

Abstract
Transforming growth factor-β1 (TGF-β1) has been proven to function primarily in mammalian immunobiological activity, but information regarding the immune role of TGF-β1 in teleosts is limited. In the present study, we describe the cDNA cloning and characterization of the TGF-β1 molecule in the topmouth culter. TGF-β1 is highly expressed in immune-related tissues of the culter, including the thymus, head kidney, and spleen. The recombinant culter TGF-β1 (cTGF-β1) was successfully expressed and purified in vitro, and the effects of cTGF-β1 on the mRNA expression of pro-inflammatory cytokines, such as TNF-α and IL-1β, in the absence or presence of LPS was determined in culter peripheral blood leukocytes. cTGF-β1 was found to have bipolar properties in inflammatory reactions. Additionally, to assess the immune role of teleost TGF-β1 in vivo, the expression of TGF-β1 in the culter thymus and spleen tissues induced by poly I:C were also examined. The expression of TGF-β1 was obviously up-regulated, as shown in the cell lines. However, the peak time of cTGF-β1 expression in the cell lines occurred significantly earlier than in the organic tissues under the same inducer, suggesting that the response of the teleost TGF-β1 molecule to exogenous infection depends on a more complicated signalling pathway in vivo than in vitro.
Transforming growth factor-β (TGF-β) has been studied largely over the past few decades and has been demonstrated to be an important multifunctional cytokine involved in the regulation of cell proliferation, differentiation, survival, migration and apoptosis under physiological and pathological conditions1. TGF-β is initially released from macrophages, fibroblasts and platelets2 as a small complex termed ‘latent TGF-β’, which consists of mature TGF-β associated with the latency-associated protein (LAP)3. Before TGF-β can perform its function, the active TGF-β must be separated from the LAP and the binding protein; subsequently, the cytokine exerts its effects via the receptor-Smad signal transduction pathway4.
Vast evidence indicates that TGF-β plays a critical immunoregulatory role in mammals both in the innate and adaptive immune pathways5. In general, TGF-β inhibits T cell proliferation by blocking interleukin-2 production and cyclin expression1 and exerts multiple stimulatory effects on B cells, natural killer cells and dendritic cells, including the activation of these cells and the regulation of chemotaxis6,7,8. TGF-β has also been shown to regulate the active and inactive states of monocytes and macrophages under specific conditions9. Finally, TGF-β can up-regulate the expression of the fibronectin receptor by monocytes10,11,12, making TGF-β a potent chemoattractant13,14.
Three TGF-β isoforms, TGF-β1, β2 and β3, have been identified in mammals, among which TGF-β1 plays a primary role in immunobiological activity. The TGF-β1 isoform is also the most studied in non-mammals, and has been cloned and characterized in several fish species, including carp15, hybrid striped bass16, sea bream17, grass carp18 and goldfish3. These studies provided new evidence for use in interpreting the immunoregulatory mechanism of the TGF-β1 gene in fish species. However, the data pertaining to the functional analysis of this cytokine are still very scarce and fragmentary, and the functional role of TGF-β1 in fish immunoregulation is still unclear.
Culter alburnus, or the topmouth culter (hereafter referred to as ‘culter’), is widely distributed throughout large rivers, reservoirs and lakes in China. Over recent decades, this fish species has become one of the most commercially important freshwater cultured fish species in China. Unfortunately, outbreaks of infectious diseases in freshwater cultured culter species have recently become a major bottleneck in the culter fishery industry. These fish appear to be vulnerable to various pathogens, such as bacteria, viruses and parasites. Thus, finding new clues regarding culter immunomodulation is critical in explaining the susceptibility of this species to pathogens.
Despite the crucial role of TGF-β in immunity, the relevant information pertaining to this cytokine in culter remains largely lacking. In the present study, we report for the first time the cloning of TGF-β1 from culter (cTGF-β1), followed by the expression and purification of the TGF-β1 protein in vitro. Furthermore, the effect of cTGF-β1 on the mRNA expression levels of pro-inflammatory cytokines, including TNF-α and IL-1β, in the absence or presence of LPS was determined in culter peripheral blood leukocytes (PBLs). The TGF-β1 molecule showed bipolar properties in the inflammatory reaction. Additionally, the expression levels of the TGF-β1 molecule were up-regulated in both the culter thymus and spleen tissues after induction with polyinosinic-polycytidylic acid (poly I:C), although TGF-β1 appeared to be more sensitive to poly I:C induction in the thymus than in the spleen. Taken together, the results presented in this study will help to improve our understanding of the role of TGF-β1 in teleost immunobiological activity.
Results
Molecular cloning and characterization of cTGF-β1
The culter TGF-β1 cDNA (GenBank accession no. KJ725122) was assembled by cDNA cloning based on 3′- and 5′-RACE. The full length cTGF-β1 cDNA was 2175 bp, including a 1134 bp opening reading frame, a 529 bp 5′ untranslated region and a 512 bp 3′ untranslated region (Fig. S1). The putative cTGF-β1 sequence, which consisted of 377 amino acids containing the precursor region and mature region, had a molecular weight of approximately 43.21 kDa. An alignment analysis showed that some structural domains that are conserved in mammals and other fish species was also present in the cTGF-β1 molecule, including a characteristic RGD integrin-binding site and a RRKR cut site in the precursor region, nine cysteine residues allowing for the formation of inter-chain and intra-chain disulfide bonds, the C-terminal cysteine knot in the mature peptide, and the conserved proline and glycine residues in the mature peptide, which are the distinguishing hallmarks of the TGF superfamily (Fig. 1).
