

Abstract
1,25-Dihydroxvitamin D3 [1,25(OH)2D3] is the hormonally active form of vitamin D. The genomic mechanism of 1,25(OH)2D3 action involves the direct binding of the 1,25(OH)2D3 activated vitamin D receptor/retinoic X receptor (VDR/RXR) heterodimeric complex to specific DNA sequences. Numerous VDR co-regulatory proteins have been identified, and genome-wide studies have shown that the actions of 1,25(OH)2D3 involve regulation of gene activity at a range of locations many kilobases from the transcription start site. The structure of the liganded VDR/RXR complex was recently characterized using cryoelectron microscopy, X-ray scattering, and hydrogen deuterium exchange. These recent technological advances will result in a more complete understanding of VDR coactivator interactions, thus facilitating cell and gene specific clinical applications. Although the identification of mechanisms mediating VDR-regulated transcription has been one focus of recent research in the field, other topics of fundamental importance include the identification and functional significance of proteins involved in the metabolism of vitamin D. CYP2R1 has been identified as the most important 25-hydroxylase, and a critical role for CYP24A1 in humans was noted in studies showing that inactivating mutations in CYP24A1 are a probable cause of idiopathic infantile hypercalcemia. In addition, studies using knockout and transgenic mice have provided new insight on the physiological role of vitamin D in classical target tissues as well as evidence of extraskeletal effects of 1,25(OH)2D3 including inhibition of cancer progression, effects on the cardiovascular system, and immunomodulatory effects in certain autoimmune diseases. Some of the mechanistic findings in mouse models have also been observed in humans. The identification of similar pathways in humans could lead to the development of new therapies to prevent and treat disease.
I. INTRODUCTION
In recent years, vitamin D has received increased attention due to the resurgence of vitamin D deficiency and rickets as a global health issue together with compelling evidence in the laboratory indicating that 1,25-dihydroxyvitamin D3 [1,25(OH)2D3], the hormonally active form of vitamin D, generates a number of extraskeletal biological responses including inhibition of breast, colon, and prostate cancer cell progression; effects on the cardiovascular system; and protection against a number of autoimmune diseases including multiple sclerosis and inflammatory bowel disease (195). This review summarizes our current understanding of vitamin D and its bioactivation and discusses new developments that have changed our understanding of the mechanism of vitamin D action in classical as well as nonclassical target tissues. This article also evaluates the suggested role of vitamin D in extraskeletal health, provides an overview of 1,25(OH)2D3 analogs that have been developed, and indicates questions that remain and need to be addressed.
II. VITAMIN D AND ITS BIOACTIVATION
A. Vitamin D and 25(OH)D3
Vitamin D3 (cholecalciferol), the natural form of vitamin D, is produced in the skin from 7-dehydrocholesterol. Upon irradiation, 7-dehydrocholesterol produces pre-vitamin D3 which undergoes a temperature-sensitive rearrangement of three double bonds to form vitamin D3. The synthesis of vitamin D in the skin is the most important source of vitamin D and depends on the intensity of the ultraviolet irradiation which is dependent on season and latitude. For example, in Boston (42.2°N), no vitamin D is produced from sun-exposed skin from November to February, while in San Juan (18°N) the skin produces vitamin D all year. Melanin and sunscreen markedly diminish the production of vitamin D (194, 483). Vitamin D can also be taken in the diet. However, vitamin D is present in only a few foods (which include fortified dairy products and fish oils). Vitamin D3 itself is not biologically active. Vitamin D is transported in the blood by vitamin D binding protein (DBP; which binds vitamin D and its metabolites in serum) to the liver. In the liver, vitamin D is hydroxylated at C-25 to produce 25-hydroxyvitamin D3 [25(OH)D3]. 25(OH)D3 is the major circulating form of vitamin D. Its concentration in the serum has served as one of the most reliable biomarkers of vitamin D status (42, 184, 195). The synthesis of 25(OH)D3 has not been reported to be highly regulated (108). Many cytochrome P-450 enzymes (CYPs) including CYP2R1, CYP27A1, and CYP2D25 have been considered as candidates for the enzyme responsible for the conversion of vitamin D to 25(OH)D3 (523). It has been suggested that CYP2R1, first identified as a microsomal vitamin D 25-hydroxylase by Cheng et al. (80), is the key vitamin D 25-hydroxylase, since patients with a mutation of the CYP2R1 have 25(OH)D3 deficiency and symptoms of vitamin D-dependent rickets (12, 73, 79, 121, 275). The crystal structure of CYP2R1 in complex with vitamin D3 has been reported showing that at the active site the 17β-aliphatic side chain of vitamin D is located above the heme plate appropriate for 25-hydroxylation (430). Further strengthening the evidence for the physiological role of CYP2R1 are recent studies using Cyp2r1 null mutant mice which demonstrate that CYP2R1 is the major enzyme responsible for 25-hydroxylation of vitamin D (524). In the Cyp2r1 null mice, although 25(OH)D3 levels are dramatically reduced, synthesis of 25(OH)D3 is not abolished, suggesting the presence of other vitamin D 25-hydroxylases yet to be identified (524).
25(OH)D3 is transported by DBP to the kidney and is filtered by the glomerulus. In the kidney megalin, a 600-kDa transmembrane protein, and a member of the low-density lipopoprotein receptor superfamily, acts as a cell surface receptor for DBP resulting in uptake of 25(OH)D in the tubular epithelial cells by endocytic internalization (89). The significance of megalin renal uptake and metabolism of 25(OH)D3 is demonstrated in studies using Megalin knockout mice. Mice lacking Megalin lose DBP and 25(OH)D3 in the urine and have defects of bone metabolism that resemble vitamin D-deficient rickets (336). Cubulin, a second surface receptor for DBP in the proximal tubule, also associates with megalin to internalize complexes of DBP and 25(OH)D3 (337). In addition, disabled 2 (Dab) 2, a cytoplasmic adaptor protein, also works in conjunction with megalin for the cellular uptake of DBP/25(OH)D3 by binding to the cytoplasmic tail of megalin enabling the proper routing of the receptor (320, 490).
B. CYP27B1
In the proximal renal tubule, 25(OH)D3 is hydroxylated at the position of carbon 1 of the A ring, resulting in the formation of 1,25(OH)2D3, the functional, hormonally active form of vitamin D which is responsible for most, if not all, of the biologic actions of vitamin D. The renal 25(OH)D 1α hydroxylase (mitochondrial CYP27B1), which metabolizes 25(OH)D3 to 1,25(OH)2D3, comprises a cytochrome P-450, a ferredoxin, and a ferredoxin reductase and is present predominantly in the kidney (proximal straight tubules) and contributes to the circulating concentrations of 1,25(OH)2D3 (215, 216). Mutations resulting in inactive or deleted CYP27B1 cause vitamin D dependency rickets type 1 (VDDRI) (also known as pseudovitamin D deficiency rickets) despite normal intake of vitamin D, indicating the importance of the CYP27B1 enzyme (234). CYP27B1 has been cloned from rat, mouse, and human (410, 425, 436). The human and rodent CYP27B1 genes comprise nine exons, extending over 5 kbp (234, 318). Cyp27b1 null mice have provided a mouse model of VDDR type 1. These mice have rickets, undetectable levels of 1,25(OH)2D3, low serum calcium, and secondary hyperparathyroidism (SHPT) (107, 351). It has been suggested that in healthy animals and humans CYP27B1 is only expressed in kidney and, during pregnancy, in placenta (108). In addition to the kidney, it has been reported that CYP27B1 is present in a number of extrarenal sites. Extrarenal production of CYP27B1 has been convincingly demonstrated in patients with sarcoidosis (4, 31). Macrophages were identified as the source of extrarenal production of 1,25(OH)2D3 resulting in hypercalcemia and hypercalciuria in these patients. In addition to sarcoidosis, hypercalcemia has also been identified in patients with Crohn's disease (51). It was suggested that activated macrophages of Crohn's granuloma are responsible for the hypercalcemia in Crohn's disease. CYP27B1 produced by macrophages, unlike renal CYP27B1, is not suppressed by elevated 1,25(OH)2D3 but is upregulated by immune stimuli [interferon-γ and lipopolysaccharide (LPS)]. Regulation by immune stimuli has been reported to involve multiple pathways (including JAK/STAT and NFκB) and to require binding of the C/EBPβ transcription factor to the mouse and human CYP27B1 genes (136, 429). Further evidence of immune derived CYP27B1 is provided by recent studies showing that reconstitution of the hematopoietic cell population from wild-type (WT) mice in Cyp27b1 knockout mice protects mice from colitis (342). Murine T-cell production of CYP27B1 was demonstrated in the CD8+ but not the CD4+ T cell population (342). Thus it is likely that murine CD8+ T cells as well as other immune cells under activation conditions can produce 1,25(OH)2D3 to resolve the immune response following antigen specific activation. Cancer cells have also been shown to express CYP27B1 (see sect. VA). In addition, CYP27B1 expression has been noted in parathyroid gland and in a number of other tissues [see Bikle (42a) for review]. However, whether there is a functional impact of CYP27B1 activity in vivo at sites other than the kidney and placenta under normal physiological conditions remains to be determined.
C. DBP
Although DBP functions as a binding protein for all vitamin D metabolites in the serum [20 times less affinity for 1,25(OH)2D3 than for 25(OH)D3], DBP also sequesters actin, can bind fatty acids, and can function as a chemotactic factor with a significant role in neutrophil recruitment (87, 453). In Dbp null mice, neutrophil recruitment has been reported to be impaired. It should be noted that although there is a marked decrease in the serum levels of 25(OH)D3 and 1,25(OH)2D3 in Dbp null mice (as expected), serum calcium, phosphorus, and PTH are equivalent in Dbp null and WT mice (390). However, Dbp null mice were more susceptible to develop osteomalacia when given a vitamin D-deficient diet, indicating that DBP may help to maintain stable stores of vitamin D. More recent studies in Dbp null mice have shown that these mice are capable of generating tissue levels of 1,25(OH)2D3 comparable to those of WT mice and that induction of vitamin D target genes is similar in WT and Dbp null mice (513). Thus the normal serum calcium levels in Dbp null mice may be caused by the ability of the vitamin D receptor (VDR) to concentrate 1,25(OH)2D3 in tissues due to its high affinity for 1,25(OH)2D3 resulting in the activation of genes involved in the maintenance of calcium homeostasis. Studies in humans have shown that DBP is highly polymorphic, with three commonly recognized variants (GC1F, GC1S, GC2) that are shown to affect protein function. The three common variants with relevance to vitamin D metabolism are determined by two SNPs in Gc; rs7041 (aspartic acid switch to glutamic acid at position 432; Gc1f vs. Gc1s) and rs4588 (threonine switch to lysine at position 436; Gc1f vs. Gc2). The resulting variations in DBP amino acid sequence appear to alter the binding affinity of DBP for vitamin D ligands, with Gc1F having the highest affinity for vitamin D metabolites and Gc2 the lowest (23, 54). Genome-wide association studies have shown that the polymorphism rs7041 and rs4588 are associated with circulating 25(OH)D3 levels (6, 302, 475); TT carriers for rs7041 (Gc1S) and AA carriers for rs4588 (Gc2) are associated with lower 25(OH)D3 levels. The prevalence of these polymorphisms differs between racial groups (97, 132, 219). Black and Asian populations are far more likely to carry the Gc1f form of DBP, while whites more frequently exhibit the Gc1s form of DBP. The Gc2 form is more frequent in people of Asian and European ancestry and rare in the black ethnic groups. An in vitro study showed that addition of DBP of higher affinity genotype (Gc1f/1f) reduced the effect of 25(OH)D3 on gene expression in monocytes, compared with lower affinity DBP polymorphic forms (Gc1s or Gc2), indicating that these DBP polymorphisms may influence 25(OH)D3 bioavailability (89). More research is needed to fully appreciate the meaning of bioavailable versus total 25(OH)D3 as well as 1,25(OH)2D3. It should be noted, however, when determining different vitamin D requirements based on circulating concentrations of DBP, that polyclonal DBP antibodies and not monoclonal antibodies (which discriminate between Gc1f and Gcls and thus could result in an underestimation of DBP concentration) should be used (52, 196).
D. CYP24A1
In the kidney, besides conversion to 1,25(OH)2D3 by CYP27B1, 25(OH)D3 can also be converted to 24,25(OH)2D3 by hydroxylation at C-24 by CYP24A1, a mitochondrial inner membrane cytochrome P-450 enzyme (214). This enzyme can hydroxylate not only 25(OH)D3 but also 1,25(OH)2D3 (Figure 1). 1,25(OH)2D3 has been suggested to be the preferred substrate for CYP24A1 (409). CYP24A1 limits the amount of 1,25(OH)2D3 when circulating 1,25(OH)2D3 is elevated by catalyzing the conversion of 1,25(OH)2D3 into 24-hydroxylated products targeted for excretion or by producing 24,25(OH)2D3 thus decreasing the pool of 25(OH)D3 available for 1-hydroxylation. CYP24A1 can also catalyze the C23 oxidation pathway resulting in the formation of 1,25(OH)2D3-26, 23 lactone from the substrate 1,25(OH)2D3 (FIGURE 1) and the formation of 25(OH)D3-26,23 lactone from 25(OH)D3 (214). CYP24A1 is present in all cells containing the VDR. Thus, in addition to regulating circulating concentrations of 1,25(OH)2D3, CYP24A1 may also modulate the levels of 1,25(OH)2D3 within the cell, resulting in an appropriate cellular response. The rat (r) Cyp24a1 gene spans ∼15 kb, is comprised of 12 exons, and is present as a single copy (340). In 2010, the crystal structure of CYP24A1 was reported (21). The crystal structure of CYP24A1 reveals that CYP24A1 has 12 α helices (A-L), and four β-sheet systems (β1-β4), as well as additional helices (A′, B′, G′ on the distal surface and K′ and K″ between β2 and the conserved heme binding motif). The CYP24A1 structure clarifies for the first time the membrane insertion elements and provides new insight on the organization of the CYP24A1 active site. These findings will be important for the design of vitamin D analogs and specific CYP24A1 inhibitors. Studies in Cyp24a1 null mice provided the first direct in vivo evidence for the role of CYP24A1 in 1,25(OH)2D3 catabolism. About 50% of homozygous mutant mice died before 3 wk of age. Cyp24a1 null mice that survive post weaning are unable to clear exogenous 1,25(OH)2D3. These animals exhibit an intramembranous bone lesion that is resolved when a double Cyp24a1/Vdr null mouse is generated, indicating that elevated 1,25(OH)2D3, acting through VDR, is responsible for the bone defect (424). Although a functional role for 24 hydroxylated metabolites in bone fracture healing has been suggested (423), a role for 24 hydroxylation other than for elimination of the vitamin D hormone has been a matter of debate (57, 188, 209, 352). Evidence for a critical role of CYP24A1 in humans was noted in recent studies which demonstrated that inactivating mutations in CYP24A1 were a probable cause of idiopathic infantile hypercalcemia (396), thus reinforcing the findings in the Cyp24a1 null mouse studies and indicating the need for careful administration of vitamin D in infants. Sequence analysis of CYP24A1 in these patients yielded five different mutations [E143del (in frame deletion of E143), E322K, R396W, L409S, and R159Q]. All mutations affect residues of structural importance. In studies in which human CYP24A1 constructs containing the mutants were transfected in cells and compared for catabolism of 1,25(OH)2D3, ablation of CYP24A1 catabolism was noted for all mutations except one mutation, L409S (distal to the active site), which retained a small level of activity. In subsequent reports, CYP24A1 mutations were identified not only in children but also in adult patients (118, 442). These patients were characterized by hypercalcemia, hypercalciuria, and recurrent nephrolithiasis. The findings in adults suggest that CYP24A1 mutations should be considered in diagnosis of long-standing hypercalcemia and hypercalciuria associated with kidney stones, particularly in patients taking vitamin D supplements.
Figure 1.

The metabolic pathway for vitamin D. CYP2R1 has been identified as a key vitamin D 25-hydroxylase. PTH, FGF23/klotho, and 1,25(OH)2D3 play key roles in the regulation of optimal levels of 1,25(OH)2D3. Only products of 1,25(OH)2D3 are represented for the C23 lactone pathway. See text for 25(OH)D products of the C23 lactone pathway.
E. Regulation of Renal CYP27B1 and CYP24A1
CYP27B1 and CYP24A1 are under stringent control. A primary signal mediating the induction of 1,25(OH)2D3 synthesis in the kidney is elevated PTH resulting from hypocalcemia (44, 187, 331, 368). The nuclear orphan receptor 4A2 (NR4A2) [also known as NURR1 (nuclear receptor related 1 protein)] which is induced in the kidney in response to PTH has been shown to be one factor mediating PTH induction of CYP27B1 transcription (525). 1,25(OH)2D3 in turn suppresses PTH production in the parathyroid gland directly at the level of transcription of the PTH gene (111, 239, 279, 387) and indirectly by increasing serum calcium levels and by upregulating the expression and transcription of the calcium sensing receptor (67). 1,25(OH)2D3 regulates its own production by inhibiting CYP27B1 (59) (Figure 1). Although negative vitamin D response elements have been identified in the PTH gene, further studies are needed to determine genome-wide mechanisms involved in 1,25(OH)2D3-mediated suppression of both PTH and CYP27B1. When compared with the regulation of CYP27B1, CYP24A1 is reciprocally regulated [stimulated by 1,25(OH)2D3 and inhibited by low calcium and PTH] (44, 187, 368). In addition to calcium, PTH, and 1,25(OH)2D3, the phosphaturic factor fibroblast growth factor 23 (FGF23), which promotes renal phosphate excretion by decreasing reabsorption in the proximal tubule, is also an important physiological regulator of vitamin D metabolism. FGF23, which belongs to the FGF19 subfamily, is an ∼32 kDa protein that is expressed predominantly in osteocytes and osteoblasts and, unlike other FGFs that act in an autocrine/paracrine fashion, acts as an endocrine factor (199, 372). 1,25(OH)2D3 and elevations in serum phosphate independently stimulate the production of FGF23 (269). αKlotho, a 130 kDa transmembrane protein that is highly expressed in the distal tubule of the kidney, acts as an obligate coreceptor for FGF23. αKlotho forms complexes with FGFR1c, FGFR3c, and FGFR4. Klotho is required for FGF23 to activate FGFRs. Together FGF23 and αklotho suppress the expression of CYP27B1 and induce CYP24A1, thereby inhibiting the synthesis and promoting the catabolism of 1,25(OH)2D3 (199) (Figure 1). Fgf23 or αklotho deficiency exhibit similar phenotypes including hyperphosphatemia, increased synthesis of 1,25(OH)2D3, ectopic calcification, and premature aging (including atherosclerosis, skin atrophy, and osteoporosis), indicating the cooperation of αklotho and FGF23 in a common signaling pathway (245, 407). Elevated FGF23 is a causative factor of tumor-induced osteomalacia and several hereditary hypophosphatemic disorders including X-linked hypophosphatemic rickets (XLH) and autosomal dominant hypophosphatemic rickets (ADHR) (1, 408, 484, 487). FGF23 is increased in chronic kidney disease (CKD). It has been suggested that increased FGF23, not 1,25(OH)2D3 insufficiency due to loss of functional renal mass, may be the initial event and thus may be an early biomarker for CKD (372). FGF23 would lead to the suppression of 1,25(OH)2D3 and thus to increased PTH observed in CKD.
Calcitonin has also been reported to regulate CYP27B1 in mammalian kidney. In addition to its known role to reduce blood calcium by shrinking osteoclasts under high calcium conditions, calcitonin has been reported to stimulate renal CYP27B1 under normocalcemic conditions (156, 225, 411). Since calcitonin levels as well as 1,25(OH)2D3 levels are elevated during lactation, early studies suggested that calcitonin may have a role to stimulate CYP27B1 resulting in increased plasma 1,25(OH)2D3 and increased intestinal calcium absorption during lactation when the need for calcium is increased (427). Studies have shown an effect of calcitonin on CYP27B1 transcription (521). In addition to calcitonin, prolactin, which is also elevated during lactation, has been reported to stimulate renal CYP27B1(382). A direct effect of prolactin in cooperation with signal transducer and activator of transcription 5 (STAT5) on renal CYP27B1 transcription has been observed (8). These findings suggest that prolactin and calcitonin can act as modulators of vitamin D-regulated calcium homeostasis during lactation when there is an increased calcium requirement for the neonate.
F. Aging and Renal Vitamin D Hydroxylases
In aging, as indicated in studies in both animals and humans, there is a decline in the ability of the kidney to synthesize 1,25(OH)2D3 (22, 454). We and others have noted that rat renal CYP24A1 increases with age (22, 295). Thus these findings suggest that the combined effect of a decline in the capacity of the kidney to convert 25(OH)D3 to 1,25(OH)2D3 and an increase in the catabolism of 1,25(OH)2D3 by CYP24A1 (and therefore a decline in intestinal calcium absorption) contribute to age-related bone loss.
G. Regulation of Placental CYP27B1
Besides kidney, placenta is also a major site for conversion of 25(OH)D3 to 1,25(OH)2D3. In the placenta CYP27B1 is expressed in both fetal trophoblasts and maternal decidua (508). Although the placenta was identified as a major site of extrarenal production of CYP27B1 over 30 years ago, the function of 1,25(OH)2D3 in the placenta was unknown. Placental expression of CYP27B1 mRNA begins early in gestation and has been reported to be highest in the first trimester (508). Recent studies have suggested that synthesis of 1,25(OH)2D3 in the placenta may play an important role in controlling placental responses to infection. Human decidual cells treated with 1,25(OH)2D3 or 25(OH)D3 show decreased synthesis of cytokines including tumor necrosis factor, granulocyte-macrophage colony stimulating factor, and interleukin-6 (137). Expression of cathelicidin, an antimicrobial peptide, is also enhanced in response to 1,25(OH)2D3 in trophoblasts and decidual cells, further indicating the importance of 1,25(OH)2D3 as a regulator of immune responses in the placenta (266). When the TLR4 ligand LPS was given in vivo, Cyp27b1 mRNA was induced in mouse placenta, indicating that placental synthesis of 1,25(OH)2D3 is also sensitive in vivo to immune challenge (267). It has been reported that CYP24A1 suppression in placenta due to excessive methylation may contribute to increased bioavailability of 1,25(OH)2D3 in human placenta (334). Together these findings suggest the importance of placental CYP27B1 during early feto-placental life as an autocrine/paracrine regulator of both acquired and innate immune responses.
III. THE VITAMIN D RECEPTOR AND GENOMIC MECHANISM OF 1,25(OH)2D3 ACTION
A. Vitamin D Receptor
1. The VDR gene and structural characterization of VDR
The biological actions of 1,25(OH)2D3 are mediated by the VDR. VDR belongs to the steroid receptor family which includes receptors for retinoic acid, thyroid hormone, sex hormones, and adrenal steroids (287). The VDR gene is evolutionarily conserved among fish, birds, and mammals (183). The human and mouse VDR genes are localized on chromosomes 12 and 15, respectively (510). Both the human and mouse genes are comprised of eight coding exons (510). Two noncoding exons are found in the mouse gene, and at least six noncoding exons are in the human gene. In the human gene there are also at least two promoters. Tissue specific promoter usage has been suggested (100, 511). VDR protein [containing 423 amino acid (mouse VDR) or 427 amino acids (human VDR)] functions as an obligate heterodimer with RXR for activation of vitamin D target genes (183, 364, 510). The two core functional domains of the VDR are the highly conserved NH2-terminal DNA binding domain (DBD) and the more variable COOH-terminal ligand binding domain (LBD). The DBD is a cysteine-rich zinc finger region. There are two zinc fingers, each of which contains a single zinc atom in a tetrahedral arrangement with four invariant cysteine residues (108, 281, 364). The LBD is comprised of at least 12 α helices [H1-H12; the ligand-dependent activation function (AF2) corresponds to H12] and 3 β sheets (S1-3) (384). 1,25(OH)2D3 binding induces a conformational change that facilitates interaction with RXR and coregulatory complexes required for the transcription of target genes. Although other coactivator interfaces in the LBD of VDR have been identified, repositioning of H12 after 1,25(OH)2D3 binding has been reported to be critical for recruitment of coactivator proteins. The DBD and the LBD are connected through a hinge region. Although the crystal structures of the isolated VDR LBD and the VDR DBD have been reported (384, 403, 404), X-ray crystallographic data of the VDR/RXR complex is currently not available. Recently, the structure of the liganded VDR/RXR DNA complex was characterized using cryoelectron microscopy (345) (Figure 2). Findings from this study suggest cooperative and allosteric effects between the LBD and the DBD in VDR-mediated regulation of gene expression. In addition, the structure reveals that the hinge region may stabilize the whole complex, thus facilitating the positioning of the LBD to make the area of H12 accessible for recruitment of coregulators (345). Recent studies using small angle X-ray scattering and hydrogen-deuterium exchange technology also enabled characterization of the VDR/RXR DNA complex and similarly indicated cooperative effects between the VDR DBD and VDR LBD, suggesting mechanisms by which ligands and DNA can act together to fine-tune regulation of gene expression (383, 514). These recent technological advances will allow the visualization of the VDR complexes that have been difficult to crystallize and will result in a more complete understanding of the structural basis for VDR and VDR coactivator action.
Figure 2.
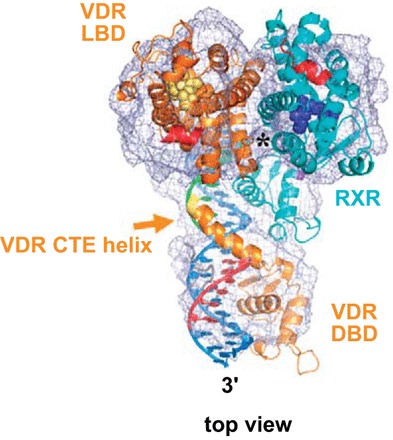
Structure of the full human RXR/VDR nuclear receptor heterodimeric complex with its target DNA. The structure of the RXR/VDR complex was determined by single particle cryo-EM and 3D reconstruction. Representation of the cryo-EM map with the fitted crystal structure of the individual RXR and VDR LBDs and DBDs resulting in a molecular model of the full RXR/VDR/DNA complex (top view of the complex). It has been suggested that the carboxy-terminal extension (CTE) of the DBD of VDR extending into the hinge region has a critical role for VDR transcriptional activity. The LBD interface contact comprising helix 4, loop 8/9 of VDR, and helix H7 of RXR is marked with a star. [Adapted from Orlov et al. (335), with permission from John Wiley and Sons.]
2. Hereditary vitamin D-resistant rickets
Hereditary vitamin D-resistant rickets (HVDRR) is a rare autosomal recessive disorder characterized by hypocalcemia, hyperparathyroidism, early-onset rickets, and organ resistance to 1,25(OH)2D3. The resistance to 1,25(OH)2D3 is caused by heterogeneous loss of function mutations in the VDR. Affected children may also exhibit alopecia (143). In addition to the findings related to the structure of VDR and the heterodimeric complex, biochemical and genetic analysis of VDR in HVDRR patients has also resulted in important insights related to the functional domains of VDR and the mechanisms involved in VDR signaling. Point mutations in the zinc finger DBD of VDR which caused HVDRR were the first disease-causing mutations identified in the steroid receptor gene superfamily (202). Since that report over 100 cases of HVDRR have been reported, and 45 unique mutations have been identified in VDR as the cause of HVDRR (143). Mutations have also been identified in the LBD that disrupt ligand binding [for example, R274H, a contact point for interaction with the 1α hydroxyl group (15)], that disrupt VDR/RXR interaction [for example, R391S, located in helix 10 (328)] or prevent coactivator recruitment [for example, E420 K located in helix 12 (282)]. The patient with the E420K mutation had rickets but did not have alopecia, indicating that coactivator recruitment by VDR is required to prevent rickets but is not required for hair growth. Treatment of HVDRR patients with intravenous or oral calcium administration has been reported to reverse the mineral and skeletal phenotype of HVDRR (191), suggesting the critical role of VDR/1,25(OH)2D3 action on intestinal calcium absorption. Recently, a humanized mouse model of HVDDR without alopecia was developed (254). Transgenic mice expressing the 1,25(OH)2D3-binding-defective hVDR-L233S mutant have no alopecia but still all the characteristics of rickets observed in Vdr null mice. Another study describes VDRgem mice, which express a VDR mutant that does not bind 1,25(OH)2D3, also not at supraphysiological doses, but can be selectively activated through the binding of the gemini vitamin D analog (200). These VDRgem mice show more impaired calcium and bone homeostasis compared with Vdr null mice. Both interesting models will give in the future new insights in signaling pathways regulated by unliganded or liganded VDR.
B. Genomic Mechanism of 1,25(OH)2D3 Action
1. Diversity of coregulators
The genomic mechanism of 1,25(OH)2D3 action involves the direct binding of 1,25(OH)2D3 activated VDR/RXR to specific DNA sequences [vitamin D response elements (VDREs)] in and around target genes resulting in either activation or repression of transcription. Although significant variability in the sequence of the VDREs has been reported, for activation of transcription, VDREs with high affinity for VDR consist of two direct imperfect repeats of hexanucleotides with a spacer of three nucleotides. The heterodimerization of 1,25(OH)2D3-VDR with RXR leads to high-affinity binding to VDREs. Following the binding of VDR-RXR heterodimer to the VDRE, changes in gene expression are mediated through the ability of the liganded receptor to recruit transcriptional coactivators. The p160 coactivators, steroid receptor activator 1, 2, and 3 (SRC-1, SRC-2, and SRC-3), that have histone acetylase (HAT) activity, are primary coactivators that bind to the AF2 domain of liganded VDR. The SRCs contain LxxL (x = any amino acid) motifs that facilitate binding to VDR and other nuclear receptors. Members of the p160 family recruit proteins as secondary coactivators, such as CBP/p300 (which also have HAT activity), resulting in a multisubunit complex that modifies chromatin and destabilizes histone/DNA interaction (83, 183, 362). In addition to acetylation, methylation also occurs on core histones. Recent studies have shown that methyltransferases may also play a fundamental role in VDR-mediated transcription (84, 402). Liganded VDR also interacts directly or indirectly with basal transcription factors [TFIIB and several TAT binding protein associated factors (TAFs)], resulting in the establishment of a stable preinitiation complex (83). VDR-mediated transcription is facilitated by Mediator, a multi-protein complex (the 205 subunit binds to VDR) which functions through recruitment of RNA polymerase II and promotes formation of the preinitiation complex (124, 505).
A number of other transcription factors have been reported to affect the transcriptional activity of VDR. Ras activated Ets transcription factor has been reported to have a critical role in induction of Cyp24a1 (126). VDR-induced Cyp24a1 and rat osteocalcin (Bglap) transcription are repressed by YY1, a multifunctional transcription factor (178, 375). There is increasing evidence that specific CAAT enhance binding protein (C/EBP) family members may be key mediators of 1,25(OH)2D3 action. C/EBPβ is induced by 1,25(OH)2D3 in kidney and osteoblastic cells and cooperates with 1,25(OH)2D3 and VDR in enhancing Cyp24a1 and Bglap transcription (113, 180). In the regulation of Bglap transcription, cooperation between C/EBPβ and VDR and between Runx2 and C/EBPβ has been reported (180). C/EBPs and Runx2 have also been reported to regulate VDR transcription (511). C/EBPα and VDR cooperate in the transcriptional regulation of the human antimicrobial peptide cathelicidin in lung epithelial cells and Runx2 and VDR cooperate in the transcriptional regulation of mouse osteopontin in osteoblastic cells (114, 405). C/EBPβ, Runx2, and VDR all contribute to the control of Mmp13 (matrix metalloproteinase 13) gene transcription (308).
The SWI/SNF complexes, that remodel chromatin using the energy of ATP hydrolysis, also contribute to transcriptional activation by VDR. C/EBPβ recruits the SWI/SNF complex to promote 1,25(OH)2D3 induction of Cyp24a1 and Bglap transcription (402, 468). An interplay between the SWI/SNF complex, C/EBPβ, and protein arginine methyltransferase 5 in epigenetic modification of VDR-mediated Cyp24a1 transcription has also recently been shown, suggesting that these are key factors involved in the regulation of 1,25(OH)2D3 catabolism and therefore in the maintenance of calcium homeostasis (402). It has been suggested that cell and gene specific functions of steroid receptors may be mediated through differential recruitment of coregulatory proteins (coactivators and their associated proteins and corepressors and their associated proteins) (426). Since VDR coregulatory proteins are master regulators of 1,25(OH)2D3 action, further studies identifying VDR coactivators and corepressors as well as epigenetic regulation of VDR function will yield significant new insight into the complex mechanisms by which 1,25(OH)2D3 acts to direct its multiple biological activities.
2. Genome-wide studies
The complexity of the molecular mechanisms involved in 1,25(OH)2D3 action is not only indicated by the diversity of coregulators and their activities but also through genome-wide studies which have shown that the actions of 1,25(OH)2D3, similar to other steroids, involve regulation of gene activity at a range of locations many kilobases upstream as well as downstream of the transcription start site (TSS) and within introns and intergenic regions. VDR binding to these sites is largely but not exclusively dependent on activation by 1,25(OH)2D3. Global networks regulated by VDR are beginning to be addressed in osteoblastic, intestinal carcinoma, immune, and hepatic stellate cells (72, 181, 361, 374). This review will focus on 1,25(OH)2D3-regulated genes involved in the regulation of calcium homeostasis. Recent ChIP-chip and ChIP-seq approaches have provided new insight into the mechanisms of regulation of 1,25(OH)2D3 targets in bone cells including VDR, RANKL which induces osteoclast differentiation from hematopoetic precursors, LRP5 (low-density lipoprotein receptor related protein 5) which facilitates β-catenin activation and is known to play a role in bone formation, and CYP24A1. With regard to regulation of VDR, 1,25(OH)2D3 has been shown to autoregulate VDR in bone cells not in intestine (253, 493). ChIP-seq analysis of MC3T3 osteoblastic cells revealed the presence of 1,25(OH)2D3-induced VDR and RXR at two intronic sites (+19 and +29) downstream and one intergenic site (−6 kb) upstream of the TSS of the mouse Vdr gene. These regulatory sites are conserved in the human and mouse VDR genes (510, 511). Genome-wide analysis of 1,25(OH)2D3 regulation of RANKL resulted in the identification of five distal VDREs upstream of the mouse Rankl (Tnfsfl 1) gene promoter (at −16, −22, −60, −69, and −75 kb). The most distal of the five enhancers was found to be the dominant mediator of 1,25(OH)2D3 activity in the mouse Rankl gene (233). Five enhancers were also identified in the human RANKL gene (between −20 and −96). However, unlike the mouse Rankl gene, the most proximal element at −20 kb was the dominant mediator of 1,25(OH)2D3 activity (326). The regulatory region responsive to 1,25(OH)2D3 in the mouse Lrp5 gene is at +19 kb (153). Although the induction by 1,25(OH)2D3 of RANKL as well as LRP5 suggests that 1,25(OH)2D3 action in bone cells can be anabolic as well as catabolic, it should be noted that 1,25(OH)2D3 was unable to induce VDR binding in the human LRP5 gene (153). With regard to CYP24A1, ChIP-chip and ChIP-seq studies confirmed the regulatory region previously defined, which is located proximal to the transcription start site of the mouse Cyp24a1 gene (at −160 and at −265), and in addition a novel intergenic region was identified at +35 and +37 kb (309). Occupancy of C/EBPβ was found to be enhanced in response to 1,25(OH)2D3 at −345 nt (confirming previous data obtained using chromatin immunoprecipitation) (113, 363). In intestine, transient potential vanilloid type 6 (TRPV6) is an epithelial calcium channel regulated at the transcriptional level by 1,25(OH)2D3. The human TRPV6 gene was found to contain multiple VDR/RXR binding sites (at −1.2, −2.1, −3.5, −4.3, and −5.5 kb) (310). The elements at −2.1 and −4.3 kb were found to be 1,25(OH)2D3 responsive. Collectively, these genome-wide studies have provided a new perspective on mechanisms involved in the regulation of gene expression by 1,25(OH)2D3 and suggest a chromatin looping mechanism whereby the regulatory regions can be brought into close proximity with the gene's promoter via protein-protein interaction.
