

Abstract
Strongly nonlinear degrade-and-fire (DF) oscillations may emerge in genetic circuits having a delayed negative feedback loop as their core element. Here we study the synchronization of DF oscillators coupled through a common repressor field. For weak coupling, initially distinct oscillators remain desynchronized. For stronger coupling, oscillators can be forced to wait in the repressed state until the global repressor field is sufficiently degraded, and then they fire simultaneously forming a synchronized cluster. Our analytical theory provides necessary and sufficient conditions for clustering and specifies the maximum number of clusters that can be formed in the asymptotic regime. We find that in the thermodynamic limit a phase transition occurs at a certain coupling strength from the weakly clustered regime with only microscopic clusters to a strongly clustered regime where at least one giant cluster has to be present.
I. INTRODUCTION
Many gene regulatory networks contain negative feedback loops as their core elements [1]. The negative feedback provides robustness and noise resistance to signaling pathways [2]. They also lie at the center of many natural rhythmic circuits (such as circadian clocks [3]) and synthetic gene oscillators [4–6]. In our recent work [7] the delayed negative feedback was identified as the mechanism of oscillations in the synthetic two-gene oscillator [5]. It is well known that delayed autorepression can lead to oscillatory gene expression even with only a single regulatory element [8–12]. Typically the period of delay-induced oscillations is comparable with the delay time, and thus this mechanism seemingly could not explain rather slow oscillations observed in gene circuits. However, it turns out that in a strongly nonlinear regime, the period of oscillations is determined by the rate of enzymatic degradation of the repressor protein and therefore can be arbitrarily longer than the transcriptional and translational delay. The essential mechanism of oscillations in this circuit is based on the alternation of two regimes: slow degradation of repressor protein, and the fast production of the repressor (firing) that occurs only when the repressor is degraded below a certain critical concentration; hence we termed these sawtooth-like oscillations “degrade-and-fire” (DF) in analogy with integrate-and-fire oscillations in neuronal circuits. Bacterial cells containing the DF circuit demonstrated robust oscillations, but due to cell-to-cell variability and stochastic effects, in the absence of cell-cell communication the bacterial colony quickly becomes desynchronized. In our subsequent work [13] we succeeded in synchronizing gene oscillators across the bacterial colony; however, the synchronized regime was achieved with a different circuit design that incorporated a quorum-sensing machinery producing small-molecule Acyl-Homoserine Lactone (AHL), which served as signaling element necessary for cell-cell synchronization.
In this paper we study synchronization of DF oscillators through a purely co-repressive interaction. To enable analytical calculations, we replace the original nonlinear delay-differential model of the DF oscillator [7] by a discontinuous piecewise linear model that assumes that the concentration x degrades from the maximum value (which can be scaled to 1) linearly with unit rate, ẋ= −1 (mimicking enzymatic decay with high enzyme affinity), until it reaches the (small) threshold value η > 0, after which repressor concentration x is immediately reset to 1 (“fire”), and the process repeats. It is easy to see that the protein concentration oscillates with period 1 − η. This model closely resembles the classical “integrate-and-fire” (IF) neuron model, which has been extensively studied in the literature [14–17]. We introduce global co-repressive coupling of DF oscillators by assuming that each DF oscillator increases the repressor field of all other oscillators in proportion to its own repressor field. Equivalently, it can be interpreted as if all DF oscillators contribute their respective repressor concentrations to a global “repressor field,” that in turn is added to the repressor field of individual oscillators and therefore delays their firing. This mechanism is qualitatively similar to the inhibitory coupling of IF neurons that has been studied by Ernst et al. [17]. However, the important difference is that the oscillators are coupled all the time, and not just during the firing events, and as we see below, the dynamics of co-repressive synchronization of DF oscillators is fundamentally different from those of inhibitory coupled IF neurons. In particular, a critical value of coupling strength exists at which a phase transition from the no-clustering regime to the strong clustering regime occurs.
Consider a population of N DF oscillators coupled through a common repressor field. We postulate that the dynamics of the concentration of repressor protein xi is given by the following rules:
If positive, xi degrades linearly with rate 1, i.e., ẋi=−1, or remains constant if it has reached 0,
When the locally averaged concentration χi(t) = (1−ε)xi(t) + εX(t) [where and 0 ≤ ε ≤ 1 is the coupling strength parameter] reaches threshold η, the ith oscillator fires, and its concentration is reset to 1, i.e., xi(t+) = 1.
With these simple evolution rules, one can readily see that all oscillators must fire indefinitely (i.e., there cannot be “oscillator death” regime for any of the oscillators). Indeed, starting from an arbitrary configuration for which all χi > η, all concentrations decay toward 0 with time (those that have reached xi = 0, remain at zero) and so do all χi. Thus the oscillator with the lowest xi (possibly more than one if several oscillators have identical concentrations xi) eventually fires when the corresponding χi reaches η. After that, the oscillator j with new lowest xj has to fire when its χj reaches η, and so on. It is also clear that if any two oscillators in a population are in sync at certain time t*, i.e., xi = xj, they will remain in sync for all t > t*. What is not obvious, however, is under which conditions oscillators that are initially out of sync will synchronize in the course of the dynamics, and what the properties of the resulting clusters are. To answer these questions, we begin with the simple case of two oscillators.