Figure 1. Alignments of the deduced amino acid sequence of cTGF-β1 with its homologues in other species.

Numbers on the right of the sequences indicate the amino acid positions. Gaps are noted with hyphens. The mature peptide portion is noted with the arrow. The signal peptide region, RGD integrin-binding site, RRKR cut site, nine conserved cysteine residues, and conserved proline and glycine at positions 36 and 46 of the mature peptide are marked in red, blue, brown, yellow, green and grey, respectively. The GenBank accession numbers of the detected species are as follows: D. rerio (NP_878293.1); C. alburnus (AIS23633.1); O. mykiss (NP_001268295.1); T. rubripes (BAM44873.1); E. coioides (ACV96791.1); C. carpio (Q9PTQ2.1); S. salar (ACN11294.1); M. musculus (NP_035707.1); H. sapiens (NP_000651.3); and A. mexicanum (ABX24523.1).
In the phylogenetic tree, cTGF-β1 first clustered with its zebrafish and rainbow trout homologues (Fig. 2), with which cTGF-β1 shared the highest sequence identity (91% and 69% amino acid identity, respectively), suggesting that cTGF-β1 had the closest phylogenetic relationship with zebrafish TGF-β1 and rainbow trout TGF-β1. Various teleost TGF-β1 molecules clustered into one distinct branch and were subsequently grouped with their congeners from frog, chicken, and mammals to form one large branch, which shared a more distant polygenetic relationship with the TGF-β2 s and TGF-β3s of fish, chicken, frogs and mammals (Fig. 2).
Figure 2. Phylogenetic analysis of cTGF-β1 with other isoforms based on the neighbour-joining method.
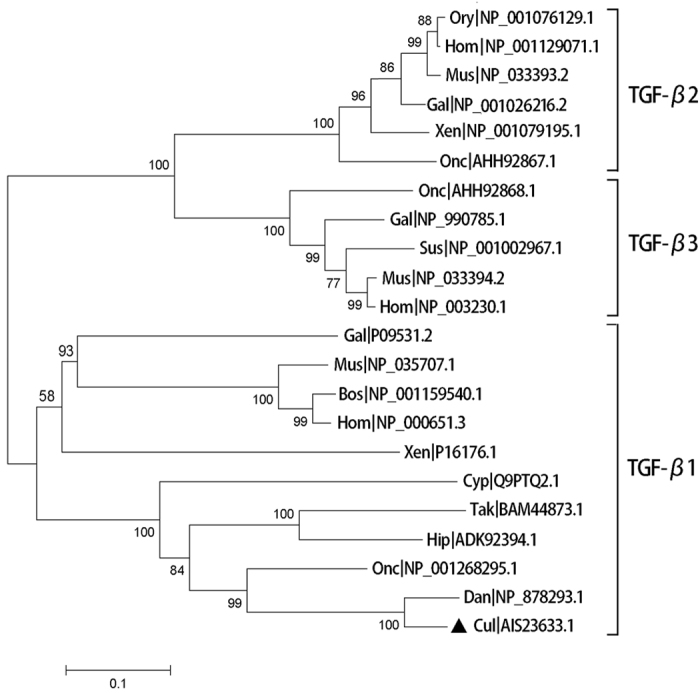
cTGF-β1 is indicated with a black triangle. The numbers near each branch represent the bootstrap values obtained with 10,000 replicates. Abbreviations of the species presented in the figure are as follows: Tak, Takifugu rubripes; Onc, Oncorhynchus mykiss; Dan, Danio rerio; Hip, Hippocampus kuda; Cul, Culter alburnus; Cyp, Cyprinus carpio; Xen, Xenopus laevis; Mus, Mus musculus; Bos, Bos taurus; Gal, Gallus gallus; Hom, Homo sapiens; Ory, Oryctolagus cuniculus; Gal, Gallus gallus; and Sus, Sus scrofa.
Tissue distribution of cTGF-β1
Eleven tissues were analysed using qPCR assays to determine the cTGF-β1 tissue distribution profile. As shown in Fig. 3, cTGF-β1 mRNA was constitutively expressed in all detected tissues. However, the expression levels varied significantly among different tissues. The cTGF-β1 transcript was most highly expressed in the thymus, head kidney, and spleen (surpassing 7-fold relative expression levels compared to the gill), to a lesser extent in the ovary, heart, liver, muscle, intestine and gill (from 1- to 3-fold relative expression levels compared to the gill), and at relatively low levels in the brain and eye (less than 1-fold relative expression levels compared to the gill).
Figure 3. Tissue distribution of TGF-β1 molecule in culter fish.
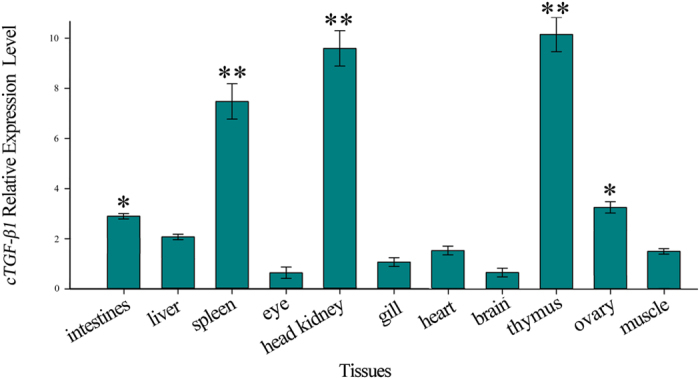
The results are represented as the mean ± SEM of 8 individual fish. The data were compared relative to β-actin (endogenous control) and were normalized to the expression in the gill.
