

Abstract
Human coronaviruses (HCoVs) are large RNA viruses that infect the human respiratory tract. The emergence of both Severe Acute Respiratory Syndrome and Middle East Respiratory syndrome CoVs as well as the yearly circulation of four common CoVs highlights the importance of elucidating the different mechanisms employed by these viruses to evade the host immune response, determine their tropism and identify antiviral compounds. Various animal models have been established to investigate HCoV infection, including mice and non-human primates. To establish a link between the research conducted in animal models and humans, an organotypic human airway culture system, that recapitulates the human airway epithelium, has been developed. Currently, different cell culture systems are available to recapitulate the human airways, including the Air-Liquid Interface (ALI) human airway epithelium (HAE) model. Tracheobronchial HAE cultures recapitulate the primary entry point of human respiratory viruses while the alveolar model allows for elucidation of mechanisms involved in viral infection and pathogenesis in the alveoli. These organotypic human airway cultures represent a universal platform to study respiratory virus-host interaction by offering more detailed insights compared to cell lines. Additionally, the epidemic potential of this virus family highlights the need for both vaccines and antivirals. No commercial vaccine is available but various effective antivirals have been identified, some with potential for human treatment. These morphological airway cultures are also well suited for the identification of antivirals, evaluation of compound toxicity and viral inhibition.
Keywords: Human coronavirus, Airway epithelium, ALI, Antiviral compound, Cell tropism
Background
Respiratory diseases caused by human coronavirus infection are of both medical and socio-economic importance. Currently, they are studied in various model systems, ranging from cell lines to animal models. Originally, the importance of HCoVs in the burden of human disease was underestimated and as a result, no general therapy exists to treat coronavirus induced disease in humans. Furthermore, no commercial vaccine is available leaving the human population vulnerable to emerging coronavirus infections. Both the Severe Acute Respiratory Syndrome and Middle East Respiratory Syndrome coronaviruses have recently crossed the species barrier and entered the human population to cause severe disease. In this review, we summarize the current knowledge on human coronavirus infection emphasizing the usefulness of organotypic human airway cultures as a model system.
Coronaviruses
Coronaviruses (CoVs), a subfamily of the Coronaviridae family, are positive strand RNA viruses with the largest genome of all known RNA viruses (≥27 Kb). The genomic RNA is capped, polyadenylated and associated with nucleocapsid proteins within an enveloped virion. The envelope is covered by the characteristic surface glycoprotein that gives the virus particles their characteristic crown-like (latin: corona) appearance [1].
All CoVs share a common genome organization where the replicase gene encompasses the 5′-two thirds of the genome and is comprised of two overlapping open reading frames (ORFs), ORF1a and ORF1b that encode for up to 16 non-structural proteins. The structural gene region, which covers the 3′-third of the genome, encodes the canonical set of structural protein genes in the order 5′ - spike (S) - envelope (E) - membrane (M) and nucleocapsid (N) – 3′. The structural gene region also harbors several ORFs that are interspersed along the structural protein coding genes. The number and location of these accessory ORFs vary between the CoV species [2, 3].
In animals, CoV infections are mainly associated with respiratory and enteric disease and can have large economical impact on the veterinary industry, e.g. Porcine Epidemic Diarrhea Virus (PEDV) causes gastrointestinal disease in pigs [4], Infectious Bronchitis Virus (IBV) causes severe kidney and respiratory disease in chicken [5] and Bovine Coronavirus (BCoV) causes both respiratory disease and diarrhea in cattle [6]. Additionally, CoV infections can have other disease manifestations, such as central nervous system (CNS) involvement, hepatitis and peritonitis [7–10].
In humans, CoV infections are mainly associated with respiratory diseases that are considered to have a large impact on the economy due to reduced productivity of the working population. Currently, 6 coronaviruses that cause disease in humans have been discovered. Four of those are commonly circulating and two have caused epidemics of severe acute respiratory disease.
Human coronaviruses
The first human coronavirus (HCoV), B814, was described in 1965. In the following years, over 30 additional strains were characterized. Ten of those strains could only be isolated from primary embryonic tracheal organ culture. Others were readily isolated from monolayer cultures and were antigenically related to the prototype strain HCoV-229E. HCoV-OC43, for organ culture 43, was isolated and found to be distinct from the 229E prototype strain [11, 12]. In the subsequent decades, research on HCoVs would center on these two distinct viruses.
However, in 2002, an unknown respiratory illness, termed Severe Acute Respiratory Syndrome (SARS), surfaced in Asia. Research determined it to be caused by a novel coronavirus [13, 14]. At the end of the epidemic, this virus had infected over 8000 people, most in China, and caused 774 deaths [15].
Following the discovery of this virus, two additional CoVs causing human disease were identified. HCoV-NL63 was isolated in the Netherlands in 2004 from an infant with bronchiolitis [16] and HCoV-HKU1 in 2005 from a patient with pneumonia in Hong Kong [17]. In 2012, another respiratory HCoV, Middle East Respiratory (MERS)–CoV, was isolated from a patient with pneumonia in Saudi-Arabia [18]. Unlike SARS-CoV, this virus is still intermittently present in the human population and most recently caused a large outbreak in South-Korea [19]. To date, there have been over 1600 cases and almost 600 deaths related to MERS-CoV infection [20].
Commonly circulating coronaviruses
Out of the 6 known human coronaviruses, HCoV-229E, HCoV-OC43, HCoV-NL63 and HCoV-HKU1 are commonly circulating in the human population and usually cause general respiratory illness and cold symptoms in healthy individuals [21–23]. Like influenza, these viruses are capable of causing more severe disease in the immunocompromised and the elderly [24]. They infect the human airway from the luminal side and progeny viruses are released from the same side facilitating spread through coughing and sneezing [25, 26]. These coronaviruses are responsible for approximately 5–10% of all upper and lower respiratory tract infections [27–29] but the interactions between them and their natural host cells are poorly understood. Currently, it is hypothesized that most of the human coronaviruses may have originated from bats [30, 31]. For example, HCoV-229E is believed to originate from African hipposiderid bats possibly using camelids as intermediate hosts [32].
Emerging coronaviruses
In the last 15 years, two coronaviruses have crossed the species barrier and caused severe and fatal disease in humans. SARS-CoV surfaced in 2002 and MERS-CoV in 2012 [13, 14, 18]. As opposed to the commonly circulating viruses, which generally only cause mild respiratory symptoms, these viruses presented with higher case fatality ratios, around 10 and 20–50% respectively [33, 34].
Currently, there is abundant phylogenetic evidence for the bat origin of SARS-CoV, based on sequences of SARS-like viruses found among bats in the recent years [35–37]. The initial transmissions of SARS-CoV from animals to humans were traced back to the live animal wet markets and it was hypothesized that the virus made its way into the human population using the civet cat as an intermediate host. However, successful isolation of SARS-like viruses from bats [38] and the fact that a contemporary bat SARS-like virus can infect human airway cultures [39] suggest that an intermediate host between humans and bat might not have been needed for the transmission of SARS-CoV.
