

Abstract
Microtubules (MTs) are involved in key processes in plant cells, including cell division, growth and development. MT-interacting proteins modulate MT dynamics and organization, mediating functional and structural interaction of MTs with other cell structures. In addition to conventional microtubule-associated proteins (MAPs) in plants, there are many other MT-binding proteins whose primary function is not related to the regulation of MTs. This review focuses on enzymes, chaperones, or proteins primarily involved in other processes that also bind to MTs. The MT-binding activity of these multifunctional MAPs is often performed only under specific environmental or physiological conditions, or they bind to MTs only as components of a larger MT-binding protein complex. The involvement of multifunctional MAPs in these interactions may underlie physiological and morphogenetic events, e.g., under specific environmental or developmental conditions. Uncovering MT-binding activity of these proteins, although challenging, may contribute to understanding of the novel functions of the MT cytoskeleton in plant biological processes.
Keywords: multifunctional MAP, MAP, microtubules, tubulin, plants
MAPs and their role in plant cells
Traditional microtubule-associated proteins (MAPs) are typically conserved in eukaryotes. However, plants possess a set of MAPs specific to plant morphology and physiology (Gardiner, 2013). A fundamental feature of MAPs is their interaction with MTs through a MT-binding domain that is manifested in MT organization, dynamics or cellular transport, which influences plant morphogenesis. The localization of MAPs in the cell is well described mainly because of their close association with MT structures. Consequently, MAPs are direct MT-interactors and their function is dependent on their MT-binding activity (Buschmann and Lloyd, 2008). MAPs are motor proteins that utilize MTs as tracks to transport cargo such as kinesins. Structural MAPs or severing proteins such as MAP65 and katanin bind, bundle or cleave MTs, and therefore, are involved in MT organization. MT dynamics, on the other hand, is influenced by MT +tip associated proteins (+TIPs), such as, e.g., CLASP, EB1, etc., through their binding and interactions at the +end of growing MT. Conventional MAPs have been reviewed in several outstanding reviews (for instance Lloyd and Hussey, 2001; Akhmanova and Steinmetz, 2008; Sedbrook and Kaloriti, 2008; Gardiner, 2013; Hamada, 2014; Hashimoto, 2015; Li et al., 2015) and will not be discussed here. MAPs identified in plants are listed in the Table 1.
Table 1.
List of MAPs described in plants.
| MAP | Characterized in | References | Notes |
|---|---|---|---|
| MAP65 | |||
| MAP65-1 | Arabidopsis thaliana | Jiang and Sonobe, 1993; Smertenko et al., 2004; Van Damme et al., 2004 | |
| MAP65-2 | Arabidopsis thaliana | Li et al., 2009 | |
| MAP65-3/PLEIADE | Arabidopsis thaliana | Muller et al., 2004 | |
| MAP65-4 | Arabidopsis thaliana | Van Damme et al., 2004 | |
| MAP65-5 | Gaillard et al., 2008; Smertenko et al., 2008 | ||
| MAP65-6 | Mao et al., 2005 | ||
| MAP65-7 | Theologis et al., 2000 | Found in silico in Arabidopsis | |
| MAP65-8 | Arabidopsis thaliana | Smertenko et al., 2008 | Does not associate with MT |
| MAP65-9 | Arabidopsis thaliana | Smertenko et al., 2008 | Pollen, does not associate with MT |
| MT +END BINDING PROTEINS | |||
| EB1 | Arabidopsis thaliana | Chan et al., 2003; Mathur et al., 2003 | |
| CLASP | Arabidopsis thaliana | Ambrose et al., 2007; Kirik et al., 2007 | |
| MOR/GEM1 | Arabidopsis thaliana | Whittington et al., 2001 | |
| TMBP200 | Nicotiana tabacum (BY-2) | Yasuhara et al., 2002; Hamada et al., 2004 | |
| AUG8 | Arabidopsis thaliana | Cao et al., 2013 | |
| KINESINS AND KINESIN-LIKE PROTEINS | |||
| ATK5 | Arabidopsis thaliana | Ambrose et al., 2005 | Binds also to MT + ends |
| KCH1 (kinesin with calponin homology 1) | Gossypium hirsutum, Oryza sativa | Preuss et al., 2004; Frey et al., 2009 | |
| KCH2 (kinesin with calponin homology 2) | Gossypium hirsutum | Xu et al., 2009 | |
| O12 | Oryza sativa | Umezu et al., 2011 | |
| kinesin 13-A | Nicotiana tabacum | Wei et al., 2005 | |
| KCBP/ZWICKEL | Arabidopsis thaliana | Krishnakumar and Oppenheimer, 1999 | |
| TBK5 | Nicotiana tabacum | Matsui et al., 2001 | |
| AtPAKRP1 | Arabidopsis thaliana, Oryza sativa | Lee and Liu, 2000 | |
| DcKRP120-2 | Daucus carota | Barroso et al., 2000 | |
| TKRP125 | Nicotiana tabacum | Asada et al., 1997 | |
| KINID1 | Physcomitrella patens | Hiwatashi et al., 2014 | |
| KatA | Arabidopsis thaliana | Liu B. et al., 1996 | |
| KatD | Arabidopsis thaliana | Tamura et al., 1999 | |
| OTHER PROTEINS | |||
| TANGLED 1 | Zea mays | Smith et al., 2001 | |
| p60 katanin subunit (AtKSS, AtKN1) | Arabidopsis thaliana | Burk et al., 2001 | |
| p80 katanin subunit | Arabidopsis thaliana | Bouquin et al., 2003 | |
| RUNKEL (RUK) | Arabidopsis thaliana | Krupnova et al., 2009 | |
| Spc98p | Arabidopsis thaliana | Erhardt et al., 2002 | |
| BPP1 | Arabidopsis thaliana | Hamada et al., 2013 | |
| NEDD1 | Arabidopsis thaliana | Zeng et al., 2009 | Acts as an anchoring factor of γ-tubulin complex, decorates spindle MTs preferentially toward theirs minus ends |
| PLANT SPECIFIC MAPS | |||
| MAP190 | Nicotiana tabacum (BY-2) | Igarashi et al., 2000 | |
| MAP70 -1 | Arabidopsis thaliana | Korolev et al., 2005; Pesquet et al., 2010 | |
| MAP70 -2 | Arabidopsis thaliana | Korolev et al., 2005 | |
| MAP70 -3 | Arabidopsis thaliana | Korolev et al., 2005 | |
| MAP70 -4 | Arabidopsis thaliana | Korolev et al., 2005 | |
| MAP70 -5 | Arabidopsis thaliana | Korolev et al., 2005, 2007 | |
| SPR1 | Arabidopsis thaliana | Nakajima et al., 2004; Sedbrook, 2004 | |
| SPR2 | Arabidopsis thaliana | Furutani et al., 2000 | |
| SB401 | Solanum berthaultii | Huang et al., 2007 | |
| SBgLR | Nicotiana tabacum | Liu et al., 2013 | Potato pollen-specific protein |
| Atg8 | Arabidopsis thaliana | Ketelaar et al., 2004 | Homolog of autophagy protein |
| AtMPB2C | Arabidopsis thaliana | Ruggenthaler et al., 2009 | Homolog of MPB2C, involved in the alignment of cortical MT |
| MDP40 | Arabidopsis thaliana | Wang et al., 2012 | Regulator of hypocotyl cell elongation |
| WVD/WDL family | Arabidopsis thaliana | Perrin et al., 2007 | |
| AIR9 | Arabidopsis thaliana | Buschmann et al., 2006 | |
Multifunctional MAPs
Additionally to numerous MAPs, MTs probably interact with other “fine tuning” factors that are most likely required for microtubular functions as well. The function of these proteins is not primarily related to MT-binding, but they may bind to MTs only under specific conditions. The cytoskeleton is a three-dimensional dynamic structure that can be thought of as a framework for compartmentalization of cytosolic regions. Binding of proteins to this scaffold may increase the efficiency of cellular processes by bringing interacting molecules together in place and time. In this case, MTs may function as a passive structure. Additionally, metabolic enzymes such as GAPDH have been shown to modulate MT cytoskeleton (Sirover, 1999). This suggests that multiple proteins may interact with MTs to integrate signaling pathways and the reorganization of microtubules. Along with MT-binding, these proteins perform other, MT-unrelated functions. For this review, we will refer to these proteins as multifunctional MAPs as a way to distinguish them from MAPs that exclusively regulate MT structure and dynamics.
It is possible that multifunctional MAPs lack a well-defined MT-binding domain, are members of larger protein complexes and, therefore, are not found using database-based sequence similarity searches for MAPs. Instead, biochemical methods are required for their detection. The existence of a large number of proteins with known MT-unrelated functions that unexpectedly interacted with MT cytoskeleton has been documented in proteomic searches for MT-interacting proteins (Chuong et al., 2004; Korolev et al., 2005; Hamada et al., 2013; Derbyshire et al., 2015). In these experiments, tens to hundreds of cytosolic proteins interacting with tubulin or MTs were identified. However, conventional MAPs represented only a minor portion of the total MT-interacting fraction. For example, Chuong et al. (2004) used tubulin-affinity chromatography to identify a set of proteins interacting with tubulin. Only 6% of proteins were predicted as known MAPs in this protein group. Similarly, liquid chromatography-tandem mass spectrometry of MAPs-enriched fraction from Arabidopsis suspension cells was used by Hamada et al. (2013) to identify hundreds of proteins. Replication, transcription and translation-associated proteins were enriched here as well (Hamada et al., 2013). Derbyshire et al. (2015) performed a MT-pull-down protein search for MT-interacting proteins exhibiting differential accumulation during tracheary element differentiation; only 3% of proteins were classified as known MAPs (Derbyshire et al., 2015).
On the other hand, the presence of the protein in MAP enriched fraction does not always indicate its direct association with MTs. For each putative multifunctional MAP, the association with MTs or tubulin indicated by biochemical isolation needs to be tested by other methods. In contrast to MAPs, investigating the role of multifunctional MAPs associated with MTs is usually challenging. Multifunctional MAPs often cannot perform their MT-related function alone; their affinity to MTs may be dependent on factors such as upstream or feedback regulations, may be phospho- or ligand-dependent or may be of short duration, e.g., as for Hsp90 (Krtkova et al., 2012). Here we review plant proteins repeatedly found to associate with MTs whose primary function is distinct from MT-binding (Table 2). If possible, we provide a short description of their physiological function in the association with MTs.
Table 2.
