

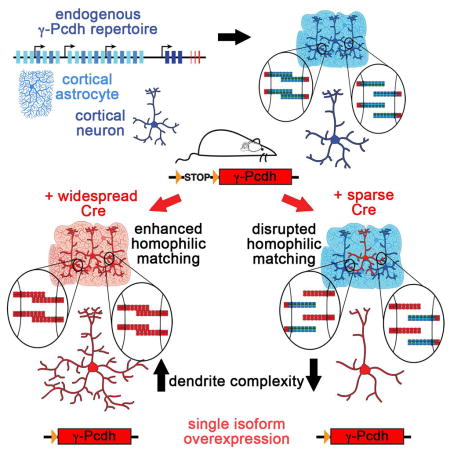
SUMMARY
Growth of a properly complex dendrite arbor is a key step in neuronal differentiation and a prerequisite for neural circuit formation. Diverse cell surface molecules, such as the clustered protocadherins (Pcdhs), have long been proposed to regulate circuit formation through specific cell-cell interactions. Here, using transgenic and conditional knockout mice to manipulate γ-Pcdh repertoire in the cerebral cortex, we show that the complexity of a neuron’s dendritic arbor is determined by homophilic interactions with other cells. Neurons expressing only one of the 22 γ-Pcdhs can exhibit either exuberant, or minimal, dendrite complexity depending only on whether surrounding cells express the same isoform. Furthermore, loss of astrocytic γ-Pcdhs, or disruption of astrocyte-neuron homophilic matching, reduces dendrite complexity cell non-autonomously. Our data indicate that γ-Pcdhs act locally to promote dendrite arborization via homophilic matching and confirm that connectivity in vivo depends on molecular interactions between neurons, and between neurons and astrocytes.
Graphical abstract
INTRODUCTION
The “chemoaffinity hypothesis”, that establishment of neuronal connectivity is aided by matching of diverse molecular cues between cells, has been very influential (Sperry, 1963; Zipursky and Sanes, 2010). A select group of immunoglobulin (Sidekicks, DSCAMs) and cadherin (Cadherin-8 and -9) cell adhesion molecules, as well as some semaphorins (Sema5A, 5B, and 6A), has been shown to specify dendritic targeting and/or formation of particular circuits (Duan et al., 2014; Krishnaswamy et al., 2015; Matsuoka et al., 2011a,b; Williams et al., 2011; Yamagata and Sanes, 2008; Yamagata et al., 2002). Candidates for generating the extraordinary molecular diversity that would be required for cell-cell interactions throughout the brain include Drosophila Dscam, with over 38,000 homophilically-interacting isoforms generated by alternative splicing (mammalian DSCAM does not exhibit such diversity), and mammalian clustered protocadherins (α-, β-, and γ-Pcdhs), which can form thousands of distinct homophilically-interacting cis-multimers (Garrett et al., 2012b; Hattori et al., 2007; Kise and Schmucker, 2013; Schreiner and Weiner, 2010; Thu et al., 2014; Wojtowicz et al., 2004; Rubinstein et al., 2015; Nicoludis et al, 2015).
In contrast to the chemoaffinity hypothesis as originally conceived (Zipursky and Sanes, 2010), fly Dscam controls the formation of dendritic arbors by mediating self-avoidance: homophilic isoneuronal matching between a neuron’s own dendrites results in repulsion, preventing self-crossing and fasciculation and allowing proper dendritic field spread (Garrett et al., 2012b; Kise and Schmucker, 2013; Zipursky and Grueber, 2013). The 22 mammalian γ-Pcdhs, encoded by the Pcdhg gene cluster (Wu and Maniatis, 1999; Figure S1A), have been shown to play an analogous, cell-autonomous self-avoidance role in retinal starburst amacrine cells (SACs), preventing isoneuronal dendrite fasciculation while allowing heteroneuronal overlap of their planar dendritic arbors (Kostadinov and Sanes, 2015; Lefebvre et al., 2012). In other CNS populations, including cortical and hippocampal neurons, hypothalamic neurons, retinal ganglion cells, and spinal interneurons, the γ-Pcdhs have been shown to regulate neuronal survival, synapse maturation, and/or axon and dendrite morphology (Garrett et al., 2012a; Garrett and Weiner, 2009; Keeler et al., 2015; Lefebvre et al., 2008; Prasad et al., 2008; Prasad and Weiner, 2011; Su et al., 2010; Suo et al., 2012; Wang et al., 2002; Weiner et al., 2005). Several of these studies indicated that the γ-Pcdhs play these roles, at least in part, cell non-autonomously via heteroneuronal and neuron-astrocyte interactions (Su et al., 2010; Garrett and Weiner, 2009; Prasad and Weiner, 2011). This indicates that cell-autonomous self-avoidance is not the only mechanism through which the γ-Pcdhs regulate neuronal development.
Cell-aggregation assays have found that the clustered Pcdhs exhibit strictly homophilic trans-interactions between their extracellular domains, with specificity mediated by extracellular cadherin domains 2 and 3 (EC2/3) (Schreiner and Weiner, 2010; Thu et al., 2014; Rubinstein et al., 2015). Based on size-exclusion chromatography and apparent molecular weight of complexes on western blots, Schreiner and Weiner (2010) proposed that the unit of γ-Pcdh trans-interaction was a cis-tetramer. Subsequent crystal structure data indicated, rather, that the clustered Pcdhs form zipper-like assemblies of cis homo- or hetero-dimers interacting in an antiparallel fashion across cell-cell junctions (Rubinstein et al., 2015). All of these studies assaying cell-aggregation in non-adhesive K562 cells are in agreement that the introduction of a single mismatching Pcdh isoform into a cell that otherwise matches surrounding cells in Pcdh repertoire is sufficient to disrupt cell aggregation (Schreiner and Weiner, 2010; Thu et al., 2014; Rubinstein et al., 2015). Though cell aggregation in K562 cells is an adhesive, rather than repulsive, phenomenon, neurons may have intracellular signaling pathways that convert an initial γ-Pcdh adhesive event into a repulsive cell response. Such a repulsive response can be inferred from the aberrant dendrite fasciculation and self-crossing in SACs and cerebellar Purkinje cells lacking the γ-Pcdhs (Lefebvre et al., 2012; Kostadinov and Sanes, 2015). However, earlier studies in fibroblast cell lines (Frank et al., 2005) and hippocampal neurons (Fernandez-Monreal et al., 2009) also showed that exogenous γ-Pcdhs can aggregate at stable cell-cell junctions and sites of axon and dendrite contact. This suggests that homophilic Pcdh trans-interactions between cells might also mediate adhesion or other positive signaling relevant to the growth of some developing neuronal subsets.
We showed previously that loss of the γ-Pcdhs throughout the developing cerebral cortex results in greatly reduced complexity of pyramidal neuron dendrite arbors in the absence of aberrant fasciculation (Garrett et al., 2012a); a similar phenotype has also been observed in hippocampal neurons lacking the α-Pcdhs (Suo et al., 2012). Because the dendrites of pyramidal neurons extend in all directions and thus, during development, will contact many surrounding γ-Pcdh-expressing neurons and astrocytes, a self-avoidance role seems less likely to explain these phenotypes. We thus tested the hypothesis that homophilic γ-Pcdh trans-interactions between neurons, and between neurons and astrocytes, provide a positive signal for dendritic growth and complexity by manipulating γ-Pcdh repertoire in discrete cell types. Across multiple in vivo and in vitro experiments, we consistently find that increasing homophilic matching between cells leads to greater dendrite complexity, while decreasing homophilic matching between cells leads to reduced dendrite complexity. Our results indicate that in the cerebral cortex, homophilic γ-Pcdh interactions between cells provide a positive cue that promotes dendrite arborization.