The evolutionary origin of MERS-CoV is less clear but it has been speculated to be bats as well. Characterization of an African bat virus closely related to MERS-CoV shows that both the human and camel strains belong to the same viral species and phylogenetic analysis suggests that MERS-CoV infection in camels predates that in humans, suggesting that camels infect humans and not the other way around. Furthermore, the bat virus roots the phylogenetic tree providing further evidence for the bat origin of MERS-CoV [40]. Additionally, human-to-human transmission, although not robust, seems to happen simultaneously as camel-to-human transmission. Therefore, any further adaptation of MERS-CoV to the human host must be monitored carefully and intermediate hosts identified [41].
Many bat coronaviruses have been identified in the recent years further highlighting the zoonotic potential of this family of viruses [30]. Given the documented history of coronaviruses overcoming the species barrier and causing severe disease in humans, it is important to investigate the zoonotic potential of close evolutionary relatives of common HCoVs in a culture model that recapitulates the aspects of the human airway, e.g. morphology and receptor distribution. It’s important to study the mechanisms of pathogenesis and the evolution of zoonotic viruses in detail in order to identify molecular determinants that affect either transmission or pathogenesis. It’s also important to elucidate whether coronaviruses currently circulating in animals are a potential danger to the human population.
Human coronavirus receptors and cell tropism
All of the known cellular receptors of HCoVs belong to the same protein family, the membrane ectopeptidases. Interestingly, the catalytic activity of these peptidases is not required for viral entry but rather the co-expression of other host peptidases activates the HCoV spike proteins [42, 43]. It has been established that the human transmembrane serine proteases TMPRSSII and HAT cleave and activate the HCoV-229E, SARS- and MERS-CoV spike proteins during viral entry [44, 45].
Out of the four commonly circulating coronaviruses, HCoV-229E is the only one that infects non-ciliated cells using the human Aminopeptidase N (hAPN) as its receptor [46]. This peptidase is predominantly expressed on non-ciliated cells in the human bronchus [47]. SARS-CoV and HCoV-NL63 both utilize the Angiotensin Converting Enzyme 2 (ACE2) for cellular binding [48, 49]. ACE2 is expressed on ciliated bronchial cells along with endothelial cells and both type I and II alveolar cells [50]. MERS-CoV was found to use a different receptor than SARS-CoV, namely the dipeptyl-peptidase 4 (DPP4) [51]. DPP4 is widely expressed in endothelial cells and various epithelial tissues in the human body [52]. In ex vivo human lung organ cultures, different tropism of SARS- and MERS-CoVs was observed. MERS-CoV can actively replicate in both bronchial and alveolar tissue while SARS-CoV primarily replicates in alveolar tissue [53]. The wide cellular tropism of MERS-CoV might contribute to its associated disease severity and high mortality rate whereas the alveolar replication of SARS-CoV would explain why it generally presents with pneumonia.
The cellular surface receptors for HCoV-OC43 and HCoV-HKU1 are currently unknown but receptor determinants for these two viruses have been identified as N-acetyl-9-O-acetylneuraminic acid and O-Acetylated Sialic acid, respectively [54, 55].
All of these viruses can be successfully cultured and investigated in HAE cultures [56, 57]. The discovery of HCoVs, their receptor usage, cell tropism and receptor binding domain (RBD) is summarized in Table 1.
Table 1.
Human coronavirus overview
| Name | Discovery | Protein Receptor | Tropism | Receptor Binding Domain (RBD) | References |
|---|---|---|---|---|---|
| HCoV-229E | 1966 | Aminopeptidase N (hAPN) | Non-ciliated cells | S407-547 | [46, 56, 57, 103, 104] |
| HCoV-OC43 | 1967 | Unknowna | Ciliated cells | Unknown | [56, 105] |
| SARS-CoV | 2003 | Angiotensin Converting Enzyme 2 (ACE2) | Ciliated cells | S303-537 | [13, 14, 48, 57, 106] |
| HCoV-NL63 | 2004 | Angiotensin Converting Enzyme 2 (ACE2) | Ciliated cells | S476 -616 | [16, 49, 56, 107] |
| HCoV-HKU1 | 2005 | Unknownb | Ciliated cells | Unknown | [17, 56] |
| MERS-CoV | 2012 | Dipeptyl-peptidase 4 (DPP4) | Non-ciliated cells | S358-588 | [18, 34, 51, 108] |
aReceptor determinant identified as N-acetyl-9-O-acetylneuraminic acid
bReceptor determinant identified as O-Acetylated Sialic acid
Furthermore, established reverse genetic systems for HCoV-229E [58], HCoV-OC43 [59] and HCoV-NL63 [60] allow for controlled virus mutation and fluorescent transgene insertion to better understand the interaction of these viruses with their pulmonary host cells.
Animal models for human coronaviruses
Traditionally, respiratory viruses are studied in animal models, usually mice and ferrets [48, 61]. However, it is not always possible to correctly recapitulate human infection and disease in animal models. The establishment of transgenic animal models for human disease is attainable when either the virus receptor has been identified, which is not the case for all HCoVs, or when viruses can be adapted to a different host. An adapted human virus may not share the same properties as the original human virus. SARS-CoV was found to replicate naturally in various strains of inbred mice but to enhance clinical signs of disease the hACE2 was introduced into these mice. This resulted in murine models with varying degree of human disease similarity. Since SARS-CoV already replicated in mouse cells, adapting it to the murine host was quite successful. This resulted in three mouse adapted strains that caused disease in mice similar to severe SARS-CoV cases in humans [62].
In an effort to establish a mouse model for HCoV-229E infection transgenic hAPN mice were created. However, the insertion of the hAPN into mouse cells is not enough to establish robust HCoV-229E infection in vivo. Nevertheless, cells isolated from these transgenic animals could be infected in vitro [63, 64].
The emergence of both SARS- and MERS-CoVs emphasized the importance of establishing animal models for human coronaviruses. Currently, a few animal models for MERS-CoV have been established. Mice carry their own variant of the viral receptor DDP4 that differs from the human in regions important for MERS-CoV spike interaction and by replacing this receptor with the human one, MERS-CoV can infect mouse cells but the method of hDPP4 insertion has an effect on the degree of pathogenesis observed in these mice [65, 66]. Various non-human primates (NHPs) can be naturally infected with both SARS- and MERS-CoVs. However, disease presentation and pathogenesis differs between the different subspecies and NHP models are expensive, although ideal to study human infection due to their genetic similarity [62].
To establish a link between the research conducted in animal models and humans, an organotypic airway culture system resembling the human airway epithelium has been developed. This model is a universal platform to study human respiratory viruses [67–70]. They have been used successfully for infection studies with all known human coronaviruses [56, 57]. Furthermore, the cultures can be inoculated with a low infectious dosage to mimic natural infection in the human airway. Whereas, animal models often require both high dosage and artificial inoculation routes.
Human airway epithelial cell cultures
Organotypic cell cultures are becoming increasingly common. Different cell culture models exist to depict different epithelial tissues [71]. These cultures closely resemble their tissue of origin and contain various different cell types with distinctive roles in the polarized tissue. Currently, various organotypic cell culture models exist to represent the different areas of the human airways. The human lungs span a long anatomical distance and carry out different functions depending on anatomical location [72, 73]. The structure of the epithelium also differs the further you descend into the airways. Tracheal and bronchial epithelium is columnar and pseudostratified, with every cell in contact with the basement membrane, while the epithelium in the alveoli is comprised of a single cell layer to facilitate air-exchange [74].