IV. CLASSICAL ROLE OF VITAMIN D
A. Intestine
1. Overall process of vitamin D-mediated intestinal calcium absorption
The principal action of 1,25(OH)2D3 and the VDR is intestinal calcium absorption. This conclusion is based on the observation that mineral and skeletal phenotypes of HVDRR patients are reversed when these patients are treated with intravenous or high oral calcium (191). In addition, when Vdr null mice (which represent an animal model of HVDRR) are fed a rescue diet high in calcium and lactose, rickets and osteomalacia are prevented (18, 258, 292), further indicating that impaired bone mineralization as a consequence of defective VDR signaling results from impaired intestinal calcium absorption. Although the studies in HVDRR patients and in Vdr null mice establish the importance of the intestine in 1,25(OH)2D3-mediated regulation of calcium and bone homeostasis, the mechanisms involved in vitamin D regulation of intestinal calcium absorption have remained incomplete. The facilitated diffusion model is the most studied mechanism of vitamin D-regulated calcium absorption. In this model, transcellular calcium transport is a saturable process comprised of three 1,25(OH)2D3 regulated steps: 1) entry of calcium through the apical membrane calcium channel TRPV6, 2) binding to the calcium binding protein calbindin-D9k, and 3) extrusion of calcium across the basolateral membrane by PMCA1b. TRPV6 and calbindin-D9k have been evaluated as the major intestinal targets of 1,25(OH)2D3. They are colocalized in the intestine (similar to VDR they are expressed in all segments of the small and large intestine) and their expression is strongly correlated to transcellular calcium absorption efficiency (85, 380, 420, 459). However, studies in Trpv6 and calbindin-D9k (S100g) KO mice show that 1,25(OH)2D3-mediated calcium transport is similar to WT in the absence of TRPV6 or calbindin, suggesting compensation by other calcium channels and other calcium binding proteins yet to be identified (9, 37). Although bone mass is comparable in the WT and Trpv6 KO mice under conditions of normal dietary calcium, when dietary calcium is low, excessive bone turnover and impaired mineralization have been observed in the Trpv6 KO mice, suggesting a role for TRPV6 under low dietary calcium conditions (262) (see model Figure 3). Transgenic mice overexpressing Trpv6 in the intestine develop hypercalciuria, hypercalcemia, and soft tissue calcification (102). Thus, although there may be compensation in the KO mouse, these findings indicate a direct role for TRPV6 in intestinal calcium absorption. Unlike the single KO mice in which active intestinal calcium absorption in response to 1,25(OH)2D3 is similar to WT, the ability of the intestine to absorb calcium in response to 1,25(OH)2D3 is reduced by 60% in the Calbindin-D9k/Trpv6 double KO mice, suggesting that TRPV6 and calbindin act together to affect calcium absorption (37). It is possible that calbindin may act to modulate TRPV6-mediated calcium influx. Calbindin may also act to buffer calcium preventing toxic levels from accumulating in intestinal cells. In the cytosol, calcium may be bound to other calcium binding proteins besides calbindin. In addition, intracellular organelles could also sequester calcium in the intestinal cell. Although the mechanism involved has been a matter of debate, it has been suggested that 1,25(OH)2D3 can also stimulate active phosphate absorption in the intestine (489).
Figure 3.

Effects of 1,25(OH)2D3 in the intestine. An important function of 1,25(OH)2D3 is the stimulation of transcellular intestinal calcium transport by increasing the expression of the apical membrane calcium channel TRPV6 and calcium binding protein calbindin-D9k. The extrusion of calcium is across the basolateral membrane by PMCA1b. This process is especially enhanced when dietary calcium intake is low. Mouse genetic studies however suggest that other calcium transporters (X) are likely involved. When calcium intake is high, the paracellular calcium transport prevails, but studies suggest that this pathway may also be regulated by 1,25(OH)2D3.
2. Vitamin D and the distal intestine
Most of what is known about the mechanisms involved in 1,25(OH)2D3 regulation of intestinal calcium absorption comes from studies that utilize the duodenum. The capacity to absorb calcium is most rapid in the duodenum. However, only 8-10% of calcium absorption takes place in the duodenum (481). Although little is known about 1,25(OH)2D3 action in other parts of the intestine, the importance of regions other than the proximal small intestine has been suggested. Vitamin D and 1,25(OH)2D3-regulated calcium transport has been reported in ileum, cecum, and colon as well as duodenum (140, 141, 250). The highest expression levels of TRPV6 are in the distal intestine (517). Studies from rats and humans show that total calcium absorption is significantly higher when the colon is preserved after extensive small bowel resection (203, 491). In addition, we recently showed that transgenic expression of VDR specifically in ileum, cecum, and colon of Vdr null mice is sufficient to prevent the abnormal calcium homeostasis phenotype of Vdr KO mice (86) (Figure 4). Together these findings indicate that the distal segments of the intestine, in addition to the duodenum, play an important role in intestinal calcium absorption and proper bone mineralization.
Figure 4.

Transgenic (TG) expression of VDR specifically in ileum, cecum, and colon of VDR null mice prevents the abnormal calcium phenotype of VDR null mice. A: the full-length hVDR cDNA was introduced into the multiple cloning cassette under control of the 9.5-kb CDX2 promoter region (9.5 kb CDX2 from E. Fearon). Mice expressing VDR exclusively in the ileum, cecum, and colon were generated by breeding VDR null mice with TG mice expressing hVDR under the control of 9.5-kb CDX2. B, top panel: expression of hVDR was restricted to ileum, cecum, and colon. Mouse (m) VDR was present in WT but not in TG mice or VDR null (KO) mice. Levels of VDR in the distal intestine in TG mice were equivalent or 1.5 upregulated compared with WT. Bottom panel: Van Kossa staining of histolocial sections of tibia showing that the expression of hVDR in the distal intestine (KO/TG1 and KO/TG2) rescues the bone defects associated with systemic VDR deficiency. Serum PTH and serum calcium are normalized in KO/TG1 and KO/TG2 mice (not shown). [From Dhawan et al. (112).]
3. Paracellular calcium transport
In addition to transcellular calcium transport, calcium is absorbed by the paracellular path that occurs between epithelial cells. The vitamin D dependency of this nonsaturable component of calcium absorption has been a subject of debate and is much less defined than vitamin D-mediated transcellular calcium transport. Early studies using cultured chick intestine as well as in vivo studies in rats provided evidence that 1,25(OH)2D3 enhanced paracellular permeability (101, 223). More recent studies have shown the paracellular associated proteins including claudin-2 and claudin-12 (transmembrane components of tight junctions), cadherin-17 (a cell adhesion protein), and aquaporin 8 (a tight junction channel) can be regulated by 1,25(OH)2D3 in the intestine, suggesting that vitamin D can regulate calcium absorption by the paracellular as well as the transcellular pathway (155, 246) (Figure 3). Further studies are needed, however, to determine the role of these intercellular adhesion molecules in intestinal physiology and the significance of the regulation by 1,25(OH)2D3 in intestinal calcium absorption.
Thus 1,25(OH)2D3-mediated intestinal calcium absorption is more complex than has been suggested by the three-step model. Future studies defining the multiple mechanisms by which 1,25(OH)2D3 acts in both proximal and distal segments of the intestine are needed to identify new therapeutic approaches to sustain calcium balance.
B. Kidney
1. Overall process of renal calcium reabsorption: role of 1,25(OH)2D3
Most of the calcium that is filtered through the glomerulus will be reabsorbed in both the proximal and distal tubule resulting in only 1% to 2% of filtered calcium appearing in the urine. Approximately 65% of the filtered calcium is passively reabsorbed at the proximal tubules in a 1,25(OH)2D3-independent way. In the distal tubules, calcium absorption is regulated by 1,25(OH)2D3 and PTH. Calcium reabsorption in the proximal tubule is passive and follows a sodium gradient, whereas calcium reabsorption in the distal tubule involves an active transcellular mechanism and resembles intestinal calcium absorption (see sect. IVA1). The model consists of calcium entry through TRPV5, transfer of calcium in the cytoplasm by binding to calbindin-D9k and calbindin-D28k, and calcium extrusion by the sodium/calcium exchanger (NCX1) and plasma membrane calcium pump 1b. Inactivation of Trpv5 results in hypercalciuria, but normocalcemia is maintained in these mice by compensatory increase of intestinal calcium absorption stimulated by high serum 1,25(OH)2D3 levels (193). These findings suggest that calcium uptake by TRPV5 is a rate-limiting step in renal calcium reabsorption. Calbindin-D28k deletion has no effect on urinary calcium excretion, but its role is mostly compensated by calbindin-D9k (520). Ablation of Calbindin-D28k in Trpv5 null mice did not worsen the Trpv5 null phenotype (165). Active renal calcium reabsorption is regulated by PTH and 1,25(OH)2D3, which both increase calcium reabsorption. Indeed, Cyp27b1 null mice show decreased expression of TRPV5, calbindin-D9k, calbindin-D28k, and NCX1 mRNAs, and this reduced expression was rescued by 1,25(OH)2D3 treatment (192). However, in the different Vdr null strains, only calbindin-D9k mRNA was consistently decreased (133, 460, 506). Nevertheless, Vdr null mice display reduced renal calcium reabsorption, as shown by the inappropriately high urinary calcium levels given the hypocalcemia (133, 259). Besides PTH and 1,25(OH)2D3, αKlotho and FGF23 can also regulate TRPV5 expression. Two pathways are proposed: the first model states that αKlotho hydrolyzes extracellular residues of TRPV5 and thereby ensures that TRPV5 is entrapped in the apical plasma membrane (74); the second model, proposed by a recent study, suggests that FGF23 signaling through the FGFR1-αKlotho complex at the basolateral membrane regulates intracellular TRPV5 trafficking and TRPV5 abundance at the apical membrane (20). In accordance with these findings, αKlotho and Fgf23 null mice exhibit hypercalciuria (14, 20) (Figure 5, bottom panel).
Figure 5.

Renal VDR actions. In the proximal tubule cells, CYP27B1 expression is suppressed by 1,25(OH)2D3 and FGF23. FGF23 also stimulates phosphate excretion by decreasing the expression of the phosphate transporters NPT2a/c in the apical membrane. FGF23 may signal by binding to the few FGFR1-Klotho complexes in the proximal tubules or by inducing a paracrine factor (factor X) in the distal tubules where abundant FGFR1-Klotho complexes are present. Renal calcium reabsorption in the distal tubule is stimulated by 1,25(OH)2D3. 1,25(OH)2D3 increases the expression of calbindin-D9k and calbindin-D28k and to a lesser extent of TRPV5. The extrusion of calcium at the basolateral side is mediated by PMCA1b and NCX1. Two models are proposed on how Klotho and FGF23 regulate TRPV5 expression: 1) secreted Klotho (sKlotho) is considered to hydrolyze sugar residues from the glycan chains on TRPV5 resulting in better entrapment of TRPV5 in the apical membrane; and 2) FGF23 binds to the basolateral FGFR1-Klotho complex, which stimulates intracellular transport of TRPV5 to the plasma membrane.
2. Regulation of 1,25(OH)2D3 synthesis and phosphate absorption
The proximal tubules of the kidney are also the major site of 1,25(OH)2D3 synthesis (see sect. IIB; CYP27B1) and of phosphate absorption. CYP27B1 expression is upregulated by PTH but downregulated by FGF23 and 1,25(OH)2D3 (see sect. IIE: regulation of renal CYP27B1 and CYP24A1) (Figure 5, top panel). Approximately 80% of filtered phosphate is reabsorbed from urine under normal dietary phosphate intake and most of it occurs within the proximal tubule (469a). Phosphate transport across the proximal tubule epithelium is mediated by sodium-phosphate cotransporters NPT2a and NPT2c and the energy derived from the transport of sodium down its gradient is used to transport phosphate into the cell (49a). This phosphate reabsorption in the proximal tubules is regulated by several factors including FGF23, PTH, and 1,25(OH)2D3. PTH and FGF23 promote renal phosphate loss by decreasing the abundance of the sodium-phosphate cotransporter (NPT2a/2c) at the apical membrane: PTH stimulates the internalization and lysosomal degradation of these transporters (27), whereas FGF23 decreases their expression (507) (Figure 5, top panel). How FGF23 signaling regulates NPT2a and NPT2c expression is not fully clarified since αKlotho is predominantly expressed in the distal tubules, whereas the effects of FGF23 on phosphate absorption are mainly observed in the proximal tubules (257). Possibly, the small number of FGFR1-αKlotho complexes found in the proximal tubules is sufficient for signaling. On the other hand, recent findings suggest that a paracrine factor is released from the distal tubules that acts on adjacent proximal tubules (139). Beside PTH and FGF23, 1,25(OH)2D3 may regulate phosphate homeostasis by increasing FGF23 expression in osteocytes (see sect. IID) and αKlotho expression in the distal tubule (150), factors that both stimulate renal phosphate loss.
Thus 1,25(OH)2D3 action regulates renal calcium reabsorption and phosphate loss, but the molecular mechanism is still incompletely characterized.
C. Bone
1. Link between bone metabolism and calcium homeostasis
Calcium and bone homeostasis are highly intertwined, as calcium is a major constituent of the bone and provides strength to the skeleton, but the bone is also the largest store of calcium in the body. The bone structural integrity thus relies on sufficient calcium supply from the serum and therefore indirectly from intestinal calcium absorption and renal calcium reabsorption, but on the other hand, calcium can be removed from the bone to preserve normal serum calcium levels in case of a negative calcium balance. In the adult, bone is continuously remodeled, and bone resorption by osteoclasts is in balance with bone formation by osteoblasts to maintain bone mass. During growth, bone lengthening is highly dependent on the coordinated growth and differentiation of chondrocytes. Studies of humans and mice lacking VDR or CYP27B1 systemically have evidenced that the bone characteristics of rickets and osteomalacia are rescued when sufficient calcium absorption is ensured by dietary (18, 107, 258, 350, 351, 446) or genetic (499) means, indicating an indirect role of VDR signaling for bone homeostasis by regulating intestinal calcium absorption and controlling phosphate homeostasis. Indeed, the low serum phosphate levels in Vdr null mice decrease apoptosis of hypertrophic chondrocytes resulting in widening and expansion of the epiphyseal growth plate (123, 389). In addition, the mineral supply for bone matrix mineralization is decreased because of the hypocalcemia and hypophosphatemia, leading to osteomalacia. Nevertheless, VDR has specific actions in osteogenic cells, and we will first discuss the contribution of VDR signaling to mineral homeostasis, next its role for bone metabolism, and finally the effects of vitamin D supplementation as part of the strategies to treat osteoporosis.
2. The paracrine role of VDR signaling in osteogenic cells for mineral homeostasis
Osteoblast VDR signaling participates in calcium metabolism primarily during a negative calcium balance. Indeed, in the situation that dietary calcium acquisition is lower than bodily calcium use and renal calcium loss, calcium is mobilized from the bone to preserve normal serum calcium levels. This skeletal response was especially evidenced in a mouse model lacking Vdr expression specifically in the intestine, resulting in markedly reduced intestinal calcium absorption (263). In this condition, serum levels of PTH and 1,25(OH)2D3 increase, which lead to a marked depletion of calcium from the bone to maintain normal serum calcium levels. The effect on the bone consisted of increased bone resorption accompanied by impaired bone mineralization (Figure 6).
Figure 6.

Skeletal effects of 1,25(OH)2D3 signaling. During a negative calcium balance, when VDR action in the intestine is impaired or dietary calcium intake is low, intestinal calcium absorption is decreased. Normal serum calcium levels can however be maintained by increased 1,25(OH)2D3 and PTH levels, which will increase bone resorption and reduce bone matrix mineralization. During a normal or positive calcium balance, normal serum 1,25(OH)2D3 levels promote intestinal calcium absorption. This pathway will deliver sufficient calcium for adequate bone matrix mineralization. VDR signaling in osteoprogenitors increases RANKL expression and stimulates osteoclastogenesis, whereas VDR action in mature osteoblasts has anticatabolic actions, by decreasing RANKL expression, and anabolic effects by stimulating LRP-5 signaling.
VDR signaling enhances bone resorption mostly indirectly by acting on osteoblasts rather than on osteoclasts. Indeed, osteoblast VDR signaling exerts direct transcriptional control on the expression of RANKL, an important osteoclastogenic factor (233) (see sect. IIIB2). RANKL binds to its cognate receptor RANK in osteoclast precursors and increases osteoclast formation and action (431) (Figure 6). This action can be blocked by the naturally occurring soluble decoy receptor of RANKL, termed osteoprotegerin (OPG). In vitro co-culture experiments have shown that osteoblast VDR signaling is necessary for 1,25(OH)2D3-induced osteoclast formation, whereas VDR activity in osteoclasts is not (435). The increase in bone resorption during a negative calcium balance is necessary to maintain normocalcemia, as evidenced by a reduction in serum calcium levels when bone resorption is pharmacologically blocked in the intestinal-specific Vdr null mice (263).
Besides stimulating bone resorption during a negative calcium balance, 1,25(OH)2D3 also inhibits bone matrix mineralization and thereby contributes to preserving normal serum calcium levels (263). These mineralization defects are characterized by abundant unmineralized bone matrix and reduced mineral content of the mineralized bone. Mechanistically, 1,25(OH)2D3 suppresses mineralization by increasing the pyrophosphate (PPi) levels and Osteopontin expression, both potent mineralization inhibitors. VDR signaling in osteoblasts regulates PPi levels by controlling the expression of several genes: increasing the expression of ectonucleotide pyrophosphatase phosphodiesterase (Ennp)1 and Ennp3, which generate PPi from trinucleotides, and increasing the levels of progressive ankylosis (Ank), which mediates the transport of PPi (169, 263, 307). VDR signaling in osteogenic cells not only influences mineral homeostasis by regulating local processes of bone mineralization and remodeling, but also controls calcium and phosphate homeostasis in an endocrine manner by stimulating FGF23 expression as has been discussed in section IIE.
3. Role of osteoblastic/osteocytic VDR signaling in bone homeostasis
As discussed, VDR signaling in osteogenic cells during a negative calcium balance is mainly directed to preserve serum calcium levels, and the increased bone resorption and impaired bone mineralization occur at the expenses of skeletal integrity. The role of VDR signaling in bone cells during a positive calcium balance is still not fully elucidated, but the specific effects likely depend on the osteoblast differentiation stage (Figure 6). VDR signaling in osteoprogenitors and osteoblasts has a positive effect on osteoclast formation and bone resorption and thus negatively regulates bone mass, as shown by Vdr inactivation in mice using the collagen type I promoter which resulted in increased bone mass (501). On the other hand, VDR activity in more mature osteoblasts has anabolic and anticatabolic activity and increases bone mass, as evidenced by Vdr overexpression using the osteocalcin promotor (29, 157). The anti-resorptive effect is mediated by decreased RANKL/OPG ratio, whereas the anabolic effect may rely on increased LRP-5 expression. The mouse Lrp-5 gene is regulated by VDR signaling. LRP-5 functions as a co-receptor in wingless (Wnt) signaling (153), a pathway known to mediate anabolic effects in osteoblasts. Finally, VDR signaling in osteocytes is redundant for bone metabolism, as Vdr inactivation in mature osteoblasts and osteocytes [using the dentin matrix protein 1 (Dmp1) promoter] has no effect on bone mass, formation, resorption, or mineralization (263). Since all these osteogenic differentiation stages coexist, the physiological relevance of the differential, and even opposing, effects of VDR signaling in osteogenic cells are still not fully defined and need further investigation.
4. VDR signaling in chondrocytes
Not only osteoblasts and osteocytes express the vitamin D machinery, but also growth plate chondroyctes express the VDR (478). Mouse genetic studies using Vdr inactivation in the chondrocytes (collagen 2 promoter) have indicated that VDR signaling in these cells is especially important during bone growth, when chondrocytes are abundantly present (293). In young mice, VDR activity in chondrocytes regulates RANKL expression and hereby trabecular bone remodeling. In addition, it indirectly contributes to FGF23 production in osteocytes and thereby vitamin D homeostasis. These effects diminish in adult mice, when VDR signaling in osteoblasts and osteocytes become more important as these cell types are then the major source of RANKL as well as FGF23 (322, 373, 496).
5. Human studies
Severe vitamin D deficiency in children, due to lack of exposure of sunlight and low vitamin D intake, is still endemic in several areas of the world (166, 369). In these children, serum 25OHD levels are usually below 10 ng/ml, which is considered as the threshold for 25OHD to control intestinal calcium absorption (55, 325). Below 10 ng/ml 25OHD there is a deficit in substrate leading to lower serum1,25(OH)2D levels and a decrease in intestinal calcium absorption (325). Extensive clinical experience and some randomized control trials indicate that daily intake of 400 IU vitamin D3 is sufficient to prevent this type of rickets in children (55, 264). In adults, clinical vitamin D-related osteomalacia is usually found in individuals with low sun exposure or in patients with impaired intestinal vitamin D absorption as part of intestinal fat malabsorption, like after bariatric surgery or with inflammatory bowel disease (395).
A negative calcium balance is often found in ageing individuals and is explained by low dietary calcium intake, vitamin D deficiency, and a decrease in VDR levels in the intestine (127) leading to reduced intestinal calcium absorption. As a consequence, SHPT develops with increased bone resorption and decreased bone mineralization (45, 241, 370). Numerous clinical trials and meta-analyses have therefore been performed to investigate the effect of vitamin D supplementation with or without calcium on fracture incidence. In general, the effect of vitamin D alone compared with placebo had no effect on fracture risk, whereas meta-analyses on the combination of vitamin D and calcium were inconclusive showing a 12–26% reduction in fracture risk in some meta-analyses, but no effect in others (264). Several factors may explain these inconsistencies. The dose of vitamin D has to be adequate (>400 IU) to reduce fracture risk, and dosage has been different between several clinical trials (46). In addition, baseline values of serum 25(OH)D3 may be different and vitamin D supplementation is mainly effective in individuals with documented or at high risk of vitamin D deficiency and low calcium intake (90, 99). Finally, compliance and persistence with calcium and vitamin D are essential, but compliance with the supplements is often low in healthy and community-dwelling individuals (439). In general, a vitamin D supplement of 600–800 IU per day in combination with calcium may reduce the incidence of nonvertebral fractures by ∼10–20% in old, vitamin D-deficient population (386).
Thus vitamin D actions control bone metabolism mainly indirectly by regulating mineral homeostasis, but the exact role of the VDR in osteogenic cells for bone homeostasis during a normal calcium balance requires further investigation. Several ongoing large-scale clinical trials will help to define the dosage of vitamin D supplement that is best for skeletal health.
V. PLEIOTROPIC ACTIONS OF VITAMIN D
Over the course of the last decades, it has become increasingly clear that the effects of 1,25(OH)2D3 are not limited to the maintenance of calcium and phosphate homeostasis. Indeed, 1,25(OH)2D3 regulates multiple cellular processes with effects on normal and malignant cell growth and differentiation (including differentiation of keratinocytes; see Ref. 43 for review), on the innate and adaptive immune function, on cardiovascular function, and on the complex interplay with other hormones. In this review we focus on effect on cancer, the cardiovascular system (including results of randomized controlled trials), and the immune system.
A. Cancer
1. VDR expression and vitamin D metabolism in cancer cells
Already more than three decades ago, Colston et al. (95) demonstrated that doubling times of melanoma cells increase after treatment with 1,25(OH)2D3. Abe et al. (3) reported shortly thereafter that HL60 leukemia cells differentiate towards the macrophage lineage upon incubation with 1,25(OH)2D3. Ever since, numerous studies have shown that 1,25(OH)2D3 and its analogs slow down cancer cell growth by arresting cells in the G0/G1 phase of the cell cycle, by inducing their differentiation or by the induction of apoptotic cell death. Furthermore, 1,25(OH)2D3 influences angiogenesis, alters cell adhesion and migration, and reduces the invasiveness of cancer cells. Interestingly, most cancer cells do not only express VDR, but also CYP27B1 and CYP24A1, which allows the cells to locally regulate 1,25(OH)2D3 metabolism. Although locally produced 1,25(OH)2D3 concentrations are considered not to contribute to calcium homeostasis, they may have significant implications for cancer cell progression (190, 433).
a) vdr. The presence of the VDR in tumor cells is a prerequisite for the antineoplastic effects of 1,25(OH)2D3. In most tumors, VDR expression is retained and, according to Narvaez et al. (323), alterations in the VDR gene are only seen in 5% of cancers in The Cancer Genome atlas. Several studies suggested that enhanced tumor VDR expression levels are correlated with a better prognosis and prolonged overall survival (119, 186). This was recently confirmed by Santagata et al. (391) who used large-scale immunohistochemistical stainings to develop a phylogenetic classification scheme. Their data showed that tumors that expressed next to the estrogen receptor (ER) and the androgen receptor (AR) also the VDR had the better prognosis and they suggested that ER/AR/VDR expression was correlated with the differentiation grade. Moreover, these observations were more pronounced at the protein level than at mRNA level (391). Interestingly, more than 900 allelic variants have been described at the VDR locus (39), and numerous studies have investigated the association of SNPs in VDR (e.g., ApaI [rs7975232], BsmI [rs1544410], FokI [rs10735810], TaqI [rs731236]) and cancer risk with inconclusive results (179, 238, 270, 516).
b) cyp27b1. Expression of CYP27B1 has been extensively studied in cancer cell lines as well as in primary tumors. In cancer cell lines, enhanced as well as reduced expression of CYP27B1 is reported (488). Interestingly, upon oncogenic transformation of a mammary epithelial cell line CYP27B1 expression levels decrease significantly which results in a reduced cellular sensitivity to 25(OH)D3 and 1,25(OH)2D3 (227). Recently, Narvaez et al. (323) reported that in The Cancer Genome Atlas datasets <2% of breast cancers exhibit genomic alterations (including amplifications, deletions, mutations, and changes in mRNA) (323). Nonetheless, multiple studies reported that in human cancer biopsies CYP27B1 mRNA expression (32, 47) and CYP27B1 protein levels (93, 272, 299) tend to be higher in well-differentiated tumors, whereas they are lower in more malignant and poorly differentiated tumors. In addition, in lung cancer, high CYP27B1 is associated with better overall survival (237). Multiple single nucleotide polymorphisms (SNP) exist in the gene that encodes for CYP27B1. Although conflicting results are reported on the associations of SNPs in the CYP27B1 gene and cancer risk, these SNPs may alter 1,25(OH)2D3 production as these SNPs may lead to a reduced enzymatic activity (207). Although associations have been reported, it is unclear at this time whether cancer cell CYP27B1 has a role in affecting disease progression.
c) cyp24a1. Not only the enzyme responsible for the production of 1,25(OH)2D3, but also the catabolic enzyme of 1,25(OH)2D3, CYP24A1, is expressed in cancer cells. CYP24A1 expression has been reported to be enhanced in the more malignant and metastatic tumors. It has been suggested that increased CYP24A1 expression is associated with increased resistance to 1,25(OH)2D3 action (154). Increased CYP24A1 levels may result from gene amplification as was demonstrated in breast tumors, where the chromosomal region 20q13.2, which contains the CYP24A1 gene, was found to be amplified (13). Also in colorectal cancer, increased CYP24A1 gene copy number is shown, whereas no differences in CYP24A1 promoter methylation are seen (189). In agreement with these findings, 10–13% of human breast cancers in the dataset from The Cancer Genome Atlas show altered CYP24A1 expression, most often due to gene amplification and characterized by enhanced mRNA levels (323). Of note, high CYP24A1 expression significantly correlates with poor survival in lung cancer cohorts (50). In addition, in the context of p53 loss, suppression of CYP24A1 caused by inhibition of the miR-17∼92 cluster was toxic in non-small cell lung cancer (50). Genetic variants in CYP2R1, 7-DHCR but also in CYP24A1 (rs6013897) are significantly correlated with vitamin D status as was reported in two recent genome-wide association studies (6, 475). Of interest, many SNPs in the CYP24A1 gene are characterized by a reduced enzymatic activity, suggesting that vitamin D catabolism may be influenced by genetic factors (207). Multiple recent studies have investigated whether polymorphisms in the CYP24A1 gene could be associated with cancer risk. However, until now, no consensus exists on the association between common variants in the CYP24A1 gene and cancer risk (316, 379, 394). This might not be surprising as the described SNPs only explain a small amount of the variation in 25(OH)D3 levels, and therefore the contribution of the specific SNPs to the predicted cancer risk may be too weak to be identified in genome-wide association studies (142).
2. In vitro antineoplastic effects of 1,25(OH)2D3
a) antiproliferative effects. One of the earliest and best-described effects of 1,25(OH)2D3 includes its growth-inhibitory and prodifferentiation effects. Indeed, differentiation along the macrophage lineage of HL60 cells is accompanied by a reduction in cell proliferation (3). However, regulation of differentiation and cell proliferation are not always coupled and seem to be cell-type dependent (38). In most cancer cells expressing a functional VDR, incubation with 1,25(OH)2D3 leads to an accumulation of cells in the G0/G1 phase of the cell cycle (211). Downregulation of the abundance of cyclins and cyclin-dependent kinases (CDKs) and/or upregulation of different CDK inhibitors, such as p21 and p27, by 1,25(OH)2D3 results in a reduced CDK activity, the formation of an intact retinoblastoma (Rb)-E2F complex, a decrease in E2F and E2F-target genes, and subsequent growth inhibition (355, 441, 469, 477) (Figure 7). However, when Rb is downregulated in prostate cells, 1,25(OH)2D3 is still able to retard the growth of these cells, suggesting that redundant growth inhibitory pathways compensate for the loss of Rb (479). In analogy, Rb-deficient murine embryonal fibroblasts (MEFs) remained sensitive to the growth-inhibitory effect of 1,25(OH)2D3, whereas the antiproliferative effect of 1,25(OH)2D3 is lost in MEFs in which the pocket proteins p107 and p130 were both deleted (467). In addition, induction of C/EBPα by 1,25(OH)2D3 and enhancement of VDR transcription by C/EBPα has been suggested as one mechanism involved in 1,25(OH)2D3-mediated inhibition of proliferation of breast cancer (115). In human colon carcinoma, 1,25(OH)2D3 antagonizes the Wnt/β-catenin signaling, which finally results in a reduced cell proliferation. Treatment with 1,25(OH)2D3 leads to a diminished interaction between β-catenin and T-cell factor (TCF) in favor of an enhanced interaction between VDR and β-catenin. Moreover, 1,25(OH)2D3 enhances E-cadherin expression, leading to the nuclear export of β-catenin, and induces the expression of Dickkopf (DKK) 1, an extracellular Wnt inhibitor. As a consequence, the transcription of TCF-target decreases, among which that of c-myc, a key regulator of cell cycle progression (246a). Very recently, Chang et al. (75) reported that 1,25(OH)2D3 induces the expression of the microRNA miR-145 in a dose- and VDR-dependent manner. Interestingly, inhibition of miR-145 abrogates the 1,25(OH)2D3-induced downregulation of E2F3 and reverses the growth-inhibitory effect of 1,25(OH)2D3 (75). In addition, 1,25(OH)2D3 also interferes with other growth regulatory pathways initiated by transforming growth factor (TGF)-β (76), epidermal growth factor (36), insulin-like growth factor (58), platelet-derived growth factor (324), and fibroblast growth factor 2 (385). Moreover, it intervenes in other mitogenic signaling pathways (e.g., ERK/mitogen-activated protein kinase pathway and c-myc) (344, 480). In BRCA1-positive breast cancer cells, liganded VDR associates with BRCA1, and this complex occupies VDREs in the p21 promoter to enhance promoter acetylation and p21 expression, revealing a novel aspect of BRCA unrelated to DNA repair (360).
Figure 7.

1,25(OH)2D3-induced signaling pathways involved in the regulation of cell proliferation, apoptosis, and inflammation in cancer. 1,25(OH)2D3 hampers the transition from the G1 to the S phase of the cell cycle either directly, through upregulation of different cyclin-dependent kinase inhibitors, or indirectly through the induction of other growth factors (e.g., TGF-β, EGF). In addition, 1,25(OH)2D3 induces apoptosis through activation of the intrinsic apoptotic pathway or by interference with other signaling pathways such as TNF-α, EGF, β-catenin, and prostaglandins. 1,25(OH)2D3 has also an immunosuppressive activity, as indicated by the repression of NFκB-mediated gene transcription, which results in a suppressed production of inflammatory cytokines, such as IL-1, IL-6, IL-8, and TNF-α.
Recent work demonstrated that VDR acts as transcriptional regulator in pancreatic stellate cells and primes them to differentiate towards a more quiescent phenotype (117). These findings are potentially very interesting because there is increasing evidence that activated pancreatic stellate cells are characterized by a pathological matrix secretion, which results in a physical barrier for chemotherapy. Moreover, they produce mitogenic factors that may promote pancreatic cancer cell proliferation, survival, and migration. Interestingly, when the vitamin D analog Calcipotriol is administered in mice with pancreatitis, less fibrosis and inflammation is observed. In addition, extensive stromal remodeling is induced, whereas tumor-supportive signaling is reduced. As a consequence, the efficacy of a co-administered chemotherapeutic agent is enhanced, and in parallel, survival increases. Previous work from the same research group on hepatic stellate cells reveals that VDR promotes quiescence of hepatic stellate cells by temporally inhibiting TGF-β1/mothers against decapentaplegic homolog 3 (SMAD3) signaling via genomic competition (406). Recent research has pointed out that 1,25(OH)2D3 is able to target the cancer stem cell population. Cancer stem cells have the capacity to continuously self-renew and retain multilineage differentiation potential. Therefore, these cells constitute a relevant target for chemoprevention and chemotherapy. Indeed, upon growth arrest and differentiation, these cells will lose their self-renewal competence and their capacity to initiate tumorigenesis. Maund et al. (300) demonstrated that upon incubation with 1,25(OH)2D3 normal adult prostate progenitor/stem cells undergo cell-cycle arrest, senescence, and differentiation. In breast cancer research, several studies have been performed on mammosphere cell cultures to enrich for mammary progenitor cells and putative breast cancer stem cells. In mammosphere cultures of SKBR3, MCF7, and HMLEH-RAS breast cancer cell lines, VDR expression is downregulated, and little growth inhibition is seen upon treatment with 1,25(OH)2D3. Upon overexpression of VDR, the ability to form mammospheres is reduced and cell differentiation increased (358). On the other hand, in MCF10DCIS mammosphere cultures, 1,25(OH)2D3 and its analogs decrease mammosphere forming efficiency and repress markers that are associated with stem cells and pluripotency such as CD44, CD49f, c-Notch1, and OCT4 (416, 417, 471). In pancreatic cancer, 1,25(OH)2D3 also suppresses cancer cell stemness through inhibition of FOXM1 signaling (261).
b) effects on apoptosis. 1,25(OH)2D3 induces apoptosis in a wide variety of cancer cells, and although the underlying mechanisms seem to be cell type-specific, several studies indicated that the nuclear VDR is required for the proapoptotic effects of 1,25(OH)2D3 (171, 526). Many studies have pointed to an activation of the intrinsic apoptotic pathway by 1,25(OH)2D3 as illustrated by suppression of the antiapoptotic B-cell lymphoma 2 (Bcl2) proteins and B-cell lymphoma-extra large (Bcl-xl) and activation of the proapoptotic protein Bax (Figure 7). This subsequently leads to the release of cytochrome c from the mitochondria and the activation of downstream caspases such as caspase 3 and 9 (171, 470, 515). In addition, 1,25(OH)2D3 may induce apoptosis by interfering with other signaling pathways such as tumor-necrosis factor (TNF)-α (168, 303). Interestingly, the induction of apoptosis by the vitamin D analog EB1089 in MCF-7 cells is suggested to occur through a pathway that involves Beclin 1-dependent autophagy (197).
