Abstract
Novel species of fungi described in the present study include the following from Australia: Neoseptorioides eucalypti gen. & sp. nov. from Eucalyptus radiata leaves, Phytophthora gondwanensis from soil, Diaporthe tulliensis from rotted stem ends of Theobroma cacao fruit, Diaporthe vawdreyi from fruit rot of Psidium guajava, Magnaporthiopsis agrostidis from rotted roots of Agrostis stolonifera and Semifissispora natalis from Eucalyptus leaf litter. Furthermore, Neopestalotiopsis egyptiaca is described from Mangifera indica leaves (Egypt), Roussoella mexicana from Coffea arabica leaves (Mexico), Calonectria monticola from soil (Thailand), Hygrocybe jackmanii from littoral sand dunes (Canada), Lindgomyces madisonensis from submerged decorticated wood (USA), Neofabraea brasiliensis from Malus domestica (Brazil), Geastrum diosiae from litter (Argentina), Ganoderma wiiroense on angiosperms (Ghana), Arthrinium gutiae from the gut of a grasshopper (India), Pyrenochaeta telephoni from the screen of a mobile phone (India) and Xenoleptographium phialoconidium gen. & sp. nov. on exposed xylem tissues of Gmelina arborea (Indonesia). Several novelties are introduced from Spain, namely Psathyrella complutensis on loamy soil, Chlorophyllum lusitanicum on nitrified grasslands (incl. Chlorophyllum arizonicum comb. nov.), Aspergillus citocrescens from cave sediment and Lotinia verna gen. & sp. nov. from muddy soil. Novel foliicolous taxa from South Africa include Phyllosticta carissicola from Carissa macrocarpa, Pseudopyricularia hagahagae from Cyperaceae and Zeloasperisporium searsiae from Searsia chirindensis. Furthermore, Neophaeococcomyces is introduced as a novel genus, with two new combinations, N. aloes and N. catenatus. Several foliicolous novelties are recorded from La Réunion, France, namely Ochroconis pandanicola from Pandanus utilis, Neosulcatispora agaves gen. & sp. nov. from Agave vera-cruz, Pilidium eucalyptorum from Eucalyptus robusta, Strelitziana syzygii from Syzygium jambos (incl. Strelitzianaceae fam. nov.) and Pseudobeltrania ocoteae from Ocotea obtusata (Beltraniaceae emend.). Morphological and culture characteristics along with ITS DNA barcodes are provided for all taxa.
Keywords: ITS DNA barcodes, LSU, novel fungal species, systematics
Overview Basidiomycota phylogeny
First of 12 equally most parsimonious trees obtained from the LSU alignment (215 parsimony-informative, 131 parsimony-uninformative and 463 constant characters) based on a heuristic analysis with simple taxon additions and tree-bisection-reconnection algorithm using PAUP v. 4.0b10 (TL = 673, CI = 0.719, RI = 0.852, RC = 0.613). GenBank accession numbers are indicated behind the species names. Bootstrap support values > 50 % from 100 000 fast replicates are shown at the nodes and the scale bar indicates the number of changes. Families, orders and classes are indicated with coloured blocks to the right of the tree. Strict consensus branches are indicated with thickened lines. The tree was rooted to Saccharomyces cerevisiae (GenBank Z73326) and the novel species described in this study for which LSU sequence data were available are indicated in bold face. The alignment and tree were deposited in TreeBASE (Submission ID 18408).

Overview Sordariomycetes phylogeny
First of 260 equally most parsimonious trees obtained from the LSU alignment (213 parsimony-informative, 115 parsimony-uninformative and 453 constant characters) based on a heuristic analysis with simple taxon additions and tree-bisection-reconnection algorithm using PAUP v. 4.0b10 (TL = 939, CI = 0.520, RI = 0.891, RC = 0.463). GenBank accession numbers are indicated behind the species names. Bootstrap support values > 50 % from 100 000 fast replicates are shown at the nodes and the scale bar indicates the number of changes. Families, orders and classes are indicated with coloured blocks to the right of the tree. Strict consensus branches are indicated with thickened lines. The tree was rooted to Saccharomyces cerevisiae (GenBank Z73326) and the novel species described in this study for which LSU sequence data were available are indicated in bold face. The alignment and tree were deposited in TreeBASE (Submission ID 18408).