List of multifunctional MAPs described in plants.
| MAP | Characterized in | References | Notes |
|---|---|---|---|
| ENZYMES OR CHAPERONS | |||
| GAPDH | Mammalian cells | Sirover, 1999; Tisdale et al., 2009 | |
| Glycolytic enzymes: lactate-dehydrogenase, pyruvate kinase, aldolase and during specific conditions also for glucose-6-phosphate isomerase and phosphoglycerate-kinase | Walsh et al., 1989 | ||
| Hsp70 | Arabidopsis thaliana | Ho et al., 2009 | Also involved in signaling |
| Hsp90 | Koyasu et al., 1986; Sanchez et al., 1988; Williams and Nelsen, 1997; Freudenreich and Nick, 1998; Petrasek et al., 1998; Pratt et al., 1999; Lange et al., 2000; de Carcer et al., 2001; Harrell et al., 2002; Wegele et al., 2004; Glover, 2005; Basto et al., 2007; Weis et al., 2010; Krtkova et al., 2012 | Also involved in signaling | |
| Plant chaperone CCT | Nicotiana tabacum | Nick et al., 2000 | |
| EF1α | Daucus carota | Durso and Cyr, 1994 | |
| EF-2 | Arabidopsis thaliana, suspension cells | Chuong et al., 2004 | |
| PLDδ | Nicotiana tabacum | Gardiner et al., 2001 | Also involved in signaling |
| THO2 | Nicotiana tabacum | Hamada et al., 2009 | Putative RNA-processing THO2 relative protein |
| PROTEINS INTERACTING WITH OTHER CELL STRUCTURES | |||
| Actin Binding Proteins | |||
| FH4 | Arabidopsis thaliana | Deeks et al., 2010 | Also involved in signaling |
| FH14 | Arabidopsis thaliana | Li et al., 2010 | aLso involved in signaling |
| FH1 | Arabidopsis thaliana | Rosero et al., 2013 | Also involved in signaling |
| ARPC2 | Nicotiana tabacum | Havelková et al., 2015 | |
| Proteins Involved in Signaling | |||
| PCaP2 (MAP18) | Arabidopsis thaliana | Wang et al., 2007; Kato et al., 2010 | |
| MDP25 (PCaP1) | Arabidopsis thaliana | Li et al., 2011 | PCAP1, MT destabilizing protein |
| MIDD1 | Arabidopsis thaliana | Oda et al., 2010 | MT-end tracking protein |
Enzymes and chaperones
Chaperone proteins and metabolic enzymes have been repeatedly found in MT-interacting protein fractions. Earlier, these proteins were considered as sample contaminates and their MT-binding activity was neglected. Recently, their interaction with MTs has shown to be of physiological relevance. In Arabidopsis, metabolism-related proteins were predicted to form 21% of the tubulin-interacting fraction (Chuong et al., 2004). Thirteen percent of metabolism-related proteins were detected while searching for MT-interacting proteins exhibiting differential accumulation during tracheary element development (Derbyshire et al., 2015). Nevertheless, only few of these proteins were well-studied. Examples discussed here are glyceraldehyde-3-phosphate-dehydrogenase (GAPDH) (Walsh et al., 1989; Chuong et al., 2004), chaperones Hsp70 and Hsp90 (Freudenreich and Nick, 1998; Ho et al., 2009), plant chaperonin complex CCTε subunit (Nick et al., 2000) and enzyme phospholipase Dδ (PLDδ) (Gardiner et al., 2001).
GAPDH
GAPDH is a conserved glycolytic enzyme that lyses glyceraldehyde-3-phosphate to 1,3 diphosphoglycerate. GAPDH was the first glycolytic enzyme found to be associated with tubulin and with MTs during polymerization/depolymerization cycles (Kumagai and Sakai, 1983; Somers et al., 1990). It was shown to interact directly with MTs in animals (Kumagai and Sakai, 1983; Walsh et al., 1989; Volker and Knull, 1997; Tisdale et al., 2009). Further, GAPDH plays role in vesicle trafficking, MT array arrangement, DNA replication and repair, nuclear RNA export, apoptosis or stress detection in animals (for review, see Sirover, 1999). GAPDH mediates MT-binding of other MT-interactors, such as Rab2 GTPase, therefore, it physically links MTs and membrane structures involved in secretory pathways of metazoans (Tisdale, 2002; Andrade et al., 2004). RabGTPases further interact with motor proteins that modulate vesicle binding to MTs (Hammer and Wu, 2002; Perez et al., 2002; Howard and Hyman, 2003). Thus, GAPDH represents a multifunctional MAP with the ability to recruit a multiprotein complex to MTs in metazoans (for suggested model, see Figure 1). In plants, GAPDH was found together with other metabolic and protein synthesis enzymes, as well as signaling proteins in the tubulin binding protein fraction, which were isolated from Arabidopsis suspension cultures (Chuong et al., 2004). Further, GAPDH interacted with MTs in maize endosperms (Azama et al., 2003). GAPDH was found in the Arabidopsis proteomic screen for MT-binding proteins (Derbyshire et al., 2015). However, the physiological role of plant GAPDH interaction with MTs and probable role in multiprotein complex recruitment to MTs remains unknown.
Figure 1.
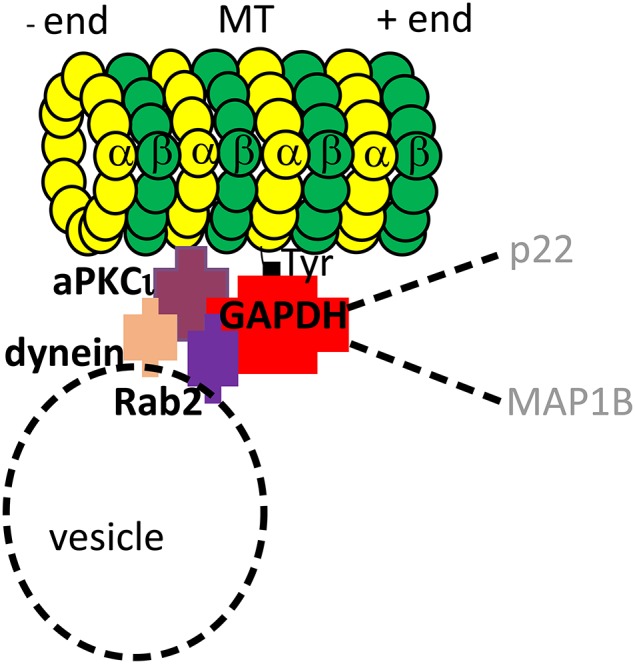
Model of GAPDH interactions in animal cells. GAPDH, athough primarily glycolytic enzyme, is an example of a multifunctional MAP that binds to MTs and recruits a multiprotein complex to them. GAPDH binds directly to C-terminus of α-tubulin (Kumagai and Sakai, 1983). Together with PKCι, an atypical protein kinase Cι, GAPDH recruits Rab2GTPase to MTs. Additionally, Rab2GTPase and PKCι recruit dynein motor protein to the complex, which presumably links the complex to vesicle trafficking (Tisdale, 2002; Tisdale et al., 2009). A broader importance of GAPDH complex is suggested by reported binding of other proteins to the complex, such as p22 (Andrade et al., 2004) and MAP1B (Cueille et al., 2007).
In animals, numerous glycolytic and sucrose metabolism enzymes were found to bind MTs: lactate-dehydrogenase, pyruvate kinase, aldolase, glucose-6-phosphate isomerase, phosphoglycerate-kinase, sucrose synthase, sucrose-UDP glucosyltransferase (Walsh et al., 1989). In plants, enzymes of folate-dependent pathways, fatty acid metabolism, pentose phosphate pathway, phosphate metabolism, amino acid biosynthesis, the tricarboxylic acid cycle, anaerobic glycolysis, and panthothenate biosynthesis enzymes were reported in the tubulin-binding fraction in Arabidopsis (Chuong et al., 2004). The significance of these interactions remains to be elucidated, but, as indicated in animals, the interactions of metabolism–related proteins with MTs signify a promising area of discoveries with high biological importance.
Heat shock protein 90
Heat shock protein 90 (Hsp90) is a highly conserved molecular chaperone essential for protein folding and stability. Along with binding various substrates in animals (Wegele et al., 2004), Hsp90 mediates switches between active and inactive states of regulatory and signaling proteins (Rutherford and Zuker, 1994). In plants, Hsp90 is involved in MAPK cascades (Takabatake et al., 2007) and R-proteins-triggered stress response (Takahashi et al., 2003; Boter et al., 2007). Hsp90 also interacts with actin and tubulin cytoskeleton (Koyasu et al., 1986; Sanchez et al., 1988; Wegele et al., 2004). Due to its numerous substrates and interacting structures including MTs, Hsp90 functions at the interface of several developmental pathways in eukaryotes (Rutherford and Lindquist, 1998).
In animal cells, Hsp90 interaction with MTs appears to be complex; it interacts with tubulin dimers (Sanchez et al., 1988; Weis et al., 2010), with polymerized MTs (Fostinis et al., 1992; Williams and Nelsen, 1997) and, Hsp90 is a subunit of the heterocomplex associated with MTs during the nuclear transport of steroid hormones (Pratt et al., 1999; Harrell et al., 2002). It is found in the centrosome (Lange et al., 2000). Together with other centrosomal proteins, Hsp90 is involved in centrosome assembly and function (de Carcer et al., 2001; Glover, 2005; Basto et al., 2007).
In plants, Hsp90 is known to associate with tubulin dimers, cortical MTs and phragmoplast MTs (Freudenreich and Nick, 1998; Petrasek et al., 1998; Krtkova et al., 2012). Tobacco Hsp90 binds directly to polymerized MTs in vitro (Krtkova et al., 2012). Since the inhibition of Hsp90 severely impairs MT re-assembly after cold-induced de-polymerization, Hsp90 interaction with MTs conceivably plays a role in rapid MT re-assembly important during environmental changes or stress (Krtkova et al., 2012; Figure 2).
Figure 2.
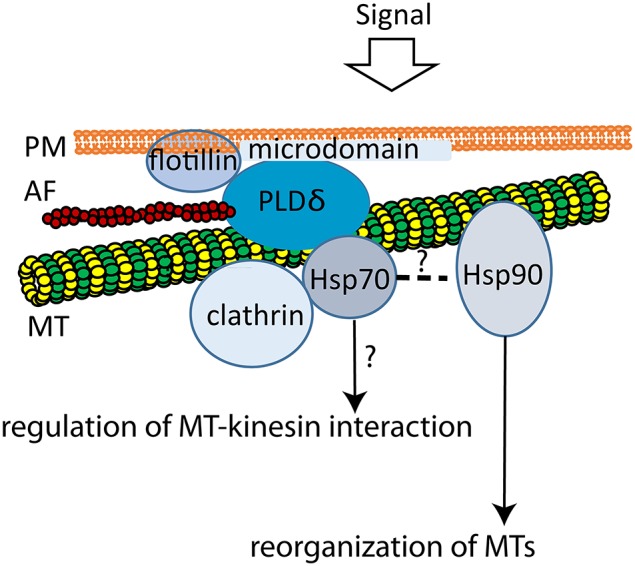
Model of plant MTs interactions with plasma membrane microdomains. The multiprotein complex composed of flotillin, PLDδ, MTs, AFs, Hsp70, and clathrin possibly creates the PM-cytoskeleton continuum and thus may be involved in cell signaling transduction and vesicle trafficking. PLDδ links PM with cortical MTs (Marc et al., 1996; Gardiner et al., 2001) at sites where cell signaling processes take place, since it binds to plant flotillin, a microdomain marker (Ho et al., 2009). Actin, tubulin, Hsp70, Hsp90, and clathrin heavy chain are further PLDδ interactors (Ho et al., 2009). Furthermore, plant MTs interact directly with both Hsp70 (Parrotta et al., 2013), and Hsp90 (Krtkova et al., 2012). Assumed heterocomplex chaperone machinery consisting of Hsp70 and Hsp90 (Pratt et al., 2001) may further control the reorganization of MTs (Hsp90, Krtkova et al., 2012) or interaction with kinesins (Hsp70, Parrotta et al., 2013). Model adapted from Ho et al. (2009).
