

Abstract
Transient receptor potential melastatin 4 (TRPM4) channels are Ca2+-activated nonselective cation channels that have been recently identified as regulators of detrusor smooth muscle (DSM) function in rodents. However, their expression and function in human DSM remain unexplored. We provide insights into the functional role of TRPM4 channels in human DSM under physiological conditions. We used a multidisciplinary experimental approach, including RT-PCR, Western blotting, immunohistochemistry and immunocytochemistry, patch-clamp electrophysiology, and functional studies of DSM contractility. DSM samples were obtained from patients without preoperative overactive bladder symptoms. RT-PCR detected mRNA transcripts for TRPM4 channels in human DSM whole tissue and freshly isolated single cells. Western blotting and immunohistochemistry with confocal microscopy revealed TRPM4 protein expression in human DSM. Immunocytochemistry further detected TRPM4 protein expression in DSM single cells. Patch-clamp experiments showed that 9-phenanthrol, a selective TRPM4 channel inhibitor, significantly decreased the transient inward cation currents and voltage step-induced whole cell currents in freshly isolated human DSM cells. In current-clamp mode, 9-phenanthrol hyperpolarized the human DSM cell membrane potential. Furthermore, 9-phenanthrol attenuated the spontaneous phasic, carbachol-induced and nerve-evoked contractions in human DSM isolated strips. Significant species-related differences in TRPM4 channel activity between human, rat, and guinea pig DSM were revealed, suggesting a more prominent physiological role for the TRPM4 channel in the regulation of DSM function in humans than in rodents. In conclusion, TRPM4 channels regulate human DSM excitability and contractility and are critical determinants of human urinary bladder function. Thus, TRPM4 channels could represent promising novel targets for the pharmacological or genetic control of overactive bladder.
Keywords: 9-phenanthrol, patch-clamp electrophysiology, smooth muscle, transient receptor potential channels, urinary bladder
overactive bladder (OAB) remains a poorly understood condition that presents a significant medical challenge (19). Although some therapeutic options are available for the treatment of OAB, there is no universally effective OAB therapy. While antimuscarinics are the primary pharmacological treatment for OAB, the clinical use of these agents provides limited efficacy and undesirable side effects (3, 37, 46). The long-term effectiveness of newer therapies such as mirabegron, a selective β3-adrenoceptor agonist (6, 9), and botulinum toxin (8, 26, 33) remains uncertain, and in some cases their use presents safety concerns (26, 28). The lack of safe and universally effective OAB treatments continues to spur the scientific community to seek novel therapeutic approaches to control OAB. Detrusor smooth muscle (DSM) cells represent the primary functional unit of the urinary bladder, and their dysregulation is responsible for a significant portion of lower urinary tract dysfunction. Ion channels expressed in DSM control urinary bladder function and, therefore, represent promising alternative targets for the pharmacological intervention of OAB (40, 41). Selective manipulation of individual ion channel subtypes in DSM could substantially alleviate various types of bladder dysfunction, such as OAB, urinary incontinence, or detrusor underactivity, while potentially minimizing collateral adverse effects elsewhere in the body.
A group of ion channels recently attracting significant interest is the transient receptor potential (TRP) cation channel family (25). As a large superfamily of 28 members, these channels are classified into seven related subfamilies: TRPC (canonical), TRPV (vanilloid), TRPM (melastatin), TRPP (polycystin), TRPML (mucolipin), TRPN [no mechanopotential (NOMP)], and TRPA (ankyrin) (34). The physiological roles of TRP channels include responding to physical and chemical stimuli such as temperature, pH, osmolality, pressure, stretch, light, and alkaloids, as well as intracellular stimuli such as Ca2+ (49).
A particularly interesting and important member of the TRP channel superfamily is the TRPM4 channel (18, 32). This channel is highly permeable to monovalent cations, such as Na+ and K+, but impermeable to anions and divalent cations, including Ca2+ (17, 32). TRPM4 channels, with a single-channel conductance of ∼25 pS, exhibit Ca2+ and voltage dependency (35, 36, 48, 50). These channels are expressed in various mammalian tissues (21, 27, 32), including smooth muscle (10, 13–17, 29, 39), and are implicated in many diverse physiological and pathophysiological conditions (7, 24, 30, 31, 42, 43). Activation of TRPM4 channels causes cell membrane depolarization via Na+ influx, which in turn activates L-type voltage-gated Ca2+ (CaV) channels (11). Alternatively, inhibition of TRPM4 channels promotes substantial cell hyperpolarization and smooth muscle relaxation (15, 16). Because of these unique channel properties, selective modulation of TRPM4 channels may have profound effects on cell excitability and may be an attractive, novel approach for the pharmacological treatment of various conditions, including OAB.
Recent studies have defined the expression and function of TRPM4 channels in rat and guinea pig DSM (39, 44, 45). These innovative studies revealed that TRPM4 channels are important regulators of DSM excitability and contractility in these species. However, the expression and function of TRPM4 channels in human DSM are completely unknown (25). Significant species-related differences (animal vs. human) in DSM excitability and contractility have been well documented (20). Thus, it is critically important to investigate the physiological role of TRPM4 channels directly in human DSM cells and tissues under physiological conditions (25).
Here, we provide the first detailed examination of TRPM4 channels in human DSM by applying a multidisciplinary experimental approach, including RT-PCR, Western blotting, immunohistochemistry, immunocytochemistry, patch-clamp electrophysiology, isometric DSM tension recordings, and native freshly isolated human DSM cells and tissue strips, as well as examination of the effects of a novel TRPM4 channel inhibitor 9-phenanthrol. 9-Phenanthrol specifically targets TRPM4 channels and has been used in a variety of biological preparations to reveal the contribution of these channels in physiological processes (18). The results from the current study suggest a critical role for the TRPM4 channels in human DSM excitability and contractility. Pharmacological inhibition of TRPM4 channels with 9-phenanthrol leads to DSM cell hyperpolarization, thus reducing the excitability and contractility of DSM cells. This study validates the TRPM4 channels as key regulators of human DSM excitability and contractility.1
MATERIALS AND METHODS
Human DSM tissue collection.
Human DSM tissue samples were obtained from donor patients undergoing open bladder surgeries, as previously described (22, 23). All procedures involving human tissue collection were reviewed and approved by the Institutional Review Board of the Medical University of South Carolina (protocol no. Pro00045232). Thirty-six patients [23 men and 13 women; 30 Caucasian, 4 African-American, and 2 other; 67.3 ± 1.3 (range 51–82) yr of age] without OAB symptoms and American Urological Association symptom score <8 were used in this study. Two types of DSM samples were collected from each patient. The first sample was stored in ice-cold Ca2+-free HEPES-buffered dissection solution (see Solutions and drugs) and used to conduct patch-clamp, Western blot, immunocytochemistry, immunohistochemistry, and functional studies on DSM contractility. The second sample was stored in RNAlater solution (Qiagen, Hilden, Germany) and used for RT-PCR experiments. The samples were immediately transported to the laboratory after surgery and further processed for DSM single-cell isolation and functional studies on DSM contractility.
Human DSM single-cell isolation.
Human DSM single cells were enzymatically isolated as previously described (22, 23). The freshly isolated DSM cells were used for patch-clamp recordings, single-cell-type RT-PCR, and immunocytochemistry within 12 h after isolation.
RNA extraction, RT-PCR, and sequencing.
