

Abstract
We report the first systematic evolution and study of tRNA variants that are able to read a set of UAGN (N = A, G, U, C) codons in a genomically recoded E. coli strain that lacks any endogenous in-frame UAGN sequences and release factor 1. Through randomizing bases in anticodon stem-loop followed by a functional selection, we identified tRNA mutants with significantly improved UAGN decoding efficiency, which will augment the current efforts on genetic code expansion through quadruplet decoding. We found that an extended anticodon loop with an extra nucleotide was required for a detectable efficiency in UAGN decoding. We also observed that this crucial extra nucleotide was converged to a U (position 33.5) in all of the top tRNA hits no matter which UAGN codon they suppress. The insertion of U33.5 in the anticodon loop likely causes tRNA distortion and affects anticodon-codon interaction, which induces +1 frameshift in the P site of ribosome. A new model was proposed to explain the observed features of UAGN decoding. Overall, our findings elevate our understanding of the +1 frameshift mechanism and provide a useful guidance for further efforts on the genetic code expansion using a non-canonical quadruplet reading frame.
While the triplet codon is the predominant form of the current genetic code, programmed frameshifts, e.g., +1 frameshift caused by the non-canonical reading of a quadruplet codon, is a natural process. These programmed +1 frameshift events generally involve certain recoding signals that are embedded in the mRNA1. On the other hand, tRNA mutants that contain extended anticodon loops (8-base instead of normal 7-base loop) could induce +1 frameshift independent of recoding signals2,3,4,5,6,7,8,9. Recent developments in genetic code engineering also demonstrated that quadruplet codons could be used to encode unnatural amino acids (unAAs) under experimental conditions10,11,12,13,14. Here we present a systematic study on UAGN (N = A, G, U, C) decoding in order to further expand the current genetic code through suppressing quadruplet codons. Efforts have been made to expand the cellular genetic code with multiple quadruplet codons as an enabling synthetic biology tool for biological investigations10,11,12,13,14. Theoretically, a quadruplet codon table provides a maximum of 256 codons, which potentially allows a significant expansion of the current genetic code to facilitate biological studies15,16,17 and to eventually enable the ribosomal synthesis of completely artificial biopolymers as new biomaterials. Besides genetic code expansion, we also intend to use UAGN decoding as a model system to study quadruplet codon decoding (+1 frameshift) mechanism.
Two major working models were proposed to explain the mechanism of quadruplet codon decoding (+1 frameshift) with tRNAs bearing extended anticodon loops: (1) the yardstick model18,19,20 (Fig. 1A) states that the anticodon loop of tRNA interacts with all four bases of quadruplet codon in the A site of the ribosome, which leads to subsequent quadruplet translocation from the A site to the P site. This model is supported by observations that a number of tRNAs with an extended anticodon loop form apparent Watson-Crick complementarity to their cognate quadruplet codons at all four anticodon positions4,11,21,22,23,24,25. A primer extension toeprint assay on tRNACCCG also supported this theory19. An NMR study showed that the anticodon stem-loop (ASL) of the tRNACCCG lacked the conserved U-turn motif and could potentially undergo conformational adjustment in order to interact with quadruplet codon19. In an altered yardstick model26, the complete quadruplet codon-anticodon interaction in the A site is not required. Instead, the extra nucleotide widens the ASL and allows the anticodon nucleotide-34 to interact with either the fourth or the third and fourth codon bases. This model is supported by reported crystal structures of the 30S ribosomal subunit of Thermus thermophilus in complex with tRNAs known to facilitate +1 frameshifting and their cognate mRNA26; (2) the slippery model7,27,28,29 (Fig. 1B) entails that tRNA makes a normal three-base codon-anticodon interaction in the A site of ribosome and translocation is always triplet. An anticodon-mRNA re-pairing subsequently occurs in the P-site with a slip of the mRNA by one base, which leads to an apparent quadruplet codon decoding. The re-pairing event usually requires that the anticodon binds to a cognate or a near cognate codon in the +1 frame. For example (Fig. S9), a well-studied CCCU suppressor (tRNASufA6)5,27,30 has a C34G35G36 anticodon that forms two G-C base pairs both before and after the re-pairing event, which does not cause significant penalty in binding energy. Evidences suggest that the “ribosomal grip” of the peptidyl-tRNA is pivotal for maintaining the reading frame31. The slippery model is also supported by observations that +1 frameshift efficiency was significantly affected by the decoding of the A-site codon following the quadruplet codon, indicating that the frameshift happens in the P site32,33,34,35. Regardless of the merits of each model, key aspects, such as the codon-anticodon interaction and translocation mechanism, of the quadruplet decoding by tRNAs with extended anticodon loops remain to be resolved.
Figure 1. Hypothetical models for +1 frameshift (quadruplet decoding) with tRNAs containing extended anticodon loop.

(A) The yardstick model features quadruplet anticodon-codon interaction in the A site followed by quadruplet translocation; (B) The Slippery model features normal triplet decoding in the A site, normal triplet translocation, and a slippage in the P site.
We14 and other36 recently reported a new approach to enhance the efficiency of quadruplet codon decoding, which is based on the engineering of tRNA anticodon stem-loop (ASL). By using this method, here we present the first systematic evolution and study of tRNA variants that decode UAGN codons in a genomically recoded E. coli strain that lacks any endogenous in-frame UAGN sequences. Most previous studies on quadruplet codon decoding10,11,12,13,14 were conducted in hosts that contain a large number of in-frame four base sequences that are identical to the quadruplet codon of interest. An efficient quadruplet decoding system would lead to genome-wide, undesirable frameshifts, which could impair protein synthesis and introduce additional difficulties to the mechanistic investigations on quadruplet codon decoding. In addition, competing recognition of the first three bases of a quadruplet codon by triplet decoding tRNAs or release factors could reduce the efficiency of quadruplet codon decoding and further complicate the study. A recent synthetic biology effort led to the successful construction of an E. coli strain, C321.ΔA, that does not express release factor 1 (RF1; decodes the UAG nonsense codon as the termination signal of protein translation) and does not contain endogenous UAG nonsense codon (replaced with ochre UAA nonsense codon)37. This genomically recoded E. coli strain represents an excellent host for the evolution and study of UAGN decoding by eliminating the pitfalls mentioned above. In fact, a recent study38 showed that a Methanocaldococcus jannaschii-derived tRNA,  , could efficiently decode UAGA codons in E. coli C321.ΔA by simply replacing the CUA anticodon of M. jannaschii-derived amber suppressor tRNA,
, could efficiently decode UAGA codons in E. coli C321.ΔA by simply replacing the CUA anticodon of M. jannaschii-derived amber suppressor tRNA,  , with UCUA anticodon. Here we report the engineering of the ASL of a Methanosarcina mazei-derived tRNA,
, with UCUA anticodon. Here we report the engineering of the ASL of a Methanosarcina mazei-derived tRNA, 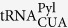 , in order to efficiently decode a set of UAGN (N = A, G, U, C) codons. We chose
, in order to efficiently decode a set of UAGN (N = A, G, U, C) codons. We chose 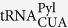 as our model tRNA since its anticodon is unlikely used as an important recognition element by its cognate PylRS according to structural data39. We also confirmed this notion by showing that
as our model tRNA since its anticodon is unlikely used as an important recognition element by its cognate PylRS according to structural data39. We also confirmed this notion by showing that 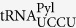 , a derivative of
, a derivative of 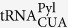 in which the anticodon CUA was replaced with UCCU, could be charged with an unAA by a PylRS variant to decode AGGA codon14. This structural feature allowed us to engineer the ASL of
in which the anticodon CUA was replaced with UCCU, could be charged with an unAA by a PylRS variant to decode AGGA codon14. This structural feature allowed us to engineer the ASL of 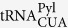 without interfering with the tRNA aminoacylation step. Our work on converting the
without interfering with the tRNA aminoacylation step. Our work on converting the 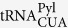 into a quadruplet codon decoding tRNA would augment the current efforts on the genetic incorporation of unAAs by PylRS variants. Since our ASL mutations in tRNAPyl unlikely affect PylRS mutants’ specificity towards their unAA substrates, evolved tRNAs can pair with PylRS mutants to incorporate a range of different unAAs.
into a quadruplet codon decoding tRNA would augment the current efforts on the genetic incorporation of unAAs by PylRS variants. Since our ASL mutations in tRNAPyl unlikely affect PylRS mutants’ specificity towards their unAA substrates, evolved tRNAs can pair with PylRS mutants to incorporate a range of different unAAs.