RESULTS
Increasing γ-Pcdh homophilic matching results in greater dendrite complexity in the cortex in vivo
In the mouse Pcdhg cluster, 22 large “variable” exons encoding 6 EC domains, a transmembrane domain, and a ~90 amino acid cytoplasmic domain are each expressed from their own promoters and spliced to three “constant” exons that encode a shared 125 amino acid C-terminal domain (Wu and Maniatis, 1999; Figure S1A). Single-cell RT-PCR (Kaneko et al., 2006; Yagi, 2012), in situ hybridization (Wang et al., 2002; Zou et al., 2007), and transcriptome (Zhang et al., 2014) data indicate that each neuron and astrocyte expresses a fraction of the Pcdhg isoforms: a few of the 19 stochastically- and monoallelically-expressed A and B subfamily members plus 2 or 3 broadly- and biallelically-expressed C subfamily members (Figure S1A). Homophilic interaction between γ-Pcdhson two adjacent cells depends on the degree of isoform repertoire matching (Schreiner and Weiner, 2010; Thu et al., 2014; Rubinstein et al., 2015).
We first sought to increase homophilic γ-Pcdh isoform matching between cells by using Emx1-Cre, which is active in all excitatory neurons and astrocytes in the cortex (Garrett et al., 2012a; Gorski et al., 2002) to broadly activate expression of mCherry-tagged γ-Pcdh-A1 or -C3 transgenes (Figure S1B,C; previously described and utilized in Lefebvre et al., 2012 and Kostadinov and Sanes, 2015). We generated mice expressing these single-isoform transgenes either in the presence (A1- or C3-overexpression [OE]) or absence (A1- or C3-Only) of endogenous γ-Pcdhs by additionally employing the Pcdhgfcon3 conditional knockout allele (Figure 1A–C and Figure S1A; Garrett et al., 2012a; Prasad et al., 2008). The A1 isoform is representative of the sparsely-expressed genes situated at the differentially-methylated 5′ end of the Pcdhg cluster, whereas the widely-expressed C3 isoform is encoded by a gene situated at the hypomethylated 3′ end of the cluster (Kaneko et al., 2006; Zou et al., 2007; Zhang et al., 2014; Toyoda et al., 2014; Lein et al., 2007; Allen Mouse Brain Atlas: http://mouse.brain-map.org). Pyramidal neuron dendrite arbors labeled by the addition of the Thy1-YFPH allele (Feng et al., 2000) were traced at 5 weeks of age from confocal stacks of somatosensory cortex, and Sholl analysis performed to measure dendrite complexity. We confirmed that the exogenous A1-mCherry and C3-mCherry proteins can co-immunoprecipitate endogenous γ-Pcdhs, indicating they integrate into multimers as expected (Figure S1D). Additionally, as expected from previous analyses of mice harboring duplications or deletions of the Pcdha or Pcdhg clusters (Noguchi et al., 2009; Chen et al., 2012), we confirmed that expression of exogenous A1-mCherry did not significantly affect levels of several representative endogenous γ-Pcdh proteins (Figure S1E).
Figure 1. Increased homophilic γ-Pcdh matching promotes dendrite arbor complexity in the cerebral cortex.

A,B) Schematics representing the cortex of controls and mice in which the A1-mCherry or C3-mCherry transgene is expressed in all primary neurons (thick lined cells) and astrocytes (thin lined cells) using Emx1-Cre. C) Cryosection from an Emx1-Cre; Pcdhgfcon3/fcon3; C3-mCherry animal showing GFP-tagged endogenous γ-Pcdhs (green, striatum [str]) and exogenous C3-mCherry (red, cortex [ctx], corpus callosum [cc]); Cre excision is limited to the cortex, and there is no leaky expression from the non-excised transgene. D,E) Representative images and traces of Thy1-YFPH-labeled layer V pyramidal neurons in the indicated genotypes; arrowheads indicate traced neurons in (D). F–H) Sholl analysis graphs showing dendrite crossings of circles drawn at 10 μm radius intervals from the soma (F,G) and area under the curve graph (H) for indicated genotypes. N=60 (control, A1-Only, C3-Only) or 120 (A1-OE, C3-OE) neurons per genotype; bar in (C)=50 μm; bar in (D)=60 μm; *p<0.05, ***p<0.001. See also Figure S1.
In the Emx1-Cre A1-Only and C3-Only single-isoform cortex, all cells should match and thus be able to interact homophilically (Figure 1B). We found that neurons traced from Emx1-Cre A1- and C3-Only cortex exhibited greatly enhanced dendrite arbor complexity compared to controls (Figure 1D–H and Figure S1F–H). In the A1-OE cortex, homophilic matching should be significantly increased due to the low, scattered expression of the endogenous A1 isoform (Zhang et al., 2014; Zou et al., 2007). Consistent with this, we observed a similar significant increase in arbor complexity (as measured by Sholl analysis) in A1-OE cortex (Figure 1E,F,H). For A1-OE and A1-Only neurons, this reflected a significant increase in both the number of branchpoints and the total dendrite length (Figure S1F,G), without any increase in the average length of each branch (Figure S1H); this latter measure was, however, also significantly increased in C3-Only neurons, suggesting that C3 may play an isoform-specific role in dendritic patterning (Figure S1H). Because most, if not all, cells already highly express the endogenous C3 isoform (Kaneko et al., 2006; Yagi, 2012; Zhang et al., 2014) activation of the C3-mCherry transgene on a wildtype background in the C3-OE cortex would not be expected to shift homophilic matching drastically. Indeed, dendrite arbor complexity in C3-OE cortex was similar to that in controls (Figure 1E,G,H and Figure S1F–H). Importantly, the normal dendrite arborization observed in C3-OE cortex indicates that a simple increase in total γ-Pcdh levels alone (documented in Figure S1C) cannot account for the enhanced arborization seen in A1-OE, A1-Only, and C3-Only cortex, in which homophilic matching between cells is expected to be increased.
Inducing γ-Pcdh homophilic mis-matching results in reduced dendrite complexity in the cortex in vivo
Next, we asked whether inducing γ-Pcdh homophilic mis-matching in the cortex in vivo would reduce dendrite arbor complexity. We utilized the Sim1-Cre line (Balthasar et al., 2005), which is active in scattered (<4%) layer V/VI pyramidal neurons in the cortex (Figure 2C and S2E), to knock out endogenous γ-Pcdhs (Figure 2A), or to express the single isoform γ-Pcdh transgenes (Figure 2B), in scattered, isolated neurons (labeled with a tdTomato Cre-reporter allele; Figure 2C). As expected from our prior work (Garrett et al., 2012a), isolated knockout neurons exhibited severely reduced dendrite arborization compared to controls (Figure 2D–F and Figure S2A–C). Strikingly, expressing the A1-mCherry transgene in isolated layer V/VI neurons, either with (Sim-A1-OE) or without (Sim-A1-Only) endogenous γ-Pcdhs, resulted in significant dendrite arborization defects indistinguishable from those of knockout neurons (Figure 2D–F; Sim-A1-OE and Sim-A1-Only results are combined here, as they were statistically indistinguishable [Figure S2F]). This reduced dendrite complexity was due to severe reductions in the number of branchpoints and total dendrite length, with a mild (but statistically significant) reduction in average branch length as well (Figure S2A–C).
Figure 2. Inducing γ-Pcdh isoform mismatching reduces dendrite arbor complexity in the cerebral cortex.

A,B) Schematics representing the cortex of mice in which endogenous γ-Pcdhs are knocked-out (grey; Sim-KO), and/or the A1-mCherry or C3-mCherry transgene is expressed (red; Sim-OE/Only) in scattered deep-layer pyramidal neurons using Sim1-Cre. C) Confocal image of isolated tdTomato+ layer V/VI neurons in a Sim1-Cre;Ai14-tdTomato cortex. D) Representative traces of tdTomato+ layer V/VI pyramidal neurons of the indicated genotypes. Sholl (E) and area under the curve (F) graphs of tdTomato+ layer V/VI pyramidal neurons of the indicated genotypes. Note that results shown here are pooled for OE and Only mice, as the results were identical [Figure S2]), N=58–60 (controls, Sim-KO), 97 (Sim-A1), or 79 (Sim-C3) neurons; Bar in C=20 μm; **p<0.01; ***p<0.001. See also Figure S2.
