.Scott RJ, Spielman M, Dickinson HG. Stamen structure and function. Plant Cell. 2004; 16:S46–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Domínguez E, Mercado JA, Quesada MA, Heredia A. Pollen sporopollenin: degradation and structural elucidation. Sex Plant Reprod. 1999; 12:171−178. [Google Scholar]
- 5.Renzaglia KS, Schuette S, Duff RJ, Ligrone R, Shaw AJ, Mishler BD, et al. Bryophyte phylogeny: advancing the molecular and morphological frontiers. Bryologist. 2007; 110:170–213. [Google Scholar]
- 6.Kim SS, Douglas CJ. Sporopollenin monomer biosynthesis in Arabidopsis. J Plant Biol. 2013; 56:1–6. [Google Scholar]
- 7.Chen W, Yu XH, Zhang K, Shi J, De Oliveira S, Schreiber L, et al. Male Sterile2 encodes a plastid-localized fatty acyl carrier protein reductase required for pollen exine development in Arabidopsis. Plant Physiol. 2011; 157:842–853. 10.1104/pp.111.181693 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wallace S, Chater CC, Kamisugi Y, Cuming AC, Wellman CH, Beerling DJ, et al. Conservation of Male Sterility 2 function during spore and pollen wall development supports an evolutionarily early recruitment of a core component in the sporopollenin biosynthetic pathway. New Phytol. 2015; 205:390–401. 10.1111/nph.13012 [DOI] [PubMed] [Google Scholar]
- 9.Morant M, Jørgensen K, Schaller H, Pinot F, Møller BL, Werck-Reichhart D, et al. CYP703 is an ancient cytochrome P450 in land plants catalyzing in-chain hydroxylation of lauric acid to provide building blocks for sporopollenin synthesis in pollen. Plant Cell. 2007; 19:1473–1487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dobritsa AA, Shrestha J, Morant M, Pinot F, Matsuno M, Swanson R, et al. CYP704B1 is a long-chain fatty acid omega-hydroxylase essential for sporopollenin synthesis in pollen of Arabidopsis. Plant Physiol. 2009; 151:574–589. 10.1104/pp.109.144469 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rozema J, Broekman RA, Blokker P, Meijkamp BB, de Bakker N, van de Staaij J, et al. UV-B absorbance and UV-B absorbing compounds (para-coumaric acid) in pollen and sporopollenin: the perspective to track historic UV-B levels. J Photochem Photobiol B. 2001; 62:108–117. [DOI] [PubMed] [Google Scholar]
- 12.de Azevedo Souza C, Kim SS, Koch S, Kienow L, Schneider K, McKim SM, et al. A novel fatty acyl-CoA synthetase is required for pollen development and sporopollenin biosynthesis in Arabidopsis. Plant Cell. 2009; 21:507–525. 10.1105/tpc.108.062513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Grienenberger E, Kim SS, Lallemand B, Geoffroy P, Heintz D, de Azevedo Souza C, et al. Analysis of TETRAKETIDE α-PYRONE REDUCTASE function in Arabidopsis thaliana reveals a previously unknown, but conserved, biochemical pathway in sporopollenin monomer biosynthesis. Plant Cell. 2010; 22:4067–4083. 10.1105/tpc.110.080036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kim SS, Grienenberger E, Lallemand B, Colpitts CC, Kim SY, de Azevedo Souza C, et al. LAP6/POLYKETIDE SYNTHASE A and LAP5/POLYKETIDE SYNTHASE B encode hydroxyalkyl α-pyrone synthases required for pollen development and sporopollenin biosynthesis in Arabidopsis thaliana. Plant Cell. 2010; 22:4045–4066. 10.1105/tpc.110.080028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jepson C, Karppinen K, Daku RM, Sterenberg BT, Suh D-Y. Hypericum perforatum hydroxyalkylpyrone synthase involved in sporopollenin biosynthesis–phylogeny, site-directed mutagenesis and expression in non-anther tissues. FEBS J. 2014; 281:3855–3868. 10.1111/febs.12920 [DOI] [PubMed] [Google Scholar]
- 16.Wellman CH, Osterloff PL, Mohiuddin U. Fragments of the earliest land plants. Nature. 2003; 425:282−285. [DOI] [PubMed] [Google Scholar]
- 17.Lang D, Zimmer AD, Rensing SA, Reski R. Exploring plant biodiversity: the Physcomitrella genome and beyond. Trends Plant Sci. 2008; 13:542–549. 10.1016/j.tplants.2008.07.002 [DOI] [PubMed] [Google Scholar]
- 18.Hohe A, Egener T, Lucht JM, Holtorf A, Reinhard C, Schween G, et al. An improved and highly standardised transformation procedure allows efficient production of single and multiple targeted gene-knockouts in a moss, Physcomitrella patens. Curr Genet. 2004; 44:339–347. [DOI] [PubMed] [Google Scholar]