The RNeasy Mini Kit (Qiagen) was used to extract total RNA from human DSM whole tissue, and a poll of enzymatically freshly isolated DSM single cells was extracted as previously described (23). The extracted total RNA was reverse-transcribed into cDNA using Moloney murine leukemia virus reverse transcriptase (Promega, Madison, WI) and oligo(dT) primers. The TRPM4 channel-specific primer pair sequences were designed on the basis of the cDNA complete sequences of human genes in GenBank and synthesized by Integrated DNA Technologies (Coralville, IA). The forward and reverse primers were 5′-ACCTTCAGGAAATCAGAAGAGGA-3′ and 5′-GGAACCAGCGGCGTAGG-3′, respectively. The cDNA product was amplified in the presence of GoTaq Green Master Mix (Promega) and specific primers for the TRPM4 channel using a Mastercycler gradient thermocycler (Eppendorf, Hamburg, Germany). The cDNA products from human DSM whole tissue, brain tissue, prostate tissue, and isolated DSM cells were heated for 5 min at 95°C and then amplified by 40 cycles (95°C for 30 s, 58°C for 30 s, and 72°C for 30 s) followed by a 5-min extension at 72°C. PCR products were loaded onto a 2% agarose-ethidium bromide-stained gel and allowed to migrate via electrophoresis. Total RNA samples, extracted from human brain and prostate, were purchased from Clontech Laboratories (Mountain View, CA) and used as positive controls. Negative control experiments were performed in the absence of reverse transcriptase. PCR products were purified using the GenElute PCR Clean-Up Kit (Sigma, St. Louis, MO) and sequenced at the University of South Carolina Environmental Genomic Core Facility for sequence confirmation.
Western blotting.
Western blot experiments were conducted on freshly isolated mucosa-free human DSM tissues as previously described (23, 44, 45). Briefly, ∼50 μg of total protein extracted from human DSM tissue were loaded into adjacent lanes, subjected to 7.5% precast SDS-PAGE for 50 min at 150 V, and transferred to Immobilon-P transfer membranes (Millipore, Bedford, MA) in transfer buffer using a Mini Trans-Blot electrophoretic transfer cell (Bio-Rad Laboratories, Hercules, CA) for 110 min at 75 V. The membrane was blocked with 5% dry milk-Tris-buffered saline-Tween 20 buffer for 1 h at room temperature and then incubated with the primary antibody anti-TRPM4 (1:200 dilution; catalog no. ACC-044, Alomone Labs, Jerusalem, Israel) overnight at 4°C. The membrane was washed four times with Tris-buffered saline-Tween 20 and incubated with goat anti-rabbit IgG conjugated with horseradish peroxidase (1:5,000 dilution; Jackson ImmunoResearch, West Grove, PA) in blocking buffer for 1 h at room temperature. Bound antibodies were detected by a Pierce enhanced chemiluminescence Western blotting substrate kit (Thermo Fisher Scientific; Fair Lawn, NJ) according to the manufacturer's instructions. The antibody specificity was verified by preincubation of the antibody with the respective competing peptide. Human embryonic kidney (HEK-293) cell lysate (TRPM4 overexpression lysate; catalog no. NBL1-17336, Novus Biologicals, Littleton, CO) was used as a positive control.
Immunohistochemistry and immunocytochemistry.
Immunohistochemistry and immunocytochemistry were carried out using mucosa-free human DSM tissues or freshly isolated human DSM single cells, as previously described (44, 45). Human DSM whole tissue, fixed with 4% paraformaldehyde, was sliced into 120-μm-thick sections using a tissue slicer (model G, Vibratome, Oxford Laboratories, Foster City, CA). Tissue sections or DSM single cells were stained with primary antibodies specific for TRPM4 (1:250 dilution; catalog no. ACC-044, Alomone Labs) and α-smooth muscle actin (1:100 dilution; catalog no. ab21027, Abcam). Nuclei were stained with 4′,6-diamidino-2-phenylindole (1:5,000 dilution). Secondary antibodies were tagged with Cy3-conjugated anti-rabbit IgG (1:200 dilution; Jackson ImmunoResearch) to stain for TRPM4 and Alexa Fluor 488 donkey anti-goat IgG (1:100 dilution; Life Technologies) to stain for α-smooth muscle actin. Tissue sections or single cells were mounted with 1,4-diazabicyclo[2.2.2]octane and visualized under a confocal microscope (LSM 700 META, Carl Zeiss, Oberkochen, Germany) with a ×63 oil-immersion objective.
Patch-clamp recordings.
The amphotericin B-perforated whole cell patch-clamp technique was used in all electrophysiological recordings. Transient inward cationic currents (TICCs) and voltage step-induced whole cell TRPM4 currents were recorded in voltage-clamp mode, as previously described (39, 44, 45). Briefly, TICCs were recorded at a holding potential of −70 mV (corrected for the junction potential), and the currents were analyzed as total open channel probability (NPo) before and after the addition of 9-phenanthrol. Voltage step-induced whole cell currents were recorded in Cs+-containing pipette and bath solutions in the presence of 10 mM tetraethylammonium chloride. Human DSM cells were held at −74 mV (corrected for junction potential), and then 400-ms steps from −94 to +96 mV in 10-mV increments were applied. Membrane potential of human DSM cells was recorded in current-clamp mode (current = 0). An Axopatch 200B amplifier (Digidata 1322A) and pCLAMP 10.2 software (Molecular Devices, Union City, CA) were used, and the currents were filtered using an eight-pole Bessel filter (model 900CT/9L8L, Frequency Devices, Ottawa, IL). The patch-clamp pipettes were made from borosilicate glass (Sutter Instruments, Novato, CA) and pulled using a vertical puller (model PP-830, Narishige, Tokyo, Japan). The pipettes were then polished with a fire polisher (Micro Forge MF-830, Narishige). All patch-clamp experiments were conducted at room temperature (22–23°C).
Isometric DSM tension recordings.
Isometric DSM tension was recorded as previously described (22, 23, 44, 45). DSM strips were separated into three experimental groups. In the first group, DSM strips exhibiting spontaneous phasic contractions were allowed to stabilize for ≥30 min before application of 30 μM 9-phenanthrol. In the second group, DSM contractions were induced by the cholinergic agonist carbachol (0.1 μM) and allowed to reach a stable level before application of 9-phenanthrol (30 μM). To minimize the potential effects of neurotransmitter release, the DSM strips exhibiting spontaneous phasic contractions and carbachol-induced contractions were treated in the presence of 1 μM tetrodotoxin, a selective blocker of neuronal voltage-gated Na+ channels. In the third group, nerve-evoked contractions were induced by electrical field stimulation (EFS) using a pair of platinum electrodes mounted in the tissue bath parallel to the DSM strip. The EFS pulses were generated using a PHM-152I stimulator (Med Associates, Georgia, VT); the EFS pulse parameters were 0.75-ms pulse width, 20-V pulse amplitude, and 3-s stimulus duration, with polarity reversed for alternating pulses. DSM strips were subjected to continuous repetitive stimulation with a frequency of 10 Hz at 1-min intervals or increasing frequencies from 3.5 to 50 Hz at 3-min intervals. Isometric tension was recorded at 37°C using the Myomed myograph system (Med Associates).
Solutions and drugs.
