

Abstract
Cadmium (Cd) is a highly toxic, nonessential heavy metal with many industrial uses that can contribute to a well-defined spectrum of diseases in animals as well as in humans. The present study examines the effect of zinc (Zn) and iron (Fe) supplementation on oxidative stress enzymes in Cd-treated rats. Wistar strain male albino rats were treated with cadmium chloride (CdCl2) at a dose of 1/10th LD50/48 h, that is, 22.5 mg/kg body weight for 7, 15, and 30 days (d) time intervals. The 15d Cd-treated rats were divided into three groups. The first group received Zn (12 mg/kg), second group Fe (40 mg/kg) alone, and third group supplemented with both Zn and Fe and observed for 7, 15, and 30d. After the specific time intervals, rats were decapitated and oxidative stress enzymes like catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx) and glutathione S-transferase (GST) were assayed in liver and kidney. Simultaneously lipid peroxidation (LPO) levels were also measured. A significant elevation in LPO levels with decreased activity levels of CAT, SOD, GPx, and GST were observed during Cd intoxication. With Zn and/or Fe supplementation, a significant reversal in the oxidative stress enzymes was observed. Our study reveals that combination of Zn and Fe supplementation is effective in detoxifying the Cd body burden from the test tissues.
Keywords: Cadmium, kidney, liver, oxidative stress, rat, trace elements supplementation
INTRODUCTION
Cadmium (Cd) is one of the most dangerous occupational and environmental pollutants, arising primarily from battery, electroplating, pigment, plastics, fertilizer industries, and cigarette smoke. It is an earth's crust natural element and found in drinking water, atmospheric air, and even in food (Moulis and Thevenod, 2010; Obaiah et al., 2013). Cd exposure has been recognized to result in a wide variety of cellular responses, oxidative stress, and body weight loss. Over the past 2 centuries, anthropogenic and industrial activities have led to high emissions of Cd into the environment at concentrations significantly exceeding those originating from natural sources (Jarup and Akesson, 2009; Cuypers et al., 2010; Dzugan et al., 2011; Nakamura et al., 2012). It has a serious toxicity in humans and animals and causes Itai-Itai disease (Nad et al., 2005) and also induces the onset of anemia, carcinogenicity, testicular atrophy, decreases red blood cell count, and hemoglobin concentration (Hartwig, 2010; Honda et al., 2010). Cd has an extremely long half-life (20–30 years) in the human body (Flora et al., 2008) and is highly cumulative, especially in the liver and kidney (Nordberg et al., 2007; Nawrot et al., 2008; Munga et al., 2010; Johri et al., 2010; Włostowski et al., 2010; Nakamura et al., 2012; Guirlet and Das, 2012).
It is a ubiquitous toxic metal and induces oxidative damage by disturbing the prooxidant–antioxidant balance in the tissues (Ognjanovic et al., 2008). Intake of Cd results in consumption of glutathione and protein binding sulfhydryl groups, and subsequently increases the levels of free radicals such as hydrogen peroxide (H2O2), hydroxide (OH–), and superoxide (O2–) anions. One of the most important effects of free radicals is oxidation of polyunsaturated fatty acids (PUFAs). As a result of free radical attack, lipids are oxidized; and hence membranes are damaged (Stohs et al., 2001; Ashraf and Kamar, 2007). Cd is readily distributed in tissues after ingestion and it inhibits oxidative stress enzymes such as catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx), and glutathione S-transferase (GST) (Asagba and Eriyamremu, 2007). As a result of this inhibition, the electron transport chain becomes highly reduced; electrons are transferred directly to available oxygen leading to enhanced formation of reactive oxygen species (ROS) (Tatrai et al., 2001). ROS may increase oxidative stress in tissues, cellular damage, peroxidation of membrane lipids, and loss of membrane bound enzymes (Liu et al., 2000). However, ROS may be used as an indicator of cell membrane injury. Cd interferes with intracellular signaling network and gene regulation at multiple levels, induces lipid peroxidation (LPO) and alterations in activity of oxidative stress enzymes (Wang et al., 2004; Sladan et al., 2005). At the cellular level, Cd induces oxidative stress in many organisms (Thevenod, 2009;Bertin and Averbeck, 2006), which might result in histological changes and physiological damage to different organs (Nawrot et al., 2008; Jarup and Akesson, 2009; Cuypers et al., 2010;Salinska et al., 2012). Antioxidants/oxidative stress enzymes prevent free radical-induced tissue damage by preventing the formation of radicals or scavenging them or by promoting their decomposition.
Trace elements supplementation has protective effect against Cd accumulation and toxicity in rats fed inorganic Cd salt (Matek et al., 2002; Piasek et al., 2004). The complex interrelationships between Cd and some essential trace elements have not been elucidated. Several essential trace elements like selenium (Se), copper (Cu), zinc (Zn), and iron (Fe) participate in controlling various metabolic and signaling pathways. In vitro studies suggests that there is a competition for transport mechanism between Cd and some essential trace elements like Zn and Cuand Fe in rats (Peraza et al., 1998; Hakan et al., 2001) and Zn and Se in Japanese quails (Nad et al., 2005) and calcium in suckling rats (Saric et al., 2002; Deepthi et al., 2010). Zn occurs in all living cells as a constituent of metalloenzymes involved in major metabolic pathways. Zn controls several enzymes of intermediary metabolism, DNA and RNA synthesis, gene expression, immunocompetence, and plays a significant role in homeostasis of hormones (Dzugan et al., 2012). Fe plays an essential role in biological processes. It is also a vital component of several enzymes and proteins (Cairo and Recalcati, 2007; Muckenthalerm et al., 2008; Qureshi et al., 2010). Fe supplementation reduces Cd retention and Cd-induced anemia in young rats (Schumann et al., 1996). Fe is a central element of the heme molecule, which is a critical part of hemoglobin and essential for oxygen transport. Heme is also a part of many essential enzymes such as CAT and cytochrome series (Brewer, 2007). In several studies, dietary Fe appeared to decrease Cd toxicity, but the exact mechanism of this interaction is unknown (Martinez et al., 2001; Park et al., 2002; Kwong et al., 2011). Hence, we made an attempt to study the interactions of Zn and/or Fe against Cd-induced toxicity in liver and kidney of a mammalian model, rat.
