

Abstract
The enzymatic modification of aminoglycosides by aminoglycoside-acetyltransferases (AAC), aminoglycoside-adenyltransferases (AAD), and aminoglycoside-phosphotransferases (APH), is the most common resistance mechanism in P. aeruginosa and these enzymes can be coded on mobile genetic elements that contribute to their dispersion. One hundred and thirty seven P. aeruginosa isolates from the University Hospital, Cumana, Venezuela (HUAPA) were evaluated. Antimicrobial susceptibility was determined by the disk diffusion method and theaac, aadB and aph genes were detected by PCR. Most of the P. aeruginosa isolates (33/137) were identified from the Intensive Care Unit (ICU), mainly from discharges (96/137). The frequency of resistant P. aeruginosaisolates was found to be higher for the aminoglycosides tobramycin and amikacin (30.7 and 29.9%, respectively). Phenotype VI, resistant to these antibiotics, was the most frequent (14/49), followed by phenotype I, resistant to all the aminoglycosides tested (12/49). The aac(6´)-Ib,aphA1 and aadB genes were the most frequently detected, and the simultaneous presence of several resistance genes in the same isolate was demonstrated. Aminoglycoside resistance in isolates ofP. aeruginosa at the HUAPA is partly due to the presence of the aac(6´)-Ib, aphA1 andaadB genes, but the high rates of antimicrobial resistance suggest the existence of several mechanisms acting together. This is the first report of aminoglycoside resistance genes in Venezuela and one of the few in Latin America.
Keywords: Aminoglycosides, AME, Resistance, PCR
INTRODUCTION
The aminoglycosides tobramycin, gentamicin, and amikacin are commonly used to treat hospital-acquired infections caused by Pseudomonas aeruginosa. These infections generally require treatment with a combination of antimicrobials in order to achieve a greater bactericidal effect and reduce the levels of resistance1. The use of a combination of antimicrobials, however, is associated with resistance mediated by aminoglycoside modifying enzymes (AME). Four of these enzymes, encoded byaac(6')-I, aac(6')-II,ant(2'')-I, and aph(3')-VI, are of particular significance because they are among the most common modifying enzymes present in P. aeruginosa, and their substrates are the most important antipseudomonal aminoglycosides2. The spread of these enzymes can occur through genetic elements exchanged between the same or different taxa, and is favored by the selective pressures present in a hospital environment where there is a constant use of antimicrobial compounds. This has led to bacterial resistance which is becoming an increasing threat to public health3.
In recent years, there has been an increased interest in studying the mechanisms of resistance associated with the different antimicrobial families used in clinical practice. According to epidemiological studies, Latin America is one of the regions with the highest incidence of hospital-associated outbreaks produced by bacteria resistant to several antibiotics4 , 5. Different patterns of resistance to betalactamics, quinolones and aminoglycosides have been reported in Mexico6, Peru7, Brazil8, and Venezuela9 , 10 , 11, and in Latin American surveys12, showing an increase in resistance in theP. aeruginosa strains.
Genes resistant to aminoglycosides in this species have, however, only been reported in Europe, where aac(6´)-II and ant(2´´)-I are the most prevalent enzymes, and Korea where the most frequent enzymes areaph(3¢)-VI, ant(2´´)-I, andaac(6´)-I, acting either on their own or in combination1. In Mexico and Brazilaac(6´)-31/aadA1, and aadA2 have also been reported in Hospital-acquired strains13 , 14. In Venezuela there have been no studies of genes that confer resistance to aminoglycosides inP. aeruginosa. However, these genes have been reported in Enterobacteria, with aadA, aadB, and aac(6)-Iqgenes being the most frequently detected in a class I integron, in strains ofKlebsiella pneumoniae from the University HospitalAntonio Patricio de Alcala (HUAPA), in Cumana, Venezuela15. Resistance in clinically important gram-negative bacteria is a growing concern which in recent years has broken the within-hospital barrier and is now also affecting outpatients. Bearing this information in mind, we evaluated the in vitro antimicrobial resistance of hospital-acquired strains of P.aeruginosa isolated from HUAPA and identified their resistant genes to aminoglycosides. We hope that this will broaden our understanding of the different aminoglycoside resistance genes, and the mechanisms of resistance in theP. aeruginosa strains present in this hospital.