II. INTERACTION OF TWO DF OSCILLATORS
In this case oscillator i (i = 1,2) fires when χi (1−ε/2)xi+εx3−i/2=η. Without loss of generality, we can assume that initially one oscillator has just fired [i.e., (x1, x2) = (x, 1) with η < x ≤ 1] and denote tf (x) > 0 the firing time associated with this configuration. The dynamics can be described through the return map R between ordered concentration pairs just after firings, (x,1) ↦ R(x,1) := (1 − tf (x),1).
The two-dimensional dynamics of two DF oscillators is contained in a parameter-dependent subset of the unit square [0,1]2 (Fig. 1). For ε = 0, firing occurs when either of the individual concentrations xi = χi touches the threshold η. A simple computation shows that R2(x,1) = (x,1) for all x; i.e., we have a continuum of periodic orbits equivalent to the rigid rotation on a two-dimensional torus [Fig. 1(a)].
FIG. 1.
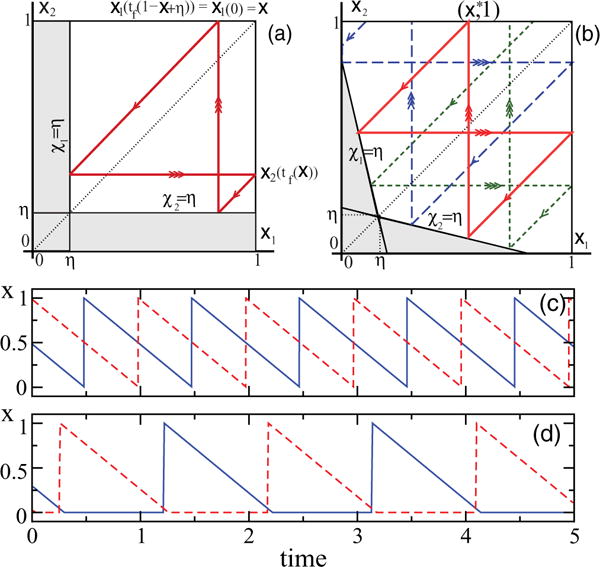
(Color online) (a, b) Typical phase trajectories of two coupled DF oscillators (solid red line indicates periodic orbits and dashed blue and green lines two transients). Simple arrows indicate motions between firings when both concentrations decay with rate 1. Triple arrows denote firings when the trajectory jumps from critical line χi = η to xi = 1. (a) ɛ = 0, every trajectory is periodic. (b) ɛ > 0, for every initial condition except x1 = x2 the system asymptotically converges to a unique periodic trajectory passing through (x*,1), where x* = 1 − 2(1 − η)/(4 – ɛ) (thick red line). (c, d) time series of two coupled DF oscillators for η = 0.01 and different values of ɛ: (c) ɛ = 0.01; (d) ɛ = 0.5.
For ε > 0, the dynamics changes drastically, as expected. Instead of a continuum of neutral periodic orbits, a single stable periodic orbit emerges that attracts all trajectories, except for the unstable periodic orbit lying on the diagonal x1= x2 [Fig. 1(b)]. These are direct consequences of Lemmas 2 and 3 below. Furthermore, it can be shown that a unique and globally stable periodic orbit exists in arbitrary systems of two coupled DF oscillators with any monotonous degradation of both concentrations. Note that this result implies a somewhat unexpected corollary that two initially distinct oscillators never synchronize and always remain distinct. As seen from Fig. 1(c) and 1(d), the two oscillators in the asymptotic regime are always in antiphase, and, for large ε, one oscillator remains repressed (xi = 0) almost the entire time when the other repressor is present (more precisely, it fires when the other concentration reaches the small value 2η/ε) and vice versa.
III. MANY COUPLED DF OSCILLATORS
Now we turn to the analysis of the clustering dynamics of a population of N co-repressively coupled DF oscillators. Before we proceed with analytical results, we illustrate the typical behavior of the system numerically. Figure 2 shows the time series of 50 oscillators with initially distinct values of x for η = 0.01 and different values of the coupling parameter ε. For small ε = 0.01 [Fig. 2(a)], all oscillators fire before they ever reach zero, and so they remain distinct (no clustering). For larger ε = 0.03 [Fig. 2(b)] some oscillators reach zero, and some of them (not all) synchronize, and thus asymptotically, only eight clusters remain. Note that this number is not universal, as it depends on specific initial conditions. For ε = 0.05 [Fig. 2(c)], only two clusters remain. Their “weights,” however, are not equal, in this example one cluster contains 24 oscillators, and another 26 oscillators. Again, this weight distribution is nonuniversal. And for very large ε = 0.5 [Fig. 2(d)] oscillators produce the repressor protein in almost exact antiphase, one repressor is present, and the other one is absent.
FIG. 2.