Construction, expression and detection of the cTGF-β1 recombinant protein
The mature peptide portion of the deduced cTGF-β1 amino acids sequence was cloned into the pET32a plasmid with the addition of the BamHI and XhoI cutting sites to construct the expression vector pET32a-cTGF-β1. The conditions were optimized, and the fusion protein was efficiently expressed and subsequently purified. An SDS-PAGE analysis showed a distinct protein band of 33.88 kDa, which corresponded to the calculated molecular weight of the recombinant protein after IPTG induction compared to the empty vector control. A western blot analysis indicated that the bacterially expressed protein showed a predominant band with an estimated molecular weight of 33.88 kDa (Fig. 4). After purification of the protein with a commercial kit, no more than 0.1 EU endotoxin residues remained in 1 ml of the purified cTGF-β1 isolate.
Figure 4. Expression of the mature peptide portion of cTGF-β1 in vitro.
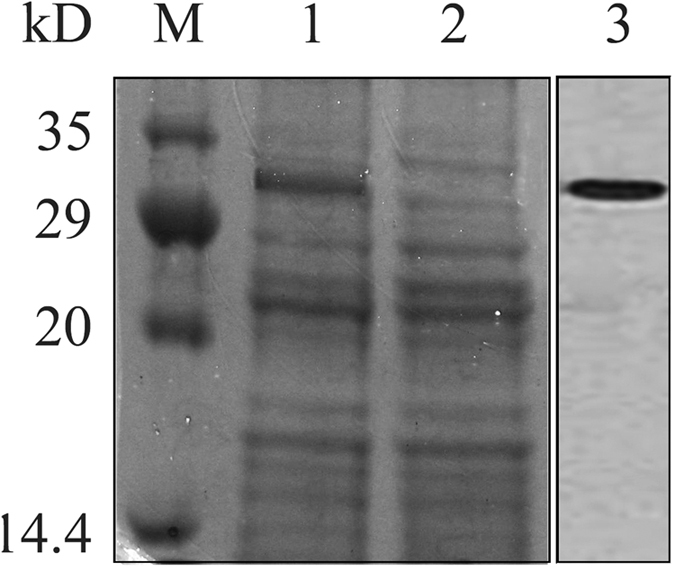
SDS-PAGE and western blot results using tissue from two testes. Each individual testis are indicated by the black frame. M, protein molecular weight marker; lane 1, lysates of transformed bacteria after induction with IPTG; lane 2, lysates of bacteria transformed with the non-inserted control; lane 3, western blot analysis of recombinant cTGF-β1 using an anti-6 × His tag mouse antibody.
Effects of cTGF-β1 on the mRNA expression of pro-inflammatory factors in PBLs
To investigate the role of cTGF-β1 in the inflammatory reaction, PBLs were isolated and cultured with 100 ng/ml of cTGF-β1, 10 μg/ml of LPS, or both of these substances for 24 h, 48 h and 96 h before the RNA was extracted. The variations in the expression levels of TNF-α and IL-1β, which both attach to pro-inflammatory factors, were chosen for examination. As shown in Fig. 5, these two factors showed expression profiles that were essentially identical after induction. At hour 24, the expression levels of TNF-α and IL-1β under the three types of induction all showed no significant difference compared to the controls. Subsequently, the expression levels sharply increased, reaching significant levels at hour 48. However, the dual induction with cTGF-β1 and LPS seemed to promote the expression of TNF-α and IL-1β much more than either cTGF-β1 or LPS alone. Notably, the expression of TNF-α increased to a highly significant level at hour 48 under the dual induction. The expression levels of TNF-α and IL-1β continually increased and peaked at hour 96 with induction by either cTGF-β1 or LPS. By contrast, the expression of these two factors showed noticeable, sharp decreases with the dual induction of cTGF-β1 and LPS after hour 48; this was especially the case for the expression level of TNF-α, which decreased to an undetectable level at hour 96.
Figure 5.

Transcript expression analysis of the pro-inflammatory factors (A) TNF-α and (B) IL-1β with the induction of cTFG-β1, LPS, or cTFG-β1 and LPS. The data presented (mean ± SEM, N = 4) are the representative results of three individual experiments. (*) Indicates statistical significance (P < 0.05).
Induction of cTGF-β1 expression by poly I:C
To assess the immunological role of TGF-β1 in vivo, the expression of this cytokine was examined in two immune-related tissues via induction by poly I:C, which was presented as a viral analogue. As shown in Fig. 6, the induction by poly I:C significantly up-regulated the expression of TGF-β1 in the thymus and the spleen. The TGF-β1 expression levels in these two culter tissues both peaked at hour 72 (11.8-fold and 185.6-fold in the spleen and thymus, respectively) and subsequently decreased gradually. However, cTGF-β1 showed a different sensitivity to poly I:C between the two tissues: in the thymus, cTGF-β1 appeared to be more sensitive to poly I:C than in the spleen, as the expression of cTGF-β1 was significantly up-regulated at hour 12 in the thymus compared to hour 48 in the spleen.
Figure 6.