In contrast, in acute myeloid leukemia (AML), treatment with 1,25(OH)2D3 results in differentiation of these cells which is accompanied by an enhanced cell survival. Changes in the anti-apoptotic protein Mcl-1 and in the Bcl2/Bad ratio contribute to these prosurvival effects (477). Recently, the increased expression of miR-32 in AML cell lines after incubation with 1,25(OH)2D3 has been implicated in the antiapoptotic effect of 1,25(OH)2D3 through suppression of the proapoptotic protein Bim (167).
c) effects on migration and invasion. Cancer invasion into the surrounding tissue is an important hallmark of cancer and is regulated by signaling pathways that regulate the cytoskeleton, induce the turnover of cell matrix, and control cell-cell junctions and cell adhesion. Interestingly, 1,25(OH)2D3 impacts each of these different processes. Indeed, in squamous cell carcinoma, 1,25(OH)2D3 leads to an altered cell morphology and actin organization (276). Moreover, 1,25(OH)2D3 inhibits expression of the cytoskeletal protein vimentin, which results in a decreased cell motility (449). Of note, in MCF-7 breast cancer cells, PDZ-LIM domain-containing protein 2 (PDLIM2), an adaptor molecule that links different components of the cytoskeleton, is identified as a direct target gene of 1,25(OH)2D3. Furthermore, the induction of PDLIM2 by 1,25(OH)2D3 mediates the proadhesion, anti-migration, and anti-invasion effects of 1,25(OH)2D3 (466). Different proteases such as the matrix metalloproteinases (MMPs), the plasminogen activators (PAs), and the cathepsins (CPs) are involved in tumor invasion as they are able to degrade the extracellular matrix (30). 1,25(OH)2D3 not only reduces the expression and secretion of metalloproteinase (MMP) 2 and 9, but also decreases cathepsin K activity, increases tissue inhibitor of MMP1 (TIMP1) and regulates different components of the plasminogen activator system (30, 204, 235). In addition, in multiple cell types treatment with 1,25(OH)2D3 leads to an altered expression of adhesion molecules. As such, the cell surface adhesion molecules α6 integrin and β4 integrin (432) as well as the intracellular adhesion molecule 1 (428) are decreased upon treatment with 1,25(OH)2D3. Expression of E-cadherin, a tumor suppressor that is inversely correlated to metastasis, is induced by 1,25(OH)2D3 in different cell types, and this induction may lead to suppression of cellular motility (66, 347).
d) effects on inflammation in cancer. Numerous studies demonstrated that 1,25(OH)2D3 has an immunosuppressive activity by stimulation of the innate immune system and by suppression of the adaptive immune system (457). Because chronic inflammation is regarded as a risk factor for the development of cancer, the suppression of inflammation by 1,25(OH)2D3 may contribute to its antineoplastic activity. In cancer, the synthesis of prostaglandins, small-molecule derivatives of arachidonic acid which play a key role in the generation of the inflammatory response, is targeted by 1,25(OH)2D3. Indeed, 1,25(OH)2D3 leads to a decrease in cyclooxygenase-2 and an increase in 15-hydroxyprostaglandin dehydrogenase resulting in a reduced prostaglandin synthesis (319, 444). In addition, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling, a major mediator of the immune response, is inhibited in various cancers by 1,25(OH)2D3 by acting on different members of this pathway (304, 455) (Figure 7). A suppressed production of inflammatory cytokines, such as interleukin (IL)-1, IL-6, IL-8, IL-17, and TNF-α is consistently demonstrated in in vitro and in vivo cancer models (5, 162, 240, 330) (Figure 7). In a preclinical SMAD3 knockout model of colitis and colon cancer, Meeker et al. (304) showed that a high vitamin D diet decreases NF-κB activation in colonic epithelial cells, which is reflected by decreased inflammatory cell infiltrates and reduced expression of proinflammatory cytokines during initiation of carcinogenesis (304). In line with this, a strong inverse association is found between 25(OH)D status and the risk of colorectal cancer with high-level lymphocytic reactions which suggests a role for vitamin D in cancer immunoprevention through modulation of the host-tumor interaction (419). A recent whole genome profiling on cells derived from normal mammary tissue and breast cancer reveals the regulation of a set of immune genes by 1,25(OH)2D3. Among these genes, soluble CD14b, which may contribute to the protection against breast cancer, seems to be consistently induced by 1,25(OH)2D3 (412). Induction of CD14 by 1,25(OH)2D3 is confirmed by a microarray study on fresh tumor slices exposed to 1,25(OH)2D3 (311).
e) effects of 1,25(oh)2d3 on cell metabolism. Recent research has focused on the role of 1,25(OH)2D3 on cellular energy metabolism in the context of cancer. In contrast to quiescent cells, which rely mainly on oxidative phosphorylation for their energy demands, most cancer cells rely on aerobic glycolysis to provide energy and biosynthetic intermediates for the production of nucleotides, amino acids, and fatty acids to sustain their rapid proliferation (464). Therefore, alterations in energy metabolism by 1,25(OH)2D3 may contribute to its antineoplastic activity. In untransformed human breast epithelial cells MCF10A as well as in ras-transformed MCF10A cells, treatment with 1,25(OH)2D3 results in reduced glycolysis. In both cell types flux of glucose to acetyl-coA and flux to oxaloacetate is decreased, indicative of a reduced tricarboxylic acid (TCA) cycle activity. However, further research is required to decipher how the 1,25(OH)2D3-induced metabolic reprogramming contributes to its chemopreventive capacity (518). Consiglio et al. (96) genetically silenced VDR expression in HaCaT keratinocytes, which are insensitive to the antiproliferative effects of 1,25(OH)2D3. In these cells, VDR silencing results in a clear growth reduction and a strong increase in mitochondrial membrane potential that leads to oxidation of metabolic intermediates that can no longer be used in biosynthetic pathways. Intriguingly, these results are indicative of a role of the VDR as an enhancer of cell proliferation (96). Microarray profiling of nontumorigenic mammary epithelial cells after treatment with 1,25(OH)2D3 revealed the regulation of a whole set of metabolic genes such as SLC1A1 and GLUL by 1,25(OH)2D3 (412). SLCA1 encodes a plasma membrane glutamate transporter that is induced by 1,25(OH)2D3, whereas GLUL, which encodes glutamine synthetase, is repressed by 1,25(OH)2D3. These changes are accompanied by an accumulation of glutathione and changes in respiratory capacity, suggesting that 1,25(OH)2D3 leads to a metabolic switch that might induce quiescence in mammary epithelial cells (323).
f) regulation of micrornas by 1,25(oh)2d3. MicroRNAs (miRNAs) are endogenous (∼22 nucleotides long) single-strand RNA molecules that target mRNAs for cleavage or translational repression and hence fulfill important regulatory roles in a wide variety of biological processes (34). miRNAs usually target sequences at the 3' end of a gene, which position facilitates the control of mRNA stability. The regulation of miRNA expression by 1,25(OH)2D3 is an emerging field of interest and has been shown to be part of the signaling cascades responsible for the effects of 1,25(OH)2D3 on cell proliferation, differentiation, apoptosis, and gene regulation (Table 1). Sing et al. (414) undertook miRNA microarray profiling in different prostate cell lines ranging from nonmalignant to highly malignant cell types and showed that miRNA expression profiles are highly cell-specific. In addition, they demonstrated in nonmalignant RWPE-1 cells that miRNAs are significantly coregulated with mRNA regulation (414). Such integration studies will be important to improve miRNA target predictions and to unravel the complex signaling pathways induced by 1,25(OH)2D3.
Table 1.
Overview of miRNAs that are involved in antineoplastic effects of 1,25(OH)2D3
| microRNA | Cell Type | Regulation by 1,25(OH)2D3 | Target of microRNA | Biological Effect | Reference Nos. |
|---|---|---|---|---|---|
| miR-22 | Colon cancer, SW480-ADH, and HCT116 cell lines | Increased expression | Implicated in antiproliferative and antimigratory effect of 1,25(OH)2D3 | 17 | |
| miR-27b mmu-miR-298 | Colon adenocarcinoma LS-180 and pancreatic PANC1 cell lines | Not investigated | VDR | Overexpression of miR-27b and mmu-miR-298 results in reduced VDR protein levels | 349 |
| miR-27b | Melanoma SK-Mel5, SK-Mel28, and IGR cell lines | Reduced expression in conjunction with 5-azacytidine | VDR | Reduced miR-27b expression correlates with increased VDR mRNA | 135 |
| miR-32 | Human myeloid leukemia HL60 and U937 cells | Increased expression | BIM | Implicated in apoptotic cell death after 1,25(OH)2D3 treatment | 167 |
| miR-98 | Prostate cancer LNCaP cells | Increased expression | CCNJ | Implicated in the antiproliferative effect of 1,25(OH)2D3 | 445 |
| miR-100 miR-125b | Prostate cancer RWPE-2 cell line | Increased expression | E2F3, PLK1 | Implicated in decreased proliferation and migration after 1,25(OH)2D3 treatment | 163 |
| miR-125b | Human breast MCF7 cell line | Not investigated | VDR, CYP24A1 | Mature miR-125 levels negatively correspond to decreased VDR protein Overexpression of miR-125b leads to reduced antiproliferative effect of 1,25(OH)2D3 CYP24 protein levels in cancer tissues are inversely associated with their miR-125b levels | 315 |
| miR-125b | Melanoma MeWo and SK-Mel28 cell lines | Not investigated | VDR | VDR protein levels are inversely associated with miR125b levels | 135 |
| miR-145 | Gastric SGC-7901 and AGS cell line | Increased expression | E2F3 | Implicated in the antiproliferative effect of 1,25(OH)2D3 | 75 |
| miR-181a miR-181b | Human myeloid leukemia HL60 and U937 cells | Reduced expression | p27KIP1 | Regulation of miR-181a is implicated in the differentiation inducing effect of 1,25(OH)2D3 | 476 |
| miR-302c miR-520c | Hematological tumor Kasumi-1 and breast cancer MDA-MB-231 cell line | Reduced expression | MICA/B and ULBP2 (NKG2D ligands) | Implicated in the immuno-attack of NK cells against malignant cells | 313 |
| miR-498 | Various ovarian, endometrium, and breast cancer cell lines | Increased expression | TERT | Implicated in decreased cell proliferation after 1,25(OH)2D3 treatment | 224 |
| miR-627 | Colorectal HT29 cell line | Increased expression | JMJD1A | Increased methylation of histone H3K9, Suppressed expression of proliferative factors | 346 |
3. In vivo anticancer effects of 1,25(OH)2D3 in animals models
a) vitamin d or vdr-deficient mouse models. Different lines of evidence indicate that an impaired vitamin D signaling, either due to absence of ligand or Vdr ablation, is correlated with decreased tumor growth.
I) Vitamin D deficiency. Several studies reported that decreased circulating serum levels of 25(OH)D in mice on a vitamin D-deficient diet, are associated with an increased tumor growth in mice that were inoculated with cancer cells (343, 376, 440). In parallel, a Western diet, low in calcium and vitamin D but high in fat, promotes colonic tumor formation (502).
II) Vdr deficiency. Several studies in Vdr−/− mice pointed out that the absence of the VDR leads to enhanced proliferation and higher susceptibility to carcinogenesis, both in genetic models as upon exposure to carcinogenic products (526). Interestingly, two different research groups investigated the effect of genetic inactivation of Vdr in Apcmin/+ mice. Both groups reported that the number of small intestinal and colonic tumors is not different between Vdr+/+Apcmin/+ and Vdr−/−Apcmin/+ mice, which suggests that not tumor formation but rather tumor growth is inhibited by 1,25(OH)2D3. However, the size of the tumors is bigger in the absence of a functional Vdr (247, 519). Tumors from Vdr−/−Apcmin/+ mice express higher levels of β-catenin/TCF target genes, indicating that upon Vdr deficiency 1,25(OH)2D3 is no longer able to relocate β-catenin from the nucleus to the plasma membrane and to suppress transcription of β-catenin/TCF target genes (247).
b) chemopreventive or therapeutic effects of vitamin d, 1,25(oh)2d3, or its analogs. I) Vitamin D supplementation. In preclinical cancer models, it was demonstrated that elevating serum 25(OH)D levels is capable to reduce tumor growth. Indeed, when rodents on a Western diet were supplemented with sufficient amounts of calcium and vitamin D3, colonic tumor formation was repressed (327). Supplementing rodent chow with 5,000 IU vitamin D3/kg is as potent as treatment with 1,25(OH)2D3 in inhibiting tumor growth in mouse xenograft models of prostate and breast cancer (433). A recent study in intestine-specific Apc mutant mice demonstrated that vitamin D3 supplementation (1,500 IU vitamin D3/kg diet) but also UV-B irradiation are able to repress outgrowth and malignant progression of primary intestinal tumors (377).
II) Therapeutic effects of 1,25(OH)2D3 or its analogs. In most rodent models for various cancer types, including but not limited to breast, colon, and prostate cancer, the use of 1,25(OH)2D3 has been proven to reduce tumor growth and metastasis. However, the adverse calcemic effects of 1,25(OH)2D3 hamper its clinical applicability, and different analogs of 1,25(OH)2D3 showed clear antitumor activity without inducing hypercalcemia (354). Several studies reported that treatment with 1,25(OH)2D3 or its analogs ameliorates the response to chemotherapy (277, 312). Recently, it also became clear that 1,25(OH)2D3 could be used as a cryosensitizing agent (392). The avenue of combined vitamin D therapy is interesting since lower doses of chemo- or cryotherapy could be given resulting in fewer side effects without losing the efficacy of the treatment.
4. Human studies
a) observational studies on vitamin d and cancer. Since the initial observation of an inverse association between sunlight exposure and colorectal cancer mortality (158), multiple studies have investigated the association between vitamin D and cancer risk. As a person's vitamin D status depends both on sunlight exposure and vitamin D intake, studies on the association between circulating 25(OH)D levels may be more informative. Indeed, synthesis of 25(OH)D is not under the strict control of calciotrophic hormones, and 25(OH)D is a stable molecule with a half-life of 3 wk. In this review, we have focused on recent meta-analyses of prospective studies where 25(OH)D in serum was measured at the initiation of the study, thus before disease onset. This is in contrast to postdiagnostic measurements, which may be prone to inverse causality. Lower postdiagnostic 25(OH)D3 levels may not be the cause but rather the consequence of the disease due to disease-related factors such as lower sun exposure and physical activity, lower food intake, or systemic effects of the cancer itself (26, 175, 176, 182, 244).
Two independent meta-analyses of prospective studies on the association between serum 25(OH)D and colorectal cancer risk identified a 30–40% decreased relative risk in patients with high circulating 25(OH)D levels, compared with those with the lowest serum concentrations (252, 278). In a meta-analysis on prediagnostic 25(OH)D and breast cancer risk, Bauer et al. (35) included nine prospective studies. Their analysis suggests no relationship between circulating 25(OH)D levels and breast cancer risk, whereas in postmenopausal women, a stepwise inverse correlation is observed in a narrow range of 25(OH)D concentrations between 27 and 35 ng/ml. Above the 35 ng/ml threshold, the association flattens with no additional benefit of higher serum 25(OH)D concentrations (35). Another recent meta-analysis on 14 prospective studies identified an overall significant inverse association between serum 25(OH)D levels and breast cancer risk. Whereas there is a protective effect of 25(OH)D levels in Americans, no statistically significant associations are observed in European studies, suggesting that ethnic differences, living environment, and dietary habits may influence the association (472). In analogy with the meta-analysis of Bauer et al. (35), an inversely statistically significant association is observed in postmenopausal women, but not in premenopausal women (472). For other cancers, such as for prostate, non-melanoma skin cancer, and bladder cancer, meta-analyses did not reveal a significant association between circulating 25(OH)D and relative cancer risk (65, 317, 498). Several recent meta-analyses investigated the impact of circulating 25(OH)D levels (at or near the time of diagnosis cancer) on patient outcome. Li et al. (256) analyzed 25 studies and found a significant inverse association between 25(OH)D levels and overall survival of colorectal and breast cancer patients and patients with lymphoma. The authors concluded that a 4 ng/ml increment in circulating 25(OH)D levels confers a hazard ratio of 0.96 for overall survival of the cancer patients (256). Two additional meta-analyses concentrated on the relationship between prediagnostic circulating 25(OH)D concentrations and all-cause and cancer-related mortality. Schottker et al. (398) performed a meta-analysis from eight prospective studies and found that lower 25(OH)D levels are inversely associated with all-cause mortality, but with respect to cancer-related mortality, an association is only observed in subjects with a history of cancer. Chowdhury et al. (81) performed a meta-analysis of primary prevention observational studies and randomized controlled trials. Analysis of the observational studies (73 cohort studies) indicates an inverse association of circulating 25(OH)D with risks of death due to cardiovascular disease, cancer, and other causes. In conclusion, the inverse association between 25(OH)D levels and cancer risk is most consistent for colorectal cancer, while for other cancers contradicting results have been described. However, several recent meta-analyses support the inverse association between circulating 25(OH)D concentrations and (cancer-related) mortality, suggesting that the effects of vitamin D may be stronger for mortality than for cancer risk.
b) randomized controlled trial of vitamin d supplements and cancer risk. More direct evidence to unequivocally assess the association between vitamin D and cancer risk is to be expected from randomized controlled trials (RCTs) with vitamin D supplements. Yet the existing RCTs, in which the effects of vitamin D are studied, have limited power, and their primary endpoint was not cancer incidence or mortality (48, 229). Therefore, awaiting results from ongoing RCTs, meta-analyses of RCTs on vitamin D (vitamin D2, vitamin D3, or calcitriol) supplementation and cancer incidence or mortality may be more informative than the results derived from individual trials. Interestingly, most of these meta-analyses agree on the finding that there is no association with vitamin D supplementation and cancer incidence but a trend for an inverse association between vitamin D supplementation and overall mortality (48, 49, 90, 229). The most elaborate meta-analyses were performed by Bjelakovic et al. who investigated the effects of vitamin D supplementation on mortality (48) and on prevention of cancer in adults (49). In their study, data from 18 RCTs with 50,623 participants were analyzed. Most of the trials included mainly elderly community-dwelling women. Vitamin D supplementation was given for a weighted mean of 6 yr. Most trials studied vitamin D3 supplementation, one trial D2 supplementation, and three trials investigated supplementation with calcitriol. The authors concluded that there is no evidence for an association between vitamin D supplementation (in whatever form) and cancer incidence. No substantial differences in the effect of vitamin D and cancer are found in a subgroup analysis of trials including participants with vitamin D levels below 20 ng/ml at entry compared with trials in which participants had levels above 20 ng/ml at entry. However, there is a trend towards a protective effect of vitamin D3 supplementation on cancer mortality and of vitamin D supplementation on overall mortality. The latter conclusions have to be taken with caution as sequential trial analysis reveals that this finding could be due to random errors (48, 49). The findings of this study are in agreement with those of the systematic review on evidence for the use of multivitamins or single nutrients for the United States Preventive Services Task Force. Indeed, in this study, no evidence is found for benefit from vitamin D supplementation for the prevention and cancer and cardiovascular disease (151). Keum and Giovannucci (229) also performed a recent analysis on vitamin D supplementation and cancer incidence and mortality, but they limited their meta-analysis to RCTs of 2–7 yr of duration, involving moderate doses of supplemental vitamin D (400-1,100 IU per day), and for reasonable numbers for total cancer incidence and mortality (n = 4). Also this analysis failed to provide evidence for an effect of vitamin D supplementation on total cancer incidence. However, the authors propose a significant benefit from vitamin D supplementation on cancer mortality (229). There are some considerations that have to be taken into account when interpreting the results of these meta-analyses. As outlined above, only a few trials had cancer occurrence as a primary end-point. Furthermore, the duration of supplementation and follow-up is rather short in some studies, certainly when taking into account that carcinogenesis is a long process (48). Meanwhile, other RCTs are ongoing among which the large VITAL trial in the United States, that assesses the effect of vitamin D supplementation (2,000 IU/day), whether or not in combination with omega-3 fatty acids (1 g/day), on the risk of developing cancer as a primary end-point in more than 20,000 participants (285). All participants are enrolled and the trial is anticipated to be finished in June 2016. In Europe, a similar but smaller RCT (DO-Health, same compounds, same dosages), is running; however, in this trial cancer is not included as a primary end-point. Analyses of these trials may provide information on the dosage, the form, and the duration of vitamin D supplementation that may or may not result in beneficial extraskeletal effects.
5. Conclusion
There is ample evidence for the in vitro antineoplastic activities of the active form of vitamin D3 as numerous studies have reported beneficial effects of 1,25(OH)2D3 on reducing the malignant behavior of cancer cells. However, upon analysis of the preclinical in vivo models of cancer, most studies seem to agree that vitamin D signaling mainly affects cancer progression rather than carcinogenesis itself. In line with this, most up to date meta-analyses of observational studies and randomized clinical trials argue for a positive effect of vitamin D status on (cancer-related) mortality but not on cancer incidence.
B. Cardiovascular System
1. Vitamin D and cardiovascular disease: associations
Interest in the role of vitamin D in cardiovascular disease (CVD) came from animal studies, but also from epidemiological studies reporting the increase in cardiovascular events in winter and at increasing distance from the equator (147, 321, 400). In general, increasing observational evidence supports an association between low 25(OH)D3 levels and cardiovascular disease, although not all observational studies find significant associations, and these relationships may be prone to confounding factors. In addition, RCTs on vitamin D supplementation show inconsistent effects in relation to cardiovascular events. We will review major findings with relation to cardiovascular disease in general and to hypertension in particular, but we will start with discussing potential mechanisms revealed by animal studies.
2. Mechanisms: in vitro data and mouse data
Data from systemic and tissue-specific Vdr null mice provide insight into the role of vitamin D signaling in the cardiovascular system. Indeed, Vdr or Cyp27B1 null mice have increased levels of renin and consequently of angiotensin II, resulting in hypertension and cardiac hypertrophy (260, 522). The renin-angiotensin system is an important endocrine system controlling vascular tone, peripheral vascular resistance, and volume homeostasis. However, serum PTH levels were still increased in normocalcemic Vdr null mice, hindering interpretation of the data as PTH has been shown to increase serum renin levels (62). On the other hand, aged normocalcemic Vdr null mice fed with a rescue diet develop endothelial dysfunction, increased arterial stiffness, increased aortic impedance, and impaired systolic and diastolic function. Mechanistically, the lack of Vdr signaling results in chronically lower bioavailability of the vasodilator nitric oxide (NO) due to reduced expression of NO synthesizing enzyme, and these effects are independent of changes in the renin-angiotensin system (19). In accordance herewith, deletion of the Vdr specifically in endothelial cells results in endothelial dysfunction evidenced by impaired blood vessel relaxation, an effect that was associated with reduced endothelial NO synthase expression (329). Moreover, mice with cardiomyocyte-selective deletion of Vdr also develop cardiac hypertrophy, independent of changes in the renin-angiotensin system and thus indicating a direct in vivo antihypertrophic effect of 1,25(OH)2D3 (78). Finally, Vdr null mice display a prothrombotic state that was associated with a decrease in antithrombin and thrombomodulin (7). The therapeutic potential of 1,25(OH)2D3 was tested in rat models that develop chronic hypertension and cardiac hypertrophy and progress to congestive heart failure. Treatment with 1,25(OH)2D3 reduced cardiac hypertrophy especially in rats fed a high-salt diet (283). Together, these findings indicate that VDR signaling has effects on several aspects of the cardiovascular system and may promote endothelial and cardiac function, although the functional significance in normal physiology requires further investigation.
3. Human studies
a) vitamin d levels and endpoints of cardiovascular disease. I) Observational studies. Several prospective observational studies investigated 25(OH)D3 levels and the risk of CVD using as endpoints myocardial infarction (MI), combined CVD, stroke, and CV mortality. Most studies observed that low 25(OH)D3 levels were associated with a high risk of CVD, although this relationship was not always significant. The Framingham Offspring Study followed 1,739 white participants free of CVD at baseline. Over an average follow-up of 5 years, the risk of cardiovascular events was 1.62 times higher in those with 25(OH)D3 levels <15 ng/ml versus the remainder (95% CI = 1.11–2.36; P = 0.01). Noteworthy, the significant increased risk of CVD with vitamin D deficiency was noted in hypertensive subjects but not in those without hypertension (474). The Health Professionals Follow-up Study with a nested case-control study design evaluated more than 18,000 men and observed that the incidence of acute myocardial infarction was 2.42 times higher in men with 25(OH)D3 levels <15 ng/ml compared with those with levels above 30 ng/ml (95% CI = 1.53–3.84) (164). An 8-year follow-up cohort study of more than 3,000 patients undergoing coronary angiography noted that subjects with low 25(OH)D3 levels (<8 ng/ml) had significantly higher CV mortality compared with patients with higher levels (>28 ng/ml) (120). On the other hand, the NHANES III study, which included data from more than 13,300 participants followed for 8.7 years, showed only a trend toward an increased risk in the lowest (<17.8 ng/ml) compared with the highest 25(OH)D3 levels (305). A 10-year follow-up study of 755 patients noted no significant association of serum 25(OH)D3 levels with MI incidence, but an association with stroke was observed (288). In a prospective cohort study, as subset of the MrOS study, 813 men were followed for 4.4 years. No significant association was found between 25(OH)D3 deficiency (<15 ng/ml) and CVD incidence (coronary heart disease and cerebrovascular attack) compared with vitamin D sufficiency (> 30 ng/ml) (HR = 1.34; 95% CI = 0.65–2.77) (306). A recent meta-analysis of prospective observational studies revealed an inverse relationship between levels of 25(OH)D3 and risk of CVD, including coronary heart disease, stroke, and total CVD mortality (473). These increased risks for coronary heart disease and ischemic stroke or cerebrovascular disease for individuals with the lowest 25(OH)D3 levels were also observed in other meta-analyses (60, 61, 82).
Several studies evaluated not only changes in risk with low serum 25(OH)D3 levels, but also the contribution of higher levels. Most studies suggest that risk does not continue to decrease with levels >30 ng/ml as shown by the Framingham Osteoporosis Study and the NHANES study (228, 305, 474). Some studies even suggested a possible U-shaped relation, with a slight increase in CVD risk at high 25(OH)D3 levels (>60 ng/ml) (125, 305, 474). The IOM report concluded from the observational data that evidence is provided for an association between low 25(OH)D3 levels and greater risk of CVD but that evidence is limited to support the view that higher levels of 25(OH)D3 are linked with a further decrease of risk (Institute of Medicine 2011).
II) Limitations of observational studies. Observational data are potentially subject to residual confounding. Many disease states will reduce activity and thus sun exposure, leading to reverse causality. In addition, several risk factors for CVD are known to lead to low 25(OH)D3 levels, including age, obesity, smoking, and physical inactivity (291). It is therefore important to investigate the independent effect of vitamin D from these other risk factors.
III) Randomized control trials. The number of RCTs investigating only vitamin D versus placebo with respect to CVD is limited, as often vitamin D supplementation is combined with calcium. In addition, no large RCTs have been published that were designed specifically to test the effect of vitamin D supplementation on cardiovascular events. Indeed, most randomized vitamin D therapy trials to date were designed to investigate its protective skeletal effects, and because of the older age group, many subjects had established cardiovascular disease or risk factors.
One RCT used 100,000 IU four times per year, and they found a nonsignificant trend toward a reduction in CV deaths (RR = 0.84; 95% CI = 0.055–1.10) (452). Another trial added vitamin D to ongoing calcium supplementation and observed a decrease in ischemic heart disease event rates (371). In the large WHI trial, 36,000 women received vitamin D and calcium supplements and were followed for 7 years. No significant effect was reported on MI, coronary heart disease death, or stroke (198). Of note, a low dose of vitamin D (400 IU) was used in this large WHI trial, which may account for the lack of relationship. A recent meta-analysis reported on the few eligible studies and found no significant effect of vitamin D on MI or stroke (131). Together, the evidence from RCTs is currently insufficient to define a relationship between vitamin D and decrease in CV events and to support recommending vitamin D supplementation for lowering CVD risk (131).
b) vitamin d and hypertension. I) Observational studies. The most convincing evidence for the involvement of vitamin D metabolism in CVD is obtained in studies on hypertension. Several cross-sectional studies reported an inverse association between levels of 25(OH)D3 and the risk of hypertension (218, 291, 401). Also numerous prospective studies have shown an inverse relationship between 25(OH)D3 levels and blood pressure (BP). A recent meta-analysis reported that 25(OH)D3 levels were inversely associated with hypertension and with a significant dose-response effect: every 16 ng/ml increase in serum 25(OH)D3 was associated with a 16% decreased risk of hypertension (64). Another recent large meta-analysis included more than 283,000 participants with a mean follow-up of 9 years (242) and reported a significant inverse association of 25(OH)D3 levels and risk of incident hypertension (RR = 0.70; 95% CI = 0.57–0.86) when comparing the highest with the lowest tertile of baseline 25(OH)D3 levels.
II) Randomized control trials. RCTs of vitamin D supplementation offer the highest clinical evidence for establishing whether vitamin D deficiency is causally related to high BP. Results of RCTs have been conflicting and remain inconclusive, with some studies, but not others, suggesting a positive effect of vitamin D supplementation (148, 248, 286, 359, 367, 492). A meta-analysis from 2009 demonstrated a modest but significant decrease in BP with vitamin D treatment especially in patients with elevated mean BP at baseline (492), whereas another meta-analysis did not find a significant effect of vitamin D supplementation on BP (495). More recently, a meta-analysis reported that vitamin D supplementation resulted in a nonsignificant reduction in systolic and diastolic BP (243). A significant decrease in diastolic BP was however observed in participants with preexisting cardiometabolic disease. A recent meta-analysis on a large number of studies found no evidence of vitamin D supplementation on BP reduction (40). Also, two recent RCT in hypertensive patients with low 25(OH)D3 levels did not find a significant effect of vitamin D supplementation on BP (24, 365). Together, these findings do not support the use of vitamin D as treatment for hypertension.
c) mechanistic insight from human studies. Some of the mechanistic findings in mouse models were also observed in humans. Several studies reported an inverse association between 25(OH)D3 and parameters of endothelial dysfunction (11, 206, 434) and arterial stiffness (161). Vitamin D deficiency may increase CVD risk by activating an inflammatory cascade, which results in endothelial dysfunction and increased arterial stiffness, both of which contribute to high BP and are risk markers for CVD (160). A small RCT reported that vitamin D supplementation improved arterial stiffness (122). In other observational studies, renin activity and hypertension have been found to be inversely associated with 25(OH)D3 levels (64, 149, 366, 450). Another possible mechanism for this association of low vitamin D and high BP is that vitamin D deficiency leads to high PTH levels that are known to be linked with myocardial hypertrophy and higher BP levels (415).
4. Conclusions and future directions
Observational evidence supports an association between low 25(OH)D3 levels and both cardiovascular risk factors and cardiovascular events. However, the major problem with epidemiological studies is that 25(OH)D status may just be a surrogate for sociodemographic risk factors and poor metabolic health, thereby confounding any observational associations with CVD. The low 25(OH)D3 levels may also be a result of cardiovascular disorders rather than the cause of the disease, as sunlight exposure is the major source of vitamin D. In addition, existing trial data show inconsistent effects on cardiovascular risk factors, and the current evidence therefore does not support routine supplementation with vitamin D to reduce cardiovascular risk at the population level.
The inconsistent results reported by clinical trials have been attributed to several reasons. These include limited sample sizes to detect incremental differences in BP, heterogeneity in study populations, short follow-up periods, and the fact that the majority of trials reported results from post hoc subgroup analyses. Useful insights are therefore expected from the ongoing VITamin D and OmegA-3 TriaL (VITAL), with over 20,000 healthy participants randomized to daily dietary supplements of vitamin D3 or omega-3 fatty acids during 5 years (285).
C. Immune System
1. Mechanism of 1,25(OH)2D3 action in the immune system
Vitamin D as an immune system regulator was suggested in early studies with the discovery of the presence of VDR in activated T cells (41). More recent studies have shown that 1,25(OH)2D3 regulates both adaptive and innate immunity but in opposite directions. 1,25(OH)2D3 inhibits the adaptive immune response and promotes the innate immune response [for reviews, see Chun et al. (88), Cantorna (68), and Baeke et al. (28)]. The immunosuppressive effect of 1,25(OH)2D3 is correlated with a decrease in inflammatory cytokines including IL-2 and interferon (IFN)-γ (255). The suppressive effect of 1,25(OH)2D3 on IL-2 transcription involves blockage of the formation of the NFAT/AP-1 complex by VDR/RXR and association of VDR/RXR with the NFAT element in the IL-2 promoter as well as sequestration of Runx1 by VDR (16, 217). It has been suggested that the repressive effect of 1,25(OH)2D3 on IFN-γ transcription is due to direct binding of VDR/RXR to a silencer region in the hIFN-γ promoter (92). 1,25(OH)2D3 also stimulates the production of IL-4 by Th2 cells (280). 1,25(OH)2D3 has also been reported to result in an enhancement of T regulatory (Treg) cells (a subset of CD4+T cells important for inhibition of inflammation) (456, 458, 462, 463) and to induce Foxp3 (a lineage specific transcription factor involved in the development and function of Treg cells) (210, 217). IL-17 and IL-9 secreting T cells (Th17 and Th9 cells; distinct from Th1, Th2, and Treg subsets) have also been shown to be 1,25(OH)2D3 targets. IL-17 and IL-9, which have been implicated in many autoimmune diseases, are repressed by 1,25(OH)2D3 (217, 357, 457). The absence of IL-10 signaling completely prevented the inhibitory effect of 1,25(OH)2D3 on Th9 cells but had no effect on 1,25(OH)2D3-mediated inhibition of Th17 cells (348). The mechanism of repression of IL-17 by 1,25(OH)2D3 involves blockage of NFAT binding to the IL-17 promoter, sequestration of Runx1 by VDR, and induction of Foxp3 (which associates with and inhibits NFAT and Runx1 function) (217). Moreover, Palmer et al. (348) demonstrated that 1,25(OH)2D3 diminishes Th17 development partially through inhibition of the transcription factor RORυt, both in the presence and absence of IL-23 signaling. The stimulatory effects of 1,25(OH)2D3 on IL-4 and IL-10 and perhaps other cytokines may be indirect, and the immune response to 1,25(OH)2D3 may be dependent on the interaction of multiple cell types and activation states. In the immune cascade, one of the main targets of 1,25(OH)2D3 is the dendritic cells. Exposure of in vitro differentiating DCs to 1,25(OH)2D3 interferes with their maturation locking the cells in a semi-mature state. The altered dendritic cells have a reduced expression of class II MHC, costimulatory molecules (CD40, CD80, CD86) and a changed IL12/IL10 ratio (145, 174, 356). The mechanism of the repression of IL-12 by 1,25(OH)2D3 has been reported to involve binding of VDR/RXR to the NF-kappaB site in the IL-12p40 promoter (104). The dendritic cells are able to alter the behavior of T lymphocytes, inducing T cell anergy and increasing apoptosis levels while shifting T cell cytokine responses from a pro-inflammatory, with T-helper(Th)1 and Th17, to a more tolerogenic one, with Th2 and T regulatory cells (145). Moreover 1,25(OH)2D3-treated mouse NOD dendritic cells exhibit an intact functional migratory capacity and successfully dampen proliferation of activated T cells in vivo (144). Recent data demonstrate that 1,25(OH)2D3 affects the phenotype and behavior of dendritic cells through its early and transcriptionally mediated reprogramming of metabolic pathways, namely, the increase of glycolysis and oxidative phosphorylation at the same time (146). We are only beginning to understand the factors involved in the regulation of the immune system by vitamin D. Although global networks regulated by VDR are beginning to be addressed in immune cells, further studies related to genome-wide, proteome, and metabolic analyses are needed to define multiple roles of vitamin D in immune function.
2. In vivo studies in mouse models of autoimmunity
The physiological significance of these effects of 1,25(OH)2D3 on the immune system has been suggested by in vivo studies in mouse models of autoimmunity. 1,25(OH)2D3 can protect against a number of experimental autoimmune diseases including inflammatory bowel disease (IBD) and experimental autoimmune encephalomyelitis (EAE; mouse model for multiple sclerosis, MS) (110). Dietary calcium has been reported to be required for the suppressive effect of 1,25(OH)2D3 on IBD and EAE (70, 110). 1,25(OH)2D3 has also been reported to reverse as well as to prevent paralysis of EAE mice (69, 217). The protective effect of 1,25(OH)2D3 in EAE is associated with inhibition of IL-12 and IL-17 and requires IL-10 signaling (217, 298, 421). Not only 1,25(OH)2D3 but also high-dose dietary vitamin D was found to attenuate EAE. The combination of high-dose dietary vitamin D (20 IU/g diet) and IFN-β was more effective than either high-dose dietary vitamin D alone or IFN-β alone in diminishing paralysis in mice with ongoing EAE (Figure 8). These findings provide a rationale for trials combining vitamin D and IFN-β in MS patients. With regard to IBD, VDR KO mice show more severe IBD which is associated with overproduced IFN-γ and IL-17 cells (68). Recently several studies demonstrated the importance of the VDR expression specifically in intestinal epithelial cells in IBD (232, 269a, 494). Treatment with 1,25(OH)2D3 of experimentally induced colitis results in inhibition of Th1 and Th17 cells, induction of Treg cells, and reduced inflammation (71, 106). Protection against autoimmune diabetes in nonobese diabetic (NOD) mice by 1,25(OH)2D3 has also been reported. Induction of Treg cells and decreased numbers of effector T cells have been suggested as the basis for this protection by 1,25(OH)2D3 (173, 294, 437). Although these findings are suggestive of a protective effect of 1,25(OH)2D3 against the pathogenesis of autoimmune inflammation, whether vitamin D supplementation or treatment with analogs of 1,25(OH)2D3 is beneficial clinically in the treatment of autoimmune diseases is not known. Adequately powered, RCTs are needed to demonstrate the suggested benefit of vitamin D.
Figure 8.

Cooperative effects of dietary vitamin D and IFN-β treatments in attenuation of EAE. On day 0, EAE was induced by immunization with myelin oligodendrocyte glycoprotein (MOG) p35-55 in C57BL/6 mice and scored daily for degree of paralysis. On day 5, the diet was changed to 20 IU vitamin D/g diet or left at 1.5 IU vitamin D/g diet. On day 7, treatment with IFN-β (a first line treatment for multiple sclerosis) was initiated and continued every other day through day 16. The combination of high-dose vitamin D and IFN-β was more effective than high dietary vitamin D alone or IFN-β alone in diminishing paralysis in EAE (R. Axtell, L. Steinman, and S. Christakos, unpublished data).
3. Effects of vitamin D on innate immunity
With regard to effects on innate immunity, in vitro studies have shown that 1,25(OH)2D3 induces the antimicrobial peptide cathelicidin in both myeloid and epithelial cells with the subsequent killing of bacteria (170, 268, 486, 504). Despite significant in vitro evidence showing a negative effect of 1,25(OH)2D3 on bacterial growth, relatively few studies have examined the effect of 1,25(OH)2D3 in vivo on host resistance against bacteria. The studies in experimental animal models of infection have not supported a consistent beneficial or adverse effect of vitamin D. Vitamin D-deficient mice are more susceptible to Mycobacterium bovis infection than WT mice due to an effect on NO production (482). Using Vdr KO mice, it was shown that VDR is not required for the clearance of L. monocytogenes following either primary or secondary infection (63). 1,25(OH)2D3 treatment was reported to impair host defense against colitis induced by Citobacter rodentium (388). 1,25(OH)2D3 treatment of infected mice resulted in increased pathogen burdens, exaggerated tissue pathology, and significantly reduced numbers of Th17 cells. Although IL-17 plays a pathological role in inflammatory diseases, this cytokine also plays a protective role against infection. IL-17 can enhance host defense by induction of antimicrobial peptides and by induction of chemokines to recruit neutrophils (222, 230, 503) It was suggested that 1,25(OH)2D3 by suppressing Th17 T cell responses in vivo resulted in impaired host defense against Citrobacter rodentium (388). Thus 1,25(OH)2D3 treatment can be a double-edged sword. Further in vivo data are needed to support a relationship between vitamin D and host resistance to infection.