Tracheobronchial cells are one of the first targets of human respiratory viruses and can be cultured in air-liquid interface (ALI) where the apical side of the cell layer is exposed to air while the basolateral side is submerged in medium. Tracheobronchial cells cultured in that way form a pseudostratified epithelial layer that both morphologically and functionally resembles the human upper conducting airway (Fig. 1a) [75, 76]. After differentiation, these cultures contain many different cell types such as basal, ciliated and goblet cells. They also produce protective mucus, much like in vivo epithelium. When compared to primary bronchial cells in submerged two-dimensional culture, the gene expression of primary ALI cultures differs significantly. However, the expression pattern of primary human bronchial ALI cultures is comparable to that of in vivo epithelium. The human bronchial cell line Calu-3 has been used as a culture model for respiratory epithelium but its gene expression in ALI cultures is more similar to submerged bronchial cell cultures than differentiated epithelium [77]. Additionally, Calu-3 cells respond differently to MERS-CoV infection compared to primary HAE cultures. During infection in Calu-3 cells, profound apoptosis was detected within 24 h of infection [78] while infection of primary HAE cultures does not result in any disruption of the cell layer [57]. Therefore, the primary tracheobronchial ALI culture model is especially fitting for human respiratory virus research since it accurately recapitulates the primary entry point for these viruses. By using these cultures, virus replication and host interactions can be studied in natural target cells. Further establishing the usefulness of this system HCoV-HKU1 was propagated for the first time in ciliated cells of bronchial HAE cultures in 2010 after culturing it in conventional cell lines had failed [26].
Fig. 1.
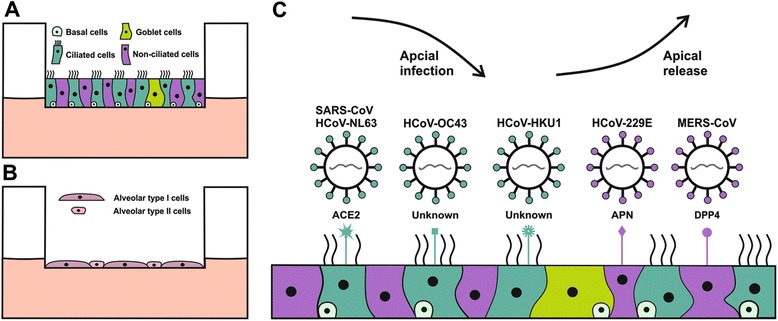
Human airway epithelial cell culture models and HCoV receptor distribution. a: Schematic representation of human tracheobronchial cells at air-liquid interface (ALI). They form a pseudostratified epithelial layer containing different cell types. b: Schematic representation of human alveolar cells at ALI that form single squamous epithelium containing only two cells types, alveolar type I and II cells. c: Illustration of the mode of infection, release and associated cell tropism of the six human coronaviruses (HCoVs) in the human airway epithelial cell culture model. SARS-CoV, HCoV-NL63, HCoV-OC43 and HCoV-HKU1 infect ciliated cells but the receptors for HCoV-HKU1 and HCoV-OC43 are currently unknown. HCoV-229E and MERS-CoV infect non-ciliated cells using different receptors
Alveolar epithelial ALI cultures (Fig. 1b) can also be used for virus-host interaction studies and are especially applicable when a viral infection causes pneumonia and alveolar damage [79]. HCoV-HKU1 has also been propagated in alveolar HAE cultures and exhibits a strong tropism for alveolar type II cells and causes large syncytia formation upon infection [80].
When compared to traditional two dimensional cell cultures, the HAE cultures are more cumbersome and their preparation is time consuming but they do have an advantage over traditional monolayer cell cultures when it comes to virus-host interaction studies. Different types of ALI cultures used for virus research are summarized in Table 2.
Table 2.
Different types of ALI cultures used in coronavirus studies
| Cell types | Origin | HCoVs | Features | References |
|---|---|---|---|---|
| Primary bronchial | Trachea, Bronchus | All HCoVs | Differentiated pseudostratified epithelium, many cell types | [56, 57] |
| Calu-3 | Sub-mucosal glands (adenocarcinoma) | MERS-CoV, SARS-CoV | Single or polarized epithelium, one cell type | [109, 110] |
| Primary alveolar | Alveoli | MERS-CoV, SARS-CoV, HCoV-HKU1 | Differentiated squamous epithelium, two cell types | [79, 80, 111] |
Innate immunity
Within the respiratory epithelium the innate immune system has a major protective role as the first line of defense against respiratory pathogens. In particular, the interferon (IFN) system orchestrates hundreds of different cellular effector proteins that (i) protect the epithelial barrier by altering the physiological and cellular environment, (ii) impair virus propagation, spread and transmission, and (iii) shape the host’s adaptive immune response. Recent publications have demonstrated that the innate immune system is functional in the HAE cell culture system and that most pathogen recognition receptors are expressed and up-regulated upon treatment with exogenous stimuli [57, 81].
In general, HCoVs do not elicit a strong innate immune response in primary target cells of the human airway early during infection. Despite the presence of all major pathogen recognition receptors, no elevated expression of IFN beta, pro-inflammatory cytokines or interferon stimulated genes can be observed up to 12 h post-infection in HAEs infected with HCoV-229E, MERS- or SARS-CoVs [57]. This is most likely due to the intrinsic CoV properties harbored in the replicative non-structural proteins that actively aid in avoiding recognition by the host innate immune system. For example, the 5′ termini of the viral mRNA are capped making them indistinguishable from the host cellular mRNAs and no longer detectable by cellular sensors. Furthermore, CoV replication is associated with the appearance of double membrane vesicles (DMVs) in the host cell cytoplasm, which might serve as a protective shield for viral RNA to prevent recognition by cytoplasmic RNA sensors [82–85].
In addition to the non-structural proteins, various CoV accessory proteins have been discovered to inhibit interferon signaling at different stages of the host innate immune response. For example, MERS-CoV accessory protein 4a inhibits innate antiviral signaling by suppressing the activation of MDA5 and RIGI [86, 87] whereas 4b inhibits the induction of the IFN-beta promoter [88]. While ORF 4a and 4b are IFN antagonists in the genome of MERS-CoV, SARS-CoV ORF3b antagonizes IFN signaling through MAVS/RIGI [89]. Whereas SARS-CoV ORF6 disrupts IFN signaling by blocking the nuclear translocation of STAT1 [89, 90]. These discoveries highlight that HCoVs employ similar yet different strategies to evade the innate immune response during infection in the respiratory epithelium.
Therapy
Despite that respiratory infections with HCoVs can result in severe respiratory disease there are currently no effective prophylactic or therapeutic treatment options available. However, the emergence of novel coronaviruses has emphasized the need to develop effective treatment options. For example, vaccines using the spike proteins of both SARS- and MERS-CoVs have proven protective in animal models [91, 92] suggesting that a vaccine against HCoVs for human use might be achievable.