As we would predict given the ubiquitous expression of the endogenous C3 isoform and the Emx1-Cre results (Figure 1), dendrite arborization of isolated neurons expressing the C3-mCherry transgene were much less affected, though their arbors were still significantly less complex than control (Figure 2D–F and Figure S2A–C). Comparing these Sim1-Cre results with the Emx1-Cre results (Figure 1) clearly shows that expression of a single γ-Pcdh isoform can be sufficient for a cortical neuron to form a complex dendrite arbor, but only if surrounding cells express the same isoform. In the Emx-A1-Only cortex, neurons exhibit exuberant dendrite arborization above control levels, likely because of perfect homophilic matching with surrounding cells (Figure 1D–F, H). In the Sim-A1-Only cortex, however, neurons activating expression of the exact same transgene fail to arborize normally, because in this case few of the surrounding cells are likely to express endogenous γ-Pcdh-A1 and γ-Pcdh homophilic matching between cells is disrupted (Figure 2D–F). It is thus unlikely that our data could reflect isoneuronal dendrite self-avoidance; the dendrites of both Emx-A1-Only and Sim-A1-Only neurons will match themselves perfectly, and yet the results in these two compound transgenics are precisely opposite. To ensure that our results did not reflect changes in isoneuronal self-avoidance within Sim1-Cre cortical neurons, we directly quantified the number of dendritic self-crossings from three-dimensional confocal stacks. Sparsely-labeled tdTomato+ layer V/VI cortical neurons exhibited few, if any, instances of dendrite self-crossings or fasciculation, and the number of these did not vary between control neurons and any knockout or transgene-expressing neurons (Figure S2D).
Evidence for local control of dendrite complexity via γ-Pcdh homophilic interaction
An intriguing question is whether the effect of γ-Pcdhs on dendrite arborization is local (i.e., specific to the homophilically-matching or mis-matching dendritic branch) or global (e.g., involving a signaling change throughout the neuron). We took advantage of the varied activity pattern of the Sim1-Cre allele to address this question. As discussed above, in layers V and VI of the somatosensory cortex, Sim1-Cre is active in only a few, scattered neurons (Figures 2C and 3A,B, purple boxes): these are the neurons that were analyzed in Figure 2 and Figure S2. However, within layer IV, there is an intermittent denser band of Sim1-Cre/tdTomato+ cells (Figure 3A,B, white boxes), none of which are labeled by the layer V-restricted Thy1-YFPH transgene (Figure 3A,B). The basal and oblique dendrite arbors of Sim1-Cre/tdTomato+ layer V/VI neurons, analyzed in Figure 2, are situated well below this intermittent band in layer IV (Figure 3A,B, purple boxes) and thus make no contact with it. However, this is not the case for the more superficially-situated Thy1-YFPH-labeled neurons: In Sim-A1-OE or Sim-A1-Only cortex, their basal dendrite arbor (Figure 3A,B, yellow boxes) will contact neurons that, as they do, express only endogenous repertoires of γ-Pcdh isoforms; however, their oblique dendrites, some of which project off the apical branch as it passes through the Sim1-Cre/tdTomato+ layer IV band (Figure 3A,B, white boxes), will encounter many cells that express the A1 transgene, and thus mis-match.
Figure 3. γ-Pcdhs control dendrite complexity through local homophilic interactions.

A,B) Schematic diagram (A) and low magnification confocal image (B) of a Sim1-Cre;Thy1-YFPH;Ai14-tdTomato;A1-mCherry cortex. Regions marked by different color boxes correspond between A and B. Purple boxes mark the basal and oblique arbors of isolated tdTomato+ layer V/VI neurons, which were analyzed in Figure 2; because these neurons overexpress the A1-mCherry transgene while their surrounding cells are wildtype, their dendrites experience mis-matches that lead to greatly reduced arborization (Figure 2). Yellow boxes mark the basal arbors of YFP+ layer V neurons, which are analyzed in D. These neurons express endogenous γ-Pcdh repertoires, as do the immediately surrounding cells, so their basal arbors should exhibit normal complexity. White boxes mark the oblique arbors of YFP+ layer V neurons, which are analyzed in C (red shaded region). These oblique arbors branch off the apical shaft within layer IV, which contains a band of denser Sim1-Cre activity and thus A1-mCherry expression. These YFP+ oblique dendrites express endogenous γ-Pcdh repertoires, but locally encounter layer IV neurons overexpressing the mis-matching A1-mCherry isoform. C,D) Sholl analysis of the oblique (mis-matching; C) and basal (normal matching; D) arbors of YFP+ neurons in control (black lines) and Sim-A1-OE or Sim-A1-Only cortex (gray lines; OE and Only results are pooled here, as they were identical; see Figure S3). E) The calculated ratio of oblique/basal dendrite complexity (oblique area under the curve divided by basal area under the curve; control normalized to 1) confirms a significant localized loss of complexity only within the oblique arbor of YFP+ neurons in the Sim-A1 cortex. N=28 YFP+ neurons per genotype; Bar=100 μm; ***p<0.001.
If this layer IV-restricted mis-matching globally affected dendrite arborization, we would expect YFP+ neurons to exhibit reduced complexity throughout their entire arbor. If, on the other hand, γ-Pcdh matching or mis-matching acts locally, we would expect only the oblique dendrites of YFP+ neurons (Figure 3A,B, white boxes) to exhibit reduced complexity, with the basal dendritic arbor (Figure 3A,B, yellow boxes) remaining unaffected. Sholl analyses clearly support the latter possibility: Oblique dendritic arbors of YFP+ neurons were specifically reduced within the mis-matching layer IV band (Figure 3C, red shading), while basal arbors of the same neurons were indistinguishable from controls (Figure 3D), resulting in a significantly decreased oblique/basal complexity ratio in YFP+ neurons within the Sim-A1-OE/Only cortex (Figure 3E shows combined results, which were identical [Figure S3]). We note that a local rather than global effect is also supported by the fact that the scattered layer V/VI Sim1-Cre/tdTomato+ neurons (Figure 3A,B, purple boxes) exhibit severely reduced arborization throughout their mis-matching basal and oblique arbors (Figure 2D–F) despite the fact that their apical shaft passes through the layer IV band, which contains Sim1-Cre/tdTomato+ neurons that match them homophilically. These results strongly imply that γ-Pcdhs regulate dendrite arborization locally, via discrete neuron-neuron contacts and/or through spatially-contained signaling events.
Astrocytes regulate neuronal dendrite arborization cell non-autonomously via γ-Pcdh homophilic interactions
Since our results thus far revealed the importance of homophilic γ-Pcdh interactions between a neuron and its surrounding cells, we next asked whether astrocytic γ-Pcdhs play a role in promoting the proper formation of dendritic arbors. Astrocyte processes, which contain γ-Pcdhs and localize them to the sites of neuronal contact (Garrett and Weiner, 2009), are extensive in the cortical neuropil. Restricted mutation of the γ-Pcdhs from astrocytes in the developing spinal cord had no effect on their survival, morphology, or release of thrombospondin, but the loss of astrocyte-neuron γ-Pcdh contacts resulted in delayed synapse development (Garrett and Weiner, 2009). We utilized a Gfap-Cre line (77.6; Garcia et al., 2004) that, unlike other lines, reproducibly restricts excision to astrocytes in the cortex (Figure 4C and Figure S4A,B) to generate mice in which wildtype neurons were surrounded by γ-Pcdh-knockout astrocytes (Figure 4A). Compared to controls, neurons surrounded by knockout astrocytes exhibited significantly reduced dendrite complexity (Figure 4D–F; reductions in branchpoints and total dendrite length, but not average branch length [Figure S4G–I]). This occurred in the absence of any changes in the number of astrocytes, the size of their somata, the number of their primary branches, or the total area covered by their processes (Figure S4C–F). These data indicate that astrocytic γ-Pcdhs regulate neuronal development in a cell non-autonomous manner: the fact that the traced neurons expressed endogenous γ-Pcdhs demonstrates that isoneuronal and/or neuron-neuron interactions alone are insufficient for normal arbor complexity in cortical neurons.