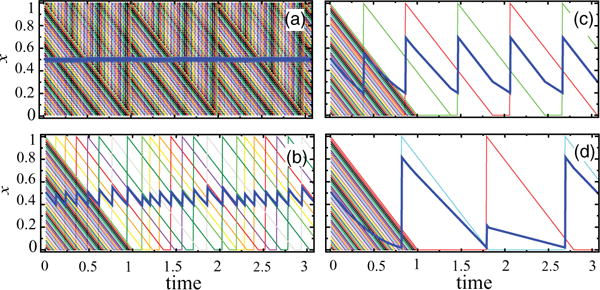
(Color online) Typical time series of 50 coupled DF oscillators and of their mean value X(t) (thick/blue line) for η = 0.01 and different values of ɛ: (a) 0.01; (b) 0.03; (c) 0.05; (d) 0.5.
By grouping oscillators with identical value of xi into one cluster, the population dynamics can be described via where nk ∈ {1,…,N} denotes the size of the cluster k and xk the corresponding repressor concentration (K ⩾ N is the total number of clusters). In this viewpoint the cluster size distribution {nk} obviously remains unaffected in time unless two clusters k and k′ fire together.
As before, we consider the map between consecutive firings. Any ordering in {(nk,xk)} is irrelevant because of the permutation symmetry in this system. However, it is more convenient to deal with ordered values of xk. Thus we assume that 0 < x1 < x2 <⋯< xK−1 < xK=1. Given {nk}, this defines the state configuration set where the firing map is effectively defined, viz., . In order to maintain the ordering of protein concentrations in time, we must include cyclic permutations of indices in the firing map dynamics. Similar considerations have been used in the analysis of the population dynamics analysis in [15]. The absence of clustering requires that, when starting in , the after-firing configuration has to lie in (where all indices are understood mod K), and this should hold for each k = 1,…,Kper where Kper is the minimal size distribution period. This property globally holds in state configuration sets provided that the coupling strength is small enough.
Lemma 1
for all k = 1,…,Kper iff
| (1) |
In particular, for εη < N/(N − 2), which is the minimal right-hand side in (1) over all cluster distributions {nk}, no clustering can ever occur, independently of the initial configuration. Notice that N/(N − 2) > 1 and → 1 as N → ∞. Lemma 1 is rigorously proven in Appendix A.
The nonclustering condition (1) can be intuitively understood as follows. The only way two clusters can merge is when one cluster reaches zero (collapses) and remains at zero at least until the following cluster also reaches zero. It is easy to see that in the worst case scenario two clusters 1 and 2 should be very close to zero when the other K − 2 clusters are very close to 1. In the limit x1,2 → 0,x3,…,xK →1, after x1 has collapsed, the value of x2 is equal to [N − εη(N − n1 − n2)]/(Nεη) when χ1 reaches η, and in order to avoid merging of n1 and n2, this value has to be positive. Of course, the same condition has to be satisfied for all consecutive pairs nk,nk+1, hence the minimum in inequality (1).
Independently of (1), we may ask about the fate of the trajectories for which no clustering occurs. Since the sequence of merging events is always finite, this actually would determine every possible asymptotic regime. In absence of clustering, the firing map in becomes , where tf still denotes the firing time. [This map is actually K − 1-dimensional because .] Since the image belongs to , one needs to iterate further until the point returns back to ; hence the dynamics to study is the return map . It can be shown (see Appendix B) that this map is always a pure contraction.
Lemma 2
For every and 0 < ε ≤ 1, there is a norm in for which is a global contraction.
It follows that, in absence of clustering, every trajectory initially in must approach a unique periodic orbit (whose single element in is the fixed point of the return map). More generally, every trajectory in a population of N oscillators must converge to the periodic orbit associated with some {nk}. Hence, the asymptotic cluster size distributions and periodic orbits are in one-to-one correspondence. Accordingly, to get asymptotically attainable cluster distributions, it suffices to compute the existence domains of the corresponding periodic orbits. These domains are given by the following statement.
Lemma 3
The periodic orbit in exists iff
| (2) |
This claim is proved in Appendix C where we also give the explicit expressions for the periodic orbit. Note that when nK = N − n1 (i.e., K = 2), we have ζ({nk})=+∞, which means that every two-cluster periodic orbit exists for any ε. In particular, for n1=n2=1 this observation implies the result for two coupled oscillators presented above.
For K > 2, the critical value ζ({nk}) is minimal for the (equi-)distribution where all nk = 1 (in this case K=N), and the corresponding minimum is ζc = 2N/(N−2). So all possible periodic orbits exist for εη up to ζc. For larger εη, some periodic orbits disappear, certainly the one associated with equidistribution, and the number of clusters K must eventually be less than N.