Additionally, various drugs that inhibit HCoV infection at different stages of the replication cycle have been reported and some could potentially serve as treatment options for HCoV associated severe respiratory disease. For example, patients presenting with severe respiratory disease, caused by SARS- or MERS-CoVs, are generally treated with steroids and interferon, sometimes in combination with the antiviral drug Ribavirin [93–96]. This treatment, however, is not especially effective highlighting the need for HCoV specific antivirals. Many different compounds have been determined to have anti-HCoV activity. For example, protease inhibitors which suppress HCoV entry [97–99], Cyclosporin A (CsA) treatment blocks the replication of coronaviruses from all subgroups [100] and non-immunosuppressive derivatives of CsA represent a possible therapeutic option for both human and animal CoV infections.
HCoV infection can also be inhibited by pre-treating HAE cultures with either recombinant IFN alpha or lambda [57]. Similar effect has also been shown for recombinant IFN alpha and beta which could inhibit MERS-CoV in ex vivo lung cultures [53]. As previously described, IFN treatment of active HCoV infection is not particularly effective in vivo. Therefore, the use of IFN in humans might be limited to prophylactic treatment of exposed persons and/or health care workers treating infected patients.
Screenings of compound libraries have also resulted in the identification of some HCoV specific antivirals. For example, a novel small compound inhibitor (K22) has been identified, and showed to be effective against a broad spectrum of CoVs and could inhibit both HCoV-229E and MERS-CoV in HAE cultures [101]. Additionally, HCoV-NL63 has been inhibited in HAE cultures with polymer-based compounds [102].
To date, most treatment and inhibitor studies have been conducted in HCoV susceptible cell lines. However, the HAE cultures represent an ideal system to test the application and efficacy of those already identified, and new, antiviral compounds against HCoVs in cells that represent the primary site of replication. Furthermore, the HAE cultures are heterogenous, containing many different cellular sub-populations, and would allow for the evaluation of compound toxicity and effect in a differentiated layer similar to human airway epithelium. Compounds already shown to inhibit HCoVs in cell lines should be applied to HAE cultures as well before any animal or human trials.
Conclusions
HCoV induced respiratory diseases are of both medical and socio-economic importance. The emergence of SARS- and MERS-CoV and the yearly circulation of the four common HCoVs highlight the importance of elucidating the different mechanisms employed by HCoVs to evade the host immune system as well as identifying antiviral compounds and human vaccine candidates. The HAE culture system is based on primary human cells offering a unique platform to study respiratory viruses in cells representing the primary entry point of these viruses, bronchial epithelial cells, or investigate the interaction of HCoVs and the distal airways, in type I and II alveolar cells. Additionally, the inclusion of airway epithelial cultures for other species enables the study of zoonosis and animal-to-human transmission. Currently, many aspects of HCoV infection and pathogenesis remain to be determined. The HAE culture system, both tracheobronchial and alveolar, represents a unique platform to study virus-host interaction in natural target cells at the molecular level. These cultures are becoming more common and more relevant to HCoV research. Especially, for those viruses for which there is no animal model, as they provide an organotypic substitute for virus – host interaction studies.
Acknowledgements
We would like to thank Prof. Dr. Volker Thiel for his careful review of the manuscript and Dr. Eveline Kindler for providing components for figures.
Funding
This work was supported by the 3R Foundation, Switzerland (project 128–11) and the University of Bern Initiator Grant.
Abbreviations
- ALI
Air-Liquid Interface
- BCoV
Bovine Coronavirus
- CNS
Central Nervous System
- CoV
Coronavirus
- CsA
Cyclosporin A
- DMV
Double Membrane Vesicles
- FDA
Food and Drug Administration
- hACE2
human Angiotensin Converting Enzyme 2
- HAE
Human Airway Epithelium
- hAPN
human Aminopeptidase N
- HAT
Human Airway Trypsin-like protease
- HCoV
Human Coronavirus
- hDPP4
human Dipeptyl Peptidase 4
- IBV
Infectious Bronchitis Virus
- IFN
Interferon
- MDA5
Melanoma Differentiation-Associated protein 5
- MERS
Middle East Respiratory Syndrome
- NHP
Non-Human Primate
- ORF
Open Reading Frame
- PEDV
Porcine Epidemic Diarrhea Virus
- RBD
Receptor Binding Domain
- RNA
Ribonucleic Acid
- SARS
Severe Acute Respiratory Syndrome
- TMPRSSII
Transmembrane Protease, Serine 2
Footnotes
Competing interests
The authors declare no competing interests.
Authors’ contribution
HRJ wrote the review, designed tables and figures. RD revised the text, tables and figures. Both authors read and approved the final manuscript.
Contributor Information
Hulda R. Jonsdottir, Email: hulda.jonsdottir@vetsuisse.unibe.ch
Ronald Dijkman, Phone: +41 31 631 25 02, Email: ronald.dijkman@vetsuisse.unibe.ch.
References
- 1.Fehr AR, Perlman S. Coronaviruses: an overview of their replication and pathogenesis. Methods Mol Biol. 2015;1282:1–23. doi: 10.1007/978-1-4939-2438-7_1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Brian DA, Baric RS. Coronavirus genome structure and replication. Curr Top Microbiol Immunol. 2005;287:1–30. doi: 10.1007/3-540-26765-4_1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.de Vries AAF, Horzinek MC, Rottier PJM, de Groot RJ. The Genome Organization of the Nidovirales: Similarities and Differences between Arteri-, Toro-, and Coronaviruses. Seminars in Virology. 1997;8:33–47. doi: 10.1006/smvy.1997.0104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pensaert MB, de Bouck P. A new coronavirus-like particle associated with diarrhea in swine. Arch Virol. 1978;58:243–7. doi: 10.1007/BF01317606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cavanagh D. A nomenclature for avian coronavirus isolates and the question of species status. Avian Pathol. 2001;30:109–15. doi: 10.1080/03079450120044506. [DOI] [PubMed] [Google Scholar]
- 6.Fulton RW, Herd HR, Sorensen NJ, Confer AW, Ritchey JW, Ridpath JF, et al. Enteric disease in postweaned beef calves associated with Bovine coronavirus clade 2. J Vet Diagn Invest. 2015;27:97–101. doi: 10.1177/1040638714559026. [DOI] [PubMed] [Google Scholar]
- 7.Le Coupanec A, Desforges M, Meessen-Pinard M, Dube M, Day R, Seidah NG, et al. Cleavage of a Neuroinvasive Human Respiratory Virus Spike Glycoprotein by Proprotein Convertases Modulates Neurovirulence and Virus Spread within the Central Nervous System. PLoS Pathog. 2015;11 doi: 10.1371/journal.ppat.1005261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Weinger JG, Marro BS, Hosking MP, Lane TE. The chemokine receptor CXCR2 and coronavirus-induced neurologic disease. Virology. 2013;435:110–7. doi: 10.1016/j.virol.2012.08.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kyuwa S, Shibata S, Tagawa Y, Iwakura Y, Machii K, Urano T. Acute hepatic failure in IFN-gamma-deficient BALB/c mice after murine coronavirus infection. Virus Res. 2002;83:169–77. doi: 10.1016/S0168-1702(01)00432-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Felten S, Weider K, Doenges S, Gruendl S, Matiasek K, Hermanns W, et al. Detection of feline coronavirus spike gene mutations as a tool to diagnose feline infectious peritonitis. J Feline Med Surg. 2015. [DOI] [PMC free article] [PubMed]
- 11.Hamre D, Procknow JJ. A new virus isolated from the human respiratory tract. Proc Soc Exp Biol Med. 1966;121:190–3. doi: 10.3181/00379727-121-30734. [DOI] [PubMed] [Google Scholar]
- 12.McIntosh K, Dees JH, Becker WB, Kapikian AZ, Chanock RM. Recovery in tracheal organ cultures of novel viruses from patients with respiratory disease. Proc Natl Acad Sci U S A. 1967;57:933–40. doi: 10.1073/pnas.57.4.933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Drosten C, Gunther S, Preiser W, van der Werf S, Brodt HR, Becker S, et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N Engl J Med. 2003;348:1967–76. doi: 10.1056/NEJMoa030747. [DOI] [PubMed] [Google Scholar]
- 14.Rota PA, Oberste MS, Monroe SS, Nix WA, Campagnoli R, Icenogle JP, et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science. 2003;300:1394–9. doi: 10.1126/science.1085952. [DOI] [PubMed] [Google Scholar]
- 15.Summary of probable SARS cases with onset of illness from 1 November 2002 to 31 July 2003. http://www.who.int/csr/sars/country/table2004_04_21/en/. Accessed 7 Dec 2015.