For the functional studies on human DSM contractility, the physiological saline solution consisted of (in mM) 119 NaCl, 4.7 KCl, 24 NaHCO3, 1.2 KH2PO4, 2.5 CaCl2, 1.2 MgSO4, and 11 d-glucose, aerated with 95% O2-5% CO2 (pH 7.4). The composition of Ca2+-free dissection solution was (in mM) 80 monosodium glutamate, 55 NaCl, 6 KCl, 2 MgCl2, 10 HEPES, and 10 d-glucose, with pH adjusted to 7.3 with NaOH. The extracellular (bath) solution used in the patch-clamp experiments in the gap-free mode (voltage and current clamp) contained (in mM) 134 NaCl, 6 KCl, 1 MgCl2, 2 CaCl2, 10 d-glucose, and 10 HEPES, with pH adjusted to 7.4 with NaOH. The patch-clamp pipette solution for these experiments contained (in mM) 110 potassium aspartate, 30 KCl, 10 NaCl, 1 MgCl2, 10 HEPES, and 0.05 EGTA, with pH adjusted to 7.2 with NaOH, and was supplemented with freshly dissolved (every 1–2 h) amphotericin B (200 μg/ml) in dimethyl sulfoxide (DMSO). The extracellular (bath) solution used in the voltage step-induced whole cell patch-clamp experiments contained (in mM) 10 tetraethylammonium chloride, 6 CsCl, 124 NaCl, 1 MgCl2, 2 CaCl2, 10 HEPES, and 10 d-glucose, with pH adjusted to 7.3 with CsOH. The composition of Cs+-containing pipette solution was (in mM) 110 CsOH, 110 aspartic acid, 10 NaCl, 1 MgCl2, 10 HEPES, 0.05 EGTA, and 30 CsCl, with pH adjusted to 7.2 with CsOH, and was supplemented with freshly dissolved (every 1–2 h) amphotericin B (200 μg/ml) in DMSO. BSA was obtained from Thermo Fisher Scientific (Fair Lawn, NJ). All other compounds were obtained from Sigma-Aldrich. 9-Phenanthrol was dissolved daily in DMSO (30 or 100 mM stock solution). Tetrodotoxin (in citrate buffer) and carbachol were dissolved in double-distilled water. The final concentration of DMSO in the bath solutions did not exceed 0.05%.
Data analysis and statistics.
MiniAnalysis software (Synaptosoft, Decatur, GA) was used to analyze the DSM phasic contraction parameters. The DSM contractile activity was quantified from measurement of average phasic contraction amplitude (the difference between the force-time baseline curve and the maximum peak of the contractions), frequency (contractions/min), muscle force integral (calculated by integrating the area under the force-time baseline curve), phasic contraction duration (defined as width of the individual phasic contraction at 50% of the amplitude), and DSM tone (the difference between the zero line and the force-time baseline curve). Statistical analyses were performed with Prism 4.03 software (GraphPad, La Jolla, CA), and CorelDRAW Graphics Suite X3 software (Corel, Ottawa, ON, Canada) was used to illustrate the data. For the EFS-induced contractions, the contraction amplitude at the EFS frequency under control conditions was taken to be 100%, and the data were normalized. For evaluation of the effect of cumulative addition of 9-phenanthrol (0.1–30 μM) on 10-Hz EFS-induced contractions, the last 5 min before addition of each concentration to the bath were analyzed. The TICCs were analyzed as NPo, as previously described (39, 44). In voltage step experiments, the last 200-ms interval was analyzed and averaged before, after, and following washout of 9-phenanthrol. Five minutes of ≥10 min of stable current-clamp recordings prior to application of 9-phenanthrol were analyzed for control data, and the last 5 min of each 10-min continuous recording after application of 9-phenanthrol were analyzed to evaluate the effect of 9-phenanthrol on the membrane potential. Data are summarized as means ± SE for the number of DSM strips or cells (n) isolated from the number of patients (N). Data were compared using a two-way ANOVA followed by Bonferroni's post test or paired or unpaired Student's t-test where appropriate. P < 0.05 was considered statistically significant.
RESULTS
TRPM4 channel mRNA detection in human DSM.
Previously, we demonstrated TRPM4 channel expression in rat and guinea pig DSM (39, 44, 45). Here, we took an additional step to elucidate TRPM4 channel expression at the cell and tissue levels in human DSM. Utilizing RT-PCR, we detected mRNA transcripts for TRPM4 channels in DSM whole tissues (n = 4, N = 4; Fig. 1). Since mRNA transcripts in DSM whole tissue may not have originated from only DSM cells, but may also have originated from other cell types in the detrusor muscle layer (such as neurons, interstitial cells, fibroblasts, and vascular and endothelial cells), additional RT-PCR experiments were carried out on RNA samples collected only from freshly isolated human DSM cells. The single-cell RT-PCR experiments on isolated DSM cells, which avoid plausible additional sources of mRNA from non-DSM cells, clearly confirmed the presence of TRPM4 channel mRNA directly in human DSM cells (n = 4, N = 4; Fig. 1). Negative control experiments performed in the absence of the reverse transcriptase enzyme did not detect the band, indicating an absence of genomic DNA contamination (Fig. 1).
Fig. 1.

Transient receptor potential melastatin 4 (TRPM4) channel mRNA expression in human detrusor smooth muscle (DSM) whole tissue and native freshly isolated human DSM single cells. Gel electrophoresis imaging illustrates RT-PCR detection of TRPM4 channel mRNA transcripts (196 bp) in DSM whole tissue and DSM single cells (n = 4, N = 4). Human brain and prostate samples were used as positive controls (+). No products were observed in the negative control (−RT), in which reverse transcriptase (RT) was not added to the reaction.
TRPM4 channel protein expression at the protein level in human DSM.
To further investigate if the TRPM4 channels are expressed at the protein level in native human DSM tissue, we performed Western blot experiments using a TRPM4 channel-specific antibody (see materials and methods). A ∼134-kDa band, consistent with the expected molecular mass of the TRPM4 channel protein, was detected (Fig. 2A). In control experiments, preabsorption of the primary antibody with its antigenic competing peptide resulted in the loss of the protein band, thereby indicating the specificity of the antibody for its intended epitope (n = 4, N = 4; Fig. 2A). The slightly different bands might be due to posttranslational modifications of native human DSM tissue and HEK-293 cells (Fig. 2A). Thus the Western blot experiments confirmed the presence of the TRPM4 channel protein in human DSM tissue.
Fig. 2.

Western blot, immunohistochemical, and immunocytochemical detections of TRPM4 channel protein in human DSM tissue and DSM single cells. A: Western blot showing TRPM4 channel protein expression in human DSM tissue. Arrow indicates ∼134-kDa band, consistent with expected molecular mass of TRPM4 channel protein. Lack of an immunoreactive band in the presence of competing peptide (CP) confirmed specificity of the primary antibody. HEK-293 cell lysate was used as a positive control. B: confocal images showing immunohistochemical detection of TRPM4 channel protein expression in human DSM tissue. Red staining (bottom left) indicates TRPM4 channels; blue staining indicates cell nuclei (top left); green staining indicates α-smooth muscle actin (α-SMA, top right); merged image (bottom right) illustrates overlap of all 3 images. C: confocal images illustrating immunocytochemical detection of TRPM4 channel protein expression in isolated human DSM cells. Red staining (bottom left) indicates TRPM4 channels; blue staining indicates cell nucleus (top left); green staining indicates α-SMA (top right); merged image (bottom right) illustrates overlap of all 3 images. Results were verified in 4 separate experiments using DSM whole tissue or multiple DSM cells isolated from 4 patients. D and E: immunohistochemistry and immunocytochemistry experiments, respectively. In control experiments, no staining was visible after absorption of the primary antibody with a competing peptide (+CP control). Merged image (bottom right) illustrates overlap of all 3 images.