Figure 4. Astrocytic γ-Pcdhs regulate dendrite arborization cell non-autonomously in the cerebral cortex.

A,B) Schematics representing neurons and astrocytes in the indicated genotypes, as in previous figures. C) Maximum projection of a confocal stack through the cortex of a Gfap-Cre;Thy1-YFPH;Ai-14-tdTomato mouse, demonstrating astrocyte-restricted excision mediated by the 77.6 Cre line. D–F) Representative traces (D), Sholl analysis (E) and area under the curve graph (F) of Thy1-YFPH+ layer V pyramidal neurons of the indicated genotypes. Bar in (C)=20 μm; N=60 neurons per genotype; **p <0.01. See also Figure S4.
We then addressed whether induced mis-matching between astrocytic and neuronal γ-Pcdhs negatively affects dendrite complexity. Using Gfap-Cre, we generated mice in which astrocytes, but not neurons, expressed the A1-mCherry transgene (Figure 4B). Wildtype neurons, few of which express endogenous γ-Pcdh-A1 and none of which overexpress A1-mCherry, indeed exhibited severely reduced dendrite arborization when surrounded by mis-matching A1-OE astrocytes (Figure 4D–F). As we found for mis-matched neurons in the Sim1-Cre analyses (Figure S2A–C), wildtype neurons encountering mis-matching astrocytes exhibited reductions in average branch length as well as in total dendrite length and number of branchpoints (Figure S4G–I). As a control, we also analyzed mice in which the C3-mCherry transgene was activated in astrocytes using Gfap-Cre. Wildtype neurons, most or all of which highly express endogenous γ-Pcdh-C3, exhibited normal dendrite complexity when surrounded by C3-OE astrocytes (Figure 4D–F and Figure S4G–I), a result entirely consistent with the Emx- and Sim-C3 results (Figures 1 and 2).
An in vitro system to manipulate dendrite complexity via γ-Pcdh repertoire matching or mis-matching
To further explore the role that homophilic γ-Pcdh matching between cells plays in dendrite complexity, we devised an in vitro assay (Figure 5A). Cells from a neonatal Emx-A1-OE or Emx-C3-OE cortex were labeled by nucleofection with a GFP-encoding plasmid, diluted at a 1:100 ratio with other cortical cells expressing either the matching, or mis-matching, mCherry-tagged isoform, and plated. Isolated GFP+/mCherry+ neurons, surrounded by GFP−/mCherry+ neurons and astrocytes (Figure 5B) were then imaged, traced, and subjected to Sholl analysis after 8 days in vitro. In all cases, dendrite arborization was significantly higher when the traced neurons were surrounded by cells overexpressing the matching γ-Pcdh (A1 on A1 and C3 on C3) compared to those surrounded by cells overexpressing the mis-matching γ-Pcdh (A1 on C3 and C3 on A1; OE results shown in Figure 5C–G; OE vs. Only results shown in Figure S5). As positive and negative controls for γ-Pcdh function, we also generated similar co-cultures using wildtype or knockout neurons (Figure 5E,G). Together, our in vitro results remarkably corroborate the in vivo data gathered using Emx1-Cre and Sim1-Cre: 1) A1-OE on A1-OE neurons exhibit greater dendrite complexity than do wildtype neurons, while C3-OE on C3-OE neurons are similar to wildtype (Figure 5E,G, c.f. Figure 1H); 2) Mis-matching neurons (A1 on C3 and C3 on A1) exhibited reduced dendrite complexity compared to wildtype (Figure 5E,G, c.f. Figure 2F); 3) Both A1-Only on A1-Only and A1-OE on A1-OE neurons exhibited similarly heightened dendrite complexity (Figure S5A, c.f. Figure 1H); and 4) C3-Only on C3-Only neurons exhibited increased complexity compared to wildtype, while C3-OE on C3-OE neurons did not (Figure S5B, c.f. Figure 1H).
Figure 5. Dendrite arborization can be increased or decreased by manipulating γ-Pcdh homophilic matching between a neuron and its environment in vitro.

A) Schematic of the neuronal co-culture assay: neonatal cortical cells from one transgenic mouse (red) are nucleofected with a plasmid encoding GFP, diluted at a 1:100 ratio with cells from another cortex expressing the same (red, top dish) or a different (purple) single-isoform transgene. B) Image of a nucleofected GFP+/mCherry+ neuron growing on GFP−/mCherry+ astrocytes in an “A1-Only on A1-Only” (perfectly matching) culture. A1-mCherry protein is concentrated at sites of contact (arrowheads), suggesting the formation of a homophilic trans-interaction complex at the cell surface. C) Representative traces of GFP+ nucleofected neurons in matching “A1 on A1” or “C3 on C3” cultures and mis-matching “A1 on C3” or “C3 on A1” cultures. Sholl (D,F) and area under the curve (E,G) graphs showing that dendrite arbor complexity is significantly higher in neurons growing in homophilically-matching co-cultures (“A1 on A1” or “C3 on C3”) than in those growing in mis-matching co-cultures (“A1 on C3” or “C3 on A1”; all results shown here are OE; OE vs. Only results shown in Figure S5). Additionally, comparison to similarly generated wildtype (WT) cultures corroborates the in vivo results shown in Figures 1, 2 and 4: “A1 on A1” OE (E) neurons exhibit significantly greater arborization than control neurons, while “C3 on C3” OE neurons do not (G). C3-Only on C3-Only neurons, again as in vivo, exhibit significantly greater arborization than do C3-OE on C3-OE neurons (Figure S5). H) Relationship of dendrites to astrocytes was quantified in fixed cultures. Each instance where a dendrite appeared to contact only the edge of an astrocyte, or to have turned to grow away from an astrocyte was counted, and this number divided by the total number of dendrite/astrocyte contacts to obtain the percentage of dendrite segments “avoiding” astrocytes(H). I) An index measuring neuron-neuron contact points was calculated by counting the number of intersections between GFP+ nucleofected dendrites and other surrounding MAP2+ dendrites, normalizing to total dendrite area and multiplying by a constant. N=57–60 (A1/C3 on A1/C3 and WT), 180 (KO), or 30–36 (avoidance and contact point index) neurons per condition; Bars in (B)=50 μm (left), 15 μm (upper, middle right), or 10 μm (lower right). **p<0.01; ***p<0.001. See also Figure S5.
We could not find evidence of repulsive avoidance in cortical co-cultures. Because dendrites often encounter astrocytes in vitro (e.g., Figure 5B), we asked whether those contacting a homophilically-matching astrocyte ever stop, grow along the astrocyte’s edge rather than onto it, or turn and grow away from the contact site. We found very few instances of dendrites that appeared to avoid growing onto astrocytes, regardless of whether they homophilically matched or mis-matched (Figure 5H). Next, we calculated a “neuronal contact point index” by counting the number of intersections or fasciculations between nucleofected GFP+ dendrites and other surrounding GFP−/MAP2+ dendrites, normalizing to total dendrite area in the field, and multiplying by a constant. We found that the extent of contact between homophilically-matching neurons (A1 on A1 and C3 on C3) was actually significantly higher than it was between mismatching neurons (A1 on C3 and C3 on A1) (Figure 5I), again failing to provide evidence for homophilic repulsion and indicating that neuron-neuron contacts contribute to the formation of complex dendritic arbors. Finally, we noted that when a neuron grew onto a homophilically matching astrocyte, mCherry-tagged γ-Pcdhs often concentrated along the length of contacts (Figure 5B, arrowheads), suggestive of homophilic trans-interactions at the cell membrane. Together, our data indicate a positive, apparently adhesive, response to γ-Pcdh homophilic matching in cortical neurons that leads to enhanced dendrite arborization.