A closer look at the formula (2) reveals that when {nk} is only composed of microscopic clusters, i.e., when , the corresponding critical value ζ({nk}) approaches 2 in the thermodynamic limit N → ∞. Therefore, such distributions do not perdure asymptotically beyond εη ≃ 2 when N is large, and at least some of the clusters size nk reach O(N) in the course of time. Moreover, distributions that contain macroscopic cluster(s) (i.e., nk ≃ ρkN with ρk > 0 for some k) all have critical value ζ({nk}) larger than 2 in the thermodynamic limit. Thus, there is a sharp transition in the system at a certain εc = ζc (which is close to 2η for large N), from a dynamical regime in which all kinds of cluster size distributions may exist asymptotically, to a regime where every asymptotic distribution contains at least one giant O(N) cluster.
IV. MAXIMAL NUMBER OF CLUSTERS
The transition at εη = ζc can be quantitatively described by estimating the number of asymptotic clusters as a function of εη. Since the precise number actually depends on the initial condition and can be as little as 2, the appropriate quantity to compute is the maximal possible number Kmax. It can be proven (see Appendix D) that for any given K ⩾ 3 and N ⩾ K (and N ≠ 6 if K = 3), the critical value in (2) reaches its maximum for the distribution consisting of K − 1 unitary clusters of weight nk = 1 and one big cluster nK = N − K + 1. In this case, we have
and therefore all distributions with K clusters exist for 0 ≤ εη < ζmax. This immediately yields the maximum number of clusters at a given N and εη:
| (3) |
This result indicates that a kind of second order phase transition takes place at εη = ζc with Kmax that behaves like slightly above the threshold. Formula (3) implies that for each 1 ≤ K≤ Kmax(εη), there exists a nonempty set of initial conditions that produces a stable periodic trajectory with K clusters. However, the “typical” number of clusters emerging from an arbitrary set of initial conditions can be much less than this upper bound. We performed numerical simulations of the full model with initial conditions randomly selected from the [0,1]N hypercube. Figure 3 shows that the typical number of clusters for a given ε can be well approximated by the number Ke corresponding to the uniformly distributed initial condition xi=i/N. However, the number of asymptotic clusters depends strongly on the width of the distribution of initial states. We performed simulations for initial conditions xi equidistributed between 1 − δ and 1 and found that the number of asymptotic clusters strongly increases as the width of the initial distribution δ gets smaller and approaches the values close to the upper bound Kmax [Fig. 3(a), inset].
FIG. 3.
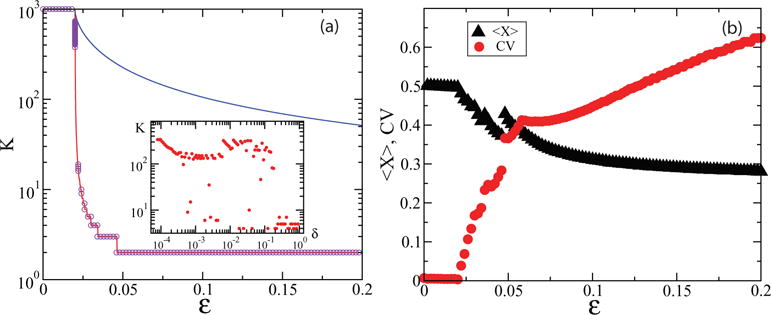
(Color online) (a) Number of clusters in the asymptotic regime for 1000 coupled oscillators with η = 0.01 and 1000 different random initial conditions for each ɛ. The solid blue line indicates the upper bound (3), and the solid red line shows the number of clusters Ke(ɛ) for the uniform initial distribution. Inset: The number of clusters as a function of the initial distribution width δ for ɛ = 0.03; (b) time-averaged mean field 〈X〉 and the coefficient of variation CV as a function of ɛ for δ = 0.01 and firing noise η = 0.05 for 10 000 oscillators and 500 periods.
We addressed the robustness of the clustering transition against noise. DF oscillators are most sensitive to noise during the firing, so we modeled this by resetting xi to a random number uniformly distributed in [1 − η, 1 + η] (η characterizes the noise strength). In this case, the clusters are not “exact,” since after firing identical xi become different; however, for small noise groups of oscillators remain closely correlated. We can characterize the degree of correlation by the coefficient of variation of the mean field X(t), CV = [〈X2〉 − 〈X〉2]1/2/〈X〉. CV is small in the nonclustered regime but increases rapidly as soon as the clustering occurs [Fig. 3(b)].
V. CONCLUDING REMARKS
We presented an analytically solvable model of co-repressively coupled degrade-and-fire oscillators in a strongly nonlinear regime. At a certain coupling strength this model exhibits a phase transition from a regime when any cluster distributions including the nonclustered state can be attained to the regime of strong clustering characterized by the appearance of at least one giant O(N) cluster. This phenomenology is reminiscent of the synchronization in the Kuramoto model of globally coupled oscillators [18] or populations of integrate-and-fire neurons [14–17]; however, the details of the transition are different. In particular, in the Kuramoto and IF models with identical oscillators synchronization occurs for any positive coupling strength, while in the array of co-repressively coupled DF oscillators the nonclustered regime is stable for sufficiently small coupling strength.