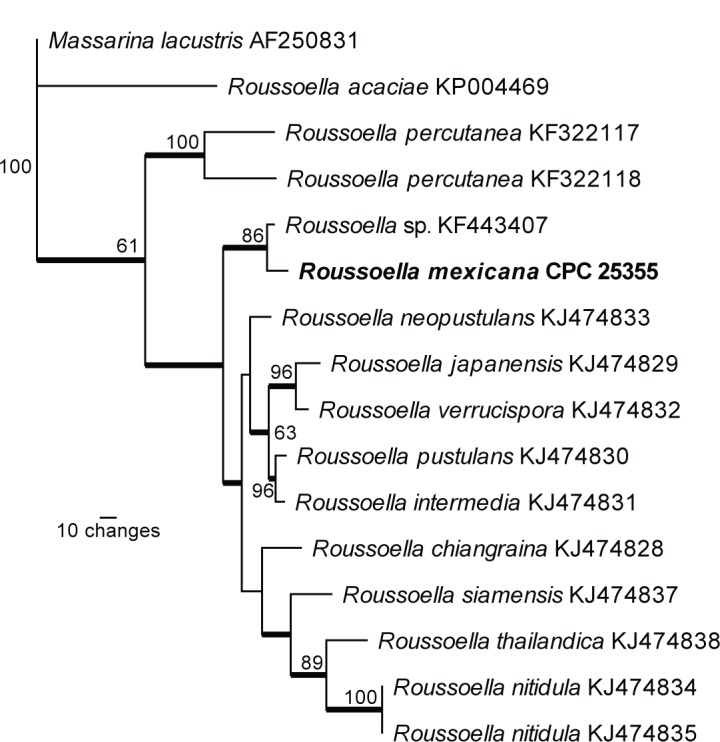
Overview Dothideomycetes and other classes phylogeny
First of 1 000 equally most parsimonious trees obtained from the LSU alignment (308 parsimony-informative, 44 parsimony-uninformative and 417 constant characters) based on a heuristic analysis with simple taxon additions and tree-bisection-reconnection algorithm using PAUP v. 4.0b10 (TL = 1 536, CI = 0.414, RI = 0.844, RC = 0.349). GenBank accession numbers are indicated behind the species names. Bootstrap support values > 50 % from 100 000 fast replicates are shown at the nodes and the scale bar indicates the number of changes. Families, orders and classes are indicated with coloured blocks to the right of the tree. Strict consensus branches are indicated with thickened lines. The tree was rooted to Saccharomyces cerevisiae (GenBank Z73326) and the novel species described in this study for which LSU sequence data were available are indicated in bold face. The alignment and tree were deposited in TreeBASE (Submission ID 18408).

Acknowledgments
Michel Heykoop, Gabriel Moreno and Pablo Alvarado wish to express their gratitude to Dr L. Örstadius for kindly sending them Swedish material of Psathyrella effibulata and P. complutensis as well as colour photographs and notes; to Dr L. Monje and Mr A. Pueblas of the Department of Drawing and Scientific Photography at the University of Alcalá for their help in the digital preparation of the photographs; to Dr J. Rejos, curator of the AH herbarium for his assistance with the specimens examined. The research of Miroslav Kolařík and Alena Nováková was supported through a grant from the Czech Science Foundation (P506-12-1064). Rosa M. V. Sanhueza, Amauri Bogo, C. André Lévesque, Tharcisse Barasubiye and Tara Rintoul acknowledge Dr Patricia Ritschel (EMBRAPA, Brazilian Corporation of Agricultural Research) and Carla Comparin (Santa Catarina State University) for their assistance with the description of Neofabraea brasiliensis. Juan Carlos Zamora thanks Yolanda Ruiz (UTAI, Real Jardín Botánico-CSIC) for her technical assistance with the SEM, Guillermo Rolón (IAA, FADU-UBA) for the photograph of the habitat, and financial support from the Consejo Superior de Investigaciones Científicas (Jae-Pre 2010). Rohit Sharma acknowledges support from the Department of Biotechnology, New Delhi, for funding the Microbial Culture Collection (MCC), NCCS, Pune, India (BT/PR10054/NDB/52/94/2007) and Mr Mitesh Khairnar (MCC, Pune) for photographs. Renée Lebeuf, Greg Thorn, David Boertmann and Andrus Voitk, owe thanks to Bill Roody for collecting samples of Hygrocybe andersonii, Hashini Puwakgolle for sequencing work and Michael Burzynski, Phyllis Mann, Henry Mann, Anne Marceau and Maria Voitk for help and companionship on the collecting trip to Labrador. We also thank the CBS technical staff, A. van Iperen (cultures), M. Vermaas (photographic plates) and M. Starink-Willemse (DNA isolation, amplification and sequencing) for their invaluable assistance.
