

Abstract
In the past century, noncommunicable diseases have surpassed infectious diseases as the principal cause of sickness and death, worldwide. Trillions of commensal microbes live in and on our body, and constitute the human microbiome. The vast majority of these microorganisms are maternally derived and live in the gut, where they perform functions essential to our health and survival, including: digesting food, activating certain drugs, producing short-chain fatty acids (which help to modulate gene expression by inhibiting the deacetylation of histone proteins), generating anti-inflammatory substances, and playing a fundamental role in the induction, training, and function of our immune system. Among the many roles the microbiome ultimately plays, it mitigates against untoward effects from our exposure to the environment by forming a biotic shield between us and the outside world. The importance of physical activity coupled with a balanced and healthy diet in the maintenance of our well-being has been recognized since antiquity. However, it is only recently that characterization of the host–microbiome intermetabolic and crosstalk pathways has come to the forefront in studying therapeutic design. As reviewed in this report, synthetic biology shows potential in developing microorganisms for correcting pathogenic dysbiosis (gut microbiota–host maladaptation), although this has yet to be proven. However, the development and use of small molecule drugs have a long and successful history in the clinic, with small molecule histone deacetylase inhibitors representing one relevant example already approved to treat cancer and other disorders. Moreover, preclinical research suggests that epigenetic treatment of neurological conditions holds significant promise. With the mouth being an extension of the digestive tract, it presents a readily accessible diagnostic site for the early detection of potential unhealthy pathogens resident in the gut. Taken together, the data outlined herein provide an encouraging roadmap toward important new medicines and companion diagnostic platforms in a wide range of therapeutic indications.
Key words: : α-lipoic acid, autism, dementia, dysbiosis, epigenetic, exercise, fatty acids, histone deacetylase, immune system, microbiota, mitochondria, neurodegenerative, probiotics, schizophrenia, synthetic biology
Introduction
In less than 100 years, noncommunicable diseases have surpassed infectious diseases as the principal cause of sickness and death, worldwide.1 One hundred trillion commensal microbes (including the fungal community referred to as the mycobiome)2 that live in and on our body constitute the human microbiome,3 although a recent study4 estimates the overall figure to be much lower. Regardless of the absolute number, the vast majority of these microorganisms live in the gut (microbiota),5 where they perform functions that are essential to our health and survival. They help us digest food6; participate in the activation of certain drugs7; produce short-chain fatty acids (SCFAs) that help modulate gene expression by inhibiting deacetylation of histone proteins8–10; generate molecules that reduce inflammation11; and play a fundamental role in the induction, basic development, training, and function of our immune system.12–14 Thus, as a whole, the microbiome becomes an integral part of our immune makeup, and is largely inherited from the mother with significant differences consequent to cesarean versus vaginal deliveries.15–20 Among the many roles the microbiome may ultimately play in health and disease, it mitigates against the untoward effects from our exposure to the environment by residing as a biotic barrier between us and the world around us.1,21–25
Neuropsychiatric disorders26 are on the increase globally and, of the noncommunicable diseases, stand out as a leading cause of disability.3,8,27,28 Accruing evidence strongly links gut dysbiosis (gut microbiota–host maladaptation) as a risk factor in a wide range of mental illnesses that include neuropsychiatric conditions,3,29–38 such as autism spectrum disorder (ASD)39–46 and schizophrenia39,40,46–49 among them. There are currently no drugs approved that treat the core symptoms of ASD.50 The pathogenic mechanisms underlying schizophrenia, a debilitating mental disorder, are unknown51 and drug therapies used to treat the associated psychotic symptoms have advanced little since the introduction of clozapine in 1960.52,53
The reported association of mental illness with digestive disturbances dates back to Hippocrates and stands as the single consistently linked comorbidity described in the medical literature from ancient times to the present.49 Although the genesis of our microbiome is predominantly our mother's,18,54 eventually our microbiome transforms into our own unique signature.55 Changes in the gut microbial composition and function constantly adapt to our diet,56,57 and the mechanistic relationships between the gut microbiota in the development of the enteric nervous system58 and the preservation of our metabolic health59,60 are only now beginning to be elucidated.
Microbiota-Induced Epigenetics
Advances in genetic editing technologies may help clarify whether it is our genetics that control our epigenome or epigenetics that control the genome—or, more likely, the relationship between the two is mutual.61 There is supporting evidence to suggest that our microbiome plays a fundamental role in this relationship.14,62,63 Numerous studies64,65 show microbe-generated metabolites are intertwined with host cell biochemistry and physiology, and SCFA-mediated cell signaling is a key pathway that gut microbes use to communicate with the host.9,44,66–68 Acetate, propionate (propionic acid is also commonly referred to as PPA), butyrate, and pentanoate, having respectively, 2, 3, 4, and 5 carbon atoms are SCFAs (Table 1), largely produced by microbial fermentation of complex polysaccharides (starches and fibers) in the colon (longer chain aliphatic acids with 6 to 12 carbons are considered to be medium-chain fatty acids [MCFAs]). SCFAs are absorbed into the colonic epithelium where, primarily, butyrate is consumed as a preferred fuel source by colonocytes.69–73 Microbiota-produced SCFAs enter the bloodstream through the portal circulation of the host and/or the distal colon and are transported to recipient tissues where they are taken up and used in a variety of cellular responses, including the regulation of gene expression.9,47,74,75
Table 1.
Chemical Structures of Fatty Acids with Two to Eight Carbon Atoms
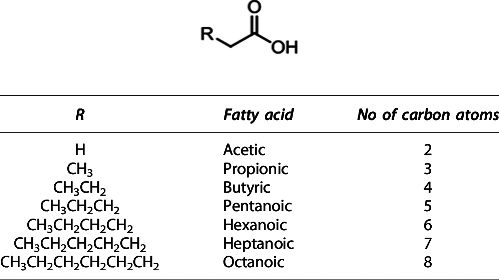 |
Many brain disorders are associated with imbalances in protein acetylation levels and transcriptional dysfunction.76 Histone deacetylase (HDAC) inhibitors represent a promising therapeutic option to correct these deficiencies, and numerous studies using butyrate, the most potent of the SCFA HDAC inhibitors,9,77 demonstrate the medicinal potential of butyrate in the intervention of neurodegenerative diseases and psychiatric disorders.26,64,76,78–85
α-Lipoic acid [(R)-5-(1,2-dithiolan-3-yl)pentanoic acid] (ALA, Fig. 1) is a naturally occurring 5-membered ring disulfide-substituted SCFA HDAC inhibitor86,87 with strong antioxidant activity.88 It is an essential cofactor in aerobic metabolism and is the central component forming the pyruvate dehydrogenase complex,89,90 which functionally links glycolysis in the cytoplasm to oxidative phosphorylation (OXPHOS) in mitochondria.91 ALA plays a role in microbial metabolism too.92 Although ALA is present in almost all food types that we eat,93 and is readily digested, absorbed, and transported to tissues, including brain,94–96 the amounts available from diet are low.93 Although the acquisition and use of ALA vary in different microbes, yeast, and animal cells,97,98 its functions are, nonetheless, essential to the organism, and in most prokaryotic and eukaryotic microorganisms, plant and animal mitochondria, and plant plastids, ALA is enzymatically synthesized endogenously from the MCFA, octanoate.97–99
FIG. 1.
