

Abstract
Background
Previous studies have developed models predicting methylation age from DNA methylation in blood and other tissues (epigenetic clock) and suggested the difference between DNA methylation and chronological ages as a marker of healthy aging. The goal of this study was to confirm and expand such observations by investigating whether different concepts of the epigenetic clocks in a population-based cohort are associated with cancer, cardiovascular, and all-cause mortality.
Results
DNA methylation age was estimated in a cohort of 1863 older people, and the difference between age predicted by DNA methylation and chronological age (Δage) was calculated. A case-cohort design and weighted proportional Cox hazard models were used to estimate associations of Δage with cancer, cardiovascular, and all-cause mortality. Hazard ratios for Δage (per 5 years) calculated using the epigenetic clock developed by Horvath were 1.23 (95 % CI 1.10–1.38) for all-cause mortality, 1.22 (95 % CI 1.03–1.45) for cancer mortality, and 1.19 (95 % CI 0.98–1.43) for cardiovascular mortality after adjustment for batch effects, age, sex, educational level, history of chronic diseases, hypertension, smoking status, body mass index, and leucocyte distribution. Associations were similar but weaker for Δage calculated using the epigenetic clock developed by Hannum.
Conclusions
These results show that age acceleration in terms of the difference between age predicted by DNA methylation and chronological age is an independent predictor of all-cause and cause-specific mortality and may be useful as a general marker of healthy aging.
Electronic supplementary material
The online version of this article (doi:10.1186/s13148-016-0228-z) contains supplementary material, which is available to authorized users.
Keywords: DNA methylation age, Epigenetic clock, Epigenetic age acceleration, Mortality risk
Background
DNA methylation is a form of epigenetic control involved in several cellular processes and in the regulation of gene expression, especially through gene silencing [1]. Modifications of DNA methylation are influenced by genetic and environmental factors, including age, a phenomenon called “epigenetic drift” [2]. Such age-related modifications are bidirectional (they both show an increase and a decrease in methylation, depending on the sites), vary between individuals of the same age, and differ across the genome [3].
Several models, also called “epigenetic clock,” have been developed, predicting chronological age from DNA methylation (DNAmage) with high accuracy [4, 5]. An Italian study showed that semi-supercentenarians had a lower epigenetic age than age-matched controls and that their offspring’s epigenetic age was systematically lower than their chronological age [6], suggesting that epigenetic processes play a role in healthy aging. An increasing number of studies indicate that the discrepancy between DNAmage and chronological age may be associated with age-related diseases and mortality. A few studies indicated that epigenetic age acceleration (i.e., predicted methylation age exceeding chronological age) was significantly associated with increased mortality risk [7–9], frailty [10], and lower cognitive performance [11], including neuropathological markers of Alzheimer’s disease [12]. Age acceleration was also observed in brain tissues of people affected by Trisomy 21, who have a well-known increased risk of developing Alzheimer’s disease [13] and there is emerging evidence that blood epigenetic age may predict cancer incidence [14] and mortality [15].
This literature seems to suggest that age acceleration in terms of the difference between age predicted by DNA methylation and chronological age could be a useful marker of healthy aging. However, in order to corroborate such findings, it is of importance to replicate the above results in different population-based cohorts and, additionally, to explore whether epigenetic age acceleration is also associated with other relevant age-related outcomes, such as cardiovascular disease (CVD) mortality.
We aimed to investigate whether the discrepancy between DNAmage as predicted by the models of Hannum et al. [4] (hereinafter also DNAmage (Hannum)) and Horvath [5] (hereinafter also DNAmage (Horvath)) and chronological age is associated with cancer, cardiovascular, and all-cause mortality in a population-based cohort of older people.
Methods
Study population and study design
Measurements of DNA methylation in blood were performed in a baseline subsample of the ESTHER cohort, which is a large population-based epidemiological study conducted in the German State of Saarland with the aim of assessing chances of prevention and early detection of chronic diseases. As described in more detail in Saum et al. [16], between July 2000 and December 2002, general practitioners recruited 9949 older adults aged 50–75 years during a regular health check-up.