The cytosolic chaperonin-containing TCP-1 complex
The cytosolic chaperonin-containing TCP-1 complex (CCT), also known as the TCP1 ring complex (TRiC), plays a role in folding of newly synthetized actin and tubulin molecule and in organization of the MT cytoskeleton in mammalian cells (Lewis et al., 1997). In plant cells, its CCTε subunit localizes along phragmoplast MTs and cortical bundles that accompany secondary-wall thickenings (Nick et al., 2000). It is possible that CCTε is involved in the reorganization of microtubular cytoskeleton by regulating tubulin folding (Moser et al., 2000).
Heat shock protein 70
Proteins of Hsp70 family are involved in a range of cellular processes, predominantly under stress conditions, such as heat. They prevent protein aggregation, assist in protein refolding, import and translocation, signal transduction and transcriptional activation (for review, see (Zhang and Glaser, 2002; Wang et al., 2004). In the plant cortical region, Hsp70 associates with MTs and tubulin, as well as with PLDδ (Ho et al., 2009). In Chlamydomonas, the failure of the Hsp70-Hsp40 chaperone system to recognize or fold the client protein(s) results in increased MT stability and resistance to the MT-destabilizing effect of the herbicides (Silflow et al., 2011). Parrotta et al. (2013) identified a Hsp70 isoform in the pollen tube of tobacco that binds to MTs in an ATP-dependent manner. Interestingly, Hsp70 binding to MTs was also dependent on the binding of a kinesin motor p90 (Parrotta et al., 2013). This raises a possibility that Hsp70 may modulate kinesin action on MTs, a phenomenon observed also in other systems (Terada et al., 2010).
Protein translation machinery proteins
The interaction of cytoskeleton with polysomes was first identified in the 1970's. Since then, many data supporting the role of actin and MTs in metazoan translation machinery localization and regulation were published (for review see Kim and Coulombe, 2010). Plant transcription machinery seems to interact with the cytoskeleton as well (Muench and Park, 2006). In proteomic screens performed in plants, large groups of proteins interacting with MTs are primarily involved in RNA transcription processes. For example, Chuong et al. (2004) predicted 21% of tubulin-binding proteins assist in RNA binding and 19% in translation. Similarly, in the screen of MT-associated proteins with changed expression during tracheary element differentiation, 13% of isolated proteins were predicted to be involved in protein synthesis, and 19% in DNA or RNA binding (Derbyshire et al., 2015). In a model proposed for plant cells, the predominant role in the transport and localization of translation machinery components is assigned to actin cytoskeleton, whereas MTs may anchor and perhaps influence the translation process (Muench and Park, 2006). Indeed, some proteins participating in the translation are repeatedly reported to associate with MTs or tubulin. Here, elongation factor 1α and THO2 proteins are discussed.
Elongation factor 1α
Elongation factor 1α (EF1α) is a translational factor that binds aminoacyl-tRNA and ribosomes in a GTP-dependent manner (Carneiro et al., 1999). Additionally, EF1α was reported to bind and to bundle actin filaments (AFs) in animal cells (Murray et al., 1996). It is believed that F-actin-bound EF1α is translationally inactive, since F-actin sequesters elongation factor 1α from the interaction with aminoacyl-tRNA in a pH-dependent reaction (Liu G. et al., 1996). This suggests the role of EF1α binding to F-actin in the regulation of proteosynthesis. In addition to this, this protein was shown to sever MTs (Shiina et al., 1994). EF1α is repeatedly present in plant MAP-enriched protein fractions (for example Durso and Cyr, 1994; Chuong et al., 2004; Hamada et al., 2013; Derbyshire et al., 2015). EF1α influences AF dynamics (Murray et al., 1996) and MT dynamics in Ca2+ and calmodulin-dependent manner (Durso and Cyr, 1994; Moore et al., 1998). Since Ca2+ and calmodulin are key players in plant cell signaling, EF1α regulation of cytoskeletal dynamics can serve as a manual transmission stick connecting the cytoskeleton and plant developmental and signaling pathways.
THO2
In animal and yeast cells, THO2 is part of the THO-TREX complex that participates in mRNA metabolism and nuclear export (Koehler and Hurt, 2007). Hamada et al. (2009) described tobacco putative THO2-related protein (NtTHO2) as a MT-associated protein which binds directly to MTs in vitro and co-localizes with cortical MTs in vivo, indicating its role in translation targeted to specific plant cell compartments.
Plasma membrane interacting proteins
In plant cells, cortical MTs underlie the plasma membrane (PM) (Dixit and Cyr, 2004). The association of cortical MTs to the PM is related to the guiding of cellulose synthase complexes (CESAs), enzyme complexes in the plasma membrane that synthesize cellulose into the extracellular space (Paredez et al., 2006). Surprisingly, only a subtle number of proteins were proven to mediate the interaction of cortical MTs with the plasma membrane. In this section, phospholipase D and developmentally-regulated plasma membrane polypeptide (DREPP) proteins are discussed. Both were first reported to participate in MT unrelated processes, however, their roles associated with MTs were revealed later. Some plant formins were reported to interact with the PM and MTs as well. Being primarily actin-associated proteins, they are discussed later in a separate chapter.
Phospholipase D
PLDs with N-terminal lipid binding domain are important signaling enzymes in plant cells (Munnik, 2001; Elias et al., 2002). Various PLD isoforms differ in their affinity to different substrates—membrane phospholipids. These are cleaved by PLDs to produce signaling molecules (Munnik, 2001; Wang, 2002).
Phospholipase D δ (PLDδ) is a central enzyme of phospholipid signaling in plants. It cleaves plasma membrane (PM) phospholipids to produce phosphatidic acid (PA) and predominantly ethanolamine and choline (for review, see Wang, 2002). PLDδ isoform strongly associates with PM (Gardiner et al., 2001; Wang and Wang, 2001) and connects it physically with cortical MTs (Marc et al., 1996; Gardiner et al., 2001). Upon stress, e.g., NaCl, hypoosmotic stress, xylanase or mastoparane treatment, PLDδ is activated and triggers MT reorganization (Dhonukshe et al., 2003). The mechanism of PLD-triggered reorganization is likely based on the activation of PLD on the plasma membrane, which leads to the release of MTs from the membrane and MTs reorientation (Dhonukshe et al., 2003). Another potential mechanism of PLD-based MT reorganization mechanism may involve the role of PLD signaling product, PA, on MT (for review see Pleskot et al., 2014).
The importance of PLDδ in plants is confirmed by the plasma membrane and MT-binding discussed above and its interaction with actin (Ho et al., 2009). Phospholipase Dδ is thus discussed hereinafter as an example of a protein potentially integrating multiple structures into a functional complex in plants.
PCaP1/MDP25 and PCaP2/MAP18
DREPP (Developmentally-Regulated Plasma membrane Polypeptide) proteins include a family of plant-specific proteins that interact with the plasma membrane (Gantet et al., 1996). Arabidopsis DREPP family contains proteins PCaP1 named also MDP25 (Ide et al., 2007; Li et al., 2011), and a divergent paralog PCaP2, first described as a Microtubule-Associated Protein 18 kDa MAP18 (Wang et al., 2007; Kato et al., 2010). PCaP1/MDP25 links calcium signaling to the regulation of cytoskeleton organization. Under normal conditions, PCaP1/MDP25 is localized to the plasma membrane. Increased calcium levels cause PCaP1/MDP25 to partially dissociate from the plasma membrane and to move into the cytosol. In the hypocotyl, cytosolic PCaP1/MDP25 binds and destabilizes cortical MTs by depolymerization and subsequently inhibits hypocotyl cell elongation (Li et al., 2011). In the subapical region of pollen tubes, PCaP1/MDP25 binds directly to actin cytoskeleton and severs individual actin filaments, thus negatively regulating pollen tube growth (Qin et al., 2014). PCaP2, previously reported as MT-binding MAP18 (Wang et al., 2007), is a plant-specific protein found only in Arabidopsis that is involved in intracellular signaling in growing root hairs and pollen tubes. PCaP2/MAP18 is localized in plasma membranes possibly via N-myristoylation, and destabilizes MTs (Keech et al., 2010). It is associated with specific PtdInsPs and it exhibits the capacity to bind calcium and calcium–calmodulin (Ca2+–CaM) complex (Kato et al., 2010). It is possible that association and/or dissociation of PCaP2/MAP18 with PtdInsPs via oscillation in Ca2+ cytosolic concentration regulate the signaling function of PtdIns(4,5)P2, which includes regulation of ion channels (Suh and Hille, 2008), cytoskeletal organization and membrane traffic (Meijer and Munnik, 2003; Lee et al., 2007; Kato et al., 2010).
Actin binding proteins as multifunctional MAPs
In plants, AFs are crucial for cell polarity, division, membrane trafficking and thus, growth and development. Their organization and dynamics is modulated by actin binding proteins, such as formins, Arp2/3 complex, profilin, cofilin, myosin etc. (for review see Thomas et al., 2009). An increasing list of proteins interacting with both actin and MTs in plants was reported (for review see Petrasek and Schwarzerova, 2009). The existence of proteins interacting with both AF and MT is not surprising, since actin-cytoskeletal functions are fulfilled in a close collaboration with MT cytoskeleton (Collings, 2008; Smertenko et al., 2010; Sampathkumar et al., 2011), e.g., during plant cell division, in PPB and phragmoplast (Traas et al., 1987; Mineyuki, 1999; Sano et al., 2005; Wu and Bezanilla, 2014). The following examples were reported to interact with both AFs and MTs: plant formins (Deeks et al., 2010; Li et al., 2010), a subunit of ARP2/3 protein complex ARPC2 (Havelková et al., 2015), conventional MAPs, such as kinesins (Preuss et al., 2004; Frey et al., 2009; Klotz and Nick, 2012; Schneider and Persson, 2015), plant specific MAPs, such as 190 kDa polypeptide (Igarashi et al., 2000) and SB401 in Solanaceae (Huang et al., 2007), enzyme PLDδ (Ho et al., 2009) or protein DREPP/AtPCaP1/MDP25 (Li et al., 2011; Qin et al., 2014). Here, proteins with primary functions related to actin cytoskeleton organization that were found to interact also with MTs (formins and ARPC2) are discussed.
AtFH4 and AFH14
Formins nucleate actin and contribute to the filament growth, thus, they participate in cell polarity, morphogenesis and division (Sagot et al., 2002; Kovar and Pollard, 2004; Pruyne et al., 2004; Ingouff et al., 2005). However, some plant formins also bind directly to MTs (Deeks et al., 2010; Li et al., 2010). Formins are characterized by the presence of formin homology-2 and formin homology-1 (FH2 and FH1, respectively) domains that are common in mammals and plants (Blanchoin and Staiger, 2010). Besides FH1 and FH2 domains important for actin nucleation, plant AtFH4 contains a plant-specific transmembrane domain, and a specific GOE domain that binds directly to MTs (Deeks et al., 2010). Thus, AtFH4 represents a protein that links both membranes, MTs and AFs in plant cells. Another plant formin called FORMIN14 (AFH14) appeared to bind directly either AF or MTs (Li et al., 2010). Unlike AtFH4, the FH2 domain of AFH14 is critical for both MT and AF binding and bundling. AFH14 localized to typical plant MT structures, such as preprophase band (PPB), spindles, or phragmoplast. MTs competed with AFs to bind AFH14, and the overexpression of AFH14 caused co-alignment of MTs with AFs, which perturbed the progress of cell division (Li et al., 2010). Therefore, actin-binding proteins formin AtFH4 and AFH14 represent multifunctional MAPs with specialized function in mediating AF and MT crosstalk.