METHODS
We studied 137 different isolates of Pseudomonas aeruginosa, from patients with diagnosis of infection, attending different units within the HUAPA, from September 2010 to December 2011. For the identification of bacteria, the isolates were grown in Luria Bertani (LB) agar and taken to the Molecular Genetics Laboratory at the Research Institute of Biomedical and Applied Science "Dra. Susan Tai", Universidad de Oriente (IIBCAUDO), where they were kept until their analysis. Isolates were taken only from those patients who had signed a written consent form, after complete disclosure of the aims, risks, and scope of the research project was given. Treatment of patients, analysis of isolates, and information generated was conducted according to the bioethical and biosafety guidelines as stated by the Bioethics and Biosafety Commission of the IIBCAUDO (CoBioBios).
To define the presence of infection, we used the criteria established by the Spanish Society of Infectious Diseases and Clinical Microbiology (http://www.seimc.org/documentoscientificos.php?mn_MP=3&mn_MS=358). In addition, an infection was considered a Healthcare-associated Infection (HAI) if the date of detection of the site-specific infection, as defined by the US National Healthcare Safety Network (NHSN), had taken place after the 3rd day of admission to an inpatient. Otherwise, they were regarded as a community-acquired infection (the infection was already present on the day of admission).
Bacteriological Diagnosis
The viability and purity of the isolates was confirmed using standard culture and biochemical tests, according to the procedures and protocols for the identification of non-fermenting gram-negative bacilli17. Microbial susceptibility was determined by the disc diffusion method18 using the following antimicrobials (BD): amikacin (30 µg), gentamicin (10 µg), netilmicin (30 µg), tobramycin (30 µg), ciprofloxacin (5 µg), cefepime (30 µg), ceftazidime (30 µg), piperacillin (30 µg), piperacillin/tazobactam (100/10 µg), meropenem (10 µg), imipenem (10 µg), and aztreonam (30 µg), according to the guidelines proposed by the Clinical Laboratory Standard Institute for P. aeruginosa 19.
Molecular identification of aminoglycoside resistance genes
Bacterial genomic DNA was extracted using the Kit Wizard(r) Genomic (Promega) according to the manufacturer instructions for gram-negative bacteria. The genes coding for aminoglycoside resistance: aac(3)-IIa(aacC2-F/R 5´-actgtgatgggatacgcgtc-3´/5´-ctccgtcagcgtttcagcta-3´)20, aphA1 (aphA1-F/R 5´-atgggctcgcgataatgtc-3´/5´-ctcaccgaggcagttccat-3´) and aphA2(aphA2-F/R 5´-gaacaagatggattgcacgc-3´/5´-gctcttcagcaatatcacgg-3´)21, aac(6´)-Ib(aac(6´)-IbF/R 5´-taggagtggctaaatcgat-3´/5´-cccgctttctcgtagca-3´)22 and aadB (aadB-F/R 5´-cgtcatggaggagttggact-3´/5´-cgcaagacctcaaccttttc-3´)23 were detected using PCR. Klebsiella pneumoniae strains Kp01, which contains theaac(3´)-IIa gene; Kp28, which contains theaac(6´)-Ib and aph genes; and KpM7, which contains the aadB gene, were used as positive controls15. The amplification products were run on a 2% agarose gel, stained with ethidium bromide (0.5 µg/ml) and buffered in TBE 1X, for 30 minutes at 100 V.
RESULTS
From the total number of P. aeruginosa isolates obtained from different patients who were admitted to the HUAPA for various treatments, 83.2% (114/137) were identified from patients with hospital-acquired infections. These infections were defined when the clinical and epidemiological information of each individual showed that the patients stayed at least 72 hours after they were admitted. Most isolates were from patients treated in the Medicine A-B unit (n = 46), in the intensive care unit (ICU) (n = 33), and in the nursery (n = 10). The patients treated were hospitalized over a long period (between 55 and 250 days, mean 109 days) and were either immunosuppressed or in a critical condition.
The isolates were mainly from discharges, and were obtained from different parts of the body (96 isolates, 70.0%), followed by catheter tips (15 isolates), sputum (13 isolates), urine (7 isolates), blood (4 isolates), and peritoneal liquid (2 isolates). It should be mentioned that the identification of P. aeruginosa from catheter tips was only undertaken when the bacterium was also isolated from a blood sample of the same patient.