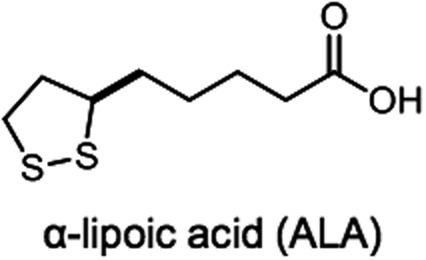
Chemical structure of R-(+)-lipoic acid.
In addition to its role in the metabolic pathways, ALA is reported in many research studies to be a potent activator of the nuclear factor (erythroid-derived 2)-like 2 (Nrf2) antioxidant response element signaling pathway that regulates the expression of genes whose protein products are involved in the detoxification and elimination of reactive oxygen species and electrophilic agents.26,96,100–102 Oxidative stress has been implicated in the pathogenesis of various neurodegenerative and neuropsychiatric disorders, including depression.103,104 Vasconcelos et al.105 showed that ALA (100 mg/kg) alone or combined with clozapine reversed schizophrenia-like alterations induced by ketamine. Ketamine is a known glutamatergic N-methyl-d-aspartate receptor antagonist that can induce psychotomimetic, perceptual, cognitive, and neuroendocrine responses in humans and in rodent models of schizophrenia.106,107 ALA also inhibits nuclear factor kappa-B (NF-κB) activation independent of its antioxidant function. NF-κB belongs to an important group of transcription factors regulated by a kinase-mediated signaling pathway that transduces signals from the cell surface to changes in gene expression.108,109
Fatty acids represent one of the body's long-term storage reservoirs and sources of fuel energy—the heart being a primary consumer.110 In the presence of respiratory oxygen, fatty acids are shuttled through the mitochondrial OXPHOS complex system, where they are degraded by two carbon units at a time to shorter-chain fatty acids (and ultimately to acetyl coenzyme A), with concomitant release of water, CO2, and ATP in the process. The cellular availability of SCFAs for use in epigenetic chromatin remodeling through their ability to inhibit HDAC activity, therefore, is closely tied to mitochondrial energy production and metabolism. Since both prokaryote and eukaryote cells share common pathways for energy production, for example, the citric acid cycle,75 it comes as no surprise that gut microbiota inexorably affect host–cell bioenergetics, which in turn fuels gene expression in the mitochondrial and nuclear genomes.75,111 Over a billion years of evolutionary history have allowed our mitochondrial DNA (mtDNA) and nuclear DNA to coevolve with a high degree of genetic compatibility.112,113 Interestingly, our microbiome and our mitogenome (mtDNA), as well as portions of our epigenome—for example, maternal silencing,114 are uniquely passed to each of us from our mother.
Gut microbiota have a profound influence on the host immune system.13 Maternal immune activation is a shared environmental risk factor for a plethora of neuropsychiatric and neurodegenerative disorders that may or may not develop into clinical symptoms in offspring.13 Evidence from an in-depth study115 of data extracted from the Danish health registry of more than 1 million children born between 1980 and 2005, focusing on cases where the mother had a viral infection with fever requiring hospitalization during the first trimester, strongly links maternal immune dysregulation with suppressed neurodevelopment and cognitive function (ASD) in their offspring.116 In other epidemiological studies,117 prenatal exposure to infection visibly stands out as a risk factor in schizophrenia and other neurodevelopmental abnormalities. The possibility that prenatal Zika virus infection from a mosquito reservoir is responsible for the current outbreak in Brazil of children born with microcephaly is a disturbing and frightening example.118–121
Synthetic Biology
With a capacity to act either naturally or by manipulation, the gut microbial ecosystem is an indispensable and constituent player in the maintenance of our well-being.7 Thus, in treating disease, adjusting the functional composition of the gut microbiome may help facilitate and even alter the outcome of therapeutic interventions.122,123 Dietary sources of probiotics such as, for example, in traditional Greek yogurt, have been used since antiquity in the Mediterranean region (Mediterranean diet)124,125 to maintain a state of wellness. However, once pathogenic dysbiosis sets in, probiotics have not proven to be remedial123,126,127 and other interventional methods are being investigated. In this effort, independent work from several groups suggests that (engineered) bacteria have potential to be an effective means for delivering, enhancing, or themselves acting as therapeutic agents (“living pills”)128 to treat certain diseases,22,129–132 including psychiatric disorders,43,48,133–136 and significant investments are being made to adapt a variety of commensal microbial species for remodeling the gut microbiota (ecobiotics) in disease-treating indications.133,137–139 A similar approach aims to utilize engineered viruses to seek and selectively destroy pathogenic bacteria.140–142 Drawing conclusions from a study of 11 children affected by ASD that showed improvement in communication and behavioral tests after being treated with vancomycin for 8 weeks, Mangiola et al.46 speculate that modulation of gut microbiota through antibiotic treatment may influence the symptoms and expression of psychiatric disorders in general. Devkota143 takes this further by underscoring a comprehensive need for more investigations into drug–microbiome interactions and the mechanisms that are involved therein.
Interestingly, during long stays in space, the configuration of the gut microbiome of astronauts is often significantly transformed relative to the one they had on Earth.144–146 The ongoing NASA Twins Study, with Scott Kelly having recently returned from a historic 340-day mission aboard the International Space Station, may shine additional light on this subject.147 Gut microbial dysregulation can alter one's immune status and cause aberrant social and cognitive behavior.35,148,149 This may result in catastrophic consequences during long space flights, as for example, to Mars, if an astronaut's ability to carry out demanding tasks at a high performance and optimal level becomes severely compromised.150 History shows that addressing the technological challenges space exploration presents has a constructive rippling effect on the technological advances made for a wide range of applications here on Earth. Synthetic biology151 has potential to deliver robust and reliable organisms that can assist on long-duration astronaut missions.152 It is anticipated that the techniques required to be developed may also be applied to engineer phage and bacteria to explore and to therapeutically modify the gut microbiome as needed.153
Brain Development and Neurological Disorders
Aging is a leading risk factor (Fig. 2)154 in progressing to dementia.155–159 Although the latest studies suggest that the prevalence of dementia may be leveling off and even decreasing in some subsets of the population,160 for the foreseeable future, dementia will continue to be a major challenge for the healthcare establishment.161
FIG. 2.

Meta analyzed estimates of dementia prevalence in the United States.
The gut's evolving capacity to adapt and maintain normal microbiota, which begins at birth and continues throughout one's life, is necessary to support the metabolic activities of the brain.59,162 This is especially so in the early childhood years through adulthood.59 Studies have shown that some of the typical behavioral and physiological abnormalities associated with neurodevelopmental disorders, including autism43–45,163–165 and schizophrenia,40,47–49 can be modulated by reconfiguring the gut microbiome composition.122,166
Acetate, propionate, and butyrate comprise the majority of SCFAs produced in the gut by microbial fermentation.21,69 Propionate and butyrate can modulate brain functioning, principally appetite and energy homeostasis, through regulation of neuropeptide production.167 Butyrate is mostly absorbed by the colonic epithelium, whereas acetate and propionate are passed into the portal circulation.168–170 In gut dysbiosis, the constitutional spectrum of SCFAs varies substantially from the host's natural healthy balance,3,32,39,60,171 and higher than normal levels of PPA have been linked to deleterious effects on brain function43,75,78,172–179 in autistic children.24,174 Given these findings, and the fact that PPA is widely used as a food preservative,180 there may be cause for some concern.