Although cell-aggregation assays have shown that all 22 γ-Pcdhs similarly exhibit strictly homophilic interactions (Schreiner and Weiner, 2010; Thu et al., 2014), it is important to confirm that the disruptive effect of isoform mis-matching on dendrite arborization is not confined to the A1 or C3 isoforms for which transgenic lines are available. In vitro assays have demonstrated that introduction of a single mis-matching isoform to K562 cells that otherwise match in γ-Pcdh repertoire significantly disrupts homophilic cell aggregation (Schreiner and Weiner, 2010; Thu et al., 2014; Rubinstein et al., 2015). We thus used low-efficiency lipofection of matching “A1 on A1” cultures to ask whether introduction of a single mis-matching γ-Pcdh isoform to a neuron would affect the formation of its arbor. Neurons transfected with control plasmid (GFP only) or a GFP-tagged γ-Pcdh-A1 (Figure 6A–C) exhibited similarly high dendrite arbor complexity, confirming the finding that increasing total levels of γ-Pcdhs without modulating matching has little effect (c.f. C3-OE results, Figures 1, 2, 4). Dendrite arborization was, in contrast, drastically and identically reduced to knockout levels in neurons expressing any one of 4 GFP-tagged mis-matching γ-Pcdhs: A3, A12, B2, and B6 (Figure 6A–C; note that N-terminally HA-tagged B2 had the same effect as C-terminally GFP-tagged B2 in this assay). For a final test of the effect of γ-Pcdh mismatching on dendrite arborization, we utilized a chimeric molecule (EC1 and EC3 from γ-Pcdh-A3; EC2 and EC4-6 from γ-Pcdh-C3; Schreiner and Weiner, 2010) that can interact with homophilically with itself but not with any endogenous γ-Pcdhs. We lipofected wildtype cortical neuron cultures with a plasmid encoding GFP, either alone or in addition to a plasmid encoding this chimeric γ-Pcdh, and assessed dendrite arborization after 8 days in vitro. As predicted from the prior literature (Schreiner and Weiner, 2010; Thu et al., 2014; Rubinstein et al., 2015) and our other experiments here, disruption of endogenous γ-Pcdh interaction by introduction of this chimeric “poison partner” significantly reduced dendrite arbor complexity (Figure 6D).
Figure 6. Disrupting γ-Pcdh homophilic interactions by overexpressing a single mis-matching isoform reduces dendrite arborization.

Neurons in “A1 on A1” homophilically matching co-cultures were lipofected at low efficiency (1–5%) with a plasmid expressing GFP alone (Control) or in addition to a plasmid encoding one of several γ-Pcdh isoforms with a C-terminal GFP tag: γ-Pcdh-A1, -A3, -A12, -B2, or –B6 (an N-terminally HA-tagged γ-Pcdh-B2 plasmid was also used to control for effects of the GFP tag). A) Representative neuronal traces in each transfection condition. B) Sholl analysis and C) area under the curve graphs. Addition of a single non-matching γ-Pcdh isoform to a neuron growing in an otherwise homophilically-matching co-culture (“A1 on A1”) is sufficient to reduce arborization to KO levels (c.f. Figure 5E,G). D) Introducing a chimeric γ-Pcdh isoform that can interact with itself but not with any endogenous γ-Pcdh isoform into wildtype neurons also significantly reduced dendrite arborization (measured by Sholl analysis at 8 days in vitro; area under the curve shown) compared to control neurons. N=80–105 (+additional isoform) or 30 (+chimera) neurons. ***p<0.001.
DISCUSSION
Here we have differentially manipulated γ-Pcdh isoform repertoire in cells of the cerebral cortex in vivo and shown that increasing the likelihood of homophilic matching between a given neuron and its surrounding neurons and astrocytes promotes dendrite arborization. Conversely, induced mis-matching of γ-Pcdh trans-interactions results, both in vivo and in vitro, in simplified arbors that resemble those of Pcdhg knockout neurons. Our data indicate that astrocytes regulate dendrite arborization cell non-autonomously in vivo via homophilic γ-Pcdh interactions, adding to the growing evidence that these cells play critical roles in neural circuit formation (Clarke and Barres, 2013). Further, we present evidence suggesting that homophilic γ-Pcdh interactions act locally, rather than through global cell signaling, to promote the growth and complexity of dendritic arbors.
Though homophilic γ-Pcdh interactions result in repulsion between adjacent SACs in the retina (Lefebvre et al., 2012; Kostadinov and Sanes, 2015), in the cortex these interactions promote dendrite growth and branching in a way that seems more consistent with an adhesive or otherwise “positive” signaling effect. In our earlier analysis of cortical Pcdhg knockouts, we found no evidence of increased dendrite self-crossing or fasciculation (Garrett et al., 2012a), in contrast to the clear self-avoidance defects shown in SACs and Purkinje cells (Lefebvre et al., 2012). In the present study, we found that neurons sharing expression of a single γ-Pcdh isoform exhibited an increased number of contacts compared to neurons that overexpressed mis-matching isoforms. Additionally, dendrites did not avoid perfectly-matching astrocytes, but rather grew along them and clustered γ-Pcdh proteins along contact sites. These data suggest that homophilic γ-Pcdh trans-interactions can result in adhesion, repulsion, or other signaling events relevant to dendrite arborization depending on cell-type specific attributes. These might include the type and shape of the dendritic arbor (e.g., planar SACs and Purkinje cells vs. bushy cortical neurons), expression of distinct cis-interactors (including the many α- and β-Pcdhs) or variations in downstream intracellular signaling components. Such context-dependent signaling would not be surprising in a family of molecules as complex as the clustered Pcdhs; in chick retina, Dscams are adhesive rather than repulsive (Yamagata and Sanes, 2008) and guidance molecules such as netrins, ephrins, and semaphorins (Dickson, 2002) can act as both attractants and repellents depending upon the presence of distinct co-receptors or signaling pathway activity states.
Although the most parsimonious interpretation of our data is that homophilic engagement of γ-Pcdhs across cell membranes directly promotes dendrite arborization, we cannot discount the possibility that γ-Pcdh over- or mis-expression affects other proteins involved in regulating arborization. Proteomic analysis of γ-Pcdh-containing protein complexes identified, in addition to other clustered Pcdhs, over 120 putative interacting proteins (Han et al., 2010). These include other adhesion-associated proteins with dendritic roles (e.g., classical cadherins, catenins) as well as proteins regulating trafficking, cytoskeleton, and intracellular signaling. As a portion of γ-Pcdh proteins are found in intracellular vesicular compartments (Fernandez-Monreal et al., 2010) it may well be that γ-Pcdhs regulate the trafficking of other cell surface proteins that are critical for dendrite arborization. Similarly, it is not yet clear whether homophilic trans-interaction of γ-Pcdhs in neurons directly induces a dendritic branch point or instructs dendrite growth along a contact site. We and others previously showed that the γ-Pcdhs can promote dendrite arborization by regulating signaling pathways involving the adhesion kinases FAK and Pyk2, PKC, MARCKS, and Rho GTPases (Garrett et al., 2012a; Suo et al., 2012; Keeler et al., 2015). These studies did not address whether γ-Pcdh regulation of such signaling required homophilic trans-interactions or could take place constitutively. If the latter, then an increase in total γ-Pcdh protein levels, as seen in our studies using the single-isoform transgenics, could conceivably be important. For several reasons noted earlier, we believe that this is unlikely to be a major driver of our results; nevertheless, in the absence of tools to accurately quantify γ-Pcdh surface levels on a cell-by-cell basis, we can’t exclude the possibility that subtle variations in γ-Pcdh expression modulate their function. Finally, we note that in vitro studies indicate that the γ-Pcdhs can undergo two cleavage events: matrix metalloproteinases such as ADAM10 mediate ectodomain shedding, while γ-secretase cuts near the membrane to release an intracellular fragment that can translocate to the nucleus (Haas et al., 2005; Reiss et al., 2006). It is not yet clear how such cleavage is regulated in vivo, whether it differs for different isoforms, or whether cis- or trans-interactions modulate it. The regulation of γ-Pcdh levels at the dendritic cell surface by this or other mechanisms will be important to address in future studies.