Expression analysis of the cTGF-β1 transcript in the spleen (A) and the thymus (B) after induction by poly I:C. The values depicted in the graphs are the mean ± SEM of 4 individual fish. (*) Indicates statistical significance (P < 0.05).
Discussion
In the present study, the full-length cDNA of TGF-β1 in culter was cloned for the first time, and the amino acid sequence of cTGF-β1, which contains 377 residues, was deduced. The alignment analysis showed that all structural domains were strictly conserved in all detected TGF-β1 counterparts from various species. Phylogenetically, cTGF-β1 was closely related to its homologue in other vertebrates. All of these results indicated that the teleost TGF-β1 molecule may be functionally similar to that of higher vertebrates. Indeed, the TGF-β system has been shown to be highly conserved; the ligands, receptors and signalling molecules associated with this system can be traced back to arthropods, suggesting that the TGF-β system is over 1 billion years old19.
The tissue distribution of cTGF-β1 showed a ubiquitous expression pattern. This result was in accordance with previous results showing that in other teleosts, such as rainbow trout20, common carp15, hybrid striped bass16, sea bream17, and grass carp18, TGF-β1 is expressed in all detected tissues. This suggests that the TGF-β1 molecule plays multiple roles in teleost physiological activities, which has been proven in mammals. However, the cTGF-β1 molecule showed the highest expression levels in the thymus, spleen and head kidney. Similar results were also observed in the grass carp, in which the expression of TGF-β1 was the highest in those three tissues18. Because the thymus, spleen and head kidney tissues have been shown to be the major lymphoid organs in fish21, these results suggested that the TGF-β1 molecule may predominantly play an immunological role in teleost physiological activities.
The TGF-β family members are ubiquitous molecules with pleiotropic functions in a wide variety of cells22,23,24. Notably, TGF-β is a key regulator of host defence that performs an essential role in immune function modulation in both the innate and adaptive immune pathways5,25,26. TGF-β1, the most important isoform of TGF-β, has been widely studied in mammals over the past few decades. However, information regarding the functional role of TGF-β1 in fish immunity has been limited. In the present study, the effect of TGF-β1 on the mRNA expression of the pro-inflammatory cytokines TNF-α and IL-1β in the absence or presence of LPS was determined in culter PBLs. The results showed that the cTGF-β1 significantly increased the mRNA levels of TNF-α and IL-1β, but significantly attenuated the LPS-enhanced mRNA levels of these two cytokines. This finding was similar to the results found in mammals, in which the cytokine TGF-β1 performs a dual role in the inflammatory reaction9,27. Mammalian TGF-β, which has been shown to be one of the first agents to appear in the inflammatory response, acts as a pro-inflammatory cytokine to trigger monocyte recruitment and cytokine expression1,5. At the end of inflammation, TGF-β functions as a terminal signal to turn off the inflammatory reaction28. Actually, parallel results have also been observed in several fish species in recent years. Jang et al. reported that mammalian rTGF-β could enhance or suppress trout macrophage respiratory burst activity29 under different conditions. In goldfish, TGF-β1 significantly blocks the TNF-α-induced activation of macrophages and induces the proliferation of the goldfish fibroblast cell line CCL713. In grass carp, TGF-β1 down-regulates the LPS/PHA-stimulated proliferation of PBLs, which is in contrast to the stimulatory effect of TGF-β1 alone in the same type of cells30. In red seabream, recombinant TGF-β1 could induce HKL and PBL migration in a dose-dependent manner, but suppressed LPS-activated HKL and PBL migration31. These findings suggest that the teleost TGF-β1 molecules may be employed in an immunoregulatory mechanism involved in the inflammation response that is analogous to the same system in mammals.
Regrettably, these previously mentioned studies that were conducted in fish species mostly focused on the interpretation of the TGF-β1 immunoregulatory mechanism at the cellular level in vitro. Few works have been concerned with the immunological characteristics of this cytokine in organic tissues in vivo. The organ-based immune system is extremely sophisticated, and the constituent components of this system are ubiquitous and systemically acted upon during a single immune event. Aiming to assess the immune role of teleost TGF-β1 in vivo, the expression of this cytokine in culter thymus and spleen tissues after induction with poly I:C was examined. In the previous in vitro studies, teleost cells such as orange spotted grouper HKLs32 and teleost epithelial cells33 were also induced with poly I:C and showed obvious up-regulation in TGF-β1 expression. Remarkably, despite the significant increase in cTGF-β1 expression observed in the thymus and spleen, the time to peak TGF-β1 expression was noticeably different in different teleost cell lines and organ tissues; TGF-β1 expression peaked significantly earlier in the cell lines than in the organ tissues (hour 4 after stimulation in orange spotted grouper head kidney lymphocytes and hour 6 in teleost epithelial cells vs hour 72 after stimulation in culter thymus and spleen tissues), suggesting that the cTGF-β1 molecule may synergistically act with other immunologic factors after the poly I:C induction in vivo. Overall, these results suggest that TGF-β1 plays an immunoregulatory role in culter and that the teleost TGF-β1 may be involved in a more complicated signalling pathway in response to exogenous infection in vivo compared to in vitro.
Interestingly, the cTGF-β1 molecule in the thymus appeared to be more sensitive to poly I:C than in the spleen. The thymus is considered to be the central lymphoid organ34 and is primarily responsible for T cell development in teleosts35. As reported, the thymus controls the differentiation of the head kidney and spleen during the histogenesis of the lymphoid organs25,36. In rainbow trout, developing B cells mature in the head kidney and then migrate to sites of activation, which are either the spleen or the posterior kidney37. Accordingly, we supposed that the culter TGF-β1 originated from the lymphocytes of the thymus and then migrated to the secondary immune organs, such as the head kidney and spleen.