REFERENCES
- Alfenas RF, Lombard L, Pereira OL, et al. 2015. Diversity and potential impact of Calonectria species in Eucalyptus plantations in Brazil. Studies in Mycology 80: 89–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnolds E. 2003 Rare and interesting species of Psathyrella. Fungi non delineati raro vel haud perspecte et explorate descripti aut definite picti. Pars XXVI. Edizioni Candusso. [Google Scholar]
- Bogo A, Maffioletti MA, Sanhueza RMV, et al. 2008 Morphological characterization of Cryptosporiopsis perennans isolates in different culture media. Tropical Plant Pathology 33: 248–251. [Google Scholar]
- Bronckers RJC. 2002a Paratrichophaea macrocystis en Paratrichophaea michiganensis, twee zeldzame pelsbekertjes op mest van grote grazers in Zuid-Limburg. PSL-Nieuws 9: 5–9. [Google Scholar]
- Bronckers RJC. 2002b Afwijkingen bij Paratrichophaea macrocystis veroorzaakt door een mosmijt. PSL-Nieuws 9: 16–17. [Google Scholar]
- Bronckers RJC. 2003. Een sleutel tot de Europese soorten van de genera Trichophaea, Trichophaeopsis en Paratrichophaea. Sterbeeckia 23: 9–27. [Google Scholar]
- Calonge FD, Donadini JC, De la Torre M, et al. 1988 Trichophaea paraphysincrustata (Ascomycotina), especie nueva para la ciencia. Boletín de la Sociedad Micológica de Madrid 12: 27–33. [Google Scholar]
- Cheewangkoon R, Groenewald JZ, Summerell BA, et al. 2009 Myrtaceae, a cache of fungal biodiversity. Persoonia 23: 55–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C, Verkley GJM, Sun G, et al. 2015 Redefining common endophytes and plant pathogens in Neofabraea, Pezicula, and related genera. Fungal Biology. In press. doi.org/10.1016/j.funbio.2015.09.013. [DOI] [PubMed] [Google Scholar]
- Chen SF, Lombard L, Roux J, et al. 2011 Novel species of Calonectria associated with Eucalyptus leaf blight in Southeast China. Persoonia 26: 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW. 1993 New and interesting fungi. 13. Foliicolous microfungi. South African Journal of Botany 59: 602–610. [Google Scholar]
- Crous PW, Groenewald JZ. 2013 A phylogenetic re-evaluation of Arthrinium. IMA Fungus 4: 133–154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Knox-Davies PS, Wingfield MJ. 1989 A list of Eucalyptus leaf fungi and their potential importance to South African forestry. South African Forestry Journal 149: 17–29. [Google Scholar]
- Crous PW, Phillips AJL, Baxter AP. 2000 Phytopathogenic fungi from South Africa. University of Stellenbosch Printers, Department of Plant Pathology Press, Stellenbosch, South Africa. [Google Scholar]
- Crous PW, Schubert K, Braun U, et al. 2007 Opportunistic, human-pathogenic species in the Herpotrichiellaceae are phenotypically similar to saprobic or phytopathogenic species in the Venturiaceae. Studies in Mycology 58: 185–217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Seifert KA, Castañeda-Ruiz RF. 1996 Microfungi associated with Podocarpus leaf litter in South Africa. South African Journal of Botany 62: 89–98. [Google Scholar]
- Crous PW, Van der Linde EJ. 1993 New and interesting fungi. 11. Eucalyptus leaf fungi. South African Journal of Botany 59: 300–304. [Google Scholar]