- 19.Rensing SA, Lang D, Zimmer AD, Terry A, Salamov A, Shapiro H, et al. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science. 2008; 319:64−69. [DOI] [PubMed] [Google Scholar]
- 20.Hiss M, Laule O, Meskauskiene RM, Arif MA, Decker EL, Erxleben A, et al. Large scale gene expression profiling data of the model moss Physcomitrella patens aid understanding developmental progression, culture and stress conditions. Plant J. 2014; 79:530–539. 10.1111/tpj.12572 [DOI] [PubMed] [Google Scholar]
- 21.Colpitts CC, Kim SS, Posehn SE, Jepson C, Kim SY, Wiedemann G, et al. PpASCL, a moss ortholog of anther-specific chalcone synthase-like enzymes, is a hydroxyalkylpyrone synthase involved in an evolutionarily conserved sporopollenin biosynthesis pathway. New Phytol. 2011; 192:855–868. 10.1111/j.1469-8137.2011.03858.x [DOI] [PubMed] [Google Scholar]
- 22.Wallace S, Fleming A, Wellman CH, Beerling DJ. Evolutionary development of the plant and spore wall. AoB Plants. 2011; plr027 10.1093/aobpla/plr027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ashton NW, Cove DJ. The isolation and preliminary characterisation of auxotrophic and analogue resistant mutants of the moss, Physcomitrella patens. Mol Gen Genet. 1977; 154:87–95. [Google Scholar]
- 24.Singer SD, Krogan NT, Ashton NW. 2007. Clues about the ancestral roles of plant MADS-box genes from a functional analysis of moss homologues. Plant Cell Rep. 2007; 26:1155–1169. [DOI] [PubMed] [Google Scholar]
- 25.Knight CD, Cove DJ, Boyd PJ, Ashton NW. The isolation of biochemical and developmental mutants in Physcomitrella patens In: Glime JM, editor. Methods in bryology. Nichinan, Miyazaki, Japan: Hattori Botanical Laboratory; 1988. pp. 47–58. [Google Scholar]
- 26.Grimsley NH, Ashton NW, Cove DJ. The production of somatic hybrids by protoplast fusion in the moss, Physcomitrella patens. Mol Gen Genet. 1977; 154:97–100. [Google Scholar]
- 27.Ashton NW, Schulze A, Hall P, Bandurski RS. Estimation of indole-3-acetic acid in gametophytes of the moss, Physcomitrella patens. Planta. 1985; 164:142–144. [PubMed] [Google Scholar]
- 28.Sakakibara K, Nishiyama T, Deguchi H, Hasebe M. Class 1 KNOX genes are not involved in shoot development in the moss Physcomitrella patens but do function in sporophyte development. Evol Dev. 2008; 10:555–566. 10.1111/j.1525-142X.2008.00271.x [DOI] [PubMed] [Google Scholar]
- 29.Ashton NW, Champagne CEM, Weiler T, Verkoczy LK. The bryophyte Physcomitrella patens replicates extrachromosomal transgenic elements. New Phytol. 2000; 146: 391–402. [Google Scholar]
- 30.Engel PP. The induction of biochemical and morphological mutants in the moss Physcomitrella patens. Am J Bot. 1968; 55:438–446. [Google Scholar]
- 31.Courtice GRM, Ashton NW, Cove DJ. Evidence for the restricted passage of metabolites into the sporophyte of the moss Physcomitrella patens (Hedw.) Br. Eur. J Bryol. 1978; 2:191–198. [Google Scholar]
- 32.Alexander MP. Differential staining of aborted and non-aborted pollen. Biotech Histochem. 1969; 44:117–122. [DOI] [PubMed] [Google Scholar]
- 33.Peterson R, Slovin JP, Chen C. A simplified method for differential staining of aborted and non-aborted pollen grains. Int J Plant Biol. 2010; 1:66–70. [Google Scholar]
- 34.Karnovsky M. 1965. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J Cell Biol. 1965; 27:137A–138A. [Google Scholar]
- 35.Singer SD, Ashton NW. Revelation of ancestral roles of KNOX genes by a functional analysis of Physcomitrella homologues. Plant Cell Rep. 2007; 26:2039–2054. [DOI] [PubMed] [Google Scholar]
- 36.Tang LK, Chu H, Yip WK, Yeung EC, Lo C. An anther-specific dihydroflavonol 4-reductase-like gene (DRL1) is essential for male fertility in Arabidopsis. New Phytol. 2009; 181:576–587. 10.1111/j.1469-8137.2008.02692.x [DOI] [PubMed] [Google Scholar]