Our ability to evolve quadruplet decoding tRNAs that are derived from the same ancestor but decode a set of UAGN codons will enable a systematic investigation on the mechanism of quadruplet codon decoding with tRNAs bearing extended anticodon loops. In addition, inferences from this work will likely bring better understanding of ribosome function in the future as well, especially how the ribosome maintains the reading frame and its relation to translocation, which remains one of the key questions to be understood in protein translation.
Results
tRNA (with an extended anticodon loop) library construction and selection for UAGN (N = A, G, U, C) codon decoding
An additional and randomized nucleotide, N (N = A, G, U, or C), was inserted between U33 and C34 in the anticodon loop of 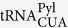 . This extra nucleotide is numbered as 33.5 (Fig. 2) so that the conventional numbering of tRNA molecule at other positions was not changed. Randomization at position 33.5 resulted in a mixture of four tRNAs, including
. This extra nucleotide is numbered as 33.5 (Fig. 2) so that the conventional numbering of tRNA molecule at other positions was not changed. Randomization at position 33.5 resulted in a mixture of four tRNAs, including 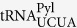 -wt,
-wt, 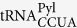 -wt,
-wt, 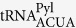 -wt, and
-wt, and 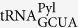 -wt. The ‘wt’ designation indicates that these tRNAs have identical sequence as their parent, wild-type
-wt. The ‘wt’ designation indicates that these tRNAs have identical sequence as their parent, wild-type 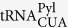 except the extra nucleotide in the anticodon loop. Each of the four tRNAs now contains an extended (eight rather than seven nucleotides) anticodon loop. We expect that such modification of tRNA can facilitate UAGN decoding since previous work suggested that tRNA mutants containing extended anticodon loops could induce +1 frameshift independent of recoding signals2,3,4.
except the extra nucleotide in the anticodon loop. Each of the four tRNAs now contains an extended (eight rather than seven nucleotides) anticodon loop. We expect that such modification of tRNA can facilitate UAGN decoding since previous work suggested that tRNA mutants containing extended anticodon loops could induce +1 frameshift independent of recoding signals2,3,4.
Figure 2. Clover leaf structures of 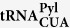 and
and 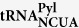 .
.

Comparing to 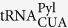 ,
, 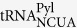 mutants contain eight rather than seven bases in its anticodon loop. The additional base, N, is numbered as 33.5. Nucleotides in the anticodon stem-loop (ASL) that were randomized are shown in bold.
mutants contain eight rather than seven bases in its anticodon loop. The additional base, N, is numbered as 33.5. Nucleotides in the anticodon stem-loop (ASL) that were randomized are shown in bold.
As mutations in the ASL of 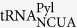 (N = A, G, U, or C) likely change the tRNA’s interaction with ribosome, mRNA, and other components of the translational machinery, we created a tRNA library (Library-NCUA11, theoretical diversity 4.2 × 106; actual library size 5.2 × 106) in which four nucleotides of the anticodon loop (32, 33, 37, 38; Fig. 2) and six nucleotides of the anticodon stem (29, 30, 31, 39, 40, 41; Fig. 2) were completely randomized in
(N = A, G, U, or C) likely change the tRNA’s interaction with ribosome, mRNA, and other components of the translational machinery, we created a tRNA library (Library-NCUA11, theoretical diversity 4.2 × 106; actual library size 5.2 × 106) in which four nucleotides of the anticodon loop (32, 33, 37, 38; Fig. 2) and six nucleotides of the anticodon stem (29, 30, 31, 39, 40, 41; Fig. 2) were completely randomized in 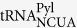 . The resulting tRNA library was subsequently subjected to positive selections against UAGA, UAGG, UAGU, and UAGC codons, respectively, in E. coli C321.ΔA.exp in order to identify
. The resulting tRNA library was subsequently subjected to positive selections against UAGA, UAGG, UAGU, and UAGC codons, respectively, in E. coli C321.ΔA.exp in order to identify 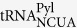 variants that can decode these quadruplet codons. Randomization of the nucleotide at position 33.5 would allow us to select
variants that can decode these quadruplet codons. Randomization of the nucleotide at position 33.5 would allow us to select 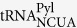 variants against fully matched or fourth-base mismatched quadruplet codons in order to examine the requirement of codon-anticodon pairing in quadruplet codon decoding. Previous data suggest that full quadruplet Watson-Crick base pairing is not absolutely required but indeed results in the highest quadruplet decoding efficiency4,7,10,22,24,25,40. With a systematic study and more suitable model system, we expect to gain more reliable and deeper insights into this question.
variants against fully matched or fourth-base mismatched quadruplet codons in order to examine the requirement of codon-anticodon pairing in quadruplet codon decoding. Previous data suggest that full quadruplet Watson-Crick base pairing is not absolutely required but indeed results in the highest quadruplet decoding efficiency4,7,10,22,24,25,40. With a systematic study and more suitable model system, we expect to gain more reliable and deeper insights into this question.
The selection was conducted in the presence of Nε-(tert-butyloxy-carbonyl)-L-lysine (Boc-Lys), BocLysRS (a pyrrolysyl-tRNA synthetase mutant that specifically charges 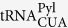 with Boc-Lys)41, chloramphenicol (concentrations ranging from 34 to 75 μg mL−1), and a chloramphenicol acetyltransferase mutant gene containing the quadruplet codon of interest at a permissive site (the codon for Gln98 is replaced with a quadruplet codon). The survivors were further screened with a range of different concentrations of chloramphenicol (50, 75, and 100 μg mL−1) in the presence and absence of Boc-Lys. It was considered a hit if the cell grew at 100 μg mL−1 chloramphenicol in the presence of Boc-Lys, but did not grow at 50 μg mL−1 chloramphenicol in the absence of Boc-Lys. Multiple hits were obtained for each of the UAGA, UAGG, and UAGU codons. We failed to identify any hit for UAGC codon in a number of attempts. Each hit was designated as UAGN-X, where N represents A, G, or U, and X is the hit number.
with Boc-Lys)41, chloramphenicol (concentrations ranging from 34 to 75 μg mL−1), and a chloramphenicol acetyltransferase mutant gene containing the quadruplet codon of interest at a permissive site (the codon for Gln98 is replaced with a quadruplet codon). The survivors were further screened with a range of different concentrations of chloramphenicol (50, 75, and 100 μg mL−1) in the presence and absence of Boc-Lys. It was considered a hit if the cell grew at 100 μg mL−1 chloramphenicol in the presence of Boc-Lys, but did not grow at 50 μg mL−1 chloramphenicol in the absence of Boc-Lys. Multiple hits were obtained for each of the UAGA, UAGG, and UAGU codons. We failed to identify any hit for UAGC codon in a number of attempts. Each hit was designated as UAGN-X, where N represents A, G, or U, and X is the hit number.
Characterization and validation of evolved tRNAs
In order to examine the efficiency and fidelity of UAGN codon decoding by the obtained tRNAs, the four quadruplet codons of interest, UAGA, UAGG, UAGU, and UAGC were introduced into GFPUV (a variant of green fluorescent protein) to replace the Asn149 codon and yielded GFPUV-Asn149UAGA, GFPUV-Asn149UAGG, GFPUV-Asn149UAGU, and GFPUV-Asn49UAGC, respectively. A series of plasmids, pGFPUV-NCUA-X (N = A, G, U, or C; X = hit number), were subsequently constructed. Each plasmid contains a tRNA mutant of interest (expressed under the control of a lpp promoter) and a GFPUV mutant (driven by a T5 promoter) containing corresponding quadruplet codon. As controls, we also constructed four pGFPUV-NCUA-wt (N = A, G, U, or C) plasmids in which 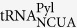 -wt (
-wt (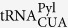 with an extra base at position 33.5) were used instead of the tRNAs mutants that were obtained from selection.
with an extra base at position 33.5) were used instead of the tRNAs mutants that were obtained from selection.