Socio-demographic, lifestyle, and health characteristics were collected using standardized self-administered questionnaires. Self-reported diagnoses of prevalent cardiovascular diseases, cancer, hypertension, and diabetes mellitus were validated through medical records. Information on survival status was obtained by record linkage with population registries. Cause of death was ascertained according to death certificate. All participants provided blood samples, which were stored at −80 °C. The Ethics Committee of the Medical Faculty of the University of Heidelberg and of the Physician Board of Saarland approved the study and participation was conditional on written informed consent.
Due to financial constraints, DNA was extracted only from a subsample of 3499 ESTHER participants who were recruited in the initial phase of the enrolment between July 2000 and March 2001 and who therefore had the longest follow-up time. Among these, epigenetic measurements were conducted in a randomly selected subcohort of 1548 participants irrespective of survival status as well as in all participants who had died during follow-up (cases) and did not overlap with the randomly selected subcohort (N = 316). In total, 1864 participants with epigenetic measurements were therefore available for the present case-cohort study.
DNA methylation measurement
Blood DNA was extracted using a salting out procedure [17] and measured using PicoGreen (Invitrogen, Darmstadt, Germany). DNA methylation profiles were measured using the Infinium HumanMethylation450 BeadChip (Illumina Inc.) in the Genomics and Proteomics Core Facility at the German Cancer Research Center, Heidelberg. As quality control, three random DNA samples for each plate were used as duplicates for replication. Methylation levels at each CpG were calculated with Illumina’s Genomestudio 2011.1, Modul M Version 1.9.0 as previously described in detail [18].
Prediction of DNAmage
DNAmage (Horvath) was calculated using the R tutorial of the pertinent publication [5] and DNAmage (Hannum) as the sum of the beta values multiplied by the respective regression coefficients reported by Hannum et al. [4] as already described by Marioni et al. [7]. For all calculations of DNAmage, we only used signals of probes with a detection p value ≤0.05.
Statistical analysis
We used descriptive statistics to show the distribution of socio-demographic variables and prevalent diseases at baseline separately among the participants of the randomly selected subcohort who survived over the follow-up and among the cases. We also differentiated the cases between those participants of the subcohort who deceased during the follow-up (cases within the subcohort) and those who were specifically selected among deceased participants (cases outside the subcohort).
Following Marioni et al. [7], we calculated the difference between age predicted by DNA methylation and chronological age (Δage; epigenetic age acceleration) and estimated hazard ratios (HR) for the association of Δage (per 5 years of age acceleration) with all-cause and cause-specific mortality. HRs were estimated using the SAS® program code implementing weighted proportional Cox hazard models as illustrated in detail by Kulathinal and colleagues [19]. Effect estimates were adjusted in model 1 for batch effects, chronological age (continuous), sex, and leucocyte distribution including the following cell types: CD56 natural killer cells, CD14 monocytes, CD4 T-cells, CD8 T-cells, CD19 B-cells, and granulocytes. Proportions of leucocyte cells were estimated from DNA methylation using the algorithm from Houseman et al. [20]. Model 2 was additionally adjusted for educational level, history of cancer, history of CVD, hypertension, diabetes mellitus, smoking status, and body mass index. Sensitivity analyses were conducted without adding leucocyte distribution as a covariate in model 1.
Scatter plots were generated in order to illustrate the relationship between chronological age and DNAmage. Pearson correlation coefficient (r) between chronological and methylation age were calculated both for DNAmage (Horvath) and DNAmage (Hannum).
In order to explore specific biological components of the epigenetic clock associated with mortality, we estimated, in Cox models adjusted for chronological age, sex, leucocyte distribution, and batch effects, which of the 353 CpGs of Horvath’s epigenetic clock were significantly associated with all-cause mortality. Multiple testing adjustments were performed both by controlling the false discovery rate (FDR) and using the Bonferroni correction.
With the exception of DNAmage (Horvath), which we calculated using R software version 3.1.1, we conducted all analyses using SAS® statistical software version 9.4 (SAS® Institute Inc., Cary, NC, USA).
Results
The subsample of the ESTHER cohort consisted of 1863 participants with 1260 (67.6 %) survivors and 602 (32.3 %) cases (316 selected cases and 286 deceased participants within the subcohort) and 1 participant with missing information relating to survival status (Table 1). Overall, we recorded 235 (12.6 %) deaths due to cancer (N = 95 among women and N = 140 among men) and 194 (10.4 %) cardiovascular deaths (N = 80 among women and N = 114 among men). The majority of deceased participants were men both among cases outside the subcohort (56.3 %) and within the subcohort (60.5 %). Cases showed more age-related diseases at baseline, such as CVD, hypertension, and diabetes mellitus, than survivors. Also, there were more current smokers among cases within (27.0 %) and outside (24.3 %) the subcohort than among surviving participants (16.4 %).