The resistance profiles of the P. aeruginosa isolates showed that 65.0% (89/137) were resistant to one or more antibiotics: 51.1% (70/137) were resistant to ciprofloxacin, 36.5% (50/137) to meropenem, 35.0% (48/137) to imipenem, 32.8% (45/137) to piperacillin, and 32.1% (44/137) to piperacillin-tazobactam (Fig. 1). In addition, 30.7% (42/137) and 29.9% (41/137) of the isolates showed resistance to the aminoglycosides tobramycin and amikacin, respectively. A total of 9 aminoglycoside resistance profiles (phenotypes) were identified in the isolates (Table 1), of which, the most frequent was the phenotype VI, which showed resistance to tobramycin and amikacin, and type I, which was resistant to all of the aminoglycosides assayed. Furthermore, 95.9% of the isolates with phenotype I also showed multidrug-resistant phenotypes.
Fig. 1. - Antimicrobial resistance in strains of P. aeruginosa isolated from patients at the University Hospital in Cumana, Venezuela. PIP: piperacillin, TZP: piperacillin/tazobactam, CAZ: ceftazidime, FEP: cefepime, ATM: aztreonam, IPM: imipenem, MEM: meropenem, GM: gentamicin, NN: tobramycin, NET: netilmicin, AK: amikacin, CIP: ciprofloxacin.
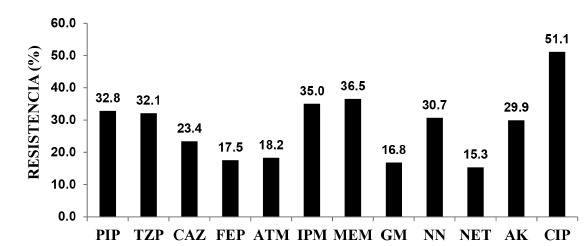
Table 1. Resistance phenotypes, number of strains (N) and resistance to aminoglycosides in P. aeruginosa strains isolated from patients at the University Hospital in Cumana, Venezuela.
| Phenotype | N | Resistance to Aminoglycosides |
|---|---|---|
| I | 12 | GM NN NET AK |
| II | 4 | GM NN NET |
| III | 7 | GM NN AK |
| IV | 4 | NN NET AK |
| V | 1 | GM NN |
| VI | 14 | NN AK |
| VII | 1 | NN |
| VIII | 1 | NET |
| IX | 5 | AK |
| Total | 49 | |
GM: gentamicin; NN: tobramicin; NET: netilmicin; AK: amikacin.
The most frequent aminoglycoside resistance gene was aac(6´)-Ib,detected in phenotypes I, II, III, IV, VI, and VII (Table 2), followed by aphA1 (phenotypes I, IX, and VIII) and aadB (phenotypes I and V). These genes were also shown to be simultaneously present in all of the phenotype I strains, and in three of the phenotype VI strains.
Table 2. Resistance phenotypes and amplified genes in Pseudomonas aeruginosa strains collected from patients at the University Hospital in Cumana, Venezuela.
| Phenotype | N | Amplified Gene | ||||
|---|---|---|---|---|---|---|
| aac(3´)-IIa | aac(6´)-Ib | aphA1 | aphA2 | aadB | ||
| I | 3 | - | X | X | - | X |
| I | 1 | - | - | X | X | X |
| I | 5 | - | - | X | - | X |
| I | 2 | - | X | - | - | X |
| I | 1 | - | X | - | X | X |
| II | 4 | - | X | - | - | - |
| III | 5 | - | X | - | - | - |
| IV | 3 | - | X | - | - | - |
| V | 1 | - | - | - | - | X |
| VI | 1 | X | - | - | - | - |
| VI | 3 | - | X | - | - | - |
| VI | 3 | - | X | - | X | - |
| VI | 2 | - | - | - | X | - |
| VII | 1 | - | X | - | - | - |
| VIII | 1 | - | - | X | - | - |
| IX | 3 | - | - | X | - | - |
| Total | 39 | 1 | 25 | 13 | 7 | 13 |
Nevertheless, the results of the aminoglycoside resistant phenotypes did not always coincide with those of the resistance genes detected. Thus, 10 of the isolates with phenotypic resistance to the aminoglycosides did not yield PCR amplification products for any of the genes evaluated in this study. Six isolates showed the presence of multidrug-resistant phenotypes (Table 3). Only two samples (phenotypes III and VII) came from outpatients (adult observation area). Isolates with phenotypes III and IV and 2 with phenotype VI showed a phenotype that suggests the presence of metallobetalactamases.
