

Somatic mutations in JAK2, CALR and MPL are found in the majority of myeloproliferative neoplasms (MPN) but many patients also harbor somatic mutations in epigenetic regulators of DNA methylation (TET2, DNMT3A and IDH1/2) or chromatin structure (ASXL1 and EZH2). In MPN patients, mutations in TET2, ASXL1 and EZH2 occur either prior to or following the acquisition of JAK2V617F 1 and, recently, the order of mutation acquisition for JAK2V617F and TET2 has been shown to influence hematopoietic stem/progenitor cell biology and clinical presentation.2 DNMT3A is the next most frequently mutated gene in MPN after TET2, affecting 7–10% of patients.3–5 However in contrast to other mutations, DNMT3A mutations (DNMT3Amut) have only been reported early in myeloid disease: prior to acquisition of JAK2V617F or in a separate clone in MPN;1,6 prior to NPM1 or FLT3 mutations in de novo acute myeloid leukemia;7 and prior to transformation of MPN or myelodysplasia into acute myeloid leukemia.8 In this study we used clonal analysis of hematopoietic colonies from DNMT3Amut MPN patients to investigate timing of mutation acquisition, subclonal evolution, and the influence of mutation order.
Thirteen DNMT3Amut MPN patients were identified in whom viable material was available for clonal analysis (Online Supplementary Table S1): nine JAK2V617F-mutated and two MPL-mutated patients identified by exome-sequencing, one JAK2exon12-mutated patient identified by whole-genome sequencing, and one CALR-mutated patient identified by targeted gene screening. MPN diagnoses conformed to British Committee for Standards in Haematology. All samples were obtained following written informed consent and ethical approval. Peripheral blood mononuclear cells were cultured to obtain burst-forming units-erythroid (BFU-E) as described previously,2 and 2991 individual colonies (average of 230 colonies/patient) were genotyped by Sanger sequencing for mutations in DNMT3A, JAK2, CALR and MPL.
DNMT3Amut occurred prior to acquisition of JAK2V617F in four patients (‘DNMT3A-first’ patients, Figure 1A) as evidenced by single-mutant colonies harboring DNMT3Amut only and double-mutant colonies with DNMT3Amut and JAK2V617F. Single-mutant colonies represented a substantial proportion of BFU-E (mean 58%, range 27%–86%), consistent with mutant allele fractions from granulocyte whole-exome sequencing, and indicating the presence of significant ‘pre-JAK2’ multi-lineage clonal hematopoiesis in these patients. These results accord with observations of clonal hematopoiesis in patients with essential thrombocythemia despite low allele burdens of JAK2V617F,9 and with reports of DNMT3Amut-associated clonal hematopoiesis in normal individuals.10,11 In three patients, DNMT3Amut and JAK2V617F colonies were mutually exclusive (‘biclonal’ patients, Figure 1B). In such patients, the two mutations arose either in separate cells downstream of a shared ancestral clone or in clonally unrelated cells. DNMT3Amut and JAK2V617F clones did not share any of the additional mutations identified by exome sequencing in the three biclonal patients (Figure 2 patients #25, #81, #27). Moreover, in one of the two female patients, DNMT3Amut and JAK2V617F clones harbored different active X-chromosomes (Online Supplementary Figure S1). These data demonstrate that DNMT3Amut and JAK2V617F clones in the same patient can represent clonally-unrelated expansions.
Figure 1.

Timing of acquisition of DNMT3A mutations. Single cell-derived hematopoietic erythroid colonies (BFU-E) were grown in vitro from peripheral blood-derived mononuclear cells and individually genotyped for mutations using Sanger sequencing. Plots in (A), (B), (C) and (D) show colony genotyping results for mutations in DNMT3A and JAK2, MPL or CALR for each patient and the order of mutation acquisition. Within each plot, each dot represents a single colony and its quadrant placement shows the corresponding genotype of DNMT3A (vertical axis) and JAK2/MPL/CALR (horizontal axis). Solid red arrows within quadrants show the confirmed path of clonal evolution. Wt, wild-type; het, heterozygous mutation; hom, homozygous mutation; PV, polycythemia vera; ET, essential thrombocythemia; PMF, primary myelofibrosis; PPV-MF, post-PV myelofibrosis (A) DNMT3A-first patients: four patients in whom mutated DNMT3A occurred prior to the acquisition of JAK2V617F. (B) Biclonal patients: three patients in whom DNMT3Amut and JAK2V617F were in separate clones. (C) JAK2/MPL-first patients: three patients in whom mutated JAK2 or MPL occurred prior to the acquisition of mutated DNMT3A. (D) Order unknown: three patients in whom the order of mutation acquisition of DNMT3A and JAK2/MPL/CALR could not be delineated as only wild-type colonies and/or double mutant colonies were detected.