Plasmids pGFPUV-NCUA-X and pGFPUV-NCUA-wt were transformed individually into E. coli C321.ΔA strain containing plasmid pBK-BocLysRS that carries a copy of BocLysRS-encoding gene. Protein expression was carried out in LB medium supplemented with or without 5 mM Boc-Lys. The Fig. 3 shows the top three tRNA mutants for decoding UAGA, UAGG, and UAGU codons. All mutants exhibited significantly enhanced quadruplet decoding efficiency in the presence of Boc-Lys relative to that of the corresponding 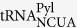 -wt. The best UAGA-, UAGG-, and UAGU-decoding tRNA mutant displayed approximately 55, 40, and 9 folds improvement over their corresponding
-wt. The best UAGA-, UAGG-, and UAGU-decoding tRNA mutant displayed approximately 55, 40, and 9 folds improvement over their corresponding 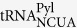 -wt (Fig. 3). Notably, the decoding efficiency of UAGA and UAGG codons was much higher than that of UAGU codon. In addition, fluorescence analyses of E. coli cultures also showed that significant amount of full-length GFPUV protein was only produced in the presence of Boc-Lys for all of our evolved
-wt (Fig. 3). Notably, the decoding efficiency of UAGA and UAGG codons was much higher than that of UAGU codon. In addition, fluorescence analyses of E. coli cultures also showed that significant amount of full-length GFPUV protein was only produced in the presence of Boc-Lys for all of our evolved 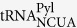 variants that were examined (Fig. 3). This result indicates that these tRNA mutants can only be charged with Boc-Lys by BocLysRS and cannot be charged with natural amino acids by BocLysRS or any endogenous aminoacyl-tRNA synthetase (aaRSs) in E. coli. While no hit was obtained to decode UAGC codon, we still examined the decoding efficiency of
variants that were examined (Fig. 3). This result indicates that these tRNA mutants can only be charged with Boc-Lys by BocLysRS and cannot be charged with natural amino acids by BocLysRS or any endogenous aminoacyl-tRNA synthetase (aaRSs) in E. coli. While no hit was obtained to decode UAGC codon, we still examined the decoding efficiency of 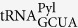 -wt against UAGC codon and no detectable decoding was observed.
-wt against UAGC codon and no detectable decoding was observed.
Figure 3. GFPUV fluorescence assays of cells expressing 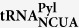 variants.
variants.
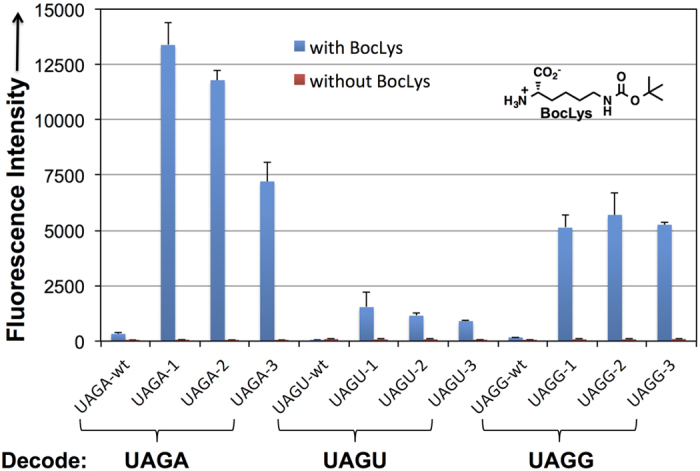
Fluorescence readings of E. coli C321.ΔA cells expressing  -wt or the evolved
-wt or the evolved  mutants, each coexpressed with BocLysRS and corresponding GFPUV-Asn149UAGN. The expressions were conducted either in the presence or in the absence of 5 mM Boc-Lys. Fluorescence intensity was normalized to cell growth. Each data point is the average of triplicate measurements with standard deviation.
mutants, each coexpressed with BocLysRS and corresponding GFPUV-Asn149UAGN. The expressions were conducted either in the presence or in the absence of 5 mM Boc-Lys. Fluorescence intensity was normalized to cell growth. Each data point is the average of triplicate measurements with standard deviation.
We further confirmed the incorporation of Boc-Lys into GFPUV-Asn149UAGN by mass spectrometry after SDS-PAGE separation and trypsin digest. As shown in the mass spectra (Figs S2, S3 and S4 in the Supporting Information), two mass peaks were observed in each sample. The (M + 2 H)2+ peak corresponds to the peptide fragment (141-LEYNYNSHBoc-LysVYITADK) from GFPUV that contains an intact Boc-Lys residue at position 149. The (M + 3 H)3+ peak corresponds to the peptide fragment that contains an lysine residue at position 149. Since the BocLysRS cannot charge  variants with lysine according to both literature report41 and our data (Fig. S5), the observed peptide that contains lysine at position 149 must be derived from the cleavage of the Boc group under the mass spectrometry conditions. The carbamate cleavage of Boc-Lys was also observed previously with electron spray ionization process in the literature42,43. Overall, our data confirmed that there was no undesirable incorporation of natural amino acids. The yields of the mutant GFPUV proteins were 54, 9, and 21 mg/L after partial purification by affinity chromatography when tRNA mutants UAGA-1, UAGU-1, and UAGG-2 were used to decode UAGA, UAGU, and UAGG codons, respectively.
variants with lysine according to both literature report41 and our data (Fig. S5), the observed peptide that contains lysine at position 149 must be derived from the cleavage of the Boc group under the mass spectrometry conditions. The carbamate cleavage of Boc-Lys was also observed previously with electron spray ionization process in the literature42,43. Overall, our data confirmed that there was no undesirable incorporation of natural amino acids. The yields of the mutant GFPUV proteins were 54, 9, and 21 mg/L after partial purification by affinity chromatography when tRNA mutants UAGA-1, UAGU-1, and UAGG-2 were used to decode UAGA, UAGU, and UAGG codons, respectively.
Incorporation of other unAAs using the evolved tRNAs
Structural data shows the lack of direct interaction between PylRS and the ALS region of 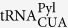 . Therefore, evolved
. Therefore, evolved 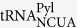 mutants in theory can be charged by PylRS mutants with unAAs other than Boc-Lys. To demonstrate the general applicability of evolved
mutants in theory can be charged by PylRS mutants with unAAs other than Boc-Lys. To demonstrate the general applicability of evolved 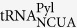 mutants, we randomly selected two PylRS variants for the incorporation of 3′-azibutyl-N-carbamoyl-lysine44 (AbK) and o-nitrobenzyl-oxycarbonyl-Nε-L-lysine45 (ONBK) in this study. We observed excellent selectivity when tRNA mutants were used to decode UAGN codons (AbK, Fig. S6; ONBK, Fig. S7). The relative incorporation efficiencies of Boc-Lys, Abk, and ONBK in response to either UAGN codons or UAG codon are consistent (Figs 3, S6, S7, and S8). The above results further confirmed that ASL mutations in tRNAPyl do not affect its recognition by PylRS mutants or the synthetases binding specificity towards their unAA substrates. Therefore, the evolved tRNAs can be potentially paired with any PylRS mutants for unAA incorporation, which is a significant augment of the current efforts in genetic code expansion.
mutants, we randomly selected two PylRS variants for the incorporation of 3′-azibutyl-N-carbamoyl-lysine44 (AbK) and o-nitrobenzyl-oxycarbonyl-Nε-L-lysine45 (ONBK) in this study. We observed excellent selectivity when tRNA mutants were used to decode UAGN codons (AbK, Fig. S6; ONBK, Fig. S7). The relative incorporation efficiencies of Boc-Lys, Abk, and ONBK in response to either UAGN codons or UAG codon are consistent (Figs 3, S6, S7, and S8). The above results further confirmed that ASL mutations in tRNAPyl do not affect its recognition by PylRS mutants or the synthetases binding specificity towards their unAA substrates. Therefore, the evolved tRNAs can be potentially paired with any PylRS mutants for unAA incorporation, which is a significant augment of the current efforts in genetic code expansion.
Analysis of the evolved tRNA mutants
We did not isolate and directly sequencing tRNA mutants. Instead, genes that encode the evolved tRNA mutants were sequenced. All tRNA-encoding genes were isolated and re-transformed into E. coli C321.ΔA for various characterizations throughout the study and consistent data were obtained. Therefore, we assume that no mutations of tRNA occurred during or after transcription and the actual tRNA sequences can be correctly deduced from their encoding genes. The observed mutations of the top three tRNAs from each of UAGA, UAGG, and UAGU selections are shown in Table 1. The most striking observation was that all of these tRNAs contained apparent UCUA (U at 33.5 position) anticodon no matter which UAGN codon they decode. In addition, some tRNAs that were selected against different UAGN codons have the same sequence, such as UAGA-2 and UAGU-3, UAGU-1 and UAGG-1, UAGU-2 and UAGG-3 (Table 1). We subsequently analysed a few less efficient tRNA variants from the selection, three, one, and one additional tRNA sequences were identified from UAGA, UAGU, and UAGG selections, respectively. As shown in Table S1, UAGU-4 and UAGG-4 have the same sequence while all UAGA hits (UAGA-4, UAGA-5, and UAGA-6) are unique. According to our data, all tRNA mutants from UAGU selection could be found from UAGA and/or UAGG selections. A large number of tRNA mutants from UAGU and UAGG selections have sequence overlap. On the other hand, most of tRNA mutants from UAGA selections are unique. Overall, the above results implicate that a fourth base-pairing between anticodon and codon is not essential for UAGN decoding.