Table 3. Resistance phenotypes, hospital areas, sample type and resistance to antibiotics in P. aeruginosa strains that did not amplify the aminoglycoside modifying enzyme genes, isolated from patients at the University Hospital in Cumana, Venezuela.
| Phenotype | N | Hospital Area | Type of Sample | Resistance to Antibiotics |
|---|---|---|---|---|
| III | 1 | Medicine B | Discharges | GM NN AK CIP |
| 1* | Adult Obs. | Discharges | PIP TZP CAZ FEP IPM MEM GM NN AK CIP | |
| IV | 1* | Nursery | Blood | PIP TZP CAZ FEP IPM MEM NN NET AK |
| VI | 2* | ICU | Discharges | PIP TZP CAZ FEP IPM MEM NN AK CIP |
| 1* | Medicine A | Discharges | ||
| 1 | ICU | Discharges | PIP IPM MEM NN AK CIP | |
| VII | 1 | Adult Obs. | Urine | PIP TZP IPM MEM NN CIP |
| IX | 1 | Medicine A | Discharges | FEP IPM MEM AK CIP |
| 1 | Medicine B | Discharges | PIP TZP ATM IPM MEM AK CIP | |
* These isolates showed phenotypes of metallobetalactamases.
DISCUSSION
Pseudomonas aeruginosa has become an important pathogen, constituting 10 to 15% of hospital-acquired infections worldwide, especially in immunocompromised patients in ICUs24. These data agree with the findings of our study. Similar results have been reported in Mexico6 and in Caracas, Venezuela9. In addition, we found that 16.8% of community-associated infections were caused by P. aeruginosa. This is comparable to a study of patients from India suffering from acquired pneumonia in the community25, where P. aeruginosa was the second most frequent bacteria.
The high frequencies of P. aeruginosa isolates resistant to tobramycin and amikacin have also been reported in other health institutions. In Venezuela, in a study of Pseudomonas isolates collected in 30 different health institutions throughout the country during the years 1988-199826, the resistance shown to gentamicin was 27%, 19% to amikacin and 23% to tobramycin. In a collaborative study, where the resistance pattern of 586 isolates from Argentina, Brazil, Chile, Colombia, Costa Rica, Ecuador, Guatemala, Mexico, Panama, Peru, and Venezuela12 were analyzed, the average resistance found for gentamicin was 32.6%, 24.6% for amikacin and 29.9% for tobramycin. Others studies in Latin American countries have shown a frequency of resistant isolates between 27.8- 42.0% for gentamicin and 16.3- 28.9% to amikacin27 , 28 , 29. In Spain, resistance frequencies of 24.2% to gentamicin and 16.7% to amikacin have been reported30.
The inactivation of aminoglycosides by modifying enzymes is the most common resistance mechanism in gram-negative bacilli. This was corroborated in this study performed at HUAPA, where the most frequent genes detected wereaphA1, aadB and aac(6´)-Ib, with N-acetylation at the 6' position catalyzed by the last enzyme, being the most usual ways in which aminoglycosides are modified by P. aeruginosa 1 , 31.
This study has also demonstrated the simultaneous presence of several aminoglycoside resistance genes in all the phenotype I isolates, and in three of the phenotype VI strains evaluated. In Korea, the presence of aph(3')-VI,aad(2'')-I, and aac(6')-I genes has been reported in 250 isolates of P. aeruginosa 32, where the genes were found either alone or combined, withaac(6')-I + aad(2'')-I being the most frequent combination (24/36 isolates). In Iran, aac(6´)-II (36%);aad(2´´)-I (28%), aph(3´)-VI (11%), andaac(6´) (7%) genes have been reported in P. aeruginosa strains, highlighting the fact that up to four different genes have been found simultaneously in the same isolate2. In this sense, integrons have been reported in Brazil13 , 33 and Mexico14, carrying multiple copies of aminoglycoside resistance genes.