Actin related protein 2/3 complex subunit 2
Arp2/3 complex represents the second mechanism of AF nucleation. This evolutionarily conserved complex of 7 subunits (Welch et al., 1997) is functional also in plants, where it controls several aspects of plant morphogenesis (for review see Deeks and Hussey, 2005; Yanagisawa et al., 2013). Recently, it has been shown that actin related protein 2/3 complex subunit 2 (ARPC2) of Arp2/3 complex binds directly to MTs. It is possible that the ARPC2 subunit of Arp2/3 complex mediates the interaction between MTs and AFs in plants (Havelková et al., 2015). Alternatively, Arp2/3-based interaction of MTs and AFs may contribute to mutual dynamic regulation of AFs and MTs. ARPC2 protein thus, represents another multifunctional MAP with the primary role unrelated to MT binding.
Multifunctional MAPs involved in signaling crosstalks
Stimuli from the outer environment are transferred into the plant cell across the rigid cellulose cell wall and lipid plasma membrane. Specific receptors on the plasma membrane may transfer stimuli by the cell wall-PM-cortical MT continuum. According to the recent studies, mediators in this physical continuum may be PLDδ and/or plant-specific formins with transmembrane domains. These proteins bind to the plasma membrane, are associated with cortical MTs and actin, and as in the case of AtFH4, possess extracellular extensin-like motifs that can anchor it to the cell wall compounds (Baluska and Hlavacka, 2005; Deeks et al., 2005, 2010; Ho et al., 2009; Cvrckova, 2013). Formins can further mediate attachment of endomembrane compartments, such as the ER or secretory vesicles, to the MT cytoskeleton (Cvrckova et al., 2015).
In addition to binding to PM phospholipids, PLDδ binds to plant flotillin homolog (Ho et al., 2009), a lipid microdomain marker (Martin et al., 2005). Lipid microdomains are PM detergent-resistant regions that are important for the assembly of multimolecular signaling complexes containing G-proteins or kinases (Martin et al., 2005; Dunkley et al., 2006; Tapken and Murphy, 2015). Therefore, PLDδ may link PM with MTs at sites where cell-signaling processes take place. However, PM and MTs are not PLDδ's sole interacting structures. F-actin (Kusner et al., 2003), Actin 7, Hsp70, ATPase and clathrin heavy chain (Ho et al., 2009) were reported as PLDδ interactors, too. Thus, by interacting with both cytoskeletal networks, PLDδ is a possible mediator in the cell wall-PM-cytoskeleton continuum. Its additional interaction with Hsp70 may contribute to the signal transduction to the cytosol (Ho et al., 2009). This interaction is probably mediated by MTs. As authors hypothesize, multiprotein complexes composed of flotillin, PLDδ, MTs, AFs, Hsp70, and clathrin indirectly bound to PLD are involved in cell signaling and vesicle trafficking (Ho et al., 2009).
Hsp70 and Hsp90 cooperate in the signaling, protein folding, stabilization, and turnover by the formation of multichaperone complexes (Pratt et al., 2001, 2010). They have been reported as tubulin interactors as well (Freudenreich and Nick, 1998; Garnier et al., 1998; Petrasek et al., 1998; Parrotta et al., 2013). Since Hsp90 localizes to the cortical MTs and was isolated as the protein interacting with both PM and MTs (Krtkova et al., 2012), it is likely that the whole complex composed of Hsp90, Hsp70, MTs and actin is linked to the lipid microdomain platforms by PLDδ. By this mean, the PM-cytoskeleton continuum involved in cell signaling may be established (Figure 2).
Formin interacting with both AFs and MTs (AtFH4) is possibly involved in cell signaling as well. Via its extracellular and transmembrane domain, it may transduce mechanical stimuli from the plant cell wall across PM to both cytoskeletal networks. According to the model for AtFH4 proposed in Deeks et al. (2010; see also Figure 3), mechanical stimuli transduced by formin-mediating PM-cytoskeleton continuum may be preferentially manifested in actin dynamics changes. In this hypothesis, MTs represent structural scaffolds enabling FH2 domain of AtFH4 to perform its actin-nucleating function. AtFH4 also co-aligns MTs with endoplasmic reticulum, suggesting a further role of AtFH4 at the interface of actin and MT cytoskeleton (Deeks et al., 2010).
Figure 3.
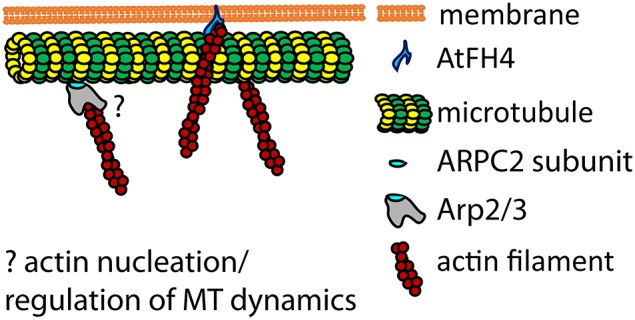
Model of plant MTs interactions with actin filaments based on actin nucleators. Formin AtFH4 is anchored in the PM and binds to AFs, providing a supportive scaffold for MTs attachment (Deeks et al., 2010). ARPC2 subunit of Arp2/3 complex binds directly to MTs (Havelková et al., 2015). The role of ARPC2-MTs interaction in the cross-linking of AFs and MTs through Arp2/3 complex, or in the regulation of AFs and MTs dynamics, remains to be elucidated (question mark). Model adapted from Deeks et al. (2010).
Physiological demonstration of multifunctional MAPs-MT interaction—future prospects
Stability, dynamics and organization of MTs is modulated by their interacting proteins. MAPs, such as +TIPs (for review, see e.g., Akhmanova and Steinmetz, 2008) or other structural MAPs (Gardiner, 2013) coordinate MT reorganization events spatiotemporally, thus controlling the localization of MTs in the plant cell during specific environmental or developmental conditions. Such events underlie plant cell shape changes and plant tissue differentiation, determining survival of the plant organism through proper growth regulation.
MTs apparently require further mediating-proteins as well. These mediators may be the traditional motor or structural MAPs, but also proteins with another primary function than MT-binding. These proteins may interact with MTs in short time limits, under specific environmental conditions or interact with MTs weakly or indirectly as members of MT-associated structures or complexes. Some of these proteins were discussed in this review. These spatiotemporally tightly regulated physiological functions, or secondary interactions, as well as functions of single complex subunits, are difficult to detect. Nevertheless, important progress has been made recently in identifying new multifunctional MAPs; new proteins will be added to the list in the future. Plant hormone signaling, stress and pathogen response, development of specific morphological structures and other plant specific processes represent areas for investigating new highly specific MT-associated proteins. Investigating into the functional interactions between MTs and both protein synthesis machinery and metabolism-related enzymes in plants is an exciting research area awaiting deeper exploration.
Author contributions
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Alexander Paredez for supportive working environment. We highly appreciate the English language and style editing made by Kelly Hennessey. We also thank Lukáš Fischer and Jan Petrášek for stimulating discussions and continuous interest in the project. The work was supported by the project NPUI (LO1417) of Czech Ministry of Education, Youth and Sports and by the grants of Charles University GAUK to JK and MB [grant numbers 82710/2010 and 990213/2012].
Glossary
Abbreviations
- CCT
cytosolic chaperonin containing TCP-1
- EF1α
elongation factor 1α
- EF2
elongation factor 2
- FH2
formin homology 2
- GAPDH
glyceraldehyde-3-phosphate-dehydrogenase
- GDA
geldanamycin
- MAP
microtubule-associated protein
- MDP
microtubule destabilizing protein
- MT
microtubule
- PA
phosphatidic acid
- PcaP
plasma membrane-associated Ca2+-binding protein
- PLD
phospholipase δ
- PPB
preprophase band
- +TIP
+tip interacting protein.