- 16.van der Hoek L, Pyrc K, Jebbink MF, Vermeulen-Oost W, Berkhout RJ, Wolthers KC, et al. Identification of a new human coronavirus. Nat Med. 2004;10:368–73. doi: 10.1038/nm1024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Woo PC, Lau SK, Chu CM, Chan KH, Tsoi HW, Huang Y, et al. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J Virol. 2005;79:884–95. doi: 10.1128/JVI.79.2.884-895.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zaki AM, van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med. 2012;367:1814–20. doi: 10.1056/NEJMoa1211721. [DOI] [PubMed] [Google Scholar]
- 19.Korea Centers for Disease, Control, Prevention Middle East Respiratory Syndrome Coronavirus Outbreak in the Republic of Korea, 2015. Osong Public Health Res Perspec. 2015;6:269–78. doi: 10.1016/j.phrp.2015.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Middle East respiratory syndrome coronavirus (MERS-CoV). http://www.who.int/emergencies/mers-cov/en/. Accessed 7 Dec 2015.
- 21.Bradburne AF, Bynoe ML, Tyrrell DA. Effects of a "new" human respiratory virus in volunteers. Br Med J. 1967;3:767–9. doi: 10.1136/bmj.3.5568.767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kraaijeveld CA, Reed SE, Macnaughton MR. Enzyme-linked immunosorbent assay for detection of antibody in volunteers experimentally infected with human coronavirus strain 229 E. J Clin Microbiol. 1980;12:493–7. doi: 10.1128/jcm.12.4.493-497.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Reed SE. The behaviour of recent isolates of human respiratory coronavirus in vitro and in volunteers: evidence of heterogeneity among 229E-related strains. J Med Virol. 1984;13:179–92. doi: 10.1002/jmv.1890130208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Falsey AR, McCann RM, Hall WJ, Criddle MM, Formica MA, Wycoff D, et al. The "common cold" in frail older persons: impact of rhinovirus and coronavirus in a senior daycare center. J Am Geriatr Soc. 1997;45:706–11. doi: 10.1111/j.1532-5415.1997.tb01474.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang G, Deering C, Macke M, Shao J, Burns R, Blau DM, et al. Human coronavirus 229E infects polarized airway epithelia from the apical surface. J Virol. 2000;74:9234–9. doi: 10.1128/JVI.74.19.9234-9239.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pyrc K, Sims AC, Dijkman R, Jebbink M, Long C, Deming D, et al. Culturing the unculturable: human coronavirus HKU1 infects, replicates, and produces progeny virions in human ciliated airway epithelial cell cultures. J Virol. 2010;84:11255–63. doi: 10.1128/JVI.00947-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Razuri H, Malecki M, Tinoco Y, Ortiz E, Guezala MC, Romero C, et al. Human Coronavirus-Associated Influenza-Like Illness in the Community Setting in Peru. Am J Trop Med Hyg. 2015;93:1038–40. doi: 10.4269/ajtmh.15-0271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lepiller Q, Barth H, Lefebvre F, Herbrecht R, Lutz P, Kessler R, et al. High incidence but low burden of coronaviruses and preferential associations between respiratory viruses. J Clin Microbiol. 2013;51:3039–46. doi: 10.1128/JCM.01078-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Talbot HK, Shepherd BE, Crowe JE, Jr, Griffin MR, Edwards KM, Podsiad AB, et al. The pediatric burden of human coronaviruses evaluated for twenty years. Pediatr Infect Dis J. 2009;28:682–7. doi: 10.1097/INF.0b013e31819d0d27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Drexler JF, Corman VM, Drosten C. Ecology, evolution and classification of bat coronaviruses in the aftermath of SARS. Antiviral Res. 2014;101:45–56. doi: 10.1016/j.antiviral.2013.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Huynh J, Li S, Yount B, Smith A, Sturges L, Olsen JC, et al. Evidence supporting a zoonotic origin of human coronavirus strain NL63. J Virol. 2012;86:12816–25. doi: 10.1128/JVI.00906-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Corman VM, Baldwin HJ, Tateno AF, Zerbinati RM, Annan A, Owusu M, et al. Evidence for an Ancestral Association of Human Coronavirus 229E with Bats. J Virol. 2015;89:11858–70. doi: 10.1128/JVI.01755-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mizumoto K, Saitoh M, Chowell G, Miyamatsu Y, Nishiura H. Estimating the risk of Middle East respiratory syndrome (MERS) death during the course of the outbreak in the Republic of Korea, 2015. International Journal of Infectious Diseases. 2015;39:7–9. doi: 10.1016/j.ijid.2015.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Milne-Price S, Miazgowicz KL, Munster VJ. The emergence of the Middle East Respiratory Syndrome coronavirus (MERS-CoV) Pathogens and disease. 2014;71:119–34. doi: 10.1111/2049-632X.12166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wu Z, Yang L, Ren X, Zhang J, Yang F, Zhang S, et al. ORF8-Related Genetic Evidence for Chinese Horseshoe Bats as the Source of Human Severe Acute Respiratory Syndrome Coronavirus. J Infect Dis. 2016;213:579–83. doi: 10.1093/infdis/jiv476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.He B, Zhang Y, Xu L, Yang W, Yang F, Feng Y, et al. Identification of diverse alphacoronaviruses and genomic characterization of a novel severe acute respiratory syndrome-like coronavirus from bats in China. J Virol. 2014;88:7070–82. doi: 10.1128/JVI.00631-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yang L, Wu Z, Ren X, Yang F, He G, Zhang J, et al. Novel SARS-like betacoronaviruses in bats, China, 2011. Emerg Infect Dis. 2013;19:989–91. doi: 10.3201/eid1906.121648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ge XY, Li JL, Yang XL, Chmura AA, Zhu G, Epstein JH, et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature. 2013;503:535–8. doi: 10.1038/nature12711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Menachery VD, Yount Jr BL, Debbink K, Agnihothram S, Gralinski LE, Plante JA, et al. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat Med. 2015. [DOI] [PMC free article] [PubMed]