VI. VITAMIN D ANALOGS
A. Analog Development: Introduction
Since the discovery that supraphysiological doses of 1,25(OH)2D3 (calcitriol) can stimulate the differentiation of promeylocytic leukemia cells towards mature macrophages (3) and can inhibit the proliferation of melanoma cells (95), academic and nonacademic institutions started to develop analogs of 1,25(OH)2D3. 1,25(OH)2D3 is a very flexible molecule that can be structurally modified to yield analogs with altered biological properties. Chemical modifications (Figure 9) were introduced in the A-ring, seco-B ring, central CD-region, or side chain of 1,25(OH)2D3 with the objective to minimize the calcemic side effects and keep or increase the antiproliferative or prodifferentiating effects of 1,25(OH)2D3 (53). Until the end of the 20st century, this new field of vitamin D analog research was based on a “trial and error” approach. Once the biological profile of some analogs was characterized, it became possible to predict the biological implications of some, but not all, chemical modifications (“educated guess”). Several modifications such as 19-nor, 16-ene, 23-yne, and 20-epi were shown to diminish the calcemic effects and/or enhance the antipoliferative effects of the mother compound. In 2000, the group of Moras and Rochel (384) was able to crystallize the VDR (see sect. IIIA1), and this VDR crystal model led to the rational design of a second generation of vitamin D analogs (carbon-2 substituted analogs; gemini analogs). In this overview we will first focus on analogs that are used for disorders linked to the classical effects of 1,25(OH)2D3 to regulate calcium and bone homeostasis. A second group of analogs are characterized by a dissociation of the calcemic effects and the nonclassical effects of 1,25(OH)2D3 on proliferation and differentiation. These analogs have the eligible profile for the treatment of hyperproliferative disorders such as psoriasis and cancer. Next we discuss the possible mechanism of action of these compounds.
Figure 9.

Chemical structure of 1,25(OH)2D3 and possible analog modifications.
B. Analogs Related to the Classical Function of 1,25(OH)2D3
1. Kidney disease
Therapy with 1,25(OH)2D3 in patients with CKD can lead to an increase in calcium and phosphorus absorption with hypercalcemia and hyperphosphatemia as a result (103). The existence of vitamin D analogs, which selectively decrease PTH levels without a major rise of serum calcium levels, opened new perspectives for the commercialization of these compounds for chronic renal failure (Figure 10). Paricalcitol and doxercalciferol are vitamin D2 analogs approved for therapeutic use of SHPT. In Japan, falecalcitriol and maxacalcitol are used to treat this disorder. Paricalcitol, maxacalcitol, and falecalcitriol bind directly to the VDR, whereas doxercalciferol needs analogous to alfacalcidol a 25-hydroxylation step in the liver to become active and is independent of renal or extrarenal 1α-hydroxylase.
Figure 10.

Overview of clinically approved vitamin D analogs.
Seventy-eight patients with end-stage renal disease (ESRD) and receiving hemodialysis were studied in three identical double-blind, placebo controlled, randomized, multicenter trials to evaluate the safety and efficacy of paricalcitol (290). Patients under calcitriol treatment received a 2-wk washout period before weekly measurements of the baseline biochemical parameters of intact PTH, serum calcium, and phosphorus for 2 wk. Biochemical parameters were also measured in patients without calcitriol therapy. After the 2-wk baseline period, patients were randomized in a placebo or paricalcitol (intravenous) treatment group during 12 wk. The dose of paricalcitol was increased as long as PTH did not decrease for more than 30%, serum calcium levels did not increase for more than 11.5 mg/dl, and calcium-phosphorus product was less than 75. PTH levels decreased from 785 ± 66 to 370 ± 73 pg/ml during the study of 12 wk in the paricalcitol group, whereas PTH levels did not change in the placebo group. The mean serum calcium levels increased significantly but remained in the normal range at the end of the study (9.56 ± 0.15 mg/dl), and serum phosphorus levels did not change significantly. There were some episodes of serum calcium levels ≥11 mg/dl but very few in number. Limited RCTs are available in which paricalcitol or other vitamin D analogs are compared with calcitriol treatment. Studies in which the dose of paricalcitol was evaluated suggested that for the same suppressive effects, paricalcitol should be dosed at a ratio of 4:1 to calcitriol (289), although other studies indicated a 3:1 ratio (271). A double-blind randomized multicenter study (422) compared the effectiveness of paricalcitol and calcitriol, both injected intravenously in ESRD patients undergoing hemodialysis. Patients with paricalcitol therapy showed a significantly faster ≥50% reduction from baseline PTH than did the calcitriol-treated patients, and only the paricalcitol-treated patients had PTH levels in the desired range between 100 and 300 mg/ml. Paricalcitol treatment induced less episodes of hypercalcemia and increased calcium-phosphorus product compared with calcitriol therapy. Also in chronic hemodialysis patients with SHPT paricalcitol intravenous therapy seemed to be superior compared with calcitriol therapy in a single-center randomized open label study (2). Moreover, paricalcitol has a small effect on intestinal calcium absorption compared with calcitriol in hemodialysis patients (273).
Doxercalciferol is mainly used for the treatment of moderate to advanced CKD (353). Intermittent intravenous or oral doxercalciferol therapy in hemodialysis patients with SHPT (152, 301) decreased PTH levels. The study consisted of a washout period (8 wk), an open-label treatment with doxercalciferol (16 wk), and a randomized, double-blinded treatment with doxercalciferol or placebo (8 wk). Baseline PTH levels (897 ± 52 pg/ml) decreased by 20 ± 3.4% by week 1 and by 55 ± 2.9% at week 16. PTH levels returned to baseline during placebo treatment but remained suppressed with doxercalciferol therapy. During open-label treatment, serum calcium levels were 9.2 ± 0.84 to 9.7 ± 1.05 mg/dl and phosphorus levels were 5.4 ± 1.10 to 5.9 ± 1.55 mg/dl. During double-blinded treatment, serum calcium levels were slightly increased with doxercalciferol compared with the placebo group, but phosphorus levels did not differ. A RCT with doxercalciferol in patients with SHPT associated with CKD stages 3 and 4 (94) demonstrated a decrease of ≥30% in PTH levels in 74% of the treated patients after 24 wk of oral therapy compared with 7% of the patients in the placebo group. No significant differences were observed between both groups in mean serum calcium or phosphorus levels or incidence of hypercalcemia or hyperphosphatemia. A prospective clinical trial was done in South Asian patients with CKD stage 4 and confirmed that doxercalciferol is effective in suppressing SHPT with an acceptable risk of hypercalcemia and hyperphosphatemia (116). A prospective study in 60 pediatric patients with CKD on peritoneal dialysis and doxercalciferol or calcitriol therapy for 8 mo (in combination with either calcium, carbonate, or sevelamer) demonstrated the same potency of both agents to reduce PTH levels, bone turnover, and FGF23 levels (485). Limited studies compared the efficacy of doxercalciferol with paricalcitol. High-dose paricalcitol (160 μg) was compared with high-dose doxercalciferol (120 μg) in a prospective study of 13 hemodialysis patients (212), and the degree of PTH suppression was similar in both groups; however, the serum phosphorus levels increased faster and higher in the doxercalciferol-treated patients compared with the paricalcitol treated patients. A dose equivalency study was performed on chronic hemodialysis patients treated with a stable dose of paricalcitol for at least 3 mo and were randomized to be treated with a dose of doxercalciferol equivalent to either 35, 50, or 65% of the paricalcitol dose for 6 wk. Serum PTH, calcium, phosphorus, and albumin were determined at baseline and monitored every 2 wk. A linear regression analysis of percent change in PTH values by dose group was performed to determine the conversion factor (527). In patients on a maintenance dose of paricalcitol, dosing doxercalciferol at 55–60% of the paricalcitol dose resulted in comparable inhibition of PTH. Paricalcitol and doxercalciferol are used in the United States to treat SHPT, whereas falecalcitriol and maxacalcitol are applied in Japan. Comparative studies with maxacalcitol and calcitriol are equally potent in reducing PTH levels and are equally safe (339, 438). Maxacalcitol (intravenous) and paricalcitol (intravenous) were recently tested for 12 wk in a double-blind controlled study in 255 Japanese CKD patients with SHPT on hemodialysis. Both intravenous paricalcitol (27.7%) and maxacalcitol (30.5%) were equally effective in reducing PTH without the presence of hypercalcemia during treatment (10).
The treatment of CKD with vitamin D derivatives (calcitriol etc.) or vitamin D analogs has primarily focused on the outcome of biochemical parameters [PTH, calcium, phosphorus, 25(OH)D3, or 1,25(OH)2D3 levels] (231) but also intermediate (vascular calcifications, bone density, and histology) or hard clinical end points (cardiovascular disease, mortality, fractures. . .) should be investigated in prospective RCTs. There have been several observational studies which demonstrated a survival benefit in CKD patients on dialysis (314); however, a recent meta-analysis of RCTs evaluating the effects of oral vitamin D supplementation (cholecalciferol, doxercalciferol, paricalcitol, alfacalcidol) compared with placebo on overall all-cause mortality (RR = 0.84; 95% CI = 0.47, 1.52), cardiovascular mortality (RR = 0.79; 95% CI = 0.26, 2.28) and serious adverse cardiovascular events (RR = 1.20; 95% CI = 0.49, 2.99) in CKD patients did not show convincing evidence (284). The studies included in this meta-analysis showed a large variation in the doses used and the treatment period. A meta-analysis of RCTs investigating the effect of vitamin D therapy in nondialysis CKD patients on proteinuria and the progression of CKD together with adverse events such as hypercalcemia (497) showed that this treatment decreased proteinuria and had no negative influence on renal function. However, there was an increased risk of developing hypercalcemia compared with the control patients. Moreover, there was no superiority found for newer vitamin D analogs (paricalcitol, doxercalciferol, maxacalcitol, falecalcitriol) versus established vitamin D compounds (ergocalciferol, cholecalciferol, 25D3/calcidiol, calcitriol). The VITAL double-blind, placebo-controlled randomized-controlled study (98) included patients with CKD stages 2–4 and demonstrated in a post hoc analysis that paricalcitol dose-dependently reduced PTH levels but with a modest increase in calcium and phosphate. Moreover, paricalcitol reduced bone-specific alkaline phosphatase which may be beneficial for inhibiting vascular calcification. In the PRIMO trial, 227 CKD stage 3 and 4 patients with mild left ventricular hypertrophy but with normal systolic function were randomized to paricalcitol or placebo treatment (443). The primary end point was a change in ventricular mass over 48 wk, but the paricalcitol treatment was not significantly different from the placebo group, although the risk of hospitalizations was lower in the paricalcitol group (1.1 vs. 8.8 per 100 persons years). Paricalcitol was able to suppress PTH to 30% of baseline in 85.7% of patients compared with 16.5% in the placebo group, but there was an increase in serum calcium and phosphorus.
In conclusion, a lot of clinical studies with vitamin D analogs were performed, and for some of them it is not yet completely clear if these compounds are indeed superior to calcitriol to diminish hypercalcemia and hyperphosphatemia. For this reason, more elaborate RCTs should be performed with several vitamin D analogs together such as maxacalcitol, paricalcitol, doxacalciferol, and compared with calcitriol while taking into account biochemical parameters but also patient-centered end points (CKD progression, fractures, mortality, cardiovascular disease).
2. Osteoporosis
Calcium (1 g) and vitamin D (cholecalciferol, 800 IU) supplementation is given to prevent or treat osteoporosis, but also pharmacological intervention with bisphosphonates, selective estrogen receptor modulators (raloxifene), denosumab, strontium relanate, and PTH peptides are frequently used for osteoporosis treatment (221). Both alfacalcidol and calcitriol are used in some countries including Japan for the treatment of osteoporosis. Alfacalcidol is metabolized to calcitriol by its 25-hydroxylation in the liver. Both compounds have been proven to be active in increasing bone mineral density (BMD) and in reducing vertebral and nonvertebral fractures in prospective, randomized, and placebo-controlled trials (335). In a randomized, double-blind, double-dummy, parallel group study alfacalcidol was superior to vitamin D plus calcium in increasing BMD in postmenopausal osteoporosis. Lumbar BMD in the alfacalcidol arm increased 2.33% compared with baseline levels after 12 mo and 2.87% after 18 mo, whereas in the vitamin D plus calcium group there was only an increase of 0.7% at both time points. At the end of the study, serum calcium concentration was slightly higher in the alfacalcidol group versus vitamin D plus calcium (9.56 ± 0.48 vs. 9.45 ± 0.49 mg/dl) but was still within the safety margin (335). Eldecalcitol is a vitamin D analog with a hydroxypropyloxy group at the carbon 2β-position of 1,25(OH)2D3 which is approved in Japan since 2011 for the treatment of osteoporosis. In a placebo-controlled randomized-controlled trial, eldecalcitol suppressed in a dose-dependent manner bone turnover and increased lumbar spine and total hip BMD in osteoporosis patients with sufficient vitamin D supply. A lower incidence of hypercalcemia was observed with 0.75 μg/day eldecalcitol compared with 1.0 μg/day (297). In a phase III randomized, double-blind comparative study, the efficacy of eldecalcitol (0.75 μg/day) was compared with alfacalcidol (1.0 μg/day) over 3 years in osteoporotic patients receiving 400 IU vitamin D if serum 25(OH)D3 was less than 20 ng/ml (296). Eldecalcitol was more potent to increase BMD and decrease the frequency of vertebral and wrist fractures compared with alfacalcidol treatment. The incidence of hypercalcemia above 11.5 mg/dl was very low in both groups (2 in eldecalcitol and 0 in alfacalcidol group), but the increase of serum and urinary calcium was significantly higher with eldecalctiol therapy.
A vitamin D analog which possesses bone anabolic effects and could possibly be used for the treatment of osteoporosis is 2-methylene-19-nor-20(S)-1,25(OH)2D3 (2MD). In ovariectomized rats 2MD restored trabecular and cortical bone mass and strength and enhanced bone-mobilizing activity (226). A randomized, double-blind, placebo-controlled trial in postmenopausal women with osteopenia demonstrated that a daily oral treatment for 1 year with 2MD (220 or 440 ng) increased bone resorption and bone formation markers; however, it did not change bone mass (109). In this study 2MD suppressed serum PTH levels dose-dependently without elevating serum calcium levels at the lowest dose of 220 ng/day. Recently a dose-finding study with 2MD in postmenopausal women confirmed the PTH suppressive effects of 2MD without calcemic side effects (509). A distribution study performed in rats showed that radioactive 2MD was accumulated in the classical target tissues of vitamin D (bone, intestine) but was most strongly localized in the thyroid/parathyroid (509). In a 5/6 nephrectomy rat model of renal failure, 2MD suppressed PTH dose-dependently, and a side-to-side experiment with paricalcitol demonstrated that the effective dose of 2MD was 40 times lower than paricalcitol without increasing serum calcium levels (509). Currently 2MD is under evaluation in phase 2 study in dialysis patients.
C. Analogs Related to the Nonclassical Effects of 1,25(OH)2D3
1. Psoriasis
The epidermis has a pivotal position in the vitamin D system since it is the site of photosynthesis of vitamin D which can be further converted locally to 25(OH)D3 (CYP27A1 and CYP2R1 enzyme) and finally to active 1,25(OH)2D3 (CYP27B1 enzyme). Moreover, the VDR is present in the skin and 1,25(OH)2D3 can induce differentiation at 10−10 M whereas it has a dual effect on the proliferation of keratinocytes. At physiological doses (picomolar), 1,25(OH)2D3 promotes proliferation and at supraphysiological concentrations 1,25(OH)2D3 inhibits proliferation. Psoriasis is a chronic autoimmune inflammatory skin disorder, and patients have erythematous scaling plaques that are the result of keratinocyte hyperproliferation and abnormal differentiation (397). Calcipotriol, tacalcitol, and the more recent approved maxacalcitol are used either as monotherapy or in combination with topical steroids to treat psoriasis. The vitamin D analogs exert prodifferentiating and antiproliferative effects on keratinocytes and also possess important anti-inflammatory properties (418). Topical calcipotriol increased the expression of VDR on epidermal keratinocytes which enhances its potent effects on cell proliferation and differentiation (378). However, not all patients with psoriasis respond well to vitamin D analog treatment, and the responsiveness can be correlated to the increase in VDR mRNA in treated skin areas (77). Pro-inflammatory cytokines TNF-α, IFN-γ, IL-2, and IL-8 play a critical role in the T-cell-mediated inflammatory process in the psoriatic skin (397). Thirty patients with moderate plaque-type psoriasis treated with calcipotriol or vehicle showed an increase in IL-10 levels (pg/mg protein) and a decrease in IL-8 levels (pg/mg protein) in skin plaques when treated with calcipotriol compared with baseline levels at the start of the treatment (220), whereas both cytokines were unaffected by vehicle treatment. Moreover, calcipotriol suppressed the Th17 cytokine-mediated production of psoriasin and koebnerisin, two antimicrobial chemoattractant peptides that amplify psoriatic inflammation (185). The cytokine lymphopoietin which induces Th2 differentiation and inhibits IL-12/23 production was upregulated by topical calcitriol or calcipotriol treatment as well as the antimicrobial peptide cathelicidin (393). Calcipotriol has a very short half-life, and it has been proven that it is very effective and safe in the topical treatment of psoriasis (451). Maxacalcitol was more potent to diminish erythema and scaling compared with calcipotriol in a double-blind, placebo-controlled comparator study (33). Topical tacalcitol ointment was very potent to reduce the psoriasis area severity index (PASI) without calcemic side effects (461). The adverse side effect noted for all vitamin D analogs is skin irritation. Combination therapies targeting immune-mediated inflammation together with epidermal changes provided improved efficacy and safety versus monotherapy (451).
2. Cancer
More than 3,000 vitamin D analogs were developed worldwide and several analogs demonstrated more potent antiproliferative and prodifferentating effects on cancer cell lines compared with 1,25(OH)2D3. Moreover, some analogs demonstrated a dissociation between anti-cancer and calcemic side effects in several mice models of cancer (xenografts, chemically induced). In view of these promising results, a limited number of analogs have been tested in cancer patients. Seocalcitol was the first analog that has been explored in advanced breast and colon cancer (177) and inoperable pancreatic cancer (138); however, no clear anti-cancer activities were observed. A very small effect was seen with seocalcitol in inoperable hepatocellular carcinoma in which 2 of 33 patients showed a complete response (105). Some of the analogs that are approved for SHPT have also been tested on their anti-tumor capacity in urogenital neoplasms. Doxercalciferol (265) or paricalcitol (399) was mainly tested in patients with androgen-independent prostate cancer, but only doxercalcifereol could stabilize the disease in 6 of the 20 patients for over 6 mo in a phase II study (265). Doxercalciferol was also evaluated in localized prostate cancer and high-grade prostatic intraepithelial neoplasia, but no beneficial effects were noticed in serum and tissue markers (159). The anti-psoriatic vitamin D analog calcipotriol was applied topically at a dose of 100 μg/day in patients with locally advanced or cutaneous metastases from breast cancer. One study reported no response after 3 mo of treatment (338), while in the other study, three patients showed a 50% reduction in the diameter of treated lesions after 6 wk (56). The 16-ene-23-yne-1,25(OH)2D3 (ILX23-7553) analog showed in in vitro and in vivo animal models potent antitumor properties with diminished calcemic side effects (333) and was recently investigated in 16 patients with advanced solid tumors, but no objective response was observed (208). Since the monotherapeutic approach was not really successful, several clinical trials were set up to combine vitamin D analogs with established protocols in cancer therapy (chemotherapy, radiotherapy, …). The 14-epi-19-nor-23-yne-1,25(OH)2D3 analog inecalcitol was given orally to patients with hormone-refractory prostate cancer in combination with docetaxel for maximum 18 wk. This study had a response rate of 85% based on a PSA decline of at least 30% within 3 mo of treatment. This analog has now successfully been tested in a phase II study in chronic lymphocytic leukemia (CLL) at a dose of 2 mg daily after which 52% of patients experienced stabilization or decrease of blood leukemic lymphocytes counts. Based on the promising results of inecalcitol in CLL a phase II study is planned with 4 mg/day given to chronic myeloid leukemia patients under treatment with oral imatinib (website Hybrigenics). A placebo-controlled study with oral doxercalciferol (10 μg/day) in combination with weekly docetaxel for 4 wk did not enhance PSA response rate or survival (25). Oral paricalcitol was recently shown to be safe in women with metastatic breast cancer receiving taxane-based chemotherapy (249).
Vitamin D analogs possess potent anti-cancer effects in cancer cell cultures and in animal cancer models, but these effects could not be confirmed in human studies. In contrast to chemotherapeutics, vitamin D analogs are not cytotoxic and should therefore be combined with established cancer therapies.
D. Mechanism of Action
The mechanisms of action of the analogs with increased antiproliferative, prodifferentiating effects and/or decreased calcemic side effects are not yet fully understood and are probably based on a combination of several phenomena. Differences in pharmacokinetics may contribute to lower calcemic activity because most of these analogs display a low affinity for DBP and therefore their free concentration approaches their total plasma concentration which results in rapid extracellular clearance. Moreover, altered pharmacokinetics may also explain the enhanced antiproliferative effects since the analog will quickly reach high peak levels in target tissues compared with the slow rise and drop of 1,25(OH)2D3. Maxacalcitol has a short half-life due to 500 times lower affinity to DBP (341). Eldecalcitol on the contrary has a fourfold stronger affinity for DBP than 1,25(OH)2D3 (381) which results in longer sustained plasma levels than 1,25(OH)2D3. A recent study manually docked calcitriol and eldecalcitol in the crystal structure of DBP (236) and demonstrated that calcitriol binds to DBP via three hydrogen bonds, whereas eldecalcitol binds to DBP via three additional hydrogen bonds that interact with the 3-hydroxypropyloxy group.
Also, the intracellular metabolism might be different from that of 1,25(OH)2D3 due to chemical modifications (side chain fluorination, 20-epimerization) that make the analogs more resistant to catabolism by CYP24A1. Hence, the cells are exposed for a longer period to the analog (213). Eldecalcitol binds more weakly to VDR [less than half of that of 1,25(OH)2D3] but is more potent to induce mRNA expression of CYP24A1 than calcitriol. However, eldecalcitol is not metabolized by CYP24A1, which may also prolong the activity of this analog in target tissues (381). At the cellular level, the antiproliferative activity seems to correlate well with the ability of an analog to promote interaction between VDR and coactivator proteins (130). The elucidation of the crystal structure of the ligand-binding domain of VDR and the determination of the exact interaction of 1,25(OH)2D3 with amino acids in the ligand-binding pocket demonstrated an expansion of this binding pocket near the position of carbon 2 of the A-ring of the ligand (384). Vanhooke et al. (465) crystallized the ligand-binding domain of the VDR with the carbon 2 analog 2MD in complex with a LXXLL containing coactivator peptide and showed that the LBD bound to 2MD was unchanged compared with the VDR-LBD-1,25(OH)2D3 complex. On the other hand, rat VDR-LBD studied in solution by NMR spectroscopy showed different chemical shifts when bound to 2MD compared with 1,25(OH)2D3 (413). The 14-epi analog inecalcitol was cocrystallized with VDR-LBD (129) and the 17-methyl D-ring analog CD 578 with VDR-LBD together with an SRC-1 coactivator peptide with second LXXLL-motif containing NR box (128). Both studies indicated that the analogs did not induce major differences in the protein conformation upon binding with the LBD compared with binding with 1,25(OH)2D3 but induce rather subtle differences such as closer or additional contacts to certain amino acid residues allowing more stable interactions with coactivators. Approximately 10-fold lower doses of inecalcitol are needed, compared with 1,25(OH)2D3, to acquire the same amount of coactivator interactions (129). The crystal structures of VDR-LBD bound to 20-epi vitamin D analogs [KH1060, 20-epi-1,25(OH)2D3], seocalcitol, or calcipotriol also demonstrated that all compounds are anchored to the same residues in the LBD via the hydroxyl groups of the A-ring and the side chain, therefore they are locked in identical positions and form the same hydrogen bonds (447, 448). New insights were gathered by the synthesis of the Gemini analog which has two identical side chains branching at carbon 20 and is characterized by less VDR binding affinity compared with 1,25(OH)2D3 but equal or more transactivation potency (332). The VDR-Gemini complex can switch from agonist to an inverse agonist confirmation in the presence of an excess of corepressor N-Cor (172). The Gemini-VDR complex revealed that the binding of a ligand with 25% increased volume did not change the overall structure of the LBD including the position of helix H12 compared with VDR-LBD-1,25(OH)2D3 complex; however, an extra channel was opened to accommodate the second side chain (91). Several derivatives (two different side chains) of the original Gemini analog were developed to enhance its biological potency (251), for example, by decreasing CYP24A1 metabolic degradation via carbon-23 unsaturation or 26,27-fluorination. The crystal structures of such Gemini derivatives (Gemini-0072, -0097) with zebrafish zVDR-LBD and a GRIP1/TIF2-coactivator peptide containing LXXLL motif, showed that the second side chains induced an extra cavity within the LBD comparable to the original Gemini and that additional interactions of the side chain fluorine atoms stabilize helix H12 allowing increased interactions with coactivator proteins (201).
More recently, ChIP studies are performed to examine VDR binding sites throughout the genome after treatment with 1,25(OH)2D3 or analogs. Such a study compared the binding sites of the VDR in intestinal tissue after 1,25(OH)2D3 or 20-epi-1,25(OH)2D3 analog treatment and showed that both compounds induce VDR binding to CYP24A1 and TRPV6 loci in the intestine, but the analog elicited a prolonged VDR binding to these genes leading to its superagonistic characteristics such as hypercalcemia in vivo (512). In osteoblast cell models, 2MD bound to the VDR was able to bind VDREs at lower concentrations compared with 1,25(OH)2D3 (500). ChIP and Re-ChIP assays revealed that eldecalcitol mediates dissociation of Williams syndrome transcription factor (WSTF) from the aromatase CYP19A1 promoter and thereby decreases the gene expression of aromatase in MCF-7 cells (274). Microarray studies demonstrated that 1,25(OH)2D3 as well as vitamin D analogs regulate the expression of genes involved in several signaling pathways in a cell type and tissue specific manner (241). Moreover, on the same tissue, 1,25(OH)2D3 or vitamin D analogs up or downregulate the expression of the same set of genes (241).
In conclusion, several analogs are used to treat SHPT, osteoporosis, or psoriasis; however, most of the clinical trials testing vitamin D analogs for cancer treatment were not really convincing. The concept that vitamin D analogs can be used for cancer therapy should be further explored by combining analogs with standard protocols for cancer therapies as well as by examining the correct duration and timing of administration. The exact working mechanism is not completely unraveled, but new findings that vitamin D can induce epigenetic modulations should be further explored.
VII. CONCLUSIONS AND FUTURE DIRECTIONS
New target genes involved in the multiple actions of 1,25(OH)2D3 in numerous different systems will undoubtedly be identified. New insight will be obtained with regard to regulatory pathways, novel transcription factors, and epigenetic modifications involved in mediating these diverse biological responses. We will also obtain an increased understanding of the structure of VDR in the presence of protein partners thus facilitating selective modulation of 1,25(OH)2D3 action in different target tissues. It is likely that future studies will reveal that posttranscriptional mechanisms are also an important mechanism controlling the expression of vitamin D target proteins. Further studies are also needed using null and transgenic mice to increase our understanding of vitamin D biology. The main function of vitamin D is increased intestinal calcium absorption. However, we are far from understanding the mechanisms involved. To identify new therapeutic approaches to sustain calcium balance, the multiple mechanisms by which 1,25(OH)2D3 acts in both proximal and distal segments of the intestine need further definition. Although there is compelling evidence from the laboratory of extraskeletal beneficial effects of 1,25(OH)2D3, conclusive clinical data for the use of vitamin D and analogs for the treatment or prevention of a number of disease processes are not yet available. Although there are many differences between the human condition and mouse models, since many genes function similarly in animals and humans, results in animals models may unravel complex signaling pathways similarly affected in humans. These findings could lead to the identification of novel targets for chemoprevention and chemotherapy.
GRANTS
S. Christakos receives funding from National Institutes of Health Grants AG044552, AI100379, and DK38961-22. A. Verstuyf is funded by Fund for Scientific Research-Flanders (FWO) Grant G.OA17.14N and by KU Leuven Grant GOA/14/010. G. Carmeliet is funded by Grant FWO G.0573.13.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
acknowledgments
Address for reprint requests and other correspondence: S. Christakos, Dept. of Microbiology, Biochemistry, and Molecular Genetics, Rutgers, The State University of New Jersey, New Jersey Medical School, 185 South Orange Ave., Newark, NJ 07103 (e-mail: christak@njms.rutgers.edu).