- Crous PW, Wingfield MJ, Guarro J, et al. 2015 Fungal Planet description sheets: 320–370. Persoonia 34: 167–266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Wingfield MJ, Schumacher RK, et al. 2014 Fungal Planet description sheets 281–319. Persoonia 33: 212–289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Gruyter J, Woudenberg JHC, Aveskamp MM, et al. 2010 Systematic reappraisal of species in Phoma section Paraphoma, Pyrenochaeta and Pleurophoma. Mycologia 102: 1066–1081. [DOI] [PubMed] [Google Scholar]
- Dougoud R. 2002 Contribution à la connaissance de quelques Discomycètes operculés rares ou méconnus. Fungi non Delineati 18: 1–70. [Google Scholar]
- Ellis MB. 1971 Dematiaceous Hyphomycetes. Commonwealth Mycological Institute, Kew Surrey, England. [Google Scholar]
- Ellis MB. 1976 More dematiaceous Hyphomycetes. Commonwealth Mycological Institute, Kew Surrey, England. [Google Scholar]
- Erwin DC, Ribeiro OK. 1996 Phytophthora diseases worldwide. APS Press, St. Paul, Minnesota. [Google Scholar]
- Gams W. 2000 Phialophora and some similar morphologically little-differentiated anamorphs of divergent ascomycetes. Studies in Mycology 45: 187–200. [Google Scholar]
- Gamundi IJ. 1973 Discomycetes de tierra de fuego II. Especies nuevas de Humariaceae. Boletín de la Sociedad Argentina de Botánica 15: 85–92. [Google Scholar]
- Glienke C, Pereira OL, Stringari D, et al. 2011 Endophytic and pathogenic Phyllosticta species, with reference to those associated with Citrus Black Spot. Persoonia 26: 47–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hangsanan S, Tian Q, Bahkali AH, et al. 2015. Zeloasperisporiales ord. nov., and two new species of Zeloasperisporium. Cryptogamie, Mycologie 36: 301–317. [Google Scholar]
- Heredia G, Arias RM, Reyes M, et al. 2002 New anamorph fungi with rhombic conidia from Mexican tropical forest litter. Fungal Diversity 11: 99–107. [Google Scholar]
- Hirayama K, Tanaka K, Raja HA, et al. 2010 A molecular phylogenetic assessment of Massarina ingoldiana sensu lato. Mycologia 102: 729–746. [DOI] [PubMed] [Google Scholar]
- Hodges CS, Barr ME. 1971. A new species of Pseudomassaria and its Beltraniella conidial state. Mycologia 63: 562–566. [Google Scholar]
- Jacobs A, Coetzee MPA, Wingfield BD, et al. 2003 Phylogenetic relationships among Phialocephala species and other ascomycetes. Mycologia 95: 637–645. [DOI] [PubMed] [Google Scholar]
- Jacobs K, Wingfield MJ. 2001 Leptographium species: tree pathogens, in-sect associates, and agents of blue-stain. APS Press, St. Paul, Minnesota, USA. [Google Scholar]
- Ju YM, Rogers JD, Huhndorf SM. 1996 Valsaria and notes on Endoxylina, Pseudothyridaria, Pseudovalsaria, and Roussoella. Mycotaxon 58: 419–481. [Google Scholar]
- Kanouse BB. 1958 Some species of the genus Trichophaea. Mycologia 50: 121–140. [Google Scholar]
- Kearse M, Moir R, Wilson A, et al. 2012 Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28: 1647–1649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kits van Waveren E. 1985 The Dutch, French and British species of Psathyrella. Persoonia Supplement 2: 1–300. [Google Scholar]