MATERIALS AND METHODS
Chemicals
Cd as cadmium chloride (CdCl2), Zn as zinc chloride (ZnCl2), and Fe as ferric chloride anhydrous (FeCl3) were purchased from Merck (Dormstadt, Germany). All other chemicals which were used in the present study were obtained from the standard chemical companies like Sigma Chemical Co. (St Louis, MO, USA) and SD Fine Chemicals, India. The chemicals used in this study were of the highest purity.
Animals
Three-months-old Wistar strain male albino rats weighing 180 ± 20 g were chosen for the present study. The animals were obtained from Sri Venkateswara Traders, Bangalore, Karnataka, India and were kept in stainless steel mesh cages, housed under standard laboratory conditions (23 ± 2°C, 50 ± 20% relative humidity, 12-h light-dark cycle) with standard rat chow (Sai Durga Feeds and Foods, Bangalore, India) and drinking water ad libitum. The rats were acclimatized to the laboratory conditions for 10 days. The protocol and animal use has been approved by the Institutional Animal Ethics Committee (Resol. No. 10(ii)/a/CPCSCA/IAEC/SVU/AUR-JO dt 22-12-2008), Sri Venkateswara University, Tirupati, Andhra Pradesh, India.
Experimental design
After acclimatization, the rats were divided into two groups, namely control and experimental. Control rats received only deionized water without Cd. The experimental rats were treated with Cd as CdCl2 at a dose of 1/10th LD50/48h, that is, 22.5 mg/kg body weight over a period of7, 15, and 30 days (d) time intervals. Then 15d Cd-treated rats were divided into three groups. Group I received supplementation of Zn (12 mg/kg) for 7, 15, and 30d. Group II received Fe supplementation (40 mg/kg) and Group III animals were supplemented with both Zn and Fe at the above said doses for 7, 15, and 30d long sojourn.
Isolation of tissues
After specific time intervals, the control and experimental rats were decapitated and tissues such as liver and kidney were quickly isolated under ice cold conditions and weighed to their nearest mg using Shimadzu electronic balance. After weighing, tissues were immediately used for the assay of oxidative stress enzymes like CAT, SOD, GST, GPx, and the levels of LPO and were stored at −80°C for future use.
Assay of oxidative stress enzymes
LPO
The LPO was determined by the thiobarbituric acid (TBA) method of Ohkawa et al., (1979). The tissues were homogenized in 1.5% KCl (20% W/V). To 1ml of tissue homogenate, 2.5 ml of 20% trichloroacetic acid (TCA) was added and the contents were centrifuged at 3,500 g for 10 min and the precipitate was dissolved in 2.5 ml of 0.05 M sulfuric acid. To this, 3 ml of TBA was added and the samples were kept in a hot water bath for 30 min. The samples were cooled and malonaldehyde (MDA) was extracted with 4 ml of n-butanol and the color was read at 530 nm in an ultraviolet (UV) spectrophotometer (Hitachi U-2000) against the reagent blank. Trimethoxy pentane (TMP) was used as external standard. Values are expressed in micromoles of MDA formed/gram tissue/hour.
CAT (EC: 1.11.1.6)
CAT activity was measured by a slightly modified method of Aebi (1984) at room temperature. The tissue was homogenized in ice-cold 50 mM phosphate buffer (pH 7.0) containing 0.1 m Methylene diamine tetra acetic acid (EDTA) to give 5% homogenate (W/V). The homogenates were centrifuged at 10,000 rpm for 10 min at 4°C in cold centrifuge. The resulting supernatant was used as enzyme source. A 10 μl of 100% ethyl alcohol (EtOH) was added to 100 μl of tissue extract and then placed in an ice bath for 30 min. After 30 min, the tubes were kept at room temperature followed by the addition of 10 μl of Triton X-100 RS. In a cuvette containing 200 μl of phosphate buffer, 50 μl of tissue extract and 250 μl of 0.006 M H2O2(in phosphate buffer) were added and the decrease in optical density was measured at 240 nm for 60s in a UV spectrophotometer (Hitachi U-2000). The molar extinction coefficient of 43.6 M cm−1 was used to determine CAT activity. One unit of activity is equal to the moles of H2O2 degraded/mg protein/min.
SOD (EC: 1.15.1.1)
SOD activity was determined according to the method of Misra and Fridovich (1972) at room temperature. The tissue was homogenized in ice-cold 50 mM phosphate buffer (pH 7.0) containing 0.1 mM EDTA to give 5% homogenate (W/V). The homogenates were centrifuged at 10,000 rpm for 10 min at 4°C in cold centrifuge. The supernatant was separated and used for enzyme assay. A 100 μl of tissue extract was added to 880 μl (0.05 M, pH 10.2, containing 0.1 mM EDTA) carbonate buffer; 20 μl of 30 mM epinephrine (in 0.05% acetic acid) was added to the mixture, and the optical density values were measured at 480 nm for 4 min using UVspectrophotometer (Hitachi U-2000). Values are expressed in superoxide anion reduced/milligram protein/minute.
GST (EC: 2.5.1.18)
GST activity was measured with its conventional substrate 1-chloro, 2,4-dinitrobenzene (CDNB) at 340 nm as per the method of Habig et al., (1974). The tissues were homogenized in 50 mM Tris-HCl buffer pH 7.4 containing 0.25 M sucrose and centrifuged at 4,000 g for 15 min at 4°C and the supernatant was again centrifuged at 16,000 g for 1 h at 4°C. The pellet was discarded and the supernatant was used as the enzyme source. The reaction mixture in a volume of 3 ml contained 2.4 ml of 0.3 M potassium phosphate buffer pH 6.9, 0.1 ml of 30 mM CDNB, 0.1 ml of 30 mM glutathione, and the appropriate enzyme source. The reaction was initiated by the addition of glutathione and the absorbance was read at 340 nm against reagent blank and the activity was expressed as micromoles of thioether formed/milligram protein/minute.
GPx (EC: 1.11.1.9)
GPx was determined by a modified method of Flohe and Gunzler (1984) at 37°C. A 5% (W/V) of tissue homogenate was prepared in 50 mM phosphate buffer (pH 7.0) containing 0.1 mM EDTA. The homogenates were centrifuged at 10,000 g for 10 min at 4°C in cold centrifuge. The resulting supernatant was used as enzyme source. The reaction mixture consisted of 500 μl of phosphate buffer, 100μl of 0.01 M GSH (reduced form), 100 μl of 1.5 mM NADPH, and 100 μl of glutathione reductase (GR,0.24 units). The 100 μl of tissue extract was added to the reaction mixture and incubated at 37°C for 10 min. Then 50 μl of 12 mM t-butyl hydroperoxide was added to 450 μl of tissue reaction mixture and measured at 340 nm for 180 s. The molar extinction coefficient of 6.22 × 103 M cm−1 was used to determine the activity. The enzyme activity was expressed in micromoles of NADPH oxidized/milligram protein/minute.