Immunohistochemistry with confocal microscopy also detected the expression and localization of TRPM4 channel protein in human DSM whole tissue (Fig. 2B), further confirming the results from the Western blot experiments. Our data reveal that TRPM4 channels are expressed in DSM cells, as evidenced by our findings that TRPM4 channels and α-smooth muscle actin are colocalized spatially within the same cell.
To determine further whether DSM cells express TRPM4 channel proteins, immunocytochemistry was carried out with TRPM4 channel-specific antibodies. As illustrated in Fig. 2C, coimmunolabeling with the TRPM4 channel-specific antibodies and the α-smooth muscle actin marker revealed the expression of TRPM4 channel protein in DSM single cells. Control experiments were further conducted by preabsorption of the primary antibody with its antigenic competing peptide to verify the specificity of the antibody for its intended epitope (Fig. 2, D–E).
Pharmacological inhibition of TRPM4 channels with 9-phenanthrol decreases TICC activity in freshly isolated human DSM cells.
The role of TRPM4 channels in human DSM cell excitability was investigated using the amphotericin B-perforated patch-clamp technique and the novel TRPM4 channel inhibitor 9-phenanthrol (18). The average cell capacitance of all human DSM cells used in this study was 21.9 ± 2.0 pF (n = 37, N = 19). At the holding potential of −70 mV, human DSM cells exhibited TICCs (Fig. 3A). Application of 30 μM 9-phenanthrol decreased TICC activity by ∼50% from NPo of 2.7 ± 0.6 to 1.3 ± 0.4 (n = 15, N = 9, P < 0.05; Fig. 3B). These results support the novel concept that TRPM4 channels are functionally active in human DSM. By regulating TICC activity, TRPM4 channels are key determinants of human DSM excitability.
Fig. 3.

Inhibition of transient inward cationic currents (TICCs) by the TRPM4 channel-selective inhibitor 9-phenanthrol in freshly isolated human DSM cells. A: original recording illustrating the inhibitory effect of 30 μM 9-phenanthrol on TICC activity in a human DSM single cell recorded at −70 mV. B: summary data illustrating inhibitory effects of 30 μM 9-phenanthrol on TICCs, analyzed as total open channel probability (NPo) before and after application of 30 μM 9-phenanthrol (n = 15, N = 9). *P < 0.05.
Pharmacological inhibition of TRPM4 channels with 9-phenanthrol decreases the amplitude of voltage step-induced whole cell currents in human DSM cells.
To further investigate the role of TRPM4 channels in human DSM excitability, we measured the current-voltage relationship of the cationic current in human DSM isolated cells at −94 to +96 mV in 10-mV increments for 400 ms, as previously described (44). Between steps, the holding potential was −74 mV (corrected for junction potential). In this experimental series, K+ channel currents were blocked by 10 mM extracellular tetraethylammonium and Cs+-containing solution (see Solutions and drugs). 9-Phenanthrol (30 μM) significantly inhibited the amplitude of the voltage step-induced whole cell currents in human DSM cells, and the effect was reversed by washout with 9-phenanthrol-free extracellular bath solution (n = 7, N = 7, P < 0.05; Fig. 4). For the depolarization step to +96 mV, 9-phenanthrol inhibited the outward current by 43.4 ± 8.2% (n = 7, N = 7, P < 0.05; Fig. 4). These results reveal the inhibitory effect of 9-phenanthrol on the whole cell currents in human DSM cells, providing further insight into TRPM4 channel properties in human DSM cells.
Fig. 4.

Inhibition of TRPM4 channels with 9-phenanthrol attenuates voltage step-induced whole cell currents in human DSM cells. A: representative recordings illustrate the inhibitory effect of 30 μM 9-phenanthrol on the amplitude of the voltage step-induced whole cell current in human DSM cell. Inhibitory effect of 9-phenanthrol was reversed after washout of the human DSM cell with fresh 9-phenanthrol-free bath solution. B: summary data of current-voltage relationships in the absence (control), in the presence, and after washout of 30 μM 9-phenanthrol (n = 7, N = 7). *P < 0.05.
TRPM4 channels regulate the membrane potential in human DSM cells.
The role of TRPM4 channels in controlling the human DSM cell resting membrane potential was investigated using the current-clamp mode of the patch-clamp technique. 9-Phenanthrol (30 μM) significantly hyperpolarized the membrane potential in human DSM cells by 24.3 ± 7.7 mV (n = 4, N = 3, P < 0.05; Fig. 5). Under control conditions, the DSM cell membrane potential was −22.7 ± 6.1 mV, which was hyperpolarized to −46.9 ± 11.9 mV after application of 9-phenanthrol (n = 4, N = 3, P < 0.05; Fig. 6). These results strongly support a key functional role for the TRPM4 channels in regulating the resting membrane potential in human DSM cells.
Fig. 5.

Inhibition of TRPM4 channels with 9-phenanthrol hyperpolarizes the resting membrane potential (RMP) in human DSM cells. A: representative current-clamp recording illustrating hyperpolarizing effects of 30 μM 9-phenanthrol on RMP in a human DSM cell. B: summary data illustrating hyperpolarizing effects of 30 μM 9-phenanthrol on the RMP in human DSM cells (n = 4, N = 3). *P < 0.05.
Fig. 6.

Inhibition of TRPM4 channels with 9-phenanthrol attenuates spontaneous phasic and tonic contractions of human DSM isolated strips. A: representative myograph recording demonstrating inhibition of spontaneous phasic contractions and tone of a human DSM isolated strip by 30 μM 9-phenanthrol. B: summary data illustrating decrease in spontaneous phasic contraction amplitude, muscle force integral, frequency, duration, and tone of human DSM strips by 30 μM 9-phenanthrol (n = 11, N = 5). ***P < 0.005.
The TRPM4 channel inhibitor 9-phenanthrol significantly decreases spontaneous phasic and tonic contractions in human DSM isolated strips.
In this experimental series, in vitro functional studies were carried out to elucidate the effects of the TRPM4 channel inhibitor 9-phenanthrol on spontaneous contractile activity of human DSM isolated strips. 9-Phenanthrol (30 μM) significantly decreased the spontaneous phasic contraction amplitude by 99.3 ± 0.7%, muscle force integral by 99.4 ± 0.5%, contraction duration by 94.2 ± 5.8%, contraction frequency by 97.3 ± 2.7%, and muscle tone by 64.8 ± 8.7% (n = 11, N = 5, P < 0.005; Fig. 6). These results suggest that TRPM4 channels are key regulators of human DSM spontaneous phasic and tonic contractions under physiological conditions.
The TRPM4 channel inhibitor 9-phenanthrol attenuates carbachol-induced contractions in human DSM isolated strips.
Initiation of bladder voiding contractions is associated with activation of parasympathetic nerves releasing acetylcholine, which subsequently activates muscarinic receptors in DSM cells (2). To study the role of the TRPM4 channels under conditions of muscarinic receptor stimulation, we performed functional studies on human DSM contractility in the presence of the muscarinic receptor agonist carbachol. Specifically, we sought to determine the effects of 9-phenanthrol on carbachol-induced contractions in human DSM isolated strips. The muscarinic receptor agonist carbachol (0.1 μM) rapidly increased the spontaneous phasic contractions of DSM strips (Fig. 7A). 9-Phenanthrol (30 μM) significantly decreased the amplitude of carbachol-induced contraction by 84.5 ± 7.7%, muscle force integral by 89.6 ± 2.9%, contraction duration by 64.1 ± 8.8%, contraction frequency by 68.9 ± 7.7%, and muscle tone by 56.9 ± 4.6% (n = 17, N = 5, P < 0.005; Fig. 7B).