We previously identified a role for γ-Pcdh-mediated astrocyte-neuron contacts in the regulation of synaptogenesis in the embryonic spinal cord (Garrett and Weiner, 2009). Here we extend the in vivo functions of astrocytic γ-Pcdhs, showing that either loss of endogenous γ-Pcdhs, or overexpression of a sparsely-expressed γ-Pcdh isoform (A1), in astrocytes cell non-autonomously disrupted dendrite arborization in cortical neurons. Several lines of evidence indicate that these results are due to loss of γ-Pcdh homophilic interactions between astrocytes and neurons, rather than undetected cellular or molecular defects in GFAP-KO or GFAP-A1 astrocytes: 1) Complete loss of γ-Pcdhs in both astrocytes and neurons using Emx1-Cre did not result in any increase in apoptosis of either cell type (Garrett et al., 2012a); 2) Overexpression of the A1 transgene in both astrocytes and neurons in EMX-A1-OE cortex (Figure 1) led to increased dendrite arborization, while restricted expression of the exact same transgene in astrocytes only (GFAP-A1-OE cortex, Figure 4) resulted in reduced dendrite arborization; 3) Astrocyte number, soma size, number of primary branches, and area covered by astrocyte branches were all identical between controls and GFAP-KO, GFAP-A1-OE, and GFAP-C3-OE cortex (Figure S4). It is important to note that the reduction in dendrite complexity observed in the Gfap-Cre knockout cortex is not as severe as that previously reported in the Emx1-Cre knockout cortex (Garrett et al., 2012a). This indicates that, while astrocyte-neuron γ-Pcdh interactions are necessary for normal dendritic arbor complexity, neuron-neuron interactions must contribute as well. This is further supported by other in vivo (Figure 3C) and in vitro (Figure 5I) data in the present study.
It is interesting that dendrite complexity in neurons encountering astrocytes overexpressing a mis-matching γ-Pcdh isoform was lower than in those encountering knockout astrocytes (Figure 4F). Though the reason for this is not clear, it might imply the collaboration of α- and/or β-Pcdhs, which have been found to interact physically and functionally in cis with γ-Pcdhs (Murata et al., 2004; Han et al. 2010; Thu et al., 2014), in homophilic adhesion between neurons and astrocytes. If adhesive interfaces in vivo indeed incorporate α-, β-, and γ-Pcdhs, introduction of a mis-matching γ-Pcdh in astrocytes could act as a “poison partner” (analogous to the experiments shown in Figure 6), and affect neuronal dendrite arborization more than simple loss of astrocytic γ-Pcdhs would. Hippocampal neurons lacking α-Pcdhs exhibit reduced dendrite complexity in vivo (Suo et al., 2012), though as yet there are no reports of phenotypes in mice lacking the β-Pcdhs. Clearly, it will be important in the future to examine neurons lacking all clustered Pcdhs to determine whether α-, β-, and γ-Pcdhs act through the same pathways to regulate neuronal morphology.
A recent model based on structural and biochemical data (Rubinstein et al., 2015) suggests that the unit of clustered Pcdh interaction in trans is a dimer of dimers, and that many of these units together could form a zipper-like structure. Cells with identical isoform repertoires would be able to form larger zipper-like Pcdh assemblies, while even a small number of mis-matches would be enough to severely abrogate such trans-interactions (Rubinstein et al., 2015). Though this model was developed to explain Pcdh-mediated self-avoidance, our results are not inconsistent with such a mis-match mechanism. Overexpression in a given neuron of a single isoform unlikely to be found in surrounding cells (Sim-A1 mice, Figure 2; transfection of A1 on A1 neurons with other isoforms, Figure 6) led to a severe reduction in dendrite arborization, while overexpression of a single isoform likely to be shared by all cells (Sim-C3 mice, Figure 2; transfection of A1 on A1 neurons with A1-GFP, Figure 6) affected arborization mildly or not at all. One caveat of our study is that we manipulated only 2 γ-Pcdh isoforms as proof-of-principle exemplars in vivo. While it is possible that manipulation of other isoforms could give different results than we see for A1 and C3, this is unlikely given that: 1) all γ-Pcdhs can interact in cis- and their trans-interactions are all strictly homophilic in K562 assays (Schreiner and Weiner, 2010; Thu et al., 2014); and 2) we showed directly that 4 other γ-Pcdh isoforms (A3, A12, B2, and B6) can, when introduced into otherwise-matching neurons, significantly disrupt dendrite arborization (Figure 6).
Given the apparently stochastic expression of the 19 endogenous γ-Pcdh A and B subfamily isoforms, on a background of near-ubiquitous expression of the 3 C subfamily isoforms (Yagi, 2012), there should be a particular statistical chance, albeit a small one, of cells with similar repertoires being in the vicinity of a neuron’s growing dendrites. In order to determine precisely how our proof-of-principle results shown here, utilizing single overexpressed isoforms, relate to the normal developmental interactions between endogenous repertoires of γ-Pcdhs, we need a better understanding of many variables. These include, but are not limited to: 1) the extent to which isoform protein levels vary cell-by-cell and if so, whether this can significantly modulate interactions; 2) whether any isoform-specific rules govern the formation of cis-complexes in vivo (e.g., Must every dimer of dimers include a C subfamily isoform? Are homodimers favored over heterodimers?); 3) whether all possible combinations of isoforms, given a cell’s transcript repertoire, are translated, delivered to the plasma membrane, and localized equivalently; 4) whether individual isoforms, through their variable cytoplasmic domains, activate unique intracellular signaling partners; and 5) whether partial homophilic matches within a dimer or other multimer (i.e., only 1 of 2 isoforms in a dimer of dimers being shared), while not allowing stable interaction, might nevertheless transiently activate a signaling event with a distinct outcome.
In any case, it seems unlikely that homophilic trans-interactions between endogenous γ-Pcdhs would provide directional or instructive signals to particular individual cells; rather, the statistical likelihood of enough homophilic interactions occurring (however many “enough” is) might place an permissive bound on the extent of a neuron’s dendrite arbor that would normalize arbor complexity across a large population. Our results allow us to predict that reduction in functional Pcdhg gene repertoire in the cortex, either through mutation or hypermethylation, should result in an increased chance of homophilic matching between cells and an increase in the complexity of dendritic arbors. A key future goal will be to demonstrate this experimentally using new animal models, as well as to consider epigenetic changes in the Pcdhg cluster that could contribute to developmental disorders in which altered dendrite morphology is observed.
EXPERIMENTAL PROCEDURES (See also Supplemental Experimental Procedures)
Mouse strains
The A1-mCherry and C3-mCherry transgenic lines (Lefebvre et al., 2012) were the kind gift of Julie Lefebvre and Joshua Sanes (Harvard University). The Pcdhgfcon3 conditional mutant allele has been described (Garrett et al., 2012a; Prasad et al., 2008). The Gfap-Cre, Emx1-Cre, Sim1-Cre,Thy1-YFPH and Ai14-TdTomato lines were obtained from The Jackson Laboratory (Bar Harbor, ME).
Cortical neuron cultures
Cultures were prepared as described (Garrett et al., 2012a). For co-cultures, cells harvested from individual animals were plated at a density of 200,000 cells per coverslip. Remaining cells were nucleofected (Amaxa) with N1-EGFP plasmid and plated at 2,000 cells per coverslip onto the previously-plated high-density cells from the same, or another, individual animal. In other experiments, low efficiency (~1–5%) transfection using Lipofectamine 2000 (Invitrogen) was performed at 2 DIV.