Idiopathic late-onset dementia (ILOD) is characterized by a series of declining daily functional competences, most often involving memory, reasoning, and sociobehavioral abilities, in the elderly.157,161 Dementia encompasses a myriad of clinical symptoms typically associated with discrete neurological disorders such as Alzheimer and Parkinson diseases, hippocampal sclerosis of aging, and Lewy body and frontotemporal dementias being the more notable ones, but not collectively manifested by any one of them. Cellular processes depend on the energy supplied by their mitochondria, and dysfunctional mitochondria can lead to an unsustainable cellular bioenergetics deficit that is detrimental to the cell's function and survival.181 In brain cells, even a small energy deficit, which is a common occurrence during the aging process, can reduce synaptic neurotransmitter release and adversely affect synaptic function.26,101 Maintaining a healthy gut microbiota state is necessary to support the metabolic activities of the brain,59,162 and Mattson157 and Bourassa et al.85 posit that some of the common pathologies leading to ILOD and other brain disorders may be amenable to therapeutic modification by diet and lifestyle changes. For example, exercise, yoga, and meditation are lifestyle activities known to improve brain blood flow—which, presumably, can enhance perfusion of the brain with micronutrients absorbed by the gut182—and are increasingly being incorporated in treatments for depression and other mental disturbances.36,127,183–185
At the other end of the age spectrum, the correlation between impaired intellectual development and a prolonged state of malnutrition in infants and young children is inescapable.186–189 Recent studies have demonstrated that the normal pattern of gut microbiota assembly is disrupted in malnourished children.162,187 To maximize the therapeutic benefit of diet and dietary supplements, preclinical evidence suggests that a healthy microbiome in these children may need to be configured as well.190,191
Concluding Remarks
The importance of physical activity coupled with a balanced and healthy diet in the maintenance of our well-being has been recognized since antiquity. However, it is only recently that characterization of the host–microbiome intermetabolic and crosstalk pathways has come to the forefront for study in therapeutic design and treatments.68,192 As reviewed in this report, synthetic biology has potential to develop microorganisms for correcting pathogenic dysbiosis, but this has yet to be proven. (For additional examples of the latest approaches to manipulating the microbiota, including illustrative figures, see Ash and Mueller,193 and articles cited therein.) In contrast, the development and use of small molecule drugs have a long and successful history in the clinical treatment of diseases. Small molecule HDAC inhibitors are already used in the clinic to treat cancer and hematological disorders,77 and preclinical research with SCFA HDAC inhibitors demonstrates significant potential in epigenetic treatment of neurological conditions.26,101 Epigenetic regulation of host–microbiota interactions by utilizing epigenomic-targeting drugs has been suggested by Alenghat and Artis.74
The mouth, being an extension of the digestive tract, presents a readily accessible diagnostic site for the early detection of potential unhealthy pathogens resident in the gut. Salivanomics is a rapidly emerging tool in the arsenal of salivary diagnostics.194 Collecting saliva or swabbing the inside cheek of the mouth is noninterventional, making it a highly attractive diagnostic procedure, particularly for infants and young children. It is not unlikely that regular visits to your dentist may soon be as important to your gut as it is to your oral hygiene, white teeth, and a nice smile.
Abbreviations Used
- ALA
α-lipoic acid
- ASD
autism spectrum disorder
- HDAC
histone deacetylase
- ILOD
idiopathic late-onset dementia
- MCFA
medium-chain fatty acid
- mtDNA
mitochondrial DNA
- NF-κB
nuclear factor kappa-B
- Nrf2
nuclear factor (erythroid-derived 2)-like 2
- OXPHOS
oxidative phosphorylation
- PPA
propionic acid
- SCFA
short-chain fatty acid
Acknowledgments
We thank the MitoCure Foundation for their generous financial support, and Drs. Carl A. Pinkert (The University of Alabama, Tuscaloosa, AL), Michael H. Irwin (Auburn University, Auburn, AL), Robert J. Zamboni (McGill University, Montreal, QC, Canada), and Susan P. Perrine (Boston University School of Medicine) for their invaluable advice and helpful discussions in the preparation of this article.
Authors’ Contributions
The article was written through contributions of all authors. All authors have given approval to the final version of the article.
Author Disclosure Statement
K.S. owns shares in PhenoMatriX. W.H.M. and K.S. have collaborated in the past on cancer projects as part of a formal agreement between SRI International and PhenoMatriX. No competing financial interests exist.
References
- 1.Dietert J, Dietert R. The sum of our parts. Scientist. 2015;29:44–49 [Google Scholar]
- 2.Ghannoum M. The mycobiome. Scientist. 2016;30:32–37 [Google Scholar]
- 3.Fond G, Boukouaci W, Chevalier G, et al. The “psychomicrobiotic”: targeting microbiota in major psychiatric disorders: a systematic review. Pathol Biol. 2015;63:35–42 [DOI] [PubMed] [Google Scholar]
- 4.Sender R, Fuchs S, Milo R. Revised estimates for the number of human and bacteria cells in the body. BioRxiv. 2016. [Epub ahead of print]; DOI: http://dx.doi.org/10.1101/036103 [DOI] [PMC free article] [PubMed]
- 5.Blekhman R, Goodrich JK, Huang K, et al. Host genetic variation impacts microbiome composition across human body sites. Genome Biol. 2015;16:191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bohórquez DV, Liddle RA. The gut connectome: making sense of what you eat. J Clin Invest. 2015;125:888–890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Koppel N, Balskus EP. Exploring and understanding the biochemical diversity of the human microbiota. Cell Chem Biol. 2016;23:18–30 [DOI] [PubMed] [Google Scholar]
- 8.Forsythe P, Sudo N, Dinan T, et al. Mood and gut feelings. Brain Behav Immun. 2010;24:9–16 [DOI] [PubMed] [Google Scholar]