Fig. 7.

Inhibition of TRPM4 channels with 9-phenanthrol significantly reduces carbachol-induced phasic and tonic contractions of human DSM isolated strips. A: representative myograph recording obtained from a human DSM strip depicting the inhibitory effect of 30 μM 9-phenanthrol on 0.1 μM carbachol-induced phasic contractions. B: summary data illustrating inhibitory effects of 30 μM 9-phenanthrol on amplitude, muscle force integral, frequency, duration, and tone of carbachol-induced contractions of human DSM isolated strips (n = 17, N = 5). ***P < 0.005.
The TRPM4 channel inhibitor 9-phenanthrol reduces nerve-evoked contractions in human DSM isolated strips.
During bladder voiding, activation of parasympathetic nerves causes release of the excitatory neurotransmitter acetylcholine, which induces forceful DSM contractions (2). In this experimental series, we sought to explore how the cumulative addition of 9-phenanthrol (0.1–30 μM) reduces DSM nerve-evoked contractions induced by continuous 10-Hz EFS in the absence of tetrodotoxin. A representative myograph recording in Fig. 8A illustrates that the cumulative addition of 9-phenanthrol (0.1–30 μM) reduced the 10-Hz EFS-induced contractions in human DSM isolated strips. Figure 8B illustrates the concentration responses and demonstrates significant inhibitory effects of 9-phenanthrol on nerve-evoked contraction amplitude and muscle force integral, with IC50 values and maximum inhibition of 3.3 (95% confidence interval = 1.1–10.1) μM and 40.5 ± 5.4% and 3.5 (95% confidence interval = 1.3–9.1) μM and 27.3 ± 7.1%, respectively (n = 16, N = 11, P < 0.05; Fig. 8).
Fig. 8.

Cumulative addition of 9-phenanthrol inhibits continuous 10-Hz electrical field stimulation (EFS)-induced contractions of human DSM isolated strips. A: representative myograph recording from a human DSM strip illustrating effects of 0.1–30 μM 9-phenanthrol on 10-Hz EFS-induced contractions. B: cumulative concentration-response curves for inhibitory effects of 9-phenanthrol on 10-Hz EFS-induced contraction amplitude and muscle force integral of human DSM isolated strips (n = 16, N = 11).
In the next series of experiments, human DSM isolated strips were stimulated by increasing EFS frequencies (3.5–50 Hz). After a frequency-response curve under control conditions, DSM strips were incubated with 30 μM 9-phenanthrol and a second frequency-response curve was generated. 9-Phenanthrol significantly decreased the contraction amplitude generated in response to EFS (n = 22, N = 16, P < 0.05; Fig. 9B). At the highest frequency of 50 Hz, 9-phenanthrol (30 μM) inhibited EFS-induced contraction amplitude in human DSM isolated strips by 30.3 ± 3.7% compared with the control responses (Fig. 9B). This experimental series further indicates that inhibition of TRPM4 channels reduces nerve-evoked contractions of human DSM isolated strips to a lesser extent than spontaneous phasic and tonic contractions (Fig. 7).
Fig. 9.
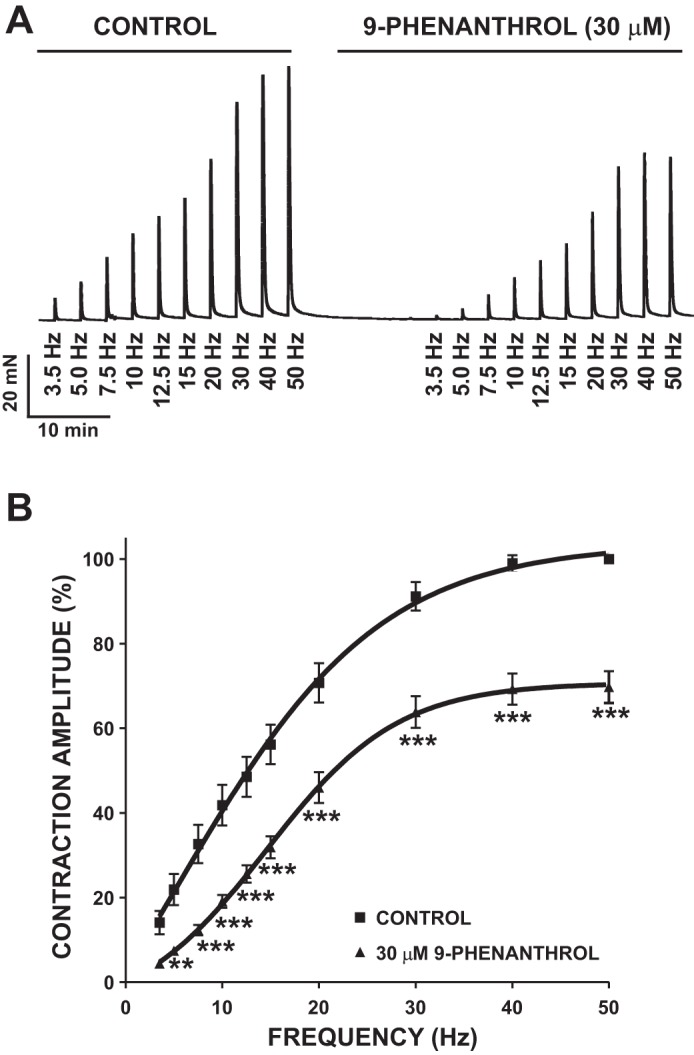
Inhibition of TRPM4 channels with 9-phenanthrol decreases the amplitude of 3.5- to 50-Hz EFS-induced contractions of human DSM isolated strips. A: representative recording of EFS-induced contractions (stimulation frequency = 3.5–50 Hz) in the absence of 9-phenanthrol (control) and 10 min after application of 30 μM 9-phenanthrol. B: frequency-response curves in the presence or absence of 30 μM 9-phenanthrol illustrating a decrease in amplitude of EFS-induced contractions of human DSM isolated strips (n = 22, N = 16). **P < 0.01, ***P < 0.005.
DISCUSSION
This study used clinically characterized human DSM tissues and isolated DSM cells obtained from donor patients without a preoperative history of OAB to investigate the functional role of TRPM4 channels in the human urinary bladder under physiological conditions. The data revealed novel findings that TRPM4 channels are important and physiologically relevant regulators of human DSM.
The physiological role of the TRPM4 channels is facilitation of membrane depolarization via nonselective influx of monovalent cations through the cell membrane (11). Given the greater driving force for Na+ than K+ under physiological conditions, pharmacological inhibition of TRPM4 channels can reduce the net Na+ influx to promote membrane potential hyperpolarization (15, 16). On the other hand, activation of TRPM4 channels would favor an increase in Na+ influx, causing membrane depolarization (11). Therefore, unlike K+ channels, which are responsible primarily for membrane hyperpolarization, TRPM4 channels contribute to the depolarization component of the ionic currents in excitable cells (15, 17, 29, 30).
Recently, TRPM4 channels have been identified in rat and guinea pig DSM, suggesting important roles for these channels in the rodent urinary bladder (39, 44, 45, 47). However, there was a lack of knowledge regarding the TRPM4 channel expression and function in human DSM (25). Since humans are the primary species of interest for therapeutic intervention, investigations on human DSM are critically important to validate previous findings in animal studies and to fully characterize TRPM4 channel properties in humans while determining key interspecies differences. Here, we revealed TRPM4 channel expression in human DSM by utilizing a combination of molecular biological techniques, which demonstrated TRPM4 channel expression in human DSM whole tissue and single cells. Our single-cell RT-PCR, Western blot, immunohistochemistry, and immunocytochemistry with confocal microscopy experiments (Figs. 1 and 2) provided the first direct evidence for TRPM4 channel expression at mRNA and protein levels in human DSM cells and further demonstrated TRPM4 channel localization at the DSM cell membrane.