Several of the P. aeruginosa isolates showing phenotypic resistance to aminoglycosides evaluated in this study did not test positive for the resistance genes evaluated, this suggests that other genes, not tested here, which confer resistance to aminoglycosides, may be present in these isolates, as reported elsewhere2. Differences in the distribution of these enzymes could be derived from differences in the selective pressures on the bacterial population with respect to aminoglycoside utilization34. There are also other mechanisms that could lead to aminoglycoside resistance, that were not evaluated here, such as: changes in the permeability of the external membrane, active efflux systems, and alterations of the 30S ribosomal subunit conferred by mutations2 , 34 , 35. Active efflux systems have been demonstrated to be involved in the resistance of P. aeruginosa to aminoglycosides in several countries35 , 36. An explanation regarding the presence of resistance genes but not of phenotypic resistance, is the presence of mutations producing non-functional proteins and the lack of promoters in the upstream region where the genes are inserted37.
The multi-resistance observed in isolates that did not amplify the AME genes evaluated, were mainly obtained from inpatients undergoing long term treatments, and the individuals had received a combination of antimicrobials that could also be partly associated with the indiscriminate use of multiple treatment schemes with broad spectrum antimicrobials, favoring the increase of bacterial resistance caused by multiple mechanisms. However, the high frequency of resistance to different antimicrobials shown by the P. aeruginosa isolates from HUAPA suggests the existence of multiple resistance mechanisms acting together.
ACKNOWLEDGEMENTS
We would like to thank the staff of the Bacteriological Laboratory at the HUAPA: Medina Belkis, Martinez Dianny, Rodríguez Lucy, Barreto José Antonio, Caña Luisa, Garcia Jose, González Diorelis, Mundaray Maylim, and Peinado Veronica, for collecting the strains evaluated in this study: your contribution was invaluable. We also want to thank Frances Osborn for the translation and revision of the manuscript.
Footnotes
FINANCIAL SUPPORT This study was partially supported by the Mission Science Project Nº G-2007001442 "Development, adaption, validation, and transfer of technologies for molecular diagnosis of communicable diseases, through the Venezuelan Network, for identification and molecular diagnosis of microorganisms" National Fund for Science and Technology (FONACIT).
REFERENCES
- 1.Strateva T, Yordanov D. Pseudomonas aeruginosa a phenomenon of bacterial resistance. J Med Microbiol. 2009;58:1133–1148. doi: 10.1099/jmm.0.009142-0. [DOI] [PubMed] [Google Scholar]
- 2.Vaziri F, Peerayeh SN, Nejad QB, Farhadian A. The prevalence of aminoglycoside-modifying enzyme genes (aac (6')-I, aac (6')-II, ant (2")-I, aph (3')-VI) in Pseudomonas aeruginosa. Clinics(Sao Paulo) 2011;66:1519–1522. doi: 10.1590/S1807-59322011000900002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hall RM, Collis CM. Antibiotic resistance in gram-negative bacteria: the role of gene cassettes and integrons. Drug Resist Updat. 1998;1:109–119. doi: 10.1016/s1368-7646(98)80026-5. [DOI] [PubMed] [Google Scholar]
- 4.Casellas JM. Antibacterial drug resistance in Latin America: consequences for infectious disease control. Rev Panam Salud Publica. 2011;30:519–528. [PubMed] [Google Scholar]
- 5.Gales AC, Castanheira M, Jones RN, Sader HS. Antimicrobial resistance among Gram-negative bacilli isolated from Latin America: results from SENTRY Antimicrobial Surveillance Program (Latin America, 2008-2010) Diagn Microbiol Infect Dis. 2012;73:354–360. doi: 10.1016/j.diagmicrobio.2012.04.007. [DOI] [PubMed] [Google Scholar]
- 6.Camacho A, Acosta G, Rositas F, Canizález J. Resistencia antimicrobiana de Pseudomonas aeruginosa en un hospital de enseñanza del norte de México. Enf Infecc Microbiol Clin. 2007;27:44–48. [Google Scholar]