- 9.Natarajan N, Pluznick JL. From microbe to man: the role of microbial short chain fatty acid metabolites in host cell biology. Am J Physiol Cell Physiol. 2014;307:C979–C985 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dietert R, Dietert J. The microbiome and sustainable healthcare. Healthcare. 2015;3:100–129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Caesar R, Tremaroli V, Kovatcheva-Datchary P, et al. Crosstalk between gut microbiota and dietary lipids aggravates WAT inflammation through TLR signaling. Cell Metabol. 2015;22:1–11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Belkaid Y, Hand TW. Role of the microbiota in immunity and inflammation. Cell. 2014;157:121–141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Estes ML, McAllister AK. Immune mediators in the brain and peripheral tissues in autism spectrum disorder. Nat Rev Neurosci. 2015;16:469–486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Grogan D. The microbes within. Nature. 2015;518:S2. [DOI] [PubMed] [Google Scholar]
- 15.Bäckhed F, Roswall J, Peng Y, et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe. 2015;17:690–703 [DOI] [PubMed] [Google Scholar]
- 16.Debnath M, Venkatasubramanian G, Berk M. Fetal programming of schizophrenia: select mechanisms. Neurosci Biobehav Rev. 2015;49:90–104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dominguez-Bello MG, Blaser MJ. Asthma: undoing millions of years of coevolution in early life? Sci Transl Med. 2015;7:307fs39. [DOI] [PubMed] [Google Scholar]
- 18.van Opstal EJ, Bordenstein SR. Rethinking heritability of the microbiome. Science. 2015;349:1172–1173 [DOI] [PubMed] [Google Scholar]
- 19.Dominguez-Bello MG, De Jesus-Laboy KM, Shen N, et al. Partial restoration of the microbiota of cesarean-born infants via vaginal microbial transfer. Nat Med. 2016;22:250–253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gomez de Agüero M, Ganal-Vonarburg SC, Fuhrer T, et al. The maternal microbiota drives early postnatal innate immune development. Science. 2016;351:1296–1302 [DOI] [PubMed] [Google Scholar]
- 21.Al-Asmakh M, Hedin L. Microbiota and the control of blood-tissue barriers. Tissue Barriers. 2015;3:e10396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.McKenney PT, Pamer EG. 2015. From hype to hope: the gut microbiota in enteric infectious disease. Cell 163:1326–1332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Scharschmidt TC, Vasquez KS, Truong H-A, et al. A wave of regulatory T cells into neonatal skin mediates tolerance to commensal microbes. Immunity. 2015;43:1011–1021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang Z, Roberts AB, Buffa JA, et al. Non-lethal inhibition of gut microbial trimethylamine production for the treatment of atherosclerosis. Cell. 2015;163:1585–1595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yang I, Corwin EJ, Brennan PA, et al. The infant microbiome. Implications for infant health and neurocognitive development. Nursing Res. 2016;65:76–88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Moos WH, Maneta E, Pinkert CA, et al. Epigenetic treatment of neuropsychiatric disorders: autism and schizophrenia. Drug Dev Res. 2016;77:53–72 [DOI] [PubMed] [Google Scholar]
- 27.Shakir R. Neurodegenerative noncommunicable diseases (neurology NCDs). Where are we now? J Neurol Sci. 2015;356:1–2 [DOI] [PubMed] [Google Scholar]
- 28.Silberberg D, Anand NP, Michels K, et al. Brain and other nervous system disorders across the lifespan—global challenges and opportunities. Nature. 2015;527:S151–S154 [DOI] [PubMed] [Google Scholar]
- 29.Borre YE, O'Keeffe GW, Clarke G, et al. Microbiota and neurodevelopmental windows: implications for brain disorders. Trends Mol Med. 2014;20:509–518 [DOI] [PubMed] [Google Scholar]
- 30.de Theije CGM, Bavelaar BM, da Silva SL, et al. Food allergy and food-based therapies in neurodevelopmental disorders. Pediatr Allergy Immunol. 2014;25:218–226 [DOI] [PubMed] [Google Scholar]
- 31.Mayer EA, Knight R, Mazmanian SK, et al. Gut microbes and the brain: paradigm shift in neuroscience. J Neurosci. 2014;34:15490–15496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Stilling RM, Dinan TG, Cryan JF. Microbial genes, brain & behaviour—epigenetic regulation of the gut–brain axis. Genes Brain Behav. 2014;13:69–86 [DOI] [PubMed] [Google Scholar]
- 33.Bienenstock J, Kunze W, Forsythe P. Microbiota and the gut–brain axis. Nutr Rev. 2015;73(S1):28–31 [DOI] [PubMed] [Google Scholar]
- 34.Dash SR. The microbiome and brain health: what's the connection? Medscape, March 24, 2015. Accessed March21, 2016, http://www.medscape.com/viewarticle/841748
- 35.Dinan TG, Stilling RM, Stanton C, et al. Collective unconscious: how gut microbes shape human behavior. J Psychiatr Res. 2015;63:1–9 [DOI] [PubMed] [Google Scholar]
- 36.Kaplan BJ, Rucklidge JJ, Romijn A, et al. The emerging field of nutritional mental health: inflammation, the microbiome, oxidative stress, and mitochondrial function. Clin Psychol Sci. 2015;3:964–980 [Google Scholar]
- 37.Sherman MP, Zaghouani H, Niklas V. Gut microbiota, the immune system, and diet influence the neonatal gut–brain axis. Pediatr Res. 2015;77:127–135 [DOI] [PubMed] [Google Scholar]
- 38.Donaldson GP, Lee SM, Mazmanian SK. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol. 2016;14:20–32 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hsiao EY, McBride SW, Hsien S, et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell. 2013;155:1451–1463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kraneveld AD, de Theije CGM, van Heesch F, et al. The neuro-immune axis: prospect for novel treatments for mental disorders. Basic Clin Pharmacol Toxicol. 2014;114:128–136 [DOI] [PubMed] [Google Scholar]
- 41.Buie T. Potential etiologic factors of microbiome disruption in autism. Clin Ther. 2015;37:976–983 [DOI] [PubMed] [Google Scholar]
- 42.Carding S, Verbeke K, Vipond DT, et al. Dysbiosis of the gut microbiota in disease. Microb Ecol Health Dis. 2015;26:26191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Frye RE, Rose S, Slattery J, et al. Gastrointestinal dysfunction in autism spectrum disorder: the role of the mitochondria and the enteric microbiome. Microb Ecol Health Dis. 2015;26:27458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Liu X, Cao S, Zhang X. Modulation of gut microbiota–brain axis by probiotics, prebiotics, and diet. J Agric Food Chem. 2015;63:7885–7895 [DOI] [PubMed] [Google Scholar]
- 45.Reddy BL, Saier MH. Autism and our intestinal microbiota. J Mol Microbiol Biotechnol. 2015;25:51–55 [DOI] [PubMed] [Google Scholar]
- 46.Mangiola F, Ianiro G, Franceschi F, et al. Gut microbiota in autism and mood disorders. World J Gastroenterol. 2016;22: 361–368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Dinan TG, Borre YE, Cryan JF. Genomics of schizophrenia: time to consider the gut microbiome? Mol Psychiatry. 2014;19:1252–1257 [DOI] [PubMed] [Google Scholar]
- 48.Nemani K, Ghomi RH, McCormick B, et al. Schizophrenia and the gut–brain axis. Prog Neuropsychopharmacol Biol Psychiatry. 2015;56:155–160 [DOI] [PubMed] [Google Scholar]