The abundant TRPM4 channel protein expression in human DSM may infer an important functional role for these channels in the human bladder. In rat and guinea pig DSM cells, inhibition of the TRPM4 channels with 9-phenanthrol attenuated TICC activity, thereby decreasing intracellular Ca2+ levels to promote DSM relaxation (39, 44, 45). The TRPM4 channel inhibitor 9-phenanthrol reduced cellular excitability to a much larger extent in human than rodent DSM (Fig. 10A) (44), indicating important species differences. The voltage-clamp recordings revealed that 9-phenanthrol significantly attenuated TICC activity at −70 mV (Fig. 3) and inhibited voltage step-induced whole cell outward currents in human DSM cells (Fig. 4). Furthermore, 9-phenanthrol substantially hyperpolarized the human DSM membrane potential by ∼24 mV (Fig. 5). The combined results from our patch-clamp studies (Figs. 3–5) support the concept that TRPM4 channels are functionally active in human DSM and play a key role in controlling human DSM excitability.
Fig. 10.

Species-related differences in inhibitory effects of 9-phenanthrol on human and rodent DSM excitability and contractility. A: effects of 30 μM 9-phenanthrol on voltage step-induced whole cell currents recorded in human and guinea pig DSM cells, represented as percentage of whole cell current (n = 7, N = 7 for human; n = 10, N = 5 for guinea pig). *P < 0.05; **P < 0.01. B: effects of 9-phenanthrol (30 μM) on spontaneous phasic DSM contractions in humans (n = 11, N = 5), guinea pigs (n = 7, N = 7), and rats (n = 8, N = 7). **P < 0.01; ***P < 0.005. Rat and guinea pig data are from Smith et al. (44, 45).
Hyperpolarization of DSM cell membrane potential promotes inhibition of L-type CaV channels and leads to DSM relaxation (41). Previous studies on DSM isolated from guinea pigs and rats showed that inhibition of TRPM4 channels with 9-phenanthrol reduced the intracellular Ca2+ levels in DSM cells to induce DSM relaxation (44, 45). We were intrigued by our finding of a higher degree of 9-phenanthrol-induced attenuation of the spontaneous phasic contraction amplitude, muscle force integral, duration, frequency, and tone of human DSM isolated strips (Fig. 6) than inhibition of DSM contractility in rats and guinea pigs (44, 45) (Fig. 10B). Furthermore, inhibition of the TRPM4 channels with 9-phenanthrol attenuated carbachol-induced (Fig. 7) and nerve-evoked (Figs. 8 and 9) contractions in human DSM isolated strips. Consistent with previous findings in rodents (39, 44, 45), our functional studies clearly support a key regulatory role of TRPM4 channels in human DSM contractility. One important observation is that 9-phenanthrol much more effectively inhibited the spontaneous phasic DSM contractions than nerve-evoked DSM contractions, suggesting that TRPM4 channel inhibition may effectively control the nonvoiding contractions associated with detrusor overactivity while having little or no effect on normal voiding.
To investigate the functional role of TRPM4 channels in human DSM, we used 9-phenanthrol as a selective pharmacological inhibitor. This agent has been successfully used to study TRPM4 channel activity in various preparations (18), including rodent DSM (39, 44, 45). However, recent studies indicated that 9-phenanthrol inhibits transmembrane protein 16A (TMEM16A) channels (5) in vascular smooth muscle and activates intermediate-conductance Ca2+-activated K+ (IK) channels (12) in endothelial cells. These nonspecific effects of 9-phenanthrol raised an important question about the plausible involvement of TMEM16A and/or IK channels in the inhibitory effects of 9-phenanthrol on human DSM. Theoretically, 9-phenanthrol could affect TMEM16A and IK channel activity and, thus, reduce human DSM excitability and contractility. However, although we cannot completely exclude this possibility, our experimental data support the concept that the majority of the inhibitory effects of 9-phenanthrol in human DSM are mediated by TRPM4 channel inhibition. The substantial hyperpolarizing effect of 9-phenanthrol on the DSM membrane potential (∼24 mV; Fig. 5) is consistent with a relatively large ionic gradient for Na+ (ENa = +65 mV) in our experimental conditions. This significant shift (Fig. 5) could be explained by TRPM4 channel inhibition by 9-phenanthrol and subsequent reduction of Na+ conductance, but not by TMEM16A channel inhibition. TMEM16A channels are Ca2+-activated Cl− channels, contributing to Cl− conductance in smooth muscle cells (4). Because of the relatively small driving force for Cl− (ECl = −32 mV, with the assumption that pipette Cl− replaces all the intracellular Cl− under our experimental conditions), we expect that inhibition of TMEM16A channels would cause only a minor, if any, shift in DSM resting membrane potential. However, the physiological role of TMEM16A channels in human DSM is largely unknown, and clarification of these important issues requires further investigation. On the other hand, it is very unlikely that 9-phenanthrol inhibitory effects are mediated by IK channel activation, since previous studies reported no functional role of IK channels in the bladder (1, 38). Most importantly, our molecular biology data clearly demonstrate an abundant TRPM4 channel mRNA and protein expression in human DSM cells (Figs. 1 and 2). Therefore, our data support the concept that the majority of 9-phenanthrol inhibitory effects in human DSM are caused by TRPM4 channel inhibition.
In the present study, we demonstrated significant species-related differences in TRPM4 channel activity in human DSM compared with rat (45) and guinea pig (44) DSM (Fig. 10). Pharmacological inhibition of TRPM4 channels with 9-phenanthrol leads to a larger decrease in human than rodent DSM excitability and contractility. Figure 10A illustrates that the whole cell 9-phenanthrol-sensitive current is much higher in human than guinea pig DSM cells. We also compared the effects of 9-phenanthrol on the spontaneous phasic contractions in human, guinea pig, and rat DSM isolated strips. Consistently, the data analysis revealed a greater inhibitory effect of 9-phenanthrol on DSM contractility in human than guinea pig and rat DSM isolated strips (Fig. 10B). However, no significant differences were noted between rats and guinea pigs (Fig. 10B). These intriguing results indicate important interspecies-related differences and reveal that TRPM4 channels have a more prominent physiological role in the regulation of DSM function in humans than rodents (Fig. 10).
A critical step for development of novel, safe, and more effective therapies for OAB is a better understanding of the mechanisms that control DSM excitability and contractility in OAB patients. Abnormal functionality of TRPM4 channels could result in substantial pathological changes in DSM and lead to OAB and associated detrusor overactivity. Dysfunction in expression and activity of certain ion channels or their regulatory mechanisms may affect DSM contractility and result in OAB (40, 41). However, the physiological role of TRPM4 channels in OAB etiology requires future investigations on DSM tissue and cells obtained from patients with confirmed OAB symptoms.
Given the prominent role of the TRPM4 channels in human DSM excitability and contractility, these channels may represent novel therapeutic targets for pharmacological or genetic treatment of OAB. The challenge of TRPM4 channel expression in other tissues (32) is not unique to therapeutic targets affecting lower urinary tract function, and, fortunately, this obstacle has been pharmacologically overcome in a number of disease states. Considering the relatively higher TRPM4 channel mRNA expression in DSM than vasculature (39), as well as the important functional roles of TRPM4 channels in DSM, pharmacological manipulation of TRPM4 channels in OAB patients may provide effective treatment with minimal adverse collateral cardiovascular effects. The potential clinical application of TRPM4 channel modulation for OAB therapy should be considered as a novel approach for OAB management and further validated in clinical trials.