Analysis of dendrite arborization
Transgenic mice were perfused with 4% PFA at 5 weeks of age. Confocal images of Vibratome sections were taken using a 20X objective on Leica SP2 AOBS or Leica TCS SPE microscopes. All collected images were blinded to condition. Fifty to 60μm Z-stacks were collected with a 0.5μm step size, imported into FIJI (NIH Image/J), and dendrites traced in 3D using the Simple Neurite Tracer plugin. Epifluorescence images cultured neurons (8 DIV) were taken using a 20X objective on a Leica TCS SPE microscope and traced in 2D. The Sholl Analysis function of Simple Neurite Tracer was used. Comparisons were performed in Prism by one-way ANOVA, corrected for multiple comparisons with Tukey’s test and reported multiplicity adjusted p values for each comparison, or by unpaired, two-tailed t-test when comparing only two samples. Asterisks denote the following significance levels: *p<0.05; **p<0.01; ***p<0.001.
Supplementary Material
Highlights.
Increasing γ-Pcdh homophilic matching promotes dendrite complexity in the cortex
Inducing γ-Pcdh homophilic mismatching reduces dendrite complexity in the cortex
Astrocytes regulate dendrite complexity cell non-autonomously via γ-Pcdh interactions
γ-Pcdhs promote dendrite complexity through local interactions
Acknowledgments
We thank Leah Fuller for mouse breeding and genotype analysis; Joshua Sanes and Julie Lefebvre for providing the A1-mCherry and C3-mCherry mouse lines and for helpful discussions; and Robert Burgess, Andrew Garrett, and Michael Dailey for helpful discussions. Supported by NIH R01 NS055272 and March of Dimes grant FY11-456 to J.A.W.
Footnotes
AUTHOR CONTRIBUTIONS
M.J.M, A.B.K, and J.A.W. conceived the experiments, collected and analyzed data, and wrote the manuscript.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T, Ferreira M, Tang V, McGovern RA, Kenny CD, et al. Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell. 2005;123:493–505. doi: 10.1016/j.cell.2005.08.035. [DOI] [PubMed] [Google Scholar]
- Chen J, Lu Y, Meng S, Han MH, Lin C, Wang X. alpha- and gamma-Protocadherins negatively regulate PYK2. J Biol Chem. 2009;284:2880. doi: 10.1074/jbc.M807417200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke LE, Barres BA. Emerging roles of astrocytes in neural circuit development. Nat Rev Neurosci. 2013;14:311–321. doi: 10.1038/nrn3484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickson BJ. Molecular mechanisms of axon guidance. Science. 2002;298:1959–1964. doi: 10.1126/science.1072165. [DOI] [PubMed] [Google Scholar]
- Duan X, Krishnaswamy A, De la Huerta I, Sanes JR. Type II cadherins guide assembly of a direction-selective retinal circuit. Cell. 2014;158:793–807. doi: 10.1016/j.cell.2014.06.047. [DOI] [PubMed] [Google Scholar]
- Feng G, Mellor RH, Bernstein M, Keller-Peck C, Nguyen QT, Wallace M, Nerbonne JM, Lichtman JW, Sanes JR. Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron. 2000;28:41–51. doi: 10.1016/s0896-6273(00)00084-2. [DOI] [PubMed] [Google Scholar]
- Fernandez-Monreal M, Kang S, Phillips GR. Gamma-protocadherin homophilic interaction and intracellular trafficking is controlled by the cytoplasmic domain in neurons. Mol Cell Neurosci. 2009;40:344. doi: 10.1016/j.mcn.2008.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez-Monreal M, Oung T, Hanson HH, O’Leary R, Janssen WG, Dolios G, Wang R, Phillips GR. Gamma-protocadherins are enriched and transported in specialized vesicles associated with the secretory pathway in neurons. Eur J Neurosci. 2010;32:921–931. doi: 10.1111/j.1460-9568.2010.07386.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank M, Ebert M, Shan W, Phillips GR, Arndt K, Colman DR, Kemler R. Differential expression of individual gamma-protocadherins during mouse brain development. Mol Cell Neurosci. 2005;29:603–16. doi: 10.1016/j.mcn.2005.05.001. [DOI] [PubMed] [Google Scholar]
- Garcia AD, Doan NB, Imura T, Bush TG, Sofroniew MV. GFAP-expressing porogenitors are the principal source of constitutive neurogenesis in adult mouse forebrain. Nat Neurosci. 2004;7:1233–1241. doi: 10.1038/nn1340. [DOI] [PubMed] [Google Scholar]
- Garrett AM, Schreiner D, Lobas MA, Weiner JA. -Protocadherins Control Cortical Dendrite Arborization by Regulating the Activity of a FAK/PKC/MARCKS Signaling Pathway. Neuron. 2012a;74:269–276. doi: 10.1016/j.neuron.2012.01.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrett AM, Tadenev AL, Burgess RW. DSCAMs: restoring balance to developmental forces. Front Mol Neurosci. 2012b;5:86. doi: 10.3389/fnmol.2012.00086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrett AM, Weiner JA. Control of CNS synapse development by {gamma}-protocadherin-mediated astrocyte-neuron contact. J Neurosci. 2009;29:11723–11731. doi: 10.1523/JNEUROSCI.2818-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorski JA, Talley T, Qiu M, Puelles L, Rubenstein JL, Jones KR. Cortical excitatory neurons and glia, but not GABAergic neurons, are produced in the Emx1-expressing lineage. J Neurosci. 2002;22:6309–6314. doi: 10.1523/JNEUROSCI.22-15-06309.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haas IG, Frank M, Veron N, Kemler R. Presenilin-dependent processing and nuclear function of g-protocadherins. J Biol Chem. 2005;280:9313–9319. doi: 10.1074/jbc.M412909200. [DOI] [PubMed] [Google Scholar]
- Han MH, Lin C, Meng S, Wang X. Proteomics analysis reveals overlapping functions of clustered protocadherins. Mol Cell Proteomics. 2010;9:71–83. doi: 10.1074/mcp.M900343-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hattori D, Demir E, Kim HW, Viragh E, Zipursky SL, Dickson BJ. Dscam diversity is essential for neuronal wiring and self-recognition. Nature. 2007;449:223–227. doi: 10.1038/nature06099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaneko R, Kato H, Kawamura Y, Esumi S, Hirayama T, Hirabayashi T, Yagi T. Allelic gene regulation of Pcdh-alpha and Pcdh-gamma clusters involving both monoallelic and biallelic expression in single Purkinje cells. J Biol Chem. 2006;281:30551–30560. doi: 10.1074/jbc.M605677200. [DOI] [PubMed] [Google Scholar]
- Keeler AB, Schreiner D, Weiner JA. Protein Kinase C Phosphorylation of a gamma-Protocadherin C-terminal Lipid Binding Domain Regulates Focal Adhesion Kinase Inhibition and Dendrite Arborization. J Biol Chem. 2015;290:20674–20686. doi: 10.1074/jbc.M115.642306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kise Y, Schmucker D. Role of self-avoidance in neuronal wiring. Curr Opin Neurobiol. 2013;23:983–989. doi: 10.1016/j.conb.2013.09.011. [DOI] [PubMed] [Google Scholar]
- Kostadinov D, Sanes JR. Protocadherin-dependent dendritic self-avoidance regulates neural connectivity and circuit function. eLife. 2015;4 doi: 10.7554/eLife.08964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krishnaswamy A, Yamagata M, Duan X, Hong YK, Sanes JR. Sidekick 2 directs formation of a retinal circuit that detects differential motion. Nature. 2015;524:466–470. doi: 10.1038/nature14682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lefebvre JL, Kostadinov D, Chen WV, Maniatis T, Sanes JR. Protocadherins mediate dendritic self-avoidance in the mammalian nervous system. Nature. 2012;488:517. doi: 10.1038/nature11305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lefebvre JL, Zhang Y, Meister M, Wang X, Sanes JR. gamma-Protocadherins regulate neuronal survival but are dispensable for circuit formation in retina. Development. 2008;135:4141–4151. doi: 10.1242/dev.027912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lein ES, et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature. 2007;445:168–176. doi: 10.1038/nature05453. [DOI] [PubMed] [Google Scholar]