- Klaubauf S, Tharreau D, Fournier E, et al. 2014. Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Studies in Mycology 79: 85–120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korf RP, Erb RW. 1972 The new genus Trichophaeopsis. Phytologia 24: 15–19. [Google Scholar]
- Landschoot PJ, Jackson N. 1989 Gaeumannomyces incrustans sp. nov., a root-infecting hyphopodiate fungus from grass roots in the United States. Mycological Research 93: 55–58. [Google Scholar]
- Larsson E, Örstadius L. 2008 Fourteen coprophilous species of Psathyrella identified in the Nordic countries using morphology and nuclear rDNA sequence data. Mycological Research 112: 1165–1185. [DOI] [PubMed] [Google Scholar]
- Lebel T, Syme A. 2012 Sequestrate species of Agaricus and Macrolepiota from Australia: new species and combinations and their position in a calibrated phylogeny. Mycologia 104: 496–520. [DOI] [PubMed] [Google Scholar]
- Lebeuf R, Thorn G, Boertmann D, et al. 2016 Hygrocybe jackmanii. Omphalina 7: 6–10. In press. <www.nlmushrooms.ca>. [Google Scholar]
- Lodge DJ, Padamsee M, Matheny PB, et al. 2014 Molecular phylogeny, morphology, pigment chemistry and ecology in Hygrophoraceae (Agaricales). Fungal Diversity 64: 1–99. [Google Scholar]
- Lombard L, Chen SF, Mou X, et al. 2015a New species, hyper-diversity and potential importance of Calonectria spp. from Eucalyptus in South China. Studies in Mycology 80: 151–188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lombard L, Van der Merwe N, Groenewald J, et al. 2015b. Generic concepts in Nectriaceae. Studies in Mycology 80: 189–245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo J, Walsh E, Zhang N. 2014 Four new species in Magnaporthaceae from grass roots in New Jersey Pine Barrens. Mycologia 106: 580–588. [DOI] [PubMed] [Google Scholar]
- Luo J, Zhang N. 2013 Magnaporthiopsis, a new genus in Magnaporthaceae (Ascomycota). Mycologia 105: 1019–1029. [DOI] [PubMed] [Google Scholar]
- Luttrell ES. 1954 An undescribed species of Pyricularia on sedges. Mycologia 46: 810–814. [Google Scholar]
- Maharachchikumbura SSN, Hyde KD, Groenewald JZ, et al. 2014 Pestalotiopsis revisited. Studies in Mycology 79: 121–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marincowitz S, Duong TA, De Beer ZW, et al. 2015 Cornuvesica: A little known mycophilic genus with a unique biology and unexpected new species. Fungal Biology 119: 615–630. [DOI] [PubMed] [Google Scholar]
- Moravec J. 1979 Trichophaeopsis latispora sp. nov., a new Discomycete from Moravia (Czechoslovakia). Ceská Mykologie 33: 13–18. [Google Scholar]
- Moreno G, Esqueda M, Pérez-Silva E, et al. 2007 Some interesting gasteroid and secotioid fungi from Sonora, Mexico. Persoonia 19: 265–280. [Google Scholar]
- Moreno-Rico O, Groenewald JZ, Crous PW. 2014 Foliicolous fungi from Arctostaphylos pungens in Mexico. IMA Fungus 5: 7–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy LG, Vágvölgyi C, Papp T. 2013 Morphological characterization of clades of the Psathyrellaceae (Agaricales) inferred from a multigene phylogeny. Mycological Progress 12: 505–517. [Google Scholar]
- Olson DM, Dinerstein E, Wikramanayake ED, et al. 2001 Terrestrial ecoregions of the world: A new map of life on earth. BioScience 51: 933–938. [Google Scholar]