- 49.Severance EG, Prandovszky E, Castiglione J, et al. Gastroenterology issues in schizophrenia: why the gut matters. Curr Psychiatry Rep. 2015;17:27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Chadman KK. Making progress in autism drug discovery. Expert Opin Drug Discov. 2014;9:1389–1391 [DOI] [PubMed] [Google Scholar]
- 51.Sekar A, Bialas AR, de Rivera H, et al. Schizophrenia risk from complex variation of complement component 4. Nature. 2016;530:177–183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Muir WJ, McKechanie AG. Can epigenetics help in the discovery of therapeutics for psychiatric disorders, especially schizophrenia? Expert Opin Drug Discov. 2009;4:621–627 [DOI] [PubMed] [Google Scholar]
- 53.Bosia M, Pigoni A, Cavallaro R. Genomics and epigenomics in novel schizophrenia drug discovery: translating animal models to clinical research and back. Expert Opin Drug Discov. 2015;10:125–139 [DOI] [PubMed] [Google Scholar]
- 54.Hinde K, Lewis ZT. Mother's littlest helpers. Breastmilk nourishes the microbes colonizing the neonatal intestinal tract. Science. 2015;348:1427–1428 [DOI] [PubMed] [Google Scholar]
- 55.Meadow JF, Altrichter AE, Bateman AC, et al. Humans differ in their personal microbial cloud. PeerJ. 2015;3:e1258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Zeevi D, Korem T, Zmora N, et al. Personalized nutrition by prediction of glycemic responses. Cell. 2015;163:1079–1094 [DOI] [PubMed] [Google Scholar]
- 57.Zuker CS. Food for the brain. Cell. 2015;161:9–11 [DOI] [PubMed] [Google Scholar]
- 58.Kabouridis PS, Pachnis V. Emerging roles of gut microbiota and the immune system in the development of the enteric nervous system. J Clin Invest. 2015;125:956–964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Goyal MS, Venkatesh S, Milbrandt J, et al. Feeding the brain and nurturing the mind: linking nutrition and the gut microbiota to brain development. Proc Natl Acad Sci U S A. 2015;112:14105–14112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Janssen AWF, Kersten S. The role of the gut microbiota in metabolic health. FASEB J. 2015;29:3111–3123 [DOI] [PubMed] [Google Scholar]
- 61.Ledford H. The genome unwrapped. Nature. 2015;528:S12–S13 [DOI] [PubMed] [Google Scholar]
- 62.Takahashi K. Influence of bacteria on epigenetic gene control. Cell Mol Life Sci. 2014;71:1045–1054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Stefano GB, Kream RM. Dysregulated mitochondrial and chloroplast bioenergetics from a translational medical perspective (Review). Int J Mol Med. 2016;37:547–555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.De Vadder F, Kovatcheva-Datchary P, Goncalves D, et al. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell. 2014;156:84–96 [DOI] [PubMed] [Google Scholar]
- 65.Jandhyala SM, Talukdar R, Subramanyam C, et al. Role of the normal gut microbiota. World J Gastroenterol. 2015;21:8787–8803 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Psichas A, Reimann F, Gribble FM. Gut chemosensing mechanisms. J Clin Invest. 2015;125:908–917 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Scheperjans F, Aho V, Pereira PAB, et al. Gut microbiota are related to Parkinson's disease and clinical phenotype. Mov Disord. 2015;30:350–358 [DOI] [PubMed] [Google Scholar]
- 68.Saint-Georges-Chaumet Y, Edeas M. Microbiota–mitochondria inter-talk: consequence for microbiota–host interaction. FEMS Pathog Dis. 2016;74:ftv096. [DOI] [PubMed] [Google Scholar]
- 69.Donohoe DR, Garge N, Zhang X, et al. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 2011;13:517–526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Donohoe DR, Collins LB, Wali A, et al. The Warburg effect dictates the mechanism of butyrate-mediated histone acetylation and cell proliferation. Mol Cell. 2012;48:612–626 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Krishnan S, Alden N, Lee K. Pathways and functions of gut microbiota metabolism impacting host physiology. Curr Opin Biotechnol. 2015;36:137–145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Velasquez-Manoff M. Gut microbiome: the peacekeepers. Nature. 2015;518:S3-S11 [DOI] [PubMed] [Google Scholar]
- 73.Raman M, Ambalam P, Doble M. Short-chain fatty acids. In: Probiotics and Bioactive Carbohydrates in Colon Cancer Management. Springer: New Delhi, India; pp. 97–115; 2016 [Google Scholar]
- 74.Alenghat T, Artis D. Epigenomic regulation of host–microbiota interactions. Trends Immunol. 2014;35:518–525 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.MacFabe DF. Enteric short-chain fatty acids: microbial messengers of metabolism, mitochondria, and mind: implications in autism spectrum disorders. Microb Ecol Health Dis. 2015;26:28177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Chuang D-M, Leng Y, Marinova Z, et al. Multiple roles of HDAC inhibition in neurodegenerative conditions. Trends Neurosci. 2009;32:591–601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Steliou K, Boosalis MS, Perrine SP, et al. Butyrate histone deacetylase inhibitors. BioRes Open Access. 2012;1:192–198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Thomas EA. Focal nature of neurological disorders necessitates isotype-selective histone deacetylase (HDAC) inhibitors. Mol Neurobiol. 2009;40:33–45 [DOI] [PubMed] [Google Scholar]
- 79.Grayson DR, Kundakovic M, Sharma RP. Is there a future for histone deacetylase inhibitors in the pharmacotherapy of psychiatric disorders? Mol Pharmacol. 2010;77:126–135 [DOI] [PubMed] [Google Scholar]
- 80.Canani RB, Di Costanzo M, Leone L. The epigenetic effects of butyrate: potential therapeutic implications for clinical practice. Clin Epigenet. 2012;4:4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Rosenstock TR. Lysine (K)-deacetylase inhibitors: the real next step to neuropsychiatric and neurodegenerative disorders? Cell Biol. 2013;2:a8 [Google Scholar]
- 82.Szyf M. Prospects for the development of epigenetic drugs for CNS conditions. Nat Rev Drug Discov. 2015;14:461–474 [DOI] [PubMed] [Google Scholar]
- 83.Vizcarra JA, Wilson-Perez HE, Espay AJ. The power in numbers: gut microbiota in Parkinson's disease. Mov Disord. 2015;30:296–298 [DOI] [PubMed] [Google Scholar]
- 84.Volmar C-H, Wahlestedt C. Histone deacetylases (HDACs) and brain function. Neuroepigenetics. 2015;1:20–27 [Google Scholar]
- 85.Bourassa MW, Alim I, Bultman SJ, et al. Butyrate, neuroepigenetics and the gut microbiome: can a high fiber diet improve brain health? Neurosci Lett. 2015. [Epub ahead of print]; DOI: 10.1016/j.neulet.2016.02.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.van de Mark K, Chen JS, Steliou K, et al. α-Lipoic acid induces p27Kip-dependent cell cycle arrest in non-transformed cell lines and apoptosis in tumor cell lines. J Cell Physiol. 2003;194:325–340 [DOI] [PubMed] [Google Scholar]