References
- Akhmanova A., Steinmetz M. O. (2008). Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat. Rev. Mol. Cell Biol. 9, 309–322. 10.1038/nrm2369 [DOI] [PubMed] [Google Scholar]
- Ambrose J. C., Li W. X., Marcus A., Ma H., Cyr R. (2005). A minus-end-directed kinesin with plus-end trackin protein activity is involved in spindle morphogenesis. Mol. Biol. Cell 16, 1584–1592. 10.1091/mbc.E04-10-0935 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ambrose J. C., Shoji T., Kotzer A. M., Pighin J. A., Wasteneys G. O. (2007). The Arabidopsis CLASP gene encodes a microtubule-associated protein involved in cell expansion and division. Plant Cell 19, 2763–2775. 10.1105/tpc.107.053777 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrade J., Pearce S. T., Zhao H., Barros M. (2004). Interactions among p22, glyceraldehyde-3-phosphate dehydrogenase and microtubules. Biochem. J. 384, 327–336. 10.1042/BJ20040622 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asada T., Kuriyama R., Shibaoka H. (1997). TKRP125, a kinesin-related protein involved in the centrosome-independent organization of the cytokinetic apparatus in tobacco BY-2 cells. J. Cell Sci. 110, 179–189. [DOI] [PubMed] [Google Scholar]
- Azama K., Abe S., Sugimoto H., Davis E. (2003). Lysine-containing proteins in maize endosperm: a major contribution from cytoskeleton-associated carbohydrate-metabolizing enzymes. Planta 217, 628–638. 10.1007/s00425-003-1016-5 [DOI] [PubMed] [Google Scholar]
- Baluska F., Hlavacka A. (2005). Plant formins come of age: something special about cross-walls. New Phytol. 168, 499–503. 10.1111/j.1469-8137.2005.01595.x [DOI] [PubMed] [Google Scholar]
- Barroso C., Chan J., Allan V., Doonan J., Hussey P., Lloyd C. (2000). Two kinesin-related proteins associated with the cold-stable cytoskeleton of carrot cells: characterization of a novel kinesin, DcKRP120-2. Plant J. 24, 859–868. 10.1046/j.1365-313x.2000.00937.x [DOI] [PubMed] [Google Scholar]
- Basto R., Gergely F., Draviam V. M., Ohkura H., Liley K., Raff J. W. (2007). Hsp90 is required to localise cyclin B and Msps/ch-TOG to the mitotic spindle in Drosophila and humans. J. Cell Sci. 120, 1278–1287. 10.1242/jcs.000604 [DOI] [PubMed] [Google Scholar]
- Blanchoin L., Staiger C. J. (2010). Plant formins: diverse isoforms and unique molecular mechanism. Biochim. Biophys. Acta. Mol. Cell Res. 1803, 201–206. 10.1016/j.bbamcr.2008.09.015 [DOI] [PubMed] [Google Scholar]
- Boter M., Amigues B., Peart J., Breuer C., Kadota Y., Casais C., et al. (2007). Structural and functional analysis of SGT1 reveals that its interaction with HSP90 is required for the accumulation of Rx, an R protein involved in plant immunity. Plant Cell 19, 3791–3804. 10.1105/tpc.107.050427 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouquin T., Mattsson O., Naested H., Foster R., Mundy J. (2003). The Arabidopsis lue1 mutant defines a katanin p60 ortholog involved in hormonal control of microtubule orientation during cell growth. J. Cell Sci. 116, 791–801. 10.1242/jcs.00274 [DOI] [PubMed] [Google Scholar]
- Burk D. H., Liu B., Zhong R. Q., Morrison W. H., Ye Z. H. (2001). A katanin-like protein regulates normal cell wall biosynthesis and cell elongation. Plant Cell 13, 807–827. 10.1105/tpc.13.4.807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buschmann H., Chan J., Sanchez-Pulido L., Andrade-Navarro M. A., Doonan J. H., Lloyd C. W. (2006). Microtubule-associated AIR9 recognizes the cortical division site at preprophase and cell-plate insertion. Curr. Biol. 16, 1938–1943. 10.1016/j.cub.2006.08.028 [DOI] [PubMed] [Google Scholar]
- Buschmann H., Lloyd C. W. (2008). Arabidopsis mutants and the network of microtubute-associated functions. Mol. Plant 1, 888–898. 10.1093/mp/ssn060 [DOI] [PubMed] [Google Scholar]
- Cao L., Wang L., Zheng M., Cao H., Ding L., Zhang X., et al. (2013). Arabidopsis AUGMIN subunit8 is a microtubule plus-end binding protein that promotes microtubule reorientation in hypocotyls. Plant Cell 25, 2187–2201. 10.1105/tpc.113.113472 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Carneiro N. P., Hughes P. A., Larkins B. A. (1999). The eEFlA gene family is differentially expressed in maize endosperm. Plant Mol. Biol. 41, 801–813. 10.1023/A:1006391207980 [DOI] [PubMed] [Google Scholar]
- Chan J., Calder G. M., Doonan J. H., Lloyd C. W. (2003). EB1 reveals mobile microtubule nucleation sites in Arabidopsis. Nat. Cell Biol. 5, 967–971. 10.1038/ncb1057 [DOI] [PubMed] [Google Scholar]
- Chuong S. D. X., Good A. G., Taylor G. J., Freeman M. C., Moorhead G. B. G., Muench D. G. (2004). Large-scale identification of tubulin-binding proteins provides insight on subcellular trafficking, metabolic channeling, and signaling in plant cells. Mol. Cell. Proteomics 3, 970–983. 10.1074/mcp.M400053-MCP200 [DOI] [PubMed] [Google Scholar]
- Collings D. A. (2008). Crossed-wires: interactions and cross-talk between the microtubule and microfilament networks in plants, in Plant Microtubules: Development and Flexibility, 2nd Edn., Vol. 11, ed Nick P. (Berlin; Heidelberg: Springer; ), 47–79. [Google Scholar]
- Cueille N., Blanc C. T., Riederer I. M., Riederer B. M. (2007). Microtubule-associated protein 1B binds glyceraldehyde-3-phosphate dehydrogenase. J. Proteome Res. 6, 2640–2647. 10.1021/pr070081z [DOI] [PubMed] [Google Scholar]
- Cvrckova F. (2013). Formins and membranes: anchoring cortical actin to the cell wall and beyond. Front. Plant Sci. 4:436. 10.3389/fpls.2013.00436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cvrckova F., Oulehlova D., Zarsky V. (2015). Formins: linking cytoskeleton and endomembranes in plant cells. Int. J. Mol. Sci. 16, 1–18. 10.3390/ijms16010001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Carcer G., Avides M. D., Lallena M. J., Glover D. M., Gonzalez C. (2001). Requirement of Hsp90 for centrosomal function reflects its regulation of Polo kinase stability. EMBO J. 20, 2878–2884. 10.1093/emboj/20.11.2878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deeks M. J., Cvrckova F., Machesky L. M., Mikitova V., Ketelaar T., Zarsky V., et al. (2005). Arabidopsis group Ie formins localize to specific cell membrane domains, interact with actin-binding proteins and cause defects in cell expansion upon aberrant expression. New Phytol. 168, 529–540. 10.1111/j.1469-8137.2005.01582.x [DOI] [PubMed] [Google Scholar]
- Deeks M. J., Fendrych M., Smertenko A., Bell K. S., Oparka K., Cvrckova F., et al. (2010). The plant formin AtFH4 interacts with both actin and microtubules, and contains a newly identified microtubule-binding domain. J. Cell Sci. 123, 1209–1215. 10.1242/jcs.065557 [DOI] [PubMed] [Google Scholar]
- Deeks M. J., Hussey P. J. (2005). Arp2/3 and scar: plants move to the fore. Nat. Rev. Mol. Cell Biol. 6, 954–964. 10.1038/nrm1765 [DOI] [PubMed] [Google Scholar]
- Derbyshire P., Menard D., Green P., Saalbach G., Buschmann H., Lloyd C. W., et al. (2015). Proteomic analysis of microtubule interacting proteins over the course of xylem tracheary element formation in Arabidopsis. Plant Cell 27, 2709–2726. 10.1105/tpc.15.00314 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhonukshe P., Laxalt A. M., Goedhart J., Gadella T. W. J., Munnik T. (2003). Phospholipase D activation correlates with microtubule reorganization in living plant cells. Plant Cell 15, 2666–2679. 10.1105/tpc.014977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dixit R., Cyr R. (2004). The cortical microtubule array: from dynamics to organization. Plant Cell 16, 2546–2552. 10.1105/tpc.104.161030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunkley T. P. J., Hester S., Shadforth I. P., Runions J., Weimar T., Hanton S. L., et al. (2006). Mapping the Arabidopsis organelle proteome. Proc. Natl. Acad. Sci. U.S.A. 103, 6518–6523. 10.1073/pnas.0506958103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durso N. A., Cyr R. J. (1994). A Calmodulin-sensitive interaction between microtubules and a higher-plant homolog of elongation factor-1-alpha. Plant Cell 6, 893–905. 10.1105/tpc.6.6.893 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elias M., Potocky M., Cvrckova F., Zarsky V. (2002). Molecular diversity of phospholipase D in angiosperms. BMC Genomics 3:2. 10.1186/1471-2164-3-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erhardt M., Stoppin-Mellet V., Campagne S., Canaday J., Mutterer J., Fabian T., et al. (2002). The plant Spc98p homologue colocalizes with gamma-tubulin at microtubule nucleation sites and is required for microtubule nucleation. J. Cell Sci. 115, 2423–2431. [DOI] [PubMed] [Google Scholar]
- Fostinis Y., Theodoropoulos P. A., Gravanis A., Stournaras C. (1992). Heat-shock protein HSP90 and its association with the cytoskeleton - a morphological-study. Biochem. Cell Biol. 70, 779–786. 10.1139/o92-118 [DOI] [PubMed] [Google Scholar]
- Freudenreich A., Nick P. (1998). Microtubular organization in tobacco cells: heat-shock protein 90 can bind to tubulin in vitro. Bot. Acta 111, 273–279. 10.1111/j.1438-8677.1998.tb00708.x [DOI] [Google Scholar]
- Frey N., Klotz J., Nick P. (2009). Dynamic bridges a calponin-domain kinesin from rice links actin filaments and microtubules in both cycling and non-cycling cells. Plant Cell Physiol. 50, 1493–1506. 10.1093/pcp/pcp094 [DOI] [PubMed] [Google Scholar]
- Furutani I., Watanabe Y., Prieto R., Masukawa M., Suzuki K., Naoi K., et al. (2000). The SPIRAL genes are required for directional central of cell elongation in Arabidopsis thaliana. Development 127, 4443–4453. [DOI] [PubMed] [Google Scholar]
- Gaillard J., Neumann E., Van Damme D., Stoppin-Mellet V., Ebel C., Barbier E., et al. (2008). Two microtubule-associated proteins of Arabidopsis MAP65s promote antiparallel microtubule bundling. Mol. Biol. Cell 19, 4534–4544. 10.1091/mbc.E08-04-0341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gantet P., Masson F., Domergue O., Marquismention M., Bauw G., Inze D., et al. (1996). Cloning of a cDNA encoding a developmentally regulated 22 kDa polypeptide from tobacco leaf plasma membrane. Biochem. Mol. Biol. Int. 40, 469–477. 10.1080/15216549600201033 [DOI] [PubMed] [Google Scholar]
- Gardiner J. (2013). The evolution and diversification of plant microtubule-associated proteins. Plant J. 75, 219–229. 10.1111/tpj.12189 [DOI] [PubMed] [Google Scholar]