Figure 2.

Clonal structures of DNMT3A-mutated MPN. Individual single cell-derived hematopoietic erythroid colonies (BFU-E) from six patients were genotyped using Sanger sequencing or Fluidigm single nucleotide polymorphism genotyping for their respective somatic variants identified previously by whole exome sequencing. Genotyping results from individual colonies were then used to construct phylogenetic trees. Circles represent the subclones; wild-type (white); mutated (brown). The earliest detectable clone is represented at the top of each structure, with subsequent subclones shown below. Somatic mutations acquired in each subclone are indicated beside respective circles, and represent those that are acquired in addition to mutations present in earlier clones. Numbers of colonies identified for each subclone are shown inside circles. Mutations in JAK2, MPL, CALR and DNMT3A are highlighted in red. Additional mutations in genes known to be recurrently mutated in myeloid malignancies are highlighted in green. ET: essential thrombocythemia; PMF: primary myelofibrosis; PV: polycythemia vera; het: heterozygous mutation; hom: homozygous mutation.
In three patients, DNMT3Amut occurred after acquisition of mutated JAK2 or MPL (JAK2/MPL-first patients, Figure 1C) but these patients were more difficult to identify. In patient #650, the order of mutation acquisition was initially unclear as the majority of colonies were double-mutant for DNMT3A and JAK2exon12, and no antecedent single-mutant colonies were detected (Figure 1C). However, we confirmed that the JAK2exon12 mutation arose first, and that heterozygous-JAK2exon12 colonies with wild-type DNMT3A became undetectable following acquisition of DNMT3Amut, because colonies grown from an earlier time-point showed heterozygous-JAK2exon12 colonies with wild-type DNMT3A (Online Supplementary Figure S2A). Furthermore, whole genome sequencing of a diagnostic sample and one taken at a later time-point demonstrated mutated JAK2exon12 at both times but DNMT3Amut at only the later time-point (Online Supplementary Figure S2B). Consistent with this mutation order, the two homozygous-JAK2exon12 subclones present in this patient (Figure 1C, h1 and h2) carried different mitotic recombination breakpoints (Online Supplementary Figure S2C). In two further JAK2/MPL-first patients, antecedent JAK2/MPL single-mutant colonies were detected but were present only at low levels (~3% of total colonies, Figure 1C). Overall, considering MPN patients with sequential acquisition of mutations within the same clone, single-mutant JAK2/MPL subclones were significantly smaller than single-mutant DNMT3Amut and double-mutant subclones (P=0.03 for both comparisons, t-test, Online Supplementary Figure S3). To exclude a confounding effect of mutations in other genes known to be recurrently mutated in myeloid malignancies, whole exome or genome sequencing data were interrogated for all 13 patients. Four patients had additional mutations in TET2, CBL or SH2B3. Delineation of full phylogenetic hierarchies using Sanger sequencing or Fluidigm single nucleotide polymorphism genotyping (see Online Supplementary Methods) of individual colonies from these patients did not identify any preferential association of these mutations with either single or double-mutant sub-clones (Figure 2). Our data therefore suggest that JAK2/MPL single-mutant subclones may have a competitive disadvantage compared with DNMT3Amut subclones, in which case JAK2/MPL-first patients may be enriched among those in whom mutation order could not be determined by colony assay (Figure 1D). To investigate competition between JAK2/MPL-mutated and DNMT3Amut sub-clones further, colonies were grown from paired samples obtained at different time-points (median separation 35 months; range, 6–179 months) in ten patients (3 biclonal, 3 JAK2/MPL-first, 3 DNMT3-first, 1 order unclear). In six patients who harbored single-mutant DNMT3Amut colonies (2 patients receiving hydroxycarbamide, 1 patient receiving pipobroman, 2 patients receiving interferon-α, and 1 patient not receiving cytoreduction), the proportions did not change significantly between time-points (Figure 3A, B; blue shading). Similarly, in seven patients with double-mutant colonies (4 patients receiving hydroxycarbamide, 1 patient receiving pipobroman and 2 patients not receiving cytoreduction) there was no significant change in the proportions over time (Figure 3A,B; purple shading). In contrast, in all six patients with single-mutant JAK2/MPL colonies (1 patient receiving hydroxycarbamide, 2 patients receiving interferon and 3 patients not receiving cytoreduction), the proportions of single-mutant colonies fell significantly over time (Figure 3A,B; red shading, P=0.027, Wilcoxon signed rank test,). While the observed reduction of JAK2/MPL colonies could have been influenced by interferon treatment in two patients (#81 and #25), two patients in whom this pattern was also observed had not received cytoreduction (#27 and #650), supporting the notion that single-mutant JAK2/MPL subclones have a competitive disadvantage. We found no associations between subclonal changes over time and treatment responses (measured in accordance with European LeukemiaNet guidelines) in patients, possibly because individual patients harbored multiple and differing combinations of the various clones.