- 87.Bassett SA, Barnett MPG. The role of dietary histone deacetylases (HDACs) inhibitors in health and disease. Nutrients. 2014;6:4273–4301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Rochette L, Ghibu S, Richard C, et al. Direct and indirect antioxidant properties of alpha-lipoic acid and therapeutic potential. Mol Nutr Food Res. 2013;57:114–125 [DOI] [PubMed] [Google Scholar]
- 89.de Boer VCJ, Houten SM. A mitochondrial expatriate: nuclear pyruvate dehydrogenase. Cell. 2014;158:9–10 [DOI] [PubMed] [Google Scholar]
- 90.Sutendra G, Kinnaird A, Dromparis P, et al. A nuclear pyruvate dehydrogenase complex is important for the generation of acetyl-CoA and histone acetylation. Cell. 2014;158:84–97 [DOI] [PubMed] [Google Scholar]
- 91.Mayr JA, Feichtinger RG, Tort F, et al. Lipoic acid biosynthesis defects. J Inherit Metab Dis. 2014. 37:553–563 [DOI] [PubMed] [Google Scholar]
- 92.Ullah MW, Khattak WA, Ul-Islam M, et al. Metabolic engineering of synthetic cell-free systems: strategies and applications. Biochem Eng J. 2016;105:391–405 [Google Scholar]
- 93.Li S. Synthetic bioactive substances. In:Handbook of Food Chemistry. Cheung PCK, Mehta BM, (eds.) Springer-Verlag: Berlin, Germany; pp. 1–20; 2015. (ISBN: 978–3-642–41609-5) [Google Scholar]
- 94.Carlson DA, Smith AR, Fischer SJ, et al. The plasma pharmacokinetics of R-(+)-lipoic acid administered as sodium R-(+)-lipoate to healthy human subjects. Altern Med Rev. 2007;12:343–351 [PubMed] [Google Scholar]
- 95.Papanas N, Ziegler D. Efficacy of α-lipoic acid in diabetic retinopathy. Expert Opin Pharmacother. 2015;15:2721–2731 [DOI] [PubMed] [Google Scholar]
- 96.Rochette L, Ghibu S, Muresan A, et al. Alpha-lipoic acid: molecular mechanisms and therapeutic potential in diabetes. Can J Physiol Pharmacol. 2015;93:1021–1027 [DOI] [PubMed] [Google Scholar]
- 97.Spalding MD, Sean T, Prigge ST. Lipoic acid metabolism in microbial pathogens. Microbiol Mol Biol Rev. 2010;74:200–228 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Zhang H, Luo Q, Gao H, et al. A new regulatory mechanism for bacterial lipoic acid synthesis. Microbiologyopen. 2015;4:282–300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Pashaj A, Xia M, Moreau R. α-Lipoic acid as a triglyceride-lowering nutraceutical. Can J Physiol Pharmacol. 2015;93:1029–1041 [DOI] [PubMed] [Google Scholar]
- 100.Dörsam B, Fahrer J. The disulfide compound α-lipoic acid and its derivatives: a novel class of anticancer agents targeting mitochondria. Cancer Lett. 2016;371:12–19 [DOI] [PubMed] [Google Scholar]
- 101.Irwin MH, Moos WH, Faller DV, et al. Epigenetic treatment of neurodegenerative disorders: Alzheimer and Parkinson diseases. Drug Dev Res. 2016;77:109–123 [DOI] [PubMed] [Google Scholar]
- 102.Prasad KN. Simultaneous activation of Nrf2 and elevation of dietary and endogenous antioxidant chemicals for cancer prevention in humans. J Am Coll Nutr. 2016;35:175–184 [DOI] [PubMed] [Google Scholar]
- 103.Lombard JH. Depression, psychological stress, vascular dysfunction, and cardiovascular disease: thinking outside the barrel. J Appl Physiol. 2010;108:1025–1026 [DOI] [PubMed] [Google Scholar]
- 104.Silva MCC, de Sousa CNS, Gomes PXL, et al. Evidence for protective effect of lipoic acid and desvenlafaxine on oxidative stress in a model depression in mice. Prog Neuropsychopharmacol Biol Psychiatry. 2016;64:142–148 [DOI] [PubMed] [Google Scholar]
- 105.Vasconcelos GS, Ximenes NC, de Sousa CNS, et al. Alpha-lipoic acid alone and combined with clozapine reverses schizophrenia-like symptoms induced by ketamine in mice: participation of antioxidant, nitrergic and neurotrophic mechanisms. Schizophr Res. 2015;165:163–170 [DOI] [PubMed] [Google Scholar]
- 106.Krystal JH, Karper LP, Seibyl JP, et al. Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Arch Gen Psychiatry. 1994;51:199–214 [DOI] [PubMed] [Google Scholar]
- 107.Lafioniatis A, Orfanidou MA, Papadopoulou ES, et al. Effects of the inducible nitric oxide synthase inhibitor aminoguanidine in two different rat models of schizophrenia. Behav Brain Res. 2016;309:14–21 [DOI] [PubMed] [Google Scholar]
- 108.Ying Z, Kampfrath T, Sun Q, et al. Evidence that alpha-lipoic acid inhibits NF-kappaB activation independent of its antioxidant function. Inflamm Res. 2011;60:219–225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Buendia I, Michalska P, Navarro E, et al. Nrf2–ARE pathway: an emerging target against oxidative stress and neuroinflammation in neurodegenerative diseases. Pharmacol Ther. 2016;157:84–104 [DOI] [PubMed] [Google Scholar]
- 110.Beauloye C, Horman S, Bertrand L. Even is better than odd: one fat may conceal another. Am J Physiol Heart Circ Physiol. 2015;309:H1112–H1114 [DOI] [PubMed] [Google Scholar]
- 111.Shenderov BA, Midtvedt T. Epigenomic programing: a future way to health? Microb Ecol Health Dis. 2014;25:24145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Hill GE. Mitonuclear ecology. Mol Biol Evol. 2015;32:1917–1927 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Johnston IG, Williams BP. Evolutionary inference across eukaryotes identifies specific pressures favoring mitochondrial gene retention. Cell Systems. 2016;2:101–111 [DOI] [PubMed] [Google Scholar]
- 114.Wilkins JF, Úbeda F, Van Cleve J. The evolving landscape of imprinted genes in humans and mice: conflict among alleles, genes, tissues, and kin. Bioessays. 2016;38:482–489 [DOI] [PubMed] [Google Scholar]
- 115.Atladóttir HÓ, Thorsen P, Østergaard L, et al. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J Autism Dev Disord. 2010;40:1423–1430 [DOI] [PubMed] [Google Scholar]
- 116.Choi GB, Yim YS, Wong H, et al. The maternal interleukin-17a pathway in mice promotes autism-like phenotypes in offspring. Science. 2016;351:933–939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Marangoni C, Hernandez M, Faedda GL. The role of environmental exposures as risk factors for bipolar disorder: a systematic review of longitudinal studies. J Affect Disord. 2016;193:165–174 [DOI] [PubMed] [Google Scholar]
- 118.Brasil P, Pereira JP, Jr., Gabaglia CR, et al. Zika virus infection in pregnant women in Rio de Janeiro—preliminary report. N Engl J Med. 2016. [Epub ahead of print]; DOI: 10.1056/NEJMoa1602412 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Broutet N, Krauer F, Riesen M, et al. Zika virus as a cause of neurologic disorders. N Engl J Med. 2016;374:1506–1509 [DOI] [PubMed] [Google Scholar]
- 120.Estes ML, McAllister AK. Maternal TH17 cells take a toll on baby's brain. Science. 2016;351:919–920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Victora CG, Schuler-Faccini L, Matijasevich A, et al. Microcephaly in Brazil: how to interpret reported numbers? Lancet. 2016;387:621–623 [DOI] [PubMed] [Google Scholar]
- 122.Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behavior. Nat Rev Neurosci. 2012;13:701–712 [DOI] [PubMed] [Google Scholar]