Table 1. Evolved
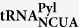 variants with improved UAGN decoding activity. Sequences of each
variants with improved UAGN decoding activity. Sequences of each
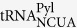 variant at randomized positions are listed.
variant at randomized positions are listed.
| Codon | tRNA variants | Positions |
||||
|---|---|---|---|---|---|---|
| 29–31 | 32,33 | 33.5 | 37, 38 | 39–41 | ||
| UAGA | UAGA-wt | G G A | C U | U | A A | U C C |
| UAGA-1 | G G G | C U | U | A U | C C U | |
| UAGA-2 | U G G | A U | U | A C | C U U | |
| UAGA-3 | C G G | A U | U | A C | C U U | |
| UAGU | UAGU-wt | G G A | C U | A | A A | U C C |
| UAGU-1 | U G G | C U | U | A U | C U U | |
| UAGU-2 | A G G | C U | U | A U | C U U | |
| UAGU-3 | U G G | A U | U | A C | C U U | |
| UAGG | UAGG-wt | G G A | C U | C | A A | U C C |
| UAGG-1 | U G G | C U | U | A U | C U U | |
| UAGG-2 | G G G | C U | U | A U | C U U | |
| UAGG-3 | A G G | C U | U | A U | C U U | |
The sequencing results also revealed three additional features of the evolved tRNA mutants: (1) no mutation was observed for the two nucleotides that are immediately 5′- or 3′-adjacent (position 33 and 37; Table 1 and Table S1) to the hypothetical four-base anticodon (NCUA). These two positions are likely essential for the function of 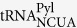 -derived mutants in E. coli; (2) all evolved tRNA mutants contain the same A31G and U39C mutations (Table 1 and Table S1). This led to the replacement of the A31-U39 with a G31-C39 base pair, which likely strengthens the part of the anticodon stem that is directly adjacent to the anticodon loop; (3) most of the evolved tRNA mutants do not have Watson-Crick base pairing at either position 29–41 or position 30–40 (Table 1 and Table S1). These mutations likely lead to structural changes in the anticodon and ASL, which allow a favorable conformational adjustment of tRNA upon its binding to the A site and subsequent translocation to the P and E sites of the ribosome.
-derived mutants in E. coli; (2) all evolved tRNA mutants contain the same A31G and U39C mutations (Table 1 and Table S1). This led to the replacement of the A31-U39 with a G31-C39 base pair, which likely strengthens the part of the anticodon stem that is directly adjacent to the anticodon loop; (3) most of the evolved tRNA mutants do not have Watson-Crick base pairing at either position 29–41 or position 30–40 (Table 1 and Table S1). These mutations likely lead to structural changes in the anticodon and ASL, which allow a favorable conformational adjustment of tRNA upon its binding to the A site and subsequent translocation to the P and E sites of the ribosome.
Cross-decoding among UAGN codons
Three top individual tRNA mutants, UAGA-1, UAGU-1, and UAGG-2, from UAGA, UAGU, and UAGG selection, respectively, were cross-tested against codons that they were not selected against. To simplify the cross-test, a series of plasmids, pGFPUV-UAGN-BocLysRS (N = A, G, or U), were constructed. Each plasmid contains a BocLysRS gene (expressed under the control of a glnS promoter) and a GFPUV mutant containing one of the UAGN (N = A, G, or U) codons. The decoding efficiencies of tRNA mutants toward each UAGN codon are correlated with the intensity of GFP fluorescence. As shown in Fig. 4, while the evolved tRNAs were among the best against the codons they were selected for, significant cross-activities were observed. Again, these results indicate that the fourth base-pairing is not essential for UAGN decoding. In addition, none of the obtained tRNA mutants could efficiently decode UAGC codon (data not shown).
Figure 4. Cross-decoding among UAGN codons.
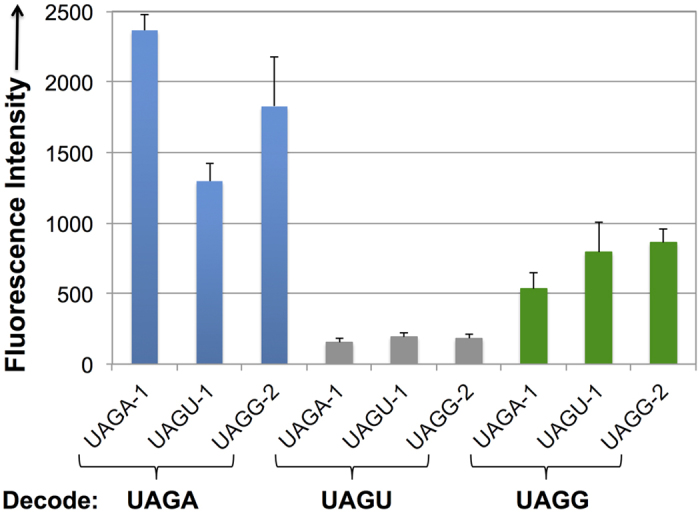
Fluorescence readings of E. coli C321.ΔA cells expressing the GFP reporter GFPUV-Asn149UAGN, BocLysRS (in plasmid pGFPUV-UAGN-BocLysRS), and tRNA mutants (in plasmid pBK-UAGN-X; X = hit number) in the presence of 5 mM Boc-Lys. Fluorescence intensity was normalized to cell growth. Each data point is the average of triplicate measurements with standard deviation.
tRNA (with regular 7-base anticodon loop) library construction and selection for UAGN (N = A, G, U, C) decoding
Since the fourth base-pairing of anticodon-codon interaction is apparently not essential for UAGN decoding, we next examined if tRNA variants with regular 7-base anticodon loop could decode UAGN codons. To this end, we constructed a tRNA library (Library-CUA10, theoretical diversity 1.0 × 106; actual library size 8.7 × 106) based on 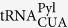 in which four nucleotides of the anticodon loop (32, 33, 37, 38; Fig. 2) and six nucleotides of the anticodon stem (29, 30, 31, 39, 40, 41; Fig. 2) were randomized. The resulting tRNA library was subsequently subjected to positive selections against UAGA, UAGG, UAGU, and UAGC, respectively, employing the same procedure as used in the Library-NCUA11 selection. No hit was obtained from these selections. The result indicates that an extra nucleotide in the anticodon loop is essential for UAGN decoding.
in which four nucleotides of the anticodon loop (32, 33, 37, 38; Fig. 2) and six nucleotides of the anticodon stem (29, 30, 31, 39, 40, 41; Fig. 2) were randomized. The resulting tRNA library was subsequently subjected to positive selections against UAGA, UAGG, UAGU, and UAGC, respectively, employing the same procedure as used in the Library-NCUA11 selection. No hit was obtained from these selections. The result indicates that an extra nucleotide in the anticodon loop is essential for UAGN decoding.
Cross-decoding against UAG codons
Three top tRNA mutants, UAGA-1, UAGU-1, and UAGG-2, from UAGA, UAGU, and UAGG selection, respectively, were examined for UAG codon decoding. To do this, a plasmid, pGFPUV-UAG-BocLysRS, was constructed and the expression of GFP (contains an amber mutation at position 149) was examined in the presence of UAGA-1, UAGU-1, or UAGG-2. As shown in Fig. 5, UAGA-1 and UAGG-2 displayed significantly higher activity toward UAGA and UAGG codons, respectively, than to UAG codon. On the other hand, UAGU-1 displayed similar activity toward UAGU and UAG codons, which might due to the low decoding efficiency of UAGU-1 against UAGU codon. All three tRNA mutants showed similar and significantly diminished UAG decoding efficiency (~20 fold lower) than that of  .
.
Figure 5. UAGN and UAG codon decoding efficiency of evolved tRNA variants.