- Gardiner J. C., Harper J. D. I., Weerakoon N. D., Collings D. A., Ritchie S., Gilroy S., et al. (2001). A 90-kD phospholipase D from tobacco binds to microtubules and the plasma membrane. Plant Cell 13, 2143–2158. 10.1105/tpc.13.9.2143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garnier C., Barbier P., Gilli R., Lopez C., Peyrot V., Briand C. (1998). Heat-shock protein 90 (hsp90) binds in vitro to tubulin dimer and inhibits microtubule formation. Biochem. Biophys. Res. Commun. 250, 414–419. 10.1006/bbrc.1998.9319 [DOI] [PubMed] [Google Scholar]
- Glover D. M. (2005). Polo kinase and progression through M phase in Drosophila: a perspective from the spindle poles. Oncogene 24, 230–237. 10.1038/sj.onc.1208279 [DOI] [PubMed] [Google Scholar]
- Hamada T. (2014). Microtubule organization and microtubule-associated proteins in plant cells. Int. Rev. Cell Mol. Biol. 312, 1–52. 10.1016/B978-0-12-800178-3.00001-4 [DOI] [PubMed] [Google Scholar]
- Hamada T., Igarashi H., Itoh T. J., Shimmen T., Sonobe S. (2004). Characterization of a 200 kDa microtubule-associated protein of tobacco BY-2 cells, a member of the XMAP215/MOR1 family. Plant Cell Physiol. 45, 1233–1242. 10.1093/pcp/pch145 [DOI] [PubMed] [Google Scholar]
- Hamada T., Igarashi H., Taguchi R., Fujiwara M., Fukao Y., Shimmen T., et al. (2009). The Putative RNA-processing protein, THO2, is a microtubule-associated protein in tobacco. Plant Cell Physiol. 50, 801–811. 10.1093/pcp/pcp024 [DOI] [PubMed] [Google Scholar]
- Hamada T., Nagasaki-Takeuchi N., Kato T., Fujiwara M., Sonobe S., Fukao Y., et al. (2013). Purification and characterization of novel microtubule-associated proteins from arabidopsis cell suspension cultures. Plant Physiol. 163, 1804–1816. 10.1104/pp.113.225607 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammer J. A., Wu X. F. S. (2002). Rabs grab motors: defining the connections between Rab GTPases and motor proteins. Curr. Opin. Cell Biol. 14, 69–75. 10.1016/S0955-0674(01)00296-4 [DOI] [PubMed] [Google Scholar]
- Harrell J. M., Kurek I., Breiman A., Radanyi C., Renoir J. M., Pratt W. B., et al. (2002). All of the protein interactions that link steroid receptor center dot hsp90 center dot immunophilin heterocomplexes to cytoplasmic dynein are common to plant and animal cells. Biochemistry 41, 5581–5587. 10.1021/bi020073q [DOI] [PubMed] [Google Scholar]
- Hashimoto T. (2015). Microtubules in plants. Arabidopsis Book 13, e0179. 10.1199/tab.0179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Havelková L., Nanda G., Martinek J., Bellinvia E., Sikorová L., Šlajcherová K., et al. (2015). Arp2/3 complex subunit ARPC2 binds to microtubules. Plant Sci. 241, 96–108. 10.1016/j.plantsci.2015.10.001 [DOI] [PubMed] [Google Scholar]
- Hiwatashi Y., Sato Y., Doonan J. H. (2014). Kinesins have a dual function in organizing microtubules during both tip growth and cytokinesis in Physcomitrella patens. Plant Cell 26, 1256–1266. 10.1105/tpc.113.121723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho A. Y. Y., Day D. A., Brown M. H., Marc J. (2009). Arabidopsis phospholipase D delta as an initiator of cytoskeleton-mediated signalling to fundamental cellular processes. Funct. Plant Biol. 36, 190–198. 10.1071/FP08222 [DOI] [PubMed] [Google Scholar]
- Howard J., Hyman A. A. (2003). Dynamics and mechanics of the microtubule plus end. Nature 422, 753–758. 10.1038/nature01600 [DOI] [PubMed] [Google Scholar]
- Huang S. L., Jin L. F., Du J. Z., Li H., Zha Q., Ou G. S., et al. (2007). SB401, a pollen-specific protein from Solanum berthaultii, binds to and bundles microtubules and F-actin. Plant J. 51, 406–418. 10.1111/j.1365-313X.2007.03153.x [DOI] [PubMed] [Google Scholar]
- Ide Y., Nagasaki N., Tomioka R., Suito M., Kamiya T., Maeshima M. (2007). Molecular properties of a novel, hydrophilic cation-binding protein associated with the plasma membrane. J. Exp. Bot. 58, 1173–1183. 10.1093/jxb/erl284 [DOI] [PubMed] [Google Scholar]
- Igarashi H., Orii H., Mori H., Shimmen T., Sonobe S. (2000). Isolation of a novel 190 kDa protein from tobacco BY-2 cells: possible involvement in the interaction between actin filaments and microtubules. Plant Cell Physiol. 41, 920–931. 10.1093/pcp/pcd015 [DOI] [PubMed] [Google Scholar]
- Ingouff M., Gerald J. N. F., Guerin C., Robert H., Sorensen M. B., Van Damme D., et al. (2005). Plant formin AtFH5 is an evolutionarily conserved actin nucleator involved in cytokinesis. Nat. Cell Biol. 7, 374–380. 10.1038/ncb1238 [DOI] [PubMed] [Google Scholar]
- Jiang C. J., Sonobe S. (1993). Identification and preliminary characterization of A 65-kDA higher-plant microtubule-associated protein. J. Cell Sci. 105, 891–901. [DOI] [PubMed] [Google Scholar]
- Kato M., Nagasaki-Takeuchi N., Ide Y., Maeshima M. (2010). An Arabidopsis hydrophilic Ca-2-binding protein with a PEVK-rich domain, PCaP2, is associated with the plasma membrane and interacts with calmodulin and phosphatidylinositol phosphates. Plant Cell Physiol. 51, 366–379. 10.1093/pcp/pcq003 [DOI] [PubMed] [Google Scholar]
- Keech O., Pesquet E., Gutierrez L., Ahad A., Bellini C., Smith S. M., et al. (2010). Leaf senescence is accompanied by an early disruption of the microtubule network in Arabidopsis. Plant Physiol. 154, 1710–1720. 10.1104/pp.110.163402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ketelaar T., Voss C., Dimmock S. A., Thumm M., Hussey P. J. (2004). Arabidopsis homologues of the autophagy protein Atg8 are a novel family of microtubule binding proteins. FEBS Lett. 567, 302–306. 10.1016/j.febslet.2004.04.088 [DOI] [PubMed] [Google Scholar]
- Kim S., Coulombe P. A. (2010). OPINION Emerging role for the cytoskeleton as an organizer and regulator of translation. Nat. Rev. Mol. Cell Biol. 11, 75–81. 10.1038/nrm2818 [DOI] [PubMed] [Google Scholar]
- Kirik V., Herrmann U., Parupalli C., Sedbrook J. C., Ehrhardt D. W., Hulskamp M. (2007). CLASP localizes in two discrete patterns on cortical microtubules and is required for cell morphogenesis and cell division in Arabidopsis. J. Cell Sci. 120, 4416–4425. 10.1242/jcs.024950 [DOI] [PubMed] [Google Scholar]
- Klotz J., Nick P. (2012). A novel actin-microtubule cross-linking kinesin, NtKCH, functions in cell expansion and division. New Phytol. 193, 576–589. 10.1111/j.1469-8137.2011.03944.x [DOI] [PubMed] [Google Scholar]
- Koehler A., Hurt E. (2007). Exporting RNA from the nucleus to the cytoplasm. Nat. Rev. Mol. Cell Biol. 8, 761–773. 10.1038/nrm2255 [DOI] [PubMed] [Google Scholar]
- Korolev A. V., Buschmann H., Doonan J. H., Lloyd C. W. (2007). AtMAP70-5, a divergent member of the MAP70 family of microtubule-associated proteins, is required for anisotropic cell growth in Arabidopsis. J. Cell Sci. 120, 2241–2247. 10.1242/jcs.007393 [DOI] [PubMed] [Google Scholar]
- Korolev A. V., Chan J., Naldrett M. J., Doonan J. H., Lloyd C. W. (2005). Identification of a novel family of 70 kDa microtubule-associated proteins in Arabidopsis cells. Plant J. 42, 547–555. 10.1111/j.1365-313X.2005.02393.x [DOI] [PubMed] [Google Scholar]
- Kovar D. R., Pollard T. D. (2004). Progressing actin: sFormin as a processive elongation machine. Nat. Cell Biol. 6, 1158–1159. 10.1038/ncb1204-1158 [DOI] [PubMed] [Google Scholar]
- Koyasu S., Nishida E., Kadowaki T., Matsuzaki F., Iida K., Harada F., et al. (1986). 2 Mammalian heat-shock proteins, HSP90 and HSP100, Are Actin-Binding Proteins. Proc. Natl. Acad. Sci. U.S.A. 83, 8054–8058. 10.1073/pnas.83.21.8054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krishnakumar S., Oppenheimer D. G. (1999). Extragenic suppressors of the Arabidopsis zwi-3 mutation identify new genes that function in trichome branch formation sand pollen tube growth. Development 126, 3079–3088. [DOI] [PubMed] [Google Scholar]
- Krtkova J., Zimmermann A., Schwarzerova K., Nick P. (2012). Hsp90 binds microtubules and is involved in the reorganization of the microtubular network in angiosperms. J. Plant Physiol. 169, 1329–1339. 10.1016/j.jplph.2012.06.010 [DOI] [PubMed] [Google Scholar]
- Krupnova T., Sasabe M., Ghebreghiorghis L., Gruber C. W., Hamada T., Dehmel, et al. (2009). Microtubule-associated kinase-like protein RUNKEL needed for cell plate expansion in Arabidopsis cytokinesis. Curr. Biol. 19, 518–523. 10.1016/j.cub.2009.02.021 [DOI] [PubMed] [Google Scholar]
- Kumagai H., Sakai H. (1983). A porcine brain protein (35K-protein) which bundles microtubules and its identification as glyceraldehyde-3-phosphate dehydrogenase. J. Biochem. 93, 1259–1269. [DOI] [PubMed] [Google Scholar]
- Kusner D. J., Barton J. A., Qin C. B., Wang X. M., Iyer S. S. (2003). Evolutionary conservation of physical and functional interactions between phospholipase D and actin. Arch. Biochem. Biophys. 412, 231–241. 10.1016/S0003-9861(03)00052-3 [DOI] [PubMed] [Google Scholar]
- Lange B. M. H., Bachi A., Wilm M., Gonzalez C. (2000). Hsp90 is a core centrosomal component and is required at different stages of the centrosome cycle in Drosophila and vertebrates. EMBO J. 19, 1252–1262. 10.1093/emboj/19.6.1252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee Y., Kim Y.-W., Jeon B. W., Park K.-Y., Suh S. J., Seo J., et al. (2007). Phosphatidylinositol 4,5-bisphosphate is important for stomatal opening. Plant J. 52, 803–816. 10.1111/j.1365-313X.2007.03277.x [DOI] [PubMed] [Google Scholar]
- Lee Y. R. J., Liu B. (2000). Identification of a phragmoplast-associated kinesin-related protein in higher plants. Curr. Biol. 10, 797–800. 10.1016/S0960-9822(00)00564-9 [DOI] [PubMed] [Google Scholar]
- Lewis S. A., Tian G. L., Cowan N. J. (1997). The alpha- and beta-tubulin folding pathways. Trends Cell Biol. 7, 479–484. 10.1016/S0962-8924(97)01168-9 [DOI] [PubMed] [Google Scholar]
- Li H., Zeng X., Liu Z. Q., Meng Q. T., Yuan M., Mao T. L. (2009). Arabidopsis microtubule-associated protein AtMAP65-2 acts as a microtubule stabilizer. Plant Mol. Biol. 69, 313–324. 10.1007/s11103-008-9426-1 [DOI] [PubMed] [Google Scholar]