- 123.Zmora N, Zeevi D, Korem T, et al. Taking it personally: personalized utilization of the human microbiome in health and disease. Cell Host Microbe. 2016;19:12–20 [DOI] [PubMed] [Google Scholar]
- 124.Katsiardanis K, Diamantaras A-A, Dessypris N, et al. Cognitive Impairment and dietary habits among elders: the Velestino study. J Med Food. 2013;16:343–350 [DOI] [PubMed] [Google Scholar]
- 125.Babio N, Becerra-Tomás N, Martínez-González MÁ, et al. Consumption of yogurt, low-fat milk, and other low-fat dairy products is associated with lower risk of metabolic syndrome incidence in an elderly Mediterranean population. J Nutr. 2015;145:2308–2316 [DOI] [PubMed] [Google Scholar]
- 126.Petra AI, Panagiotidou S, Hatziagelaki E, et al. Gut-microbiota-brain axis and its effect on neuropsychiatric disorders with suspected immune dysregulation. Clin Ther. 2015;37:984–995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Klein N, Kemper KJ. Integrative approaches to caring for children with autism. Curr Probl Pediatr Adolesc Health Care. 2016. [Epub ahead of print]; DOI: 10.1016/j.cppeds.2015.12.004 [DOI] [PubMed] [Google Scholar]
- 128.Eisenstein M. Living factories of the future. Nature. 2016;531:401–403 [DOI] [PubMed] [Google Scholar]
- 129.Bel S, Hooper LV. A bacterial nudge to T-cell function. Nature. 2015;526:328–330 [DOI] [PubMed] [Google Scholar]
- 130.Leslie M. Microbes aid cancer drugs. Science. 2015;350:614–615 [DOI] [PubMed] [Google Scholar]
- 131.Piñero-Lambea C, Ruano-Gallego D, Fernández LA. Engineered bacteria as therapeutic agents. Curr Opin Biotechnol. 2015;35:94–102 [DOI] [PubMed] [Google Scholar]
- 132.Vétizou M, Pitt JM, Daillère R, et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015;350:1079–1084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Garber K. Drugging the gut microbiome. Nat Biotechnol. 2015;33:228–231 [DOI] [PubMed] [Google Scholar]
- 134.Kali A. Human microbiome engineering: the future and beyond. J Clin Diagn Res. 2015;9:DEO1–DEO4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Zhou L, Foster JA. Psychobiotics and the gut–brain axis: in the pursuit of happiness. Neuropsychiatr Dis Treat. 2015;11:715–723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Escoll P, Mondino S, Rolando M, et al. Targeting of host organelles by pathogenic bacteria: a sophisticated subversion strategy. Nat Rev Microbiol. 2016;14:5–19 [DOI] [PubMed] [Google Scholar]
- 137.Hayden EC. Tech investors bet on synthetic biology. Nature. 2015;527:19. [DOI] [PubMed] [Google Scholar]
- 138.Jarvis LM. Harnessing the hordes in the microbiome. Chem Eng News. 2015;93(38):12–19 [Google Scholar]
- 139.Khanna S, Pardi DS, Kelly CR, et al. A novel microbiome therapeutic increases gut microbial diversity and prevents recurrent Clostridium difficile Infection. J Infect Dis. 2016. [Epub ahead of print]; DOI: 10.1093/infdis/jiv766 [DOI] [PubMed] [Google Scholar]
- 140.Byrd AL, Segre JA. Adapting Koch's postulates. Criteria for disease causation must take microbial interactions into account. Science. 2016;351:224–226 [DOI] [PubMed] [Google Scholar]
- 141.Madhusoodanan J. Viral soldiers. Scientist. 2016;30:27–33 [Google Scholar]
- 142.Pfeiffer JK, Virgin HW. Transkingdom control of viral infection and immunity in the mammalian intestine. Science. 2016;351:aad5872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Devkota S. Prescription drugs obscure microbiome analyses. Patient populations should be stratified for medications when looking for gut microbial signatures of disease. Science. 2016;351:452–453 [DOI] [PubMed] [Google Scholar]
- 144.Wilson JW, Ott CM, Quick L, et al. Media ion composition controls regulatory and virulence response of Salmonella in spaceflight. PLoS One. 2008;3:e3923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Li P, Shi J, Zhang P, et al. Simulated microgravity disrupts intestinal homeostasis and increases colitis susceptibility. FASEB J. 2015;29:3263–3273 [DOI] [PubMed] [Google Scholar]
- 146.Ritchie LE, Taddeo SS, Weeks BR, et al. Space environmental factor impacts upon murine colon microbiota and mucosal homeostasis. PLoS One. 2015;10:e0125792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Schierholz S, Huot D. NASA astronaut Scott Kelly returns safely to Earth after one-year mission. March 2, Release 16-023. Accessed March20, 2016, http://www.nasa.gov/press-release/nasa-astronaut-scott-kelly-returns-safely-to-earth-after-one-year-mission
- 148.Snyder C, Kream RM, Ptacek R, et al. Mitochondria, microbiome and their potential psychiatric modulation. Autism Open Access. 2015;5:2 [Google Scholar]
- 149.Parashar A, Udayabanu M. Gut microbiota regulates key modulators of social behavior. Eur Neuropsychopharmacol. 2016;26:78–91 [DOI] [PubMed] [Google Scholar]
- 150.Taylor RW. Impact of space flight on bacterial virulence and antibiotic susceptibility. Infect Drug Resist. 2015;8:249–262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Hutchison CA, III, Chuang R-Y, Noskov VN, et al. Science. 2016;351:aad6253. [DOI] [PubMed] [Google Scholar]
- 152.Menezes AA, Montague MG, Cumbers J, et al. Grand challenges in space synthetic biology. JRS Interface. 2015;12: 20150803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Ciechonska M, Grob A, Isalan M. From noise to synthetic nucleoli: can synthetic biology achieve new insights? Integr Biol. 2016;8:383–393 [DOI] [PubMed] [Google Scholar]
- 154.Prince M, Bryce R, Albanese E, et al. The global prevalence of dementia: a systematic review and metaanalysis. Alzheimer's Dement. 2013;9:63–75 [DOI] [PubMed] [Google Scholar]
- 155.Lin M, Beal MF. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature. 2006;443:787–795 [DOI] [PubMed] [Google Scholar]
- 156.Currais A. Ageing and inflammation–A central role for mitochondria in brain health and disease. Ageing Res Rev. 2015;21:30–42 [DOI] [PubMed] [Google Scholar]
- 157.Mattson MP. Late-onset dementia: a mosaic of prototypical pathologies modifiable by diet and lifestyle. Nat Aging Mech Dis. 2015;1:15003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Quaglio G, Brand H, Dario C. Fighting dementia in Europe: the time to act is now. Lancet Neurol. 2016;15:452–454 [DOI] [PubMed] [Google Scholar]
- 159.Satizabal CL, Beiser AS, Chouraki V, et al. Incidence of dementia over three decades in the Framingham Heart Study. N Engl J Med. 2016;374:523–532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Wu Y-T, Fratiglioni L, Matthews FE, et al. Dementia in western Europe: epidemiological evidence and implications for policy making. Lancet Neurol. 2016;15:116–124 [DOI] [PubMed] [Google Scholar]