Taken together, the results presented in this study may provide valuable information regarding the potential functional mechanism of TGF-β1 in teleosts.
Methods
Animal uses
Healthy C. alburnus specimens were obtained from the Chidonghu fishery, Hubei Province, China and were maintained at the Aquatic Facility of the Fishery college, Huazhong Agricultural University. All animal use and studies were approved by the Institutional Animal Care and Use Committee of Hubei province, Wuhan, P. R. China. All procedures were carried out in accordance with the approved guidelines.
Sample collection and nucleic acid extraction
All of the fish were approximately one year old and ranged from 100–150 g. The fish were kept at 25 °C in a flowthrough water system for two weeks before treatment. During the experiment, the fish were fed with commercial feed.
The heart, intestine, eye, head kidney, brain, spleen, muscle, liver, thymus, ovary and gill were removed from 8 freshly killed culter fish using a lethal dose of MS-222 at 200 mg/L and frozen immediately in liquid nitrogen and stored at −80 °C until RNA extraction.
The total RNA was extracted from the frozen tissues using TRIzol reagent (Life Technologies, USA) following the manufacturer’s instructions in combination with DNase I treatment. The RNA quality was assessed by running the samples on 1% agarose gels and visualizing by staining with ethidium bromide. The RNA concentration was measured using a NanoDrop 2000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA) with an absorption at 260 nm.
Amplification of cTGF-β1 cDNA
Two pairs of degenerate primers (T1 and T2, Table 1) for amplification of the cTGF-β1 cDNA partial sequence were designed at the conserved regions of teleost TGF-β1 cDNA using the Primer Premier 5.0 software (Premier Biosoft International, USA). Reverse transcription polymerase chain reaction (RT-PCR) was performed using the total RNA from C. alburnus spleen as the template.
Table 1. Sequence of primers used for amplification of partial cDNA, 3′ and 5′ RACE of topmouth culter TGF-β1 and qPCR analysis for cTGF-β1,TNF-α and IL-1β.
| Primer | Sequence (5′–3′) | Usage |
|---|---|---|
| T-1 | GAYCTGGRNTGGAARTGGAT | TGF-β1 partial |
| GARGCBATTMGRRGHCAGAT | ||
| T-2 | GCMGAKGCWCCDGGRTTGTG | TGF-β1 partial |
| GTTTTCAGCATTCCAGATGTAGGTG | ||
| 5′RACE | CCTTGTGTTGTCTTCC | TGF-β1 5′RACE |
| GACTGCAGGTACGCAACATGG | ||
| GAGTATTTGTTCTCAGCATTCC | ||
| 3′RACE | GTGCATCTGCTCAGCCATGTTGCGTACCT | TGF-β1 3′RACE |
| CCTGCAGTCCTCGACCCTCTACCAATTCT | ||
| T-y | CAGGATCCGATGTTGGCACTGATGTAACTTG | TGF-β1 expression |
| GACCTCGAGACTGCACTTGCAGCTCCTCAC | ||
| T-q | GGAGGTGCACAAGTTCATTAT | TGF-β1 qPCR |
| GGCAGATATGATGGTGAAGCC | ||
| TNF-q | ACTGTCTCCTTCACGCTCCA | TNF-α qPCR |
| CTGTGAGGTGCCATTCGCTT | ||
| IL-q | ACGCAGGCCGGAGCCTCTGT | IL-1β qPCR |
| CCTGTAGATGAGGCGGCTGTC | ||
| β-actin | TGCGTGACATCAAGGAGAAG | internal control |
| GCTGGAAGGTGGACAGAGAG |
The first-strand cDNAs were synthesised using the RevertAidTM First Strand cDNA Synthesis Kit (Fermentas, Canada) and oligo(dT)18 primers. PCR was carried out with 2 μl synthesized cDNA in a 50-μl reaction volume containing 10 pmol of each specific primer and 2 units of Taq DNA polymerase (Takara). The PCRs were all performed on a Veriti thermocycler (Life Technologies, USA). The PCR conditions were as follows: an initial denaturation of 94 °C for 5 min; 35 cycles of denaturation at 94 °C for 30 s, annealing for 30 s, and extension at 72 °C for 60 s; and a final extension at 72 °C for 7 min. The PCR products were detected on a 1.5% agarose gel and were then subcloned into pMD18-T vectors (Takara). The confirmed recombinant plasmid DNA was used as the template for automated sequencing using an ABI 3730 automated DNA sequencer. The nucleotide sequences of the sense strands of the plasmid inserts of three independent clones were determined and were then assembled using the CAP3 online software (http://pbil.univ-lyon1.fr/cap3.php).
To obtain the 5′ and 3′ ends of cTGF-β1 cDNA, a number of transcript-specific sense or antisense primers (5′RACE and 3′RACE, Table 1) were designed from the partial cDNA sequence. The 5′- and 3′-rapid amplifications of the cDNA ends (5′- and 3′-RACE) were performed using the RACE cDNA Amplification Kit (Life Technologies, USA) with splenic total RNA as the template according to the manufacturer’s protocol. The nested PCR products were subcloned and sequenced as described above. Following the generation of the 5′- and 3′-RACE fragments, the core fragments were assembled to obtain the cTGF-β1 cDNA full length.