GRANTS
This work was supported by National Institute of Diabetes and Digestive and Kidney Diseases Grant R01 DK-106964, South Carolina Clinical and Translational Research Institute Pilot Project National Center for Advancing Translational Sciences Grant UL1 TR-000062, and University of South Carolina ASPIRE-I Grant 11110-E406 to G. V. Petkov.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
K.L.H., E.S.R., and G.V.P. developed the concept and designed the research; K.L.H., A.C.S., S.P.P., J.M., and G.V.P. performed the experiments; K.L.H., A.C.S., S.P.P., J.M., and G.V.P. analyzed the data; K.L.H., J.M., E.S.R., and G.V.P. interpreted the results of the experiments; K.L.H., A.C.S., J.M., and G.V.P. prepared the figures; K.L.H., A.C.S., and G.V.P. drafted the manuscript; K.L.H., A.C.S., S.P.P., J.M., E.S.R., and G.V.P. edited and revised the manuscript; K.L.H., A.C.S., S.P.P., J.M., E.S.R., and G.V.P. approved the final version of the manuscript.
ACKNOWLEDGMENTS
We thank Aaron Provence for performing some of the experiments; Medical University of South Carolina (MUSC) urology staff surgeons Drs. Thomas Keane, Harry Clarke, Stephen Savage, Ross Rames, Sandip Prasad, and Jonathan Picard, as well as MUSC urology residents Taylor Vaughan, Samuel Walker Nickles, Matthew Young, Erin Burns, Justin Ellett, Ryan Levey, Austin Younger, Mark Currin, Nima Baradaran, Olugbemisola McCoy, and Tracy Tipton for help with human tissue collection; and Drs. Wenkuan Xin and Vítor Fernandes and Aaron Provence for critical evaluation of the manuscript.
Footnotes
This article is the topic of an Editorial Focus by Kirk L. Hamilton (18a).
REFERENCES
- 1.Afeli SA, Rovner ES, Petkov GV. SK but not IK channels regulate human detrusor smooth muscle spontaneous and nerve-evoked contractions. Am J Physiol Renal Physiol 303: F559–F568, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Andersson KE, Arner A. Urinary bladder contraction and relaxation: physiology and pathophysiology. Physiol Rev 84: 935–986, 2004. [DOI] [PubMed] [Google Scholar]
- 3.Armstrong RB, Dmochowski RR, Sand PK, MacDiarmid S. Safety and tolerability of extended-release oxybutynin once daily in urinary incontinence: combined results from two phase 4 controlled clinical trials. Int Urol Nephrol 39: 1069–1077, 2007. [DOI] [PubMed] [Google Scholar]
- 4.Bulley S, Jaggar JH. Cl− channels in smooth muscle cells. Pflügers Arch 466: 861–872, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Burris SK, Wang Q, Bulley S, Neeb ZP, Jaggar JH. 9-Phenanthrol inhibits recombinant and arterial myocyte TMEM16A channels. Br J Pharmacol 172: 2459–2468, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chapple CR, Cardozo L, Nitti VW, Siddiqui E, Michel MC. Mirabegron in overactive bladder: a review of efficacy, safety, and tolerability. Neurourol Urodyn 33: 17–30, 2014. [DOI] [PubMed] [Google Scholar]
- 7.Cho CH, Lee YS, Kim E, Hwang EM, Park JY. Physiological functions of the TRPM4 channels via protein interactions. BMB Rep 48: 1–5, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Conte A, Giannantoni A, Proietti S, Giovannozzi S, Fabbrini G, Rossi A, Porena M, Berardelli A. Botulinum toxin A modulates afferent fibers in neurogenic detrusor overactivity. Eur J Neurol 19: 725–732, 2012. [DOI] [PubMed] [Google Scholar]
- 9.Cui Y, Zong H, Yang C, Yan H, Zhang Y. The efficacy and safety of mirabegron in treating OAB: a systematic review and meta-analysis of phase III trials. Int Urol Nephrol 46: 275–284, 2014. [DOI] [PubMed] [Google Scholar]
- 10.Dwyer L, Rhee PL, Lowe V, Zheng H, Peri L, Ro S, Sanders KM, Koh SD. Basally activated nonselective cation currents regulate the resting membrane potential in human and monkey colonic smooth muscle. Am J Physiol Gastrointest Liver Physiol 301: G287–G296, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fliegert R, Glassmeier G, Schmid F, Cornils K, Genisyuerek S, Harneit A, Schwarz JR, Guse AH. Modulation of Ca2+ entry and plasma membrane potential by human TRPM4b. FEBS J 274: 704–713, 2007. [DOI] [PubMed] [Google Scholar]
- 12.Garland CJ, Smirnov SV, Bagher P, Lim CS, Huang CY, Mitchell R, Stanley C, Pinkney A, Dora KA. TRPM4 inhibitor 9-phenanthrol activates endothelial cell intermediate conductance calcium-activated potassium channels in rat isolated mesenteric artery. Br J Pharmacol 172: 1114–1123, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gonzales AL, Amberg GC, Earley S. Ca2+ release from the sarcoplasmic reticulum is required for sustained TRPM4 activity in cerebral artery smooth muscle cells. Am J Physiol Cell Physiol 299: C279–C288, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gonzales AL, Earley S. Endogenous cytosolic Ca2+ buffering is necessary for TRPM4 activity in cerebral artery smooth muscle cells. Cell Calcium 51: 82–93, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gonzales AL, Earley S. Regulation of cerebral artery smooth muscle membrane potential by Ca2+-activated cation channels. Microcirculation 20: 337–347, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gonzales AL, Garcia ZI, Amberg GC, Earley S. Pharmacological inhibition of TRPM4 hyperpolarizes vascular smooth muscle. Am J Physiol Cell Physiol 299: C1195–C1202, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Guinamard R, Demion M, Launay P. Physiological roles of the TRPM4 channel extracted from background currents. Physiology 25: 155–164, 2010. [DOI] [PubMed] [Google Scholar]
- 18.Guinamard R, Hof T, Del Negro CA. The TRPM4 channel inhibitor 9-phenanthrol. Br J Pharmacol 171: 1600–1613, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18a.Hamilton KL. New life in overactive bladder. Focus on “Novel regulatory mechanism in human urinary bladder: central role of transient receptor potential melastatin 4 channels in detrusor smooth muscle function.” Am J Physiol Cell Physiol (February 17, 2016). doi: 10.1152/ajpcell.00039.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hanna-Mitchell AT, Kashyap M, Chan WV, Andersson KE, Tannenbaum C. Pathophysiology of idiopathic overactive bladder and the success of treatment: a systematic review from ICI-RS 2013. Neurourol Urodyn 33: 611–617, 2014. [DOI] [PubMed] [Google Scholar]