- Li J., Wang X., Qin T., Zhang Y., Liu X., Sun J., et al. (2011). MDP25, A novel calcium regulatory protein, mediates hypocotyl cell elongation by destabilizing cortical microtubules in Arabidopsis. Plant Cell 23, 4411–4427. 10.1105/tpc.111.092684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S., Sun T., Ren H. (2015). The functions of the cytoskeleton and associated proteins during mitosis and cytokinesis in plant cells. Front. Plant Sci. 6:282. 10.3389/fpls.2015.00282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y. H., Shen Y. A., Cai C., Zhong C. C., Zhu L., Yuan M., et al. (2010). The type II Arabidopsis formin14 interacts with microtubules and microfilaments to regulate cell division. Plant Cell 22, 2710–2726. 10.1105/tpc.110.075507 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu B., Cyr R. J., Palevitz B. A. (1996). A kinesin-like protein, KatAp, in the cells of arabidopsis and other plants. Plant Cell 8, 119–132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C., Qi X., Zhao Q., Yu J. (2013). Characterization and functional analysis of the potato pollen-specific microtubule-associated protein SBgLR in tobacco. PLoS ONE 8:e60543. 10.1371/journal.pone.0060543 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu G., Tang J. Z., Edmonds B. T., Murray J., Levin S., Condeelis J. (1996). F-actin sequesters elongation factor 1 alpha from interaction with aminoacyl-tRNA in a pH-dependent reaction. J. Cell Biol. 135, 953–963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lloyd C., Hussey P. (2001). Microtubule-associated proteins in plants - why we need a map. Nat. Rev. Mol. Cell Biol. 2, 40–47. 10.1038/35048005 [DOI] [PubMed] [Google Scholar]
- Mao G. J., Chan J., Calder G., Doonan J. H., Lloyd C. W. (2005). Modulated targeting of GFP-AtMAP65-1 to central spindle microtubules during division. Plant J. 43, 469–478. 10.1111/j.1365-313X.2005.02464.x [DOI] [PubMed] [Google Scholar]
- Marc J., Sharkey D. E., Durso N. A., Zhang M., Cyr R. J. (1996). Isolation of a 90-kD microtubule-associated protein from tobacco membranes. Plant Cell 8, 2127–2138. 10.1105/tpc.8.11.2127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin S. W., Glover B. J., Davies J. M. (2005). Lipid microdomains - plant membranes get organized. Trends Plant Sci. 10, 263–265. 10.1016/j.tplants.2005.04.004 [DOI] [PubMed] [Google Scholar]
- Mathur J., Mathur N., Kernebeck B., Srinivas B. P., Hulskamp M. (2003). A novel localization pattern for an EB1-like protein links microtubule dynamics to endomembrane organization. Curr. Biol. 13, 1991–1997. 10.1016/j.cub.2003.10.033 [DOI] [PubMed] [Google Scholar]
- Matsui K., Collings D., Asada T. (2001). Identification of a novel plant-specific kinesin-like protein that is highly expressed in interphase tobacco BY-2 cells. Protoplasma 215, 105–115. 10.1007/BF01280307 [DOI] [PubMed] [Google Scholar]
- Meijer H. J. G., Munnik T. (2003). Phospholipid-based signaling in plants. Annu. Rev. Plant Biol. 54, 265–306. 10.1146/annurev.arplant.54.031902.134748 [DOI] [PubMed] [Google Scholar]
- Mineyuki Y. (1999). The preprophase band of microtubules: its function as a cytokinetic apparatus in higher plants. Int. Rev. Cytol. 187, 1–49. 10.1016/S0074-7696(08)62415-8 [DOI] [Google Scholar]
- Moore R. C., Durso N. A., Cyr R. J. (1998). Elongation factor-1 alpha stabilizes microtubules in a calcium/calmodulin-dependent manner. Cell Motil. Cytoskeleton 41, 168–180. [DOI] [PubMed] [Google Scholar]
- Moser M., Schafer E., Ehmann B. (2000). Characterization of protein and transcript levels of the chaperonin containing tailless complex protein-1 and tubulin during light-regulated growth of oat seedlings. Plant Physiol. 124, 313–320. 10.1104/pp.124.1.313 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muench D. G., Park N.-I. (2006). Messages on the move: the role of the cytoskeleton in mRNA localization and translation in plant cells. Can. J. Bot. 572–580. 10.1139/b05-167 [DOI] [Google Scholar]
- Muller S., Smertenko A., Wagner V., Heinrich M., Hussey P. J., Hauser M. T. (2004). The plant microtubule-associated protein AtMAP65-3/PLE is essential for cytokinetic phragmoplast function. Curr. Biol. 14, 412–417. 10.1016/j.cub.2004.02.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munnik T. (2001). Phosphatidic acid: an emerging plant lipid second messenger. Trends Plant Sci. 6, 227–233. 10.1016/S1360-1385(01)01918-5 [DOI] [PubMed] [Google Scholar]
- Murray J. W., Edmonds B. T., Liu G., Condeelis J. (1996). Bundling of actin filaments by elongation factor 1 alpha inhibits polymerization at filament ends. J. Cell Biol. 135, 1309–1321. 10.1083/jcb.135.5.1309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakajima K., Furutani I., Tachimoto H., Matsubara H., Hashimoto T. (2004). SPIRAL1 encodes a plant-specific microtubule-localized protein required for directional control of rapidly expanding Arabidopsis cells. Plant Cell 16, 1178–1190. 10.1105/tpc.017830 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nick P., Heuing A., Ehmann B. (2000). Plant chaperonins: a role in microtubule-dependent wall formation? Protoplasma 211, 234–244. 10.1007/bf01304491 [DOI] [Google Scholar]
- Oda Y., Iida Y., Kondo Y., Fukuda H. (2010). Wood cell-wall structure requires local 2D-microtubule disassembly by a novel plasma membrane-anchored protein. Curr. Biol. 20, 1197–1202. 10.1016/j.cub.2010.05.038 [DOI] [PubMed] [Google Scholar]
- Paredez A. R., Somerville C. R., Ehrhardt D. W. (2006). Visualization of cellulose synthase demonstrates functional association with microtubules. Science 312, 1491–1495. 10.1126/science.1126551 [DOI] [PubMed] [Google Scholar]
- Parrotta L., Cresti M., Cai G. (2013). Heat-shock protein 70 binds microtubules and interacts with kinesin in tobacco pollen tubes. Cytoskeleton 70, 522–537. 10.1002/cm.21134 [DOI] [PubMed] [Google Scholar]
- Perez F., Pernet-Gallay K., Nizak C., Goodson H. V., Kreis T. E., Goud B. (2002). CLIPR-59, a new trans-Golgi/TGN cytoplasmic linker protein belonging to the CLIP-170 family. J. Cell Biol. 156, 631–642. 10.1083/jcb.200111003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrin R. M., Wang Y., Yuen C. Y. L., Will J., Masson P. H. (2007). WVD2 is a novel microtubule-associated protein in Arabidopsis thaliana. Plant J. 49, 961–971. 10.1111/j.1365-313X.2006.03015.x [DOI] [PubMed] [Google Scholar]
- Pesquet E., Korolev A. V., Calder G., Lloyd C. W. (2010). The microtubule-associated protein AtMAP70-5 regulates secondary wall patterning in Arabidopsis wood cells. Curr. Biol. 20, 744–749. 10.1016/j.cub.2010.02.057 [DOI] [PubMed] [Google Scholar]
- Petrasek J., Freudenreich A., Heuing A., Opatrny Z., Nick P. (1998). Heat-shock protein 90 is associated with microtubules in tobacco cells. Protoplasma 202, 161–174. 10.1007/BF01282544 [DOI] [Google Scholar]
- Petrasek J., Schwarzerova K. (2009). Actin and microtubule cytoskeleton interactions. Curr. Opin. Plant Biol. 12, 728–734. 10.1016/j.pbi.2009.09.010 [DOI] [PubMed] [Google Scholar]
- Pleskot R., Pejchar P., Staiger C. J., Potocky M. (2014). When fat is not bad: the regulation of actin dynamics by phospholipid signaling molecules. Front. Plant Sci. 5:5. 10.3389/fpls.2014.00005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pratt W. B., Krishna P., Olsen L. J. (2001). Hsp90-binding immunophilins in plants: the protein movers. Trends Plant Sci. 6, 54–58. 10.1016/S1360-1385(00)01843-4 [DOI] [PubMed] [Google Scholar]
- Pratt W. B., Morishima Y., Peng H. M., Osawa Y. (2010). Proposal for a role of the Hsp90/Hsp70-based chaperone machinery in making triage decisions when proteins undergo oxidative and toxic damage. Exp. Biol. Med. 235, 278–289. 10.1258/ebm.2009.009250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pratt W. B., Silverstein A. M., Galigniana M. D. (1999). A model for the cytoplasmic trafficking of signalling proteins involving the hsp90-binding immunophilins and p50(cdc37). Cell. Signal. 11, 839–851. 10.1016/S0898-6568(99)00064-9 [DOI] [PubMed] [Google Scholar]
- Preuss M. L., Kovar D. R., Lee Y. R. J., Staiger C. J., Delmer D. P., Liu B. (2004). A plant-specific kinesin binds to actin microfilaments and interacts with cortical microtubules in cotton fibers. Plant Physiol. 136, 3945–3955. 10.1104/pp.104.052340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pruyne D., Gao L., Bi E. F., Bretscher A. (2004). Stable and dynamic axes of polarity use distinct formin isoforms in budding yeast. Mol. Biol. Cell 15, 4971–4989. 10.1091/mbc.E04-04-0296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin T., Liu X., Li J., Sun J., Song L., Mao T. (2014). Arabidopsis microtubule-destabilizing protein 25 functions in pollen tube growth by severing actin filaments. Plant Cell 26, 325–339. 10.1105/tpc.113.119768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosero A., Zarsky V., Cvrckova F. (2013). AtFH1 formin mutation affects actin filament and microtubule dynamics in Arabidopsis thaliana. J. Exp. Bot. 64, 585–597. 10.1093/jxb/ers351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruggenthaler P., Fichtenbauer D., Krasensky J., Jonak C., Waigmann E. (2009). Microtubule-associated protein AtMPB2C plays a role in organization of cortical microtubules, stomata patterning, and tobamovirus infectivity. Plant Physiol. 149, 1354–1365. 10.1104/pp.108.130450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rutherford S. L., Lindquist S. (1998). Hsp90 as a capacitor for morphological evolution. Nature 396, 336–342. 10.1038/24550 [DOI] [PubMed] [Google Scholar]
- Rutherford S. L., Zuker C. S. (1994). Protein-folding and the regulation of signaling pathways. Cell 79, 1129–1132. 10.1016/0092-8674(94)90003-5 [DOI] [PubMed] [Google Scholar]
- Sagot I., Klee S. K., Pellman D. (2002). Yeast formins regulate cell polarity by controlling the assembly of actin cables. Nat. Cell Biol. 4, 42–50. 10.1038/ncb719 [DOI] [PubMed] [Google Scholar]
- Sampathkumar A., Lindeboom J. J., Debolt S., Gutierrez R., Ehrhardt D. W., Ketelaar T., et al. (2011). Live cell imaging reveals structural associations between the actin and microtubule cytoskeleton in Arabidopsis. Plant Cell 23, 2302–2313. 10.1105/tpc.111.087940 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez E. R., Redmond T., Scherrer L. C., Bresnick E. H., Welsh M. J., Pratt W. B. (1988). Evidence that the 90-kilodalton heat-shock protein is associated with tubulin-containing complexes in L-cell cytosol and in intact Ptk cells. Mol. Endocrinol. 2, 756–760. 10.1210/mend-2-8-756 [DOI] [PubMed] [Google Scholar]
- Sano T., Higaki T., Oda Y., Hayashi T., Hasezawa S. (2005). Appearance of actin microfilament ‘twin peaks’ in mitosis and their function in cell plate formation, as visualized in tobacco BY-2 cells expressing GFP-fimbrin. Plant J. 44, 595–605. 10.1111/j.1365-313X.2005.02558.x [DOI] [PubMed] [Google Scholar]