REFERENCES
- 1.Autosomal dominant hypophosphataemic rickets is associated with mutations in FGF23. Nature Genet 26: 345–348, 2000. [DOI] [PubMed] [Google Scholar]
- 2.Abdul Gafor AH, Saidin R, Loo CY, Mohd R, Zainudin S, Shah SA, Norella KC. Intravenous calcitriol versus paricalcitol in haemodialysis patients with severe secondary hyperparathyroidism. Nephrology 14: 488–492, 2009. [DOI] [PubMed] [Google Scholar]
- 3.Abe E, Miyaura C, Sakagami H, Takeda M, Konno K, Yamazaki T, Yoshiki S, Suda T. Differentiation of mouse myeloid leukemia cells induced by 1 alpha,25-dihydroxyvitamin D3. Proc Natl Acad Sci USA 78: 4990–4994, 1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Adams JS, Sharma OP, Gacad MA, Singer FR. Metabolism of 25-hydroxyvitamin D3 by cultured pulmonary alveolar macrophages in sarcoidosis. J Clin Invest 72: 1856–1860, 1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Adorini L, Penna G, Amuchastegui S, Cossetti C, Aquilano F, Mariani R, Fibbi B, Morelli A, Uskokovic M, Colli E, Maggi M. Inhibition of prostate growth and inflammation by the vitamin D receptor agonist BXL-628 (elocalcitol). J Steroid Biochem Mol Biol 103: 689–693, 2007. [DOI] [PubMed] [Google Scholar]
- 6.Ahn J, Yu K, Stolzenberg-Solomon R, Simon KC, McCullough ML, Gallicchio L, Jacobs EJ, Ascherio A, Helzlsouer K, Jacobs KB, Li Q, Weinstein SJ, Purdue M, Virtamo J, Horst R, Wheeler W, Chanock S, Hunter DJ, Hayes RB, Kraft P, Albanes D. Genome-wide association study of circulating vitamin D levels. Hum Mol Genet 19: 2739–2745, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Aihara K, Azuma H, Akaike M, Ikeda Y, Yamashita M, Sudo T, Hayashi H, Yamada Y, Endoh F, Fujimura M, Yoshida T, Yamaguchi H, Hashizume S, Kato M, Yoshimura K, Yamamoto Y, Kato S, Matsumoto T. Disruption of nuclear vitamin D receptor gene causes enhanced thrombogenicity in mice. J Biol Chem 279: 35798–35802, 2004. [DOI] [PubMed] [Google Scholar]
- 8.Ajibade DV, Dhawan P, Fechner AJ, Meyer MB, Pike JW, Christakos S. Evidence for a role of prolactin in calcium homeostasis: regulation of intestinal transient receptor potential vanilloid type 6, intestinal calcium absorption, and the 25-hydroxyvitamin D(3) 1alpha hydroxylase gene by prolactin. Endocrinology 151: 2974–2984, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Akhter S, Kutuzova GD, Christakos S, DeLuca HF. Calbindin D9k is not required for 1,25-dihydroxyvitamin D3-mediated Ca2+ absorption in small intestine. Arch Biochem Biophys 460: 227–232, 2007. [DOI] [PubMed] [Google Scholar]
- 10.Akizawa T, Akiba T, Hirakata H, Kinugasa E, Tominaga Y, Fukagawa M, Yokoyama K, Zhang W, Linde PG, Suzuki M. Comparison of paricalcitol with maxacalcitol injection in Japanese hemodialysis patients with secondary hyperparathyroidism. Ther Apher Dial 19: 225–234, 2015. [DOI] [PubMed] [Google Scholar]
- 11.Al Mheid I, Patel R, Murrow J, Morris A, Rahman A, Fike L, Kavtaradze N, Uphoff I, Hooper C, Tangpricha V, Alexander RW, Brigham K, Quyyumi AA. Vitamin D status is associated with arterial stiffness and vascular dysfunction in healthy humans. J Am Coll Cardiol 58: 186–192, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Al Mutair AN, Nasrat GH, Russell DW. Mutation of the CYP2R1 vitamin D 25-hydroxylase in a Saudi Arabian family with severe vitamin D deficiency. J Clin Endocrinol Metab 97: E2022–2025, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Albertson DG, Ylstra B, Segraves R, Collins C, Dairkee SH, Kowbel D, Kuo WL, Gray JW, Pinkel D. Quantitative mapping of amplicon structure by array CGH identifies CYP24 as a candidate oncogene. Nature Genet 25: 144–146, 2000. [DOI] [PubMed] [Google Scholar]
- 14.Alexander RT, Woudenberg-Vrenken TE, Buurman J, Dijkman H, van der Eerden BC, van Leeuwen JP, Bindels RJ, Hoenderop JG. Klotho prevents renal calcium loss. J Am Soc Nephrol 20: 2371–2379, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Aljubeh JM, Wang J, Al-Remeithi SS, Malloy PJ, Feldman D. Report of two unrelated patients with hereditary vitamin D resistant rickets due to the same novel mutation in the vitamin D receptor. J Pediatr Endocrinol Metab 24: 793–799, 2011. [DOI] [PubMed] [Google Scholar]
- 16.Alroy I, Towers TL, Freedman LP. Transcriptional repression of the interleukin-2 gene by vitamin D3: direct inhibition of NFATp/AP-1 complex formation by a nuclear hormone receptor. Mol Cell Biol 15: 5789–5799, 1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Alvarez-Diaz S, Valle N, Ferrer-Mayorga G, Lombardia L, Herrera M, Dominguez O, Segura MF, Bonilla F, Hernando E, Munoz A. MicroRNA-22 is induced by vitamin D and contributes to its antiproliferative, antimigratory and gene regulatory effects in colon cancer cells. Hum Mol Genet 21: 2157–2165, 2012. [DOI] [PubMed] [Google Scholar]
- 18.Amling M, Priemel M, Holzmann T, Chapin K, Rueger JM, Baron R, Demay MB. Rescue of the skeletal phenotype of vitamin D receptor-ablated mice in the setting of normal mineral ion homeostasis: formal histomorphometric and biomechanical analyses. Endocrinology 140: 4982–4987, 1999. [DOI] [PubMed] [Google Scholar]
- 19.Andrukhova O, Slavic S, Zeitz U, Riesen SC, Heppelmann MS, Ambrisko TD, Markovic M, Kuebler WM, Erben RG. Vitamin D is a regulator of endothelial nitric oxide synthase and arterial stiffness in mice. Mol Endocrinol 28: 53–64, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Andrukhova O, Smorodchenko A, Egerbacher M, Streicher C, Zeitz U, Goetz R, Shalhoub V, Mohammadi M, Pohl EE, Lanske B, Erben RG. FGF23 promotes renal calcium reabsorption through the TRPV5 channel. EMBO J 33: 229–246, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Annalora AJ, Goodin DB, Hong WX, Zhang Q, Johnson EF, Stout CD. Crystal structure of CYP24A1, a mitochondrial cytochrome P450 involved in vitamin D metabolism. J Mol Biol 396: 441–451, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Armbrecht HJ, Zenser TV, Davis BB. Effect of age on the conversion of 25-hydroxyvitamin D3 to 1,25-dihydroxyvitamin D3 by kidney of rat. J Clin Invest 66: 1118–1123, 1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Arnaud J, Constans J. Affinity differences for vitamin D metabolites associated with the genetic isoforms of the human serum carrier protein (DBP). Hum Genet 92: 183–188, 1993. [DOI] [PubMed] [Google Scholar]
- 24.Arora P, Song Y, Dusek J, Plotnikoff G, Sabatine MS, Cheng S, Valcour A, Swales H, Taylor B, Carney E, Guanaga D, Young JR, Karol C, Torre M, Azzahir A, Strachan SM, O'Neill DC, Wolf M, Harrell F, Newton-Cheh C, Wang TJ. Vitamin D therapy in individuals with prehypertension or hypertension: the DAYLIGHT trial. Circulation 131: 254–262, 2015. [DOI] [PubMed] [Google Scholar]
- 25.Attia S, Eickhoff J, Wilding G, McNeel D, Blank J, Ahuja H, Jumonville A, Eastman M, Shevrin D, Glode M, Alberti D, Staab MJ, Horvath D, Straus J, Marnocha R, Liu G. Randomized, double-blinded phase II evaluation of docetaxel with or without doxercalciferol in patients with metastatic, androgen-independent prostate cancer. Clin Cancer Res 14: 2437–2443, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Autier P, Boniol M, Pizot C, Mullie P. Vitamin D status and ill health: a systematic review. Lancet Diabetes Endocrinol 2: 76–89, 2014. [DOI] [PubMed] [Google Scholar]
- 27.Bacic D, Lehir M, Biber J, Kaissling B, Murer H, Wagner CA. The renal Na+/phosphate cotransporter NaPi-IIa is internalized via the receptor-mediated endocytic route in response to parathyroid hormone. Kidney Int 69: 495–503, 2006. [DOI] [PubMed] [Google Scholar]
- 28.Baeke F, Takiishi T, Korf H, Gysemans C, Mathieu C. Vitamin D: modulator of the immune system. Curr Opin Pharmacol 10: 482–496, 2010. [DOI] [PubMed] [Google Scholar]
- 29.Baldock PA, Thomas GP, Hodge JM, Baker SU, Dressel U, O'Loughlin PD, Nicholson GC, Briffa KH, Eisman JA, Gardiner EM. Vitamin D action and regulation of bone remodeling: suppression of osteoclastogenesis by the mature osteoblast. J Bone Miner Res 21: 1618–1626, 2006. [DOI] [PubMed] [Google Scholar]
- 30.Bao BY, Yeh SD, Lee YF. 1Alpha,25-dihydroxyvitamin D3 inhibits prostate cancer cell invasion via modulation of selective proteases. Carcinogenesis 27: 32–42, 2006. [DOI] [PubMed] [Google Scholar]
- 31.Barbour GL, Coburn JW, Slatopolsky E, Norman AW, Horst RL. Hypercalcemia in an anephric patient with sarcoidosis: evidence for extrarenal generation of 1,25-dihydroxyvitamin D. N Engl J Med 305: 440–443, 1981. [DOI] [PubMed] [Google Scholar]
- 32.Bareis P, Bises G, Bischof MG, Cross HS, Peterlik M. 25-Hydroxy-vitamin d metabolism in human colon cancer cells during tumor progression. Biochem Biophys Res Commun 285: 1012–1017, 2001. [DOI] [PubMed] [Google Scholar]
- 33.Barker JN, Ashton RE, Marks R, Harris RI, Berth-Jones J. Topical maxacalcitol for the treatment of psoriasis vulgaris: a placebo-controlled, double-blind, dose-finding study with active comparator. Br J Dermatol 141: 274–278, 1999. [DOI] [PubMed] [Google Scholar]
- 34.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, function. Cell 116: 281–297, 2004. [DOI] [PubMed] [Google Scholar]
- 35.Bauer SR, Hankinson SE, Bertone-Johnson ER, Ding EL. Plasma vitamin D levels, menopause, and risk of breast cancer: dose-response meta-analysis of prospective studies. Medicine 92: 123–131, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Belochitski O, Ariad S, Shany S, Fridman V, Gavrilov V. Efficient dual treatment of the hormone-refractory prostate cancer cell line DU145 with cetuximab and 1,25-dihydroxyvitamin D3. In Vivo 21: 371–376, 2007. [PubMed] [Google Scholar]
- 37.Benn BS, Ajibade D, Porta A, Dhawan P, Hediger M, Peng JB, Jiang Y, Oh GT, Jeung EB, Lieben L, Bouillon R, Carmeliet G, Christakos S. Active intestinal calcium transport in the absence of transient receptor potential vanilloid type 6 and calbindin-D9k. Endocrinology 149: 3196–3205, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bergh G, Telleus A, Fritzon A, Kornfalt S, Johnson E, Olsson I, Gullberg U. Forced expression of the cyclin-dependent kinase inhibitor p16(INK4A) in leukemic U-937 cells reveals dissociation between cell cycle and differentiation. Exp Hematol 29: 1382–1391, 2001. [DOI] [PubMed] [Google Scholar]
- 39.Berlanga-Taylor AJ, Knight JC. An integrated approach to defining genetic and environmental determinants for major clinical outcomes involving vitamin D. Mol Diagnosis Ther 18: 261–272, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Beveridge LA, Struthers AD, Khan F, Jorde R, Scragg R, Macdonald HM, Alvarez JA, Boxer RS, Dalbeni A, Gepner AD, Isbel NM, Larsen T, Nagpal J, Petchey WG, Stricker H, Strobel F, Tangpricha V, Toxqui L, Vaquero MP, Wamberg L, Zittermann A, Witham MD, Collaboration DP. Effect of vitamin D supplementation on blood pressure: a systematic review and meta-analysis incorporating individual patient data. JAMA Intern Med 175: 745–754, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bhalla AK, Amento EP, Clemens TL, Holick MF, Krane SM. Specific high-affinity receptors for 1,25-dihydroxyvitamin D3 in human peripheral blood mononuclear cells: presence in monocytes and induction in T lymphocytes following activation. J Clin Endocrinol Metab 57: 1308–1310, 1983. [DOI] [PubMed] [Google Scholar]
- 42.Bikle D, Adams J, Christakos S. Vitamin D: production, metabolism and clinical requirements. In: Primer Metab Bone Dis, edited by Rosen C. Hoboken, NJ: Wiley, 2013, p. 235–245. [Google Scholar]
- 42a.Bikle DD. Extra renal synthesis of 1,25-dihydroxyvitamin D and its health implications. Clin Rev Bone Miner Metab, 7: 114–125, 2009. [Google Scholar]
- 43.Bikle DD. Vitamin D and the skin: physiology and pathophysiology. Rev Endocr Metab Disorders 13: 3–19, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bikle DD. Vitamin D metabolism, mechanism of action, and clinical applications. Chem Biol 21: 319–329, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bischoff-Ferrari HA, Dietrich T, Orav EJ, Dawson-Hughes B. Positive association between 25-hydroxy vitamin D levels and bone mineral density: a population-based study of younger and older adults. Am J Med 116: 634–639, 2004. [DOI] [PubMed] [Google Scholar]
- 46.Bischoff-Ferrari HA, Willett WC, Orav EJ, Lips P, Meunier PJ, Lyons RA, Flicker L, Wark J, Jackson RD, Cauley JA, Meyer HE, Pfeifer M, Sanders KM, Stahelin HB, Theiler R, Dawson-Hughes B. A pooled analysis of vitamin D dose requirements for fracture prevention. N Engl J Med 367: 40–49, 2012. [DOI] [PubMed] [Google Scholar]
- 47.Bises G, Kallay E, Weiland T, Wrba F, Wenzl E, Bonner E, Kriwanek S, Obrist P, Cross HS. 25-Hydroxyvitamin D3-1alpha-hydroxylase expression in normal and malignant human colon. J Histochem Cytochem 52: 985–989, 2004. [DOI] [PubMed] [Google Scholar]
- 48.Bjelakovic G, Gluud LL, Nikolova D, Whitfield K, Krstic G, Wetterslev J, Gluud C. Vitamin D supplementation for prevention of cancer in adults. Cochrane Database Syst Rev 6: CD007469, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bjelakovic G, Gluud LL, Nikolova D, Whitfield K, Wetterslev J, Simonetti RG, Bjelakovic M, Gluud C. Vitamin D supplementation for prevention of mortality in adults. Cochrane Database Syst Rev 1: CD007470, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49a.Blaine J, Chonchol M, Levi M. Renal control of calcium, phosphate, and magnesium homeostasis. Clin J Am Soc Nephrol 10: 1257–1272, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Borkowski R, Du L, Zhao Z, McMillan E, Kosti A, Yang CR, Suraokar M, Wistuba II, Gazdar AF, Minna JD, White MA, Pertsemlidis A. Genetic mutation of p53 and suppression of the miR-17 approximately 92 cluster are synthetic lethal in non-small cell lung cancer due to upregulation of vitamin D signaling. Cancer Res 75: 666–675, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bosch X. Hypercalcemia due to endogenous overproduction of 1,25-dihydroxyvitamin D in Crohn's disease. Gastroenterology 114: 1061–1065, 1998. [DOI] [PubMed] [Google Scholar]
- 52.Bouillon R, Jones K, Schoenmakers I. Vitamin D-binding protein and vitamin D in blacks and whites. N Engl J Med 370: 879, 2014. [DOI] [PubMed] [Google Scholar]
- 53.Bouillon R, Okamura WH, Norman AW. Structure-function relationships in the vitamin D endocrine system. Endocr Rev 16: 200–257, 1995. [DOI] [PubMed] [Google Scholar]
- 54.Bouillon R, van Baelen H, de Moor P. Comparative study of the affinity of the serum vitamin D-binding protein. J Steroid Biochem 13: 1029–1034, 1980. [DOI] [PubMed] [Google Scholar]
- 55.Bouillon R, Van Schoor NM, Gielen E, Boonen S, Mathieu C, Vanderschueren D, Lips P. Optimal vitamin D status: a critical analysis on the basis of evidence-based medicine. J Clin Endocrinol Metab 98: E1283–1304, 2013. [DOI] [PubMed] [Google Scholar]
- 56.Bower M, Colston KW, Stein RC, Hedley A, Gazet JC, Ford HT, Combes RC. Topical calcipotriol treatment in advanced breast cancer. Lancet 337: 701–702, 1991. [DOI] [PubMed] [Google Scholar]
- 57.Boyan BD, Hurst-Kennedy J, Denison TA, Schwartz Z. 24R,25-dihydroxyvitamin D3 [24R,25(OH)2D3] controls growth plate development by inhibiting apoptosis in the reserve zone and stimulating response to 1alpha,25(OH)2D3 in hypertrophic cells. J Steroid Biochem Mol Biol 121: 212–216, 2010. [DOI] [PubMed] [Google Scholar]
- 58.Boyle BJ, Zhao XY, Cohen P, Feldman D. Insulin-like growth factor binding protein-3 mediates 1alpha,25-dihydroxyvitamin D3 growth inhibition in the LNCaP prostate cancer cell line through p21/WAF1. J Urol 165: 1319–1324, 2001. [PubMed] [Google Scholar]
- 59.Brenza HL, DeLuca HF. Regulation of 25-hydroxyvitamin D3 1alpha-hydroxylase gene expression by parathyroid hormone and 1,25-dihydroxyvitamin D3. Arch Biochem Biophys 381: 143–152, 2000. [DOI] [PubMed] [Google Scholar]
- 60.Brondum-Jacobsen P, Benn M, Jensen GB, Nordestgaard BG. 25-Hydroxyvitamin D levels and risk of ischemic heart disease, myocardial infarction, and early death: population-based study and meta-analyses of 18 and 17 studies. Arteriosclerosis Thrombosis Vasc Biol 32: 2794–2802, 2012. [DOI] [PubMed] [Google Scholar]
- 61.Brondum-Jacobsen P, Nordestgaard BG, Schnohr P, Benn M. 25-Hydroxyvitamin D and symptomatic ischemic stroke: an original study and meta-analysis. Ann Neurol 73: 38–47, 2013. [DOI] [PubMed] [Google Scholar]
- 62.Broulik PD, Horky K, Pacovsky V. Effect of parathyroid hormone on plasma renin activity in humans. Hormone Metab Res 18: 490–492, 1986. [DOI] [PubMed] [Google Scholar]
- 63.Bruce D, Whitcomb JP, August A, McDowell MA, Cantorna MT. Elevated non-specific immunity and normal Listeria clearance in young and old vitamin D receptor knockout mice. Int Immunol 21: 113–122, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Burgaz A, Orsini N, Larsson SC, Wolk A. Blood 25-hydroxyvitamin D concentration and hypertension: a meta-analysis. J Hypertens 29: 636–645, 2011. [DOI] [PubMed] [Google Scholar]
- 65.Caini S, Boniol M, Tosti G, Magi S, Medri M, Stanganelli I, Palli D, Assedi M, Marmol VD, Gandini S. Vitamin D and melanoma and non-melanoma skin cancer risk and prognosis: a comprehensive review and meta-analysis. Eur J Cancer 50: 2649–2658, 2014. [DOI] [PubMed] [Google Scholar]
- 66.Campbell MJ, Elstner E, Holden S, Uskokovic M, Koeffler HP. Inhibition of proliferation of prostate cancer cells by a 19-nor-hexafluoride vitamin D3 analogue involves the induction of p21waf1, p27kip1 and E-cadherin. J Mol Endocrinol 19: 15–27, 1997. [DOI] [PubMed] [Google Scholar]
- 67.Canaff L, Hendy GN. Human calcium-sensing receptor gene. Vitamin D response elements in promoters P1 and P2 confer transcriptional responsiveness to 1,25-dihydroxyvitamin D. J Biol Chem 277: 30337–30350, 2002. [DOI] [PubMed] [Google Scholar]
- 68.Cantorna MT. Mechanisms underlying the effect of vitamin D on the immune system. Proc Nutr Soc 69: 286–289, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Cantorna MT, Hayes CE, DeLuca HF. 1,25-Dihydroxyvitamin D3 reversibly blocks the progression of relapsing encephalomyelitis, a model of multiple sclerosis. Proc Natl Acad Sci USA 93: 7861–7864, 1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cantorna MT, Humpal-Winter J, DeLuca HF. Dietary calcium is a major factor in 1,25-dihydroxycholecalciferol suppression of experimental autoimmune encephalomyelitis in mice. J Nutr 129: 1966–1971, 1999. [DOI] [PubMed] [Google Scholar]
- 71.Cantorna MT, McDaniel K, Bora S, Chen J, James J. Vitamin D, immune regulation, the microbiota, and inflammatory bowel disease. Exp Biol Med 239: 1524–1530, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Carlberg C. Genome-wide (over)view on the actions of vitamin D. Frontiers Physiol 5: 167, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Casella SJ, Reiner BJ, Chen TC, Holick MF, Harrison HE. A possible genetic defect in 25-hydroxylation as a cause of rickets. J Pediatr 124: 929–932, 1994. [DOI] [PubMed] [Google Scholar]
- 74.Chang Q, Hoefs S, van der Kemp AW, Topala CN, Bindels RJ, Hoenderop JG. The beta-glucuronidase klotho hydrolyzes and activates the TRPV5 channel. Science 310: 490–493, 2005. [DOI] [PubMed] [Google Scholar]
- 75.Chang S, Gao L, Yang Y, Tong D, Guo B, Liu L, Li Z, Song T, Huang C. miR-145 mediates the antiproliferative and gene regulatory effects of vitamin D3 by directly targeting E2F3 in gastric cancer cells. Oncotarget 6: 7675–7685, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Chen A, Davis BH, Sitrin MD, Brasitus TA, Bissonnette M. Transforming growth factor-beta 1 signaling contributes to Caco-2 cell growth inhibition induced by 1,25(OH)2D3. Am J Physiol Gastrointest Liver Physiol 283: G864–G874, 2002. [DOI] [PubMed] [Google Scholar]
- 77.Chen ML, Perez A, Sanan DK, Heinrich G, Chen TC, Holick MF. Induction of vitamin D receptor mRNA expression in psoriatic plaques correlates with clinical response to 1,25-dihydroxyvitamin D3. J Invest Dermatol 106: 637–641, 1996. [DOI] [PubMed] [Google Scholar]
- 78.Chen S, Law CS, Grigsby CL, Olsen K, Hong TT, Zhang Y, Yeghiazarians Y, Gardner DG. Cardiomyocyte-specific deletion of the vitamin D receptor gene results in cardiac hypertrophy. Circulation 124: 1838–1847, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Cheng JB, Levine MA, Bell NH, Mangelsdorf DJ, Russell DW. Genetic evidence that the human CYP2R1 enzyme is a key vitamin D 25-hydroxylase. Proc Natl Acad Sci USA 101: 7711–7715, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Cheng JB, Motola DL, Mangelsdorf DJ, Russell DW. De-orphanization of cytochrome P450 2R1: a microsomal vitamin D 25-hydroxylase. J Biol Chem 278: 38084–38093, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Chowdhury R, Kunutsor S, Vitezova A, Oliver-Williams C, Chowdhury S, Kiefte-de-Jong JC, Khan H, Baena CP, Prabhakaran D, Hoshen MB, Feldman BS, Pan A, Johnson L, Crowe F, Hu FB, Franco OH. Vitamin D and risk of cause specific death: systematic review and meta-analysis of observational cohort and randomised intervention studies. BMJ 348: g1903, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Chowdhury R, Stevens S, Ward H, Chowdhury S, Sajjad A, Franco OH. Circulating vitamin D, calcium and risk of cerebrovascular disease: a systematic review and meta-analysis. Eur J Epidemiol 27: 581–591, 2012. [DOI] [PubMed] [Google Scholar]
- 83.Christakos S. Vitamin D gene regulation In: Principles of Bone Biology, edited by Bilezikian J Raisz LG, Martin TJ. New York: Elsevier-Academic, 2008, p. 779–794. [Google Scholar]
- 84.Christakos S, Dhawan P, Benn B, Porta A, Hediger M, Oh GT, Jeung EB, Zhong Y, Ajibade D, Dhawan K, Joshi S. Vitamin D: molecular mechanism of action. Ann NY Acad Sci 1116: 340–348, 2007. [DOI] [PubMed] [Google Scholar]
- 85.Christakos S, Lieben L, Masuyama R, Carmeliet G. Vitamin D endocrine system and the intestine. Bone Key Reports 3: 496, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Christakos S, Seth T, Hirsch J, Porta A, Moulas A, Dhawan P. Vitamin D biology revealed through the study of knockout and transgenic mouse models. Annu Rev Nutr 33: 71–85, 2013. [DOI] [PubMed] [Google Scholar]
- 87.Chun RF. New perspectives on the vitamin D binding protein. Cell Biochem Funct 30: 445–456, 2012. [DOI] [PubMed] [Google Scholar]
- 88.Chun RF, Liu PT, Modlin RL, Adams JS, Hewison M. Impact of vitamin D on immune function: lessons learned from genome-wide analysis. Front Physiol 5: 151, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Chun RF, Peercy BE, Orwoll ES, Nielson CM, Adams JS, Hewison M. Vitamin D and DBP: the free hormone hypothesis revisited. J Steroid Biochem Mol Biol 144: 132–137, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Chung M, Lee J, Terasawa T, Lau J, Trikalinos TA. Vitamin D with or without calcium supplementation for prevention of cancer and fractures: an updated meta-analysis for the US Preventive Services Task Force. Ann Internal Med 155: 827–838, 2011. [DOI] [PubMed] [Google Scholar]
- 91.Ciesielski F, Rochel N, Moras D. Adaptability of the Vitamin D nuclear receptor to the synthetic ligand Gemini: remodelling the LBP with one side chain rotation. J Steroid Biochem Mol Biol 103: 235–242, 2007. [DOI] [PubMed] [Google Scholar]
- 92.Cippitelli M, Santoni A. Vitamin D3: a transcriptional modulator of the interferon-gamma gene. Eur J Immunol 28: 3017–3030, 1998. [DOI] [PubMed] [Google Scholar]
- 93.Clinckspoor I, Gerard AC, Van Sande J, Many MC, Verlinden L, Bouillon R, Carmeliet G, Mathieu C, Verstuyf A, Decallonne B. The vitamin D receptor in thyroid development and function. Eur Thyroid J 1: 168–175, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Coburn JW, Maung HM, Elangovan L, Germain MJ, Lindberg JS, Sprague SM, Williams ME, Bishop CW. Doxercalciferol safely suppresses PTH levels in patients with secondary hyperparathyroidism associated with chronic kidney disease stages 3 and 4. Am J Kidney Dis 43: 877–890, 2004. [DOI] [PubMed] [Google Scholar]
- 95.Colston K, Colston MJ, Feldman D. 1,25-Dihydroxyvitamin D3 and malignant melanoma: the presence of receptors and inhibition of cell growth in culture. Endocrinology 108: 1083–1086, 1981. [DOI] [PubMed] [Google Scholar]
- 96.Consiglio M, Destefanis M, Morena D, Foglizzo V, Forneris M, Pescarmona G, Silvagno F. The vitamin D receptor inhibits the respiratory chain, contributing to the metabolic switch that is essential for cancer cell proliferation. PloS One 9: e115816, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Constans J, Hazout S, Garruto RM, Gajdusek DC, Spees EK. Population distribution of the human vitamin D binding protein: anthropological considerations. Am J Physical Anthropol 68: 107–122, 1985. [DOI] [PubMed] [Google Scholar]
- 98.Coyne DW, Andress DL, Amdahl MJ, Ritz E, de Zeeuw D. Effects of paricalcitol on calcium and phosphate metabolism and markers of bone health in patients with diabetic nephropathy: results of the VITAL study. Nephrol Dialysis Transplant 28: 2260–2268, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Cranney A, Horsley T, O'Donnell S, Weiler H, Puil L, Ooi D, Atkinson S, Ward L, Moher D, Hanley D, Fang M, Yazdi F, Garritty C, Sampson M, Barrowman N, Tsertsvadze A, Mamaladze V. Effectiveness and safety of vitamin D in relation to bone health. Evidence Report Technol Assessment 1–235, 2007. [PMC free article] [PubMed] [Google Scholar]
- 100.Crofts LA, Hancock MS, Morrison NA, Eisman JA. Multiple promoters direct the tissue-specific expression of novel N-terminal variant human vitamin D receptor gene transcripts. Proc Natl Acad Sci USA 95: 10529–10534, 1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Cross HS, Corradino RA, Peterlik M. Calcitriol-dependent, paracellular sodium transport in the embryonic chick intestine. Mol Cell Endocrinol 53: 53–58, 1987. [DOI] [PubMed] [Google Scholar]
- 102.Cui M, Li Q, Johnson R, Fleet JC. Villin promoter-mediated transgenic expression of transient receptor potential cation channel, subfamily V, member 6 (TRPV6) increases intestinal calcium absorption in wild-type and vitamin D receptor knockout mice. J Bone Miner Res 27: 2097–2107, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Cunningham J, Locatelli F, Rodriguez M. Secondary hyperparathyroidism: pathogenesis, disease progression, and therapeutic options. Clin J Am Soc Nephrol 6: 913–921, 2011. [DOI] [PubMed] [Google Scholar]
- 104.D'Ambrosio D, Cippitelli M, Cocciolo MG, Mazzeo D, Di Lucia P, Lang R, Sinigaglia F, Panina-Bordignon P. Inhibition of IL-12 production by 1,25-dihydroxyvitamin D3. Involvement of NF-kappaB downregulation in transcriptional repression of the p40 gene. J Clin Invest 101: 252–262, 1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Dalhoff K, Dancey J, Astrup L, Skovsgaard T, Hamberg KJ, Lofts FJ, Rosmorduc O, Erlinger S, Bach Hansen J, Steward WP, Skov T, Burcharth F, Evans TR. A phase II study of the vitamin D analogue Seocalcitol in patients with inoperable hepatocellular carcinoma. Br J Cancer 89: 252–257, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Daniel C, Sartory NA, Zahn N, Radeke HH, Stein JM. Immune modulatory treatment of trinitrobenzene sulfonic acid colitis with calcitriol is associated with a change of a T helper (Th) 1/Th17 to a Th2 and regulatory T cell profile. J Pharmacol Exp Ther 324: 23–33, 2008. [DOI] [PubMed] [Google Scholar]
- 107.Dardenne O, Prud'homme J, Arabian A, Glorieux FH, St-Arnaud R. Targeted inactivation of the 25-hydroxyvitamin D(3)-1(alpha)-hydroxylase gene (CYP27B1) creates an animal model of pseudovitamin D-deficiency rickets. Endocrinology 142: 3135–3141, 2001. [DOI] [PubMed] [Google Scholar]
- 108.DeLuca HF. Evolution of our understanding of vitamin D. Nutr Rev 66: S73–87, 2008. [DOI] [PubMed] [Google Scholar]
- 109.DeLuca HF, Bedale W, Binkley N, Gallagher JC, Bolognese M, Peacock M, Aloia J, Clagett-Dame M, Plum L. The vitamin D analogue 2MD increases bone turnover but not BMD in postmenopausal women with osteopenia: results of a 1-year phase 2 double-blind, placebo-controlled, randomized clinical trial. J Bone Miner Res 26: 538–545, 2011. [DOI] [PubMed] [Google Scholar]
- 110.DeLuca HF, Cantorna MT. Vitamin D: its role and uses in immunology. FASEB J 15: 2579–2585, 2001. [DOI] [PubMed] [Google Scholar]
- 111.Demay MB, Kiernan MS, DeLuca HF, Kronenberg HM. Sequences in the human parathyroid hormone gene that bind the 1,25-dihydroxyvitamin D3 receptor and mediate transcriptional repression in response to 1,25-dihydroxyvitamin D3. Proc Natl Acad Sci USA 89: 8097–8101, 1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Dhawan P, Hsaio C, Yehia H, Porta P, LL, Carmeliet G, Christakos S. Transgenic expression of the vitamin D receptor restricted to the ileum, cecum and colon of VDR knockout mice rescues VDR dependent rickets. Bone Miner Res S34, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Dhawan P, Peng X, Sutton AL, MacDonald PN, Croniger CM, Trautwein C, Centrella M, McCarthy TL, Christakos S. Functional cooperation between CCAAT/enhancer-binding proteins and the vitamin D receptor in regulation of 25-hydroxyvitamin D3 24-hydroxylase. Mol Cell Biol 25: 472–487, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Dhawan P, Wei R, Sun C, Gombart AF, Koeffler HP, Diamond G, Christakos S. C/EBPalpha and the vitamin D receptor cooperate in the regulation of cathelicidin in lung epithelial cells. J Cell Physiol 230: 464–472, 2015. [DOI] [PubMed] [Google Scholar]
- 115.Dhawan P, Wieder R, Christakos S. CCAAT enhancer-binding protein alpha is a molecular target of 1,25-dihydroxyvitamin D3 in MCF-7 breast cancer cells. J Biol Chem 284: 3086–3095, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Dheerendra PC, Sakhuja V, Kohli HS, Jha V. Efficacy and safety of oral doxercalciferol in the management of secondary hyperparathyroidism in chronic kidney disease stage 4. Indian J Nephrol 23: 271–275, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Ding N, Yu RT, Subramaniam N, Sherman MH, Wilson C, Rao R, Leblanc M, Coulter S, He M, Scott C, Lau SL, Atkins AR, Barish GD, Gunton JE, Liddle C, Downes M, Evans RM. A vitamin D receptor/SMAD genomic circuit gates hepatic fibrotic response. Cell 153: 601–613, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Dinour D, Beckerman P, Ganon L, Tordjman K, Eisenstein Z, Holtzman EJ. Loss-of-function mutations of CYP24A1, the vitamin D 24-hydroxylase gene, cause long-standing hypercalciuric nephrolithiasis and nephrocalcinosis. J Urol 190: 552–557, 2013. [DOI] [PubMed] [Google Scholar]
- 119.Ditsch N, Toth B, Mayr D, Lenhard M, Gallwas J, Weissenbacher T, Dannecker C, Friese K, Jeschke U. The association between vitamin D receptor expression and prolonged overall survival in breast cancer. J Histochem Cytochem 60: 121–129, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Dobnig H, Pilz S, Scharnagl H, Renner W, Seelhorst U, Wellnitz B, Kinkeldei J, Boehm BO, Weihrauch G, Maerz W. Independent association of low serum 25-hydroxyvitamin D and 1,25-dihydroxyvitamin D levels with all-cause and cardiovascular mortality. Arch Internal Med 168: 1340–1349, 2008. [DOI] [PubMed] [Google Scholar]
- 121.Dong Q, Miller WL. Vitamin D 25-hydroxylase deficiency. Mol Genet Metab 83: 197–198, 2004. [DOI] [PubMed] [Google Scholar]
- 122.Dong Y, Stallmann-Jorgensen IS, Pollock NK, Harris RA, Keeton D, Huang Y, Li K, Bassali R, Guo DH, Thomas J, Pierce GL, White J, Holick MF, Zhu H. A 16-week randomized clinical trial of 2000 international units daily vitamin D3 supplementation in black youth: 25-hydroxyvitamin D, adiposity, and arterial stiffness. J Clin Endocrinol Metab 95: 4584–4591, 2010. [DOI] [PubMed] [Google Scholar]
- 123.Donohue MM, Demay MB. Rickets in VDR null mice is secondary to decreased apoptosis of hypertrophic chondrocytes. Endocrinology 143: 3691–3694, 2002. [DOI] [PubMed] [Google Scholar]
- 124.Dowd P, Dowd DR. Coregulators of VDR mediated gene expression. In: Vitamin D, edited by Feldman D, Pike JW, Adams JS. New York: Elsevier, 2011, p. 193–209. [Google Scholar]
- 125.Durup D, Jorgensen HL, Christensen J, Schwarz P, Heegaard AM, Lind B. A reverse J-shaped association of all-cause mortality with serum 25-hydroxyvitamin D in general practice: the CopD study. J Clin Endocrinol Metab 97: 2644–2652, 2012. [DOI] [PubMed] [Google Scholar]
- 126.Dwivedi PP, Omdahl JL, Kola I, Hume DA, May BK. Regulation of rat cytochrome P450C24 (CYP24) gene expression. Evidence for functional cooperation of Ras-activated Ets transcription factors with the vitamin D receptor in 1,25-dihydroxyvitamin D3-mediated induction. J Biol Chem 275: 47–55, 2000. [DOI] [PubMed] [Google Scholar]
- 127.Ebeling PR, Sandgren ME, DiMagno EP, Lane AW, DeLuca HF, Riggs BL. Evidence of an age-related decrease in intestinal responsiveness to vitamin D: relationship between serum 1,25-dihydroxyvitamin D3 and intestinal vitamin D receptor concentrations in normal women. J Clin Endocrinol Metab 75: 176–182, 1992. [DOI] [PubMed] [Google Scholar]
- 128.Eelen G, Valle N, Sato Y, Rochel N, Verlinden L, De Clercq P, Moras D, Bouillon R, Munoz A, Verstuyf A. Superagonistic fluorinated vitamin D3 analogs stabilize helix 12 of the vitamin D receptor. Chem Biol 15: 1029–1034, 2008. [DOI] [PubMed] [Google Scholar]
- 129.Eelen G, Verlinden L, Rochel N, Claessens F, De Clercq P, Vandewalle M, Tocchini-Valentini G, Moras D, Bouillon R, Verstuyf A. Superagonistic action of 14-epi-analogs of 1,25-dihydroxyvitamin D explained by vitamin D receptor-coactivator interaction. Mol Pharmacol 67: 1566–1573, 2005. [DOI] [PubMed] [Google Scholar]
- 130.Eelen G, Verlinden L, Van Camp M, Claessens F, De Clercq P, Vandewalle M, Bouillon R, Verstuyf A. Altered Vitamin D receptor-coactivator interactions reflect superagonism of Vitamin D analogs. J Steroid Biochem Mol Biol 97: 65–68, 2005. [DOI] [PubMed] [Google Scholar]
- 131.Elamin MB, Abu Elnour NO, Elamin KB, Fatourechi MM, Alkatib AA, Almandoz JP, Liu H, Lane MA, Mullan RJ, Hazem A, Erwin PJ, Hensrud DD, Murad MH, Montori VM. Vitamin D and cardiovascular outcomes: a systematic review and meta-analysis. J Clin Endocrinol Metab 96: 1931–1942, 2011. [DOI] [PubMed] [Google Scholar]
- 132.Engelman CD, Fingerlin TE, Langefeld CD, Hicks PJ, Rich SS, Wagenknecht LE, Bowden DW, Norris JM. Genetic and environmental determinants of 25-hydroxyvitamin D and 1,25-dihydroxyvitamin D levels in Hispanic and African Americans. J Clin Endocrinol Metab 93: 3381–3388, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Erben RG, Soegiarto DW, Weber K, Zeitz U, Lieberherr M, Gniadecki R, Moller G, Adamski J, Balling R. Deletion of deoxyribonucleic acid binding domain of the vitamin D receptor abrogates genomic and nongenomic functions of vitamin D. Mol Endocrinol 16: 1524–1537, 2002. [DOI] [PubMed] [Google Scholar]