- 40.Corman VM, Ithete NL, Richards LR, Schoeman MC, Preiser W, Drosten C, et al. Rooting the phylogenetic tree of middle East respiratory syndrome coronavirus by characterization of a conspecific virus from an African bat. J Virol. 2014;88:11297–303. doi: 10.1128/JVI.01498-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Cotten M, Watson SJ, Zumla AI, Makhdoom HQ, Palser AL, Ong SH, et al. Spread, circulation, and evolution of the Middle East respiratory syndrome coronavirus. MBio. 2014;5:e01062–13. doi: 10.1128/mBio.01062-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Heald-Sargent T, Gallagher T. Ready, set, fuse! The coronavirus spike protein and acquisition of fusion competence. Viruses. 2012;4:557–80. doi: 10.3390/v4040557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bosch BJ, Smits SL, Haagmans BL. Membrane ectopeptidases targeted by human coronaviruses. Current Opinion in Virology. 2014;6:55–60. doi: 10.1016/j.coviro.2014.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bertram S, Dijkman R, Habjan M, Heurich A, Gierer S, Glowacka I, et al. TMPRSS2 activates the human coronavirus 229E for cathepsin-independent host cell entry and is expressed in viral target cells in the respiratory epithelium. J Virol. 2013;87:6150–60. doi: 10.1128/JVI.03372-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Glowacka I, Bertram S, Muller MA, Allen P, Soilleux E, Pfefferle S, et al. Evidence that TMPRSS2 activates the severe acute respiratory syndrome coronavirus spike protein for membrane fusion and reduces viral control by the humoral immune response. J Virol. 2011;85:4122–34. doi: 10.1128/JVI.02232-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yeager CL, Ashmun RA, Williams RK, Cardellichio CB, Shapiro LH, Look AT, et al. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature. 1992;357:420–2. doi: 10.1038/357420a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.van der Velden VH, Wierenga-Wolf AF, Adriaansen-Soeting PW, Overbeek SE, Moller GM, Hoogsteden HC, et al. Expression of aminopeptidase N and dipeptidyl peptidase IV in the healthy and asthmatic bronchus. Clin Exp Allergy. 1998;28:110–20. doi: 10.1046/j.1365-2222.1998.00198.x. [DOI] [PubMed] [Google Scholar]
- 48.Tseng CT, Huang C, Newman P, Wang N, Narayanan K, Watts DM, et al. Severe acute respiratory syndrome coronavirus infection of mice transgenic for the human Angiotensin-converting enzyme 2 virus receptor. J Virol. 2007;81:1162–73. doi: 10.1128/JVI.01702-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hofmann H, Pyrc K, van der Hoek L, Geier M, Berkhout B, Pohlmann S. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci U S A. 2005;102:7988–93. doi: 10.1073/pnas.0409465102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hamming I, Timens W, Bulthuis ML, Lely AT, Navis G, van Goor H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J Pathol. 2004;203:631–7. doi: 10.1002/path.1570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Raj VS, Mou H, Smits SL, Dekkers DH, Muller MA, Dijkman R, et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature. 2013;495:251–4. doi: 10.1038/nature12005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lambeir AM, Durinx C, Scharpe S, De Meester I. Dipeptidyl-peptidase IV from bench to bedside: an update on structural properties, functions, and clinical aspects of the enzyme DPP IV. Crit Rev Clin Lab Sci. 2003;40:209–94. doi: 10.1080/713609354. [DOI] [PubMed] [Google Scholar]
- 53.Chan RW, Chan MC, Agnihothram S, Chan LL, Kuok DI, Fong JH, et al. Tropism of and innate immune responses to the novel human betacoronavirus lineage C virus in human ex vivo respiratory organ cultures. J Virol. 2013;87:6604–14. doi: 10.1128/JVI.00009-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Huang X, Dong W, Milewska A, Golda A, Qi Y, Zhu QK, et al. Human Coronavirus HKU1 Spike Protein Uses O-Acetylated Sialic Acid as an Attachment Receptor Determinant and Employs Hemagglutinin-Esterase Protein as a Receptor-Destroying Enzyme. J Virol. 2015;89:7202–13. doi: 10.1128/JVI.00854-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Krempl C, Schultze B, Herrler G. Analysis of cellular receptors for human coronavirus OC43. Adv Exp Med Biol. 1995;380:371–4. doi: 10.1007/978-1-4615-1899-0_60. [DOI] [PubMed] [Google Scholar]
- 56.Dijkman R, Jebbink MF, Koekkoek SM, Deijs M, Jonsdottir HR, Molenkamp R, et al. Isolation and characterization of current human coronavirus strains in primary human epithelial cell cultures reveal differences in target cell tropism. J Virol. 2013;87:6081–90. doi: 10.1128/JVI.03368-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kindler E, Jonsdottir HR, Muth D, Hamming OJ, Hartmann R, Rodriguez R, et al. Efficient replication of the novel human betacoronavirus EMC on primary human epithelium highlights its zoonotic potential. MBio. 2013;4:e00611–12. doi: 10.1128/mBio.00611-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Thiel V, Herold J, Schelle B, Siddell SG. Infectious RNA transcribed in vitro from a cDNA copy of the human coronavirus genome cloned in vaccinia virus. J Gen Virol. 2001;82:1273–81. doi: 10.1099/0022-1317-82-6-1273. [DOI] [PubMed] [Google Scholar]
- 59.St-Jean JR, Desforges M, Almazan F, Jacomy H, Enjuanes L, Talbot PJ. Recovery of a neurovirulent human coronavirus OC43 from an infectious cDNA clone. J Virol. 2006;80:3670–4. doi: 10.1128/JVI.80.7.3670-3674.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Donaldson EF, Yount B, Sims AC, Burkett S, Pickles RJ, Baric RS. Systematic assembly of a full-length infectious clone of human coronavirus NL63. J Virol. 2008;82:11948–57. doi: 10.1128/JVI.01804-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Barnard DL. Animal models for the study of influenza pathogenesis and therapy. Antiviral Res. 2009;82:A110–22. doi: 10.1016/j.antiviral.2008.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Gretebeck LM, Subbarao K. Animal models for SARS and MERS coronaviruses. Curr Opin Virol. 2015;13:123–9. doi: 10.1016/j.coviro.2015.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lassnig C, Sanchez CM, Egerbacher M, Walter I, Majer S, Kolbe T, et al. Development of a transgenic mouse model susceptible to human coronavirus 229E. Proc Natl Acad Sci U S A. 2005;102:8275–80. doi: 10.1073/pnas.0408589102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Wentworth DE, Tresnan DB, Turner BC, Lerman IR, Bullis B, Hemmila EM, et al. Cells of human aminopeptidase N (CD13) transgenic mice are infected by human coronavirus-229E in vitro, but not in vivo. Virology. 2005;335:185–97. doi: 10.1016/j.virol.2005.02.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhao J, Li K, Wohlford-Lenane C, Agnihothram SS, Fett C, Zhao J, et al. Rapid generation of a mouse model for Middle East respiratory syndrome. Proc Natl Acad Sci U S A. 2014;111:4970–5. doi: 10.1073/pnas.1323279111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Agrawal AS, Garron T, Tao X, Peng BH, Wakamiya M, Chan TS, et al. Generation of a transgenic mouse model of Middle East respiratory syndrome coronavirus infection and disease. J Virol. 