- Matsuoka RL, Nguyen-Ba-Charvet KT, Parray A, Badea TC, Chédotal A, Kolodkin AL. Transmembrane semaphorin signalling controls laminar stratification in the mammalian retina. Nature. 2011a;470:259. doi: 10.1038/nature09675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsuoka RL, Chivatakarn O, Badea TC, Samuels IS, Cahill H, Katayama K, Kumar SR, Suto F, Chédotal A, Peachey NS, Nathans J, Yoshida Y, Giger RJ, Kolodkin AL. Class 5 transmembrane semaphorins control selective Mammalian retinal lamination and function. Neuron. 2011b;71:460–73. doi: 10.1016/j.neuron.2011.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murata Y, Hamada S, Morishita H, Mutoh T, Yagi T. Interaction with protocadherin-gamma regulates the cell surface expression of protocadherin-alpha. J Biol Chem. 2004;279:49508–49516. doi: 10.1074/jbc.M408771200. [DOI] [PubMed] [Google Scholar]
- Nicoludis JM, Lau SY, Schärfe CPI, Marks DS, Weihofen WA, Gaudet R. Structure and sequence analyses of clustered protocadherins reveal antiparallel interactions that mediate homophilic specificity. Structure. 2015;23:1–12. doi: 10.1016/j.str.2015.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noguchi Y, Hirabayashi T, Katori S, Kawamura Y, Sanbo M, Hirabayashi M, et al. Total expression and dual gene-regulatory mechanisms maintained in deletions and duplications of the Pcdha cluster. J Biol Chem. 2009;284:32002–14. doi: 10.1074/jbc.M109.046938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasad T, Wang X, Gray PA, Weiner JA. A differential developmental pattern of spinal interneuron apoptosis during synaptogenesis: insights from genetic analyses of the protocadherin-gamma gene cluster. Development. 2008;135:4153–4164. doi: 10.1242/dev.026807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasad T, Weiner JA. Direct and Indirect Regulation of Spinal Cord Ia Afferent Terminal Formation by the -Protocadherins. Frontiers in Molecular Neuroscience. 2011;4 doi: 10.3389/fnmol.2011.00054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reiss K, Maretzky T, Haas IG, Schulte M, Ludwig A, Frank M, Saftig P. Regulated ADAM10-dependent ectodomain shedding of γ-protocadherin C3 modulates cell-cell signaling. J Biol Chem. 2006;281:21735–21744. doi: 10.1074/jbc.M602663200. [DOI] [PubMed] [Google Scholar]
- Rubinstein R, Thu CA, Goodman KM, Wolcott HN, Bahna F, Mannepalli S, Ahlsen G, Chevee M, Halim A, Clausen H, Maniatis T, Shapiro L, Honig B. Molecular logic of neuronal self-recognition through protocadherin domain interactions. Cell. 2015;163:629–642. doi: 10.1016/j.cell.2015.09.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schreiner D, Weiner JA. Combinatorial homophilic interaction between gamma-protocadherin multimers greatly expands the molecular diversity of cell adhesion. Proc Natl Acad Sci U S A. 2010;107:14893–14898. doi: 10.1073/pnas.1004526107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sperry RW. Chemoaffinity in The Orderly Growth Of Nerve Fiber Patterns And Connections. Proc Natl Acad Sci U S A. 1963;50:703–10. doi: 10.1073/pnas.50.4.703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su H, Marcheva B, Meng S, Liang FA, Kohsaka A, Kobayashi Y, Xu AW, Bass J, Wang X. Gamma-protocadherins regulate the functional integrity of hypothalamic feeding circuitry in mice. Developmental biology. 2010;339:38. doi: 10.1016/j.ydbio.2009.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suo L, Lu H, Ying G, Capecchi MR, Wu Q. Protocadherin clusters and cell adhesion kinase regulate dendrite complexity through Rho GTPase. J Mol Cell Biol. 2012;4:362–376. doi: 10.1093/jmcb/mjs034. [DOI] [PubMed] [Google Scholar]
- Thu CA, Chen WV, Rubinstein R, Chevee M, Wolcott HN, Felsovalyi KO, Tapia JC, Shapiro L, Honig B, Maniatis T. Single-Cell Identity Generated by Combinatorial Homophilic Interactions between alpha, beta, and gamma Protocadherins. Cell. 2014;158:1045–1059. doi: 10.1016/j.cell.2014.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toyoda S, Kawaguchi M, Kobayashi T, Tarusawa E, Toyama T, Okano M, Oda M, Nakauchi, Yoshimura HY, Sanbo M, Hirabayashi M, Hirayama T, Hirabayashi T, Yagi T. Developmental epigenetic modification regulates stochastic expression of clustered protocadherin genes, generating single neuron diversity. Neuron. 2014;82:94–108. doi: 10.1016/j.neuron.2014.02.005. [DOI] [PubMed] [Google Scholar]
- Wang X, Weiner J, Levi S, Craig A, Bradley A, Sanes J.Anonymous. Gamma protocadherins are required for survival of spinal interneurons. Neuron. 2002;36:843. doi: 10.1016/s0896-6273(02)01090-5. [DOI] [PubMed] [Google Scholar]
- Weiner JA, Wang X, Tapia JC, Sanes JR. Gamma protocadherins are required for synaptic development in the spinal cord. Proc Natl Acad Sci U S A. 2005;102:8–14. doi: 10.1073/pnas.0407931101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams ME, Wilke SA, Daggett A, Davis E, Otto S, Ravi D, Ripley B, Bushong EA, Ellisman MH, Klein G, Ghosh A. Cadherin-9 regulates synapse-specific differentiation in the developing hippocampus. Neuron. 2011;71:640–655. doi: 10.1016/j.neuron.2011.06.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wojtowicz WM, Flanagan JJ, Millard SS, Zipursky SL, Clemens JC. Alternative splicing of Drosophila Dscam generates axon guidance receptors that exhibit isoform-specific homophilic binding. Cell. 2004;118:619–633. doi: 10.1016/j.cell.2004.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu Q, Maniatis T. A striking organization of a large family of human neural cadherin-like cell adhesion genes. Cell. 1999;97:779–790. doi: 10.1016/s0092-8674(00)80789-8. [DOI] [PubMed] [Google Scholar]
- Yagi T. Molecular codes for neuronal individuality and cell assembly in the brain. Front Mol Neurosci. 2012;5:45. doi: 10.3389/fnmol.2012.00045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamagata M, Sanes JR. Dscam and Sidekick proteins direct lamina-specific synaptic connections in vertebrate retina. Nature. 2008;451:465–469. doi: 10.1038/nature06469. [DOI] [PubMed] [Google Scholar]
- Yamagata M, Weiner JA, Sanes JR. Sidekicks: synaptic adhesion molecules that promote lamina-specific connectivity in the retina. Cell. 2002;110:649–660. doi: 10.1016/s0092-8674(02)00910-8. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Chen K, Sloan SA, Bennett ML, Scholze AR, O’Keeffe S, Phatnani HP, Guarnieri P, Caneda C, Ruderisch N, et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J Neurosci. 2014;34:11929–11947. doi: 10.1523/JNEUROSCI.1860-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zipursky SL, Grueber WB. The molecular basis of self-avoidance. Annu Rev Neurosci. 2013;36:547–568. doi: 10.1146/annurev-neuro-062111-150414. [DOI] [PubMed] [Google Scholar]
- Zipursky SL, Sanes JR. Chemoaffinity revisited: dscams, protocadherins, and neural circuit assembly. Cell. 2010;143:343–353. doi: 10.1016/j.cell.2010.10.009. [DOI] [PubMed] [Google Scholar]
- Zou C, Huang W, Ying G, Wu Q. Sequence analysis and expression mapping of the rat clustered protocadherin gene repertoires. Neuroscience. 2007;144:579–603. doi: 10.1016/j.neuroscience.2006.10.011. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.