- 134.Essa S, Denzer N, Mahlknecht U, Klein R, Collnot EM, Tilgen W, Reichrath J. VDR microRNA expression and epigenetic silencing of vitamin D signaling in melanoma cells. J Steroid Biochem Mol Biol 121: 110–113, 2010. [DOI] [PubMed] [Google Scholar]
- 135.Essa S, Reichrath S, Mahlknecht U, Montenarh M, Vogt T, Reichrath J. Signature of VDR miRNAs and epigenetic modulation of vitamin D signaling in melanoma cell lines. Anticancer Res 32: 383–389, 2012. [PubMed] [Google Scholar]
- 136.Esteban L, Vidal M, Dusso A. 1alpha-Hydroxylase transactivation by gamma-interferon in murine macrophages requires enhanced C/EBPbeta expression and activation. J Steroid Biochem Mol Biol 89–90: 131–137, 2004. [DOI] [PubMed] [Google Scholar]
- 137.Evans KN, Nguyen L, Chan J, Innes BA, Bulmer JN, Kilby MD, Hewison M. Effects of 25-hydroxyvitamin D3 and 1,25-dihydroxyvitamin D3 on cytokine production by human decidual cells. Biol Reprod 75: 816–822, 2006. [DOI] [PubMed] [Google Scholar]
- 138.Evans TR, Colston KW, Lofts FJ, Cunningham D, Anthoney DA, Gogas H, de Bono JS, Hamberg KJ, Skov T, Mansi JL. A phase II trial of the vitamin D analogue Seocalcitol (EB1089) in patients with inoperable pancreatic cancer. Br J Cancer 86: 680–685, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Farrow EG, Davis SI, Summers LJ, White KE. Initial FGF23-mediated signaling occurs in the distal convoluted tubule. J Am Soc Nephrol 20: 955–960, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Favus MJ. Factors that influence absorption and secretion of calcium in the small intestine and colon. Am J Physiol Gastrointest Liver Physiol 248: G147–G157, 1985. [DOI] [PubMed] [Google Scholar]
- 141.Favus MJ, Angeid-Backman E. Effects of 1,25(OH)2D3 and calcium channel blockers on cecal calcium transport in the rat. Am J Physiol Gastrointest Liver Physiol 248: G676–G681, 1985. [DOI] [PubMed] [Google Scholar]
- 142.Feldman D, Krishnan AV, Swami S, Giovannucci E, Feldman BJ. The role of vitamin D in reducing cancer risk and progression. Nature Rev Cancer 14: 342–357, 2014. [DOI] [PubMed] [Google Scholar]
- 143.Feldman D, Malloy PJ. Mutations in the vitamin D receptor and hereditary vitamin D-resistant rickets. Bone Key Reports 3: 510, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Ferreira GB, Gysemans CA, Demengeot J, da Cunha JP, Vanherwegen AS, Overbergh L, Van Belle TL, Pauwels F, Verstuyf A, Korf H, Mathieu C. 1,25-Dihydroxyvitamin D3 promotes tolerogenic dendritic cells with functional migratory properties in NOD mice. J Immunol 192: 4210–4220, 2014. [DOI] [PubMed] [Google Scholar]
- 145.Ferreira GB, van Etten E, Verstuyf A, Waer M, Overbergh L, Gysemans C, Mathieu C. 1,25-Dihydroxyvitamin D3 alters murine dendritic cell behaviour in vitro and in vivo. Diabetes Metab Res Rev 2 7: 933–941, 2011. [DOI] [PubMed] [Google Scholar]
- 146.Ferreira GB, Vanherwegen AS, Eelen G, Gutierrez AC, Van Lommel L, Marchal K, Verlinden L, Verstuyf A, Nogueira T, Georgiadou M, Schuit F, Eizirik DL, Gysemans C, Carmeliet P, Overbergh L, Mathieu C. Vitamin D3 induces tolerance in human dendritic cells by activation of intracellular metabolic pathways. Cell Reports. 10: 711–725, 2015. [DOI] [PubMed] [Google Scholar]
- 147.Fleck A. Latitude and ischaemic heart disease. Lancet 1: 613, 1989. [DOI] [PubMed] [Google Scholar]
- 148.Forman JP, Scott JB, Ng K, Drake BF, Suarez EG, Hayden DL, Bennett GG, Chandler PD, Hollis BW, Emmons KM, Giovannucci EL, Fuchs CS, Chan AT. Effect of vitamin D supplementation on blood pressure in blacks. Hypertension 61: 779–785, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Forman JP, Williams JS, Fisher ND. Plasma 25-hydroxyvitamin D and regulation of the renin-angiotensin system in humans. Hypertension 55: 1283–1288, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Forster RE, Jurutka PW, Hsieh JC, Haussler CA, Lowmiller CL, Kaneko I, Haussler MR, Kerr Whitfield G. Vitamin D receptor controls expression of the anti-aging klotho gene in mouse and human renal cells. Biochem Biophys Res Commun 414: 557–562, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Fortmann SP, Burda BU, Senger CA, Lin JS, Whitlock EP. Vitamin and mineral supplements in the primary prevention of cardiovascular disease and cancer: an updated systematic evidence review for the US Preventive Services Task Force. Ann Intern Med 159: 824–834, 2013. [DOI] [PubMed] [Google Scholar]
- 152.Frazao JM, Elangovan L, Maung HM, Chesney RW, Acchiardo SR, Bower JD, Kelley BJ, Rodriguez HJ, Norris KC, Robertson JA, Levine BS, Goodman WG, Gentile D, Mazess RB, Kyllo DM, Douglass LL, Bishop CW, Coburn JW. Intermittent doxercalciferol (1alpha-hydroxyvitamin D2) therapy for secondary hyperparathyroidism. Am J Kidney Dis 36: 550–561, 2000. [DOI] [PubMed] [Google Scholar]
- 153.Fretz JA, Zella LA, Kim S, Shevde NK, Pike JW. 1,25-Dihydroxyvitamin D3 regulates the expression of low-density lipoprotein receptor-related protein 5 via deoxyribonucleic acid sequence elements located downstream of the start site of transcription. Mol Endocrinol 20: 2215–2230, 2006. [DOI] [PubMed] [Google Scholar]
- 154.Friedrich M, Rafi L, Mitschele T, Tilgen W, Schmidt W, Reichrath J. Analysis of the vitamin D system in cervical carcinomas, breast cancer and ovarian cancer. Recent Results Cancer Res 164: 239–246, 2003. [DOI] [PubMed] [Google Scholar]
- 155.Fujita H, Sugimoto K, Inatomi S, Maeda T, Osanai M, Uchiyama Y, Yamamoto Y, Wada T, Kojima T, Yokozaki H, Yamashita T, Kato S, Sawada N, Chiba H. Tight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytes. Mol Biol Cell 19: 1912–1921, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Galante L, Colston KW, MacAuley SJ, MacIntyre I. Effect of calcitonin on vitamin D metabolism. Nature 238: 271–273, 1972. [DOI] [PubMed] [Google Scholar]
- 157.Gardiner EM, Baldock PA, Thomas GP, Sims NA, Henderson NK, Hollis B, White CP, Sunn KL, Morrison NA, Walsh WR, Eisman JA. Increased formation and decreased resorption of bone in mice with elevated vitamin D receptor in mature cells of the osteoblastic lineage. FASEB J 14: 1908–1916, 2000. [DOI] [PubMed] [Google Scholar]
- 158.Garland CF, Garland FC. Do sunlight and vitamin D reduce the likelihood of colon cancer? Int J Epidemiol 9: 227–231, 1980. [DOI] [PubMed] [Google Scholar]
- 159.Gee J, Bailey H, Kim K, Kolesar J, Havighurst T, Tutsch KD, See W, Cohen MB, Street N, Levan L, Jarrard D, Wilding G. Phase II open label, multi-center clinical trial of modulation of intermediate endpoint biomarkers by 1alpha-hydroxyvitamin D2 in patients with clinically localized prostate cancer and high grade pin. The Prostate 73: 970–978, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Geleijnse JM. Vitamin D and the prevention of hypertension and cardiovascular diseases: a review of the current evidence. Am J Hypertens 24: 253–262, 2011. [DOI] [PubMed] [Google Scholar]
- 161.Giallauria F, Milaneschi Y, Tanaka T, Maggio M, Canepa M, Elango P, Vigorito C, Lakatta EG, Ferrucci L, Strait J. Arterial stiffness and vitamin D levels: the Baltimore longitudinal study of aging. J Clin Endocrinol Metab 97: 3717–3723, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Giangreco AA, Dambal S, Wagner D, Van der Kwast T, Vieth R, Prins GS, Nonn L. Differential expression and regulation of vitamin D hydroxylases and inflammatory genes in prostate stroma and epithelium by 1,25-dihydroxyvitamin D in men with prostate cancer and an in vitro model. J Steroid Biochem Mol Biol 148: 156–165, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Giangreco AA, Vaishnav A, Wagner D, Finelli A, Fleshner N, Van der Kwast T, Vieth R, Nonn L. Tumor suppressor microRNAs, miR-100 and -125b, are regulated by 1,25-dihydroxyvitamin D in primary prostate cells and in patient tissue. Cancer Prevention Res 6: 483–494, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Giovannucci E, Liu Y, Hollis BW, Rimm EB. 25-Hydroxyvitamin D and risk of myocardial infarction in men: a prospective study. Arch Internal Med 168: 1174–1180, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Gkika D, Hsu YJ, van der Kemp AW, Christakos S, Bindels RJ, Hoenderop JG. Critical role of the epithelial Ca2+ channel TRPV5 in active Ca2+ reabsorption as revealed by TRPV5/calbindin-D28K knockout mice. J Am Soc Nephrol : JASN 17: 3020–3027, 2006. [DOI] [PubMed] [Google Scholar]
- 166.Glorieux FH, Pettifor JM. Vitamin D/dietary calcium deficiency rickets and pseudo-vitamin D deficiency rickets. Bone Key Reports 3: 524, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Gocek E, Wang X, Liu X, Liu CG, Studzinski GP. MicroRNA-32 upregulation by 1,25-dihydroxyvitamin D3 in human myeloid leukemia cells leads to Bim targeting and inhibition of AraC-induced apoptosis. Cancer Res 71: 6230–6239, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Golovko O, Nazarova N, Tuohimaa P. Vitamin D-induced up-regulation of tumour necrosis factor alpha (TNF-alpha) in prostate cancer cells. Life sciences 77: 562–577, 2005. [DOI] [PubMed] [Google Scholar]
- 169.Golub EE. Role of matrix vesicles in biomineralization. Biochim Biophys Acta 1790: 1592–1598, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Gombart AF, Borregaard N, Koeffler HP. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J 19: 1067–1077, 2005. [DOI] [PubMed] [Google Scholar]
- 171.Gonzalez-Pardo V, Suares A, Verstuyf A, De Clercq P, Boland R, de Boland AR. Cell cycle arrest and apoptosis induced by 1alpha,25(OH)2D3 and TX 527 in Kaposi sarcoma is VDR dependent. J Steroid Biochem Mol Biol 144: 197–200, 2014. [DOI] [PubMed] [Google Scholar]
- 172.Gonzalez MM, Samenfeld P, Perakyla M, Carlberg C. Corepressor excess shifts the two-side chain vitamin D analog Gemini from an agonist to an inverse agonist of the vitamin D receptor. Mol Endocrinol 17: 2028–2038, 2003. [DOI] [PubMed] [Google Scholar]
- 173.Gregori S, Giarratana N, Smiroldo S, Uskokovic M, Adorini L. A 1alpha,25-dihydroxyvitamin D3 analog enhances regulatory T-cells and arrests autoimmune diabetes in NOD mice. Diabetes 51: 1367–1374, 2002. [DOI] [PubMed] [Google Scholar]
- 174.Griffin MD, Lutz W, Phan VA, Bachman LA, McKean DJ, Kumar R. Dendritic cell modulation by 1alpha,25 dihydroxyvitamin D3 and its analogs: a vitamin D receptor-dependent pathway that promotes a persistent state of immaturity in vitro and in vivo. Proc Natl Acad Sci USA 98: 6800–6805, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Guallar E, Miller ER 3rd, Ordovas JM, Stranges S. Vitamin D supplementation in the age of lost innocence. Ann Intern Med 152: 327–329, 2010. [DOI] [PubMed] [Google Scholar]
- 176.Guallar E, Stranges S, Mulrow C, Appel LJ, Miller ER 3rd. Enough is enough: stop wasting money on vitamin and mineral supplements. Ann Intern Med 159: 850–851, 2013. [DOI] [PubMed] [Google Scholar]
- 177.Gulliford T, English J, Colston KW, Menday P, Moller S, Coombes RC. A phase I study of the vitamin D analogue EB 1089 in patients with advanced breast and colorectal cancer. Br J Cancer 78: 6–13, 1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Guo B, Aslam F, van Wijnen AJ, Roberts SG, Frenkel B, Green MR, DeLuca H, Lian JB, Stein GS, Stein JL. YY1 regulates vitamin D receptor/retinoid X receptor mediated transactivation of the vitamin D responsive osteocalcin gene. Proc Natl Acad Sci USA 94: 121–126, 1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Guo Z, Wen J, Kan Q, Huang S, Liu X, Sun N, Li Z. Lack of association between vitamin D receptor gene FokI and BsmI polymorphisms and prostate cancer risk: an updated meta-analysis involving 21,756 subjects. Tumour Biol 34: 3189–3200, 2013. [DOI] [PubMed] [Google Scholar]
- 180.Gutierrez S, Javed A, Tennant DK, van Rees M, Montecino M, Stein GS, Stein JL, Lian JB. CCAAT/enhancer-binding proteins (C/EBP) beta and delta activate osteocalcin gene transcription and synergize with Runx2 at the C/EBP element to regulate bone-specific expression. J Biol Chem 277: 1316–1323, 2002. [DOI] [PubMed] [Google Scholar]
- 181.Handel AE, Sandve GK, Disanto G, Berlanga-Taylor AJ, Gallone G, Hanwell H, Drablos F, Giovannoni G, Ebers GC, Ramagopalan SV. Vitamin D receptor ChIP-seq in primary CD4+ cells: relationship to serum 25-hydroxyvitamin D levels and autoimmune disease. BMC Med 11: 163, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Harvey NC, Cooper C. Vitamin D: some perspective please. BMJ 345: e4695, 2012. [DOI] [PubMed] [Google Scholar]
- 183.Haussler MR, Whitfield GK, Kaneko I, Haussler CA, Hsieh D, Hsieh JC, Jurutka PW. Molecular mechanisms of vitamin D action. Calcified Tissue Int 92: 77–98, 2013. [DOI] [PubMed] [Google Scholar]
- 184.Heaney RP, Horst RL, Cullen DM, Armas LA. Vitamin D3 distribution and status in the body. J Am Coll Nutr 28: 252–256, 2009. [DOI] [PubMed] [Google Scholar]
- 185.Hegyi Z, Zwicker S, Bureik D, Peric M, Koglin S, Batycka-Baran A, Prinz JC, Ruzicka T, Schauber J, Wolf R. Vitamin D analog calcipotriol suppresses the Th17 cytokine-induced proinflammatory S100 “alarmins” psoriasin (S100A7) and koebnerisin (S100A15) in psoriasis. J Invest Dermatol 132: 1416–1424, 2012. [DOI] [PubMed] [Google Scholar]
- 186.Hendrickson WK, Flavin R, Kasperzyk JL, Fiorentino M, Fang F, Lis R, Fiore C, Penney KL, Ma J, Kantoff PW, Stampfer MJ, Loda M, Mucci LA, Giovannucci E. Vitamin D receptor protein expression in tumor tissue and prostate cancer progression. J Clin Oncol 29: 2378–2385, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Henry HL. Regulation of vitamin D metabolism. Best practice & research. Clin Endocrinol Metab 25: 531–541, 2011. [DOI] [PubMed] [Google Scholar]
- 188.Henry HL, Norman AW. Vitamin D: two dihydroxylated metabolites are required for normal chicken egg hatchability. Science 201: 835–837, 1978. [DOI] [PubMed] [Google Scholar]
- 189.Hobaus J, Hummel DM, Thiem U, Fetahu IS, Aggarwal A, Mullauer L, Heller G, Egger G, Mesteri I, Baumgartner-Parzer S, Kallay E. Increased copy-number and not DNA hypomethylation causes overexpression of the candidate proto-oncogene CYP24A1 in colorectal cancer. Int J Cancer 133: 1380–1388, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Hobaus J, Thiem U, Hummel DM, Kallay E. Role of calcium, vitamin D, and the extrarenal vitamin D hydroxylases in carcinogenesis. Anti-cancer Agents Med Chem 13: 20–35, 2013. [PMC free article] [PubMed] [Google Scholar]
- 191.Hochberg Z, Tiosano D, Even L. Calcium therapy for calcitriol-resistant rickets. J Pediatr 121: 803–808, 1992. [DOI] [PubMed] [Google Scholar]
- 192.Hoenderop JG, Dardenne O, Van Abel M, Van Der Kemp AW, Van Os CH, St-Arnaud R, Bindels RJ. Modulation of renal Ca2+ transport protein genes by dietary Ca2+ and 1,25-dihydroxyvitamin D3 in 25-hydroxyvitamin D3-1alpha-hydroxylase knockout mice. FASEB J 16: 1398–1406, 2002. [DOI] [PubMed] [Google Scholar]
- 193.Hoenderop JG, van Leeuwen JP, van der Eerden BC, Kersten FF, van der Kemp AW, Merillat AM, Waarsing JH, Rossier BC, Vallon V, Hummler E, Bindels RJ. Renal Ca2+ wasting, hyperabsorption, and reduced bone thickness in mice lacking TRPV5. J Clin Invest 112: 1906–1914, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Holick MF. Resurrection of vitamin D deficiency and rickets. J Clin Invest 116: 2062–2072, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Holick MF. Vitamin D deficiency. N Engl J Med 357: 266–281, 2007. [DOI] [PubMed] [Google Scholar]
- 196.Hollis BW, Bikle DD. Vitamin D-binding protein and vitamin D in blacks and whites. N Engl J Med 370: 879–880, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Hoyer-Hansen M, Bastholm L, Mathiasen IS, Elling F, Jaattela M. Vitamin D analog EB1089 triggers dramatic lysosomal changes and Beclin 1-mediated autophagic cell death. Cell Death Differentiation 12: 1297–1309, 2005. [DOI] [PubMed] [Google Scholar]
- 198.Hsia J, Heiss G, Ren H, Allison M, Dolan NC, Greenland P, Heckbert SR, Johnson KC, Manson JE, Sidney S, Trevisan M, Women's Health Initiative I . Calcium/vitamin D supplementation and cardiovascular events. Circulation 115: 846–854, 2007. [DOI] [PubMed] [Google Scholar]
- 199.Hu MC, Shiizaki K, Kuro-o M, Moe OW. Fibroblast growth factor 23 and Klotho: physiology and pathophysiology of an endocrine network of mineral metabolism. Annu Rev Physiol 75: 503–533, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Huet T, Laverny G, Ciesielski F, Molnar F, Ramamoorthy TG, Belorusova AY, Antony P, Potier N, Metzger D, Moras D, Rochel N. A vitamin D receptor selectively activated by gemini analogs reveals ligand dependent and independent effects. Cell Reports 10: 516–526, 2015. [DOI] [PubMed] [Google Scholar]
- 201.Huet T, Maehr H, Lee HJ, Uskokovic MR, Suh N, Moras D, Rochel N. Structure-function study of gemini derivatives with two different side chains at C-20, Gemini-0072 and Gemini-0097. Med Chem Comm 2: 424–429, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Hughes MR, Malloy PJ, Kieback DG, Kesterson RA, Pike JW, Feldman D, O'Malley BW. Point mutations in the human vitamin D receptor gene associated with hypocalcemic rickets. Science 242: 1702–1705, 1988. [DOI] [PubMed] [Google Scholar]
- 203.Hylander E, Ladefoged K, Jarnum S. Calcium absorption after intestinal resection. The importance of a preserved colon. Scand J Gastroenterol 25: 705–710, 1990. [DOI] [PubMed] [Google Scholar]
- 204.Iglesias-Gato D, Zheng S, Flanagan JN, Jiang L, Kittaka A, Sakaki T, Yamamoto K, Itoh T, Lebrasseur NK, Norstedt G, Chen TC. Substitution at carbon 2 of 19-nor-1alpha,25-dihydroxyvitamin D3 with 3-hydroxypropyl group generates an analogue with enhanced chemotherapeutic potency in PC-3 prostate cancer cells. J Steroid Biochem Mol Biol 127: 269–275, 2011. [DOI] [PubMed] [Google Scholar]
- 206.Jablonski KL, Chonchol M, Pierce GL, Walker AE, Seals DR. 25-Hydroxyvitamin D deficiency is associated with inflammation-linked vascular endothelial dysfunction in middle-aged and older adults. Hypertension 57: 63–69, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Jacobs ET, Van Pelt C, Forster RE, Zaidi W, Hibler EA, Galligan MA, Haussler MR, Jurutka PW. CYP24A1 and CYP27B1 polymorphisms modulate vitamin D metabolism in colon cancer cells. Cancer Res 73: 2563–2573, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Jain RK, Trump DL, Egorin MJ, Fernandez M, Johnson CS, Ramanathan RK. A phase I study of the vitamin D3 analogue ILX23-7553 administered orally to patients with advanced solid tumors. Invest New Drugs 29: 1420–1425, 2011. [DOI] [PubMed] [Google Scholar]
- 209.Jarnagin K, Brommage R, DeLuca HF, Yamada S, Takayama H. 1- but not 24-hydroxylation of vitamin D is required for growth and reproduction in rats. Am J Physiol Endocrinol Metab 244: E290–E297, 1983. [DOI] [PubMed] [Google Scholar]
- 210.Jeffery LE, Burke F, Mura M, Zheng Y, Qureshi OS, Hewison M, Walker LS, Lammas DA, Raza K, Sansom DM. 1,25-Dihydroxyvitamin D3 and IL-2 combine to inhibit T cell production of inflammatory cytokines and promote development of regulatory T cells expressing CTLA-4 and FoxP3. J Immunol 183: 5458–5467, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Jensen SS, Madsen MW, Lukas J, Binderup L, Bartek J. Inhibitory effects of 1alpha,25-dihydroxyvitamin D3 on the G1-S phase-controlling machinery. Mol Endocrinol 15: 1370–1380, 2001. [DOI] [PubMed] [Google Scholar]
- 212.Joist HE, Ahya SN, Giles K, Norwood K, Slatopolsky E, Coyne DW. Differential effects of very high doses of doxercalciferol and paricalcitol on serum phosphorus in hemodialysis patients. Clin Nephrol 65: 335–341, 2006. [DOI] [PubMed] [Google Scholar]
- 213.Jones G. Analog metabolism. In: Vitamin D, edited by Feldman D. San Diego: Academic, 1997, p. 973–994. [Google Scholar]
- 214.Jones G, Prosser DE, Kaufmann M. 25-Hydroxyvitamin D-24-hydroxylase (CYP24A1): its important role in the degradation of vitamin D. Arch Biochem Biophys 523: 9–18, 2012. [DOI] [PubMed] [Google Scholar]
- 215.Jones G, Prosser DE, Kaufmann M. Cytochrome P450-mediated metabolism of vitamin D. J Lipid Res 55: 13–31, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Jones G, Prosser DG. The activating enzymes of vitamin D. Vitamin D 23–42, 2011. [Google Scholar]
- 217.Joshi S, Pantalena LC, Liu XK, Gaffen SL, Liu H, Rohowsky-Kochan C, Ichiyama K, Yoshimura A, Steinman L, Christakos S, Youssef S. 1,25-Dihydroxyvitamin D3 ameliorates Th17 autoimmunity via transcriptional modulation of interleukin-17A. Mol Cell Biol 31: 3653–3669, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Judd SE, Nanes MS, Ziegler TR, Wilson PW, Tangpricha V. Optimal vitamin D status attenuates the age-associated increase in systolic blood pressure in white Americans: results from the third National Health and Nutrition Examination Survey. Am J Clin Nutr 87: 136–141, 2008. [DOI] [PubMed] [Google Scholar]
- 219.Kamboh MI, Ferrell RE. Ethnic variation in vitamin D-binding protein (GC): a review of isoelectric focusing studies in human populations. Hum Genet 72: 281–293, 1986. [DOI] [PubMed] [Google Scholar]
- 220.Kang S, Yi S, Griffiths CE, Fancher L, Hamilton TA, Choi JH. Calcipotriene-induced improvement in psoriasis is associated with reduced interleukin-8 and increased interleukin-10 levels within lesions. Br J Dermatol 138: 77–83, 1998. [DOI] [PubMed] [Google Scholar]
- 221.Kanis JA, McCloskey EV, Johansson H, Cooper C, Rizzoli R, Reginster JY, Scientific Advisory Board of the European Society for Cancer, Economic Aspects of Osteoporosis, Osteoarthritis, and the Committee of Scientific Advisors of the International Osteoporosis. European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporosis Int 24: 23–57, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Kao CY, Chen Y, Thai P, Wachi S, Huang F, Kim C, Harper RW, Wu R. IL-17 markedly up-regulates beta-defensin-2 expression in human airway epithelium via JAK and NF-kappaB signaling pathways. J Immunol 173: 3482–3491, 2004. [DOI] [PubMed] [Google Scholar]
- 223.Karbach U. Paracellular calcium transport across the small intestine. J Nutr 122: 672–677, 1992. [DOI] [PubMed] [Google Scholar]
- 224.Kasiappan R, Shen Z, Tse AK, Jinwal U, Tang J, Lungchukiet P, Sun Y, Kruk P, Nicosia SV, Zhang X, Bai W. 1,25-Dihydroxyvitamin D3 suppresses telomerase expression and human cancer growth through microRNA-498. J Biol Chem 287: 41297–41309, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Kawashima H, Torikai S, Kurokawa K. Calcitonin selectively stimulates 25-hydroxyvitamin D3-1 alpha-hydroxylase in proximal straight tubule of rat kidney. Nature 291: 327–329, 1981. [DOI] [PubMed] [Google Scholar]
- 226.Ke HZ, Qi H, Crawford DT, Simmons HA, Xu G, Li M, Plum L, Clagett-Dame M, DeLuca HF, Thompson DD, Brown TA. A new vitamin D analog, 2MD, restores trabecular and cortical bone mass and strength in ovariectomized rats with established osteopenia. J Bone Miner Res 20: 1742–1755, 2005. [DOI] [PubMed] [Google Scholar]
- 227.Kemmis CM, Welsh J. Mammary epithelial cell transformation is associated with deregulation of the vitamin D pathway. J Cell Biochem 105: 980–988, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Kendrick J, Targher G, Smits G, Chonchol M. 25-Hydroxyvitamin D deficiency is independently associated with cardiovascular disease in the Third National Health and Nutrition Examination Survey. Atherosclerosis 205: 255–260, 2009. [DOI] [PubMed] [Google Scholar]
- 229.Keum N, Giovannucci E. Vitamin D supplements and cancer incidence and mortality: a meta-analysis. Br J Cancer 111: 976–980, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Khader SA, Gaffen SL, Kolls JK. Th17 cells at the crossroads of innate and adaptive immunity against infectious diseases at the mucosa. Mucosal Immunol 2: 403–411, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Kidney Disease: Improving Global Outcomes. KDIGO clinical practice guideline for the diagnosis, evaluation, prevention, and treatment of Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD). Kidney Int Suppl S1–130, 2009. [DOI] [PubMed] [Google Scholar]
- 232.Kim JH, Yamaori S, Tanabe T, Johnson CH, Krausz KW, Kato S, Gonzalez FJ. Implication of intestinal VDR deficiency in inflammatory bowel disease. Biochim Biophys Acta 1830: 2118–2128, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Kim S, Yamazaki M, Zella LA, Shevde NK, Pike JW. Activation of receptor activator of NF-kappaB ligand gene expression by 1,25-dihydroxyvitamin D3 is mediated through multiple long-range enhancers. Mol Cell Biol 26: 6469–6486, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Kitanaka S, Takeyama K, Murayama A, Sato T, Okumura K, Nogami M, Hasegawa Y, Niimi H, Yanagisawa J, Tanaka T, Kato S. Inactivating mutations in the 25-hydroxyvitamin D3 1alpha-hydroxylase gene in patients with pseudovitamin D-deficiency rickets. N Engl J Med 338: 653–661, 1998. [DOI] [PubMed] [Google Scholar]
- 235.Koli K, Keski-Oja J. 1Alpha,25-dihydroxyvitamin D3 and its analogues down-regulate cell invasion-associated proteases in cultured malignant cells. Cell Growth Differ 11: 221–229, 2000. [PubMed] [Google Scholar]
- 236.Kondo S, Takano T, Ono Y, Saito H, Matsumoto T. Eldecalcitol reduces osteoporotic fractures by unique mechanisms. J Steroid Biochem Mol Biol 148: 232–238, 2015. [DOI] [PubMed] [Google Scholar]
- 237.Kong J, Xu F, Qu J, Wang Y, Gao M, Yu H, Qian B. Genetic polymorphisms in the vitamin D pathway in relation to lung cancer risk and survival. Oncotarget 6: 2573–2582, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Kostner K, Denzer N, Muller CS, Klein R, Tilgen W, Reichrath J. The relevance of vitamin D receptor (VDR) gene polymorphisms for cancer: a review of the literature. Anticancer Res 29: 3511–3536, 2009. [PubMed] [Google Scholar]
- 239.Koszewski NJ, Alimov AP, Park-Sarge OK, Malluche HH. Suppression of the human parathyroid hormone promoter by vitamin D involves displacement of NF-Y binding to the vitamin D response element. J Biol Chem 279: 42431–42437, 2004. [DOI] [PubMed] [Google Scholar]
- 240.Kovalenko PL, Zhang Z, Cui M, Clinton SK, Fleet JC. 1,25 Dihydroxyvitamin D-mediated orchestration of anticancer, transcript-level effects in the immortalized, non-transformed prostate epithelial cell line, RWPE1. BMC Gen 11: 26, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Kuchuk NO, Pluijm SM, van Schoor NM, Looman CW, Smit JH, Lips P. Relationships of serum 25-hydroxyvitamin D to bone mineral density and serum parathyroid hormone and markers of bone turnover in older persons. J Clin Endocrinol Metab 94: 1244–1250, 2009. [DOI] [PubMed] [Google Scholar]
- 242.Kunutsor SK, Apekey TA, Steur M. Vitamin D and risk of future hypertension: meta-analysis of 283,537 participants. Eur J Epidemiol 28: 205–221, 2013. [DOI] [PubMed] [Google Scholar]
- 243.Kunutsor SK, Burgess S, Munroe PB, Khan H. Vitamin D and high blood pressure: causal association or epiphenomenon? Eur J Epidemiol 29: 1–14, 2014. [DOI] [PubMed] [Google Scholar]
- 244.Kupferschmidt K. Uncertain verdict as vitamin D goes on trial. Science 337: 1476–1478, 2012. [DOI] [PubMed] [Google Scholar]
- 245.Kuro-o M, Matsumura Y, Aizawa H, Kawaguchi H, Suga T, Utsugi T, Ohyama Y, Kurabayashi M, Kaname T, Kume E, Iwasaki H, Iida A, Shiraki-Iida T, Nishikawa S, Nagai R, Nabeshima YI. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature 390: 45–51, 1997. [DOI] [PubMed] [Google Scholar]
- 246.Kutuzova GD, Deluca HF. Gene expression profiles in rat intestine identify pathways for 1,25-dihydroxyvitamin D3 stimulated calcium absorption and clarify its immunomodulatory properties. Arch Biochem Biophys 432: 152–166, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246a.Larriba MJ, Gonzalez-Sancho JM, Barbachano A, Niell N, Ferrer-Mayorga G, Munoz A. Vitamin D is a multilevel repressor of Wnt/b-catenin signaling in cancer cells. Cancers (Basel) 5: 1242–1260, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Larriba MJ, Ordonez-Moran P, Chicote I, Martin-Fernandez G, Puig I, Munoz A, Palmer HG. Vitamin D receptor deficiency enhances Wnt/beta-catenin signaling and tumor burden in colon cancer. PloS One 6: e23524, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Larsen T, Mose FH, Bech JN, Hansen AB, Pedersen EB. Effect of cholecalciferol supplementation during winter months in patients with hypertension: a randomized, placebo-controlled trial. Am J Hypertens 25: 1215–1222, 2012. [DOI] [PubMed] [Google Scholar]
- 249.Lawrence JA, Akman SA, Melin SA, Case LD, Schwartz GG. Oral paricalcitol (19-nor-1,25-dihydroxyvitamin D2) in women receiving chemotherapy for metastatic breast cancer: a feasibility trial. Cancer Biol Ther 14: 476–480, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Lee DB, Walling MM, Levine BS, Gafter U, Silis V, Hodsman A, Coburn JW. Intestinal and metabolic effect of 1,25-dihydroxyvitamin D3 in normal adult rat. Am J Physiol Gastrointest Liver Physiol 240: G90–G96, 1981. [DOI] [PubMed] [Google Scholar]
- 251.Lee HJ, Paul S, Atalla N, Thomas PE, Lin X, Yang I, Buckley B, Lu G, Zheng X, Lou YR, Conney AH, Maehr H, Adorini L, Uskokovic M, Suh N. Gemini vitamin D analogues inhibit estrogen receptor-positive and estrogen receptor-negative mammary tumorigenesis without hypercalcemic toxicity. Cancer Prevention Res 1: 476–484, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Lee JE, Li H, Chan AT, Hollis BW, Lee IM, Stampfer MJ, Wu K, Giovannucci E, Ma J. Circulating levels of vitamin D and colon and rectal cancer: the Physicians' Health Study and a meta-analysis of prospective studies. Cancer Prevention Res 4: 735–743, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Lee SM, Bishop KA, Goellner JJ, O'Brien CA, Pike JW. Mouse and human BAC transgenes recapitulate tissue-specific expression of the vitamin D receptor in mice and rescue the VDR-null phenotype. Endocrinology 155: 2064–2076, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Lee SM, Goellner JJ, O'Brien CA, Pike JW. A humanized mouse model of hereditary 1,25-dihydroxyvitamin D-resistant rickets without alopecia. Endocrinology 155: 4137–4148, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Lemire JM, Adams JS, Kermani-Arab V, Bakke AC, Sakai R, Jordan SC. 1,25-Dihydroxyvitamin D3 suppresses human T helper/inducer lymphocyte activity in vitro. J Immunol 134: 3032–3035, 1985. [PubMed] [Google Scholar]
- 256.Li M, Chen P, Li J, Chu R, Xie D, Wang H. Review: the impacts of circulating 25-hydroxyvitamin D levels on cancer patient outcomes: a systematic review and meta-analysis. J Clin Endocrinol Metab 99: 2327–2336, 2014. [DOI] [PubMed] [Google Scholar]
- 257.Li SA, Watanabe M, Yamada H, Nagai A, Kinuta M, Takei K. Immunohistochemical localization of Klotho protein in brain, kidney, and reproductive organs of mice. Cell Struct Funct 29: 91–99, 2004. [DOI] [PubMed] [Google Scholar]
- 258.Li YC, Amling M, Pirro AE, Priemel M, Meuse J, Baron R, Delling G, Demay MB. Normalization of mineral ion homeostasis by dietary means prevents hyperparathyroidism, rickets, and osteomalacia, but not alopecia in vitamin D receptor-ablated mice. Endocrinology 139: 4391–4396, 1998. [DOI] [PubMed] [Google Scholar]
- 259.Li YC, Bolt MJ, Cao LP, Sitrin MD. Effects of vitamin D receptor inactivation on the expression of calbindins and calcium metabolism. Am J Physiol Endocrinol Metab 281: E558–E564, 2001. [DOI] [PubMed] [Google Scholar]
- 260.Li YC, Kong J, Wei M, Chen ZF, Liu SQ, Cao LP. 1,25-Dihydroxyvitamin D3 is a negative endocrine regulator of the renin-angiotensin system. J Clin Invest 110: 229–238, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Li Z, Jia Z, Gao Y, Xie D, Wei D, Cui J, Mishra L, Huang S, Zhang Y, Xie K. Activation of vitamin D receptor signaling downregulates the expression of nuclear FOXM1 protein and suppresses pancreatic cancer cell stemness. Clin Cancer Res 21: 844–853, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Lieben L, Benn BS, Ajibade D, Stockmans I, Moermans K, Hediger MA, Peng JB, Christakos S, Bouillon R, Carmeliet G. Trpv6 mediates intestinal calcium absorption during calcium restriction and contributes to bone homeostasis. Bone 47: 301–308, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Lieben L, Masuyama R, Torrekens S, Van Looveren R, Schrooten J, Baatsen P, Lafage-Proust MH, Dresselaers T, Feng JQ, Bonewald LF, Meyer MB, Pike JW, Bouillon R, Carmeliet G. Normocalcemia is maintained in mice under conditions of calcium malabsorption by vitamin D-induced inhibition of bone mineralization. J Clin Invest 122: 1803–1815, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Lips P, Gielen E, van Schoor NM. Vitamin D supplements with or without calcium to prevent fractures. Bone Key Reports 3: 512, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Liu G, Wilding G, Staab MJ, Horvath D, Miller K, Dresen A, Alberti D, Arzoomanian R, Chappell R, Bailey HH. Phase II study of 1alpha-hydroxyvitamin D2 in the treatment of advanced androgen-independent prostate cancer. Clinical Cancer Res 9: 4077–4083, 2003. [PubMed] [Google Scholar]
- 266.Liu N, Kaplan AT, Low J, Nguyen L, Liu GY, Equils O, Hewison M. Vitamin D induces innate antibacterial responses in human trophoblasts via an intracrine pathway. Biol Reprod 80: 398–406, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.Liu NQ, Kaplan AT, Lagishetty V, Ouyang YB, Ouyang Y, Simmons CF, Equils O, Hewison M. Vitamin D and the regulation of placental inflammation. J Immunol 186: 5968–5974, 2011. [DOI] [PubMed] [Google Scholar]
- 268.Liu PT, Stenger S, Tang DH, Modlin RL. Cutting edge: vitamin D-mediated human antimicrobial activity against Mycobacterium tuberculosis is dependent on the induction of cathelicidin. J Immunol 179: 2060–2063, 2007. [DOI] [PubMed] [Google Scholar]
- 269.Liu S, Tang W, Zhou J, Stubbs JR, Luo Q, Pi M, Quarles LD. Fibroblast growth factor 23 is a counter-regulatory phosphaturic hormone for vitamin D. J Am Soc Nephrol 17: 1305–1315, 2006. [DOI] [PubMed] [Google Scholar]
- 269a.Liu W, Chen Y, Golan MA, Annunziata ML, Du J, Dougherty U, Kong J, Musch M, Huang Y, Pekow J, Zheng C, Bissonnette M, Hanauer SB, Li YC. Intestinal epithelial vitamin D receptor signaling inhibits experimental colitis. J Clin Invest 123: 3983–3996, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270.Liu Y, Li C, Chen P, Li X, Li M, Guo H, Li J, Chu R, Wang H. Polymorphisms in the vitamin D Receptor (VDR) and the risk of ovarian cancer: a meta-analysis. PloS One 8: e66716, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Llach F, Yudd M. Paricalcitol in dialysis patients with calcitriol-resistant secondary hyperparathyroidism. Am J Kidney Dis 38: S45–50, 2001. [DOI] [PubMed] [Google Scholar]
- 272.Lopes N, Sousa B, Martins D, Gomes M, Vieira D, Veronese LA, Milanezi F, Paredes J, Costa JL, Schmitt F. Alterations in vitamin D signalling and metabolic pathways in breast cancer progression: a study of VDR, CYP27B1 and CYP24A1 expression in benign and malignant breast lesions. BMC Cancer 10: 483, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.Lund RJ, Andress DL, Amdahl M, Williams LA, Heaney RP. Differential effects of paricalcitol and calcitriol on intestinal calcium absorption in hemodialysis patients. Am J Nephrol 31: 165–170, 2010. [DOI] [PubMed] [Google Scholar]