- Schneider R., Persson S. (2015). Connecting two arrays: the emerging role of actin-microtubule cross-linking motor proteins. Front. Plant Sci. 6:415. 10.3389/fpls.2015.00415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sedbrook J. C. (2004). MAPs in plant cells: delineating microtubule growth dynamics and organization. Curr. Opin. Plant Biol. 7, 632–640. 10.1016/j.pbi.2004.09.017 [DOI] [PubMed] [Google Scholar]
- Sedbrook J. C., Kaloriti D. (2008). Microtubules, MAPs and plant directional cell expansion. Trends Plant Sci. 13, 303–310. 10.1016/j.tplants.2008.04.002 [DOI] [PubMed] [Google Scholar]
- Shiina N., Gotoh Y., Kubomura N., Iwamatsu A., Nishida E. (1994). Microtubule severing by elongation-factor 1-alpha. Science 266, 282–285. 10.1126/science.7939665 [DOI] [PubMed] [Google Scholar]
- Silflow C. D., Sun X., Haas N. A., Foley J. W., Lefebvre P. A. (2011). The Hsp70 and Hsp40 chaperones influence microtubule stability in Chlamydomonas. Genetics 189, 1249–1260. 10.1534/genetics.111.133587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sirover M. A. (1999). New insights into an old protein: the functional diversity of mammalian glyceraldehyde-3-phosphate dehydrogenase. Biochim. Biophys. Acta 1432, 159–184. 10.1016/S0167-4838(99)00119-3 [DOI] [PubMed] [Google Scholar]
- Smertenko A. P., Chang H. Y., Wagner V., Kaloriti D., Fenyk S., Sonobe S., et al. (2004). The Arabidopsis microtubule-associated protein AtMAP65-1: molecular analysis of its microtubule bundling activity. Plant Cell 16, 2035–2047. 10.1105/tpc.104.023937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smertenko A. P., Deeks M. J., Hussey P. J. (2010). Strategies of actin reorganisation in plant cells. J. Cell Sci. 123, 3029–3029. 10.1242/jcs.079749 [DOI] [PubMed] [Google Scholar]
- Smertenko A. P., Kaloriti D., Chang H. Y., Fiserova J., Opatrny Z., Hussey P. J. (2008). The C-terminal variable region specifies the dynamic properties of arabidopsis microtubule-associated protein MAP65 isotypes. Plant Cell 20, 3346–3358. 10.1105/tpc.108.063362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith L. G., Gerttula S. M., Han S. C., Levy J. (2001). TANGLED1: a microtubule binding protein required for the spatial control of cytokinesis in maize. J. Cell Biol. 152, 231–236. 10.1083/jcb.152.1.231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somers M., Engelborghs Y., Baert J. (1990). Analysis of the binding of glyceraldehyde-3-phosphate dehydrogenase to microtubules, the mechanism of bundle formation and the linkage effect. Eur. J. Biochem. 193, 437–444. 10.1111/j.1432-1033.1990.tb19357.x [DOI] [PubMed] [Google Scholar]
- Suh B.-C., Hille B. (2008). PIP(2) is a necessary cofactor for ion channel function: how and why? Annu. Rev. Biophys. 37, 175–195. 10.1146/annurev.biophys.37.032807.125859 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takabatake R., Ando Y., Seo S., Katou S., Tsuda S., Ohashi Y., et al. (2007). MAP kinases function downstream of HSP90 and upstream of mitochondria in TMV resistance grene N-mediated hypersensitive cell death. Plant Cell Physiol. 48, 498–510. 10.1093/pcp/pcm021 [DOI] [PubMed] [Google Scholar]
- Takahashi H., Hirota K., Kawahara A., Hayakawa E., Inoue Y. (2003). Randomization of cortical microtubules in root epidermal cells induces root hair initiation in lettuce (Lactuca sativa L.) seedlings. Plant Cell Physiol. 44, 350–359. 10.1093/pcp/pcg043 [DOI] [PubMed] [Google Scholar]
- Tamura K., Nakatani K., Mitsui H., Ohashi Y., Takahashi H. (1999). Characterization of katD, a kinesin-like protein gene specifically expressed in floral tissues of Arabidopsis thaliana. Gene 230, 23–32. 10.1016/S0378-1119(99)00070-0 [DOI] [PubMed] [Google Scholar]
- Tapken W., Murphy A. S. (2015). Membrane nanodomains in plants: capturing form, function, and movement. J. Exp. Bot. 66, 1573–1586. 10.1093/jxb/erv054 [DOI] [PubMed] [Google Scholar]
- Terada S., Kinjo M., Aihara M., Takei Y., Hirokawa N. (2010). Kinesin-1/Hsc70-dependent mechanism of slow axonal transport and its relation to fast axonal transport. EMBO J. 29, 843–854. 10.1038/emboj.2009.389 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theologis A., Ecker J. R., Palm C. J., Federspiel N. A., Kaul S., White O., et al. (2000). Sequence and analysis of chromosome 1 of the plant Arabidopsis thaliana. Nature 408, 816–820. 10.1038/35048500 [DOI] [PubMed] [Google Scholar]
- Thomas C., Tholl S., Moes D., Dieterle M., Papuga J., Moreau F., et al. (2009). Actin bundling in plants. Cell Motil. Cytoskeleton 66, 940–957. 10.1002/cm.20389 [DOI] [PubMed] [Google Scholar]
- Tisdale E. J. (2002). Glyceraldehyde-3-phosphate dehydrogenase is phosphorylated by protein kinase c mu lambda and plays a role in microtubule dynamics in the early secretory pathway. J. Biol. Chem. 277, 3334–3341. 10.1074/jbc.M109744200 [DOI] [PubMed] [Google Scholar]
- Tisdale E. J., Azizi F., Artalejo C. R. (2009). Rab2 utilizes glyceraldehyde-3-phosphate dehydrogenase and protein kinase C iota to associate with microtubules and to recruit dynein. J. Biol. Chem. 284, 5876–5884. 10.1074/jbc.M807756200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traas J. A., Doonan J. H., Rawlins D. J., Shaw P. J., Watts J., Lloyd C. W. (1987). An actin network is present in the cytoplasm throughout the cell-cycle of carrot cells and associates with the dividing nucleus. J. Cell Biol. 105, 387–395. 10.1083/jcb.105.1.387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umezu N., Umeki N., Mitsui T., Kondo K., Maruta S. (2011). Characterization of a novel rice kinesin O12 with a calponin homology domain. J. Biochem. 149, 91–101. 10.1093/jb/mvq122 [DOI] [PubMed] [Google Scholar]
- Van Damme D., Van Poucke K., Boutant E., Ritzenthaler C., Inze D., Geelen D. (2004). In vivo dynamics and differential microtubule-binding activities of MAP65 proteins. Plant Physiol. 136, 3956–3967. 10.1104/pp.104.051623 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volker K. W., Knull H. R. (1997). A glycolytic enzyme binding domain on tubulin. Arch. Biochem. Biophys. 338, 237–243. 10.1006/abbi.1996.9819 [DOI] [PubMed] [Google Scholar]
- Walsh J. L., Keith T. J., Knull H. R. (1989). Glycolytic enzyme interactions with tubulin and microtubules. Biochim. Biophys. Acta 999, 64–70. 10.1016/0167-4838(89)90031-9 [DOI] [PubMed] [Google Scholar]
- Wang C. X., Wang X. M. (2001). A novel phospholipase D of arabidopsis that is activated by oleic acid and associated with the plasma membrane. Plant Physiol. 127, 1102–1112. 10.1104/pp.010444 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X. M. (2002). Phospholipase D in hormonal and stress signaling. Curr. Opin. Plant Biol. 5, 408–414. 10.1016/S1369-5266(02)00283-2 [DOI] [PubMed] [Google Scholar]
- Wang X., Vinocur B., Shoseyov O., Altman A. (2004). Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 9, 244–252. 10.1016/j.tplants.2004.03.006 [DOI] [PubMed] [Google Scholar]
- Wang X., Zhang J., Yuan M., Ehrhardt D. W., Wang Z., Mao T. (2012). Arabidopsis microtubule destabilizing protein40 is involved in brassinosteroid regulation of hypocotyl elongation. Plant Cell 24, 5193–5193. 10.1105/tpc.112.103838 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X., Zhu L., Liu B. Q., Wang C., Jin L. F., Zhao Q., et al. (2007). Arabidopsis Microtubule-associated protein18 functions in directional cell growth by destabilizing cortical microtubules. Plant Cell 19, 877–889. 10.1105/tpc.106.048579 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wegele H., Muller L., Buchner J. (2004). Hsp70 and Hsp90 - a relay team for protein folding. Rev. Physiol. Biochem. Pharmacol. 151, 1–44. 10.1007/s10254-003-0021-1 [DOI] [PubMed] [Google Scholar]
- Wei L. Q., Liu B., Li Y. (2005). Distribution of a kinesin-related protein on Golgi apparatus of tobacco pollen tubes. Chinese Sci. Bull. 50, 2175–2181. 10.1007/bf03182668 [DOI] [Google Scholar]
- Weis F., Moullintraffort L., Heichette C., Chretien D., Garnier C. (2010). The 90-kDa heat shock protein Hsp90 protects tubulin against thermal denaturation. J. Biol. Chem. 285, 9525–9534. 10.1074/jbc.M109.096586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welch M. D., Depace A. H., Verma S., Iwamatsu A., Mitchison T. J. (1997). The human Arp2/3 complex is composed of evolutionarily conserved subunits and is localized to cellular regions of dynamic actin filament assembly. J. Cell Biol. 138, 375–384. 10.1083/jcb.138.2.375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whittington A. T., Vugrek O., Wei K. J., Hasenbein N. G., Sugimoto K., Rashbrooke M. C., et al. (2001). MOR1 is essential for organizing cortical microtubules in plants. Nature 411, 610–613. 10.1038/35079128 [DOI] [PubMed] [Google Scholar]
- Williams N. E., Nelsen E. M. (1997). HSP70 and HSP90 homologs are associated with tubulin in hetero-oligomeric complexes, cilia and the cortex of Tetrahymena. J. Cell Sci. 110, 1665–1672. [DOI] [PubMed] [Google Scholar]
- Wu S.-Z., Bezanilla M. (2014). Myosin VIII associates with microtubule ends and together with actin plays a role in guiding plant cell division. Elife 3:e03498. 10.7554/eLife.03498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu T., Qu Z., Yang X., Qin X., Xiong J., Wang Y., et al. (2009). A cotton kinesin GhKCH2 interacts with both microtubules and microfilaments. Biochem. J. 421, 171–180. 10.1042/BJ20082020 [DOI] [PubMed] [Google Scholar]
- Yanagisawa M., Zhang C., Szymanski D. B. (2013). ARP2/3-dependent growth in the plant kingdom: SCARs for life. Front. Plant Sci. 4:166. 10.3389/fpls.2013.00166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuhara H., Muraoka M., Shogaki H., Mori H., Sonobe S. (2002). TMBP200, a microtubule bundling polypeptide isolated from telophase tobacco BY-2 cells is a MOR1 homologue. Plant Cell Physiol. 43, 595–603. 10.1093/pcp/pcf074 [DOI] [PubMed] [Google Scholar]
- Zeng C. J. T., Lee Y. R. J., Liu B. (2009). The WD40 repeat protein NEDD1 functions in microtubule organization during cell division in Arabidopsis thaliana. Plant Cell 21, 1129–1140. 10.1105/tpc.109.065953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X. P., Glaser E. (2002). Interaction of plant mitochondrial and chloroplast signal peptides with the Hsp70 molecular chaperone. Trends Plant Sci. 7, 14–21. 10.1016/S1360-1385(01)02180-X [DOI] [PubMed] [Google Scholar]