Table 1.
Distribution of socio-demographic variables and prevalent diseases at baseline among participants of the case cohort (ESTHER study 2000–2001)
| Variable | Survivors | Cases within the subcohorta | Cases outside the subcohortb |
|---|---|---|---|
| Sex | |||
| Female | 721 (57.2) | 113 (39.5) | 138 (43.7) |
| Men | 539 (42.8) | 173 (60.5) | 178 (56.3) |
| Age | |||
| 50–65 | 905 (71.8) | 123 (43.0) | 163 (51.6) |
| 66–75 | 355 (28.2) | 163 (57.0) | 153 (48.4) |
| Years of school education | |||
| >9 years | 334 (27.0) | 56 (20.4) | 63 (20.5) |
| ≤9 years | 902 (73.0) | 218 (80.6) | 244 (79.5) |
| History of cancer disease | |||
| No | 1201 (95.3) | 254 (88.8) | 268 (84.8) |
| Yes | 59 (4.7) | 32 (11.1) | 48 (15.2) |
| History of cardiovascular disease | |||
| No | 1048 (83.2) | 171 (59.8) | 206 (65.2) |
| Yes | 212 (16.8) | 115 (40.2) | 110 (34.8) |
| Hypertension | |||
| No | 714 (56.7) | 142 (50.5) | 154 (49.2) |
| Yes | 545 (43.3) | 139 (49.5) | 159 (50.8) |
| Diabetes mellitus | |||
| No | 1068 (85.9) | 216 (76.1) | 227 (73.0) |
| Yes | 176 (14.2) | 68 (23.9) | 84 (27.0) |
| Body mass index (kg/m2) | |||
| <25 | 341 (27.1) | 79 (27.6) | 97 (30.7) |
| 25–<30 | 594 (47.1) | 124 (43.4) | 132 (41.8) |
| ≥30 | 325 (25.8) | 83 (29.0) | 87 (27.5) |
| Smoking | |||
| Never/former | 1034 (83.6) | 203 (73.0) | 233 (75.7) |
| Current | 203 (16.4) | 75 (27.0) | 75 (24.3) |
aParticipants of the randomly selected subcohort who were deceased during the follow-up (2000–2013)
bParticipants specifically selected among deceased participants
Mean chronological age at baseline in the total study population was 62.5 (SD 6.6; min 48.0, max 75.0). Mean of DNAmage (Horvath) was 63.0 (SD = 7.5; min = 38.9 max 95.7) and mean of DNAmage (Hannum) was 68.6 (SD = 7.1; min = 47.1 max 95.9). Pearson correlation coefficient between chronological and methylation age was 0.73 (p < 0.0001) for DNAmage (Horvath) and 0.77 (p < 0.0001) for DNAmage (Hannum). Panels a and b of Fig. 1 display scatter plots showing the relationship between chronological and DNAmage both among deceased participants (red circles) and those alive at last follow-up (black circles). Also, there was a strong linear relationship between DNAmage (Horvath) and DNAmage (Hannum) (r = 0.80; p < 0.0001).
Fig. 1.