- 37.Dobritsa AA, Lei Z, Nishikawa S-I, Urbanczyk-Wochniak E, Huhman DV, Preuss D, et al. LAP5 and LAP6 encode anther-specific proteins with similarity to chalcone synthase essential for pollen exine development in Arabidopsis. Plant Physiol. 2010; 153:937–955. 10.1104/pp.110.157446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Punt W, Hoen PP, Blackmore S, Nilsson S, Le Thomas A. Glossary of pollen and spore terminology. Rev Palaeobot Palynol. 2007; 143:1–81. [Google Scholar]
- 39.Schuette S, Wood AJ, Geisler M, Geisler-Lee J, Ligrone R, Renzaglia KS. Novel localization of callose in the spores of Physcomitrella patens and phylogenomics of the callose synthase gene family. Ann Bot. 2009; 103:749–756. 10.1093/aob/mcn268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.O’Donoghue T-M, Chater C, Wallace S, Gray JE, Beerling DJ, Fleming AJ. Genome-wide transcriptomic analysis of the sporophyte of the moss Physcomitrella patens. J Exp Bot. 2013; 64:3567–3581. 10.1093/jxb/ert190 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Aya K, Hiwatashi Y, Kojima M, Sakakibara H, Ueguchi-Tanaka M, Hasebe M, Matsuoka M. The Gibberellin perception system evolved to regulate a pre-existing GAMYB-mediated system during land plant evolution. Nat Commun. 2011; 2:544 10.1038/ncomms1552 [DOI] [PubMed] [Google Scholar]
- 42.Fraser WT, Scott AC, Forbes AES, Glasspool IJ, Plotnick RE, Kenig F, et al. Evolutionary stasis of sporopollenin biochemistry revealed by unaltered Pennsylvanian spores. New Phytol. 2012; 196:397–401. 10.1111/j.1469-8137.2012.04301.x [DOI] [PubMed] [Google Scholar]
- 43.Wang Y, Lin Y-C, So J, Du Y, Lo C. Conserved metabolic steps for sporopollenin precursor formation in tobacco and rice. Physiol Plant. 2013; 149:13–24. 10.1111/ppl.12018 [DOI] [PubMed] [Google Scholar]
- 44.Aarts MGM, Hodge R, Kalantidis K, Florack D, Wilson ZA, Mulligan BJ, et al. The Arabidopsis MALE STERILITY 2 protein shares similarity with reductases in elongation/condensation complexes. Plant J. 1997; 12:615–623. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Southern blot of AclI restricted pabB4 control and ascl-2 gDNA, hybridized with a DIG-labelled probe specific to the npt-II resistance cassette. Probe hybridization is denoted by an arrowhead. The positions of DNA size markers are indicated at left. The expected size of the AclI restriction fragment from a recombinant, i.e. mutant, PpASCL allele is 5991 bp.
(PDF)
Six weeks old, vegetatively propagated colonies of pabB4 and ascl-2. Scale bars = 5 mm.
(PDF)
(a–d) Photomicrographs of spores isolated from mature sporophytes. Fractured ascl spore walls are clearly visible in (b) and (c). Ascl plants produced smaller spores. The diameter of control spores was 28 ± 2.0 μm (n = 132, mean ± S.D.); the longest dimension of ascl spores was 20 ± 1.7 μm (n = 63), 18 ± 1.9 (n = 146) and 19 ± 2.2 (n = 39) in ascl-1, ascl-2 and ascl-3, respectively. The ascl spores were significantly smaller than the control spores (ANOVA, p < 0.0001). (e–l) SEM images of fixed, gold-coated spores. All scale bars = 10 μm.
(PDF)
Cross sections of control (a) and ascl-2 (b,c) spores from orange sporophytes were examined by TEM. In the control, the spore wall is intact and the spore is filled with cytoplasm, which here shows little structural resolution except for the faint outlines of oil droplets. The integrity of the spore wall in ascl-2 has been compromised and the arrows in (b) and (c) indicate the position of an opening or fissure in the wall. The ascl-2 spores are only partially filled with cytoplasm, presumably because of leakage of cytoplasm from the spores via the fissure in the mutant spore wall. There also appears to be uneven deposition of perine or perine-like material at the surface of the remaining cytoplasm within the ascl-2 spores. The perine projections on the outside of ascl-2 spores are smoother and less pointed than those on the outside of control spores. Magnified images of areas outlined in red in (a) and (c) are shown in Fig 7A and 7B, respectively. Scale bars = 2 μm.
(PDF)
(PDF)
(PDF)
The number of sporophytes at each developmental stage is given as a percentage of the total number in a culture tube.
(PDF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information file.