Sequence analysis and phylogenetic relationships
The nucleotide and amino acid sequences were identified using the BLAST program (GenBank, NCBI). Conserved TGF-β1 motifs were identified using the analytical tools provided by the ExPASy Molecular Biology Server (http://www.expasy.org). Multiple alignments were performed using ClustalX 1.8338. Phylogenetic relationships were investigated using the Neighbour-Joining (NJ) method in the Mega 5.0 program (Molecular Evolutionary Genetic Analysis)39. A bootstrap analysis was conducted using 10,000 replicates.
Construction of the pET32a- cTGF-β1 expression vector
Culter TGF-β1 constructs were designed for expression using the pET32a prokaryotic expression system. The culter TGF-β1 transcript encoding the mature peptide portion of the predicted amino acid sequence was amplified by PCR using the primers T-y (Table 1). PCR was conducted on a Veriti thermocycler with Taq DNA polymerase (Takara). The amplification procedure consisted of 35 cycles of 94 °C for 30 s, 58 °C for 30 s, and 72 °C for 60 s, followed by a final extension at 72 °C for 7 min. The PCR product was digested with BamHI and XhoI and then cloned into the pET-32a vector to express a fusion protein of cTGF-β1 with an N-terminal 6 × His tag. This recombinant plasmid was sequenced and named pET32a-cTGF-β1.
Production, purification and analysis of recombinant culter TGF-β1
The pET32a-cTGF-β1 vector was transformed into BL21 competent cells (Life Technologies) to express the fusion protein. The bacteria were inoculated in LB media with vigorous shaking at 37 °C. The bacterial optical density was measured using a NanoDrop 2000 spectrophotometer with absorption at 600 nm every two hours. When the OD600 reached approximately 0.6, the protein expression was induced by isopropyl-beta-D-thiogalactopyranoside to a final concentration of 1.0 mM. Eight hours after induction under the previously described conditions, the bacteria were harvested by centrifugation at 5000 × g for 15 min and resuspended with binding buffer. Then, the soluble protein was purified using the 6 × His-Tagged Protein Purification Kit (Cwbio, China) according to the manufacturer’s specifications. The removal and monitoring of endotoxin residues were achieved using the ToxinEraserTM Endotoxin Removal Kit and the ToxinSensorTM Chromogenic LAL Endotoxin Assay Kit (GenScript), respectively, following the user manuals. The expression of the 6 × His-Tagged cTGF-β1 was confirmed on a 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and was visualized by staining with Coomassie brilliant blue. The protein concentration was measured using a NanoDrop 2000 spectrophotometer with an absorption at 280 nm.
A western blot assay was conducted to identify the fusion protein. Briefly, the purified sample was separated by SDS-PAGE using a 12.5% polyacrylamide gel and transferred to a nitrocellulose membrane (Life Technologies, USA) at 150 mA for 1.5 h in transfer buffer. The membrane was incubated overnight at 4 °C in the presence of anti-6 × His mouse antibody (Life Technologies, USA) after blocking with 5% (w/v) nonfat dry milk. The membrane was subsequently washed with TBST buffer, incubated with AP-goat anti-Mouse IgG and developed using NBT-BCIP.
Isolation of culter peripheral blood leukocytes (PBLs)
Blood samples were obtained from the caudal vein of culter using a heparinized syringe. The blood samples were then diluted with an equal volume Hanks’ balanced salt solution and layered onto a Percoll (Sigma) discontinuous density gradient of 60%/67%. The samples were then centrifuged at 400 × g for 30 min at 4 °C. The cell layers at the interphase were transferred to another tube, an equal volume of Hanks’ solution was added, and the samples were centrifuged at 200 × g for 10 min at 4 °C. The cells were collected, counted using a haemocytometer under a light microscope, and seeded in a 24-well plate at 5 × 106 cells/well.
Drug treatment assay
Drug treatments were performed using different test substances, including lipopolysaccharide (LPS) (10 μg/ml) (Sigma-Aldrich) and recombinant culter TGF-β1 (100 ng/ml). After treatment with the drug for 6 hours, the cells were collected from the culture plates, centrifuged (10,000 × g for 1 min) and immediately frozen until further use. In the control, no inducer was added.
To assess the role of the TGF-β1 molecule in the culter immunity in vivo, an intraperitoneal injection experiment was conducted, as previously described40. Briefly, eighty fish were equally divided into two groups: in one group, each fish was induced with approximately 2.5 mg/kg fish poly I:C; in the other group, each fish was injected with the same volume of 0.65% physiological saline as the control. Four fish from each group were euthanized at 0 h, 4 h, 12 h, 24 h, 48 h, 72 h, 96 h and 120 h after injection. At each time point, the spleen and thymus samples were dissected and frozen.
qPCR and expression analysis
The specific primers for qPCR of TGF-β1, TNF-α and IL-1β in the topmouth culter were designed using Primer Premier 5.0. To verify the specificity, the primers were first tested in a normal PCR amplification. The PCR products were visualized on a 2% agarose gel. The qPCR were carried out in a final volume of 10 μl consisting of 1 × Platinum® SYBR® Green qPCR SuperMix-UDG (Life Technologies), 0.2 μM of each primer, and 10 ng of cDNA template. The reactions were run in triplicate. The ABI PRISM 7500 HT platform (Life Technologies, USA) was used for all real-time assays. The reactions were initially denatured at 95 °C for 10 sec, followed by 40 cycles of 95 °C for 10 sec and annealing (60 °C for TGF-β1 and IL-1β, 62 °C for TNF-α) 30 sec. A melting curve analysis was performed at the end of assay to assess the amplification specificity. The relative expression levels were measured using the 2−ΔΔCt method41 with β-action as an internal reference. Statistical significance of the data was analysed using a one-way ANOVA, and the results were deemed to be significant at P < 0.05.