Estimation of protein content
Protein content of the tissues was estimated by the method of Lowry et al., (1951). One percent (W/V) homogenates of the tissues were prepared in 0.25 M ice-cold sucrose solution. To 0.5 ml of homogenate, 1 ml 10% TCA was added, and the samples were centrifuged at 1,000 g for 15 min. Supernatant was discarded and the residues were dissolved in 1 ml of 1N sodium hydroxide. To this, 4 ml of alkaline copper reagent was added followed by 0.4 ml of Folin-phenol reagent (1:1folin: H2O). The color was measured at 600 nm in a UV spectrophotometer (Hitachi U-2000) against reagent blank. The protein content of the tissues was calculated using a standard protein (bovine serum albumin (BSA)) graph.
Data analysis
The data was subjected to statistical analysis such as mean, standard deviation (SD), and analysis of variance (ANOVA) using standard statistical software, Statistical Package for Social Sciences (SPSS; version 16). All values are expressed as mean ± SD of six individual samples. Significant differences were indicated at P < 0.05 level.
RESULTS
The data on the alterations in the oxidative stress enzymes such as CAT, SOD, GST, and GPx as well as LPO in Cd-treated rat liver and kidney both before and after supplementation with Zn and/or Fe are tabulated in Tables 1–4. The decreased enzyme levels (CAT, SOD, GST, and GPx) in the Cd-treated rats were statistically significant (P < 0.05). The supplementation of Zn and/or Fe significantly reversed the Cd-induced alterations in the oxidative stress enzymes.
Table 1.
Alterations in the activity levels of selected oxidative stress enzymes in the liver of Cd-treated rats
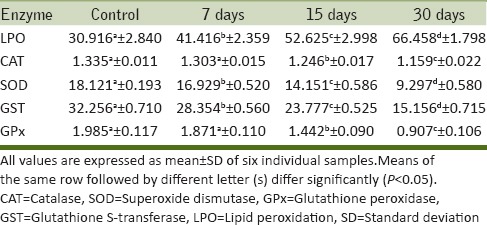
Table 4.
Alterations in the activity levels of selected oxidative stress enzymes in the kidney of Cd-treated rats after Zn and/or Fe supplementation
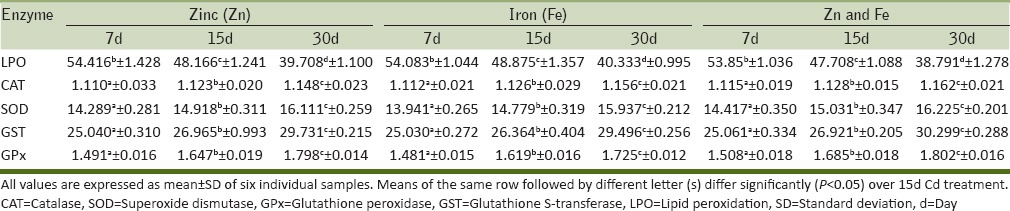
Our results revealed that LPO levels were increased markedly in both liver and kidney of Cd-treated rats at all the test periods when compared to the controls. The MDA levels were increased with the time intervals of Cd treatment compared to controls and were maximum for 30 d kidney (71.083 ± 2.113 µmole of MDA formed/g tissue/h). After supplementation with Zn and/or Fe, the LPO levels were progressively decreased at all time periods in both the test tissues. Maximum depletion was observed in 30d Zn and Fe mixture supplemented rat kidney (38.791 ± 1.278 µmole of MDA formed/g tissue/h).
CAT activity levels progressively decreased during Cd treatment. However, the activity reversed after supplementation with Zn and/or Fe. CAT activity levels were significantly reduced in 30d Cd-treated kidney (0.982 ± 0.029 µmole of H2O2 metabolized/mg protein/min) when compared to the liver tissue. However, supplementation of both Zn and Fe showed maximum elevation in CAT activity (1.324 ± 0.017 µmole of H2O2 metabolized/mg protein/min) levels for 30d liver tissue.
There was significant decrease in SOD activity during all time intervals of Cd treatment and it was maximum for 30d rat kidney (8.810 ± 0.567 superoxide anion reduced/mg protein/min). However with 30d Zn + Fe supplementation, rat liver showed maximum elevation in its activity (17.067 ± 0.188 superoxide anion reduced/mg protein/min) when compared to other time intervals of supplementation.
GST activity levels also showed a progressive decrement at all the time intervals of Cd treatment with a maximum depletion in 30d rat liver (15.146 ± 0.715 µmole of thioether formed/mg protein/min). Further supplementation with both Zn and Fe, the GST activity reached to normalcy in 30d rat kidney (30.299 ± 0.288 µmole of thioether formed/mg protein/min), suggesting the protective role of trace elements Zn and Fe.
GPx activity levels also showed a significant decrease at all the time intervals of Cd treatment with a maximum decrease in 30d kidney (0.822 ± 0.097 µmole of NADPH oxidized/mg protein/min). However, the aforesaid Cd-inhibited GPx activity levels were markedly elevated in both the test tissues after supplementation with the trace elements Zn and Fe individually as well as in combination. The combination of Zn and Fe at the time interval 30d as supplement was more effective in elevating the GPx activity levels in the liver tissue of rats (1.266 ± 0.008 µmole of NADPH oxidized/mg protein/min).
DISCUSSION
The results of the present study revealed that Cd induces oxidative stress as evidenced by increase in LPO and alterations in selected antioxidant enzyme status in liver and kidney of male albino rat at specific time intervals. However, supplementation of Zn and/or Fe reverses the Cd-induced changes in the LPO and antioxidant enzyme status.
Several mechanisms have been proposed for Cd-induced various abnormalities, but none have yet been defined explicitly. Disruption of a variety of biochemical processes have been proposed rather than a single mechanism responsible for the toxicity. Oxidative stress has recently been reported as one of the important mechanisms of toxic effects of Cd (Matovic et al., 2011).