- 7.Luján-Roca D, Ibarra J, Mamani E. Resistencia a los antibióticos en aislados clínicos de Pseudomonas aeruginosa en un hospital universitario en Lima, Perú. Rev Biomed. 2008;19:156–160. [Google Scholar]
- 8.Machado G, Lago A, Fuentefria SR, Fuentefria DB. Occurrence and the susceptibility to antimicrobial agents in Pseudomonas aeruginosa and Acinetobacter sp: at a tertiary hospital in southern Brazil. Rev Soc Bras Med Trop. 2011;44:168–172. doi: 10.1590/s0037-86822011005000015. [DOI] [PubMed] [Google Scholar]
- 9.Andrade E, Navarro P, Rodríguez J, Rodríguez P, Villarroel E. Evaluación bacteriológica de Infecciones por Pseudomonas aeruginosa. Antibiot Infect. 2002;1:29–32. [Google Scholar]
- 10.Briceño I, Suarez M. Resistencia bacteriana en la Unidad de Cuidados Intensivos del Hospital Universitario de los Andes. MEDICRIT. 2006;3:30–42. [Google Scholar]
- 11.Ysacis J, Salazar J, Ollarves L, Girón C. Comparación de la sensibilidad bacteriana in vitro: amikacina versus netilmicina. Actual Infectol(Caracas) 2002;2:15–17. [Google Scholar]
- 12.Jones RN, Guzman-Blanco M, Gales AC, Gallegos B, Castro AL, Martino MD. Susceptibility rates in Latin American nations: report from a regional resistance surveillance program (2011) Braz J Infect Dis. 2013;17:672–681. doi: 10.1016/j.bjid.2013.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mendes RE, Castanheira M, Toleman MA, Sader HS, Jones RN, Walsh TR. Characterization of an integron carrying blaIMP-1 and a new aminoglycoside resistance gene, aac(6')-31, and its dissemination among genetically unrelated clinical isolates in a Brazilian hospital. Antimicrob Agents Chemother. 2007;51:2611–2614. doi: 10.1128/AAC.00838-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sánchez-Martinez G, Garza-Ramos UJ, Reyna-Flores FL, Gaytán-Martínez J, Lorenzo-Bautista IG, Silva-Sanchez J. In169, a new class 1 integron that encoded bla(IMP-18) in a multidrug-resistant Pseudomonas aeruginosa isolate from Mexico. Arch Med Res. 2010;41:235–239. doi: 10.1016/j.arcmed.2010.05.006. [DOI] [PubMed] [Google Scholar]
- 15.Guzmán M, Alonso G. Characterization of the variable region within class 1 integrons in Klebsiella pneumoniae nosocomials strains. Rev Med Chil. 2010;138:322–329. [PubMed] [Google Scholar]
- 16.Horan TC, Andrus M, Dudeck MA. CDC/NHSN surveillance definition of health care-associated infection and criteria for specific types of infections in the acute care setting. Am J Infect Control. 2008;36:309–332. doi: 10.1016/j.ajic.2008.03.002. [DOI] [PubMed] [Google Scholar]
- 17.Winn W, Allen S, Janda W, Koneman E, Procop G, Schreckenberger P, Woods G. Koneman´s color atlas and textbook of Diagnostic microbiology. 6. Philadelphia: Lippincoff Williams & Wilkins; 2006. [Google Scholar]
- 18.Bauer A, Kirby W, Sherris J, Turck M. Antibiotic susceptibility testing by a standardized single disk method. A J Clin Pathol. 1966;45:493–496. [PubMed] [Google Scholar]
- 19.Clinical and Laboratory Standards Institute . Performance standards for antimicrobial susceptibility testing. Wayne: CLSI; 2014. (Supplement M100-S20) [Google Scholar]
- 20.Van de Klundert JA, Vliegenthart JS. Persing DH, Smith TF, Tenover FC, White TJ. Diagnostic molecular epidemiology. Washington: American Society for Microbiology; 1993. PCR detection of genes coding for aminoglycoside-modifing enzymes; pp. 547–552. [Google Scholar]
- 21.Maynard C, Fairbrother J, Bekal S, Sanschagrin F, Levesque R, Brousseau R. Antimicrobial resistance genes in enterotoxigenic Escherichia coli O149:K91 isolates obtained over a 23-year period from pigs. Antimicrob Agents Chemother. 2003;47:3214–3221. doi: 10.1128/AAC.47.10.3214-3221.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Senda K, Arawaka Y, Ichiyama S, Nakashima K, Ito H, Ohsuka S. PCR detection of metallo-b-lactamase gene (blaIPM) in Gran-negative rods resistance to broad- spectrum b-lactamas. J Clin Microbiol. 1996;34:2904–2913. doi: 10.1128/jcm.34.12.2909-2913.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Díaz P, Bello H, Domínguez M, Trabal N, Mella S, Zemelman R. Resistencia a gentamicina, amikacina y ciprofloxacina en cepas hospitalarias de Klebsiella pneeumoniae productoras de b-lactamasas de espectro extendido. Rev Med Chile. 2004;132:1173–1178. doi: 10.4067/s0034-98872004001000003. [DOI] [PubMed] [Google Scholar]