- Phillips AJL, Alves A, Abdollahzadeh J, et al. 2013 The Botryosphaeriaceae: genera and species known from culture. Studies in Mycology 76: 51–167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pirozynski KA. 1963 Beltrania and related genera. Mycological Papers 90: 1–37. [Google Scholar]
- Posada D. 2008 jModelTest: Phylogenetic Model Averaging. Molecular Biology and Evolution 25: 1253–1256. [DOI] [PubMed] [Google Scholar]
- Punithalingam E. 1974 New species of Phomopsis. Transactions of the British Mycological Society 63: 229–236. [Google Scholar]
- Quaedvlieg W, Verkley GJM, Shin H-D, et al. 2013 Sizing up Septoria. Studies in Mycology 75: 307–390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raja HA, Oberlies NH, El-Elimat T, et al. 2013 Lindgomyces angustiascus, (Lindgomycetaceae, Pleosporales, Dothideomycetes), a new lignicolous species from freshwater habitats in the USA. Mycoscience 54: 353–361. [Google Scholar]
- Raja HA, Tanaka K, Hirayama K, et al. 2011 Freshwater ascomycetes: two new species of Lindgomyces (Lindgomycetaceae, Pleosporales, Dothideomycetes) from Japan and USA. Mycologia 103: 1421–1432. [DOI] [PubMed] [Google Scholar]
- Rao DPC, Agrawal SC, Seksena SB. 1976 Phomopsis destructum on Psidium guajava fruits in India. Mycologia 68: 1132–1134. [PubMed] [Google Scholar]
- Raper KB, Fennell DI. 1965. The genus Aspergillus. Williams & Wilkins Co., Baltimore, USA. [Google Scholar]
- Rayner RW. 1970 A mycological colour chart.Commonwealth Mycological Institute and British Mycological Society, Kew. [Google Scholar]
- Rossman AY. 1996 Morphological and molecular perspectives on systematics of the Hypocreales. Mycologia 88: 1–9. [Google Scholar]
- Rossman AY, Aime MC, Farr DF, et al. 2004 The coelomycetous genera Chaetomella and Pilidium represent a newly discovered lineage of inoperculate discomycetes. Mycological Progress 3: 275–290. [Google Scholar]
- Samerpitak K, Van der Linde E, Choi H-J, et al. 2014 Taxonomy of Ochroconis, genus including opportunistic pathogens on humans and animals. Fungal Diversity 65: 89–126. [Google Scholar]
- Samson RA, Visagie CM, Houbraken J, et al. 2014 Phylogeny, identification and nomenclature of the genus Aspergillus. Studies in Mycology 78: 141–173. [DOI] [PMC free article] [PubMed] [Google Scholar]
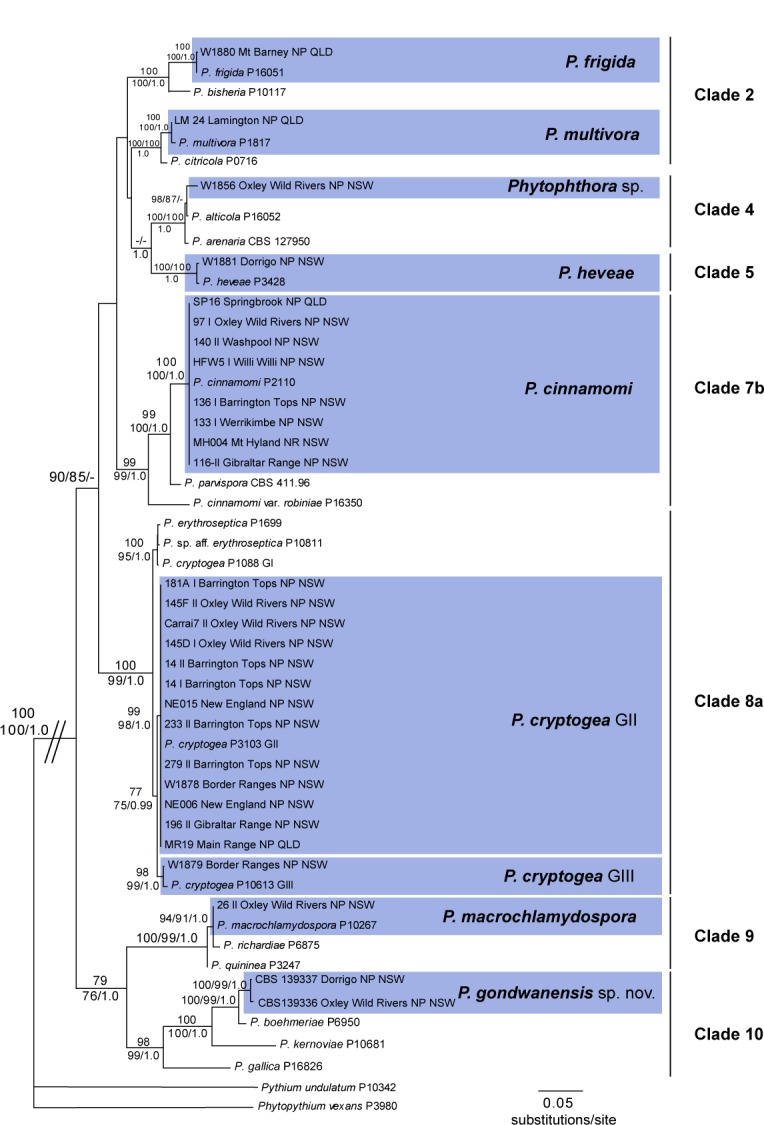
- Scarlett K, Daniel R, Shuttleworth LA, et al. 2015 Phytophthora in the Gondwana rainforests of Australia world heritage area. Australasian Plant Pathology 44: 335–348. [Google Scholar]
- Schoch CL, Robbertse B, Robert V, et al. 2014 Finding needles in haystacks: linking scientific names, reference specimens and molecular data for fungi. Database 2014: 1–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma R, Kulkarni G, Sonawane MS, et al. 2013 A new endophytic species of Arthrinium (Apiosporaceae) from Jatropha podagrica. Mycoscience 55: 118–123. [Google Scholar]
- Shear CL. 1902 Mycological notes and new species. Bulletin Torrey Botanical Club 29: 449–457. [Google Scholar]
- Slippers B, Boissin E, Phillips AJL, et al. 2013 Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Studies in Mycology 76: 31–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spegazzini C. 1898 Fungi Argentini novi vel critici. Anales del Museo Nacional de Historia Natural Buenos Aires 6: 81–288. [Google Scholar]