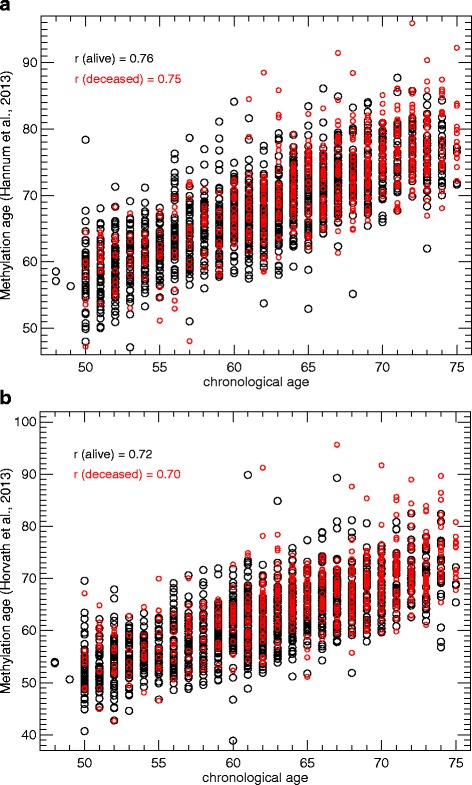
a Relationship between DNA methylation age estimated with Hannum’s predictor and chronological age. b Relationship between DNA methylation age estimated with Horvath’s predictor and chronological age
Weighted proportional Cox hazard models revealed that irrespective of which predictor was used, a DNA methylation age higher than chronological age was associated with higher mortality. In Cox models adjusted for age, sex, batch effects, and leucocyte distribution HRs for the association of Δage (per 5 years) with all-cause mortality showed a statistically significant increase in mortality by 22 % using DNAmage (Horvath) and an increase in mortality by 16 % using DNAmage (Hannum) (Table 2). In the fully adjusted models, HR estimates for Δage were stronger for DNAmage (Horvath) (HR = 1.23; 95 % CI 1.10–1.38) than for DNAmage (Hannum), which were no longer statistically significant (HR = 1.10; 95 % CI 0.94–1.29).
Table 2.
Associations of differences between Δage (per 5 years) according to different predictors of DNAmage with all-cause and cause-specific mortality
| Causes of death (N of events; %) |
Predictor | Cox model 1a
(Hazard ratios) |
Cox model 2b
(Hazard ratios) |
|---|---|---|---|
| All-cause mortality | |||
| (602; 32.3) | DNAmage (Horvath) | 1.22 (1.10–1.36) | 1.23 (1.10–1.38) |
| (602; 32.3) | DNAmage (Hannum) | 1.16 (1.00–1.34)* | 1.10 (0.94–1.29) |
| Cancer mortality | |||
| (235; 12.6) | DNAmage (Horvath) | 1.20 (1.03–1.39) | 1.22 (1.03–1.45) |
| (235; 12.6) | DNAmage (Hannum) | 1.08 (0.87–1.35) | 1.03 (0.80–1.33) |
| Cardiovascular disease mortality | |||
| (194; 10.4) | DNAmage (Horvath) | 1.20 (1.02–1.42) | 1.19 (0.98–1.43) |
| (194; 10.4) | DNAmage (Hannum) | 1.13 (0.89–1.44) | 1.00 (0.79–1.29) |
*p = 0.0408
aModel 1: adjusted for chronological age (continuous), sex, batch effects, and leucocyte distribution
bModel 2: additionally adjusted for educational level, history of cancer diseases, history of CVD, hypertension, diabetes mellitus, smoking status (never/former vs. current), and body mass index (continuous)
Epigenetic age acceleration was also associated with higher cancer and CVD mortality in Cox models using Horvath’s predictor and adjusted for age, sex, batch effects, and leucocyte distribution (HR = 1.20; 95 % CI 1.03–1.39 and HR = 1.20; 95 % CI 1.02–1.42, respectively). Cox models for DNAmage (Hannum) also showed an increased mortality both for cancer and CVD mortality in partially adjusted models but estimates were weaker and they were not statistically significant (HR = 1.08; 95 % CI 0.87–1.35 and HR = 1.13; 95 % CI 0.89–1.44, respectively).
Associations of epigenetic age acceleration with all-cause and cause-specific mortality were very similar among women and men (Table 3).
Table 3.