- 161.Winblad B, Amouyel P, Andrieu S, et al. Defeating Alzheimer's disease and other dementias: a priority for European science and society. Lancet Neurol. 2016;15:455–532 [DOI] [PubMed] [Google Scholar]
- 162.Subramanian S, Blanton LV, Frese SA, et al. Cultivating healthy growth and nutrition through the gut microbiota. Cell. 2015;161:36–48 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Koufaris C, Sismani C. Modulation of the genome and epigenome of individuals susceptible to autism by environmental risk factors. Int J Mol Sci. 2015;16:8699–8718 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Sampson TR, Mazmanian SK. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe. 2015;17:565–576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Schmidt C. Mental health: thinking from the gut. Nature. 2015;518:S12–S15 [DOI] [PubMed] [Google Scholar]
- 166.Millan MJ, Andrieux A, Bartzokis G, et al. Altering the course of schizophrenia: progress and perspectives. Nat Rev Drug Discov. 2016. [Epub ahead of print]; DOI: 10.1038/nrd.2016.28 [DOI] [PubMed] [Google Scholar]
- 167.Oriach CS, Robertson RC, Stanton C, et al. Food for thought: the role of nutrition in the microbiota-gut-brain axis. Clin Nutr Exp. 2016;6:25–38 [Google Scholar]
- 168.Maslowski KM, Mackay CR. Diet, gut microbiota and immune responses. Nat Immunol. 2011;12:5–9 [DOI] [PubMed] [Google Scholar]
- 169.Forslund K, Hildebrand F, Nielsen T, et al. Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature. 2015;528:262–266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Lee SH, de La Serre CB. Gut microbiome-brain communications regulate host physiology and behavior. J Nutr Health Food Sci. 2015;3:1–12 [Google Scholar]
- 171.Marette A, Jobin C. SCFAs take a toll en route to metabolic syndrome. Cell Metab. 2015;22:954–956 [DOI] [PubMed] [Google Scholar]
- 172.MacFabe DF, Cain DP, Rodriguez-Capote K, et al. Neurobiological effects of intraventricular propionic acid in rats: possible role of short chain fatty acids on the pathogenesis and characteristics of autism spectrum disorders. Behav Brain Res. 2007;176:149–169 [DOI] [PubMed] [Google Scholar]
- 173.MacFabe DF Short-chain fatty acid fermentation products of the gut microbiome: implications in autism spectrum disorders. Microb Ecol Health Dis. 2012;23:19260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.MacFabe D. Autism: metabolism, mitochondria, and the microbiome. Global Adv Health Med. 2013;2:52–66 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Galland L. The gut microbiome and the brain. J Med Food, 2014;17:1261–1272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Goldani AAS, Downs SR, Widjaja F, et al. Biomarkers in autism. Front Psychiatry. 2014;5:100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Foley KA, MacFabe DF, Kavaliers M, et al. Sexually dimorphic effects of prenatal exposure to lipopolysaccharide, and prenatal and postnatal exposure to propionic acid, on acoustic startle response and prepulse inhibition in adolescent rats: relevance to autism spectrum disorders. Behav Brain Res. 2015;278:244–256 [DOI] [PubMed] [Google Scholar]
- 178.Frye RE, Slattery J, MacFabe DF, et al. Approaches to studying and manipulating the enteric microbiome to improve autism symptoms. Microb Ecol Health Dis. 2015;26:26878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Mezzelani A, Landini M, Facchiano F, et al. Environment, dysbiosis, immunity and sex-specific susceptibility: a translational hypothesis for regressive autism pathogenesis. Nutr Neurosci. 2015;18:145–161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Shultz SR, Aziz NAB, Yang L, et al. Intracerebroventricular injection of propionic acid, an enteric metabolite implicated in autism, induces social abnormalities that do not differ between seizure-prone (FAST) and seizure-resistant (SLOW) rats. Behav Brain Res. 2015;278:542–548 [DOI] [PubMed] [Google Scholar]
- 181.Oyarzabal A, Bravo-Alonso I, Sánchez-Aragó M, et al. Mitochondrial response to the BCKDK-deficiency: some clues to understand the positive dietary response in this form of autism. BBA-Mol Basis Dis. 2016;1862:592–600 [DOI] [PubMed] [Google Scholar]
- 182.da Silva PGC, Domingues DD, de Carvalho LA, et al. Neurotrophic factors in Parkinson's disease are regulated by exercise: evidence-based practice. J Neurol Sci. 2016;363:5–15 [DOI] [PubMed] [Google Scholar]
- 183.Gwynette MF, Warren NJ, Warthen J, et al. Yoga as an intervention for patients with autism spectrum disorder: a review of the evidence and future directions. Autism Open Access. 2015;5:3 [Google Scholar]
- 184.Duzel E, van Praag H, Sendtner M. Can physical exercise in old age improve memory and hippocampal function? Brain. 2016;139:662–673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Luu K, Hall PA. Hatha yoga and executive function: a systematic review. J Altern Complement Med. 2016;22:125–133 [DOI] [PubMed] [Google Scholar]
- 186.O'Mahony SM, Clarke G, Dinan TG, et al. Early-life adversity and brain development: is the microbiome a missing piece of the puzzle? Neuroscience. 2015;322 [Epub ahead of print]; DOI: 10.1016/j.neuroscience.2015.09.068 [DOI] [PubMed] [Google Scholar]
- 187.Charbonneau MR, O'Donnell D, Blanton LV, et al. Sialylated milk oligosaccharides promote microbiota-dependent growth in models of infant undernutrition. Cell. 2016;164:859–871 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Cowan CSM, Callaghan BL, Kan JM, et al. The lasting impact of early-life adversity on individuals and their descendants: potential mechanisms and hope for intervention. Genes Brain Behav. 2016;15:155–168 [DOI] [PubMed] [Google Scholar]
- 189.Pennisi E. The right gut microbes help infants grow. Science. 2016;351:582. [DOI] [PubMed] [Google Scholar]
- 190.Blanton LV, Charbonneau MR, Salih T, et al. Gut bacteria that prevent growth impairments transmitted by microbiota from malnourished children. Science. 2016;351:aad3311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Schwarzer M, Makki K, Storelli G, et al. Lactobacillus plantarum strain maintains growth of infant mice during chronic undernutrition. Science. 2016;351:854–857 [DOI] [PubMed] [Google Scholar]
- 192.Thaiss CA, Elinav E. Exploring new horizons in microbiome research. Cell Host Microbe. 2014;15:662–667 [DOI] [PubMed] [Google Scholar]
- 193.Ash C, Mueller K. Manipulating the microbiota. Science. 2016;352:531. [DOI] [PubMed] [Google Scholar]
- 194.Zhang Y, Sun J, Lin CC, et al. The emerging landscape of salivary diagnostics. Periodontol 2000. 2016;70:38–52 [DOI] [PubMed] [Google Scholar]
References
Cite this article as: Moos WH, Faller DV, Harpp DN, Kanara I, Pernokas J, Powers WR, Steliou K (2016) Microbiota and neurological disorders: a gut feeling, BioResearch Open Access 5:1, 137–145, DOI: 10.1089/biores.2016.0010.