- Sunhede S. 1989 Geastraceae (Basidiomycotina). Morphology, ecology and systematics with special emphasis on the North European species. Synopsis Fungorum 1: 1–534. [Google Scholar]
- Sutton BC. 1980 The coelomycetes: fungi imperfecti with pycnidia, acervuli, and stromata. Kew, Commonwealth Mycological Institute. [Google Scholar]
- Swart HJ. 1982 Australian leaf-inhabiting fungi. XII. Semifissispora gen. nov. on dead Eucalyptus leaves. Transactions of the British Mycological Society 78: 259–264. [Google Scholar]
- Tanaka K, Hirayama K, Yonezawa H, et al. 2015 Revision of the Massarineae (Pleosporales, Dothideomycetes). Studies in Mycology 82: 75–136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trigaux G. 1985 Paratrichophaea macrocystis genre et espece noveaux. Documents Mycologiques 6: 1–6. [Google Scholar]
- Uecker FA. 1988 A world list of Phomopsis names with notes on nomenclature, morphology and biology. Mycologia Memoir 13: 1–231. [Google Scholar]
- Van der Aa HA, Vanev S. 2002 A revision of the species described in Phyllosticta. CBS, Utrecht, The Netherlands. [Google Scholar]
- Vellinga EC. 2002 New combinations in Chlorophyllum. Mycotaxon 83: 415–417. [Google Scholar]
- Vellinga EC. 2003 Notes on Chlorophyllum and Macrolepiota (Agaricacea) in Australia. Australian Systematic Botany 16: 361–370. [Google Scholar]
- Vellinga EC, De Kok RPJ, Bruns TD. 2003 Phylogeny and taxonomy of Macrolepiota (Agaricaceae). Mycologia 95: 442–456. [PubMed] [Google Scholar]
- Whitton SR, McKenzie EHR, Hyde KD. 2012 Fungi associated with Pandanaceae. Fungal Diversity Research Series 21: 1–457. [Google Scholar]
- Wikee S, Lombard L, Nakashima C, et al. 2013 A phylogenetic re-evaluation of Phyllosticta (Botryosphaeriales). Studies in Mycology 76: 1–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wingfield MJ, Van Wyk PS, Wingfield BD. 1987 Reclassification of Phialocephala based on conidial development. Transactions in British Mycological Society 89: 509–520. [Google Scholar]
- Wong PTW, Walker J. 1975 Germinating phialidic conidia of Gaeumannomyces graminis and Phialophora-like fungi from Gramineae. Transactions of the British Mycological Society 65: 41–47. [Google Scholar]
- Xu J-J, Qin S-Y, Hao Y-Y, et al. 2012 A new species of Calonectria causing leaf disease of water lily in China. Mycotaxon 112: 177–185. [Google Scholar]
- Yang CS, Korf R. 1985 Monograph of the genus Tricharina and of a new, segregate genus, Wilcoxina (Pezizales). Mycotaxon 24: 467–531. [Google Scholar]
- Zamora JC, Calonge FD, Hosaka K, et al. 2014 Systematics of the genus Geastrum (Fungi: Basidiomycota) revisited. Taxon 63: 477–497. [Google Scholar]
- Zamora JC, Calonge FD, Martín MP. 2015 Integrative taxonomy reveals an unexpected diversity in Geastrum section Geastrum (Geastrales, Basidiomycota). Persoonia 34: 130–165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Zhang X, Fournier J, et al. 2014 Lindgomyces griseosporus, a new aquatic ascomycete from Europe including new records. Mycoscience 55: 43–48. [Google Scholar]