Our model neglected the variability in parameters of individual oscillators. It also ignores the spatial localization of coupling or the time delays in exchange of the repressor concentrations among the oscillators. It is well known that these effects may play significant role in the synchronization phenomena [17,19,20]. We plan to address these effects in our future work. The experimental realization of coupled gene oscillators [13] incorporated a different (co-excitatory) mechanism of coupling through a quorum-sensing mechanism. However, we believe that co-repressive coupling may also be realized experimentally, if, for example, oscillators produce an enzyme that degrades a freely diffusing inducer or the quorum sensing molecule activates a repressor protein. Our results demonstrate that a strong clustering can be expected in this case, and large out-of-phase synchronized clusters are likely to emerge.
Supplementary Material
Acknowledgments
LT acknowledges useful discussions with Jeff Hasty and William Mather. He is also grateful to CPT for hospitality during his stay at the University of Provence, Marseille. BF thanks Lai-Sang Young for comments and suggestions and the Courant Institute (NYU) for hospitality. This work was supported by EU Marie Curie fellowship PIOF-GA-2009-235741 and NIH grants RO1-GM69811 and P50-GM085764.
APPENDIX A: PROOF OF LEMMA 1
Recall that denotes the mapping in the ordered configuration set that defines the new (ordered) configuration after firing. Recall also that εη := ε/η.
We first prove that (1) is a sufficient condition. Assume that initially . Prior to firing, we have . Accordingly, the quantity χ1 reaches η at time tf defined by
The quantity tf is the actual firing time provided that x1(tf) ⩾ 0, viz., tf ≤ x1. The latter is equivalent to
| (A1) |
Using that 0 ≤ xk − x1 ≤ 1 for state configurations in , a sufficient condition for this inequality is . Applying cyclic permutations, we conclude that, for every configuration in any of the , the lowest cluster of oscillators fires before (or exactly when) it reaches 0 if
| (A2) |
When this condition is violated, cluster configurations with all coordinates xk, k⩾2, sufficiently close to 1 and x1 sufficiently close to 0 satisfy the inequality Accordingly, the concentration x1 reaches 0 before χ1 reaches η, and, to prevent clustering, we have to make sure that χ2(tf) > η i.e., tf < x2 if tf still denotes the time when x1 fires. For t⩾x1, we have . The condition χ1(tf) = η defines the time firing as follows , where ηɛ=η/ɛ. Now, the inequality tf < x2 turns out to be equivalent to
| (A3) |
which is certainly satisfied when . Solving this inequality for all permutations {nk+l} gives the desired condition (1) on the coupling parameter.
We now show that maps into when (1) holds. This consists in checking three conditions:
First, we have x2 − tf > 0 either because x1 − tf ⩾0 and x2 > x1 or simply because of (A3) when (A2) fails.
Then, the strict ordering x2 < ⋯ < xK−1<1 implies a similar ordering for the coordinates of R{nk}x where x = (x1,…,x).
- Finally, we have
where we used x2 > x1 and 1 ⩾ x1 – tf to obtain the inequality. Thus the last inequality in the definition of is satisfied for , and the proof is complete.(A4)
Finally, that (1) is a necessary condition is easy to check. Indeed, when , initial conditions in with x1 and x2 sufficiently close to 0 and the other xi close enough to 1 simultaneously violate (A1) and (A3). Accordingly, the clusters n1 and n2 fire simultaneously, and a K′-cluster distribution results with K′≤ K−1. In particular, the image cannot belong to . The proof is complete.
APPENDIX B: PROOF OF LEMMA 2
Recall that denotes the firing map acting in in the absence of clustering. The proof of Lemma 2 essentially consists in showing that, due to the dissipative nature of the coupling, all eigenvalues of the linear parts of each map lie inside the unit circle.
We first prove contraction for the individual map . Its form depends upon whether the initial condition x = (x1,…,xK) satisfies (A1) or not.
Stability in the domain where x satisfies (A1)
When only considering the first K − 1 coordinates, we obtain
for all k = 1,…,K − 1. Applying the change of variable x ↦ y defined by
the linear part of the previous expression becomes where
(We used .) The corresponding (K − 1) × (K −1) matrix is a companion matrix whose characteristic polynomial immediately follows by reading the bottom line, namely,
The polynomial coefficients are positive and decaying (as l decreases) when 0 < ɛ ≤ 1. By a classical result in numerical analysis (see, e.g., Ref. [21], p. 116), this property implies that is Schur stable, viz., all its roots lie inside the unit disk.
Stability in the domain where x does not satisfy (A1)
In this case, the map reads
for all k = 1,…,K − 1. After applying the same change of variable as before the corresponding linear part becomes
now based on . The associated characteristic polynomial in this case reads
As before, the polynomial has positive and decaying coefficients; hence all roots of lie inside the unit disk.
Proof that the return map is a contraction
Given that both spectral radii and are less than 1, the end of the proof is quite standard. Take any norm |·| in ℝK−1. We have (see, e.g., Ref. [22]). Take δ > 0 sufficiently small so that and let tδ enough be large so that
Consider the analogous for the operator and take [this requires electing δ so that both inequalities and simultaneously hold]. By choosing in ℝK−1, we conclude that the linear parts of have norm ‖·‖ less than 1, viz., is a global contraction.