2015;89:3659–70. doi: 10.1128/JVI.03427-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Schaap-Nutt A, Liesman R, Bartlett EJ, Scull MA, Collins PL, Pickles RJ, et al. Human parainfluenza virus serotypes differ in their kinetics of replication and cytokine secretion in human tracheobronchial airway epithelium. Virology. 2012;433:320–8. doi: 10.1016/j.virol.2012.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Deng X, Li Y, Qiu J. Human bocavirus 1 infects commercially available primary human airway epithelium cultures productively. J Virol Methods. 2014;195:112–9. doi: 10.1016/j.jviromet.2013.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Thompson CI, Barclay WS, Zambon MC, Pickles RJ. Infection of human airway epithelium by human and avian strains of influenza a virus. J Virol. 2006;80:8060–8. doi: 10.1128/JVI.00384-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Dijkman R, Koekkoek SM, Molenkamp R, Schildgen O, van der Hoek L. Human bocavirus can be cultured in differentiated human airway epithelial cells. J Virol. 2009;83:7739–48. doi: 10.1128/JVI.00614-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Gordon S, Daneshian M, Bouwstra J, Caloni F, Constant S, Davies DE, et al. Non-animal models of epithelial barriers (skin, intestine and lung) in research, industrial applications and regulatory toxicology. Altex. 2015;32:327–78. doi: 10.14573/altex.1510051. [DOI] [PubMed] [Google Scholar]
- 72.Crystal RG, Randell SH, Engelhardt JF, Voynow J, Sunday ME. Airway epithelial cells: current concepts and challenges. Proc Am Thorac Soc. 2008;5:772–7. doi: 10.1513/pats.200805-041HR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Whitsett JA, Weaver TE. Alveolar development and disease. Am J Respir Cell Mol Biol. 2015;53:1–7. doi: 10.1165/rcmb.2015-0128PS. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Rackley CR, Stripp BR. Building and maintaining the epithelium of the lung. J Clin Invest. 2012;122:2724–30. doi: 10.1172/JCI60519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.de Jong PM, van Sterkenburg MA, Kempenaar JA, Dijkman JH, Ponec M. Serial culturing of human bronchial epithelial cells derived from biopsies. In Vitro Cell Dev Biol Anim. 1993;29a:379–87. doi: 10.1007/BF02633985. [DOI] [PubMed] [Google Scholar]
- 76.de Jong PM, van Sterkenburg MA, Hesseling SC, Kempenaar JA, Mulder AA, Mommaas AM, et al. Ciliogenesis in human bronchial epithelial cells cultured at the air-liquid interface. Am J Respir Cell Mol Biol. 1994;10:271–7. doi: 10.1165/ajrcmb.10.3.8117445. [DOI] [PubMed] [Google Scholar]
- 77.Pezzulo AA, Starner TD, Scheetz TE, Traver GL, Tilley AE, Harvey BG, et al. The air-liquid interface and use of primary cell cultures are important to recapitulate the transcriptional profile of in vivo airway epithelia. Am J Physiol Lung Cell Mol Physiol. 2011;300:L25–31. doi: 10.1152/ajplung.00256.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Tao X, Hill TE, Morimoto C, Peters CJ, Ksiazek TG, Tseng CT. Bilateral entry and release of Middle East respiratory syndrome coronavirus induces profound apoptosis of human bronchial epithelial cells. J Virol. 2013;87:9953–8. doi: 10.1128/JVI.01562-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Qian Z, Travanty EA, Oko L, Edeen K, Berglund A, Wang J, et al. Innate immune response of human alveolar type II cells infected with severe acute respiratory syndrome-coronavirus. Am J Respir Cell Mol Biol. 2013;48:742–8. doi: 10.1165/rcmb.2012-0339OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Dominguez SR, Travanty EA, Qian Z, Mason RJ. Human coronavirus HKU1 infection of primary human type II alveolar epithelial cells: cytopathic effects and innate immune response. PLoS One. 2013;8 doi: 10.1371/journal.pone.0070129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lauber C, Vieyres G, Terczynska-Dyla E. Anggakusuma, Dijkman R, Gad HH, et al.: Transcriptome analysis reveals a classical interferon signature induced by IFNlambda4 in human primary cells. Genes Immun. 2015;16:414–21. doi: 10.1038/gene.2015.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.van Hemert MJ, van den Worm SH, Knoops K, Mommaas AM, Gorbalenya AE, Snijder EJ. SARS-coronavirus replication/transcription complexes are membrane-protected and need a host factor for activity in vitro. PLoS Pathog. 2008;4 doi: 10.1371/journal.ppat.1000054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Knoops K, Kikkert M, Worm SH, Zevenhoven-Dobbe JC, van der Meer Y, Koster AJ, et al. SARS-coronavirus replication is supported by a reticulovesicular network of modified endoplasmic reticulum. PLoS Biol. 2008;6 doi: 10.1371/journal.pbio.0060226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Daffis S, Szretter KJ, Schriewer J, Li J, Youn S, Errett J, et al. 2'-O methylation of the viral mRNA cap evades host restriction by IFIT family members. Nature. 2010;468:452–6. doi: 10.1038/nature09489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zust R, Cervantes-Barragan L, Habjan M, Maier R, Neuman BW, Ziebuhr J, et al. Ribose 2'-O-methylation provides a molecular signature for the distinction of self and non-self mRNA dependent on the RNA sensor Mda5. Nat Immunol. 2011;12:137–43. doi: 10.1038/ni.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Siu KL, Yeung ML, Kok KH, Yuen KS, Kew C, Lui PY, et al. Middle east respiratory syndrome coronavirus 4a protein is a double-stranded RNA-binding protein that suppresses PACT-induced activation of RIG-I and MDA5 in the innate antiviral response. J Virol. 2014;88:4866–76. doi: 10.1128/JVI.03649-13. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 87.Niemeyer D, Zillinger T, Muth D, Zielecki F, Horvath G, Suliman T, et al. Middle East respiratory syndrome coronavirus accessory protein 4a is a type I interferon antagonist. J Virol. 2013;87:12489–95. doi: 10.1128/JVI.01845-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Matthews KL, Coleman CM, van der Meer Y, Snijder EJ, Frieman MB. The ORF4b-encoded accessory proteins of Middle East respiratory syndrome coronavirus and two related bat coronaviruses localize to the nucleus and inhibit innate immune signalling. J Gen Virol. 2014;95:874–82. doi: 10.1099/vir.0.062059-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Kopecky-Bromberg SA, Martinez-Sobrido L, Frieman M, Baric RA, Palese P. Severe acute respiratory syndrome coronavirus open reading frame (ORF) 3b, ORF 6, and nucleocapsid proteins function as interferon antagonists. J Virol. 