Figure 3.
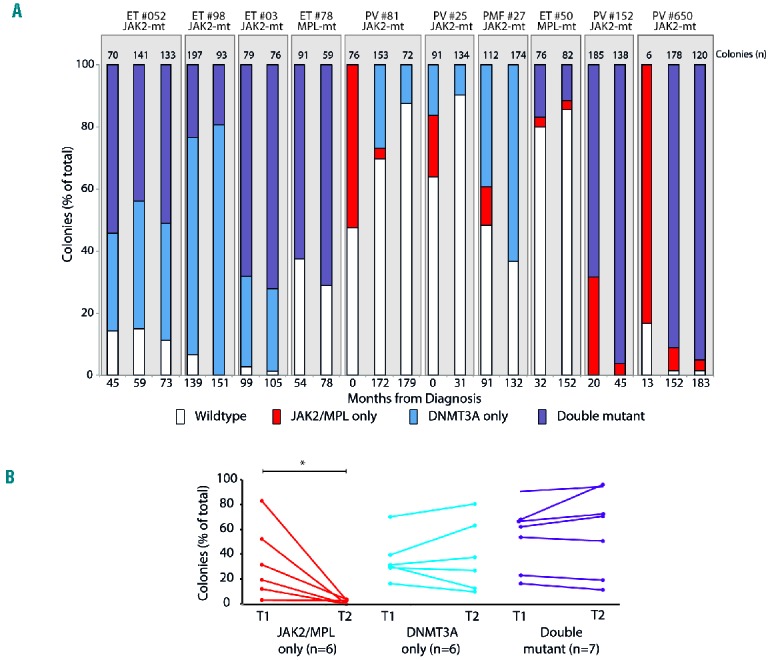
Evolution of subclones in DNMT3A-mutated MPN. (A) Colonies grown from paired samples obtained at different time-points (median separation 35 months; range, 6–179 months) in ten patients. Vertical axis shows the percentage of total colonies and columns are shaded to represent the proportions of the different genotypes (red, JAK2/MPL-only; blue, DNMT3A-only; purple, double mutant). Numbers of colonies genotyped per patient are shown above columns and the timings of sample acquisition (months from diagnosis) are shown below. (B) Changes in subclonal proportions over time for the ten patients in (A) for a total of 19 subclones. red, JAK2/MPL-only; blue, DNMT3A-only; purple, double mutant; T1 and T2 represent the earliest and latest time-points sampled for the patients. The median interval between T1 and T2 did not differ significantly between the different subclones (one-way analysis of variance). * <0.05 Wilcoxon ranked sum test.
We next assessed whether order of acquisition of DNMT3Amut influenced JAK2V617F-mutated MPN phenotype. In the eight patients in whom mutation order was established, all four DNMT3Amut-first patients had essential thrombocythemia, and of four patients in whom JAK2V617F occurred on a DNMT3A-non-mutated background (1 JAK2V617F–first and 3 biclonal patients), three had polycythemia vera and one had myelofibrosis. To expand this cohort, an additional 918 patients were screened by targeted gene sequencing, and a further 33 patients with DNMT3Amut and JAK2V617F were identified. Copy-number corrected variant allele fractions were used to determine the order of mutation acquisition as recently described,2 and mutation order was unambiguously assigned in eight further patients: two DNMT3A-first patients had essential thrombocythemia, and six JAK2-first patients comprised three with polycythemiva vera, one with post-polycythemia vera myelofibrosis, one primary myelofibrosis and one essential thrombocythemia. Combining both cohorts, all six DNMT3A-first patients presented with essential thrombocythemia. By contrast, of ten patients in whom JAK2V617F arose on a wild-type DNMT3A background, seven presented with polycythemia vera and only one with essential thrombocythemia (P=0.003, chi-squared test). There were no significant differences in other clinical features (Online Supplementary Table S2) in this cohort. Our results therefore indicate that mutation order influences clinical presentation, not only in MPN patients with JAK2V617F and TET2 mutations, as previously described,2 but also in patients with JAK2V617F and DNMT3A mutations. Mutations in either DNMT3A or TET2 are associated with an essential thrombocythemia phenotype when acquired prior to JAK2V617F. By contrast, acquisition of JAK2V617F prior to mutation of DNMT3A or TET2 is associated with polycythemia vera. ‘TET2-first’ patients were older at presentation in our previous study,2 but no difference in age was identified between DNMT3A-first and JAK2-first patients. This may be due to the smaller number of patients in the current study or may reflect a real difference in the age at which TET2 and DNMT3A mutations are acquired.