Finally, the return map will be contracting for the norm ‖xs‖1/s where s is any integer larger than each sδ of the and δ is such that all these maps are contracting. The proof is complete.
APPENDIX C: PROOF OF LEMMA 3
Lemma 3 is a consequence of existence conditions for the periodic orbit associated with each cluster size distribution {nk}. This analysis of these conditions directly follows from the explicit computation of periodic orbits coordinates.
Without loss of generality, we study the solution in of the equation
| (C1) |
for any K-cluster configuration . For the sake of notation, we use the symbol Rℓ instead of and R0 = Id.
Periodic orbits have a property that remarkably simplifies their analysis; namely, all coordinates fire from the same value. In particular, this implies that they either all fire before reaching 0 or all reach 0 before firing. To see this, recall that tf(x) denotes the firing time for x. The firing map definition obviously implies (RKx)K = 1, but also (RKx)K−1 = 1 − tf(RK−1x) since (RK−1x)K−1 = 1 and tf (RK−1x) is the firing time of the configuration RK−1x. By repeating the argument for the other coordinates (RKx)k, we easily obtain the following expression:
| (C2) |
Together with the periodicity assumption RKx=x and the firing map definition, this expression implies
By induction, we conclude that successive firing levels (Rℓx)1 − tf(Rℓx) do not depend on l = 0,…,K−1, and the announced alternative follows. To proceed, we consider both cases separately.
Case 1: All coordinates fire before reaching 0
In this case, we assume that tf(Rℓx) ≤ (Rℓx)1 for all l = 0,…,K−1, and we accordingly solve the equation (C1), i.e., RK(x) = x.
Using the notation τk = xk+1 − xk, it is convenient to rewrite the periodic orbit coordinates as follows:
which in particular yields xK=1. In addition, periodicity conditions (C1) and (C2) imply
| (C3) |
To proceed, we use the assumption to rewrite as follow:
where
Accordingly, the definition of the firing map implies for all k = 1,…,K and then since we are assuming tf(Rx) ≤ (Rx)1 Repeating this argument inductively, we obtain
| (C4) |
and
| (C5) |
From equation (C3), it results that for all l = 1,…, K−1, Which is equivalent to
Using again the firing map definition we have
and, together with the relation , the previous relation thus simplifies to
for ℓ = 0,…,K−2. For ℓ = 1,…,K−2 we can use relations (C4) and (C5) to get (Rℓx)2−(Rℓx)1 = τℓ+1 and (Rℓx)1= τℓ+Δℓ−1. Accordingly, the previous relation results in
For ℓ = 0, using the expressions of x1 and tf(x) the previous equality becomes
Since all Δℓ = Δ the are equal, we finally get the condensed expression for all 1 ,…, K−1.
In order to compute Δ we introduce the latter into the definition of Δ0 via the coordinates xk. This gives a first-order equation for Δ, whose solution is
where . Direct combinatorics implies
showing that . Notice that Δ is well defined when ɛ < 1 since ΣK < 1. Moreover, we have Δ < 1 when η < 1. From the expression of Δ, we immediately get
To conclude the analysis in the present case, it remains to check the conditions on parameters, for which we have
Using (C4) and (C5), the first inequality amounts to Δ > 0, viz., . As for the second conditions, the only nontrivial restriction is (Rℓx)1 > 0 for all = 0,…,K−1, since the other ones are automatically satisfied from the definition of R and the nonclustering assumption. Again, (C4) and (C5) show that for all l = 1,…,K−1. For ℓ = 0, we have , i.e., x1 > 0. Summarizing, the periodic orbit passing exists with all coordinates firing before they reach 0 iff
Case 2: All coordinates reach 0 before firing
This case can only occur when . We claim that the solution reads
| (C6) |
for all k = 1,…,K − 1 and obviously xK = 1. To check this assertion, according to (C2), it suffices to verify that
We shall indeed show by induction that this relation holds for all k = 0,…,K − 1. Since we now assume that tf (x) > x1 we have . Using (C6) and the definition of ΣK we obtain
which gives the desired result for k = 0. The other cases proceed similarly by induction.
The existence conditions now become
and
The inequalities xk < xk+1 for all k = 1,…, K − 1 are equivalent to the following ones:
which certainly hold when . Moreover, x1 > 0 is equivalent to , and naturally, we obtain x1 < tf(x) iff ηɛ < ΣK.
For ℓ > 0, the only nontrivial constraint in is (Rℓx)1 > 0, i.e., (Rℓ−1x)2 > tf(Rℓ−1x), since all other constraints follow from the definition of the firing map and the nonclustering assumption. Therefore, all that remains to be checked is tf(Rℓx) ≤ (Rℓx)2 for all ℓ = 0,…, K − 1.