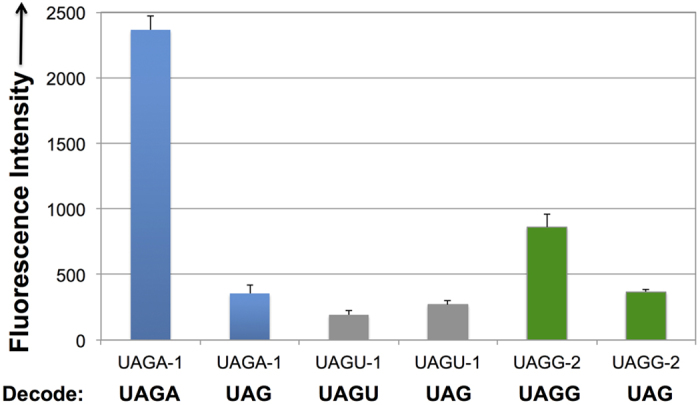
Fluorescence readings of E. coli C321.ΔA cells expressing the GFP reporter GFPUV-Asn149UAGN (or GFPUV-Asn149UAG), BocLysRS (in plasmid pGFPUV-UAGN-BocLysRS), and tRNA mutants (in plasmid pBK-UAGN-X; X = hit number) in the presence of 5 mM Boc-Lys. Fluorescence intensity was normalized to cell growth. Each data point is the average of triplicate measurements with standard deviation.
Discussion
This work represents the first example of a systematic evolution and study of tRNAs that decode a quartet of quadruplet codons (UAGN, N = A, U, G, or C) in a genomically recoded host with a clean background. The lack of endogenous UAG nonsense codons eliminates any in-frame UAGN sequences, which rules out the possibility of missing highly efficient UAGN-decoding tRNAs that might be cytotoxic due to undesirable reading-through of stop signals. Utilization of this host strain ensures that the most efficient UAGN-decoding tRNA variants can be identified. We targeted the evolution and study of tRNAs that are derived from the same ancestor, which enables informative and reliable comparison among obtained tRNAs. By introducing mutations into the ASL of parent tRNAs, we identified  variants that displayed significant improvement in UAGN decoding efficiency in comparison to their corresponding parents. The best hit (UAGA-1) showed 55-fold improvement in UAGA decoding efficiency than that of
variants that displayed significant improvement in UAGN decoding efficiency in comparison to their corresponding parents. The best hit (UAGA-1) showed 55-fold improvement in UAGA decoding efficiency than that of  -wt. Several sequence convergences were observed among the evolved tRNAs, which likely shift their interaction favorably with other components of the translational machinery for higher efficiency in quadruplet codon decoding. The observed improvement could also be partially attributed to the lack of competition from amber UAG codon-recognizing release factor-1 in the host strain. In addition to Boc-Lys, we showed that the evolved tRNA variants could be used to incorporate AbK and ONBK as well. It is likely that the evolved tRNA variants can be applied to the genetic incorporation of a range of unAAs using PylRS mutants in response to UAGN codons.
-wt. Several sequence convergences were observed among the evolved tRNAs, which likely shift their interaction favorably with other components of the translational machinery for higher efficiency in quadruplet codon decoding. The observed improvement could also be partially attributed to the lack of competition from amber UAG codon-recognizing release factor-1 in the host strain. In addition to Boc-Lys, we showed that the evolved tRNA variants could be used to incorporate AbK and ONBK as well. It is likely that the evolved tRNA variants can be applied to the genetic incorporation of a range of unAAs using PylRS mutants in response to UAGN codons.
Our data suggest that an extra nucleotide in the anticodon loop of tRNA is essential for high-efficiency decoding of UAGN codons within the natural translational machinery. However, the complete four-nucleotide Watson-Crick base-pairing between anticodon and codon is apparently not required for UAGN decoding. In fact, the best tRNA hit that read UAGG and UAGU codons do not have the potential to form four-nucleotide Watson-Crick base-pairing with these codons. In addition, cross activities were observed among tRNA mutants that were selected against different UAGN codons. These results implicated that reading of quadruplet codons is less likely to happen in the A site, which is consistent with the two recent structural investigations on the ASLs of frameshift suppressor tRNAs (tRNASufJ and tRNASufA6) bound to the Thermus thermophiles 70S A site30,46. These two structural studies suggested that +1 frameshift suppressor tRNAs do not form four-nucleotide Watson-Crick base-pairing with codons. Instead, ASLSufJ and ASLSufA6 form a cognate and a near cognate triplet codon–anticodon interaction in the A site, respectively. It is likely that the evolved 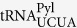 variants could engage a similar cognate or near cognate triplet interaction with the first three bases of UAGN sequences. Such cognate or a near cognate triplet codon–anticodon interactions would sufficiently lead to conformational changes of three bases, A1492, A1493, and G530, in 16S rRNA at ribosome A site47 and allow tRNAs to pass this critical checkpoint for decoding specificity. Since UAGN decoding does not require suppressor tRNA to occupy all four bases of the codon at the decoding center of A site, the overall and apparent quadruplet UAGN codon decoding process may be more accurately described as the induction of +1 frameshift by the evolved
variants could engage a similar cognate or near cognate triplet interaction with the first three bases of UAGN sequences. Such cognate or a near cognate triplet codon–anticodon interactions would sufficiently lead to conformational changes of three bases, A1492, A1493, and G530, in 16S rRNA at ribosome A site47 and allow tRNAs to pass this critical checkpoint for decoding specificity. Since UAGN decoding does not require suppressor tRNA to occupy all four bases of the codon at the decoding center of A site, the overall and apparent quadruplet UAGN codon decoding process may be more accurately described as the induction of +1 frameshift by the evolved 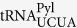 variants.
variants.
While both ours and others’ results indicate that direct quadruplet codon recognition does not happen in the A site, a question remains on how +1 frameshift happens. In the slippery model7,27,28,29, a re-pairing of tRNA to the +1 frame of mRNA (Figs 1B and S9A) occurs after the translocation of tRNA–mRNA complex from A to P site. The tRNA–mRNA re-pairing was suggested to be initiated by the P-site interaction between ribosome and the ASL of tRNA27. A recent structural study showed that the insertion of G37.5 in tRNASufA6 destabilized the non-canonical U32–A38 base pair, which likely affected EF-G-dependent translocation and therefore facilitated +1 frameshifting30. The insertion of an extra nucleotide-33.5 into 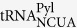 likely impaired the non-canonical C32-A38 interaction in
likely impaired the non-canonical C32-A38 interaction in 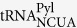 in a similar way and led to +1 frameshift. In addition, all of our tRNA hits contain either an A38U or an A38C mutation, which likely changed the finely tuned nucleotide identity of the 32–38 pair and influenced the decoding process48,49.
in a similar way and led to +1 frameshift. In addition, all of our tRNA hits contain either an A38U or an A38C mutation, which likely changed the finely tuned nucleotide identity of the 32–38 pair and influenced the decoding process48,49.
Although our data on UAGN decoding are more consistent with the slippery model7,27,28,29, this model cannot explain why a much higher decoding efficiency was observed for UAGA codon by the evolved 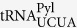 variants that have potential to form a four-nucleotide Watson-Crick base-pairing with the UAGA codon. In addition, the evolved
variants that have potential to form a four-nucleotide Watson-Crick base-pairing with the UAGA codon. In addition, the evolved 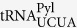 variants do not have an alternative cognate or near-cognate site for re-pairing in the P site (Fig. S9B), which is a hallmark of the slippery model. Even re-pairing would happen, we would observe that the efficiency of UAGG decoding would be higher than that of UAGA decoding since the re-pairing of the triplet anticodon of
variants do not have an alternative cognate or near-cognate site for re-pairing in the P site (Fig. S9B), which is a hallmark of the slippery model. Even re-pairing would happen, we would observe that the efficiency of UAGG decoding would be higher than that of UAGA decoding since the re-pairing of the triplet anticodon of 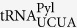 is energetically more favored for UAGG. However, we observed the opposite. Here we propose a modified slippery model for UAGN decoding (Fig. 6). The tRNA rearrangement in the P site does not necessarily involve a slippage event. It is possible for a suppressor tRNA to interact with all four bases of a quadruplet sequence in the P site and facilitate +1 frameshifting. In the cases of UAGN decoding, the extra U33.5 in
is energetically more favored for UAGG. However, we observed the opposite. Here we propose a modified slippery model for UAGN decoding (Fig. 6). The tRNA rearrangement in the P site does not necessarily involve a slippage event. It is possible for a suppressor tRNA to interact with all four bases of a quadruplet sequence in the P site and facilitate +1 frameshifting. In the cases of UAGN decoding, the extra U33.5 in 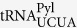 may engage certain level of interaction with the fourth base of the UAGN sequence, which creates steric interference with the incoming tRNA in the A site and induces +1 frameshift. This hypothesis is consistent with our observations on the efficiencies of UAGN suppression. The U33.5-A interaction in UAGA decoding is the most favorable one and led to the highest decoding efficiency. The U33.5-G interaction in UAGG decoding is also favorable and led to the second highest decoding efficiency. On the other hand, the U33.5-U interaction in UAGU decoding and the U33.5-C interaction in UAGC decoding are less favorable and led to the lowest decoding efficiency. In fact, we did not obtain a hit that is able to suppress UAGC codon. Other mutations in ASL of
may engage certain level of interaction with the fourth base of the UAGN sequence, which creates steric interference with the incoming tRNA in the A site and induces +1 frameshift. This hypothesis is consistent with our observations on the efficiencies of UAGN suppression. The U33.5-A interaction in UAGA decoding is the most favorable one and led to the highest decoding efficiency. The U33.5-G interaction in UAGG decoding is also favorable and led to the second highest decoding efficiency. On the other hand, the U33.5-U interaction in UAGU decoding and the U33.5-C interaction in UAGC decoding are less favorable and led to the lowest decoding efficiency. In fact, we did not obtain a hit that is able to suppress UAGC codon. Other mutations in ASL of 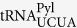 mutants may augment such interaction between tRNA and UAGN sequences by changing the conformation of engineered tRNAs. While this new model may not be suitable to explain all other +1 frameshifting cases, it provides reasonable molecular details on +1 frameshifting involving UAGN sequences.