Associations of differences between Δage (per 5 years) according to different predictors of DNAmage with all-cause and cause-specific mortality by sex
| Number of events; % | Predictor | Cox model 1a
(Hazard ratios) |
Cox model 1b
(Hazard ratios) |
|
|---|---|---|---|---|
| All-cause mortality | ||||
| Women | 251; 41.7 | DNAmage (Horvath) | 1.27 (1.07–1.52) | 1.24 (1.02–1.52) |
| DNAmage (Hannum) | 1.19 (0.94–1.51) | 1.05 (0.82–1.36) | ||
| Men | 351; 58.3 | DNAmage (Horvath) | 1.23 (1.05–1.43) | 1.28 (1.09–1.51) |
| DNAmage (Hannum) | 1.14 (0.94–1.40) | 1.14 (0.92–1.41) | ||
| Cancer mortality | ||||
| Women | 95; 40.4 | DNAmage (Horvath) | 1.20 (0.93–1.54) | 1.21 (0.88–1.65) |
| DNAmage (Hannum) | 1.02 (0.69–1.50) | 0.89 (0.58–1.36) | ||
| Men | 140; 59.6 | DNAmage (Horvath) | 1.18 (0.95–1.47) | 1.25 (0.98–1.59) |
| DNAmage (Hannum) | 1.08 (0.80–1.46) | 1.12 (0.79–1.58) | ||
| CVD mortality | ||||
| Women | 80; 41.2 | DNAmage (Horvath) | 1.17 (0.91–1.51) | 1.13 (0.82–1.55) |
| DNAmage (Hannum) | 1.21 (0.85–1.72) | 1.01 (0.64–1.61) | ||
| Men | 114; 58.8 | DNAmage (Horvath) | 1.25 (0.98–1.59) | 1.29 (0.99–1.68) |
| DNAmage (Hannum) | 1.06 (0.77–1.47) | 1.00 (0.71–1.42) |
CVD cardiovascular disease
aModel 1: adjusted for chronological age (continuous), sex, batch effects, and leucocyte distribution
bModel 2: additionally adjusted for educational level, history of cancer diseases, history of CVD, hypertension, diabetes mellitus, smoking status (never/former vs. current), and Body Mass Index (continuous)
Sensitivity analyses conducted without adding leucocyte distribution as covariate in model 1 revealed weaker point estimates for DNAmage (Horvath) (HR(all-cause mortality) = 1.17; 95 % CI 1.06–1.30; HR (cancer mortality) = 1.15; 95 % CI 0.99–1.34; HR(CVD mortality) = 1.15; 95 % CI 0.98–1.34) and stronger estimates for DNAmage (Hannum) (HR(all-cause mortality) = 1.21; 95 % CI 1.06–1.38; HR (cancer mortality) = 1.11; 95 % CI 0.91–1.37; HR(CVD mortality) = 1.20; 95 % CI 0.96–1.50) .
Cox regression analyses for the 353 CpGs of Horvath’s epigenetic clock revealed that 6 CpGs (cg19724470 in CD27; cg15804973 in MAP3K5, cg25564800 in KPNA1; cg01820374 in LAG3; cg01511567 in SSRP1; cg26614073 in SCAP) showed a statistically significant association with mortality after FDR adjustment and 3 CpGs after Bonferroni correction (Additional file 1: Table S1). Without adjustment for multiple testing, we found that 33 CpGs were associated with mortality.
Discussion
In this cohort of older adults, epigenetic age acceleration in terms of the difference between methylation age and chronological age was associated with increased total, cancer, and CVD mortality even after adjustment for a large number of potential confounders. These findings strengthen suggestions that epigenetic age acceleration may be a general (bio-)marker of healthy aging.
Being the first study addressing CVD mortality, we cannot compare our results relating to such outcome with other studies, but our findings relating to all-cause mortality were quite comparable with those from a recent publication. In particular, Marioni and colleagues [7] found that Δage (per 5 years) was significantly associated with a 21 % greater mortality risk in a model adjusted for chronological age and sex using DNAmage (Hannum) (HR = 1.21; 95 % CI 1.14–1.29) and with a 11 % increase using DNAmage (Horvath) (1.11; 95 % CI 1.05–1.18). The consistent results obtained for all-cause mortality reinforce suggestions that epigenetic age acceleration may be regarded as a possible candidate biomarker of healthy aging.
In our main analysis with adjustment for leucocyte distribution, the association between Δage and mortality was stronger for DNAmage (Horvath) than for DNAmage (Hannum). In sensitivity analyses without adjustment for leucocyte distribution, point estimates for DNAmage (Hannum), but not for DNAmage (Horvath), were stronger than the corresponding point estimates adjusted for leukocyte distribution. This seems to suggest that Horvath’s predictor, being developed from several human tissues and cell types and not only from blood, might be more robust than the blood-based predictor from Hannum and also less sensitive to changes in blood cell composition, which could mediate the association between Δage and mortality [7]. However, given that the direction of the association remained stable and that there was only a moderate change in the strength of the association for both predictors, we can infer that leucocyte distribution did not have a major influence on the association between epigenetic clocks and mortality in our cohort.