By induction we get . Using the explicit expression above, we obtain
for all ℓ = 0,…, K − 1. Altogether, we conclude that the existence condition in the case when all coordinates reach 0 before firing reduces to
APPENDIX D: MAXIMIZING CONFIGURATIONS
Lemma 4
Given K ⩾ 3 and N ⩾ K (and N ≠ 6 if K = 3), the K-cluster configuration that maximizes the critical value ζ({nk}) is the one defined by
| (D1) |
This result is not as obvious as it may first appear. Indeed, collapsing extensive clusters together and splitting off unitary clusters in a way to keep the total number K constant increases the left term in the expression of ζ({nk}) [see (2)] but simultaneously decreases the right term; so the overall shift of the critical value needs to be carefully evaluated. In addition, Lemma 4 does not hold in the case K = 3 and N = 6 since one can check that ζ({1, 1, 4}) < ζ({2,2,2}).
Proof
To show that the configuration (D1) minimizes the denominator in ζ ({nk}), we can assume without loss of generality that {nk} is a permutation of the configuration where 1 ≤ qk ≤ qk+1 and . Let ℓ ≤ K − 1 be the largest index for which qk = 1. We separate the cases and .
If , then we have ℓ > K − ℓ; i.e., the configuration {qk} has more clusters with a single unit than it has clusters with more than one unit. Thus every permutation {nk} must have two consecutive clusters with a single unit. In this case, the quantity
reaches its global maximum. Moreover, adding units to the largest cluster qK by taking off units from smaller clusters qk has the effect to increase the quantity . Thus the configuration (D1) minimizes the denominator in ζ ({nk}) over all permutations of increasing configurations {qk} with . The corresponding minimum is
To deal with the case , we begin by showing that for every K-cluster configuration we have . Consider the following alternative. Either all or there is a pair (nk,nk+1) for which . In the second case, by contradiction there must be another pair (nk′,nk′+1) where (otherwise we would have Σk nk > N which is impossible) implying that
In the first case, if , then we have
Otherwise there must be k such that (otherwise we would again have Σk nk > N) and then again
Thus in all cases, we have , which implies the desired inequality.
Still for , the configuration that maximizes the sum is
[Notice that the assumption that has exactly ℓ sites for which qk = 1 implies that N − 2(K − 1) + ℓ ⩾ 2.] Altogether we conclude that when , the denominator of ζ ({nk}) is certainly not smaller than
Now, the quantity Qℓ − (K − 1)(2N − K) increases with N provided that . This inequality holds for every when K ⩾ 5. Moreover, the quantity also increases with N for K ⩾ 2. Thus the quantity Qℓ − Q increases with N, and to ascertain that Q is a global minimum, it suffices to check that each Qℓ − Q is non-negative for N = 2K − ℓ. This amounts to verify that the following inequalities hold:
where
and
The right-hand side in the inequality above is a third-order polynomial in ℓ that is decreasing between 0 and when K ⩾ 5. Moreover, it is easy to check that
for every K ⩾ 4; i.e., we indeed have Qℓ ⩾ Q for every when K ⩾ 5.
For the cases K = 4 and K = 3, the proof needs to be improved because the quantity actually decreases with N when this integer is large. We begin with assuming K = 4 and check separately cases j = 0, 1, and 2.
For K = 4 and j = 0, the inequality 2(K − 2)⩾K − 1 shows that Q0 − Q increases with N. Moreover, we have just checked in the general case that Q0 ⩾ Q for all N ⩾ 2K when K = 4; thus Q is also a global minimum in this case.
For K = 4 and j = 1, since , we still have Q1 − Q increases with N. By numerically computing the values Q1 and Q for N = 7 = 2K − 1, we conclude that Q remains a global minimum in this case.
For K = 4 and j = 2, we claim that given an increasing configuration (1, 1,q,N − q − 2) with , the minimum S is maximal for the permutation (1, N − q − 2,1,q). Indeed, up to a cyclic permutation, there are three distinct permutations, and (1, N − q − 2, 1, q) is the only one that has nonconsecutive “1.” For (1, N − q − 2, 1, q), we have due to the assumptions on q, and the corresponding denominator of ζ({nk}) reads
This quantity is convex for q between 2 and (as a product of two positive functions, one increasing with decreasing derivative and the other one decreasing with decreasing derivative). Thus we only have to check that the values for q = 2 and are not smaller than the value for q = 1. Direct calculations reveal that the former quantity eventually grows with N faster than the latter when N is large enough. A numerical check shows that these values for q = 2 and are not smaller than the value for q = 1 provided that N ⩾ 6 (which is the minimal N for j = 2), and so the proof is complete for K = 4.
For K = 3, there are two types of configurations to consider, namely, the ordered ones (q1,q2,q3) and those of the form (q1,q3,q2). For (q1, q2, q3), due to the assumption qi ≤ qi+1, the minimum S is given by . For (q1,q3,q2), depending on the sign of , the minimum S is either or . Since both these quantities are never smaller than , the nonordered configuration always minimizes the denominator of ζ({nk}).
Using that for the configuration (q, N − 2q − p, q + p) (where and ), we get that the denominator of the critical value is at least
| (D2) |
Using similar arguments as above, one concludes that this quantity is convex for p between 0 and . Therefore, it reaches its minimum at the boundaries of this interval.