Additional Information
How to cite this article: Qi, P. et al. Dissecting the role of transforming growth factor-β1 in topmouth culter immunobiological activity: a fundamental functional analysis. Sci. Rep. 6, 27179; doi: 10.1038/srep27179 (2016).
Supplementary Material
Acknowledgments
This research was supported by the National Key Technology R&D Program Foundation of China (2012AA10A403) and National Key Technology Support Program of China (2011BAD13B08).
Footnotes
Author Contributions P.Q., C.X. and C.W. designed research; P.Q. and B.G. performed research; P.Q. and B.G. analyzed data; and P.Q. wrote the paper.
References
- Li M. O., Wan Y. Y., Sanjabi S., Robertson A. K. & Flavell R. A. Transforming growth factor-beta regulation of immune responses. Annu Rev Immunol 24, 99–146 (2006). [DOI] [PubMed] [Google Scholar]
- Takehara K. Growth regulation of skin fibroblasts. J Dermatol Sci 24 (Suppl 1), 70–77 (2000). [DOI] [PubMed] [Google Scholar]
- Haddad G., Hanington P. C., Wilson E. C., Grayfer L. & Belosevic M. Molecular and functional characterization of goldfish (Carassius auratus L.) transforming growth factor beta. Dev Comp Immunol 32, 654–663 (2008). [DOI] [PubMed] [Google Scholar]
- Massagué J. & Wotton D. Transcriptional control by the TGF-β/Smad signaling system. EMBO J 19, 1745–54 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGeachy M. J. & Cua D. J. T cells doing it for themselves: TGF-beta regulation of Th1 and Th17 cells. Immunity 26, 547–549 (2007). [DOI] [PubMed] [Google Scholar]
- Strobl H. & Knapp W. TGF-beta1 regulation of dendritic cells. Microbes. Infect 1, 1283–1290 (1999). [DOI] [PubMed] [Google Scholar]
- Cazac B. B. & Roes J. TGF-beta receptor controls B cell responsiveness and induction of IgA in vivo. Immunity 13, 443–451 (2000). [DOI] [PubMed] [Google Scholar]
- Laouar Y., Sutterwala F. S., Gorelik L. & Flavell R. A. Transforming growth factor-beta controls T helper type 1 cell development through regulation of natural killer cell interferon-gamma. Nat Immunol 6, 600–607 (2005). [DOI] [PubMed] [Google Scholar]
- Ashcroft G. S. Bidirectional regulation of macrophage function by TGF-beta. Microbes Infect 1, 1275–1282 (1999). [DOI] [PubMed] [Google Scholar]
- Bauvois B., Rouillard D., Sanceau J. & Wietzerbin J. IFN-gammaand transforming growth factor-beta 1 differently regulate fibronectin and laminin receptors of human differentiating monocytic cells. J Immunol 148, 3912–3919 (1992). [PubMed] [Google Scholar]
- Wahl S. M., Costa G. L., Corcoran M., Wahl L. M. & Berger A. E. Transforming growth factor-beta mediates IL-1-dependentinduction of IL-1 receptor antagonist. J Immunol 150, 3553–3560 (1993a). [PubMed] [Google Scholar]
- Wahl S. M. et al. Role of transforming growth factor beta in the pathophysiology of chronic inflammation. J Periodontol 64, 450–455 (1993b). [PubMed] [Google Scholar]
- Wahl S. M. et al. Transforming growth factor type beta induces monocyte chemotaxis and growth factor production. Proc Natl Acad Sci USA 84, 5788–5792 (1987). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiseman D. M., Polverini P. J., Kamp D. W. & Leibovich S. J. Transforming growth factor-beta (TGF beta) is chemotactic for human monocytes and induces their expression of angiogenic activity. Biochem Biophys Res Commun 157, 793–800 (1988). [DOI] [PubMed] [Google Scholar]
- Zhan Y. & Jimmy K. Molecular isolation and characterization of carp transforming growth factor beta 1 from activated leucocytes. Fish Shellfish Immunol 10, 309–318 (2000). [DOI] [PubMed] [Google Scholar]
- Harms C. A. Kennedy-stoskopf, S., Horne, W. A., Fuller, F. J. & Tompkins, W. A. F. Cloning and sequencing hybrid striped bass (Morone saxatilis × M. chrysops) transforming growth factor-beta (TGF-beta), and development of a reverse transcription quantitative competitive polymerase chain reaction (RT-qPCR) assay to measure TGF-beta mRNA of teleost fish. Fish Shellfish Immunol 10, 61–85 (2000). [DOI] [PubMed] [Google Scholar]
- Tafalla C. et al. Molecular characterisation of sea bream (Sparus aurata) transforming growth factor β1. Fish Shellfish Immunol 14, 405–421 (2003). [DOI] [PubMed] [Google Scholar]
- Yang M. & Zhou H. Grass carp transforming growth factor-beta 1 (TGF-β1): molecular cloning, tissue distribution and immunobiological activity in teleost peripheral blood lymphocytes. Mol Immunol 45, 1792–1798 (2008). [DOI] [PubMed] [Google Scholar]