mutants may augment such interaction between tRNA and UAGN sequences by changing the conformation of engineered tRNAs. While this new model may not be suitable to explain all other +1 frameshifting cases, it provides reasonable molecular details on +1 frameshifting involving UAGN sequences.
Figure 6. A new model for quadruplet decoding (+1 frameshift).

This model features a triplet translocation followed by a quadruplet anticodon-codon interaction in the P site of ribosome.
Our current data does not pinpoint the reason for the sequence convergence to U at position 33.5 in evolved tRNAs for UAGN suppression. It is possible that only U33.5 could preserve a characteristic U-turn structure of the anticodon loop that is essential for tRNA translocation50,51. Either U33 or U33.5 interact with A37 (no mutation) or U38/C38 (A38 in the wild-type) at other side of the anticodon loop. Future structural studies would be needed to obtain more defined answers. Besides conformational factors discussed above, posttranscriptional modifications of tRNA could also play a role. The primary sequence of wild-type and evolved pyrrolysyl tRNAs may not accurately reflect the anticodon base-pairing capability for quadruplet decoding. This is because extensive and chemically diverse nucleoside modifications fine-tune the biophysical and biochemical properties of transcribed tRNA molecules52,53. In addition, certain tRNA modifications influence reading frame maintenance of the ribosome and may facilitate +1 frameshift54,55. The insertion of U33.5 may impair the normal posttranscriptional modifications of tRNA and induces reading frame shift. Currently, we are investigating the posttranscriptional modification of M. maize tRNA in E. coli host and the implication in UAGN suppression.
In summary, we have evolved a collection of tRNA mutants that are able to incorporate unAAs in response to quadruplet UAGN sequences. Our work is an augment to the current efforts in genetic code expansion. While, theoretically, a quadruplet codon table provides 256 blank codons, our data suggest that cross-recognition among quadruplet codons may significantly reduce the number of usable quadruplet codons that are intended to be used simultaneously for unAA incorporation. We are currently examining whether we are able to engineer orthogonal tRNA variants for different UAGN sequences in order to truly allow a significant expansion of the genetic code using quadruplet suppression mechanism. The present study also represents a significant step toward the elucidation of the quadruplet suppression mechanism with tRNAs bearing extended anticodon loop. Unlike most of the previous studies that only focused on limited number of +1 frameshift suppressor tRNAs, this study involves a collection of tRNAs that decode a set of UAGN codons. The inferences from this work will likely bring better understanding to how the ribosome maintains the reading frame in the future.
Methods
Materials and General Methods
Boc-Lys was purchased from Bachem. Primers were ordered from Sigma. Restriction enzymes, Antarctic phosphatase (AP) and T4 DNA ligase were purchased from New England Biolabs. KOD hot start DNA polymerase was purchased from EMD Millipore. Protein mass spectrometric data was collected on a Waters Synapt® G2 mass spectrometer. Data was processed using MasslynxTM software (Waters). Standard molecular biology techniques56 were used throughout. Site-directed mutagenesis was carried out using overlapping PCR. E. coli GeneHogs were used for routine cloning and DNA propagation. E. coli C321.ΔA.exp (Addgene) and C321.ΔA (Addgene) were used for quadruplet decoding tRNA selection and evaluation. All solutions were prepared in deionized water further treated by Barnstead Nanopure® ultrapure water purification system (Thermo Fisher Scientific Inc). Antibiotics were added where appropriate to following final concentrations: ampicillin, 100 mg L−1; kanamycin, 50 mg L−1; tetracycline, 12.5 mg L−1.
Plasmid construction
Plasmid pBK-BocLysRS was constructed by inserting BocLysRS-encoding gene behind the constitutive glnS promoter (PglnS) on pBK vector57.
Plasmid pREP-BocLysRS-UAGN (for positive selection) was constructed by modifying plasmid pRepCM12b58. Specifically, the UAG codon (position 98) in the chloramphenicol acetyltransferase-encoding gene on pRepCM12b was changed to UAGN by site-directed mutagenesis. The primers that were used for mutagenesis can be found in supporting information. The resulting plasmid, pRepCM12b-UAGN, was digested with XbaI and ligated to a DNA fragment containing PglnS-BocLysRS cassette, which was amplified from template pBK-BocLysRS, to yield plasmid pREP-BocLysRS-UAGN. Individual plasmids with four different quadruplet codons, pREP-BocLysRS-UAGA, pREP-BocLysRS-UAGG, pREP-BocLysRS-UAGU, and pREP-BocLysRS-UAGC, were isolated from single E. coli colonies and verified by DNA sequencing.
Plasmids pGFPUV-NCUA-wt (N = A, G, U, or C) were obtained by first introducing the desirable UAGN mutation into plasmid pLei-GFPUV-Asn149UAG58 by overlapping PCR. The primers that were used for mutagenesis can be found in supporting information. The resulting plasmid, pGFPUV-UAGN, was subsequently modified by replacing the original tRNA with 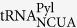 -wt and yielded plasmids, pGFPUV-NCUA-wt (N = A, G, U, or C).
-wt and yielded plasmids, pGFPUV-NCUA-wt (N = A, G, U, or C).
Variants of pGFPUV-NCUA-X (N = A, G, U, or C; X = hit number) were constructed by replacing the wild-type 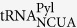 -wt on pGFPUV-NCUA-wt with evolved tRNA mutants.
-wt on pGFPUV-NCUA-wt with evolved tRNA mutants.
Plasmids pGFPUV-UAGN-BocLysRS (N = A, G, U, or C) were obtained by deleting tRNA fragment and inserting DNA fragment containing PglnS-BocLysRS cassette into PstI and SpeI sites of plasmids pGFPUV-UCUA-wt. The PglnS-BocLysRS cassette was amplified from template pBK-BocLysRS. Individual plasmids, each of which contains a specific UAGN codon of interest, were isolated from single E. coli colonies and verified by DNA sequencing.
Plasmids pGFPUV-UAG-BocLysRS was obtained by deleting the tRNA fragment and inserting DNA fragment containing PglnS-BocLysRS cassette into PstI and SpeI sites of plasmids pLei-GFPUV-Asn149UAG58. The PglnS-BocLysRS cassette was amplified from template pBK-BocLysRS.
tRNA library construction
Procedures for library construction were adapted from Noren and Noren59. Mutants of tRNA were obtained by overlapping PCR using pBK-mmPylT45 as the template. Resulting PCR products contain DNA cassettes of lpp promoter-tRNA mutant-rrnC terminator. Digestion of PCR products with NcoI and XhoI followed by ligation between NcoI and XhoI sites of pBK vector resulted in the tRNA library. The primers that were used for library construction can be found in supporting information.