- 274.Lundqvist J, Hansen SK, Lykkesfeldt AE. Vitamin D analog EB1089 inhibits aromatase expression by dissociation of comodulator WSTF from the CYP19A1 promoter-a new regulatory pathway for aromatase. Biochim Biophys Acta 1833: 40–47, 2013. [DOI] [PubMed] [Google Scholar]
- 275.Levine MA, Dang A, Ding C, Fischer PR, Singh R, Thacher T. Tropical rickets in Nigeria: mutation of the CYP2R1 gene encoding vitamin D 25-hydroxylase as a cause of vitamin D dependent rickets. Bone 40: s60–s61, 2007. [Google Scholar]
- 276.Ma Y, Yu WD, Su B, Seshadri M, Luo W, Trump DL, Johnson CS. Regulation of motility, invasion, and metastatic potential of squamous cell carcinoma by 1alpha,25-dihydroxycholecalciferol. Cancer 119: 563–574, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Ma Y, Yu WD, Trump DL, Johnson CS. 1,25D3 enhances antitumor activity of gemcitabine and cisplatin in human bladder cancer models. Cancer 116: 3294–3303, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278.Ma Y, Zhang P, Wang F, Yang J, Liu Z, Qin H. Association between vitamin D and risk of colorectal cancer: a systematic review of prospective studies. J Clin Oncol 29: 3775–3782, 2011. [DOI] [PubMed] [Google Scholar]
- 279.Mackey SL, Heymont JL, Kronenberg HM, Demay MB. Vitamin D receptor binding to the negative human parathyroid hormone vitamin D response element does not require the retinoid x receptor. Mol Endocrinol 10: 298–305, 1996. [DOI] [PubMed] [Google Scholar]
- 280.Mahon BD, Wittke A, Weaver V, Cantorna MT. The targets of vitamin D depend on the differentiation and activation status of CD4 positive T cells. J Cell Biochem 89: 922–932, 2003. [DOI] [PubMed] [Google Scholar]
- 281.Malloy PJ, Pike JW, Feldman D. The vitamin D receptor and the syndrome of hereditary 1,25-dihydroxyvitamin D-resistant rickets. Endocr Rev 20: 156–188, 1999. [DOI] [PubMed] [Google Scholar]
- 282.Malloy PJ, Xu R, Peng L, Clark PA, Feldman D. A novel mutation in helix 12 of the vitamin D receptor impairs coactivator interaction and causes hereditary 1,25-dihydroxyvitamin D-resistant rickets without alopecia. Mol Endocrinol 16: 2538–2546, 2002. [DOI] [PubMed] [Google Scholar]
- 283.Mancuso P, Rahman A, Hershey SD, Dandu L, Nibbelink KA, Simpson RU. 1,25-Dihydroxyvitamin-D3 treatment reduces cardiac hypertrophy and left ventricular diameter in spontaneously hypertensive heart failure-prone (cp/+) rats independent of changes in serum leptin. J Cardiovasc Pharmacol 51: 559–564, 2008. [DOI] [PubMed] [Google Scholar]
- 284.Mann MC, Hobbs AJ, Hemmelgarn BR, Roberts DJ, Ahmed SB, Rabi DM. Effect of oral vitamin D analogs on mortality and cardiovascular outcomes among adults with chronic kidney disease: a meta-analysis. Clin Kidney J 8: 41–48, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Manson JE, Bassuk SS, Lee IM, Cook NR, Albert MA, Gordon D, Zaharris E, Macfadyen JG, Danielson E, Lin J, Zhang SM, Buring JE. The VITamin D and OmegA-3 TriaL (VITAL): rationale and design of a large randomized controlled trial of vitamin D and marine omega-3 fatty acid supplements for the primary prevention of cancer and cardiovascular disease. Contemp Clin Trials 33: 159–171, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Margolis KL, Ray RM, Van Horn L, Manson JE, Allison MA, Black HR, Beresford SA, Connelly SA, Curb JD, Grimm RH Jr, Kotchen TA, Kuller LH, Wassertheil-Smoller S, Thomson CA, Torner JC, Women's Health Initiative I . Effect of calcium and vitamin D supplementation on blood pressure: the Women's Health Initiative Randomized Trial. Hypertension 52: 847–855, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Margolis RN, Christakos S. The nuclear receptor superfamily of steroid hormones and vitamin D gene regulation. An update. Ann NY Acad Sci 1192: 208–214, 2010. [DOI] [PubMed] [Google Scholar]
- 288.Marniemi J, Alanen E, Impivaara O, Seppanen R, Hakala P, Rajala T, Ronnemaa T. Dietary and serum vitamins and minerals as predictors of myocardial infarction and stroke in elderly subjects. Nutr Metab Cardiovasc Dis 15: 188–197, 2005. [DOI] [PubMed] [Google Scholar]
- 289.Martin KJ, Gonzalez EA, Gellens M, Hamm LL, Abboud H, Lindberg J. 19-Nor-1-alpha-25-dihydroxyvitamin D2 (Paricalcitol) safely and effectively reduces the levels of intact parathyroid hormone in patients on hemodialysis. J Am Soc Nephrol 9: 1427–1432, 1998. [DOI] [PubMed] [Google Scholar]
- 290.Martin KJ, Gonzalez EA, Gellens ME, Hamm LL, Abboud H, Lindberg J. Therapy of secondary hyperparathyroidism with 19-nor-1alpha,25-dihydroxyvitamin D2. Am J Kidney Dis 32: S61–66, 1998. [DOI] [PubMed] [Google Scholar]
- 291.Martins D, Wolf M, Pan D, Zadshir A, Tareen N, Thadhani R, Felsenfeld A, Levine B, Mehrotra R, Norris K. Prevalence of cardiovascular risk factors and the serum levels of 25-hydroxyvitamin D in the United States: data from the Third National Health and Nutrition Examination Survey. Arch Internal Med 167: 1159–1165, 2007. [DOI] [PubMed] [Google Scholar]
- 292.Masuyama R, Nakaya Y, Katsumata S, Kajita Y, Uehara M, Tanaka S, Sakai A, Kato S, Nakamura T, Suzuki K. Dietary calcium and phosphorus ratio regulates bone mineralization and turnover in vitamin D receptor knockout mice by affecting intestinal calcium and phosphorus absorption. J Bone Miner Res 18: 1217–1226, 2003. [DOI] [PubMed] [Google Scholar]
- 293.Masuyama R, Stockmans I, Torrekens S, Van Looveren R, Maes C, Carmeliet P, Bouillon R, Carmeliet G. Vitamin D receptor in chondrocytes promotes osteoclastogenesis and regulates FGF23 production in osteoblasts. J Clin Invest 116: 3150–3159, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 294.Mathieu C, Waer M, Laureys J, Rutgeerts O, Bouillon R. Prevention of autoimmune diabetes in NOD mice by 1,25 dihydroxyvitamin D3. Diabetologia 37: 552–558, 1994. [DOI] [PubMed] [Google Scholar]
- 295.Matkovits T, Christakos S. Variable in vivo regulation of rat vitamin D-dependent genes (osteopontin, Ca,Mg-adenosine triphosphatase, and 25-hydroxyvitamin D3 24-hydroxylase): implications for differing mechanisms of regulation and involvement of multiple factors. Endocrinology 136: 3971–3982, 1995. [DOI] [PubMed] [Google Scholar]
- 296.Matsumoto T, Ito M, Hayashi Y, Hirota T, Tanigawara Y, Sone T, Fukunaga M, Shiraki M, Nakamura T. A new active vitamin D3 analog, eldecalcitol, prevents the risk of osteoporotic fractures–a randomized, active comparator, double-blind study. Bone 49: 605–612, 2011. [DOI] [PubMed] [Google Scholar]
- 297.Matsumoto T, Miki T, Hagino H, Sugimoto T, Okamoto S, Hirota T, Tanigawara Y, Hayashi Y, Fukunaga M, Shiraki M, Nakamura T. A new active vitamin D, ED-71, increases bone mass in osteoporotic patients under vitamin D supplementation: a randomized, double-blind, placebo-controlled clinical trial. J Clin Endocrinol Metab 90: 5031–5036, 2005. [DOI] [PubMed] [Google Scholar]
- 298.Mattner F, Smiroldo S, Galbiati F, Muller M, Di Lucia P, Poliani PL, Martino G, Panina-Bordignon P, Adorini L. Inhibition of Th1 development and treatment of chronic-relapsing experimental allergic encephalomyelitis by a non-hypercalcemic analogue of 1,25-dihydroxyvitamin D3. Eur J Immunol 30: 498–508, 2000. [DOI] [PubMed] [Google Scholar]
- 299.Matusiak D, Benya RV. CYP27A1 and CYP24 expression as a function of malignant transformation in the colon. J Histochem Cytochem 55: 1257–1264, 2007. [DOI] [PubMed] [Google Scholar]
- 300.Maund SL, Barclay WW, Hover LD, Axanova LS, Sui G, Hipp JD, Fleet JC, Thorburn A, Cramer SD. Interleukin-1alpha mediates the antiproliferative effects of 1,25-dihydroxyvitamin D3 in prostate progenitor/stem cells. Cancer Res 71: 5276–5286, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Maung HM, Elangovan L, Frazao JM, Bower JD, Kelley BJ, Acchiardo SR, Rodriguez HJ, Norris KC, Sigala JF, Rutkowski M, Robertson JA, Goodman WG, Levine BS, Chesney RW, Mazess RB, Kyllo DM, Douglass LL, Bishop CW, Coburn JW. Efficacy and side effects of intermittent intravenous and oral doxercalciferol (1alpha-hydroxyvitamin D2) in dialysis patients with secondary hyperparathyroidism: a sequential comparison. Am J Kidney Dis 37: 532–543, 2001. [PubMed] [Google Scholar]
- 302.McGrath JJ, Saha S, Burne TH, Eyles DW. A systematic review of the association between common single nucleotide polymorphisms and 25-hydroxyvitamin D concentrations. J Steroid Biochem Mol Biol 121: 471–477, 2010. [DOI] [PubMed] [Google Scholar]
- 303.McGuire TF, Trump DL, Johnson CS. Vitamin D3-induced apoptosis of murine squamous cell carcinoma cells. Selective induction of caspase-dependent MEK cleavage and up-regulation of MEKK-1. J Biol Chem 276: 26365–26373, 2001. [DOI] [PubMed] [Google Scholar]
- 304.Meeker S, Seamons A, Paik J, Treuting PM, Brabb T, Grady WM, Maggio-Price L. Increased dietary vitamin D suppresses MAPK signaling, colitis, and colon cancer. Cancer Res 74: 4398–4408, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Melamed ML, Michos ED, Post W, Astor B. 25-Hydroxyvitamin D levels and the risk of mortality in the general population. Arch Internal Med 168: 1629–1637, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306.Messenger W, Nielson CM, Li H, Beer T, Barrett-Connor E, Stone K, Shannon J. Serum and dietary vitamin D and cardiovascular disease risk in elderly men: a prospective cohort study. Nutr Metab Cardiovasc Dis 22: 856–863, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Meyer MB, Benkusky NA, Lee CH, Pike JW. Genomic determinants of gene regulation by 1,25-dihydroxyvitamin D3 during osteoblast-lineage cell differentiation. J Biol Chem 289: 19539–19554, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Meyer MB, Benkusky NA, Pike JW. Selective distal enhancer control of the Mmp13 gene identified through clustered regularly interspaced short palindromic repeat (CRISPR) genomic deletions. J Biol Chem 290: 11093–11107, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309.Meyer MB, Goetsch PD, Pike JW. A downstream intergenic cluster of regulatory enhancers contributes to the induction of CYP24A1 expression by 1alpha,25-dihydroxyvitamin D3. J Biol Chem 285: 15599–15610, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Meyer MB, Watanuki M, Kim S, Shevde NK, Pike JW. The human transient receptor potential vanilloid type 6 distal promoter contains multiple vitamin D receptor binding sites that mediate activation by 1,25-dihydroxyvitamin D3 in intestinal cells. Mol Endocrinol 20: 1447–1461, 2006. [DOI] [PubMed] [Google Scholar]
- 311.Milani C, Katayama ML, de Lyra EC, Welsh J, Campos LT, Brentani MM, Maciel Mdo S, Roela RA, del Valle PR, Goes JC, Nonogaki S, Tamura RE, Folgueira MA. Transcriptional effects of 1,25 dihydroxyvitamin D3 physiological and supra-physiological concentrations in breast cancer organotypic culture. BMC Cancer 13: 119, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312.Milczarek M, Rosinska S, Psurski M, Maciejewska M, Kutner A, Wietrzyk J. Combined colonic cancer treatment with vitamin D analogs and irinotecan or oxaliplatin. AntiCancer Res 33: 433–444, 2013. [PubMed] [Google Scholar]
- 313.Min D, Lv XB, Wang X, Zhang B, Meng W, Yu F, Hu H. Downregulation of miR-302c and miR-520c by 1,25(OH)2D3 treatment enhances the susceptibility of tumour cells to natural killer cell-mediated cytotoxicity. Br J Cancer 109: 723–730, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314.Moe SM. Vitamin D, cardiovascular disease, and survival in dialysis patients. J Bone Miner Res 22 Suppl 2: V95–99, 2007. [DOI] [PubMed] [Google Scholar]
- 315.Mohri T, Nakajima M, Takagi S, Komagata S, Yokoi T. MicroRNA regulates human vitamin D receptor. Int J Cancer 125: 1328–1333, 2009. [DOI] [PubMed] [Google Scholar]
- 316.Mondul AM, Shui IM, Yu K, Weinstein SJ, Tsilidis KK, Joshi AD, Agudo A, Berg CD, Black A, Buring JE, Chasman DI, Gaudet MM, Haiman C, Hankinson SE, Henderson BE, Hoover RN, Hunter DJ, Khaw KT, Kuhn T, Kvaskoff M, Le Marchand L, Lindstrom S, McCullough ML, Overvad K, Peeters PH, Riboli E, Ridker PM, Stram DO, Sund M, Trichopoulos D, Tumino R, Weiderpass E, Willett W, Kraft P, Ziegler RG, Albanes D. Vitamin D-associated genetic variation and risk of breast cancer in the Breast and Prostate Cancer Cohort Consortium (BPC3). Cancer Epidemiol Biomarkers Prevention 24: 627–630, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317.Mondul AM, Weinstein SJ, Horst RL, Purdue M, Albanes D. Serum vitamin D and risk of bladder cancer in the Prostate, Lung, Colorectal, and Ovarian (PLCO) Cancer Screening trial. Cancer Epidemiol Biomarkers Prevention 21: 1222–1225, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 318.Monkawa T, Yoshida T, Wakino S, Shinki T, Anazawa H, Deluca HF, Suda T, Hayashi M, Saruta T. Molecular cloning of cDNA and genomic DNA for human 25-hydroxyvitamin D3 1 alpha-hydroxylase. Biochem Biophys Res Commun 239: 527–533, 1997. [DOI] [PubMed] [Google Scholar]
- 319.Moreno J, Krishnan AV, Swami S, Nonn L, Peehl DM, Feldman D. Regulation of prostaglandin metabolism by calcitriol attenuates growth stimulation in prostate cancer cells. Cancer Res 65: 7917–7925, 2005. [DOI] [PubMed] [Google Scholar]
- 320.Morris SM, Tallquist MD, Rock CO, Cooper JA. Dual roles for the Dab2 adaptor protein in embryonic development and kidney transport. EMBO J 21: 1555–1564, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Motiwala SR, Wang TJ. Vitamin D and cardiovascular risk. Curr Hypertens Reports 14: 209–218, 2012. [DOI] [PubMed] [Google Scholar]
- 322.Nakashima T, Hayashi M, Fukunaga T, Kurata K, Oh-Hora M, Feng JQ, Bonewald LF, Kodama T, Wutz A, Wagner EF, Penninger JM, Takayanagi H. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nature Med 17: 1231–1234, 2011. [DOI] [PubMed] [Google Scholar]
- 323.Narvaez CJ, Matthews D, LaPorta E, Simmons KM, Beaudin S, Welsh J. The impact of vitamin D in breast cancer: genomics, pathways, metabolism. Front Physiol 5: 213, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Nazarova N, Golovko O, Blauer M, Tuohimaa P. Calcitriol inhibits growth response to Platelet-Derived Growth Factor-BB in human prostate cells. J Steroid Biochem Mol Biol 94: 189–196, 2005. [DOI] [PubMed] [Google Scholar]
- 325.Need AG, O'Loughlin PD, Morris HA, Coates PS, Horowitz M, Nordin BE. Vitamin D metabolites and calcium absorption in severe vitamin D deficiency. J Bone Miner Res 23: 1859–1863, 2008. [DOI] [PubMed] [Google Scholar]
- 326.Nerenz RD, Martowicz ML, Pike JW. An enhancer 20 kilobases upstream of the human receptor activator of nuclear factor-kappaB ligand gene mediates dominant activation by 1,25-dihydroxyvitamin D3. Mol Endocrinol 22: 1044–1056, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Newmark HL, Yang K, Kurihara N, Fan K, Augenlicht LH, Lipkin M. Western-style diet-induced colonic tumors and their modulation by calcium and vitamin D in C57Bl/6 mice: a preclinical model for human sporadic colon cancer. Carcinogenesis 30: 88–92, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Nguyen M, d'Alesio A, Pascussi JM, Kumar R, Griffin MD, Dong X, Guillozo H, Rizk-Rabin M, Sinding C, Bougneres P, Jehan F, Garabedian M. Vitamin D-resistant rickets and type 1 diabetes in a child with compound heterozygous mutations of the vitamin D receptor (L263R and R391S): dissociated responses of the CYP-24 and rel-B promoters to 1,25-dihydroxyvitamin D3. J Bone Miner Res 21: 886–894, 2006. [DOI] [PubMed] [Google Scholar]
- 329.Ni W, Watts SW, Ng M, Chen S, Glenn DJ, Gardner DG. Elimination of vitamin D receptor in vascular endothelial cells alters vascular function. Hypertension 64: 1290–1298, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 330.Nonn L, Peng L, Feldman D, Peehl DM. Inhibition of p38 by vitamin D reduces interleukin-6 production in normal prostate cells via mitogen-activated protein kinase phosphatase 5: implications for prostate cancer prevention by vitamin D. Cancer Res 66: 4516–4524, 2006. [DOI] [PubMed] [Google Scholar]
- 331.Norman AW. From vitamin D to hormone D: fundamentals of the vitamin D endocrine system essential for good health. Am J Clin Nutr 88: 491S–499S, 2008. [DOI] [PubMed] [Google Scholar]
- 332.Norman AW, Manchand PS, Uskokovic MR, Okamura WH, Takeuchi JA, Bishop JE, Hisatake JI, Koeffler HP, Peleg S. Characterization of a novel analogue of 1alpha,25(OH)2-vitamin D3 with two side chains: interaction with its nuclear receptor and cellular actions. J Med Chem 43: 2719–2730, 2000. [DOI] [PubMed] [Google Scholar]
- 333.Norman AW, Zhou JY, Henry HL, Uskokovic MR, Koeffler HP. Structure-function studies on analogues of 1 alpha,25-dihydroxyvitamin D3: differential effects on leukemic cell growth, differentiation, and intestinal calcium absorption. Cancer Res 50: 6857–6864, 1990. [PubMed] [Google Scholar]
- 334.Novakovic B, Sibson M, Ng HK, Manuelpillai U, Rakyan V, Down T, Beck S, Fournier T, Evain-Brion D, Dimitriadis E, Craig JM, Morley R, Saffery R. Placenta-specific methylation of the vitamin D 24-hydroxylase gene: implications for feedback autoregulation of active vitamin D levels at the fetomaternal interface. J Biol Chem 284: 14838–14848, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 335.Nuti R, Bianchi G, Brandi ML, Caudarella R, D'Erasmo E, Fiore C, Isaia GC, Luisetto G, Muratore M, Oriente P, Ortolani S. Superiority of alfacalcidol compared to vitamin D plus calcium in lumbar bone mineral density in postmenopausal osteoporosis. Rheumatol Int 26: 445–453, 2006. [DOI] [PubMed] [Google Scholar]
- 336.Nykjaer A, Dragun D, Walther D, Vorum H, Jacobsen C, Herz J, Melsen F, Christensen EI, Willnow TE. An endocytic pathway essential for renal uptake and activation of the steroid 25-(OH)vitamin D3. Cell 96: 507–515, 1999. [DOI] [PubMed] [Google Scholar]
- 337.Nykjaer A, Fyfe JC, Kozyraki R, Leheste JR, Jacobsen C, Nielsen MS, Verroust PJ, Aminoff M, de la Chapelle A, Moestrup SK, Ray R, Gliemann J, Willnow TE, Christensen EI. Cubilin dysfunction causes abnormal metabolism of the steroid hormone 25(OH)vitamin D3. Proc Natl Acad Sci USA 98: 13895–13900, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 338.O'Brien ME, Talbot D, Maclennan K, Smith IE. Inefficacy of calcipotriol in skin metastases from breast cancer. Lancet 342: 994, 1993. [DOI] [PubMed] [Google Scholar]
- 339.Ogata H, Koiwa F, Shishido K, Takahashi K, Ito H, Kinugasa E, Taguchi S. Effects of 22-oxacalcitriol and calcitriol on PTH secretion and bone mineral metabolism in a crossover trial in hemodialysis patients with secondary hyperparathyroidism. Ther Apher Dialysis 11: 202–209, 2007. [DOI] [PubMed] [Google Scholar]
- 340.Ohyama Y, Noshiro M, Eggertsen G, Gotoh O, Kato Y, Bjorkhem I, Okuda K. Structural characterization of the gene encoding rat 25-hydroxyvitamin D3 24-hydroxylase. Biochemistry 32: 76–82, 1993. [DOI] [PubMed] [Google Scholar]
- 341.Okano T, Tsugawa N, Masuda S, Takeuchi A, Kobayashi T, Nishii Y. Protein-binding properties of 22-oxa-1 alpha,25-dihydroxyvitamin D3, a synthetic analogue of 1 alpha,25-dihydroxyvitamin D3. J Nutr Sci Vitaminol 35: 529–533, 1989. [DOI] [PubMed] [Google Scholar]
- 342.Ooi JH, McDaniel KL, Weaver V, Cantorna MT. Murine CD8+ T cells but not macrophages express the vitamin D 1alpha-hydroxylase. J Nutr Biochem 25: 58–65, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 343.Ooi LL, Zhou H, Kalak R, Zheng Y, Conigrave AD, Seibel MJ, Dunstan CR. Vitamin D deficiency promotes human breast cancer growth in a murine model of bone metastasis. Cancer Res 70: 1835–1844, 2010. [DOI] [PubMed] [Google Scholar]
- 344.Ordonez-Moran P, Larriba MJ, Palmer HG, Valero RA, Barbachano A, Dunach M, de Herreros AG, Villalobos C, Berciano MT, Lafarga M, Munoz A. RhoA-ROCK and p38MAPK-MSK1 mediate vitamin D effects on gene expression, phenotype, and Wnt pathway in colon cancer cells. J Cell Biol 183: 697–710, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 345.Orlov I, Rochel N, Moras D, Klaholz BP. Structure of the full human RXR/VDR nuclear receptor heterodimer complex with its DR3 target DNA. EMBO J 31: 291–300, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 346.Padi SK, Zhang Q, Rustum YM, Morrison C, Guo B. MicroRNA-627 mediates the epigenetic mechanisms of vitamin D to suppress proliferation of human colorectal cancer cells and growth of xenograft tumors in mice. Gastroenterology 145: 437–446, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 347.Palmer HG, Gonzalez-Sancho JM, Espada J, Berciano MT, Puig I, Baulida J, Quintanilla M, Cano A, de Herreros AG, Lafarga M, Munoz A. Vitamin D3 promotes the differentiation of colon carcinoma cells by the induction of E-cadherin and the inhibition of beta-catenin signaling. J Cell Biol 154: 369–387, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348.Palmer MT, Lee YK, Maynard CL, Oliver JR, Bikle DD, Jetten AM, Weaver CT. Lineage-specific effects of 1,25-dihydroxyvitamin D3 on the development of effector CD4 T cells. J Biol Chem 286: 997–1004, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 349.Pan YZ, Gao W, Yu AM. MicroRNAs regulate CYP3A4 expression via direct and indirect targeting. Drug Metab Disposition 37: 2112–2117, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350.Panda DK, Miao D, Bolivar I, Li J, Huo R, Hendy GN, Goltzman D. Inactivation of the 25-hydroxyvitamin D 1alpha-hydroxylase and vitamin D receptor demonstrates independent and interdependent effects of calcium and vitamin D on skeletal and mineral homeostasis. J Biol Chem 279: 16754–16766, 2004. [DOI] [PubMed] [Google Scholar]
- 351.Panda DK, Miao D, Tremblay ML, Sirois J, Farookhi R, Hendy GN, Goltzman D. Targeted ablation of the 25-hydroxyvitamin D 1alpha-hydroxylase enzyme: evidence for skeletal, reproductive, and immune dysfunction. Proc Natl Acad Sci USA 98: 7498–7503, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 352.Parfitt AM, Mathews CH, Brommage R, Jarnagin K, DeLuca HF. Calcitriol but no other metabolite of vitamin D is essential for normal bone growth and development in the rat. J Clin Invest 73: 576–586, 1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 353.Park J, Rhee CM, Lau WL, Kalantar-Zadeh K. Clinical uses of 1-alpha-hydroxy-ergocalciferol. Curr Vasc Pharmacol 12: 306–312, 2014. [DOI] [PubMed] [Google Scholar]
- 354.Park MR, Lee JH, Park MS, Hwang JE, Shim HJ, Cho SH, Chung IJ, Bae WK. Suppressive effect of 19-nor-1alpha-25-dihydroxyvitamin D2 on gastric cancer cells and peritoneal metastasis model. J Korean Medical Sci 27: 1037–1043, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 355.Park WH, Seol JG, Kim ES, Jung CW, Lee CC, Binderup L, Koeffler HP, Kim BK, Lee YY. Cell cycle arrest induced by the vitamin D3 analog EB1089 in NCI-H929 myeloma cells is associated with induction of the cyclin-dependent kinase inhibitor p27. Exp Cell Res 254: 279–286, 2000. [DOI] [PubMed] [Google Scholar]
- 356.Penna G, Adorini L. 1 Alpha,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol 164: 2405–2411, 2000. [DOI] [PubMed] [Google Scholar]
- 357.Penna G, Amuchastegui S, Cossetti C, Aquilano F, Mariani R, Sanvito F, Doglioni C, Adorini L. Treatment of experimental autoimmune prostatitis in nonobese diabetic mice by the vitamin D receptor agonist elocalcitol. J Immunol 177: 8504–8511, 2006. [DOI] [PubMed] [Google Scholar]
- 358.Pervin S, Hewison M, Braga M, Tran L, Chun R, Karam A, Chaudhuri G, Norris K, Singh R. Down-regulation of vitamin D receptor in mammospheres: implications for vitamin D resistance in breast cancer and potential for combination therapy. PloS One 8: e53287, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 359.Pfeifer M, Begerow B, Minne HW, Nachtigall D, Hansen C. Effects of a short-term vitamin D3 and calcium supplementation on blood pressure and parathyroid hormone levels in elderly women. J Clin Endocrinol Metab 86: 1633–1637, 2001. [DOI] [PubMed] [Google Scholar]
- 360.Pickholtz I, Saadyan S, Keshet GI, Wang VS, Cohen R, Bouwman P, Jonkers J, Byers SW, Papa MZ, Yarden RI. Cooperation between BRCA1 and vitamin D is critical for histone acetylation of the p21waf1 promoter and growth inhibition of breast cancer cells and cancer stem-like cells. Oncotarget 5: 11827–11846, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 361.Pike JW, Lee SM, Meyer MB. Regulation of gene expression by 1,25-dihydroxyvitamin D3 in bone cells: exploiting new approaches and defining new mechanisms. Bone Key Reports 3: 482, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 362.Pike JW, Meyer MB. Fundamentals of vitamin D hormone-regulated gene expression. J Steroid Biochem Mol Biol 144: 5–11, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 363.Pike JW, Meyer MB. Regulation of mouse Cyp24a1 expression via promoter-proximal and downstream-distal enhancers highlights new concepts of 1,25-dihydroxyvitamin D3 action. Arch Biochem Biophys 523: 2–8, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 364.Pike JW, Meyer MB. The vitamin D receptor: new paradigms for the regulation of gene expression by 1,25-dihydroxyvitamin D3. Endocrinol Metab Clin N Am 39: 255–269, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 365.Pilz S, Gaksch M, Kienreich K, Grubler M, Verheyen N, Fahrleitner-Pammer A, Treiber G, Drechsler C, Hartaigh BO, Obermayer-Pietsch B, Schwetz V, Aberer F, Mader J, Scharnagl H, Meinitzer A, Lerchbaum E, Dekker JM, Zittermann A, Marz W, Tomaschitz A. Effects of vitamin D on blood pressure and cardiovascular risk factors: a randomized controlled trial. Hypertension 65: 1195–1201, 2015. [DOI] [PubMed] [Google Scholar]
- 366.Pilz S, Tomaschitz A, Ritz E, Pieber TR. Vitamin D status and arterial hypertension: a systematic review. Nature Rev Cardiol 6: 621–630, 2009. [DOI] [PubMed] [Google Scholar]
- 367.Pittas AG, Chung M, Trikalinos T, Mitri J, Brendel M, Patel K, Lichtenstein AH, Lau J, Balk EM. Systematic review: vitamin D and cardiometabolic outcomes. Ann Intern Med 152: 307–314, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 368.Plum LA, DeLuca HF. Vitamin D, disease and therapeutic opportunities. Nature Rev Drug Discovery 9: 941–955, 2010. [DOI] [PubMed] [Google Scholar]
- 369.Prentice A. Nutritional rickets around the world. J Steroid Biochem Mol Biol 136: 201–206, 2013. [DOI] [PubMed] [Google Scholar]
- 370.Priemel M, von Domarus C, Klatte TO, Kessler S, Schlie J, Meier S, Proksch N, Pastor F, Netter C, Streichert T, Puschel K, Amling M. Bone mineralization defects and vitamin D deficiency: histomorphometric analysis of iliac crest bone biopsies and circulating 25-hydroxyvitamin D in 675 patients. J Bone Miner Res 25: 305–312, 2010. [DOI] [PubMed] [Google Scholar]
- 371.Prince RL, Austin N, Devine A, Dick IM, Bruce D, Zhu K. Effects of ergocalciferol added to calcium on the risk of falls in elderly high-risk women. Arch Internal Med 168: 103–108, 2008. [DOI] [PubMed] [Google Scholar]
- 372.Quarles LD. Role of FGF23 in vitamin D and phosphate metabolism: implications in chronic kidney disease. Exp Cell Res 318: 1040–1048, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 373.Quarles LD. Skeletal secretion of FGF-23 regulates phosphate and vitamin D metabolism. Nature Rev Endocrinol 8: 276–286, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 374.Ramagopalan SV, Heger A, Berlanga AJ, Maugeri NJ, Lincoln MR, Burrell A, Handunnetthi L, Handel AE, Disanto G, Orton SM, Watson CT, Morahan JM, Giovannoni G, Ponting CP, Ebers GC, Knight JC. A ChIP-seq defined genome-wide map of vitamin D receptor binding: associations with disease and evolution. Genome Res 20: 1352–1360, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 375.Raval-Pandya M, Dhawan P, Barletta F, Christakos S. YY1 represses vitamin D receptor-mediated 25-hydroxyvitamin D3 24-hydroxylase transcription: relief of repression by CREB-binding protein. Mol Endocrinol 15: 1035–1046, 2001. [DOI] [PubMed] [Google Scholar]
- 376.Ray R, Banks M, Abuzahra H, Eddy VJ, Persons KS, Lucia MS, Lambert JR, Holick MF. Effect of dietary vitamin D and calcium on the growth of androgen-insensitive human prostate tumor in a murine model. AntiCancer Res 32: 727–731, 2012. [PMC free article] [PubMed] [Google Scholar]
- 377.Rebel H, der Spek CD, Salvatori D, van Leeuwen JP, Robanus-Maandag EC, de Gruijl FR. UV exposure inhibits intestinal tumor growth and progression to malignancy in intestine-specific Apc mutant mice kept on low vitamin D diet. Int J Cancer 136: 271–277, 2015. [DOI] [PubMed] [Google Scholar]
- 378.Reichrath J, Muller SM, Kerber A, Baum HP, Bahmer FA. Biologic effects of topical calcipotriol (MC 903) treatment in psoriatic skin. J Am Acad Dermatol 36: 19–28, 1997. [DOI] [PubMed] [Google Scholar]
- 379.Reimers LL, Crew KD, Bradshaw PT, Santella RM, Steck SE, Sirosh I, Terry MB, Hershman DL, Shane E, Cremers S, Dworakowski E, Teitelbaum SL, Neugut AI, Gammon MD. Vitamin D-related gene polymorphisms, plasma 25-hydroxyvitamin D, and breast cancer risk. Cancer Causes Control 26: 187–203, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 380.Replogle RA, Li Q, Wang L, Zhang M, Fleet JC. Gene-by-diet interactions influence calcium absorption and bone density in mice. J Bone Miner Res 29: 657–665, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 381.Ritter CS, Brown AJ. Suppression of PTH by the vitamin D analog eldecalcitol is modulated by its high affinity for the serum vitamin D-binding protein and resistance to metabolism. J Cell Biochem 112: 1348–1352, 2011. [DOI] [PubMed] [Google Scholar]
- 382.Robinson CJ, Spanos E, James MF, Pike JW, Haussler MR, Makeen AM, Hillyard CJ, MacIntyre I. Role of prolactin in vitamin D metabolism and calcium absorption during lactation in the rat. J Endocrinol 94: 443–453, 1982. [DOI] [PubMed] [Google Scholar]
- 383.Rochel N, Ciesielski F, Godet J, Moman E, Roessle M, Peluso-Iltis C, Moulin M, Haertlein M, Callow P, Mely Y, Svergun DI, Moras D. Common architecture of nuclear receptor heterodimers on DNA direct repeat elements with different spacings. Nature Struct Mol Biol 18: 564–570, 2011. [DOI] [PubMed] [Google Scholar]
- 384.Rochel N, Wurtz JM, Mitschler A, Klaholz B, Moras D. The crystal structure of the nuclear receptor for vitamin D bound to its natural ligand. Mol Cell 5: 173–179, 2000. [DOI] [PubMed] [Google Scholar]
- 385.Rosli SN, Shintani T, Toratani S, Usui E, Okamoto T. 1Alpha, 25(OH)2D3 inhibits FGF-2 release from oral squamous cell carcinoma cells through down-regulation of HBp17/FGFBP-1. In Vitro Cell Dev Biol 50: 802–806, 2014. [DOI] [PubMed] [Google Scholar]
- 386.Ross AC, Manson JE, Abrams SA, Aloia JF, Brannon PM, Clinton SK, Durazo-Arvizu RA, Gallagher JC, Gallo RL, Jones G, Kovacs CS, Mayne ST, Rosen CJ, Shapses SA. The 2011 report on dietary reference intakes for calcium and vitamin D from the Institute of Medicine: what clinicians need to know. J Clin Endocrinol Metab 96: 53–58, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 387.Russell J, Ashok S, Koszewski NJ. Vitamin D receptor interactions with the rat parathyroid hormone gene: synergistic effects between two negative vitamin D response elements. J Bone Miner Res 14: 1828–1837, 1999. [DOI] [PubMed] [Google Scholar]
- 388.Ryz NR, Patterson SJ, Zhang Y, Ma C, Huang T, Bhinder G, Wu X, Chan J, Glesby A, Sham HP, Dutz JP, Levings MK, Jacobson K, Vallance BA. Active vitamin D (1,25-dihydroxyvitamin D3) increases host susceptibility to Citrobacter rodentium by suppressing mucosal Th17 responses. Am J Physiol Gastrointest Liver Physiol 303: G1299–G1311, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 389.Sabbagh Y, Carpenter TO, Demay MB. Hypophosphatemia leads to rickets by impairing caspase-mediated apoptosis of hypertrophic chondrocytes. Proc Natl Acad Sci USA 102: 9637–9642, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 390.Safadi FF, Thornton P, Magiera H, Hollis BW, Gentile M, Haddad JG, Liebhaber SA, Cooke NE. Osteopathy and resistance to vitamin D toxicity in mice null for vitamin D binding protein. J Clin Invest 103: 239–251, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 391.Santagata S, Thakkar A, Ergonul A, Wang B, Woo T, Hu R, Harrell JC, McNamara G, Schwede M, Culhane AC, Kindelberger D, Rodig S, Richardson A, Schnitt SJ, Tamimi RM, Ince TA. Taxonomy of breast cancer based on normal cell phenotype predicts outcome. J Clin Invest 124: 859–870, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 392.Santucci KL, Snyder KK, Baust JM, Van Buskirk RG, Mouraviev V, Polascik TJ, Gage AA, Baust JG. Use of 1,25alpha dihydroxyvitamin D3 as a cryosensitizing agent in a murine prostate cancer model. Prostate Cancer Prostatic Dis 14: 97–104, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 393.Sato-Deguchi E, Imafuku S, Chou B, Ishii K, Hiromatsu K, Nakayama J. Topical vitamin D3 analogues induce thymic stromal lymphopoietin and cathelicidin in psoriatic skin lesions. Br J Dermatol 167: 77–84, 2012. [DOI] [PubMed] [Google Scholar]
- 394.Schafer A, Emmert S, Kruppa J, Schubert S, Tzvetkov M, Mossner R, Reich K, Berking C, Volkenandt M, Pfohler C, Schon MP, Vogt T, Konig IR, Reichrath J. No association of vitamin D metabolism-related polymorphisms and melanoma risk as well as melanoma prognosis: a case-control study. Arch Dermatol Res 304: 353–361, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 395.Schafer AL, Weaver CM, Black DM, Wheeler AL, Chang H, Szefc GV, Stewart L, Rogers SJ, Carter JT, Posselt AM, Shoback DM, Sellmeyer DE. Intestinal calcium absorption decreases dramatically after gastric bypass surgery despite optimization of vitamin D status. J Bone Miner Res. 30: 1377–1385, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 396.Schlingmann KP, Kaufmann M, Weber S, Irwin A, Goos C, John U, Misselwitz J, Klaus G, Kuwertz-Broking E, Fehrenbach H, Wingen AM, Guran T, Hoenderop JG, Bindels RJ, Prosser DE, Jones G, Konrad M. Mutations in CYP24A1 and idiopathic infantile hypercalcemia. N Engl J Med 365: 410–421, 2011. [DOI] [PubMed] [Google Scholar]
- 397.Schon MP, Boehncke WH. Psoriasis. N Engl J Med 352: 1899–1912, 2005. [DOI] [PubMed] [Google Scholar]
- 398.Schottker B, Jorde R, Peasey A, Thorand B, Jansen EH, Groot L, Streppel M, Gardiner J, Ordonez-Mena JM, Perna L, Wilsgaard T, Rathmann W, Feskens E, Kampman E, Siganos G, Njolstad I, Mathiesen EB, Kubinova R, Pajak A, Topor-Madry R, Tamosiunas A, Hughes M, Kee F, Bobak M, Trichopoulou A, Boffetta P, Brenner H, Consortium on Ageing: Network of Cohorts in Europe and the United States. Vitamin D and mortality: meta-analysis of individual participant data from a large consortium of cohort studies from Europe and the United States. BMJ 348: g3656, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 399.Schwartz GG, Hall MC, Stindt D, Patton S, Lovato J, Torti FM. Phase I/II study of 19-nor-1alpha-25-dihydroxyvitamin D2 (paricalcitol) in advanced, androgen-insensitive prostate cancer. Clin Cancer Res 11: 8680–8685, 2005. [DOI] [PubMed] [Google Scholar]