Mean DNAmage predicted by the Hannum’s predictor was approximately 6 years higher than mean chronological age, an overestimation also observed in the cohorts analyzed by Marioni and colleagues [7]. Given the very good correlation between methylation and chronological age and the strong linear relationship between DNAmage (Horvath) and DNAmage (Hannum), we did not deem it necessary to account for the overestimation.
Our findings indicating associations between Δage and cancer mortality are in line with studies showing that methylation changes might represent an early event in the development of cancerous cells [2, 3] and be able to predict cancer incidence [14, 15]. Our results suggest that epigenetic age acceleration might also be an indicator for a more aggressive course of tumor disease with increased risk of a cancer-related fatal event, which supports the results of Zheng et al. [15] who also found that the discrepancy between epigenetic and chronological ages predicted cancer mortality.
Our findings revealing that epigenetic age acceleration was also associated with a 20 % greater CVD mortality risk (for estimates based on Horvath’s predictor) reveal a magnitude of association comparable to the observed association related to cancer mortality, even if CVD mortality was less frequent than cancer mortality (194 vs 235, respectively) and therefore CVD mortality models had less statistical power.
A possible explanation for the observed similar magnitude of association is that, similar to cancer, the development of CVD also involves focal proliferative events [3, 21] and this might point to a relationship between aberrant methylation in older age and focal proliferation processes. In particular, on the one hand, an abnormal epigenetic drift might lead to an aggravation of focal proliferative events leading, inter alia, to cardiovascular and tumor diseases, and on the other hand, focal proliferative events might lead to further abnormal epigenetic drift [3] with exacerbation of the cardiovascular and cancer pathologies leading eventually to higher mortality. The design of our study with epigenetic measurements obtained several years prior to the event supports a role of epigenetic changes leading to the exacerbation of cancer and CVD disease, but it cannot definitively exclude reciprocal influences, as the event leading to greater mortality might have already affected the epigenetic status at an early stage of the disease and be reflected in the epigenetic measurements.
Lin and colleagues [9] tested associations of single CpG-derived age predictions with mortality in the LBC1936 cohort. Two CpGs that showed significant results before adjustment for multiple testing were overlapping with our findings (cg19724470 and cg15804973). Further studies should explore biological properties of such genes and the possible pathways leading to early mortality.
Strengths of our study were analyses conducted with two different predictors, the large and representative sample size, and adjustment for important confounders; main limitations were the lack of differential blood counts and the impossibility to ascertain in our population the exact temporal relationship between epigenetic age acceleration and focal proliferative events.
Conclusions
In conclusion, our study further confirms the validity of estimates of DNAmage based on Horvath’s and Hannum’s epigenetic clocks, it expands previous observations by including CVD mortality, and it indicates that epigenetic age acceleration might be both a useful marker of healthy aging among older people as well as a prognostic marker.
Acknowledgements
We are deeply indebted to Utz Bescheid for his support in the preparation and analysis of epigenetic data.
Availability of data and materials
Data protection standards, which were part of the informed consent procedure of the ESTHER study, preclude that data be deposited in publicly available repositories. Individual data access may be granted within a framework of scientific cooperation.