2007;81:548–57. doi: 10.1128/JVI.01782-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Frieman M, Yount B, Heise M, Kopecky-Bromberg SA, Palese P, Baric RS. Severe acute respiratory syndrome coronavirus ORF6 antagonizes STAT1 function by sequestering nuclear import factors on the rough endoplasmic reticulum/Golgi membrane. J Virol. 2007;81:9812–24. doi: 10.1128/JVI.01012-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Bisht H, Roberts A, Vogel L, Bukreyev A, Collins PL, Murphy BR, et al. Severe acute respiratory syndrome coronavirus spike protein expressed by attenuated vaccinia virus protectively immunizes mice. Proc Natl Acad Sci U S A. 2004;101:6641–6. doi: 10.1073/pnas.0401939101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Volz A, Kupke A, Song F, Jany S, Fux R, Shams-Eldin H, et al. Protective efficacy of recombinant modified vaccinia virus Ankara delivering Middle East respiratory syndrome coronavirus spike glycoprotein. J Virol. 2015;89:8651–6. doi: 10.1128/JVI.00614-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Al-Tawfiq JA, Momattin H, Dib J, Memish ZA. Ribavirin and interferon therapy in patients infected with the Middle East respiratory syndrome coronavirus: an observational study. Int J Infect Dis. 20:42–6. [DOI] [PMC free article] [PubMed]
- 94.Zumla A, Azhar EI, Arabi Y, Alotaibi B, Rao M, McCloskey B, et al. Host-directed therapies for improving poor treatment outcomes associated with the middle east respiratory syndrome coronavirus infections. Int J Infect Dis. 40:71–4. [DOI] [PMC free article] [PubMed]
- 95.Omrani AS, Saad MM, Baig K, Bahloul A, Abdul-Matin M, Alaidaroos AY, et al. Ribavirin and interferon alfa-2a for severe Middle East respiratory syndrome coronavirus infection: a retrospective cohort study. Lancet Infect Dis. 14:1090–5. [DOI] [PMC free article] [PubMed]
- 96.Shalhoub S, Farahat F, Al-Jiffri A, Simhairi R, Shamma O, Siddiqi N, et al. IFN-alpha2a or IFN-beta1a in combination with ribavirin to treat Middle East respiratory syndrome coronavirus pneumonia: a retrospective study. J Antimicrob Chemother. 2015;70:2129–32. doi: 10.1093/jac/dkv085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kawase M, Shirato K, van der Hoek L, Taguchi F, Matsuyama S. Simultaneous treatment of human bronchial epithelial cells with serine and cysteine protease inhibitors prevents severe acute respiratory syndrome coronavirus entry. J Virol. 2012;86:6537–45. doi: 10.1128/JVI.00094-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Simmons G, Zmora P, Gierer S, Heurich A, Pohlmann S. Proteolytic activation of the SARS-coronavirus spike protein: cutting enzymes at the cutting edge of antiviral research. Antiviral Res. 2013;100:605–14. doi: 10.1016/j.antiviral.2013.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Baez-Santos YM, St John SE, Mesecar AD. The SARS-coronavirus papain-like protease: structure, function and inhibition by designed antiviral compounds. Antiviral Res. 2015;115:21–38. doi: 10.1016/j.antiviral.2014.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Pfefferle S, Schopf J, Kogl M, Friedel CC, Muller MA, Carbajo-Lozoya J, et al. The SARS-coronavirus-host interactome: identification of cyclophilins as target for pan-coronavirus inhibitors. PLoS Pathog. 2011;7:e1002331. doi: 10.1371/journal.ppat.1002331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Lundin A, Dijkman R, Bergstrom T, Kann N, Adamiak B, Hannoun C, et al. Targeting membrane-bound viral RNA synthesis reveals potent inhibition of diverse coronaviruses including the Middle East respiratory syndrome virus. PLoS Pathog. 2014;10:e1004166. doi: 10.1371/journal.ppat.1004166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Milewska A, Ciejka J, Kaminski K, Karewicz A, Bielska D, Zeglen S, et al. Novel polymeric inhibitors of HCoV-NL63. Antiviral Res. 2013;97:112–21. doi: 10.1016/j.antiviral.2012.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Kapikian AZ, James HD, Jr, Kelly SJ, Dees JH, Turner HC, McIntosh K, et al. Isolation from man of "avian infectious bronchitis virus-like" viruses (coronaviruses) similar to 229E virus, with some epidemiological observations. J Infect Dis. 1969;119:282–90. doi: 10.1093/infdis/119.3.282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Breslin JJ, Mork I, Smith MK, Vogel LK, Hemmila EM, Bonavia A, et al. Human coronavirus 229E: receptor binding domain and neutralization by soluble receptor at 37 degrees C. J Virol. 2003;77:4435–8. doi: 10.1128/JVI.77.7.4435-4438.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.McIntosh K, Becker WB, Chanock RM. Growth in suckling-mouse brain of "IBV-like" viruses from patients with upper respiratory tract disease. Proc Natl Acad Sci U S A. 1967;58:2268–73. doi: 10.1073/pnas.58.6.2268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Xiao X, Chakraborti S, Dimitrov AS, Gramatikoff K, Dimitrov DS. The SARS-CoV S glycoprotein: expression and functional characterization. Biochem Biophys Res Commun. 2003;312:1159–64. doi: 10.1016/j.bbrc.2003.11.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Lin HX, Feng Y, Wong G, Wang L, Li B, Zhao X, et al. Identification of residues in the receptor-binding domain (RBD) of the spike protein of human coronavirus NL63 that are critical for the RBD-ACE2 receptor interaction. J Gen Virol. 2008;89:1015–24. doi: 10.1099/vir.0.83331-0. [DOI] [PubMed] [Google Scholar]
- 108.Mou H, Raj VS, van Kuppeveld FJ, Rottier PJ, Haagmans BL, Bosch BJ. The receptor binding domain of the new Middle East respiratory syndrome coronavirus maps to a 231-residue region in the spike protein that efficiently elicits neutralizing antibodies. J Virol. 2013;87:9379–83. doi: 10.1128/JVI.01277-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Sims AC, Burkett SE, Yount B, Pickles RJ. SARS-CoV replication and pathogenesis in an in vitro model of the human conducting airway epithelium. Virus Res. 2008;133:33–44. doi: 10.1016/j.virusres.2007.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Josset L, Menachery VD, Gralinski LE, Agnihothram S, Sova P, Carter VS, et al. Cell host response to infection with novel human coronavirus EMC predicts potential antivirals and important differences with SARS coronavirus. MBio. 2013;4:e00165–13. doi: 10.1128/mBio.00165-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Meyerholz DK, Lambertz AM, McCray PB., Jr Dipeptidyl peptidase 4 distribution in the human respiratory tract: implications for the Middle East respiratory syndrome. Am J Pathol. 2016;186:78–86. doi: 10.1016/j.ajpath.2015.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]