Positive selection
Library DNAs were transformed into E. coli C321.ΔA.exp or C321.ΔA electrocompetent cells containing plasmid pREP-BocLysRS-UAGN (N = A, G, U, or C). Transformants were cultivated in LB media containing kanamycin and tetracycline. After 12 h of cultivation, cells were harvested. Based on calculation, a certain number of cells (>4.6× the size of the library) were plated on LB agar containing kanamycin, tetracycline, Boc-Lys (5 mM), and chloramphenicol (concentrations range from 34 mg L−1 to 75 mg L−1). The selection plates were incubated at 37 °C (C321.ΔA.exp) or 30 °C (C321.ΔA) for 48 h. Selected numbers of single colonies were further screened by replication onto plates with certain concentrations (50, 75, or 100 μg mL−1) of chloramphenicol in the presence and absence of Boc-Lys. Only the ones that grew in the presence of Boc-Lys and did not grew in the absence of Boc-Lys were selected for further evaluation.
Fluorescence analysis of bacterial culture
E. coli C321.ΔA strain harboring plasmid pBK-BocLysRS and a pGFPUV-NCUA variant (wt or evolved tRNA mutant) was cultured in 1 mL LB media containing ampicillin, kanamycin, and chloramphenicol at 30 °C. After 18 h cultivation, 50 μL culture was sub-cultured in 1 mL LB medium containing ampicillin, kanamycin, chloramphenicol, IPTG (0.1 mM), and 5 mM Boc-Lys (or 5 mM Lys, or none). Following cultivation at 30 °C for an additional 16 h, 1 mL of cell culture were collected, washed, then resuspended in 1 mL of potassium phosphate buffer (50 mM, pH 7.4). The processed cells were directly used for fluorescence and cell density measurements using a SynergyTM H1 Hybrid plate reader (BioTek Instruments). The fluorescence of GFPUV was monitored at λEx = 390 nm and λEm = 510 nm. The cell density was estimated by measuring the sample absorbance at 600 nm. Values of fluorescence intensity were normalized to cell growth. The incorporations of AbK and ONBK followed the same procedure using previously reported PylRS mutants44,60. Reported data are the average of three biological replicates with standard deviations. The cross test among different UAGN codons were conducted using the plasmid combination of pBK-tRNA-hit and pGFPUV-UAGN-BocLysRS.
Protein expression and purification
E. coli C321.ΔA strain harboring plasmid pBK-BocLysRS and a pGFPUV-NCUA variant was cultured in 5 mL LB media containing kanamycin and chloramphenicol at 30 °C. After 18 hours of cultivation, 0.5 mL culture was sub-cultured in 50 mL medium. The protein expression was induced at the OD600nm of 0.6 by additions of IPTG (0.5 mM) and Boc-Lys (5 mM). Cells were collected by centrifugation at 5 000 g and 4 °C for 15 min. Harvested cells were resuspended in lysis buffer containing potassium phosphate (20 mM, pH 7.4), NaCl (150 mM) and imidazole (10 mM). Cells were subsequently disrupted by sonication. Cellular debris was removed by centrifugation (21 000 g, 30 min, 4 °C). The cell-free lysate was applied to Ni Sepharose 6 Fast Flow resin (GE Healthcare). Protein purification followed manufacture's instructions. Protein concentrations were determined by Bradford assay (Bio-Rad). Purified protein was isolated by SDS-PAGE and digested with trypsin prior to MS analysis.
Additional Information
How to cite this article: Wang, N. et al. Systematic Evolution and Study of UAGN Decoding tRNAs in a Genomically Recoded Bacteria. Sci. Rep. 6, 21898; doi: 10.1038/srep21898 (2016).
Supplementary Material
Acknowledgments
This work is supported by the New Faculty Startup Fund to J. G. from the Chemistry Department of University of Nebraska – Lincoln and by Grant 1R01AI111862 (to J. G.) from DHHS-NIH-NIAID. C321.ΔA.exp was a gift from George Church (Addgene plasmid #49018). C321.ΔA was a gift from George Church (Addgene plasmid #48998).
Footnotes
Author Contributions N.W. constructed tRNA library and plasmids, performed the selection and characterization of the hits. X.S. synthesized amino acids. R.C. performed mass spectrometry analyses. W.N. designed the study and helped to construct tRNA library and plasmids, and wrote the manuscript. J.G. designed the study and wrote the manuscript.
References
- Atkins J. F. & Bjoerk G. R. A gripping tale of ribosomal frameshifting: extragenic suppressors of frameshift mutations spotlight P-site realignment. Microbiol. Mol. Biol. Rev. 73, 178–210 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riddle D. L. & Roth J. R. Suppressors of frameshift mutations in Salmonella typhimurium. J. Mol. Biol. 54, 131–144 (1970). [DOI] [PubMed] [Google Scholar]
- Yourno J. & Kohno T. Externally suppressible proline quadruplet CCCU. Science 175, 650–652 (1972). [DOI] [PubMed] [Google Scholar]
- Riddle D. L. & Carbon J. Frameshift suppression: a nucleotide addition in the anticodon of a glycine transfer RNA. Nat. New Biol. 242, 230–234 (1973). [DOI] [PubMed] [Google Scholar]
- Sroga G. E., Nemoto F., Kuchino Y. & Bjoerk G. R. Insertion (sufB) in the anticodon loop or base substitution (sufC) in the anticodon stem of tRNAPro2 from Salmonella typhimurium induces suppression of frameshift mutations. Nucleic Acids Res. 20, 3463–3469 (1992). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cummins C. M., Donahue T. F. & Culbertson M. R. Nucleotide sequence of the SUF2 frameshift suppressor gene of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 79, 3565–3569 (1982). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaber R. F. & Culbertson M. R. Codon recognition during frameshift suppression in Saccharomyces cerevisiae. Mol. Cell. Biol. 4, 2052–2061 (1984). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bossi L. & Roth J. R. Four-base codons ACCA, ACCU and ACCC are recognized by frameshift suppressor sufJ. Cell 25, 489–496 (1981). [DOI] [PubMed] [Google Scholar]
- Bossi L. & Smith D. M. Suppressor sufJ: a novel type of tRNA mutant that induces translational frameshifting. Proc. Natl. Acad. Sci. USA 81, 6105–6109 (1984). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson J. C. et al. An expanded genetic code with a functional quadruplet codon. Proc. Natl. Acad. Sci. USA 101, 7566–7571 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taki M., Matsushita J. & Sisido M. Expanding the genetic code in a mammalian cell line by the introduction of four-base codon/anticodon pairs. ChemBioChem 7, 425–428 (2006). [DOI] [PubMed] [Google Scholar]
- Neumann H., Wang K., Davis L., Garcia-Alai M. & Chin J. W. Encoding multiple unnatural amino acids via evolution of a quadruplet-decoding ribosome. Nature 464, 441–444 (2010). [DOI] [PubMed] [Google Scholar]
- Rodriguez E. A., Lester H. A. & Dougherty D. A. In vivo incorporation of multiple unnatural amino acids through nonsense and frameshift suppression. Proc. Natl. Acad. Sci. USA 103, 8650–8655 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niu W., Schultz P. G. & Guo J. An expanded genetic code in mammalian cells with a functional quadruplet codon. ACS Chem. Biol. 8, 1640–1645 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C. C. & Schultz P. G. Adding new chemistries to the genetic code. Annu. Rev. Biochem. 79, 413–444 (2010). [DOI] [PubMed] [Google Scholar]
- Chin J. W. Reprogramming the genetic code. Science 336, 428–429 (2012). [DOI] [PubMed] [Google Scholar]
- Niu W. & Guo J. Expanding the chemistry of fluorescent protein biosensors through genetic incorporation of unnatural amino acids. Mol. BioSyst. 9, 2961–2970 (2013). [DOI] [PubMed] [Google Scholar]
- Curran J. F. & Yarus M. Reading frame selection and transfer RNA anticodon loop stacking. Science 238, 1545–1550 (1987). [DOI] [PubMed] [Google Scholar]
- Phelps S. S. et al. Translocation of a tRNA with an extended anticodon through the ribosome. J. Mol. Biol. 360, 610–622 (2006). [DOI] [PubMed] [Google Scholar]