- Newfeld S. J., Wisotzkey R. G. & Kumar S. Molecular evolution of a developmental pathway: phylogenetic analyses of transforming growth factor-beta family ligands, receptors and Smadsignal transducers. Genetics 152, 783–95 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardie L. J. et al. Isolation of the first teleost transforming growth factor beta gene: analysis reveals tissue specific expression and a potential regulatory sequence in rainbow trout (Oncorhynchus mykiss). Cytokine 10, 555–563 (1998). [DOI] [PubMed] [Google Scholar]
- Zapata A., Diez B., Cejalvo C., Gutierrez-de F. & Cortes A. Ontogeny of the immune system of fish. Fish Shellfish Immunol 20, 126–136 (2006). [DOI] [PubMed] [Google Scholar]
- Letterio J. J. & Roberts A. B. Regulation of immune responses by TGF-β. Annu Rev Immunol 16, 137–161 (1998). [DOI] [PubMed] [Google Scholar]
- Le Y. Y. et al. The immunopharmacological properties of transforming growth factor beta. Int Immunopharmacol 5, 1771–1782 (2005). [DOI] [PubMed] [Google Scholar]
- Attisano L. & Wrana J. L. Signal transduction by the TGF-beta superfamily. Science 296, 1646–7 (2002). [DOI] [PubMed] [Google Scholar]
- Wahl S. M. TGF-beta in the evolution and resolution of in flammatory and immune processes. Introduction. Microbes Infect 1, 1247–1249 (1999). [DOI] [PubMed] [Google Scholar]
- Letterio J. J. & Roberts A. B. TGF-beta: a critical modulator of immune cell function. Clin. Immunol Immunopathol 84, 244–250 (1997). [DOI] [PubMed] [Google Scholar]
- Wan Y. Y. & Flavell R. A. ‘Yin-Yang’ functions of transforming growth factor-beta and T regulatory cells in immune regulation. Immunol Rev 220, 199–213 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wahl S. M. Transforming growth factor-beta: innately bipolar. Curr Opin Immunol 19, 55–62 (2007). [DOI] [PubMed] [Google Scholar]
- Jang S. I., Hardie L. J. & Secombes C. J. Effects of transforming growth factor beta 1 onrainbow trout Oncorhynchus mykiss macrophage respiratory burst activity. Dev Comp Immunol 18, 315–323 (1994). [DOI] [PubMed] [Google Scholar]
- Yang M., Wang Y., Wang X., Chen C. & Zhou H. Characterization of grass carp (Ctenopharyngodon idellus) Foxp1a/1b/2: evidence for their involvement in the activation of peripheral blood lymphocyte subpopulations. Fish Shellfish Immunol 28, 289–295 (2009). [DOI] [PubMed] [Google Scholar]
- Cai Z. H., Gao C. P., Li L. & Xing K. Z. Bipolar properties of red seabream (Pagrus major) transforming growth factor-β in induction of the leucocytes migration. Fish Shellfish Immunol 28, 695–700 (2010). [DOI] [PubMed] [Google Scholar]
- Ping H. L., Wu J. Y., Xu S. W., Hu K. S. & Duan Z. G. Molecular Cloning and Expression analysis of Átransforming growth factorĈβ1 (TGF-β1) from Orange Spotted Grouper (Epinephelus coioides). Journal of Sun Yetsen University 50, 92–8 (Chinese with English abstract) (2011). [Google Scholar]
- Holopainen R., Tapiovaara H. & Honkanen J. Expression analysis of immune response genes in fish epithelial cells following ranavirus infection. Fish Shellfish Immunol 32, 1095–1105 (2012). [DOI] [PubMed] [Google Scholar]
- Chilmonczyk S. The thymus in fish: development and possible function in the immune response. Annual Review Fish Diseases 2, 181–200 (1992). [Google Scholar]
- Boehm T. & Bleul C. C. The evolutionary history of lymphoid organs. Nat Immunol 8, 131–135 (2007). [DOI] [PubMed] [Google Scholar]
- Jósefsson S. & Tatner M. F. Histogenesis of the lymphoid organs in sea bream (Sparus aurata L.). Fish Shellfish Immunol 3, 35–49 (1993). [Google Scholar]
- Zwollo P., Cole S., Bromage E. & Kaattari S. B cell heterogeneity in the teleost kidney: evidence for a maturation gradient from anterior to posterior kidney. J Immunol 174, 6608–6616 (2005). [DOI] [PubMed] [Google Scholar]
- Thompson J. D., Gibson T. J., Plewniak F., Jeanmougin F. & Higgins D. G. The CLUSTAL-X windows interface, flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl Acids Res 25, 4876–4882 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamura K. et al. MEGA 5, molecular evolutionary genetics analysis using maximum Likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28, 2731–2739 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi P. Z., Guo B. Y., Zhu A. Y., Wu C. W. & Liu C. L. Identification and comparative analysis of the (Pseudosciaena crocea) microRNA transcriptome response to poly (I:C) infection using a deep sequencing approach. Fish Shellfish Immunol 39, 483–491 (2014). [DOI] [PubMed] [Google Scholar]
- Livak K. H. & Schmittgen T. D. Analysis of the realtime gene expression data using Realtime Quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408 (2001). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.