Authors’ contributions
LP designed the study and drafted the manuscript. YZ und LP carried out the statistical analysis. UM contributed to data acquisition and participated in data analysis. BH and KUS contributed to the coordination of the ESTHER study. HB conducted the ESTHER study, contributed to the conception of the study, and supervised the analysis. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Additional file
Associations of single CpG sites with all-cause mortality. (XLSX 12 kb)
References
- 1.Simmons D. Epigenetic influences and disease. Nat Educ. 2008;1:6. [Google Scholar]
- 2.Teschendorff AE, West J, Beck S. Age-associated epigenetic drift: implications, and a case of epigenetic thrift? Hum Mol Genet. 2013;22:R7–15. doi: 10.1093/hmg/ddt375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Issa JP. Aging and epigenetic drift: a vicious cycle. J Clin Invest. 2014;124:24–9. doi: 10.1172/JCI69735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hannum G, Guinney J, Zhao L, Zhang L, Hughes G, Sadda S, et al. Genome-wide methylation profiles reveal quantitatives views of human aging rates. Mol Cell. 2013;49:359–67. doi: 10.1016/j.molcel.2012.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Horvath S. DNA methylation age of human tissues and cell types. Genome Biol. 2013;14:R115. doi: 10.1186/gb-2013-14-10-r115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Horvath S, Pirazzini C, Bacalini MG, Gentilini D, Di Blasio AM, Delledonne M, et al. Decreased epigenetic age of PBMCs from Italian semi-supercentenarians and their offspring. Aging. 2015;7:1159–70. doi: 10.18632/aging.100861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Marioni RE, Shah S, McRae AF, Chen BH, Colicino E, Harris SE, et al. DNA methylation age of blood predicts all-cause mortality in later life. Genome Biol. 2015;16:25. doi: 10.1186/s13059-015-0584-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Christiansen L, Lenart A, Tan Q, Vaupel JW, Aviv A, McGue M, et al. DNA methylation age is associated with mortality in a longitudinal Danish twin study. Aging Cell. 2016;15:149–54. doi: 10.1111/acel.12421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lin Q, Weidner CI, Costa IG, Marioni RE, Ferreira MRP, Deary IJ, et al. DNA methylation levels at individual age-associated CpG sites can be indicative for life expectancy. Aging. 2016;8:394–401. doi: 10.18632/aging.100908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Breitling LP, Saum KU, Perna L, Schöttker B, Holleczek B, Brenner H. Frailty is associated with the epigenetic clock but not with telomere length in a German cohort. Clin Epigenetics. 2016;8:21. doi: 10.1186/s13148-016-0186-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Marioni RE, Shah S, McRae AF, Ritchie SJ, Muniz-Terrera G, Harris SE, et al. The epigenetic clock is correlated with physical and cognitive fitness in the Lothian Birth Cohort 1936. Int J Epidemiol. 2015;44:1388–96. doi: 10.1093/ije/dyu277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Levine ME, Lu AT, Bennett DA, Horvath S. Epigenetic age of the pre-frontal cortex is associated with neuritic plaques, amyloid load, and Alzheimer’s disease related cognitive functioning. Aging. 2015;7:1198–211. doi: 10.18632/aging.100864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Horvath S, Garagnani P, Bacalini MG, Pirazzini C, Salvioli S, Gentilini D, et al. Accelerated epigenetic aging in Down syndrome. Aging Cell. 2015;14:491–5. doi: 10.1111/acel.12325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Levine ME, Hosgood HD, Chen B, Absher D, Assimes T, Horvath S. DNA methylation age of blood predicts future onset of lung cancer in the women’s initiative. Aging. 2016;7:690–700. doi: 10.18632/aging.100809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zheng Y, Joyce BT, Colicino E, Liu L, Zhang W, Dai Q, et al. Blood epigenetic age may predict cancer incidence and mortality. EBioMedicine. doi:10.1016/j.ebiom.2016.02.008. [DOI] [PMC free article] [PubMed]
- 16.Saum KU, Dieffenbach AK, Jansen EH, Schöttker B, Holleczek B, Hauer K, Brenner H. Association between oxidative stress and frailty in an elderly German population: results from the ESTHER cohort study. Gerontology. 2015;61:407–15. doi: 10.1159/000380881. [DOI] [PubMed] [Google Scholar]
- 17.Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988;16:1215. doi: 10.1093/nar/16.3.1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Florath I, Butterbach K, Müller H, Bewerunge-Hudler M, Brenner H. Cross-sectional and longitudinal changes in DNA methylation with age: an epigenome-wide analysis revealing over 60 novel-age associated CpG sites. Hum Mol Genet. 2014;23:1186–201. doi: 10.1093/hmg/ddt531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kulathinal S, Karvanen J, Saarela O, Kuulasmaa K, for the MORGAM Project Case-cohort design in practice—experiences from the MORGAM Project. Epidemiol Perspect Innov. 2007;4:15. doi: 10.1186/1742-5573-4-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Houseman EA, Molitor J, Marsit CM. Reference-free cell mixture adjustments in analysis of DNA methylation data. Bioinformatics. 2014;30:1431–9. doi: 10.1093/bioinformatics/btu029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lenzi S, Descovich GC. Atherosclerosis and cardiovascular diseases. Lancaster: MTP Press Limited; 1986. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data protection standards, which were part of the informed consent procedure of the ESTHER study, preclude that data be deposited in publicly available repositories. Individual data access may be granted within a framework of scientific cooperation.