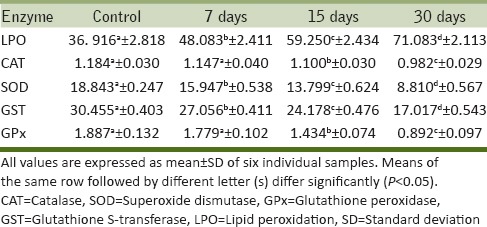
From the results, it is evident that LPO increased markedly in both liver and kidney of Cd-treated rats when compared to the control [Table 1 and 3]. MDA levels in the tissues are used as an index of oxidative stress. In our study, elevated MDA levels were observed in Cd-treated rats as an indication of increased oxidative stress. The increased MDA levels were more prominent in 30 d Cd treated rats when compared to that of other scheduled time intervals. Cd may induce oxidative damage in different tissues by enhancing peroxidation of membrane lipids and altering the antioxidant defense system of the cells. It may also decrease the absorption of trace elements in the tissues. As liver contains more amounts of unsaturated lipids, it undergoes oxidative damage by ROS than the kidney. Cd triggers the generation of ROS in the organism (Lucia et al., 2010;Olalekan et al., 2011; Jovanovic et al., 2012). In the present study, Cd-treated rats showed not only a significant increase in LPO but also significant decrease in the activity levels of antioxidant enzymes such as CAT, SOD, GST, and GPx [Tables 1–4]. These findings are in agreement with earlier reports of Stohs et al., (2001) and Kim et al., (2011) in rats and Carp, respectively, under Cd stress.
Table 3.
Alterations in the activity levels of selected oxidative stress enzymes in the kidney of Cd-treated rats
During oxidative stress the CAT activity levels progressively decreased due to high accumulation of H2O2 in the tissues, and thereby more peroxidation of lipids is favored. This could be the reason for increased LPO levels observed in Cd intoxicated rats. The significant decrease in CAT activity may be due to its inactivation by superoxide radical or due to decrease in the rate of reaction as a result of excess production of H2O2 to water and oxygen. It may alsobe due to the decreased absorption of essential trace element, Fe, required for the activity of this enzyme (Beytut and Aksakal, 2002; Jurczuk et al., 2004). The decrement in the CAT activity levels with Cd treatment indicates inefficient scavenging of hydrogen peroxide due to oxidative inactivation of enzyme.
SOD is an important antioxidant enzyme that inhibits oxyradical formation and is usually used as a biomarker to indicate oxidative stress (Zhang et al., 2004). The decrease in SOD activity may be due to its inhibition by the excess production of ROS as evidenced by LPO in the present study. Excessive production of ROS may result in alterations in the biological activity of cellular macromolecules. Therefore, the reduction in the activity of SOD may result in a number of deleterious effects due to the accumulation of superoxide radicals. Administration of Cd into rats in the present study may lead to generation of peroxy radical, O2−, which is associated with inactivation of SOD and CAT. This may be the reason for the significant reduction in the activities of CAT and SOD in liver and kidney of Cd-treated rats when compared to control. It is in accordance with results from earlier studies in which there was a decrease in SOD activity in rats (Nagaraj et al., 2000) and fish (Obaiah and Usha Rani, 2013) subjected to heavy metal, Cd.
GPx is a hydrogen peroxide degrading enzyme. Its activity was significantly decreased in both liver and kidney under Cd body burden at all the time intervals. The decreased GPx activity in the current study may be due to impairment in GSH homeostasis in liver and kidney tissues. As a result of this, liver and kidney tissue damage might have occurred under Cd insult. Recently, Waisberg et al., (2003), Ognjanovic et al., (2008), and Messaoudi et al., (2009) also reported decreased GPx activity in the liver and kidney tissues of rats under Cd stress. It may be due to either free radical dependent inactivation of enzyme or depletion of its cosubstrate, that is, GSH and NADPH in the Cd-treated rat liver and kidney. Cd-administered rat tissues showed decreased GSH content due to overutilization by the cells in the tissues. Due to nonbioavailability of GSH under Cd burden, decrement in the activity levels of GPx has also been observed in experimental tissues. Depletion of GSH may render in GPx inactivation and/or less activity (Mahendran and Shyamala Devi, 2001).
The reduced GST activity in the tissues may be due to overconsumption of the enzyme GST to escape from the toxicity of peroxides under Cd insult. GST catalyzes the reaction of the thiol(-SH) group of GSH with electrophilic reagents such as those generated by microsomal metabolism of xenobiotics; thereby, neutralizing their electrophilic sites and rendering the products more water soluble (Han et al., 2006). The decrease in GST activity might have resulted with Cd effect on GSH because of its high affinity to this molecule where a sulfhydryl acid, an amino acid, and two carboxylic acid groups, as well as two peptide linkages represent reactive sites for metals. Reactions of metals with glutathione might lead to either the formation of complexes or the oxidation of glutathione. The decreased GST activity in the test tissues is in agreement with El-Missiry and Shalaby (2000) in Cd-treated rat brain and testis. Moreover, the decrease in the activity of each of them would induce increased free radicals, thus injuring the corresponding tissues.
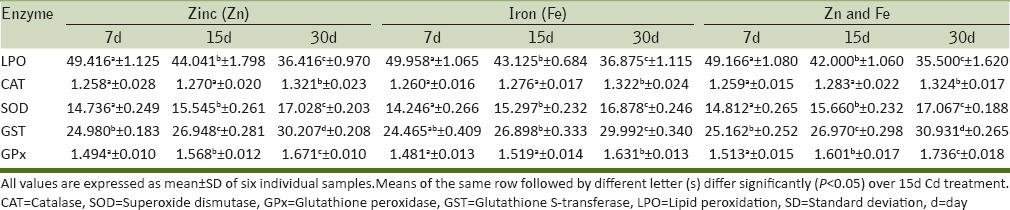
After supplementation with trace elements Zn and Fe individually and in combination there was significant reversal in the Cd-induced alterations in both liver and kidney of rats [Tables 2 and 4]. Zn provides an antioxidant function to cells by different mechanisms and decreases ROS and LPO (Joshi et al., 2004; Formigari et al., 2007; Matovic et al., 2011). The most compelling reason for the protective effects of Zn against Cd toxicity is that Zn induces the production of the metal-binding protein, MT (Bonda et al., 2004; Amara et al., 2008). Zn supplementation could compete with binding sites for Cd uptake. The amelioration effect of Zn may be in part, mediated by induction and synthesis of MT and by providing thio (-SH) groups which can scavenge hydroxyl radicals and singlet oxygen. The protective role of Zn may be due to its ability to maintain normal GSH level and CAT activity. It appears that Zn has a direct antioxidant activity through scavenging of free radicals and indirect effects via prevention of the depletion of blood GSH level and hepatic CAT resulting from Cd treatment or by interfering with Cd transport into the cell rather than its activity to stimulate MT synthesis.