- 400.Scragg R. Seasonality of cardiovascular disease mortality and the possible protective effect of ultra-violet radiation. Int J Epidemiol 10: 337–341, 1981. [DOI] [PubMed] [Google Scholar]
- 401.Scragg R, Sowers M, Bell C. Serum 25-hydroxyvitamin D, ethnicity, and blood pressure in the Third National Health and Nutrition Examination Survey. Am J Hypertens 20: 713–719, 2007. [DOI] [PubMed] [Google Scholar]
- 402.Seth-Vollenweider T, Joshi S, Dhawan P, Sif S, Christakos S. Novel mechanism of negative regulation of 1,25-dihydroxyvitamin D3-induced 25-hydroxyvitamin D3 24-hydroxylase (Cyp24a1) transcription: epigenetic modification involving cross-talk between protein-arginine methyltransferase 5 and the SWI/SNF complex. J Biol Chem 289: 33958–33970, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 403.Shaffer PL, Gewirth DT. Structural basis of VDR-DNA interactions on direct repeat response elements. EMBO J 21: 2242–2252, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 404.Shaffer PL, McDonnell DP, Gewirth DT. Characterization of transcriptional activation and DNA-binding functions in the hinge region of the vitamin D receptor. Biochemistry 44: 2678–2685, 2005. [DOI] [PubMed] [Google Scholar]
- 405.Shen Q, Christakos S. The vitamin D receptor, Runx2, and the Notch signaling pathway cooperate in the transcriptional regulation of osteopontin. J Biol Chem 280: 40589–40598, 2005. [DOI] [PubMed] [Google Scholar]
- 406.Sherman MH, Yu RT, Engle DD, Ding N, Atkins AR, Tiriac H, Collisson EA, Connor F, Van Dyke T, Kozlov S, Martin P, Tseng TW, Dawson DW, Donahue TR, Masamune A, Shimosegawa T, Apte MV, Wilson JS, Ng B, Lau SL, Gunton JE, Wahl GM, Hunter T, Drebin JA, O'Dwyer PJ, Liddle C, Tuveson DA, Downes M, Evans RM. Vitamin D receptor-mediated stromal reprogramming suppresses pancreatitis and enhances pancreatic cancer therapy. Cell 159: 80–93, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 407.Shimada T, Kakitani M, Yamazaki Y, Hasegawa H, Takeuchi Y, Fujita T, Fukumoto S, Tomizuka K, Yamashita T. Targeted ablation of Fgf23 demonstrates an essential physiological role of FGF23 in phosphate and vitamin D metabolism. J Clin Invest 113: 561–568, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 408.Shimada T, Mizutani S, Muto T, Yoneya T, Hino R, Takeda S, Takeuchi Y, Fujita T, Fukumoto S, Yamashita T. Cloning and characterization of FGF23 as a causative factor of tumor-induced osteomalacia. Proc Natl Acad Sci USA 98: 6500–6505, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 409.Shinki T, Jin CH, Nishimura A, Nagai Y, Ohyama Y, Noshiro M, Okuda K, Suda T. Parathyroid hormone inhibits 25-hydroxyvitamin D3-24-hydroxylase mRNA expression stimulated by 1 alpha,25-dihydroxyvitamin D3 in rat kidney but not in intestine. J Biol Chem 267: 13757–13762, 1992. [PubMed] [Google Scholar]
- 410.Shinki T, Shimada H, Wakino S, Anazawa H, Hayashi M, Saruta T, DeLuca HF, Suda T. Cloning and expression of rat 25-hydroxyvitamin D3-1alpha-hydroxylase cDNA. Proc Natl Acad Sci USA 94: 12920–12925, 1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 411.Shinki T, Ueno Y, DeLuca HF, Suda T. Calcitonin is a major regulator for the expression of renal 25-hydroxyvitamin D3-1alpha-hydroxylase gene in normocalcemic rats. Proc Natl Acad Sci USA 96: 8253–8258, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 412.Simmons KM, Beaudin SG, Narvaez CJ, Welsh J. Gene signatures of 1,25-dihydroxyvitamin D exposure in normal and transformed mammary cells. J Cell Biochem. 116: 1693–1671, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 413.Singarapu KK, Zhu J, Tonelli M, Rao H, Assadi-Porter FM, Westler WM, DeLuca HF, Markley JL. Ligand-specific structural changes in the vitamin D receptor in solution. Biochemistry 50: 11025–11033, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 414.Singh PK, Long MD, Battaglia S, Hu Q, Liu S, Sucheston-Campbell LE, Campbell MJ. VDR regulation of microRNA differs across prostate cell models suggesting extremely flexible control of transcription. Epigenetics 10: 40–49, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 415.Snijder MB, Lips P, Seidell JC, Visser M, Deeg DJ, Dekker JM, van Dam RM. Vitamin D status and parathyroid hormone levels in relation to blood pressure: a population-based study in older men and women. J Internal Med 261: 558–565, 2007. [DOI] [PubMed] [Google Scholar]
- 416.So JY, Lee HJ, Smolarek AK, Paul S, Wang CX, Maehr H, Uskokovic M, Zheng X, Conney AH, Cai L, Liu F, Suh N. A novel Gemini vitamin D analog represses the expression of a stem cell marker CD44 in breast cancer. Mol Pharmacol 79: 360–367, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 417.So JY, Wahler J, Das Gupta S, Salerno DM, Maehr H, Uskokovic M, Suh N. HES1-mediated inhibition of Notch1 signaling by a Gemini vitamin D analog leads to decreased CD44(+)/CD24(−/low) tumor-initiating subpopulation in basal-like breast cancer. J Steroid Biochem Mol Biol 148: 111–121, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 418.Soleymani T, Hung T, Soung J. The role of vitamin D in psoriasis: a review. Int J Dermatol 54: 383–392, 2015. [DOI] [PubMed] [Google Scholar]
- 419.Song M, Nishihara R, Wang M, Chan AT, Qian ZR, Inamura K, Zhang X, Ng K, Kim SA, Mima K, Sukawa Y, Nosho K, Fuchs CS, Giovannucci EL, Wu K, Ogino S. Plasma 25-hydroxyvitamin D and colorectal cancer risk according to tumour immunity status. Gut. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 420.Song Y, Peng X, Porta A, Takanaga H, Peng JB, Hediger MA, Fleet JC, Christakos S. Calcium transporter 1 and epithelial calcium channel messenger ribonucleic acid are differentially regulated by 1,25 dihydroxyvitamin D3 in the intestine and kidney of mice. Endocrinology 144: 3885–3894, 2003. [DOI] [PubMed] [Google Scholar]
- 421.Spach KM, Nashold FE, Dittel BN, Hayes CE. IL-10 signaling is essential for 1,25-dihydroxyvitamin D3-mediated inhibition of experimental autoimmune encephalomyelitis. J Immunol 177: 6030–6037, 2006. [DOI] [PubMed] [Google Scholar]
- 422.Sprague SM, Llach F, Amdahl M, Taccetta C, Batlle D. Paricalcitol versus calcitriol in the treatment of secondary hyperparathyroidism. Kidney Int 63: 1483–1490, 2003. [DOI] [PubMed] [Google Scholar]
- 423.St-Arnaud R. CYP24A1-deficient mice as a tool to uncover a biological activity for vitamin D metabolites hydroxylated at position 24. J Steroid Biochem Mol Biol 121: 254–256, 2010. [DOI] [PubMed] [Google Scholar]
- 424.St-Arnaud R, Arabian A, Travers R, Barletta F, Raval-Pandya M, Chapin K, Depovere J, Mathieu C, Christakos S, Demay MB, Glorieux FH. Deficient mineralization of intramembranous bone in vitamin D-24-hydroxylase-ablated mice is due to elevated 1,25-dihydroxyvitamin D and not to the absence of 24,25-dihydroxyvitamin D. Endocrinology 141: 2658–2666, 2000. [DOI] [PubMed] [Google Scholar]
- 425.St-Arnaud R, Messerlian S, Moir JM, Omdahl JL, Glorieux FH. The 25-hydroxyvitamin D 1-alpha-hydroxylase gene maps to the pseudovitamin D-deficiency rickets (PDDR) disease locus. J Bone Miner Res 12: 1552–1559, 1997. [DOI] [PubMed] [Google Scholar]
- 426.Stashi E, York B, O'Malley BW. Steroid receptor coactivators: servants and masters for control of systems metabolism. Trends Endocrinol Metab 25: 337–347, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 427.Stevenson JC, Hillyard CJ, MacIntyre I, Cooper H, Whitehead MI. A physiological role for calcitonin: protection of the maternal skeleton. Lancet 2: 769–770, 1979. [DOI] [PubMed] [Google Scholar]
- 428.Stio M, Martinesi M, Simoni A, Zuegel U, Steinmeyer A, Santi R, Treves C, Nesi G. The novel vitamin D analog ZK191784 inhibits prostate cancer cell invasion. AntiCancer Res 31: 4091–4098, 2011. [PubMed] [Google Scholar]
- 429.Stoffels K, Overbergh L, Bouillon R, Mathieu C. Immune regulation of 1alpha-hydroxylase in murine peritoneal macrophages: unravelling the IFNgamma pathway. J Steroid Biochem Mol Biol 103: 567–571, 2007. [DOI] [PubMed] [Google Scholar]
- 430.Strushkevich N, Usanov SA, Plotnikov AN, Jones G, Park HW. Structural analysis of CYP2R1 in complex with vitamin D3. J Mol Biol 380: 95–106, 2008. [DOI] [PubMed] [Google Scholar]
- 431.Suda T, Takahashi N, Udagawa N, Jimi E, Gillespie MT, Martin TJ. Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr Rev 20: 345–357, 1999. [DOI] [PubMed] [Google Scholar]
- 432.Sung V, Feldman D. 1,25-Dihydroxyvitamin D3 decreases human prostate cancer cell adhesion and migration. Mol Cell Endocrinol 164: 133–143, 2000. [DOI] [PubMed] [Google Scholar]
- 433.Swami S, Krishnan AV, Wang JY, Jensen K, Horst R, Albertelli MA, Feldman D. Dietary vitamin D3 and 1,25-dihydroxyvitamin D3 (calcitriol) exhibit equivalent anticancer activity in mouse xenograft models of breast and prostate cancer. Endocrinology 153: 2576–2587, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 434.Syal SK, Kapoor A, Bhatia E, Sinha A, Kumar S, Tewari S, Garg N, Goel PK. Vitamin D deficiency, coronary artery disease, and endothelial dysfunction: observations from a coronary angiographic study in Indian patients. J Invasive Cardiol 24: 385–389, 2012. [PubMed] [Google Scholar]
- 435.Takeda S, Yoshizawa T, Nagai Y, Yamato H, Fukumoto S, Sekine K, Kato S, Matsumoto T, Fujita T. Stimulation of osteoclast formation by 1,25-dihydroxyvitamin D requires its binding to vitamin D receptor (VDR) in osteoblastic cells: studies using VDR knockout mice. Endocrinology 140: 1005–1008, 1999. [DOI] [PubMed] [Google Scholar]
- 436.Takeyama K, Kitanaka S, Sato T, Kobori M, Yanagisawa J, Kato S. 25-Hydroxyvitamin D3 1alpha-hydroxylase and vitamin D synthesis. Science 277: 1827–1830, 1997. [DOI] [PubMed] [Google Scholar]
- 437.Takiishi T, Ding L, Baeke F, Spagnuolo I, Sebastiani G, Laureys J, Verstuyf A, Carmeliet G, Dotta F, Van Belle TL, Gysemans CA, Mathieu C. Dietary supplementation with high doses of regular vitamin D3 safely reduces diabetes incidence in NOD mice when given early and long term. Diabetes 63: 2026–2036, 2014. [DOI] [PubMed] [Google Scholar]
- 438.Tamura S, Ueki K, Mashimo K, Tsukada Y, Naitoh M, Abe Y, Kawai H, Tsuchida A, Wakamatsu R, Nojima Y. Comparison of the efficacy of an oral calcitriol pulse or intravenous 22-oxacalcitriol therapies in chronic hemodialysis patients. Clin Exp Nephrol 9: 238–243, 2005. [DOI] [PubMed] [Google Scholar]
- 439.Tang BM, Eslick GD, Nowson C, Smith C, Bensoussan A. Use of calcium or calcium in combination with vitamin D supplementation to prevent fractures and bone loss in people aged 50 years and older: a meta-analysis. Lancet 370: 657–666, 2007. [DOI] [PubMed] [Google Scholar]
- 440.Tangpricha V, Spina C, Yao M, Chen TC, Wolfe MM, Holick MF. Vitamin D deficiency enhances the growth of MC-26 colon cancer xenografts in Balb/c mice. J Nutr 135: 2350–2354, 2005. [DOI] [PubMed] [Google Scholar]
- 441.Tavera-Mendoza L, Wang TT, Lallemant B, Zhang R, Nagai Y, Bourdeau V, Ramirez-Calderon M, Desbarats J, Mader S, White JH. Convergence of vitamin D and retinoic acid signalling at a common hormone response element. EMBO Reports 7: 180–185, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 442.Tebben PJ, Milliner DS, Horst RL, Harris PC, Singh RJ, Wu Y, Foreman JW, Chelminski PR, Kumar R. Hypercalcemia, hypercalciuria, and elevated calcitriol concentrations with autosomal dominant transmission due to CYP24A1 mutations: effects of ketoconazole therapy. J Clin Endocrinol Metab 97: E423–427, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 443.Thadhani R, Appelbaum E, Pritchett Y, Chang Y, Wenger J, Tamez H, Bhan I, Agarwal R, Zoccali C, Wanner C, Lloyd-Jones D, Cannata J, Thompson BT, Andress D, Zhang W, Packham D, Singh B, Zehnder D, Shah A, Pachika A, Manning WJ, Solomon SD. Vitamin D therapy and cardiac structure and function in patients with chronic kidney disease: the PRIMO randomized controlled trial. JAMA 307: 674–684, 2012. [DOI] [PubMed] [Google Scholar]
- 444.Thill M, Becker S, Fischer D, Cordes T, Hornemann A, Diedrich K, Salehin D, Friedrich M. Expression of prostaglandin metabolising enzymes COX-2 and 15-PGDH and VDR in human granulosa cells. AntiCancer Res 29: 3611–3618, 2009. [PubMed] [Google Scholar]
- 445.Ting HJ, Messing J, Yasmin-Karim S, Lee YF. Identification of microRNA-98 as a therapeutic target inhibiting prostate cancer growth and a biomarker induced by vitamin D. J Biol Chem 288: 1–9, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 446.Tiosano D, Hadad S, Chen Z, Nemirovsky A, Gepstein V, Militianu D, Weisman Y, Abrams SA. Calcium absorption, kinetics, bone density, and bone structure in patients with hereditary vitamin D-resistant rickets. J Clin Endocrinol Metab 96: 3701–3709, 2011. [DOI] [PubMed] [Google Scholar]
- 447.Tocchini-Valentini G, Rochel N, Wurtz JM, Mitschler A, Moras D. Crystal structures of the vitamin D receptor complexed to superagonist 20-epi ligands. Proc Natl Acad Sci USA 98: 5491–5496, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 448.Tocchini-Valentini G, Rochel N, Wurtz JM, Moras D. Crystal structures of the vitamin D nuclear receptor liganded with the vitamin D side chain analogues calcipotriol and seocalcitol, receptor agonists of clinical importance. Insights into a structural basis for the switching of calcipotriol to a receptor antagonist by further side chain modification. J Med Chem 47: 1956–1961, 2004. [DOI] [PubMed] [Google Scholar]
- 449.Tokar EJ, Webber MM. Cholecalciferol (vitamin D3) inhibits growth and invasion by up-regulating nuclear receptors and 25-hydroxylase (CYP27A1) in human prostate cancer cells. Clin Exp Metastasis 22: 275–284, 2005. [DOI] [PubMed] [Google Scholar]
- 450.Tomaschitz A, Pilz S, Ritz E, Grammer T, Drechsler C, Boehm BO, Marz W. Independent association between 1,25-dihydroxyvitamin D, 25-hydroxyvitamin D and the renin-angiotensin system: The Ludwigshafen Risk and Cardiovascular Health (LURIC) study. Clin Chim Acta 411: 1354–1360, 2010. [DOI] [PubMed] [Google Scholar]
- 451.Tremezaygues L, Reichrath J. Vitamin D analogs in the treatment of psoriasis: where are we standing and where will we be going? Dermato-Endocrinol 3: 180–186, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 452.Trivedi DP, Doll R, Khaw KT. Effect of four monthly oral vitamin D3 (cholecalciferol) supplementation on fractures and mortality in men and women living in the community: randomised double blind controlled trial. BMJ 326: 469, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 453.Trujillo G, Habiel DM, Ge L, Ramadass M, Cooke NE, Kew RR. Neutrophil recruitment to the lung in both C5a- and CXCL1-induced alveolitis is impaired in vitamin D-binding protein-deficient mice. J Immunol 191: 848–856, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 454.Tsai KS, Heath H, Kumar R 3rd, Riggs BL. Impaired vitamin D metabolism with aging in women. Possible role in pathogenesis of senile osteoporosis. J Clin Invest 73: 1668–1672, 1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 455.Tse AK, Zhu GY, Wan CK, Shen XL, Yu ZL, Fong WF. 1alpha,25-Dihydroxyvitamin D3 inhibits transcriptional potential of nuclear factor kappa B in breast cancer cells. Mol Immunol 47: 1728–1738, 2010. [DOI] [PubMed] [Google Scholar]
- 456.Unger WW, Laban S, Kleijwegt FS, van der Slik AR, Roep BO. Induction of Treg by monocyte-derived DC modulated by vitamin D3 or dexamethasone: differential role for PD-L1. Eur J Immunol 39: 3147–3159, 2009. [DOI] [PubMed] [Google Scholar]
- 457.Van Belle TL, Gysemans C, Mathieu C. Vitamin D in autoimmune, infectious and allergic diseases: a vital player? Best Practice Res Clin Endocrinol Metab 25: 617–632, 2011. [DOI] [PubMed] [Google Scholar]
- 458.Van Belle TL, Vanherwegen AS, Feyaerts D, De Clercq P, Verstuyf A, Korf H, Gysemans C, Mathieu C. 1,25-Dihydroxyvitamin D3 and its analog TX527 promote a stable regulatory T cell phenotype in T cells from type 1 diabetes patients. PloS One 9: e109194, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 459.Van Cromphaut SJ, Dewerchin M, Hoenderop JG, Stockmans I, Van Herck E, Kato S, Bindels RJ, Collen D, Carmeliet P, Bouillon R, Carmeliet G. Duodenal calcium absorption in vitamin D receptor-knockout mice: functional and molecular aspects. Proc Natl Acad Sci USA 98: 13324–13329, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 460.Van Cromphaut SJ, Stockmans I, Torrekens S, Van Herck E, Carmeliet G, Bouillon R. Duodenal calcium absorption in dexamethasone-treated mice: functional and molecular aspects. Arch Biochem Biophys 460: 300–305, 2007. [DOI] [PubMed] [Google Scholar]
- 461.Van de Kerkhof PC, Berth-Jones J, Griffiths CE, Harrison PV, Honigsmann H, Marks R, Roelandts R, Schopf E, Trompke C. Long-term efficacy and safety of tacalcitol ointment in patients with chronic plaque psoriasis. Br J Dermatol 146: 414–422, 2002. [DOI] [PubMed] [Google Scholar]
- 462.Van Halteren AG, Tysma OM, van Etten E, Mathieu C, Roep BO. 1alpha,25-dihydroxyvitamin D3 or analogue treated dendritic cells modulate human autoreactive T cells via the selective induction of apoptosis. J Autoimmun 23: 233–239, 2004. [DOI] [PubMed] [Google Scholar]
- 463.Van Halteren AG, van Etten E, de Jong EC, Bouillon R, Roep BO, Mathieu C. Redirection of human autoreactive T-cells upon interaction with dendritic cells modulated by TX527, an analog of 1,25 dihydroxyvitamin D3. Diabetes 51: 2119–2125, 2002. [DOI] [PubMed] [Google Scholar]
- 464.Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 324: 1029–1033, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 465.Vanhooke JL, Benning MM, Bauer CB, Pike JW, DeLuca HF. Molecular structure of the rat vitamin D receptor ligand binding domain complexed with 2-carbon-substituted vitamin D3 hormone analogues and a LXXLL-containing coactivator peptide. Biochemistry 43: 4101–4110, 2004. [DOI] [PubMed] [Google Scholar]
- 466.Vanoirbeek E, Eelen G, Verlinden L, Carmeliet G, Mathieu C, Bouillon R, O'Connor R, Xiao G, Verstuyf A. PDLIM2 expression is driven by vitamin D and is involved in the pro-adhesion, and anti-migration and -invasion activity of vitamin D. Oncogene 33: 1904–1911, 2014. [DOI] [PubMed] [Google Scholar]
- 467.Verlinden L, Eelen G, Van Hellemont R, Engelen K, Beullens I, Van Camp M, Marchal K, Mathieu C, Bouillon R, Verstuyf A. 1alpha,25-Dihydroxyvitamin D3-induced down-regulation of the checkpoint proteins, Chk1 and Claspin, is mediated by the pocket proteins p107 and p130. J Steroid Biochem Mol Biol 103: 411–415, 2007. [DOI] [PubMed] [Google Scholar]
- 468.Villagra A, Cruzat F, Carvallo L, Paredes R, Olate J, van Wijnen AJ, Stein GS, Lian JB, Stein JL, Imbalzano AN, Montecino M. Chromatin remodeling and transcriptional activity of the bone-specific osteocalcin gene require CCAAT/enhancer-binding protein beta-dependent recruitment of SWI/SNF activity. J Biol Chem 281: 22695–22706, 2006. [DOI] [PubMed] [Google Scholar]
- 469.Wade WN, Willingham MC, Koumenis C, Cramer SD. p27Kip1 is essential for the antiproliferative action of 1,25-dihydroxyvitamin D3 in primary, but not immortalized, mouse embryonic fibroblasts. J Biol Chem 277: 37301–37306, 2002. [DOI] [PubMed] [Google Scholar]
- 469a.Wagner CA, Hernando N, Forster IC, Biber J. The SLC34 family of sodium-dependent phosphate transporters. Pflügers Arch 466: 139–153, 2014. [DOI] [PubMed] [Google Scholar]
- 470.Wagner N, Wagner KD, Schley G, Badiali L, Theres H, Scholz H. 1,25-Dihydroxyvitamin D3-induced apoptosis of retinoblastoma cells is associated with reciprocal changes of Bcl-2 and bax. Exp Eye Res 77: 1–9, 2003. [DOI] [PubMed] [Google Scholar]
- 471.Wahler J, So JY, Cheng LC, Maehr H, Uskokovic M, Suh N. Vitamin D compounds reduce mammosphere formation and decrease expression of putative stem cell markers in breast cancer. J Steroid Biochem Mol Biol 148: 148–155, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 472.Wang D, Velez de-la-Paz OI, Zhai JX, Liu DW. Serum 25-hydroxyvitamin D and breast cancer risk: a meta-analysis of prospective studies. Tumour Biol 34: 3509–3517, 2013. [DOI] [PubMed] [Google Scholar]
- 473.Wang L, Song Y, Manson JE, Pilz S, Marz W, Michaelsson K, Lundqvist A, Jassal SK, Barrett-Connor E, Zhang C, Eaton CB, May HT, Anderson JL, Sesso HD. Circulating 25-hydroxy-vitamin D and risk of cardiovascular disease: a meta-analysis of prospective studies. Circulation Cardiovasc Quality Outcomes 5: 819–829, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 474.Wang TJ, Pencina MJ, Booth SL, Jacques PF, Ingelsson E, Lanier K, Benjamin EJ, D'Agostino RB, Wolf M, Vasan RS. Vitamin D deficiency and risk of cardiovascular disease. Circulation 117: 503–511, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 475.Wang TJ, Zhang F, Richards JB, Kestenbaum B, van Meurs JB, Berry D, Kiel DP, Streeten EA, Ohlsson C, Koller DL, Peltonen L, Cooper JD, O'Reilly PF, Houston DK, Glazer NL, Vandenput L, Peacock M, Shi J, Rivadeneira F, McCarthy MI, Anneli P, de Boer IH, Mangino M, Kato B, Smyth DJ, Booth SL, Jacques PF, Burke GL, Goodarzi M, Cheung CL, Wolf M, Rice K, Goltzman D, Hidiroglou N, Ladouceur M, Wareham NJ, Hocking LJ, Hart D, Arden NK, Cooper C, Malik S, Fraser WD, Hartikainen AL, Zhai G, Macdonald HM, Forouhi NG, Loos RJ, Reid DM, Hakim A, Dennison E, Liu Y, Power C, Stevens HE, Jaana L, Vasan RS, Soranzo N, Bojunga J, Psaty BM, Lorentzon M, Foroud T, Harris TB, Hofman A, Jansson JO, Cauley JA, Uitterlinden AG, Gibson Q, Jarvelin MR, Karasik D, Siscovick DS, Econs MJ, Kritchevsky SB, Florez JC, Todd JA, Dupuis J, Hypponen E, Spector TD. Common genetic determinants of vitamin D insufficiency: a genome-wide association study. Lancet 376: 180–188, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 476.Wang X, Gocek E, Liu CG, Studzinski GP. MicroRNAs181 regulate the expression of p27Kip1 in human myeloid leukemia cells induced to differentiate by 1,25-dihydroxyvitamin D3. Cell Cycle 8: 736–741, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 477.Wang X, Studzinski GP. Antiapoptotic action of 1,25-dihydroxyvitamin D3 is associated with increased mitochondrial MCL-1 and RAF-1 proteins and reduced release of cytochrome c. Exp Cell Res 235: 210–217, 1997. [DOI] [PubMed] [Google Scholar]
- 478.Wang Y, Zhu J, DeLuca HF. Identification of the vitamin D receptor in osteoblasts and chondrocytes but not osteoclasts in mouse bone. J Bone Miner Res 29: 685–692, 2014. [DOI] [PubMed] [Google Scholar]
- 479.Washington MN, Kim JS, Weigel NL. 1Alpha,25-dihydroxyvitamin D3 inhibits C4-2 prostate cancer cell growth via a retinoblastoma protein (Rb)-independent G1 arrest. The Prostate 71: 98–110, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 480.Washington MN, Weigel NL. 1α,25-Dihydroxyvitamin D3 inhibits growth of VCaP prostate cancer cells despite inducing the growth-promoting TMPRSS2:ERG gene fusion. Endocrinology 151: 1409–1417, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 481.Wasserman RH. Vitamin D and the dual processes of intestinal calcium absorption. J Nutr 134: 3137–3139, 2004. [DOI] [PubMed] [Google Scholar]
- 482.Waters WR, Palmer MV, Nonnecke BJ, Whipple DL, Horst RL. Mycobacterium bovis infection of vitamin D-deficient NOS2−/− mice. Microbial Pathogenesis 36: 11–17, 2004. [DOI] [PubMed] [Google Scholar]
- 483.Webb AR, Kline L, Holick MF. Influence of season and latitude on the cutaneous synthesis of vitamin D3: exposure to winter sunlight in Boston and Edmonton will not promote vitamin D3 synthesis in human skin. J Clin Endocrinol Metab 67: 373–378, 1988. [DOI] [PubMed] [Google Scholar]
- 484.Weber TJ, Liu S, Indridason OS, Quarles LD. Serum FGF23 levels in normal and disordered phosphorus homeostasis. J Bone Miner Res 18: 1227–1234, 2003. [DOI] [PubMed] [Google Scholar]
- 485.Wesseling-Perry K, Pereira RC, Sahney S, Gales B, Wang HJ, Elashoff R, Juppner H, Salusky IB. Calcitriol and doxercalciferol are equivalent in controlling bone turnover, suppressing parathyroid hormone, and increasing fibroblast growth factor-23 in secondary hyperparathyroidism. Kidney Int 79: 112–119, 2011. [DOI] [PubMed] [Google Scholar]
- 486.White JH. Vitamin D as an inducer of cathelicidin antimicrobial peptide expression: past, present and future. J Steroid Biochem Mol Biol 121: 234–238, 2010. [DOI] [PubMed] [Google Scholar]
- 487.White KE, Carn G, Lorenz-Depiereux B, Benet-Pages A, Strom TM, Econs MJ. Autosomal-dominant hypophosphatemic rickets (ADHR) mutations stabilize FGF-23. Kidney Int 60: 2079–2086, 2001. [DOI] [PubMed] [Google Scholar]
- 488.Whitlatch LW, Young MV, Schwartz GG, Flanagan JN, Burnstein KL, Lokeshwar BL, Rich ES, Holick MF, Chen TC. 25-Hydroxyvitamin D-1alpha-hydroxylase activity is diminished in human prostate cancer cells and is enhanced by gene transfer. J Steroid Biochem Mol Biol 81: 135–140, 2002. [DOI] [PubMed] [Google Scholar]
- 489.Williams KB, DeLuca HF. Characterization of intestinal phosphate absorption using a novel in vivo method. Am J Physiol Endocrinol Metab 292: E1917–E1921, 2007. [DOI] [PubMed] [Google Scholar]
- 490.Willnow TE, Nykjaer A. Cellular uptake of steroid carrier proteins–mechanisms and implications. Mol Cell Endocrinol 316: 93–102, 2010. [DOI] [PubMed] [Google Scholar]
- 491.Wilson HD, Schedl HP. Rat large bowel maintains calcium transport after small bowel resection. Digestion 26: 224–230, 1983. [DOI] [PubMed] [Google Scholar]
- 492.Witham MD, Price RJ, Struthers AD, Donnan PT, Messow CM, Ford I, McMurdo ME. Cholecalciferol treatment to reduce blood pressure in older patients with isolated systolic hypertension: the VitDISH randomized controlled trial. JAMA Internal Med 173: 1672–1679, 2013. [DOI] [PubMed] [Google Scholar]
- 493.Wood RJ, Fleet JC, Cashman K, Bruns ME, Deluca HF. Intestinal calcium absorption in the aged rat: evidence of intestinal resistance to 1,25(OH)2 vitamin D. Endocrinology 139: 3843–3848, 1998. [DOI] [PubMed] [Google Scholar]
- 494.Wu S, Zhang YG, Lu R, Xia Y, Zhou D, Petrof EO, Claud EC, Chen D, Chang EB, Carmeliet G, Sun J. Intestinal epithelial vitamin D receptor deletion leads to defective autophagy in colitis. Gut 64: 1082–1094, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 495.Wu SH, Ho SC, Zhong L. Effects of vitamin D supplementation on blood pressure. Southern Med J 103: 729–737, 2010. [DOI] [PubMed] [Google Scholar]
- 496.Xiong J, Onal M, Jilka RL, Weinstein RS, Manolagas SC, O'Brien CA. Matrix-embedded cells control osteoclast formation. Nature Med 17: 1235–1241, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 497.Xu L, Wan X, Huang Z, Zeng F, Wei G, Fang D, Deng W, Li Y. Impact of vitamin D on chronic kidney diseases in non-dialysis patients: a meta-analysis of randomized controlled trials. PloS One 8: e61387, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 498.Xu Y, Shao X, Yao Y, Xu L, Chang L, Jiang Z, Lin Z. Positive association between circulating 25-hydroxyvitamin D levels and prostate cancer risk: new findings from an updated meta-analysis. J Cancer Res Clin Oncol 140: 1465–1477, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 499.Xue Y, Fleet JC. Intestinal vitamin D receptor is required for normal calcium and bone metabolism in mice. Gastroenterology 136: 1317–1327, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 500.Yamamoto H, Shevde NK, Warrier A, Plum LA, DeLuca HF, Pike JW. 2-Methylene-19-nor-(20S)-1,25-dihydroxyvitamin D3 potently stimulates gene-specific DNA binding of the vitamin D receptor in osteoblasts. J Biol Chem 278: 31756–31765, 2003. [DOI] [PubMed] [Google Scholar]
- 501.Yamamoto Y, Yoshizawa T, Fukuda T, Shirode-Fukuda Y, Yu T, Sekine K, Sato T, Kawano H, Aihara K, Nakamichi Y, Watanabe T, Shindo M, Inoue K, Inoue E, Tsuji N, Hoshino M, Karsenty G, Metzger D, Chambon P, Kato S, Imai Y. Vitamin D receptor in osteoblasts is a negative regulator of bone mass control. Endocrinology 154: 1008–1020, 2013. [DOI] [PubMed] [Google Scholar]
- 502.Yang K, Kurihara N, Fan K, Newmark H, Rigas B, Bancroft L, Corner G, Livote E, Lesser M, Edelmann W, Velcich A, Lipkin M, Augenlicht L. Dietary induction of colonic tumors in a mouse model of sporadic colon cancer. Cancer Res 68: 7803–7810, 2008. [DOI] [PubMed] [Google Scholar]
- 503.Ye P, Rodriguez FH, Kanaly S, Stocking KL, Schurr J, Schwarzenberger P, Oliver P, Huang W, Zhang P, Zhang J, Shellito JE, Bagby GJ, Nelson S, Charrier K, Peschon JJ, Kolls JK. Requirement of interleukin 17 receptor signaling for lung CXC chemokine and granulocyte colony-stimulating factor expression, neutrophil recruitment, and host defense. J Exp Med 194: 519–527, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 504.Yim S, Dhawan P, Ragunath C, Christakos S, Diamond G. Induction of cathelicidin in normal and CF bronchial epithelial cells by 1,25-dihydroxyvitamin D3. J Cystic Fibrosis 6: 403–410, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 505.Yin JW, Wang G. The Mediator complex: a master coordinator of transcription and cell lineage development. Development 141: 977–987, 2014. [DOI] [PubMed] [Google Scholar]
- 506.Yoshizawa T, Handa Y, Uematsu Y, Takeda S, Sekine K, Yoshihara Y, Kawakami T, Arioka K, Sato H, Uchiyama Y, Masushige S, Fukamizu A, Matsumoto T, Kato S. Mice lacking the vitamin D receptor exhibit impaired bone formation, uterine hypoplasia and growth retardation after weaning. Nature Genet 16: 391–396, 1997. [DOI] [PubMed] [Google Scholar]
- 507.Yu X, Ibrahimi OA, Goetz R, Zhang F, Davis SI, Garringer HJ, Linhardt RJ, Ornitz DM, Mohammadi M, White KE. Analysis of the biochemical mechanisms for the endocrine actions of fibroblast growth factor-23. Endocrinology 146: 4647–4656, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 508.Zehnder D, Evans KN, Kilby MD, Bulmer JN, Innes BA, Stewart PM, Hewison M. The ontogeny of 25-hydroxyvitamin D3 1alpha-hydroxylase expression in human placenta and decidua. Am J Pathol 161: 105–114, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 509.Zella JB, Plum LA, Plowchalk DR, Potochoiba M, Clagett-Dame M, DeLuca HF. Novel, selective vitamin D analog suppresses parathyroid hormone in uremic animals and postmenopausal women. Am J Nephrol 39: 476–483, 2014. [DOI] [PubMed] [Google Scholar]
- 510.Zella LA, Kim S, Shevde NK, Pike JW. Enhancers located within two introns of the vitamin D receptor gene mediate transcriptional autoregulation by 1,25-dihydroxyvitamin D3. Mol Endocrinol 20: 1231–1247, 2006. [DOI] [PubMed] [Google Scholar]
- 511.Zella LA, Meyer MB, Nerenz RD, Lee SM, Martowicz ML, Pike JW. Multifunctional enhancers regulate mouse and human vitamin D receptor gene transcription. Mol Endocrinol 24: 128–147, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 512.Zella LA, Meyer MB, Nerenz RD, Pike JW. The enhanced hypercalcemic response to 20-epi-1,25-dihydroxyvitamin D3 results from a selective and prolonged induction of intestinal calcium-regulating genes. Endocrinology 150: 3448–3456, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 513.Zella LA, Shevde NK, Hollis BW, Cooke NE, Pike JW. Vitamin D-binding protein influences total circulating levels of 1,25-dihydroxyvitamin D3 but does not directly modulate the bioactive levels of the hormone in vivo. Endocrinology 149: 3656–3667, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 514.Zhang J, Chalmers MJ, Stayrook KR, Burris LL, Wang Y, Busby SA, Pascal BD, Garcia-Ordonez RD, Bruning JB, Istrate MA, Kojetin DJ, Dodge JA, Burris TP, Griffin PR. DNA binding alters coactivator interaction surfaces of the intact VDR-RXR complex. Nature Struct Mol Biol 18: 556–563, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 515.Zhang J, Yao Z. Effect of 1,25(OH)2D3 on the growth and apoptosis of breast cancer cell line MCF-7. Chinese Med J 113: 124–128, 2000. [PubMed] [Google Scholar]
- 516.Zhang K, Song L. Association between vitamin D receptor gene polymorphisms and breast cancer risk: a meta-analysis of 39 studies. PloS One 9: e96125, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 517.Zhang W, Na T, Wu G, Jing H, Peng JB. Down-regulation of intestinal apical calcium entry channel TRPV6 by ubiquitin E3 ligase Nedd4-2. J Biol Chem 285: 36586–36596, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 518.Zheng W, Tayyari F, Gowda GA, Raftery D, McLamore ES, Shi J, Porterfield DM, Donkin SS, Bequette B, Teegarden D. 1,25-Dihydroxyvitamin D regulation of glucose metabolism in Harvey-ras transformed MCF10A human breast epithelial cells. J Steroid Biochem Mol Biol 138: 81–89, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 519.Zheng W, Wong KE, Zhang Z, Dougherty U, Mustafi R, Kong J, Deb DK, Zheng H, Bissonnette M, Li YC. Inactivation of the vitamin D receptor in APC(min/+) mice reveals a critical role for the vitamin D receptor in intestinal tumor growth. Int J Cancer 130: 10–19, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 520.Zheng W, Xie Y, Li G, Kong J, Feng JQ, Li YC. Critical role of calbindin-D28k in calcium homeostasis revealed by mice lacking both vitamin D receptor and calbindin-D28k. J Biol Chem 279: 52406–52413, 2004. [DOI] [PubMed] [Google Scholar]
- 521.Zhong Y, Armbrecht HJ, Christakos S. Calcitonin, a regulator of the 25-hydroxyvitamin D3 1alpha-hydroxylase gene. J Biol Chem 284: 11059–11069, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 522.Zhou C, Lu F, Cao K, Xu D, Goltzman D, Miao D. Calcium-independent and 1,25(OH)2D3-dependent regulation of the renin-angiotensin system in 1alpha-hydroxylase knockout mice. Kidney Int 74: 170–179, 2008. [DOI] [PubMed] [Google Scholar]
- 523.Zhu J, DeLuca HF. Vitamin D 25-hydroxylase: four decades of searching, are we there yet? Arch Biochem Biophys 523: 30–36, 2012. [DOI] [PubMed] [Google Scholar]
- 524.Zhu JG, Ochalek JT, Kaufmann M, Jones G, Deluca HF. CYP2R1 is a major, but not exclusive, contributor to 25-hydroxyvitamin D production in vivo. Proc Natl Acad Sci USA 110: 15650–15655, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 525.Zierold C, Nehring JA, DeLuca HF. Nuclear receptor 4A2 and C/EBPbeta regulate the parathyroid hormone-mediated transcriptional regulation of the 25-hydroxyvitamin D3-1alpha-hydroxylase. Arch Biochem Biophys 460: 233–239, 2007. [DOI] [PubMed] [Google Scholar]
- 526.Zinser GM, McEleney K, Welsh J. Characterization of mammary tumor cell lines from wild type and vitamin D3 receptor knockout mice. Mol Cell Endocrinol 200: 67–80, 2003. [DOI] [PubMed] [Google Scholar]
- 527.Zisman AL, Ghantous W, Schinleber P, Roberts L, Sprague SM. Inhibition of parathyroid hormone: a dose equivalency study of paricalcitol and doxercalciferol. Am J Nephrol 25: 591–595, 2005. [DOI] [PubMed] [Google Scholar]