- 24.Blanc D, Petignat C, Janin B, Bille J, Francioli P. Frequency and molecular diversity of Pseudomonas aeruginosa upon admission and during hospitalization: a prospective epidemiologic study. Clin Microbiol Infect. 1998;4:242–247. doi: 10.1111/j.1469-0691.1998.tb00051.x. [DOI] [PubMed] [Google Scholar]
- 25.Menon R, George A, Menon U. Etiology and anti-microbial sensitivity of organisms causing community acquired pneumonia: a single hospital study. J Family Med Prim Care. 2013;2:244–249. doi: 10.4103/2249-4863.120728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Brito A, Landaeta JM, Roldán Y, Marcano M, Santos JR, Guzmán M. Resistencia de Pseudomonas aeruginosa a la gentamicina, tobramicina amikacina en Venezuela. Bol Soc Ven Microbiol. 2000;20:42–45. [Google Scholar]
- 27.Perez F, Camejo L, Rojas E. Comportamiento de la resistencia antimicrobiana de gérmenes aislados en heridas por quemaduras. Rev Cubana Cir. 2009;48(3):6–6. [Google Scholar]
- 28.Robledo J, Calle D, Realpe T, Mejía G, Trujillo H, Marín G. Un programa de vigilancia de resistencia a antibióticos utilizando WHONET en seis hospitales de Medellín: resultados de los años 2001-2002. Infectio. 2003;7:107–107. [Google Scholar]
- 29.Villa LM, Cortés JA, Leal AL, Meneses A, Meléndez MP, GREBO Pseudomonas aeruginosa resistente a antimicrobianos en hospitales colombianos. Rev Chilena Infectol. 2013;30:605–610. doi: 10.4067/S0716-10182013000600005. [DOI] [PubMed] [Google Scholar]
- 30.Gamero M, García-Mayorgas A, Rodríguez F, Ibarra A, Casal M. Sensibilidad y resistencia de Pseudomonas aeruginosa a los antimicrobianos. Rev Esp Quimioter. 2007;20:230–233. [PubMed] [Google Scholar]
- 31.Kitao T, Miyoshi-Akiyama T, Kirikae T. AAC(6´)-Iaf, a novel aminoglycoside 6´-N-Acetyltransferase from multidrug-resistant Pseudomonas aeruginosa clinical isolates. Antimicrob Agents Chemother. 2009;6:2327–2334. doi: 10.1128/AAC.01360-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kim J, Park Y, Kwon H, Han K, Kang M, Woo G. Occurrence and mechanisms of amikacin resistance and its association with ß-lactamases in Pseudomonas aeruginosa: a Korean nationwide study. J Antimicrob Chemother. 2008;62:479–483. doi: 10.1093/jac/dkn244. [DOI] [PubMed] [Google Scholar]
- 33.Mendes RE, Toleman MA, Ribeiro J, Sader HS, Jones RN, Walsh TR. Integron carrying a novel metallo-beta-lactamase gene, blaIMP-16, and a fused form of aminoglycoside-resistant gene aac(6')-30/aac(6')-Ib': report from the SENTRY Antimicrobial Surveillance Program. Antimicrob Agents Chemother. 2004;48:4693–4702. doi: 10.1128/AAC.48.12.4693-4702.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Vakulenko S, Mobashery S. Versatility of aminoglycosides and prospects for their future. Clin Microbiol Rev. 2003;3:430–450. doi: 10.1128/CMR.16.3.430-450.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gad G, Mohamed H, Ashour H. Aminoglycoside resistance rates, phenotypes, and mechanisms of Gram-negative bacteria from infected patients in upper Egypt. PLoS ONE. 2011;6:13. doi: 10.1371/journal.pone.0017224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hocquet D, Vogne C, El Garch F, Vejux A, Gotoh N, Lee A. MexXY OprM efflux pump is necessary for an adaptive resistance of Pseudomonas aeruginosa to aminoglycosides. Antimicrob Agents Chemother. 2003;47:1371–1375. doi: 10.1128/AAC.47.4.1371-1375.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Mella S, Sepulveda M, Gonzalez G, Bellot H, Dominguez M, Zemelman R. Aminoglucósidos-aminociclitoles: características estructurales y nuevos aspectos sobre su resistencia. Rev Chil Infect. 2004;21:330–338. [Google Scholar]