Table 2.
Alterations in the activity levels of selected oxidative stress enzymes in the liver of Cd-treated rats after Zn and/or Fe supplementation
Fe is an essential trace element for many biological processes and it is important to maintain Fe concentration within its narrow normal range in the body (Turgut et al., 2007) and also a vital component of several enzymes and proteins. The nutritional Fe status has a clear effect on Cd absorption (Akesson et al., 2002: Ryu et al., 2004). Fe and Cd most probably compete at the binding sites of the Fe intestinal transfer system (Bhattacharya et al., 2000; Tallkvist et al., 2001; Park et al., 2002). In the present study Cd concentrations were significantly depleted in the test tissues with Fe supplementation suggesting that Fe can reduce the Cd-induced alterations in the oxidative stress enzymes (Obaiah et al., 2009; Usha Rani et al., 2010). In one study, it was observed that the intestinal absorption of dietary Cd in women decreased when the body Fe stores and fiber intake were increased (Berguland et al., 1994). In rats, dietary Fe supplements have shown to decrease the Cd retention after oral Cd exposure (Groten et al., 1992; Schumann et al., 1996).
Zn and Fe most effectively protects against intestinal absorption of Cd from Cd-MT, thereby decreasing Cd uptake by 80% than the other essential elements (Piasek et al., 2004). Further Casalino et al., (1997) reported that Fe supplementation corrects the anemia caused by Cd exposure in rats. It has also been suggested that higher gastrointestinal absorption of Cd is due to lower body Fe stores as measured by the concentrations of serum ferritins (Vahter et al., 1996). The overall discussion suggests that Fe may reduce the Cd body burden in the organisms by eliminating Cd from the tissues. Moreover, in the present study the Cd-induced alterations in the oxidative stress/antioxidant enzymes were recovered with specific supplements (as mentioned in Materials and Methods) of Zn and Fe individually and in combination. Our work is significant and has got applied value because the intake of trace element supplements like Zn and Fe aids in therapeutic indices of Cd and other heavy metal toxicity.
Financial support and sponsorship
Nil.
Conflicts of interest
There are no conflicts of interest
Acknowledgments
The authors are highly thankful to the UGC, New Delhi for the financial support rendered with the award of Major Research Project (No. F. 34 – 476/2008(SR)) to Dr. A. Usha Rani, Professor, Department of Zoology, Sri Venkateswara University, Tirupati.
SUGGESTED READINGS
- 1.Aebi H. Catalase. In: Packer L, editor. Methods in enzymology. 2nd edition. Vol. 105. Orlando: Academic Press; 1984. pp. 121–6. [Google Scholar]
- 2.Akesson A, Berglund M, Schutz A, Bjellerup P, Bremme K, Vahter M. Cadmium exposure in pregnancy and lactation in relation to iron status. Am J Public Health. 2002;92:284–7. doi: 10.2105/ajph.92.2.284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Salinska A, Włostowski T, Zambrzycka E. Effect of dietary cadmium and/or lead on histopathological changes in the kidneys and liver of bank voles Myodes glareolus kept in different group densities. Ecotoxicology. 2012;21:2235–43. doi: 10.1007/s10646-012-0979-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Asagba SO, Eriyamremu GE. Oral cadmium exposure and levels of superoxide dismutase, catalase, lipid peroxidation and ATPases in the eye. Res J Environ Toxicol. 2007;1:204–9. [Google Scholar]
- 5.Berguland M, Akesson A, Nermell B, Vahter M. Intestinal absorption of dietary cadmium in women depends on body iron stores and fiber intake. Environ Health Perspect. 1994;102:1058–66. doi: 10.1289/ehp.941021058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bertin G, Averbeck D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review) Biochimie. 2006;88:1549–59. doi: 10.1016/j.biochi.2006.10.001. [DOI] [PubMed] [Google Scholar]
- 7.Bhattacharya MH, Wilson AK, Rajan SS, Jonath M. Biochemical pathways in cadmium toxicity. In: Zalups RK, Koropatnick J, editors. Molecular Biology and Toxicology of Metals. London/Newyork: Taylor and Francis; 2000. pp. 34–74. [Google Scholar]
- 8.Bonda E, Wlostowki T, Krasowska T. Testicular toxicity induced by dietary cadmium is associated with decreased testicular zinc and increased hepatic and renal metallothionein and zinc in the bank vole (Clethrionomysglareolus) Biometals. 2004;17:615–24. doi: 10.1007/s10534-004-1226-8. [DOI] [PubMed] [Google Scholar]
- 9.Brewer JG. Iron and copper toxicity in diseases of ageing, particularly atherosclerosis and Alzheimer's disease. Exp Biol Med (Maywood) 2007;232:323–35. [PubMed] [Google Scholar]
- 10.Cairo G, Recalcati S. Iron-regulatory proteins: Molecular biology and pathophysiological implications. Expert Rev Mol Med. 2007;9:1–13. doi: 10.1017/S1462399407000531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Casalino E, Sblano S, Landriscina C. Enzyme activity alteration by cadmium administration to rats: The possibility of iron involvement in lipid peroxidation. Arch Biochem Biophys. 1997;346:171–9. doi: 10.1006/abbi.1997.0197. [DOI] [PubMed] [Google Scholar]