- Walker S. E. & Fredrick K. Recognition and positioning of mRNA in the ribosome by tRNAs with expanded anticodons. J. Mol. Biol. 360, 599–609 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernhardt H. S. & Tate W. P. Evidence from glycine transfer RNA of a frozen accident at the dawn of the genetic code. Biol. Direct 3, 53 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill C. W., Combriato G., Steinhart W., Riddle D. L. & Carbon J. Nucleotide sequence of the GGG-specific glycine transfer ribonucleic acid of Escherichia coli and of Salmonella typhimurium. J. Biol. Chem. 248, 4252–4262 (1973). [PubMed] [Google Scholar]
- Moore B., Persson B. C., Nelson C. C., Gesteland R. F. & Atkins J. F. Quadruplet codons: implications for code expansion and the specification of translation step size. J. Mol. Biol. 298, 195–209 (2000). [DOI] [PubMed] [Google Scholar]
- Magliery T. J., Anderson J. C. & Schultz P. G. Expanding the genetic code: selection of efficient suppressors of four-base codons and identification of “shifty’’ four-base codons with a library approach in Escherichia coli. J. Mol. Biol. 307, 755–769 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson J. C., Magliery T. J. & Schultz P. G. Exploring the limits of codon and anticodon size. Chem. Biol. 9, 237–244 (2002). [DOI] [PubMed] [Google Scholar]
- Dunham C. M. et al. Structures of tRNAs with an expanded anticodon loop in the decoding center of the 30S ribosomal subunit. RNA 13, 817–823 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qian Q. et al. A new model for phenotypic suppression of frameshift mutations by mutant tRNAs. Mol. Cell 1, 471–482 (1998). [DOI] [PubMed] [Google Scholar]
- Pedersen S. Escherichia coli ribosomes translate in vivo with variable rate. EMBO J. 3, 2895–2898 (1984). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qian Q. & Bjork G. R. Structural alterations far from the anticodon of the tRNAGGGPro of Salmonella typhimurium induce +1 frameshifting at the peptidyl-site. J. Mol. Biol. 273, 978–992 (1997). [DOI] [PubMed] [Google Scholar]
- Maehigashi T., Dunkle J. A., Miles S. J. & Dunham C. M. Structural insights into +1 frameshifting promoted by expanded or modification-deficient anticodon stem loops. Proc. Natl. Acad. Sci. USA 111, 12740–12745 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naesvall S. J., Nilsson K. & Bjoerk G. R. The ribosomal grip of the peptidyl-tRNA is critical for reading frame maintenance. J. Mol. Biol. 385, 350–367 (2009). [DOI] [PubMed] [Google Scholar]
- Farabaugh P. J. & Bjork G. R. How translational accuracy influences reading frame maintenance. EMBO J. 18, 1427–1434 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallant J. et al. On the role of the starved codon and the takeoff site in ribosome bypassing in Escherichia coli. J. Mol. Biol. 342, 713–724 (2004). [DOI] [PubMed] [Google Scholar]
- Belcourt M. F. & Farabaugh P. J. Ribosomal frameshifting in the yeast retrotransposon Ty: tRNAs induce slippage on a 7 nucleotide minimal site. Cell 62, 339–352 (1990). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farabaugh P. J. Translational frameshifting: Implications for the mechanism of translational frame maintenance. Prog. Nucleic Acid Res. Mol. Biol. 64, 131–170 (2000). [DOI] [PubMed] [Google Scholar]
- Wang K. et al. Optimized orthogonal translation of unnatural amino acids enables spontaneous protein double-labeling and FRET. Nat. Chem. 6, 393–403 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lajoie M. J. et al. Genomically recoded organisms expand biological functions. Science 342, 357–360 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chatterjee A., Lajoie M. J., Xiao H., Church G. M. & Schultz P. G. A bacterial strain with a unique quadruplet codon specifying non-native amino acids. ChemBioChem 15, 1782–1786 (2014). [DOI] [PubMed] [Google Scholar]
- Nozawa K. et al. Pyrrolysyl-tRNA synthetase-tRNAPyl structure reveals the molecular basis of orthogonality. Nature 457, 1163–1167 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss R. B., Dunn D. M., Atkins J. F. & Gesteland R. F. Ribosomal frameshifting from −2 to +50 nucleotides. Prog. Nucleic Acid Res. Mol. Biol. 39, 159–183 (1990). [DOI] [PubMed] [Google Scholar]
- Yanagisawa T. et al. Multistep engineering of pyrrolysyl-tRNA synthetase to genetically encode Nϵ-(o-azidobenzyloxycarbonyl) lysine for site-specific protein modification. Chem. Biol. 15, 1187–1197 (2008). [DOI] [PubMed] [Google Scholar]
- Nguyen D. P. et al. Genetic encoding of photocaged cysteine allows photoactivation of TEV protease in live mammalian cells. J. Am. Chem. Soc. 136, 2240–2243 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmied W. H., Elsasser S. J., Uttamapinant C. & Chin J. W. Efficient multisite unnatural amino acid incorporation in mammalian cells via optimized Pyrrolysyl tRNA Synthetase/tRNA expression and engineered eRF1. J. Am. Chem. Soc. 136, 15577–15583 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ai H.-W., Shen W., Sagi A., Chen P. R. & Schultz P. G. Probing protein-protein interactions with a genetically encoded photo-crosslinking amino acid. ChemBioChem 12, 1854–1857 (2011). [DOI] [PubMed] [Google Scholar]
- Chen P. R. et al. A facile system for encoding unnatural amino acids in mammalian cells. Angew. Chem., Int. Ed. 48, 4052–4055 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fagan C. E., Maehigashi T., Dunkle J. A., Miles S. J. & Dunham C. M. Structural insights into translational recoding by frameshift suppressor tRNASufJ. RNA 20, 1944–1954 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steitz T. A. A structural understanding of the dynamic ribosome machine. Nat. Rev. Mol. Cell Biol. 9, 242–253 (2008). [DOI] [PubMed] [Google Scholar]
- Olejniczak M. & Uhlenbeck O. C. tRNA residues that have coevolved with their anticodon to ensure uniform and accurate codon recognition. Biochimie 88, 943–950 (2006). [DOI] [PubMed] [Google Scholar]
- Ledoux S., Olejniczak M. & Uhlenbeck O. C. A sequence element that tunes Escherichia coli tRNAAlaGGC to ensure accurate decoding. Nat. Struct. Mol. Biol. 16, 359–364 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phelps S. S., Jerinic O. & Joseph S. Universally conserved interactions between the ribosome and the anticodon stem-loop of A site tRNA important for translocation. Mol. Cell 10, 799–807 (2002). [DOI] [PubMed] [Google Scholar]
- Phelps S. S. & Joseph S. Non-bridging phosphate oxygen atoms within the tRNA anticodon stem-loop are essential for ribosomal A site binding and translocation. J. Mol. Biol. 349, 288–301 (2005). [DOI] [PubMed] [Google Scholar]
- Jackman J. E. & Alfonzo J. D. Transfer RNA modifications: nature's combinatorial chemistry playground. Wiley Interdiscip. Rev.: RNA 4, 35–48 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- El Y. B., Bailly M. & de Crecy-Lagard V. Biosynthesis and function of posttranscriptional modifications of transfer RNAs. Annu. Rev. Genet. 46, 69–95 (2012). [DOI] [PubMed] [Google Scholar]
- Bjork G. R. et al. Transfer RNA modification: influence on translational frameshifting and metabolism. FEBS Lett. 452, 47–51 (1999). [DOI] [PubMed] [Google Scholar]
- Yarian C. et al. Accurate translation of the genetic code depends on tRNA modified nucleosides. J. Biol. Chem. 277, 16391–16395 (2002). [DOI] [PubMed] [Google Scholar]
- Sambrook J. F. & Russell D. W. Editors. Molecular cloning: A laboratory manual, third edition. (Cold Spring Harbor Laboratory Press, 2000). [Google Scholar]
- Wang L., Brock A., Herberich B. & Schultz P. G. Expanding the genetic code of Escherichia coli. Science 292, 498–500 (2001). [DOI] [PubMed] [Google Scholar]
- Guo J., Melancon C. E. III, Lee H. S., Groff D. & Schultz P. G. Evolution of amber suppressor tRNAs for efficient bacterial production of proteins containing nonnatural amino acids. Angew. Chem., Int. Ed. 48, 9148–9151 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noren K. A. & Noren C. J. Construction of high-complexity combinatorial phage display peptide libraries. Methods 23, 169–178 (2001). [DOI] [PubMed] [Google Scholar]
- Lang K. et al. Genetic encoding of bicyclononynes and trans-cyclooctenes for site-specific protein labeling in vitro and in live mammalian cells via rapid fluorogenic diels-alder reactions. J. Am. Chem. Soc. 134, 10317–10320 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.