- 20.Hashitani H, Brading AF. Electrical properties of detrusor smooth muscles from the pig and human urinary bladder. Br J Pharmacol 140: 146–158, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hof T, Salle L, Coulbault L, Richer R, Alexandre J, Rouet R, Manrique A, Guinamard R. TRPM4 non-selective cation channels influence action potentials in rabbit Purkinje fibres. J Physiol 594: 295–306, 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hristov KL, Afeli SAY, Parajuli SP, Cheng QP, Rovner ES, Petkov GV. Neurogenic detrusor overactivity is associated with decreased expression and function of the large conductance voltage- and Ca2+-activated K+ channels. PLos One 8: e68052, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hristov KL, Chen M, Kellett WF, Rovner ES, Petkov GV. Large-conductance voltage- and Ca2+-activated K+ channels regulate human detrusor smooth muscle function. Am J Physiol Cell Physiol 301: C903–C912, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jacobs G, Oosterlinck W, Dresselaers T, Geenens R, Kerselaers S, Himmelreich U, Herijgers P, Vennekens R. Enhanced β-adrenergic cardiac reserve in Trpm4−/− mice with ischaemic heart failure. Cardiovasc Res 105: 330–339, 2015. [DOI] [PubMed] [Google Scholar]
- 25.Kaneko Y, Szallasi A. Transient receptor potential (TRP) channels: a clinical perspective. Br J Pharmacol 171: 2474–2507, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Karsenty G, Baverstock R, Carlson K, Diaz DC, Cruz F, Dmochowski R, Fulford S, Giannantoni A, Heesakkers J, Kaufmann A, Peyrat L, Thavaseelan J, Dasgupta P. Technical aspects of botulinum toxin type A injection in the bladder to treat urinary incontinence: reviewing the procedure. Int J Clin Pract 68: 731–742, 2014. [DOI] [PubMed] [Google Scholar]
- 27.Kruse M, Pongs O. TRPM4 channels in the cardiovascular system. Curr Opin Pharmacol 15: 68–73, 2014. [DOI] [PubMed] [Google Scholar]
- 28.Kuo HC, Liao CH, Chung SD. Adverse events of intravesical botulinum toxin a injections for idiopathic detrusor overactivity: risk factors and influence on treatment outcome. Eur Urol 58: 919–926, 2010. [DOI] [PubMed] [Google Scholar]
- 29.Launay P, Fleig A, Perraud AL, Scharenberg AM, Penner R, Kinet JP. TRPM4 is a Ca2+-activated nonselective cation channel mediating cell membrane depolarization. Cell 109: 397–407, 2002. [DOI] [PubMed] [Google Scholar]
- 30.Little SC, Mohler PJ. TRPM4 modulates sinus node diastolic depolarization. Heart Rhythm 10: 1690–1691, 2013. [DOI] [PubMed] [Google Scholar]
- 31.Loh KP, Ng G, Yu CY, Fhu CK, Yu D, Vennekens R, Nilius B, Soong TW, Liao P. TRPM4 inhibition promotes angiogenesis after ischemic stroke. Pflügers Arch 466: 563–576, 2014. [DOI] [PubMed] [Google Scholar]
- 32.Mathar I, Jacobs G, Kecskes M, Menigoz A, Philippaert K, Vennekens RR. Trpm4. Hand Exp Pharmacol 222: 461–487, 2014. [DOI] [PubMed] [Google Scholar]
- 33.Mohee A, Khan A, Harris N, Eardley I. Long-term outcome of the use of intravesical botulinum toxin for the treatment of overactive bladder (OAB). BJU Int 111: 106–113, 2013. [DOI] [PubMed] [Google Scholar]
- 34.Nilius B, Owsianik G. The transient receptor potential family of ion channels. Genome Biol 12: 218, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nilius B, Prenen J, Janssens A, Owsianik G, Wang CB, Zhu MX, Voets T. The selectivity filter of the cation channel TRPM4. J Biol Chem 280: 22899–22906, 2005. [DOI] [PubMed] [Google Scholar]
- 36.Nilius B, Prenen J, Tang JS, Wang CB, Owsianik G, Janssens A, Voets T, Zhu MX. Regulation of the Ca2+ sensitivity of the nonselective cation channel TRPM4. J Biol Chem 280: 6423–6433, 2005. [DOI] [PubMed] [Google Scholar]
- 37.Oefelein MG. Safety and tolerability profiles of anticholinergic agents used for the treatment of overactive bladder. Drug Saf 34: 733–754, 2011. [DOI] [PubMed] [Google Scholar]
- 38.Parajuli SP, Hristov KL, Soder RP, Kellett WF, Petkov GV. NS309 decreases rat detrusor smooth muscle membrane potential and phasic contractions by activating SK3 channels. Br J Pharmacol 168: 1611–1625, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Parajuli SP, Hristov KL, Sullivan MN, Xin W, Smith AC, Earley S, Malysz J, Petkov GV. Control of urinary bladder smooth muscle excitability by the TRPM4 channel modulator 9-phenanthrol. Channels (Austin) 7: 537–540, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Petkov GV. Central role of the BK channel in urinary bladder smooth muscle physiology and pathophysiology. Am J Physiol Regul Integr Comp Physiol 307: R571–R584, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Petkov GV. Role of potassium ion channels in detrusor smooth muscle function and dysfunction. Nat Rev Urol 9: 30–40, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Schattling B, Steinbach K, Thies E, Kruse M, Menigoz A, Ufer F, Flockerzi V, Bruck W, Pongs O, Vennekens R, Kneussel M, Freichel M, Merkler D, Friese MA. TRPM4 cation channel mediates axonal and neuronal degeneration in experimental autoimmune encephalomyelitis and multiple sclerosis. Nat Med 18: 1805–1811, 2012. [DOI] [PubMed] [Google Scholar]
- 43.Simard C, Hof T, Keddache Z, Launay P, Guinamard R. The TRPM4 non-selective cation channel contributes to the mammalian atrial action potential. J Mol Cell Cardiol 59: 11–19, 2013. [DOI] [PubMed] [Google Scholar]
- 44.Smith AC, Hristov KL, Cheng Q, Xin W, Parajuli SP, Earley S, Malysz J, Petkov GV. Novel role for the transient potential receptor melastatin 4 channel in guinea pig detrusor smooth muscle physiology. Am J Physiol Cell Physiol 304: C467–C477, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Smith AC, Parajuli SP, Hristov KL, Cheng Q, Soder RP, Afeli SA, Earley S, Xin W, Malysz J, Petkov GV. TRPM4 channel: a new player in urinary bladder smooth muscle function in rats. Am J Physiol Renal Physiol 304: F918–F929, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Staskin DR, Dmochowski RR, Sand PK, Macdiarmid SA, Caramelli KE, Thomas H, Hoel G. Efficacy and safety of oxybutynin chloride topical gel for overactive bladder: a randomized, double-blind, placebo controlled, multicenter study. J Urol 181: 1764–1772, 2009. [DOI] [PubMed] [Google Scholar]
- 47.Syam N, Rougier JS, Abriel H. Glycosylation of TRPM4 and TRPM5 channels: molecular determinants and functional aspects. Front Cell Neurosci 8: 52, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ullrich ND, Voets T, Prenen J, Vennekens R, Talavera K, Droogmans G, Nilius B. Comparison of functional properties of the Ca2+-activated cation channels TRPM4 and TRPM5 from mice. Cell Calcium 37: 267–278, 2005. [DOI] [PubMed] [Google Scholar]
- 49.Wu LJ, Sweet TB, Clapham DE. International Union of Basic and Clinical Pharmacology. LXXVI. Current progress in the mammalian TRP ion channel family. Pharmacol Rev 62: 381–404, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yoo JC, Yarishkin OV, Hwang EM, Kim E, Kim DG, Park N, Hong SG, Park JY. Cloning and characterization of rat transient receptor potential-melastatin 4 (TRPM4). Biochem Biophys Res Commun 391: 806–811, 2010. [DOI] [PubMed] [Google Scholar]