- 12.Cuypers A, Plusquin M, Remans T, Jozefczak M, Keunen A, Gielen H, et al. Cadmium stress: An oxidative challenge. Biometals. 2010;23:927–40. doi: 10.1007/s10534-010-9329-x. [DOI] [PubMed] [Google Scholar]
- 13.Dzugan M, Lis M, Droba M, Niedziolka JW. Effect of cadmium injected in vivo on hatching results and the activity of plasma hydrolytic enzymes in newly hatched chicks. Acta Vet Hung. 2011;59:337–47. doi: 10.1556/AVet.2011.020. [DOI] [PubMed] [Google Scholar]
- 14.Dzugan M, Lis M, Droba M, Niedziolka JW. Protective effect of zinc on cadmium embryotoxicity and antioxidant status of blood plasma in newly hatched chicks. J Environ Sci Health Part ATox Hazard Subst Environ Eng. 2012;47:1288–93. doi: 10.1080/10934529.2012.672133. [DOI] [PubMed] [Google Scholar]
- 15.El-Missiry MA, Shalaby F. Role of β-carotene in ameliorating the cadmium-induced oxidative stress in rat brain and testis. J Biochem Mol Toxicol. 2000;14:238–43. doi: 10.1002/1099-0461(2000)14:5<238::AID-JBT2>3.0.CO;2-X. [DOI] [PubMed] [Google Scholar]
- 16.Guirlet E, Das K. Cadmium toxicokinetics and bioaccumulation in turtles: Trophic exposure of Trachemys scripta elegans. Ecotoxicology. 2012;21:18–26. doi: 10.1007/s10646-011-0758-2. [DOI] [PubMed] [Google Scholar]
- 17.Flohe L, Gunzler WA. Assays of glutathione peroxidase. Methods Enzymol. 1984;105:114–21. doi: 10.1016/s0076-6879(84)05015-1. [DOI] [PubMed] [Google Scholar]
- 18.Flora SJ, Mittal M, Mehta A. Heavy metal induced oxidative stress and its possible reversal by chelation therapy. Indian J Med Res. 2008;128:501–23. [PubMed] [Google Scholar]
- 19.Formigari A, Irato P, Santon A. Zinc, antioxidant systems and metallothionein in metal mediated-apoptosis: Biochemical and cytochemical aspects. Comp Biochem Physiol C Toxicol Pharmacol. 2007;146:443–59. doi: 10.1016/j.cbpc.2007.07.010. [DOI] [PubMed] [Google Scholar]
- 20.Habig WH, Pabst MJ, Jacoby WB. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J Biol Chem. 1974;249:7130–9. [PubMed] [Google Scholar]
- 21.Hakan AH, Canan C, Biltan E. In vivo interaction between cadmium and essential trace elements copper and zinc in rats. Turk J Med Sci. 2001;31:127–9. [Google Scholar]
- 22.Han XY, Xu ZR, Wang YZ, Huang QC. Effect of cadmium on lipid peroxidation and activities of antioxidant enzymes in growing pigs. Biol Trace Elem Res. 2006;110:251–63. doi: 10.1385/BTER:110:3:251. [DOI] [PubMed] [Google Scholar]
- 23.Hartwig A. Mechanism in cadmium-induced carcinogenicity: Recent insights. Biometals. 2010;23:951–60. doi: 10.1007/s10534-010-9330-4. [DOI] [PubMed] [Google Scholar]
- 24.Honda A, Komuro H, Shimada A, Hasegawa T, Seko Y, Shimada A, et al. Attenuation of cadmium-induced testicular injury in metallothionein - III null mice. Life Sci. 2010;87:545–50. doi: 10.1016/j.lfs.2010.09.009. [DOI] [PubMed] [Google Scholar]
- 25.Jarup L, Akesson A. Current status of cadmium as an environmental health problem. Toxicol Appl Pharmcol. 2009;238:201–8. doi: 10.1016/j.taap.2009.04.020. [DOI] [PubMed] [Google Scholar]
- 26.Johri N, Jacquillet G, Unwin R. Heavy metal poisoning: The effects of cadmium on kidney. Biometals. 2010;23:783–92. doi: 10.1007/s10534-010-9328-y. [DOI] [PubMed] [Google Scholar]
- 27.Joshi S, Hasan SK, Chandra R, Husain MM, Srivastava RC. Scavenging action of zinc and green tea polyphenol on cisplatin and nickel induced nitric oxide generation and lipid peroxidation in rats. Biomed Environ Sci. 2004;17:402–9. [PubMed] [Google Scholar]
- 28.Jovanovic MJ, Nikolic RS, Kocic GM, Krstic SN, Krsmanovic MM. Glutathione protectsliver and kidney tissue from cadmium - and lead - provoked lipid peroxidation. J Serb Chem Soc. 2012;77:1–7. [Google Scholar]
- 29.Kim SG, Dai W, Xu Z, Li G. Effects of montmorillonite on alleviating diertary Cd-induced oxidative damage in carp (Carassiusauratus) Biol Trace Elem Res. 2011;141:200–6. doi: 10.1007/s12011-010-8735-9. [DOI] [PubMed] [Google Scholar]
- 30.Kwong RW, Andres JA, Niyogi S. Effects of dietary cadmium exposure on tissue-specific cadmium accumulation, iron status and expression of iron-handling and stress - inducible genes in rainbow trout: Influence of elevated dietary iron. Aquat Toxicol. 2011;102:1–9. doi: 10.1016/j.aquatox.2010.12.010. [DOI] [PubMed] [Google Scholar]
- 31.Olalekan Lawal A, Lawal AF, Ologundudu A, Adeniran OY, Omonkhua A, Obi F. Antioxidant effects of heated garlic juice on cadmium-induced liver damage in rats as compared to ascorbic acid. J Toxicol Sci. 2011;36:549–57. doi: 10.2131/jts.36.549. [DOI] [PubMed] [Google Scholar]
- 32.Liu J, Yeo HC, Overvik-Douki E, Hagen T, Doniger SJ, Chu DW, et al. Chronically and acutely exercised rats: Biomarkers of oxidative stress and endogenous antioxidants. J Appl Physiol (1985) 2000;89:21–8. doi: 10.1152/jappl.2000.89.1.21. [DOI] [PubMed] [Google Scholar]
- 33.Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with Folin phenol reagent. J Biol Chem. 1951;193:265–75. [PubMed] [Google Scholar]
- 34.Lucia M, André JM, Gonzalez P, Baudrimont M, Bernadet MD, Gontier K, et al. Effect of dietary cadmium on lipid metabolism and storage of aquatic bird Cairinamoschata. Ecotoxicology. 2010;19:163–70. doi: 10.1007/s10646-009-0401-7. [DOI] [PubMed] [Google Scholar]