In summary, we demonstrate that in MPN, DNMT3Amut can follow JAK2 and MPL mutations, and that JAK2/MPL single-mutant subclones have a competitive disadvantage in vivo compared with DNMT3Amut subclones. This concept is consistent with observations that DNMT3A and TET2 mutations confer an advantage to hematopoietic stem/progenitor cells,2,12–14 whereas this is not the case for JAK2V617F in some mouse models.15 Furthermore, we show that mutation order of JAK2V617F and DNMT3Amut is associated with differences in MPN phenotype. This emphasizes the importance of the pattern of acquisition of JAK2V617F with respect to mutations in epigenetic modifiers in influencing the phenotype of JAK2V617F-mutated MPN.
Acknowledgments
We thank the Cambridge Blood and Stem Cell Biobank (Cambridge University) and Anthony Bench, Haemato-Oncology Diagnostics Service (Addenbrooke’s hospital) for sample collection and storage, and the Cancer Genome Project (Wellcome Trust Sanger Institute, Hinxton) for next-generation sequencing. Work in Professor Green’s laboratory is supported by Leukemia and Lymphoma Research, the Wellcome Trust, the Medical Research Council, the Kay Kendall Leukaemia Fund, the Cambridge NIHR Biomedical Research Centre, the Cambridge Experimental Cancer Medicine Centre and the Leukemia and Lymphoma Society of America. ALG and JN were supported by Kay Kendall Leukaemia Fund clinical fellowships. PJC is a Wellcome Trust senior clinical fellow.
Footnotes
The online version of this letter has a Supplementary Appendix.
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Lundberg P, Karow A, Nienhold R, et al. Clonal evolution and clinical correlates of somatic mutations in myeloproliferative neoplasms. Blood 2014;123(14):2220–2228. [DOI] [PubMed] [Google Scholar]
- 2.Ortmann CA, Kent DG, Nangalia J, et al. Effect of mutation order on myeloproliferative neoplasms. N Engl J Med 2015;372(7):601–612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Stegelmann F, Bullinger L, Schlenk RF, et al. DNMT3A mutations in myeloproliferative neoplasms. Leukemia 2011;25(7):1217–1219. [DOI] [PubMed] [Google Scholar]
- 4.Abdel-Wahab O, Pardanani A, Rampal R, Lasho TL, Levine RL, Tefferi A. DNMT3A mutational analysis in primary myelofibrosis, chronic myelomonocytic leukemia and advanced phases of myeloproliferative neoplasms. Leukemia 2011;25(7):1219–1220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nangalia J, Massie CE, Baxter EJ, et al. Somatic CALR mutations in myeloproliferative neoplasms with nonmutated JAK2. N Engl J Med 2013;369(25):2391–2405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rao N, Butcher CM, Lewis ID, et al. Clonal and lineage analysis of somatic DNMT3A and JAK2 mutations in a chronic phase polycythemia vera patient. Br J Haematol 2012;156(2):268–270. [DOI] [PubMed] [Google Scholar]
- 7.Shlush LI, Zandi S, Mitchell A, et al. Identification of pre-leukaemic haematopoietic stem cells in acute leukaemia. Nature 2014;506(7488):328–333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fried I, Bodner C, Pichler MM, et al. Frequency, onset and clinical impact of somatic DNMT3A mutations in therapy-related and secondary acute myeloid leukemia. Haematologica 2012;97(2):246–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kralovics R, Teo S-S, Li S, et al. Acquisition of the V617F mutation of JAK2 is a late genetic event in a subset of patients with myeloproliferative disorders. Blood 2006;108(4):1377–1380. [DOI] [PubMed] [Google Scholar]
- 10.Jaiswal S, Fontanillas P, Flannick J, et al. Age-related clonal hematopoiesis associated with adverse outcomes. N Engl J Med 2014;371(26):2488–2498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Genovese G, Kähler AK, Handsaker RE, et al. Clonal hematopoiesis and blood-cancer risk inferred from blood DNA sequence. N Engl J Med 2014;371(26):2477–2487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Challen GA, Sun D, Jeong M, et al. Dnmt3a is essential for hematopoietic stem cell differentiation. Nat Genet 2012;44(1):23–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ko M, Bandukwala HS, An J, et al. Ten-eleven-translocation 2 (TET2) negatively regulates homeostasis and differentiation of hematopoietic stem cells in mice. Proc Natl Acad Sci USA 2011;108(35):14566–14571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Moran-Crusio K, Reavie L, Shih A, et al. Tet2 loss leads to increased hematopoietic stem cell self-renewal and myeloid transformation. Cancer Cell 2011;20(1):11–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Li J, Kent DG, Chen E, Green AR. Mouse models of myeloproliferative neoplasms: JAK of all grades. Dis Model Mech 2011;4(3):311–317. [DOI] [PMC free article] [PubMed] [Google Scholar]