For p = 0, the quantity (D2) becomes
which is again convex with q between 1 and . Moreover, the value for grows faster with N than the value for q = 1 does. A numerical investigation reveals that the former is not smaller than the former provided that N ⩾ 7, i.e., ζ(1, N − 2, 1) ≤ ζ(q, N − 2q, q) for all when N ⩾ 7.
For , (D2) becomes
Using once again the convexity argument, we obtain that this quantity reaches its minimum for either q = 1 or . In the first case we want to check the inequality
Again, as N increases, the l.h.s grows faster than the right-hand side, and numerics show that the inequality holds for all N ⩾ 10. For , one can prove that the corresponding inequality holds for all N ⩾ 7. Therefore, we have shown that (1, 1, N − 2) maximizes ζ(n1, n2, N − n1 − n2) provided that N ⩾ 10. For N between 4 and 9, we have listed all three-cluster configurations and checked that the property remains valid, except for N = 6, as stated above.
Footnotes
PACS number(s): 87.18.Cf, 05.45.Xt, 87.16.Yc
References
- 1.Alon U. An Introduction to Systems Biology: Design Principles of Biological Circuits. Chapman & Hall/CRC Press; Boca Raton: 2007. [Google Scholar]
- 2.Stelling J, Sauer U, Szallasi Z, Doyle F, III, Doyle J. Cell. 2004;118:675. doi: 10.1016/j.cell.2004.09.008. [DOI] [PubMed] [Google Scholar]
- 3.Panda S, Hogenesch JB, Kay SA. Nature (London) 2002;417:329. doi: 10.1038/417329a. [DOI] [PubMed] [Google Scholar]
- 4.Elowitz MB, Leibler S. Nature (London) 2000;403:335. doi: 10.1038/35002125. [DOI] [PubMed] [Google Scholar]
- 5.Stricker J, Cookson S, Bennett MR, Mather WH, Tsimring LS, Hasty J. Nature (London) 2008;456:516. doi: 10.1038/nature07389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Tigges M, Marquez-Lago T, Stelling J, Fussenegger M. Nature (London) 2009;457:309. doi: 10.1038/nature07616. [DOI] [PubMed] [Google Scholar]
- 7.Mather W, Bennett MR, Hasty J, Tsimring LS. Phys Rev Lett. 2009;102:068105. doi: 10.1103/PhysRevLett.102.068105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.MacDonald N. J Theor Biol. 1977;67:549. doi: 10.1016/0022-5193(77)90056-x. [DOI] [PubMed] [Google Scholar]
- 9.Bliss RD, Painter PR, Marr AG. J Theor Biol. 1982;97:177. doi: 10.1016/0022-5193(82)90098-4. [DOI] [PubMed] [Google Scholar]
- 10.Mackey MC, Nechaeva IG. Phys Rev E. 1995;52:3366. doi: 10.1103/physreve.52.3366. [DOI] [PubMed] [Google Scholar]
- 11.Bratsun D, Volfson D, Tsimring LS. J Hasty, Proc Natl Acad Sci USA. 2005;102:14593. doi: 10.1073/pnas.0503858102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Coutinho R, Fernandez B, Lima R, Meyroneinc A. J Math Biol. 2006;52:524. doi: 10.1007/s00285-005-0359-x. [DOI] [PubMed] [Google Scholar]
- 13.Danino T, Mondragon-Palomino O, Tsimring L, Hasty J. Nature (London) 2010;463:326. doi: 10.1038/nature08753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tuckwell H. Introduction to Theoretical Neurobiology: Linear Cable Theory and Dendritic Structure. Vol. 1. Cambridge University Press; Cambridge: 1988. [Google Scholar]
- 15.Mirollo R, Strogatz S. SIAM J Appl Math. 1990;50:1645. [Google Scholar]
- 16.Bottani S. Phys Rev E. 1996;54:2334. doi: 10.1103/physreve.54.2334. [DOI] [PubMed] [Google Scholar]
- 17.Ernst U, Pawelzik K, Geisel T. Phys Rev Lett. 1995;74:1570. doi: 10.1103/PhysRevLett.74.1570. [DOI] [PubMed] [Google Scholar]
- 18.Kuramoto Y. Chemical Oscillations, Waves and Turbulence. Springer; Berlin: 1984. [Google Scholar]
- 19.Timme M, Wolf F, Geisel T. Phys Rev Lett. 2002;89:258701. doi: 10.1103/PhysRevLett.89.258701. [DOI] [PubMed] [Google Scholar]
- 20.Barahona M, Pecora LM. Phys Rev Lett. 2002;89:054101. doi: 10.1103/PhysRevLett.89.054101. [DOI] [PubMed] [Google Scholar]
- 21.Jury EI. Theory and Applications of the z-Transform Method. Wiley; New York: 1964. [Google Scholar]
- 22.Reed M, Simon B. Functional Analysis I. Academic Press; San Diego: 1980. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.