- 35.Piasek M, Blanusa M, Kostial K, Laskey JW. Low iron diet and parenteral cadmium exposure in pregnant rats: The effects on trace elements and fetal viability. Biometals. 2004;17:1–14. doi: 10.1023/a:1024426931676. [DOI] [PubMed] [Google Scholar]
- 36.Martinez R, Brassard P, Mwanjewe J, Grover AK. Iron promotes cadmium binding to citrate. Mol Cell Biochem. 2001;225:93–6. doi: 10.1023/a:1012257802703. [DOI] [PubMed] [Google Scholar]
- 37.Matovic V, Buha A, Bulat Z, Dukic-Cosic D. Cadmium toxicity revisited: Focus on oxidative stress induction and interaction with zinc and magnesium. Arh Hig Rada Toksikol. 2011;62:65–76. doi: 10.2478/10004-1254-62-2011-2075. [DOI] [PubMed] [Google Scholar]
- 38.Misra HP, Fridovich I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J Biol Chem. 1972;247:3170–5. [PubMed] [Google Scholar]
- 39.Moulis JM, Thevenod F. New perspectives in cadmium toxicity: An introduction. Biometals. 2010;23:763–8. doi: 10.1007/s10534-010-9365-6. [DOI] [PubMed] [Google Scholar]
- 40.Muckenthalerm MU, Galy B, Hentze MW. Systemic iron homeostasis and the iron-responsive element/iron-regulatory protein (IRE/IRP) regulatory network. Annu Rev Nutr. 2008;28:197–213. doi: 10.1146/annurev.nutr.28.061807.155521. [DOI] [PubMed] [Google Scholar]
- 41.Munga A, Xhaxhiu D, Laci D, Dova I. Cadmium accumulation in the main organs during a chronic exposure. Natura Montegrina Podgorica. 2010;9:719–30. [Google Scholar]
- 42.Nad P, Massanyi P, Skalicka M, Korenekova B, Cigankova V, Almasiova V. The effect of cadmium in combination with zinc and selenium on ovarian structure in Japanese quails. J Environ Sci Health A Tox Hazard Subst Environ Eng. 2007;42:2017–22. doi: 10.1080/10934520701629716. [DOI] [PubMed] [Google Scholar]
- 43.Nakamura Y, Ohba K, Suzuk K, Ohta H. Health effects of low-level cadmium intake and the role of metallothionein on cadmium transport from mother rats to fetus. J Toxicol Sci. 2012;37:149–56. doi: 10.2131/jts.37.149. [DOI] [PubMed] [Google Scholar]
- 44.Nordberg GF, Bigawam K, Nordberg M, Friedmann JM. Cadmium. In: Nordberg GF, Fowler BA, Nordberg M, Friberg L, editors. Hand Book on the Toxicology of Metals. 4th edition. Vol. 1. Amsterdam: Elsevier Publications; 2007. pp. 445–86. [Google Scholar]
- 45.Obaiah J, Bhavani G, Usha Rani A. Effect of calcium on cadmium induced bioaccumulation in selected tissues of fresh water teleost, Oreochromis mossambicus (Tilapia) Indian Streams Res J. 2013;3:1–4. [Google Scholar]
- 46.Obaiah J, Usha Rani A, Kumar Babu D. Trace element supplementation against cadmium induced toxicity in male albino rats: Protective role of zinc and iron. Indian J Comp Anim Physiol. 2009;27:99–104. [Google Scholar]
- 47.Obaiah J, Usha Rani A. Calcium impact on cadmium induced alterations in selected oxidative stress enzymes in the fresh water teleost, Oreochromis mossambicus (Tilapia) Gold Res Thoughts. 2013;3:1–5. [Google Scholar]
- 48.Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95:351–8. doi: 10.1016/0003-2697(79)90738-3. [DOI] [PubMed] [Google Scholar]
- 49.Park JD, Cherrington NJ, Klaassen CD. Intestinal absorption of cadmium is associated with divalent metal transporter 1 in rats. Toxicol Sci. 2002;68:288–94. doi: 10.1093/toxsci/68.2.288. [DOI] [PubMed] [Google Scholar]
- 50.Qureshi MI, D’Amici GM, Fagioni M, Rinalducci S, Zolla L. Iron stabilizes thylakoid protein-pigment complexes in Indian mustard during Cd-phytoremidiation as revealed BN-SDS-PAGE and ESI-MS/MS. J Plant Physiol. 2010;167:761–70. doi: 10.1016/j.jplph.2010.01.017. [DOI] [PubMed] [Google Scholar]
- 51.Ryu DY, Lee SJ, Park DW, Choi BS, Klassen CD, Park JD. Dietary iron regulates intestinal cadmium absorption through iron transporters in rats. ToxicolLett. 2004;152:19–25. doi: 10.1016/j.toxlet.2004.03.015. [DOI] [PubMed] [Google Scholar]
- 52.Amara S, Abdelmelek H, Garrel C, Guiraud P, Douki T, Ravanat JL, et al. Preventive effect of zinc against cadmium-induced oxidative stress in the rat testis. J Reprod Dev. 2008;54:129–34. doi: 10.1262/jrd.18110. [DOI] [PubMed] [Google Scholar]
- 53.Saric MM, Blanusa M, Piasek M, Varnai VM, Juresa D, Kostial K. Effect of dietary cadmium on cadmium absorption and retention in suckling rats. Biometals. 2002;15:175–82. doi: 10.1023/a:1015212929481. [DOI] [PubMed] [Google Scholar]
- 54.Schumann K, Friebel P, Schmolke G, Elsenhans B. State of iron repletion and cadmiumtissue accumulation as a function of growth in young rats after oral cadmium exposure. Arch Environ Contam Toxicol. 1996;31:483–7. [PubMed] [Google Scholar]
- 55.Sladan ZP, Branka IO, Andras SS, Radoslav VZ, Ratko MR, Mihajlo BS, et al. A possible protective role of coenzyme Q10 on antioxidant defense system in the heart of rats treated with cadmium. Jugoslov Med Biochem. 2005;24:121–7. [Google Scholar]
- 56.Stohs SJ, Bagchi D, Hassoun E, Bagchi M. Oxidative mechanisms in the toxicity of chromium and cadmium ions. J Environ PatholToxicolOncol. 2001;20:77–88. [PubMed] [Google Scholar]
- 57.Włostowski T, Dmowski K, Bonda-Ostaszewska E. Cadmium accumulation, metallothionein and glutathione levels, and histopathological changes in the kidneys and liver of magpie (Pica pica) from a zinc smelter area. Ecotoxicology. 2010;19:1066–73. doi: 10.1007/s10646-010-0488-x. [DOI] [PubMed] [Google Scholar]
- 58.Tallkvist J, Bowlus CL, Lonnerdal B. DMT1 gene expression and cadmium absorption in human absorptive enterocytes. Toxicol Lett. 2001;122:171–7. doi: 10.1016/s0378-4274(01)00363-0. [DOI] [PubMed] [Google Scholar]
- 59.Tatrai E, Kovacikova Z, Hudak A, Adamis Z, Ungvary G. Comparative in vitro toxicity of cadmium and lead on redox cycling in type II puemocytes. J Appl Toxicol. 2001;21:479–83. doi: 10.1002/jat.784. [DOI] [PubMed] [Google Scholar]
- 60.Thevenod F. Cadmium and cellular signaling cascades: To be or not to be? Toxicol Appl Pharmacol. 2009;238:221–39. doi: 10.1016/j.taap.2009.01.013. [DOI] [PubMed] [Google Scholar]
- 61.Nawrot TS, Van Hecke E, Thijs L, Richert T, Kuznestsova T, Jin Y, et al. Cadmium- related mortality and long-term secular trends in the cadmium body burden of an environmentally exposed population. Environ Health Perspect. 2008;116:1620–8. doi: 10.1289/ehp.11667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Turgut S, Polat A, Inan M, Turgut G, Emmungil G, Bican M, et al. Interaction between anemia and blood levels of iron, zinc, copper, cadmium and lead in children. Indian J Pediatr. 2007;74:827–30. doi: 10.1007/s12098-007-0147-2. [DOI] [PubMed] [Google Scholar]
- 63.Usha Rani A, Obaiah J, Kumar Babu D. Impact of four essential trace elements in combating cadmium bioaccumulation in liver and kidney of rat. J Nature Conserv. 2010;22:29–36. [Google Scholar]
- 64.Vahter M, Berglund M, Nermell B, Akesson A. Bioavailability of cadmium from shellfish and mixed diet in women. Toxicol Appl Pharmacol. 1996;136:332–41. doi: 10.1006/taap.1996.0040. [DOI] [PubMed] [Google Scholar]
- 65.Wang Y, Fang J, Leonard SS, Rao KM. Cadmium inhibits the electron transfer chain and induces reactive oxygen species. Free Radic Biol Med. 2004;36:1434–43. doi: 10.1016/j.freeradbiomed.2004.03.010. [DOI] [PubMed] [Google Scholar]
- 66.Matek SM, Blanusa M, Piasek M, Varnai VM, Juresa D, Kostial K. Effect of dietary calcium on cadmium absorption and retention in suckling rats. Biometals. 2002;15:175–82. doi: 10.1023/a:1015212929481. [DOI] [PubMed] [Google Scholar]
- 67.Beytut E, Aksakal M. The effect of long-term supplemental dietary cadmium on lipid peroxidation and the antioxidant system in the liver and kidneys of rabbits. Turk J Vet Anim Sci. 2002;26:1055–60. [Google Scholar]
- 68.Jurczuk M, Brzoska MM, Moniuszko-Jakoniuk J, Galazyn-Sidorczuk M, Eulikowska-Karpinska E. Antioxidant enzymes activity and lipid peroxidation in liver and kidney of rats exposed to cadmium and ethanol. Food Chem Toxicol. 2004;42:429–38. doi: 10.1016/j.fct.2003.10.005. [DOI] [PubMed] [Google Scholar]
- 69.Nagaraj M, Sunitha S, Varalakshmi P. Effect of lupeol, a pentacyclic triterpene, on lipid peroxidation and antioxidant status in rat kidney after chronic cadmium exposure. J Applied Toxicol. 2000;20:413–17. doi: 10.1002/1099-1263(200009/10)20:5<413::AID-JAT706>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 70.Waisberg M, Joseph P, Hale B, Beyersmann D. Molecular and cellular mechanisms of cadmium carcinogenesis: A review. Toxicology. 2003;192:95–117. doi: 10.1016/s0300-483x(03)00305-6. [DOI] [PubMed] [Google Scholar]
- 71.Ognjanovic BI, Markovic SD, Pavlovic SZ, Zikic RV, Stajn AS, Saicic ZS. Effect of chronic cadmium exposure on antioxidant defense system in some tissues of rats: Protective effect of Selenium. Physiol Res. 2008;57:403–11. doi: 10.33549/physiolres.931197. [DOI] [PubMed] [Google Scholar]
- 72.Messaoudi I, EI Heni J, Hammouda F. Protective effects of selenium, zinc, or their combination on cadmium-induced oxidative stress in rat kidney. Boil Trace Elem Res. 2009;130:152–161. doi: 10.1007/s12011-009-8324-y. [DOI] [PubMed] [Google Scholar]
- 73.Mahendran P, Shyamala Devi CS. The modulating effect of Garciniacambogia extract on ethanol induced peroxidative damage in rats. Ind Pharmacol. 2001;33:87–91. [Google Scholar]
- 74.Groten JP, Luten JB, Van Bladeren PJ. Dietary iron lowers the intestinal uptake of cadmium metallothionein in rats. Eur J Pharmacol. 1992;228:23–28. doi: 10.1016/0926-6917(92)90007-y. [DOI] [PubMed] [Google Scholar]
- 75.Ashraf MAM, Kamar MS. Acute effect of cadmium treatment on the kidney of rats: Biochemical and ultra structural studies. Pakistan Journal of Biological Sciences. 2007;10:3497–506. doi: 10.3923/pjbs.2007.3497.3506. [DOI] [PubMed] [Google Scholar]
- 76.Deepthi S, Ramlingam BS, Datta M, Flora SJS. Protective effect of selenium, calcium and magnesium against arsenic induced oxidative stress in male rats. Arh Hig Rada Tosikol. 2010;61:153–9. doi: 10.2478/10004-1254-61-2010-1993. [DOI] [PubMed] [Google Scholar]
- 77.Peraza MA, Ayala-Fierro F, Barber DS, Casarez E, Rael LT. Effects of micronutrients o n metal toxicity. Environmental Health Perspectives Supplements. 1998;106:203–16. doi: 10.1289/ehp.98106s1203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Zhang J, Shen H, Wang X, Wu J, Xue Y. Effects of chronic exposure of 2, 4-dichlorophenol on the antioxidant system in liver of fresh water fish Carassius auratus. Chemosphere. 2004;55:167–74. doi: 10.1016/j.chemosphere.2003.10.048. [DOI] [PubMed] [Google Scholar]